

Abstract
Protective role of the skin is against external insults and maintenance of electrolyte homeostasis of the body. Cutaneous dysfunction can account for the development of both cutaneous and systemic disorders. Thus, improvements in cutaneous functions can benefit a number of extracutaneous and cutaneous functions. Resveratrol, a natural ingredient, displays multiple benefits for various systems/organs, including the skin. The benefits of resveratrol for cutaneous functions include stimulation of keratinocyte differentiation and antimicrobial peptide expression, inhibition of keratinocyte proliferation and cutaneous inflammation, UV protection, anticancer, antiaging, and inhibition of melanogenesis. The mechanisms of action of resveratrol include activation of sirtuin 1 and nuclear factor erythroid 2-related factor 2, and inhibition of mitogen-activated protein kinase signaling. Evidence suggests that topical resveratrol could be a valuable alternative not only for daily skin care, but also for the prevention and treatment of various cutaneous disorders. This review summarizes the benefits of resveratrol for cutaneous functions.
1. Introduction
In the traditional view, the skin serves as a protective barrier between the body and the external environment. Yet, more and more evidence suggests that cutaneous function extends far beyond mere protection. In fact, cutaneous function regulates a wide spectrum of cutaneous and systemic functions. Compromised epidermal function has been linked to the development of a variety of cutaneous and extracutaneous disorders. For example, disruption of epidermal permeability barrier not only provokes the release and production of proinflammatory cytokines [1–3], but also induces infiltration and activation of inflammatory cells in the skin [4–7], suggesting that defective epidermal permeability barrier predisposes to the development of inflammatory dermatoses [8–11]. But barrier disruption also stimulates barrier homeostasis responses, including epidermal proliferation and lipid synthesis [12, 13]. Moreover, defects in epidermal permeability barrier allow the penetration of microbial pathogens into the skin [14–16].
Importantly, recent studies showed that the epidermal dysfunction-induced elevations in cutaneous cytokines lead to increased levels of proinflammatory cytokines not only in the skin, but also in circulation, supporting not only a pathogenic role for epidermal function in cutaneous and extracutaneous inflammation, but also suggesting a link between cutaneous function and inflammation-associated systemic disorders [17, 18]. In addition to epidermal permeability barrier homeostasis, other epidermal functions, such as pH and stratum corneum hydration, also regulate cutaneous functions [19–22]. Accordingly, improvements in epidermal function could benefit multiple cutaneous and extracutaneous functions [17, 23–25].
Because of the importance of cutaneous function, much recent attention has focused on the identification of active ingredients that could lead to the development of products that improve cutaneous function. In comparison with synthetic chemicals, natural ingredients are generally considered to be cheaper and more widely available, but still exhibit comparable benefits [26–28]. Among natural ingredients, bioflavonoids, including hesperidin, apigenin, epigallocatechin gallate, and resveratrol, exhibit a wide spectrum of biological activities, including antioxidative, anti-inflammatory, anticancer, antiaging, and UV protection, while improving cutaneous functions. In this article, we review the current in frontier about the regulatory role of resveratrol in cutaneous functions.
2. Properties and Sources of Resveratrol
Resveratrol (3,5,4′-trihydroxy-trans-stilbene) is a stilbenoid, belonging to a group of phytoalexins, with a molecular weight of 228.247 g/M and a melting point of 254°C (Figure 1 from http://www.streamstime.com), first identified in 1939 [29]. Resveratrol dissolves well in both ethanol and acetone (50 mg/ml), but barely dissolves in water (3 mg/1000 ml). It can be synthesized via Wittig reaction of protected 3,5-dihydroxybenzaldehyde and methylenetriphenylphosphorane, using reagent aldehyde : sodium hydride : CH3P(C6H5)Br at a ratio of 1 : 4 : 4 [30, 31] (Figure 2). Yields of resveratrol can be increased by using unprotected 3,5-dihydroxybenzaldehyde and aldehyde : sodium hydride : CH3P(C6H5)Br at a ratio of 1 : 4 : 4 [32]. In contrast, the biosynthesis of resveratrol in plants can begin with acetyl-CoA, tyrosine, or phenylalanine in response to external stressors, such as insults from insects, microbial pathogens, animals, or adverse weather conditions [33–35] (Figure 3). Resveratrol exists as both trans and cis isomers in plants. Trans-resveratrol can be isomerized to the cis form by UV irradiation [36].
Figure 1.
Structure of resveratrol.
Figure 2.
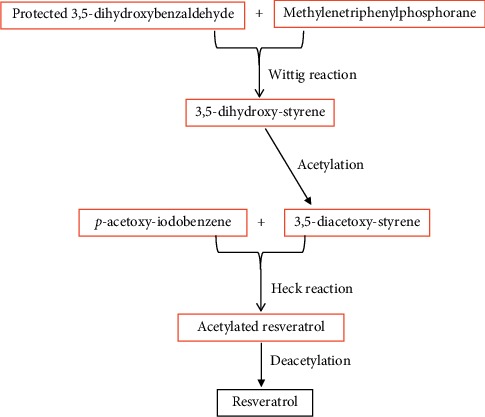
Synthesis of resveratrol [30].
Figure 3.
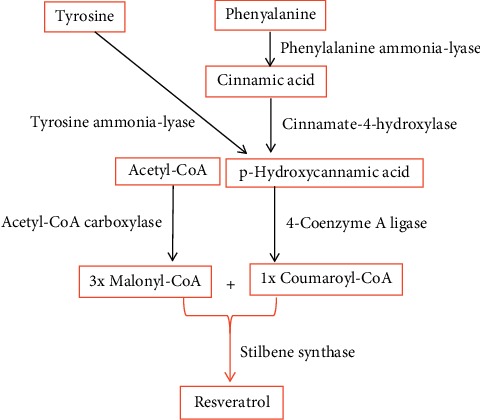
Biosynthesis of natural resveratrol.
