

Abstract
Common buckwheat is recognized as a healthy food because its seed contains large amounts of protein, minerals, and rutin. However, the yielding ability of common buckwheat is lower than that of other major crops. The short growing period, moisture injury, occurrence of sterile seeds due to lack of flower-visiting insects, and yield loss due to lodging and shattering cause low and unstable grain yield. Therefore, many common buckwheat breeders have tried to increase yielding ability by improving various characteristics. Recently, new breeding objectives for improving yielding ability by increasing preharvest sprouting resistance; reducing shattering loss; introducing self-compatibility; the ecotype, and semidwarf have been reported. In this review, we introduce the research on the important agronomic characteristics, preharvest sprouting resistance, ecotype and ecological differentiation, shattering resistance, and lodging resistance in common buckwheat.
Keywords: agronomic characteristics, ecotype, lodging resistance, preharvest sprouting resistance, shattering resistance
Introduction
Common buckwheat (Fagopyrum esculentum (L.) Moench) is recognized as a healthy food because its seed contains large amounts of protein, minerals, and the flavonoid rutin. Demand for common buckwheat grain is increasing (Japan-Buckwheat-Association 2019), but the yielding ability of common buckwheat is lower than that of other major crops. One reason for this low yielding ability is due to the short growing period, since there is significant correlation between grain yield and the length of the growing period (Morishita and Tetsuka 2001). While a short growing period (early maturity) is an important breeding objective for early harvest, improving yielding ability becomes a difficult problem. Many common buckwheat breeders have tried to increase yielding ability by developing cultivars with early maturity through overcoming inverse correlation between yield and growing period (Inuyama et al. 1994, Matsui et al. 2013, Morishita et al. 2013), lodging resistance (Matsui et al. 2013, Morishita et al. 2013, Yui et al. 2012), large seed (Nagatomo et al. 1982), downy mildew resistance (Honda et al. 1994), and determinate type (Funatsuki et al. 1996, Honda et al. 2009, Wagatsuma 2004). Although the yielding abilities of these cultivars improved on the original cultivars, they were unable to reach 100 kg/10 a in most buckwheat production areas (Statistics of Japan 2019). This can be explained by the fact that these cultivars were developed by minor improvement of original cultivars, so the yielding ability of these cultivars was improved only slightly. Furthermore, excess moisture injury (Sugimoto and Sato 2000), deterioration of soil fertility and poor manuring practice (Hayashi et al. 1994, Murayama et al. 1998), poor growth due to weed competition (Hatakenaka et al. 2015), occurrence of sterile seeds due to lack of the flower-visiting insects (Kasajima et al. 2017a), and yield loss due to lodging and shattering (Funatsuki et al. 2000) cause low and unstable grain yield. Recently, new breeding objectives for improving yielding ability by increasing preharvest sprouting resistance (Hara et al. 2012), reducing shattering loss (Suzuki et al. 2012a, 2012b); introducing self-compatibility (Matsui et al. 2008, Mukasa et al. 2010); the ecotype (Hara and Ohsawa 2013); and semidwarf (Morishita et al. 2015) have been reported. These new approaches would be beneficial for common buckwheat breeding. Here, we introduce preharvest sprouting resistance, ecotype and ecological differentiation, shattering resistance, and lodging resistance in common buckwheat.
Ecotype and ecological differentiation
Common buckwheat, a short-day plant, is a highly adaptable crop grown widely from low to high latitudes, although the cultivation season and cultivars are limited by latitude and altitude (Sugawara 2001). There are many native strains suitable for each climate in various places in Japan. Onda and Takeuchi (1942) categorized Japanese native common buckwheat strains by their ecological characteristics, such as time of maturation and nutrient growth characteristics in the production area, as northern type (early maturing and less vegetative growth), southern type (late maturing and vigorous vegetative growth), and central type (intermediate features). In flowering response to sowing season and photoperiod in this classification, the northern type shows no delay in flowering under long-day conditions, but the southern type shows prolonged flowering and increased vegetative growth under those conditions. In addition, Yamazaki (1947) classified Japanese common buckwheat into autumn, summer, and intermediate ecotypes according to earliness of flowering, vegetative growth amount, and yield when sowing at regular intervals from spring onward. In the autumn ecotype, stems and leaves flourish, flowering is delayed, seed set percentage decreases, and yield is markedly less when sowing from May to June (long-day condition) compared with that from August to September (short-day condition). In contrast, in the summer ecotype, early flowering occurs with sowing from May to June (long-day condition), seed set percentage is high, the difference in yield between the short-day and long-day condition is low, and the yield is higher in a long-day than in a short-day condition. The intermediate ecotype shows intermediate properties between autumn and summer ecotypes, and is further divided into intermediate autumn ecotype and intermediate summer ecotype. On the basis of these studies, the ecotype of current common buckwheat is classified by investigating the response to sowing time at regular intervals. Selection of ecotypes appropriate for each cultivation area and sowing time is important for a high and stable yield, since the use of an ecotype inappropriate for the environment at a particular site causes a significant decrease in yield (Nagase 2001).
The center of species in common buckwheat was reported as the southern part of China (Ohnishi 1990). It is considered to have been transmitted from the place of origin to the Kyushu region of Japan (Ujihara and Matano 1978) through North China and the Korean Peninsula (Murai and Ohnishi 1996). The following has been shown to be related to ecotype differentiation within Japan. Long-day treatment significantly delays flowering of common buckwheat compared to short-day treatment (Tabata et al. 1931); the critical photoperiod that delays flowering is 12 to 13 h. Low temperatures also delay flowering, and variation in flowering time within a population becomes large under long-day and high temperature conditions (Hsu 1936, 1938). On the basis of earliness of flowering, Ujihara and Matano (1975) identified many populations of the summer ecotype in the Tohoku to Hokkaido regions in the north of Japan, the autumn and intermediate ecotypes in the southwest, and all ecotypes from the Chubu to Kanto regions in central Japan. Although the summer ecotype tends to be favored in the northern districts and the autumn ecotype in the southwest areas, Ujihara and Matano (1975) presumed that the ecotypes differentiated owing to various factors such as altitude or sowing time, and that the present distribution of native strains is due to the distribution of various ecotypes at the same latitude (Ujihara and Matano 1975).