Stilbene synthase is a key enzyme in the synthesis of resveratrol in the plants. Studies demonstrated that both expression levels of stilbene synthase and resveratrol content are regulated by a transcription factor, Myb14, which binds to Box L5 motif, leading to elevated stilbene synthase gene expression [37]. A number of other factors also regulate stilbene synthase expression and resveratrol production. For example, leaves contain the highest level, while the shoot tips have a relatively low level of stilbene synthase protein and mRNA in 1-year-old potted grapevines of Vitis vinifera L. cv. Cabernet Sauvignon [38]. Moreover, during grape development, expression levels of stilbene synthase mRNA increase continuously in the grape skin until they became ripe [39]. Likewise, old leaves of Cabernet Sauvignon exhibit higher levels of stilbene synthase mRNA than young leaves [39].
Infections of grapes can also change the expression of stilbene synthase mRNA. Dai et al. reported that incubation of cabernet sauvignon leaves with powdery mildew fungal spores for 24 hours significantly increased expression levels of stilbene synthase mRNA [39]. Additionally, irradiation of grape leaves with UVC not only increased the expression levels of both stilbene synthase protein and mRNA, but also increased resveratrol content, starting as early as 8 hours after irradiation [38]. In grape skin, UVB (312 nm) irradiation mainly increased expression levels of stilbene synthase mRNA in unripe grapes [40]. Wilting grapes at 28°C increased trans-resveratrol content, peaking at day 60, followed by a decline [40]. Incubation of grape leaves with 50 mM calcium chloride (CaCl2) for 24 hours induced greater than 2-fold increase in trans-resveratrol content in comparison with vehicle control [41]. Irradiation with UVC could further increase trans-resveratrol content in comparison with CaCl2 alone [41]. UVC irradiation primarily increased trans-resveratrol content in young grapes [42]. Seasonal changes in resveratrol content were also observed in grape roots, with a higher content (103–123 mg/kg dry weight) in the months of May and August to October, and a lower content (61–93 mg/kg dry weight) in the remaining months of the year [43]. Fungicides and methyl jasmonate treatments can markedly reduce resveratrol content in grapes and strawberry, respectively [44, 45]. Taken together, environmental conditions and the stages of plant development affect resveratrol synthesis and content.
Among plants, resveratrol is most abundant in the skin of red grapes [46]. But some other foods and beverages also contain fair amounts of resveratrol [34, 47–56]. Moreover, it appears that the age of the beverage determines its resveratrol content. For instance, 15-year-old champagne wine only contains 1 mg/L trans-resveratrol while the same wine, aged for 8 years, contains 45 mg/L trans-resveratrol [53]. Resveratrol content also varies with different parts of plants. The highest content of resveratrol was found in the stem phloems, but the lowest in the leaves in 1-year-old potted grapevines [38]. Likewise, resveratrol content in the grape skin is higher than that in the leaves and seeds for some grapes [57, 58]. But some grape seeds contain a higher content of resveratrol than do the skins [58]. Moreover, resveratrol content varies greatly with cultivars [33, 57–62]. For example, resveratrol content in small white Spanish peanuts is over 20-fold higher than that in White's runner peanuts [33]. Similarly, resveratrol content in the skin of rootstock grapes is over 100 times that in table grapes [57]. Moreover, resveratrol content in the skin of Berlandier Resseguier grapes is over 170 times of that in the skin of Dog Ridge grapes (366 vs. 20 μg/g fresh weight) [57], the same as in tomatoes, in which the total resveratrol content is 18.4 μg/g fresh weight in Micro Toms, and 0.34 and 0.38 μg/g fresh weight, respectively, in Plum Toms and Ugly Ripes [63]. Thus, resveratrol content varies with the age of beverage, plant parts, and the cultivars.
Other factors, such as cultivation sites and nitrogen availability, can also influence the resveratrol content in plants [35, 62, 64, 65]. For example, the trans-resveratrol content in V. vinifera grape cane grown in Yantai is lower than that grown in Yangling, China (755 vs. 938 mg/kg fresh weight) [62]. Likewise, knotweed from Zunyi, China, contains 4.27 ± 0.09 mg/g of resveratrol. But knotweed from Yunnan only contains 2.06 ± 0.03 mg/g of resveratrol, while resveratrol was undetectable in knotweed grown in Dunstaffnage, Canada [65]. Similarly, the total resveratrol content is as high as 3.11 mg/L in merlot red wines made from grapes cultivated in Nagano, Japan, while the same wines only contain 0.88 mg/L of total resveratrol, if the grapes are cultivated in Iwate [35].
Variation of resveratrol content in plants is likely due to different environment/weather conditions [40–43, 64] and chemical compositions of soils [45, 66]. In addition, processing methods of foods and beverages can affect resveratrol content. Studies have shown that neither raw nor dry-roasted peanuts contain detectable trans-resveratrol [67, 68], but trans-resveratrol content was ≈1.6 mg/100g in boiled peanuts [67] and 10 to 50 μg/100 g in peanut butter [68]. Moreover, preparation and/or processing methods can influence resveratrol content in beverages. For example, ultrasonic cleaning of grapes could increase resveratrol by as much as over 50% in comparison with that without ultrasonic cleaning [69]. Similarly, heating highbush Michigan blueberries at 190°C for 18 min reduced trans-resveratrol by over 40% as compared to raw blueberries [55]. Moreover, methanol extraction of grapes yields resveratrol as much as two times of acetone extraction [70]. In summary, plants and fruits contain a fair amount of resveratrol. But environmental conditions, developmental stage, and the parts of plants or fruits largely determine resveratrol content, while processing methods and storage conditions can affect the resveratrol content of foods and beverages. The content of resveratrol in certain foods, plants, and beverages is summarized in Supplemental .
3. Benefits of Resveratrol for Cutaneous Functions
The benefits of resveratrol for health have been demonstrated in multiple systems and/or organs, including cardiovascular system, diabetes, and immune as well as the neural system [71–76]. A large body of evidence, summarized here, has also indicated the benefits of resveratrol for the skin (Table 1) [77–126].
Table 1.
Benefits of resveratrol for cutaneous functions.
| Models | Treatments | Benefits | Mechanisms | Ref. |
|---|---|---|---|---|
| Keratinocyte proliferation/differentiation | ||||
| Keratinocytes | Cells cultured with 0.25–100 μM resveratrol for 24–72 hr | ↓Proliferation | ND | [77] |
| Cells cultured with 20 or 40 μM resveratrol for 24 hr | ↓Proliferation | ↑SIRT1 | [78] | |
| Cells cultured with 50 μM resveratrol for 12 hr | ↓Proliferation | ND | [79] | |
| Cells cultured with 25–100 μM resveratrol for 24 hr | ↓Proliferation | ND | [80, 81] | |
| Cells treated with 0.197 μM resveratrol for 2 weeks | ↓Proliferation | ND | [82] | |
| Cells cultured with 3 μM resveratrol until 3 days postconfluence | ↓Differentiation | ↑SIRT1 | [83] | |
| Cells cultured with 100 μM resveratrol for 24 hr | ↓Proliferation ↓Differentiation |
↓Protein kinase D | [84] | |
|
| ||||
| Anti-UV irradiation | ||||
| Keratinocytes | Immediately after irradiation with UVA (5 J/cm2), cells were treated with 0.01–0.1 mM resveratrol for 24 hr | ↑Cell viability ↓MDA content |
↑SOD and GSH-Px expression | [85] |
| Either before or after irradiation with UVA (2.796 J/cm2), cells were treated with 2.5 and 5.0 mg/l resveratrol, respectively | ↑Cell viability ↑SOD and GST |
↑NRF2 in nuclear translocation ↓Keap1 |
[86] | |
| Cells first treated with 10 μM resveratrol for 1 hr, followed by UVB irradiation (30 mJ/cm2) | ↑Cell viability ↓Apoptosis |
↑Activation of SIRT1 | [87] | |
| Cells first treated with 2% of resveratrol for 2 hr, followed by UVB irradiation (5–100 mJ/cm2) | ↑Cell viability | ND | [88] | |
| Cells treated with 5–25 μM resveratrol for 24 hr, followed by irradiation with UVB (40 mJ/cm2) | ↓NF-κB content and activity | ↑IκBα ↓IKKα |
[89] | |
| Cells treated with 5–10 μM pterostilbene, analog of resveratrol, for 24 hr, followed by irradiation with UVB (30 mJ/cm2). | ↑Cell viability ↓ROS ↓DNA damage |
↑Nrf2 | [90] | |
| Cells treated with 50 μM resveratrol 30 min prior to irradiation with 1 J/cm2 UVA +0.1 J/cm2 UVB | ↓IL-6, MCP-1, and TNF-α mRNA | ↑ARH | [91] | |
| Cells treated with 25 or 100 μM resveratrol for 2 or 24 hr, followed by irradiation with UVB (10, 20, 40, or 100 mJ/cm2) | ↑ROS ↓Autophagy |
↑ERK activation ↑Bax/Bcl2 ratio |
[92] | |
| Cells first irradiated with 1 J/cm2 UVA +0.1 J/cm2 UVB, followed by treatment with 10 μM resveratrol. | ↓CYP1A1, CYP1B1, IL-1β, IL-6, and COX2 mRNA levels | ↓Peroxide content | [93] | |
| Dermal fibroblasts | Immediately after UVB irradiation (144 mJ/cm2), fibroblasts were treated with 10 or 100 μg/L of resveratrol-enriched rice extract at various concentrations for 24–72 hr | ↑Cell viability ↓ROS ↓MMP1, p53, Bax, TNF-α, IL-6, iNOS, and COX2 ↑PIP1 and TGF-β protein |
ND | [94] |
| Reconstructed human skin | Reconstructed human skin was treated with 1% of resveratrol-enriched rice extract for 24 hr, followed by irradiation with UVB (100 mJ/cm2) | ↓MMP1 ↑PIP1, type I procollagen, and collagen fibers |
ND | [94] |
| Mice | Mice were treated topically with 25 μM resveratrol in 200 μl acetone 30 min prior to irradiation with 180 mJ/cm2 UVB | ↓Ear weight and edema ↓Inflammatory infiltrate ↓ODC activity and COX2 activity ↓Lipid peroxidation |
ND | [95] |
| Mice were treated topically with 10 μM resveratrol in 200 μl acetone 30 min prior to irradiation with 180 mJ/cm2 UVB | ↑Cell proliferation ↓COX2 and ODC expression |
↓Survivin | [96] | |
| Mice were treated topically with 0.48% resveratrol 20 min prior to irradiation with 360 mJ/cm2 UVB | ↓Skin edema in mice treated with resveratrol either before or after UVB irradiation ↓Epidermal thickness in mice treated with resveratrol before UVB irradiation |
↑NRF2 | [97] | |
| Mice were treated topically with 0.48% resveratrol 20 min prior to irradiation with 180 mJ/cm2 UVB, 3 irradiation/week for a total of 30 weeks | ↓Lipid, DNA, and protein peroxidation ↑Activity and expression of antioxidant enzymes |
[97] | ||
|
| ||||
| Antioxidant defense | ||||
| Keratinocytes | Cells cultured with 20 and 60 μM resveratrol for 24 hr | ↑GST activity | ↑NRF2 expression and activation | [98] |
| Cells pretreated with 10 or 20 μM resveratrol for 16 hr | ↑NQO1 and GSH-Px mRNA ↑GSH protein synthesis |
↑NRF2 activation | [99] | |
| Cells treated with both 0.3–3 mM sodium nitroprusside and 1–30 μM resveratrol for 24 hr | ↑Cell viability ↓Caspase 3 and 9 activity |
↓IL-8, NOS3, and NADPH dehydrogenase mRNA ↑GSH-Px mRNA |
[100] | |
| Cells pretreated with 25 or 100 μM resveratrol 24 hr prior to addition of 200, 400, or 800 μM H2O2 and cells were harvested 48 hr postaddition of H2O2 | ↓ROS | ND | [92] | |
| Cells pretreated with 140 μM resveratrol for 1 h, then 500 μM H2O2 was added, and incubated for additional 2–16 h | ↓DNA damage and HSP70 expression | ND | [101] | |
| Cells treated with 10 μM resveratrol for 24 hr, and then 100 μM H2O2 was added and incubated for additional 30 min | ↓ROS ↓MDA |
ND | [102] | |
| Cells pretreated with 0.5–10 μM resveratrol for 24 hr, followed by removal of media, and then exposed to cigarette smoking for 50 min | ↑Scavenger receptor class B type I protein and mRNA ↓4-Hydroxynonenal adducts |