Based on these studies, Sugawara (2001) proposed that the flowering of common buckwheat is influenced by temperature but is dominated primarily by photoperiod, and that ecotype differentiation depends on reactivity to photoperiod: i.e., natural selection (Matano and Ujihara 1979, Ujihara and Matano 1978) and artificial selection (Minami and Namai 1986) were due primarily to differences in photoperiod in the culture season as the cultivation area of the autumn ecotype population spread northwards in the Japanese archipelago. In this proposal the summer ecotype is derived from the autumn ecotype.
Iwata et al. (2005) showed that genetic diversity tends to be lower in populations with early flowering (summer ecotype), and the genetic structure differs between autumn and summer ecotype populations. Hara and Ohsawa (2013) accurately evaluated photoperiod sensitivity and genetic diversity under a controlled environment. Their results support the hypothesis that summer ecotypes were derived from autumn ecotypes by adaptation to the climate in northern Japan, i.e., adaptation from short-day to long-day cultivation, as outlined in Fig. 1.
Fig. 1.

Hypothesis of ecological differentiation from autumn ecotype to summer ecotype. This hypothesis was considered based on phenotypic selection under fixed 15.0 h photoperiod conditions (JSPS KAKENHI, grant number JP16K18642) and several previous studies. a) In the long-day condition, the phenotypic variation in photoperiod sensitivity within the population is expanded (Hara and Ohsawa 2013, Minami and Namai 1986). b) On one individual, the seed set percentage and pollen fertility decrease in florets flowering after the full flowering time (Nagato et al. 1951, Nagatomo 1961, Ohsawa et al. 2001, Sugawara and Sugiyama 1954). Therefore, reproductive isolation occurs owing to asynchrony of flowering time, and assortative mating occurs between individuals close to flowering day rather than random mating (Ohsawa et al. 2001). c) In late-flowering individuals, the incidence of malformed flower increases significantly owing to long-day or high-temperature effects (Nagatomo 1961, Nakamura and Nakayama 1950, Sugawara 1958). Adaptability differs between individuals with early versus late flowering, with late-flowering individuals culled by natural selection. Harvesting after a certain cultivation period (about 80–90 days after sowing) artificially selects late-flowering individuals (Minami and Namai 1986). d) In this experiment, cultivation under long-day condition (fixed 15.0-h photoperiod) was carried out three times.
The inability to develop genetically fixed lines and cultivars due to outcrossing of common buckwheat has delayed breeding programs (Nakayama 1975). Ecotype breeding based on mass selection using phenotype values has been conducted, but if it becomes possible to predict the ecotype from the genotype, ecotype breeding could be performed without the influence of the environment of the breeding site or cultivation area. Furthermore, we consider that by fixing the photoperiod sensitivity genes, it is thought that ecotype breeding could be carried out more efficiently even in allogamous buckwheat. In recent years, data required for molecular breeding of common buckwheat, such as the draft genome (Yasui et al. 2016), buckwheat genome database (http://buckwheat.kazusa.or.jp), and a high-density linkage map (Yabe et al. 2014), have been used to increase the yielding ability by genomic selection (Yabe et al. 2018). Genomic studies of ecotype and difference in ecotype were conducted, and quantitative trait locus and ortholog genes (FeCRY1, FeELF3, FeGI, FePHY3 and FeTOC1) related to photoperiod sensitivity have been estimated (Hara et al. 2011, 2020). On the other hand, uniformity of the maturity time is also important in ecotype breeding of common buckwheat. Although the ortholog gene (GmGIa, highly homologous to FeGI) in soybean has been shown to be associated with adaptiveness, flowering and maturity (Watanabe et al. 2011), these relationships are not clear in common buckwheat. Thus, further genomic studies are expected to elucidate the details of the ecotypes, ecological differentiation and ecotype breeding.
Preharvest sprouting
Rain between grain maturity and harvest sometimes induces preharvest sprouting, as reported in Australia, Korea, and Japan (Bluett 2001, Choi et al. 1992, Hara et al. 2007, 2008, Morishita and Tetsuka 2001). Sprouting buckwheat grain has little market value (Bluett 2001). In most parts of Japan, buckwheat can be grown twice a year, i.e., in summer and autumn, although the latter is far more popular. Summer cultivation is increasing, because it can provide fresh grain when the demand for buckwheat soars in the Japanese market (Shibata 1981, Vinning 2001), and because it stabilizes production by hedging the risk of loss due to bad weather e.g., typhoon, early frost and excess rainfall. However, preharvest sprouting occurs especially in summer cultivation (Choi et al. 1992, Hara et al. 2007, Morishita and Tetsuka 2001), since the harvest time corresponds to the rainy season.
Buckwheat flour is processed into various foods (Ikeda 2002), among which cooked noodles are the most popular in Japan. Buckwheat noodles are often made from a mixture of buckwheat flour and wheat flour, egg, or yam to reduce their fragility; wheat flour is the most effective adjuvant (Ikeda 2002, Tsutsumi et al. 1990). Firmness of buckwheat noodles plays an important role in their palatability (Kimura et al. 2000, 2002). Hara et al. (2007, 2009a) studied the effect of preharvest sprouting on the textural characteristics of buckwheat noodles and pasting viscosity: sprouting decreased the peak force and peak strain when noodles made from buckwheat flour or buckwheat plus wheat flour were cut. Longer cooking also decreases peak force and peak strain of buckwheat noodles (Hara et al. 2009a, Kimura et al. 2002, Tsuji 1984), and is considered less preferable (Kimura et al. 2000, 2002, Tsuji 1984).
Hara et al. (2007) reported that preharvest sprouting lowered RVA (Rapid Visco Analyzer) peak viscosity because it digested buckwheat starch by activating alfa-amylase (Miyake et al. 2004). The decrease in peak viscosity caused by sprouting is not alleviated by adding wheat flour (Hara et al. 2007, Sugimoto 2003), because α-amylase in sprouted buckwheat flour also digests wheat starch (Sugimoto 2003).