ND | [103] | |
| Cells pretreated with 10 μM resveratrol for 24 hr, followed by removal of media, and then exposed to cigarette smoking for 50 min | ↓ROS and carbonyl groups | ↑Methionine sulfoxide reductase A mRNA ↓Transient receptor potential cation channel subfamily A member 1 mRNA and protein |
[104] | |
| Cells pretreated with 0.5 μM resveratrol for 3 hr, followed by addition of 0.1–20 μM arsenic and incubation for additional 20 hr | ↓Arsenic-induced increase in metabolic activity and expression of DNA polymerase beta ↑Arsenic-induced reduction in Y419 phosphorylation and Src protein |
ND | [105] | |
| Reconstructed human skin | Keratinocytes were pretreated with 20 or 100 μM resveratrol for 16 hr, followed by removal of media, and then exposed to 800 μM cumen hydroperoxide for 8 hr | ↑GSH expression ↓Apoptosis |
↑NRF2 activation | [99] |
| Mice | Mice were treated topically with 16 μM resveratrol, and four hours later, skin samples were collected | ↑GST activity and content | ↑NRF2 activation | [98] |
| Mice were treated topically with 8 or 16 μM resveratrol, and twenty-four hours later, skin samples were collected | ↑Glucuronosyltransferase and NADPH:quinone oxidoreductase activity | ND | [106] | |
| Humans | Stratum corneum was collected with tape strip 24 hr after single application of resveratrol at a dose of 537 μg/cm2 on the ventral forearm | ↓Production of free radical | ND | [107] |
|
| ||||
| Anticancer | ||||
| Melanoma cell line | Cells were treated with 20–40 μg/ml resveratrol for up to 5 days | ↓Proliferation ↓Apoptosis and necrosis |
↑Cells in S phase arrest ↓Cells in G1 and G2/M phase |
[108] |
| A431 human skin carcinoma cells | A431 cells were treated with 20–100 mg/L resveratrol for 24 hr | ↑Apoptosis ↓Proliferation |
↑Activation of MAPK pathway | [81] |
| A431 cells were treated with 20, 50, and 100 μM resveratrol for up to 72 hr | ↓DNA synthesis and proliferation ↓Cells in G2/M phase ↓Cell cycle regulatory proteins |
↓DNA-binding activity of AP-1 ↓ERK1/2 signaling pathway |
[109] | |
| Human squamous cell carcinoma cell lines | HSC2 cells were treated with both resveratrol and benzoxazinotropone at various concentrations for 48 hr | Resveratrol and benzoxazinotropone synergistically inhibited proliferation | ND | [110] |
| Head and neck squamous cell carcinoma cells | Head and neck squamous cell carcinoma cells were treated with 15 and 50 μM resveratrol for up to 72 hr | ↓Proliferation ↑Apoptosis |
↑H2AX ser-139 phosphorylation | [111] |
| Mice | Mice with squamous cell tumor graft were gavaged orally with 10 and 50 mg/kg body weight of resveratrol for 30 days | ↓Tumor weight and volume per mouse | ||
|
| ||||
| Anti-inflammation | ||||
| Keratinocytes | Cells treated with 20 ng/ml TNF-α for 6 hr 10 μM, followed by incubation with resveratrol for additional 16 hr | ↓IL-6 and MCP-1 | ↓Phosphorylation of IκBα | [102] |
| Cells treated with 7.5 μg/mL lipopolysaccharide for 12 hr, followed by incubation with 10–50 μM resveratrol for additional 12 hr | ↑Proliferation ↑Apoptosis ↓IL-6, IL-8, and TNF-α mRNA and protein |
↑miR-17 expression | [112] | |
| Cells treated with 50 μM resveratrol 1 hr prior to addition of 2.5 μg/mL lipopolysaccharide | ↓IL-6, IL-8, MCP-1, and COX2 mRNA | ↓EGFR-ERK signaling pathway | [91] | |
| Cells pretreated with 25 and 50 μM resveratrol for 30 min, followed by incubation with 25 μg/ml cetuximab or 2 μM gefitinib for additional 3 hr | ↓CCL2 and CXCL10 mRNA and protein | ↓Interferon regulatory factor 1 and phosphorylated STAT1 | [113] | |
| Cells pretreated with 50 μM resveratrol for 1 hr, followed by incubation with for 10 ng/ml IFN-γ and TNF-α 24 hr | ↓IL-6 | ND | [114] | |
| Cells pretreated with 44 μM resveratrol for 24 hr, followed by exposure to heat stress for 40 min | ↓IL-6, IL-8, and TNF-α | ND | [115] | |
| Mast cells | RBL-2H3 mast cells were pretreated with 1–25 μM resveratrol for 2 hr, followed by exposure to 200 ng/ml dinitrophenyl-human serum albumin | ↓IL-3, IL-4, IL-13, and TNF-α ↓Fc epsilon receptor I expression |
↓P38-MAPK, ERK1/2, JNK | [116] |
| Reconstructed human skin | 3D skin was treated 10 ng/ml IFN-γ and TNF-α twice a week, followed by treatment with 1% resveratrol thrice weekly for 2 weeks | ↓IL-6 | ND | [114] |
| Mice | BALB/c mouse ears were treated topically with 10 mM resveratrol 2 hr prior to DNFB challenge | ↓Ear thickness ↓CD3-positive cells ↓ICAM-1, CCL2, and CXCL10 expression |
↓Interferon regulatory factor 1 and phosphorylated STAT1 | [113] |
| Following induction of allergic contact dermatitis, NC/Nga mice were treated topically with 2.5% resveratrol or resveratrol-enriched rice extract twice weekly for 5 weeks | ↓Epidermal thickness ↓Dermatitis score ↓Serum IgE ↓TEWL ↑Skin hydration |
ND | [114] | |
| BALB/c mice were orally treated with resveratrol at a dose of 10 mg/kg body weight 1 hr prior to intravenous challenge with 200 μg dinitrophenyl-human serum albumin | ↓IL-4 and TNF-α ↓CD11b-positive cells |
↓Tyk2-STAT1 activation | [116] | |
| Atopic dermatitis-like lesions were induced by topical applications of DNFB to the back of BALB/c mice for 5 weeks, followed by orally given resveratrol at a daily dose of 30 mg/kg body weight for 1 week | ↓Dermatitis scores ↓Epidermal thickness ↓Cytokine mRNA |
ND | [117] | |
| Atopic dermatitis-like lesions were induced by topical applications of dermatophagoides farinae to the back of NC/Nga mice for 2 weeks, followed by orally given resveratrol at a daily dose of 20 mg/kg body weight for 2 weeks | ↓Dermatitis scores ↓Cytokine mRNA and protein |
↓High mobility group box 1 expression | [118] | |
| Psoriasis-like skin lesions were induced by topical applications of imiquimod to the back of BALB/c mice, which were orally given resveratrol at a daily dose of 400 mg/kg body weight, for 7 days. | ↓Erythema and scale scores ↓Skin thickness ↓Cytokine mRNA |
ND | [119] | |
|
| ||||
| Accelerating wound healing | ||||
| Rats | Rats were fed with resveratrol at a daily dose of 0.5 mg/kg body weight 7 days prior to operation and continued throughout the whole experiment period | ↑Collagen deposition ↑Neovascularization ↑Fibroblast maturation |
ND | [120] |
| Following induction of full-thickness skin wound, wound was treated topically with 225 μL of 50 μM once daily for 17 days | ↑Epithelialization ↓Wound size ↑Collagen deposition ↑Vascularization |
↑AMPK pathway and SIRT1 | [121] | |
| Mice | Immediately after wound, a wound dressing containing 0.04% resveratrol was applied to full-thickness skin wound for 10 days | ↓Wound size ↑Collagen fibers ↓Inflammation |
ND | [122] |
| Placing scaffolds containing 5% resveratrol on the wound for 7 days | ↓Wound size | ↑Expression of thioredoxin-1, heme oxygenase-1, and VEGF | [123] | |
| Diabetic models | 0.5% resveratrol ointment was applied to wound area in diabetic rats once daily for 21 days | ↓Wound size | ↑Activity of antioxidant enzymes | [124] |
| 10 μM resveratrol was applied to the cutaneous wound area in diabetic mice, and wound healing was assessed 7 days later | ↓Wound size ↑Endothelial cell proliferation |
Sirt1 activation | [125] | |
| Fourteen days after topical application of resveratrol (0.1 mg/ml) to wound area in diabetic rats once, wound healing was assessed | No benefit | [126] | ||
Abbreviations: ND, not determined; AQP3, aquaporin 3; SOD, superoxide dismutase; MDA, malondialdehyde; GSH-Px, glutathione peroxidase; GST, glutathione S-transferase; GSH, reduced glutathione; NQO, NAD(P)H:quinone oxidoreductase; ROS: reactive oxygen species; CYP1A1, cytochrome P540 family 1 subfamily A member 1; IKKα, ΙκB kinase α; CYP1B1, cytochrome P540 family 1 subfamily B member 1; COX, cyclooxygenase; ODC, ornithine decarboxylase; NRF2, nuclear factor erythroid 2-related factor 2; ERK: extracellular signal-regulated kinase; MAPK, mitogen-activated protein kinase; JNK, c-Jun NH2-terminal kinase; TPA, 12-O-tetradecanoyl13-phorbol acetate; DNFB, 2,4-dinitro-1-fluorobenzene; NOS, nitric oxide synthase; VEGF, vascular endothelial growth factor; AMPK, adenosine monophosphate‐activated protein kinase.
3.1. Keratinocyte Proliferation and Differentiation
Both keratinocyte proliferation and differentiation are required for the epidermis to reach its ultimate goal of the formation of the stratum corneum, an essential structure for epidermal permeability barrier. Several studies have demonstrated that resveratrol inhibits keratinocyte proliferation while stimulating differentiation. Resveratrol at a concentration as low as 2 μM markedly inhibits DNA synthesis in keratinocyte cultures [77]. Inhibitory effect of resveratrol on DNA synthesis occurred as early as 24 hr after addition to cultures, with an IC50 range of 2–8 μM [77]. Incubation of keratinocytes with resveratrol, at concentrations as low as 0.25 μM for 72 hr, could induce a dose-dependent reduction in the number of living cells. Wu et al. reported that 20 μM resveratrol inhibited keratinocyte proliferation by over 80%, using the 5-bromo-2-deoxyuridine assay [78]. With long-term treatments (e.g., two weeks), even 0.197 μM resveratrol was sufficed to inhibit keratinocyte proliferation by 80% [82].
The activities of keratinocyte proliferation and differentiation are coordinated in an inverse manner, while terminal differentiation is crucial for the formation of permeability barrier. In contrast to proliferation, resveratrol stimulates keratinocyte differentiation. For example, treatment of keratinocytes with 3 μM resveratrol until 3 days after postconfluence increased involucrin expression by 1.5-fold [83]. Moreover, formation of cornified envelopes, critical structures for epidermal permeability barrier, requires transglutaminase [127]. Treatment of keratinocytes with resveratrol induced a dose-dependent increase in transglutaminase activity, while simultaneously inhibiting DNA synthesis, with a half-maximal concentration of 35 μM. Finally, resveratrol and 1, 25(OH)2D3 synergistically increased transglutaminase activity [84]. Thus, resveratrol inhibits keratinocyte proliferation, while stimulating differentiation.