Hara et al. (2007, 2009a) reported that buckwheat cultivars differ in their tendency to undergo preharvest sprouting. We evaluated cultivar difference in preharvest sprouting, by counting sprouted grain harvested after frequent natural rainfall in the rainy season. The ratio of the preharvest sprouted grain ranged from 28% to 81%. Pasting viscosity varies widely among cultivars that have undergone preharvest sprouting, whereas the pasting properties of purified starch vary little among cultivars (Yoshimoto et al. 2004). Hara et al. (2008) reported that cultivar differences in buckwheat preharvest sprouting are highly correlated with seed dormancy assayed by a petri dish experiment. Because this petri dish assay can be used to evaluate the propensity for preharvest sprouting of buckwheat cultivars, it allows selection in buckwheat breeding programs even when preharvest sprouting is not induced naturally. In the petri dish assay, seeds that did not germinate immediately after sampling were considered dormant, since nearly all seeds germinated after 6 months’ storage. These findings suggest that cultivar differences in buckwheat preharvest sprouting are explained mainly by seed dormancy.
In 2008, buckwheat breeders registered ‘Harunoibuki’ (mass-selected from the cultivar ‘Hashikamiwase’) as a new cultivar with improved preharvest sprouting resistance (Hara et al. 2009b) that is suitable for summer cultivation and has the highest yield among cultivars grown at the same time. As far as we are aware, this is the first reported trial in improving preharvest sprouting tolerance in buckwheat. We consider that the success of this breeding effort is partly due to intra-varietal diversity in preharvest sprouting. In 2018, a cultivar ‘NARO-FE-1’ with lower preharvest sprouting than ‘Harunoibuki’ was developed. This cultivar was mass-selected from the progeny of crosses between eight cultivars. We demonstrated that preharvest sprouting, possibly leading to yield loss, can be reduced by breeding effort, by exploiting intra- and inter-varietal diversity in seed dormancy in buckwheat. Diversity in preharvest sprouting should be further exploited to develop cultivars with even lower preharvest sprouting.
Hara et al. (2008) reported that preharvest sprouting and main stem length are negatively correlated. Diversity in main stem length of buckwheat is related to ecotype (Michiyama et al. 2005), main stem length being longer in autumn ecotype cultivars than in summer ecotype cultivars (Michiyama and Hayashi 1998, Michiyama et al. 2005). Thus, autumn ecotype cultivars might be more tolerant to preharvest sprouting. The summer ecotype of buckwheat is considered to have derived from the autumn ecotype (Hara and Ohsawa 2013, Iwata et al. 2005, Minami and Namai 1986). In double cropping, summer ecotype cultivars could show a greater yield than autumn ecotype cultivars, the yield of which is greatly reduced by long-day conditions (Michiyama and Hayashi 1998, Yamazaki 1947). Double cropping of buckwheat in the history of buckwheat cultivation in Japan is summarized in Onda and Takeuchi (1942) and Sotokawa et al. (1988). Generally cultivated species are under automatic selection pressure to lose dormancy (Harlan et al. 1973). Many years of double cropping using summer ecotype cultivars could hasten the selection of lower seed dormancy, since the duration between harvesting and the next sowing is shorter than that in single cropping.
Further improvement in PHS tolerance in buckwheat would be necessary, because higher air temperatures, expected in global warming, raise the risk of preharvest sprouting (Hara et al. 2008). In addition to the cultivars with low preharvest sprouting mentioned above, a wild species F. homotropicum, known for its very strong seed dormancy (Wang and Campbell 2000) and compatibility to F. esculentum, should be a promising gene donor, although its extreme shattering habit (brittle pedicel) would need to be removed (Matsui et al. 2003). In other crops like wheat, barley, maize, and rice, genetic analysis of preharvest sprouting and dormancy has discovered many QTLs and some causal genes (Nakamura 2018). These advances are contributing to breeding efforts through marker-assisted selection and pyramiding of QTLs and genes for several different traits (Kumar et al. 2010, Tyagi et al. 2014). We are currently conducting a genetic analysis of preharvest sprouting and dormancy in common buckwheat.
Shattering resistance
Seed shattering is an important problem because it causes considerable yield loss. As buckwheat easily shatters and is resulting in yield loss (Morishita and Suzuki 2017, Suzuki et al. 2012a), shattering resistant cultivars are required. The two cultivated species of buckwheat (F. esculentum and F. tartaricum) do not have an abscission layer across the pedicels (Oba et al. 1998). Therefore, pedicel breaking is the most important cause of shattering. However, the breaking tensile strengths of buckwheat are much lower than those of other crops, such as rice (Ji et al. 2006), and correlate well with pedicel diameter (Fujimura et al. 2001, Oba et al. 1999).
Alekseeva et al. (1988) reported that the strong pedicel of the ‘green-flower’ trait is controlled by a single recessive gene. Mukasa et al. (2008) also reported that both traits are controlled by a single recessive gene and proposed that green-flower may be useful as breeding material for shattering resistance. Because of its single-gene inheritance, it is easy to introduce the green-flower trait by cross breeding. On the other hand, Mukasa et al. (2008) reported that a few whit: flowers show the same tensile strength level of the green-flower type, and this shows the possibility that a gene unrelated to the green-flower type exists. Nagai et al. (2016a, 2016b) also reported a shattering resistant gene with a strong linkage to green-flower and some shattering resistant genes of unrelated flower color in anti-tension type shattering by QTL analysis. These reports suggest that further improvement is possible by genetic accumulation.
Suzuki et al. (2012a) developed a green-flower line named ‘W/SK86GF’ in 1999, which was developed from progeny of hybridization between ‘Kitawasesoba’ × ‘Skorosperaya 86’ (Fig. 2). Green-flower breeding line ‘W/SK86GF’ showed better shattering resistance than ‘Kitawasesoba’ in field tests in 2002, 2003, 2004 (a heavy typhoon year) and 2011. Thus, green-flower is a promising trait for reducing shattering loss. Furthermore, Suzuki et al. (2012b) investigated varietal differences of breaking tensile strength in 23 buckwheat cultivars and breeding lines including 3 green-flower lines. The green-flower lines had higher breaking tensile strengths (Fig. 3). Consequently, green-flower appears to be promising as breeding material for shatter resistant cultivars.
Fig. 2.

Image of flowers, seeds and a pedicel section of buckwheat cultivars ‘W/SK86GF’ (left) and ‘Kitawasesoba’ (right). Scale bars, 100 μm. This figure is rearranged from Suzuki et al. (2012a, 2012b).
Fig. 3.