3.2. Anti-UV Irradiation
While suberythemogenic doses of UVB irradiation instead enhance epidermal function, including improvements in epidermal permeability barrier function, stimulation of epidermal lipid synthesis and keratinocyte differentiation, and antimicrobial defense [128], excessive exposure to UV irradiation can damage the skin, causing sunburns, skin cancers, and photoaging. Both in vivo and in vitro studies have demonstrated that resveratrol protected the skin from UV irradiation. For example, addition of 0.1 μM resveratrol to the culture medium immediately postirradiation provided 100% protection against UVA-induced reductions in keratinocyte proliferation and an increase in malondialdehyde (MDA) content. In parallel, both superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px) contents in UVA-irradiated keratinocytes were normalized by treatment with resveratrol [85]. Either prior to or post-UVA irradiation, treatment of keratinocytes with resveratrol decreased lipid peroxidation. But only pretreatment, not posttreatment with resveratrol, increased SOD and glutathione S-transferase (GST) protein levels [86]. In addition to UVA irradiation, resveratrol can also protect keratinocytes from UVB-induced damages. For example, treatment of keratinocytes with resveratrol 1 hr prior to UVB irradiation decreased apoptosis rates by over 50% [87]. Moreover, treatment of keratinocytes with resveratrol prior to UV irradiation decreased the expression levels of proinflammatory cytokines, such as IL-6, IL-8, and TNF-α, by ≈50% from untreated keratinocytes [91]. Likewise, addition of 10 μM resveratrol immediately after UVA and UVB irradiation also lowered the expression levels of IL-1-beta and IL-6 [93]. In addition, treatment of dermal fibroblasts with resveratrol after UVB irradiation decreased the contents of (i) reactive oxygen species, (ii) TNF-α and IL-6, (iii) inducible nitric oxide synthase (iNOS), and (iv) expression levels of matrix metallopeptidase I, while increasing procollagen 1 and elastin [94]. It appeared that pretreatment of keratinocytes with low concentration of resveratrol did not inhibit UV irradiation-induced inflammation [93].
The protective effects of resveratrol against UV irradiation have also been demonstrated in vivo. Afag et al. reported that topical application of 25 μM resveratrol 30 min prior to UVB irradiation (180 mJ/cm2) decreased skin thickness and punch weight of ear by over 50 and 81%, respectively, in comparison with vehicle-treated controls [95]. Moreover, cutaneous inflammatory infiltration and ornithine decarboxylase protein levels decreased significantly in mouse skin treated with topical resveratrol prior to UVB irradiation [96]. Yet, topical applications of resveratrol post-UVB irradiation decreased whole skin thickness, but not epidermal thickness [97]. Some studies showed that resveratrol and UV irradiation synergistically enhanced expression levels of IL-8 mRNA and protein, and DNA damage in keratinocytes as compared to UV irradiation alone [129, 130]. These discrepant results could be attributable to variations, including the doses of resveratrol and UV irradiation, treatment timing, and/or the status of growth and differentiation in these cells. Thus, further studies are needed to clarify the effects of resveratrol on cutaneous function.
3.3. Antioxidant Defense
A number of factors, including psychological stress, cigarette smoke, air pollution, and UV irradiation, can cause oxidative stress, contributing to the development of aging and a line of disorders such as dermatoses, inflammation, cardiovascular diseases, cancer, and neurodegenerative diseases [131–139]. Accordingly, antioxidants have been utilized to prevent and treat certain disorders [140–142]. Among the antioxidants, benefits of resveratrol have also been studied extensively both in vitro and in vivo. Phase II enzymes, including GSH-Px, quinone dehydrogenase (NQO), GST, SOD, and GSH, are responsible for antioxidation in living organisms. Soeur et al. reported that 10 μM resveratrol induced an over 2-fold increase in expression levels of mRNA for GSH-Px and NQO1, along with over 1-fold increase in glutathione synthesis, in keratinocytes, in addition to an over 7-fold increase in expression levels of mRNA for GSH-Px and NQO1 in reconstructed human skin [99]. Resveratrol can also protect keratinocytes from damages caused by oxidative stressors, including nitric oxide (NO), H2O2, cigarette smoke, and arsenic [92, 101–105]. Surprisingly, topical resveratrol did not mitigate UVB irradiation-induced reductions in both activity and expression levels of antioxidant enzymes, including catalase, SOD, and GSH-Px, in a mouse model of chronic UVB irradiation [97]. NO is a free radical required for maintenance of normal function, but excessive NO can induce oxidative stress, leading to apoptosis [136]. Topical applications of human skin with NO donor (nitrite) caused cutaneous inflammation and keratinocyte apoptosis [137]. Likewise, 3 mM sodium nitroprusside induced ≈70% inhibition of keratinocyte proliferation [100]. Treatment of keratinocytes with the combination of 30 μM resveratrol and 3 mM sodium nitroprusside reduced caspase 3 and 9 activity as well as apoptotic rate by 80% from cultures treated with sodium nitroprusside alone. H2O2 is commonly used as stressor to induce oxidative stress. Following the treatment of keratinocytes with 400 μM H2O2, reactive oxygen species (ROS) content increased 1.5-fold over the controls [92]. Resveratrol at concentrations of 25 μM and 100 μM lowered ROS contents by 25% and 40%, respectively, in comparison with cells treated with H2O2 alone. Finally, the protective effects of resveratrol against oxidative stress have also been demonstrated in keratinocytes treated with either arsenic or cigarette smoke [103–105].
The skin, an interface between the body and external environment, is vulnerable to environmental insults, including oxidative stress, leading to acceleration of skin aging and the development of a variety of skin disorders [138, 139, 143]. Therefore, enhancement of antioxidant defense of the skin could help manage various cutaneous dermatoses. Krajka-Kuźniak et al. showed that a single topical treatment of mouse skin with 16 μM resveratrol for 4 hr increased GST activity by 63% and GST content by 22% over controls [98]. Similarly, a greater than 1-fold increase in GST activity was observed in mouse epidermis 24 hr after topical application of 16 μM resveratrol [106]. Likewise, production of free radicals was reduced in human stratum corneum following topical treatment with resveratrol for 24 hr [107]. Collectively, resveratrol exhibits antioxidant properties both in vitro and in vivo.