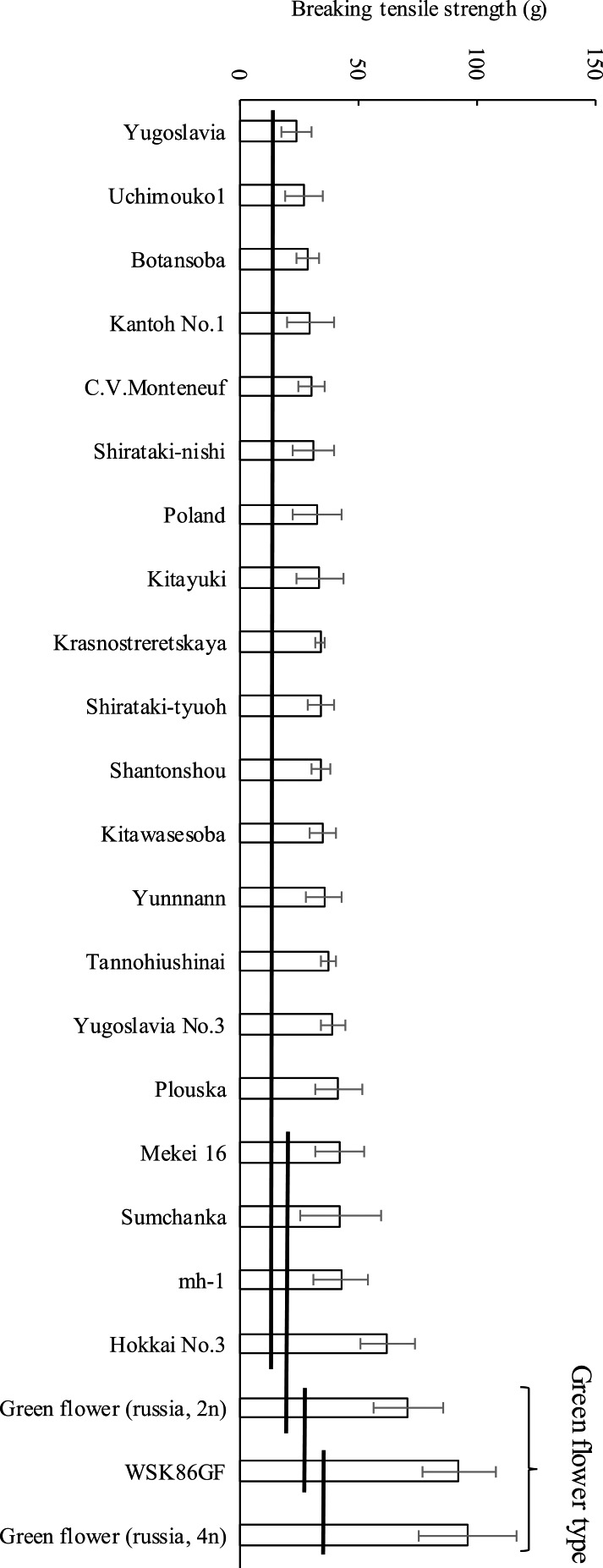
Cultivar difference in breaking tensile strength measured at maturation. Data are means ± SD of 15 individuals. The means of cultivars joined by the same horizontal rule are not significantly different at p < 0.05 (Ryan’s multiple range test). This figure is modified Suzuki et al. (2012b).
Yamaguchi et al. (2010) reported that the ‘threshing and sorting loss’ of shattering-resistant soybean was increased and suggested that the decreased pre-harvest shatter loss was offset. Therefore, it is necessary to carry out a practical examination to inspect whether the green-flower breeding line is suitable for practical use. Morishita and Suzuki (2017) performed a large-scale practical test of the green-flower shattering-resistant breeding line ‘Mekei 35’ by using a seeding machine and combine harvester. Yield loss was divided into four factors; pre-harvest shatter loss, ‘head loss’ (due to collision with the front of the combine harvester), ‘threshing and sorting loss’ (due to grain being passed by the combine harvester and thrown away) and ‘unfinished cutting loss’ (due to incomplete harvest). At the normal harvest time with the combine harvester, the total loss of ‘Mekei 35’ was greater than that of ‘Kitawasesoba’ because of a much higher ‘threshing and sorting loss’. On the other hand, in late harvesting with the combine harvester, despite an increased ‘head loss’ in ‘Mekei 35’, the ‘threshing and sorting loss’ of ‘Mekei 35’ decreased and the ‘pre-harvest shatter loss’ of ‘Kitawasesoba’ increased dramatically, so the total loss ratio of the ‘Mekei 35’ was lower than that of ‘Kitawasesoba’. Furthermore, the use of ‘Mekei 35’ was effective for reducing yield loss under conditions when shattering occurs heavily, such as in rain, strong wind, and late to extremely late harvesting. These experiments indicated that it is necessary to reduce ‘threshing and sorting loss’ by adjustment of the operating conditions of the combine harvester and choosing a suitable harvest time with the combine harvester for practical use with shattering-resistant cultivars. It seems likely that a shattering-resistant cultivar will be developed in the near future.
Lodging resistance
The tendency of buckwheat to lodge also reduces yield and product quality. Therefore, lodging resistance has been an important breeding objective. High plant height can increase lodging (Michiyama and Sakurai 1999, Murayama et al. 1998, 2004). Consistent with this, short plant height has been noted for improving lodging resistance. Determinate type cultivar ‘Kitanomashu’ has higher lodging resistance than indeterminate type cultivar ‘Kitawasesoba’ because of short plant height (Honda et al. 2009) and following cultivar ‘Reranokaori’ has higher lodging resistance than ‘Kitawasesoba’ because of slightly shorter plant height (Morishita et al. 2013). However, the lodging resistance of these two cultivars was insufficient for improving yielding ability. On the other hand, Tetsuka and Morishita (1999) reported that stem diameter at the ground is an important factor affecting lodging. Therefore, there is a demand for new common buckwheat types with short plant height and large stem diameter, that is to say dwarf or semidwarf type. In this section, we discuss the possibilities of the semidwarf trait.
Semidwarf rice and wheat have been researched from the latter half of the 20th century and the introduction of semidwarf rice and wheat cultivars contributed to the massive increase in grain yield of the Green Revolution (Athwal 1971, Hedden 2003). The use of semidwarf buckwheat cultivars may improve not only lodging resistance but also yielding ability. Ohnishi and Nagakubo (1982) discovered dwarf types of common buckwheat from farmers’ fields and identified 6 single recessive dwarf genes, dwA, dwB, dwC, dwD, dwE and dwF. Furthermore, Minami et al. (1999) reported line ‘G410’, with an incompletely dominant dwarfing gene. However, these reports did not discuss the relationship between semidwarf traits and yielding ability. In addition, the yield of the ‘semidwarf line’ was lower than that of wild type cultivars (Gubbels and Campbell 1986). For these reasons, practical semidwarf common buckwheat cultivars have not been developed. On the other hand, two semidwarf genes, sdA and sdB have been identified in Tartary buckwheat (Fagopyrum tartaricum) (Morishita et al. 2010); the yielding ability of an sdA line group was higher than that of the sdB line group, and an sdA line was registered as a new cultivar, ‘Daruma Dattan’ (Ministry of Agriculture, Forestry and Fisheries 2013). Thus, breeding of semidwarf common buckwheat has lagged behind that of semidwarf Tartary buckwheat. Recently, Morishita et al. (2015) discovered novel semidwarf materials in their breeding population of common buckwheat (Fig. 4). This semidwarf trait was controlled by a single recessive gene that was different from those reported previously. Morishita and Tetsuka (2001) reported a significant positive correlation between plant height and seed yield in common buckwheat. The ratios of plant height and seed yield for ‘Kitawasesoba’ of ‘semidwarf material’ are 60% and 84%, respectively (Table 1). Although, the plant height of this semidwarf is low, it has a relatively high ratio of seed yield. Furthermore, this ‘semidwarf material’ responded well to nitrogen fertilizer (Kasajima et al. 2017b). The yield of ‘semidwarf material’ may increase according to quantity of fertilization. It is necessary to establish suitable cultivation technology for showing the potency of yielding ability. Thus, it is anticipated that semidwarf common buckwheat will show promise for improving lodging resistance and yielding ability.
Fig. 4.

‘Semidwarf material’ (top) and wild type ‘Kitawasesoba’ (bottom). This figure is cited from Morishita et al. (2015).
Table 1.
Characteristics of semidwarf material
| Line and variety | Plant height (cm) | Stem diameter (mm) | Seed yield (kg/10 a) |
|---|---|---|---|
| Semidwarf | 65 (60) | 6.7 (104) | 140 (84) |
| Kitawasesoba | 109 (100) | 6.4 (100) | 165 (100) |
Average from 2011 to 2013.
Parentheses show the ratio of ‘Kitawasesoba’.
Literature Cited
- Alekseeva E.S., Malikov V.G. and Falendysh L.G. (1988) Green flower form of buckwheat. Fagopyrum 8: 79–82. [Google Scholar]
- Athwal D.S. (1971) Semidwarf rice and wheat in global food needs. Q. Rev. Biol. 46: 1–34. [DOI] [PubMed] [Google Scholar]
- Bluett, C. (2001) Managing buckwheat production in Australia. Rural Industries Research and Development Corporation, Canberra. [Google Scholar]
- Choi B.H., Park K.Y. and Park R.K. (1992) A study of cultural methods for summer buckwheat sown in spring. Kor. J. Crop Sci. 37: 149–154. [Google Scholar]
- Fujimura Y., Oba S. and Horiuchi T. (2001) Effects of fertilization and polyploidy on grain shedding habit of cultivated buckwheat (Fagopyrum spp.). Jpn. J. Crop Sci. 70: 221–225. [Google Scholar]
- Funatsuki H., Suvorova G.N. and Sekimura K. (1996) Determinate type variants in Japanese buckwheat lines. Breed. Sci. 46: 275–277. [Google Scholar]
- Funatsuki H., Maruyama-Funatsuki W., Fujino K. and Agatsuma M. (2000) Ripening habit of buckwheat. Crop Sci. 40: 1103–1108. [Google Scholar]
- Gubbels G.H. and Campbell C.G. (1986) Effect of seeding rate on height, yield and quality of large-seeded and semi-dwarf buckwheat genotypes. Can. J. Plant Sci. 66: 61–66. [Google Scholar]
- Hara T., Matsui K., Noda T. and Tetsuka T. (2007) Effects of preharvest sprouting on flour pasting viscosity in common buckwheat (Fagopyrum esculentum Moench). Plant Prod. Sci. 10: 361–366. [Google Scholar]
- Hara T., Tetsuka T., Matsui K., Ikoma H. and Sugimoto A. (2008) Evaluation of cultivar differences in preharvest sprouting of common buckwheat (Fagopyrum esculentum Moench). Plant Prod. Sci. 11: 82–87. [Google Scholar]
- Hara T., Sasaki T., Tetsuka T., Ikoma H. and Kohyama K. (2009a) Effects of sprouting on texture of cooked buckwheat (Fagopyrum esculentum Moench) noodles. Plant Prod. Sci. 12: 492–496. [Google Scholar]
- Hara T., Matsui K., Ikoma H. and Tetsuka T. (2009b) Cultivar difference in grain yield and preharvest sprouting in buckwheat (Fagopyrum esculentum Moench). Jpn. J. Crop Sci. 78: 189–195. [Google Scholar]
- Hara T., Iwata H., Okuno K., Matsui K. and Ohsawa R. (2011) QTL analysis of photoperiod sensitivity in common buckwheat by using markers for expressed sequence tags and photoperiod-sensitivity candidate genes. Breed. Sci. 61: 394–404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hara T., Tetsuka T. and Matsui K. (2012) New buckwheat cultivar “Harunoibuki”. Bull. NARO Kyushu Okinawa Agric. Res. Cent. 58: 37–47. [Google Scholar]
- Hara T. and Ohsawa R. (2013) Accurate evaluation of photoperiodic sensitivity and genetic diversity in common buckwheat under a controlled environment. Plant Prod. Sci. 16: 247–254. [Google Scholar]
- Hara T., Shima T., Nagai H. and Ohsawa R. (2020) Genetic analysis of photoperiod sensitivity associated with difference in ecotype in common buckwheat. Breed. Sci. 70: 101–111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harlan J.R., de Wet J. M. J. and Price E.G. (1973) Comparative evolution of cereals. Evolution 27: 311–325. [DOI] [PubMed] [Google Scholar]