3.4. Anticancer
Oxidative stress has been considered as a major contributor to the development of skin cancers, such as melanoma [144–146]. Accordingly, antioxidants, including resveratrol, have been used to prevent and treat skin cancer [147–149], although exacerbation and increased risk of cancers have been reported [150, 151]. Melanoma is the most serious type of skin cancer, with mortality rates of 3.1/100,000 in the US [152] and 1.4–1.9/100,000 in Germany [153]. Antimelanoma benefits of resveratrol have been demonstrated both in vivo and in vitro. Studies in melanoma cell cultures showed that resveratrol inhibited proliferation of melanoma cells, with a concentration of 7 μg/ml inducing a 50% inhibition of cell growth [108]. It seems that melanoma cells are more sensitive to resveratrol than keratinocytes, which instead required a concentration of 20 μg/ml to cause 50% inhibition of cell growth. Moreover, resveratrol dose-dependently increased both apoptotic and necrotic cells after 24 hr treatment of two melanoma cell lines [108]. In addition, resveratrol also inhibited the growth of squamous cell carcinoma cells (A431 cell line) [81]. Resveratrol-induced apoptosis and inhibition of cell proliferation were also observed in human head and neck squamous cell carcinoma cell lines, e.g., FaDu, Cal27, and Det562 cell lines [111]. Moreover, rates of DNA synthesis were reduced by 78% following the treatment of carcinoma cells with 25 μM resveratrol for 72 hr [109]. In vivo studies showed that oral administration of resveratrol resulted in a dose- and time-dependent inhibition of carcinoma cell growth, with over 50% reductions in both tumor volume and weight per mouse following 30-day treatment with resveratrol at a dose of 50 mg/kg body weight [111]. Taken together, resveratrol demonstrates potential anticancer activity both in vivo and in vitro.
3.5. Anti-Inflammation
Inflammatory dermatoses prove the most common clinical problems in dermatology. Because of the severe side effects of immune modulators, such as glucocorticoids and tacrolimus, safe and effective alternatives are highly demanded. Studies have shown that resveratrol is safe in both animals and humans [154, 155], but it can effectively inhibit inflammation both in vitro and in vivo. For example, incubation of keratinocytes with 50 μM resveratrol for 3 hr lowered the expression levels of mRNA for IL-6, IL-8, TNF-α, and macrophage inflammatory protein 1 (MIF-1) by over 50% [91]. While production of proinflammatory cytokines markedly increased in keratinocytes exposed to TNF-α, lipopolysaccharide, interferon γ (IFN-γ), and UV irradiation [91, 102, 112–114], addition of 50 μM resveratrol 1 hr prior to lipopolysaccharide treatment dramatically decreased mRNA levels (>40% reduction) for MIF-1, IL-6, and cyclooxygenase 2 in comparison with keratinocytes treated with lipopolysaccharide alone [91]. Yet, resveratrol also dose-dependently stimulated IL-8 production in keratinocyte cultures [91], suggesting that resveratrol differentially regulates cytokine production.
Although there is still little or no evidence that resveratrol can inhibit cutaneous inflammation in humans, anti-inflammatory benefits of resveratrol have been well demonstrated in murine models of inflammatory dermatoses. For example, in acute allergic contact dermatitis model, pretreatment of mouse ears with resveratrol reduced the density of CD3+ cells by≈90% in parallel with significant reduction of ear thickness and expression levels of intercellular adhesion molecule 1 (ICAM-1), C-X-C motif chemokine ligand 10 (CXCL10), C-C motif chemokine ligand 2 (CCL2), and IFN-κ in the epidermis [113], suggesting preventive benefits of topical resveratrol in acute allergic contact dermatitis. Moreover, the therapeutic effects of topical resveratrol have also been demonstrated in an atopic dermatitis-like disease model. Kang et al. showed that topical applications of either 2.5% resveratrol or resveratrol-enriched rice extract for five weeks markedly reduced dermatitis score and serum IgE levels in a dermatitis model induced by topical dinitrochlorobenzene [114]. Likewise, both intravenously and orally given resveratrol significantly alleviated dermatitis score and decreased cytokine expression [117, 118].
Psoriasis is another common, inflammatory skin disorder. Kjaer et al. showed that oral administrations of trans-resveratrol at a daily dose of 400 mg/kg body weight induced over 50% reductions in both erythema and scale scores, along with 15% decrease in skinfold thickness of the back skin, in an imiquimod-induced psoriasis-like mouse model [119]. In addition, topical resveratrol also decreased the expression levels of IL-17a and IL-19 mRNA by ≈60% in comparison with mice treated with imiquimod alone [119]. Together, these results show that both topical and systemic administration of resveratrol can mitigate cutaneous inflammation.
3.6. Cutaneous Wounding
Cutaneous wound healing involves the proliferation of both fibroblast and keratinocytes, as well as collagen deposition. A number of observations suggested that, in murine models of full-thickness wound, resveratrol stimulates cell proliferation and collagen, leading to acceleration in cutaneous wound healing. Application of wound dressings containing resveratrol to full-thickness skin wounds induced marked reductions in wound areas in comparison with the controls [122]. Similar acceleration in wound closure occurs following the placement of scaffolds containing resveratrol over the wound [123]. This evidence indicates that topical resveratrol accelerates wound healing in normal mice.
Management of slow wound healing in diabetics has been a substantial challenge. Yes, studies have shown that either topical or systemic administrations of resveratrol can improve cutaneous wound healing in animal model of diabetes. Moreover, the efficacy of topical resveratrol ointment for wound healing was superior to that of topical β-sitosterol, conventional wound healing product [124]. Huang et al. also reported that a single application of resveratrol accelerated cutaneous wound healing in diabetic mice [125]. However, other studies have shown little or no benefit of resveratrol in wound healing following a single application in diabetic rats [126]. Thus, the potential benefits of resveratrol for wound healing in diabetic models still need to be validated.
3.7. Others
Studies in both humans and murine models reveal that resveratrol also regulates other cutaneous functions, including skin aging, melanogenesis, and antimicrobial defense. In human keratinocyte cultures, resveratrol reduced 90% reduction in expression levels of beta-galactosidase, a biomarker of senescence, in an aging model induced by oxidative stress [156, 157]. Clinical trials have also demonstrated the antiaging properties of resveratrol. A study in 50 humans with clinical signs of aging showed that oral fruit extracts that contain resveratrol markedly improved multiple aging-associated parameters, including increased stratum corneum hydration and skin elasticity, decreased skin roughness and wrinkle depth, as well as reductions in the intensity of pigmented solar lentigines [158]. In parallel, levels of plasma derivatives of ROS dramatically declined, while skin ferric-reducing ability increased. In addition, topical applications of resveratrol-containing products also improved aging-associated signs, such as skin wrinkles, stratum corneum hydration, and pigmentation, in aged humans [159]. But in one clinical trial in 30 subjects, oral supplement of product containing trans-resveratrol did not appreciably improve skin aging, despite reductions in cutaneous MDA content and elevations in SOD content [160]. These discrepant results suggest that additional trials are still needed to determine whether resveratrol benefits skin aging.