- Hatakenaka K., Sekinuma M., Okabe M. and Kasuga S. (2015) Examination of seeding time and amount of seeding in buckwheat ‘SUNRUTIN’ cultivation on the Nobeyama Plateau. Hokuriku Crop Sci. 50: 64–66. [Google Scholar]
- Hayashi Y., Kobayashi T., Kuwahara M., Ichimura T. and Kiriyama H. (1994) Some factors affecting low yield of buckwheat in liyama pilot farm. 1. The actual state of buckwheat cultivation in Iiyama pilot farm. Hokuriku Crop Sci. 29: 85–87. [Google Scholar]
- Hedden P. (2003) The genes of the Green Revolution. Trends Genet. 19: 5–9. [DOI] [PubMed] [Google Scholar]
- Honda Y., Inuyama S., Kimura M., Furuyama S. and Kasano H. (1994) The breeding and characteristics of a buckwheat cultivar, “Kitayuki”. Res. Bull. Hokkaido Natl. Agric. Exp. Stn. 159: 11–21. [Google Scholar]
- Honda Y., Mukasa Y., Suzuki T., Maruyama-Funatsuki W., Funatsuki H., Sekimura K., Kato S. and Agatsuma M. (2009) The breeding and characteristics of a commo buckwheat cultivar, “Kitanomashu”. Res. Bull. Natl. Res. Cent. for Hokkaido Reg. 191: 41–52. [Google Scholar]
- Hsu C. (1936) A study on the effect of seasonal variation of sunshine hours and temperature on the reproductive stage of crops. Nougyo Oyobi Engei 11: 2155–2163. [Google Scholar]
- Hsu C. (1938) A study on the effect of seasonal variation of sunshine hours and temperature on the reproductive stage of crops (2nd report). A study on the flowering period and the correctness of flowering of buckwheat. Nougyo Oyobi Engei 13: 1601–1612. [Google Scholar]
- Ikeda K. (2002) Buckwheat: composition, chemistry, and processing. Adv. Food Nutr. Res. 44: 395–434. [DOI] [PubMed] [Google Scholar]
- Inuyama S., Honda Y., Furuyama S., Kimura M. and Kasano H. (1994) The breeding and characteristics of a buckwheat cultivar, “Kitawasesoba”. Res. Bull. Hokkaido Natl. Agric. Exp. Stn. 159: 1–10. [Google Scholar]
- Iwata H., Imon K., Tsumura Y. and Ohsawa R. (2005) Genetic diversity among Japanese indigenous common buckwheat (Fagopyrum esculentum) cultivars as determined from amplified fragment length polymorphism and simple sequence repeat markers and quantitative agronomic traits. Genome 48: 367–377. [DOI] [PubMed] [Google Scholar]
- Japan-Buckwheat-Association (2019) http://nihon-soba-kyokai.or.jp/uploads/1/2/3/9/123974412/jyukyuu29.pdf (accessed May 28 2019).
- Ji H.S., Chu S.H., Jiang W.Z., Cho Y.I., Hahn J.H., Eun M.Y., McCouch S.R. and Koh H.J. (2006) Characterization and mapping of a shattering mutant in rice that corresponds to a block of domestication genes. Genetics 173: 995–1005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kasajima S., Katagiri C., Ito H., Morishita T., Suzuki T. and Mukasa Y. (2017a) Growth and yield of self-compatible and hybrid common buckwheat lines pollinated with and without flies. Plant Prod. Sci. 20: 384–388. [Google Scholar]
- Kasajima S., Hatae K. and Morishita T. (2017b) Seed-setting habit of a semidwarf common buckwheat line. Fagopyrum 34: 5–12. [Google Scholar]
- Kimura T., Sugahara T., Sasaki H., Kameda K. and Nanba T. (2000) Effects of addition of konnyaku (Amorphophallus konjak) powder on the quality of soba noodles. J. Cookery Sci. Jpn. 33: 307–314. [Google Scholar]
- Kimura T., Sasaki H., Kameda K., Sugahara T. and Ishikawa K. (2002) Effects of tobiko (Konnyaku powder) on the quality of soba (Buckwheat) noodles. J. Cookery Sci. Jpn. 35: 266–274. [Google Scholar]
- Kumar J., Mir R.R., Kumar N., Kumar A., Mohan A., Prabhu K.V., Balyan H.S. and Gupta P.K. (2010) Marker-assisted selection for pre-harvest sprouting tolerance and leaf rust resistance in bread wheat. Plant Breed. 129: 617–621. [Google Scholar]
- Matano T. and Ujihara A. (1979) Agroecological classification and geographical distribution of the common buckwheat, Fagopyrum esculentum M. in the East Asia. Jpn. Agric. Res. Q. 13: 157–162. [Google Scholar]
- Matsui K., Tetsuka T. and Hara T. (2003) Two independent gene loci controlling non-brittle pedicels in buckwheat. Euphytica 134: 203–208. [Google Scholar]
- Matsui K., Tetsuka T., Hara T. and Morishita T. (2008) Breeding and characterization of a new self-compatible common buckwheat parental line, “Buckwheat Norin-PL-1”. Bull. NARO Kyushu Okinawa Agric. Res. Cent. 49: 11–17. [Google Scholar]
- Matsui K., Hara T., Tetsuka T. and Morishita T. (2013) New common buckwheat cultivar ‘Sachi-izumi’ for warm area in Japan. Bull. NARO Kyushu Okinawa Agric. Res. Cent. 59: 23–37. [Google Scholar]
- Michiyama H. and Hayashi H. (1998) Differences of growth and development between summer and autumn-type cultivars in common buckwheat (Fagopyrum esculentum Moench). Jpn. J. Crop Sci. 67: 323–330. [Google Scholar]
- Michiyama H. and Sakurai S. (1999) Effect of day and night temperatures on the growth and development of common buckwheat (Fagopyrum esculentum Moench). Jpn. J. Crop Sci. 68: 401–407. [Google Scholar]
- Michiyama H., Tsuchimoto K., Tani K., Hirano T., Hayashi H. and Campbell C. (2005) Influence of day length on stem growth, flowering, morphology of flower clusters, and seed-set in buckwheat (Fagopyrum esculentum Moench). Plant Prod. Sci. 8: 44–50. [Google Scholar]
- Minami H. and Namai H. (1986) Populational change in flowering time caused by different harvesting date observed in a late-summer type cultivar Miyazakizairai of buckwheat (Fagopyrum esculentum Moench). Japan. J. Breed. 36: 155–162. [Google Scholar]
- Minami M., Ujihara A. and Campbell C.G. (1999) Morphology and inheritance of dwarfism in common buckwheat line, G410, and its stability under different growth condition. Breed. Sci. 49: 27–32. [Google Scholar]
- Ministry of Agriculture, Forestry and Fisheries (2013) Plant Variety Protection. DARUMADATTAN. http://www.hinshu2.maff.go.jp/vips/cmm/apCMM112.aspx?TOUROKU_NO=22633&LANGUAGE=English (accessed June 5, 2019).
- Miyake K., Morita R., Handoyo T., Maeda T. and Morita N. (2004) Characterization of graded buckwheat flours and some properties of germinated ‘Mancan’ buckwheat grains. Fagopyrum 21: 91–97. [Google Scholar]
- Morishita T. and Tetsuka T. (2001) Year-to-year variation and varietal difference of agronomic characters of common buckwheat in the kyushu area. Jpn. J. Crop Sci. 70: 379–386. [Google Scholar]
- Morishita T., Mukasa Y., Suzuki T., Shimizu A., Yamaguchi H., Degi K., Aii J., Hase Y., Shikazono N., Tanaka A. et al. (2010) Characteristics and inheritance of the semidwarf mutants of Tartary buckwheat (Fagopyrum tartaricum Gaertn.) induced by gamma ray and ion beam irradiation. Breed. Res. 12: 39–43. [Google Scholar]
- Morishita T., Suzuki T., Mukasa Y. and Honda Y. (2013) Breeding of a new common buckwheat cultivar ‘Reranokaori’, and its characteristics. Res. Bull. NARO Hokkaido Agric. Res. Cent. 199: 1–12. [Google Scholar]
- Morishita T., Suzuki T. and Mukasa Y. (2015) Characteristics of a novel ‘semidwarf materials’ in common buckwheat. Fagopyrum 32: 9–14. [Google Scholar]
- Morishita T. and Suzuki T. (2017) The evaluation of harvest loss occurring in the ripening period using a combine harvester in a shattering-resistant line of common buckwheat. Jpn. J. Crop Sci. 86: 62–69. [Google Scholar]
- Mukasa Y., Suzuki T. and Honda Y. (2008) Inheritance of Green-flower trait and the accompanying strong pedicel in common buckwheat. Fagopyrum 25: 15–20. [Google Scholar]
- Mukasa Y., Suzuki T. and Honda Y. (2010) A methology for heterosis breeding of common buckwheat involving the use of the self-compatibilty gene derived from Fagopyrum homotropicum. Euphytica 172: 207–214. [Google Scholar]
- Murai M. and Ohnishi O. (1996) Population genetics of cultivated common buckwheat, Fagopyrum esculentum Moench. X. Diffusion routes revealed by RAPD markers. Genes Genet. Syst. 71: 211–218. [DOI] [PubMed] [Google Scholar]
- Murayama S., Oana Y., Nakayama T. and Ogihara H. (1998) Effect of long term continuous application of organic fertilizer and three major nutrients (NPK elements) on the growth and yield of buckwheat (Fagopyrum esculentum M.). Hokuriku Crop Sci. 33: 118–120. [Google Scholar]
- Murayama S., Miyamoto K. and Yanokuchi Y. (2004) Effects of variety, amount of fertilizer application and seeding rate on lodging of common buckwheat. Hokuriku Crop Sci. 40: 78–81. [Google Scholar]
- Nagai, H., R. Ohsawa and T. Hara (2016a) Genetic analysis of seed shattering in common buckwheat. The 13th International Symposium on Buckwheat, pp. 223–227. [Google Scholar]
- Nagai H., Morishita T., Ohsawa R. and Hara T. (2016b) Genetic analysis of seed shattering without the formation of an abscission layer in common buckwheat. Breed. Res. 18: 232. [Google Scholar]
- Nagase, Y. (2001) Soba no kigen to tokusei. Rural Culture Association Japan. Tensakuzensyo 3zakkoku. Tokyo, pp. 447–467. [Google Scholar]
- Nagato K., Sato T. and Sugahara K. (1951) On the fruiting of buckwheat. Jpn. J. Crop Sci. 19: 299–302. [Google Scholar]
- Nagatomo T. (1961) Studies on physiology of reproduction and some cases of inheritance in buckwheat. Research Report of plant Breeding Laboratory, Faculty of Agriculture, Miyazaki University 1: 1–213. [Google Scholar]
- Nagatomo T., Adachi T. and Yabuya T. (1982) A new buckwheat cultivar ‘Miyazakiootsubu’. Bull. Fac. Agric. Miyazaki Univ. 29: 293–305. [Google Scholar]
- Nakamura M. and Nakayama H. (1950) On the enervative sterility in buckwheat. Jpn. J. Crop Sci. 19: 122–125. [Google Scholar]
- Nakamura S. (2018) Grain dormancy genes responsible for preventing pre-harvest sprouting in barley and wheat. Breed. Sci. 68: 295–304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakayama K. (1975) Genecology and future breeding prospects of buckwheat. Nougyo Oyobi Engei 50: 497–502. [Google Scholar]
- Oba S., Suzuki Y. and Fujimoto F. (1998) Breaking strength of pedicel and grain shattering habit in two species of buckwheat (Fagopyrum spp.). Plant Prod. Sci. 1: 62–66. [Google Scholar]
- Oba S., Ohta A. and Fujimoto F. (1999) Breaking strength of pedicel as an index of grain-shattering habit in autotetraploid and diploid buckwheat (Fagopyrum esculentum Moench.) cultivars. Plant Prod. Sci. 2: 190–195. [Google Scholar]
- Ohnishi O. and Nagakubo T. (1982) Population genetics of cultivated common buckwheat, Fagopyrum esculentum Moench. II. Frequency of dwarf mutants in Japanese populations. Jpn. J. Genet. 57: 641–650. [Google Scholar]
- Ohnishi O. (1990) Discovery of the wild ancestor of common buckwheat. Fagopyrum 11: 5–10. [Google Scholar]
- Ohsawa, R., K. Ishikawa and H. Namai (2001) Assortative mating in the population of intermediate ecotype of common buckwheat with special reference to flowering time, pollen fertility and a rate of malformed pistil. Advance in Buckwheat Research, pp. 240–247. [Google Scholar]
- Onda S. and Takeuchi T. (1942) Ecotype of Japanese buckwheat varieties. Nougyo Oyobi Engei 17: 971–974. [Google Scholar]
- Shibata S. (1981) Komugi, Soba no chozo to Kako. Nougyo Oyobi Engei 56: 131–136. [Google Scholar]
- Sotokawa M., Tada H., Fujita M., Kumagai K. and Takemura T. (1988) Cultivation technique for stable and high yield of buckwheat and trial production of compact harvesters. Bull. Aomori Field Crops Hort. Exp. Stn. 6: 85–114. [Google Scholar]
- Statistics of Japan (2019) https://www.e-stat.go.jp/stat-search/files?page=1&layout=datalist&toukei=00500215&tstat=000001013427&cycle=7&year=20180&month=0&tclass1=000001032288&tclass2=000001032753&tclass3=000001125936 (accessed May 28 2019).
- Sugawara K. and Sugiyama K. (1954) An ecological study on the flowering and seed setting of buckwheat. The annual report of the Department of Liberal Arts, the Iwate University 6: 55–68. [Google Scholar]
- Sugawara K. (1958) On the injury of buckwheat pistil: retardation of pistil growth as influenced by day-length. Jpn. J. Crop Sci. 26: 269–270. [Google Scholar]
- Sugawara, K. (2001) Nihonjin to soba. Rural Culture Association Japan. Tensakuzensyo 3zakkoku. Tokyo, pp. 427–435. [Google Scholar]
- Sugimoto H. and Sato T. (2000) Effects of excessive soil moisture at different growth stages on seed yield of summer buckwheat. Jpn. J. Crop Sci. 69: 189–193. [Google Scholar]
- Sugimoto M. (2003) Hatsuga soba yurai no α-amylase niyoru soba men no bussei teika. Syokuhin to Gijutsu 381: 10–12. [Google Scholar]
- Suzuki T., Mukasa Y., Morishita T., Takigawa S. and Noda T. (2012a) Traits of shattering resistant buckwheat ‘W/SK86GF’. Breed. Sci. 62: 360–364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki T., Mukasa Y., Morishita T., Takigawa S. and Noda T. (2012b) Varietal differences in annual variation of breaking tensile strength in common buckwheat. Fagopyrum 29: 13–16. [Google Scholar]
- Tabata K., Ogata K. and Sukegawa K. (1931) On the influence of long and short days on growth, flowering and seed setting of common buckwheat and soybean. Jpn. J. Crop Sci. 3: 188–202. [Google Scholar]
- Tetsuka T. and Morishita T. (1999) Relationships between lodging and characteristics of Japanese landraces of buckwheat. Rep. Kyusyu Br. Crop Sci. Soc. Japan 65: 79–80. [Google Scholar]
- Tsuji S. (1984) Textural measurement of mouthfeel of cooked “soba” by using the textural instrument—Investigations on the fracture properties. Nippon Shokuhin Kogyo Gakkaishi 31: 61–65. [Google Scholar]
- Tsutsumi C., Yamagishi J., Motoyama Y., Mitsuhashi F. and Yoshinaka T. (1990) Time change of the buckwheat noodles by the difference of the thickening (TSUNAGI). J. Cookery Sci. Jpn. 23: 373–381. [Google Scholar]
- Tyagi S., Mir R.R., Kaur H., Chhuneja P., Ramesh B., Balyan H.S. and Gupta P.K. (2014) Marker-assisted pyramiding of eight QTLs/genes for seven different traits in common wheat (Triticum aestivum L.). Mol. Breed. 34: 167–175. [Google Scholar]
- Ujihara A. and Matano T. (1975) Characteristics of flowering, fertilization and seed setting of buckwheat—Approach to analysis of yield establishment process. Nougyogijyutu 30: 406–408. [Google Scholar]
- Ujihara A. and Matano T. (1978) Tsushima’s buckwheat—on the propagation and ecotype differentiation of Japanese buckwheat—. Noukounogijyutu 1: 43–59. [Google Scholar]
- Vinning, G. (2001) Buckwheat: Demand-supply analysis. Rural Industries Research and Development Corporation, Canberra. [Google Scholar]
- Wagatsuma T. (2004) Selection of determinate type plants from common buckwheat cultivar ‘Kitawasesoba’ and development of a new line, Horokei 3. J. Rakuno Gakuen Univ. 29: 1–7. [Google Scholar]
- Wang Y.J. and Campbell C.G. (2000) Breaking dormancy in buckwheat. Fagopyrum 17: 45–50. [Google Scholar]
- Watanabe S., Xia Z., Hideshima R., Tsubokura Y., Sato S., Yamanaka N., Takahashi R., Anai T., Tabata S., Kitamura K. et al. (2011) A map-based cloning strategy employing a residual heterozygous line reveals that the GIGANTEA gene is involved in soybean maturity and flowering. Genetics 188: 395–407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yabe S., Hara T., Ueno M., Enoki H., Kimura T., Nishimura S., Yasui Y., Ohsawa R. and Iwata H. (2014) Rapid genotyping with DNA micro-arrays for high-density linkage mapping and QTL mapping in common buckwheat (Fagopyrum esculentum Moench). Breed. Sci. 64: 291–299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yabe S., Hara T., Ueno M., Enoki H., Kimura T., Nishimura S., Yasui Y., Ohsawa R. and Iwata H. (2018) Potential of genomic selection in mass selection breeding of an allogamous crop: an empirical study to increase yield of common buckwheat. Front. Plant Sci. 9: 276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi M., Nagashima H., Yuzawa M., Yamada T., Hajika M. and Funatsuki H. (2010) Yield and quality losses to late machine harvest of soybean introduced shuttering resistance gene. Jpn. J. Crop Sci. 79 (Ex.1): 206–207. [Google Scholar]
- Yamazaki Y. (1947) Buckwheat. Nogyo 778: 16–32. [Google Scholar]
- Yasui Y., Hirakawa H., Ueno M., Matsui K., Tanaka T.K., Yang S.J., Aii J., Sato S. and Mori M. (2016) Assembly of the draft genome of buckwheat and its applications in identifying agronomically useful genes. DNA Res. 23: 215–224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshimoto Y., Egashira T., Hanashiro I., Ohinata H., Takase Y. and Takeda Y. (2004) Molecular structure and some physicochemical properties of buckwheat starches. Cereal Chem. 81: 515–520. [Google Scholar]
- Yui M., Yamamori M., Honda H., Kato M. and Kawasaki M. (2012) ‘Nijiyutaka’, a new buckwheat cultivar. Bull. Tohoku Agric. Res. Cent. 114: 11–12. [Google Scholar]