Other studies suggest that resveratrol exhibits antimicrobial properties. Cathelicidin antimicrobial peptides (CAMP) are a family of polypeptides, produced by keratinocytes, macrophages, and polymorphonuclear leukocytes, that display antibacterial, antifungal, and antiviral activities. Park et al. reported that incubation of keratinocytes with resveratrol for 24 hours increased the expression level of CAMP mRNA by over 4-fold [161]. But resveratrol may also directly inhibit microbial growth because incubation with resveratrol induced time- and dose-dependent reductions in Propionibacterium acnes colony-forming units, possibly due to disruption of the bacterial membrane [162]. Studies have shown that resveratrol exhibits several bactericidal and bacteriostatic activity against several pathogens, including S. pyogenes, S. aureus, C. glabrata, and C. albicans, with minimum inhibitory concentrations as low as 1.25 mg/ml [163]. Moreover, topical applications of 25% resveratrol cream markedly decreased lesion scores for herpes simplex infection, with an efficacy comparable to 5% acyclovir ointment, in a mouse model of herpes simplex infections [164]. Similar results were also obtained with topical applications of oxyresveratrol in mice infected by herpes simplex virus [165, 166]. Furthermore, studies also suggest benefits of resveratrol for keloids. For example, resveratrol induced apoptosis of fibroblasts from keloids, in parallel with reductions in the expression levels of mRNA for collagen 1 and procollagen 3, while increasing expression of SIRT1, suggesting a potential application of resveratrol for the treatment of keloids and hypertrophic scars [167–169]. Other studies have demonstrated that topical resveratrol improves epidermal permeability barrier function and stratum corneum hydration in sodium dodecyl sulfate-damaged human skin [170]. Additionally, both in vitro and in vivo studies have shown that resveratrol reduces skin pigmentation via inhibition of tyrosinase activity, cytokine production, and melanocytic microphthalmia-associated transcription factor (MITF) expression [171]. Yet, all of these putative benefits of resveratrol for cutaneous function still lack sufficient clinical validation. Therefore, well-designed clinical trials are still required before resveratrol can be widely utilized in clinical settings.
4. Mechanisms of Action
Evidence of resveratrol for multiple cutaneous functions has been well demonstrated, but the underlying mechanisms whereby resveratrol acts remain unclear. A line of evidence suggests that the actions of resveratrol could be via multiple mechanisms such as upregulation of nuclear factor erythroid 2-related factor 2 (Nrf2), activation of sirtuin 1 (SIRT1), and mitogen-activated protein kinase (MAPK) signaling pathway, depending on which function is regulated. The major putative mechanisms by which resveratrol regulates cutaneous function are illustrated in Figure 4.
Figure 4.
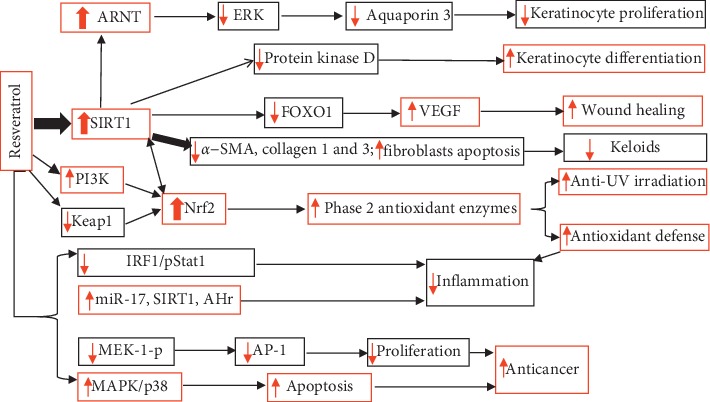
Schematic diagram of the mechanisms by which resveratrol regulates cutaneous functions. α-SMA: α-smooth muscle actin.
4.1. Keratinocyte Proliferation and Differentiation
Keratinocyte proliferation and differentiation, which are inversely regulated, are both required to form the stratum corneum, the outmost layers of the skin, providing multiple cutaneous protective functions because resveratrol can stimulate keratinocyte differentiation while inhibiting proliferation, resulting in acceleration of epidermal maturation. The inhibitory effects of resveratrol on keratinocyte proliferation occur via two mechanisms: (i) activation/upregulation of SIRT1 and (ii) inhibition of protein kinase D. In keratinocyte cultures, resveratrol upregulated expression of SIRT1, leading to elevation in aryl hydrocarbon receptor nuclear translocator (ARNT), resulting in downregulation of aquaporin 3, and consequently inhibiting cell proliferation [78]. Lee et al. showed that resveratrol increased the expression level and deacetylase activity of SIRT1, resulting in apoptosis [172]. Other studies suggest that resveratrol inhibits DNA synthesis, while increasing transglutaminase activity via inhibition of protein kinase D activity [84]. Moreover, activation of SIRT1 by resveratrol could also increase keratinocyte differentiation [83]. Activation of SIRT1 by resveratrol is likely via enhancement of the binding of specific substrates to SIRT1 [173]. Thus, resveratrol could inhibit keratinocyte proliferation and stimulate differentiation via both activation of SIRT1 and/or inhibition of protein kinase D.
4.2. Protection from UV Irradiation and Antioxidant Defense
Although the precise mechanisms by which resveratrol protects the skin against UV irradiation and oxidative stress are unclear, a handful of evidence points to a central role of Nrf2. This transcription factor regulates phase 2 antioxidant enzymes, which protect against UV irradiation- and other oxidative stress-induced damage to the skin. Nrf2 deficiency accelerated UV irradiation-induced photoaging and inflammation [174, 175], while conversely activation of Nrf2 protects against UV irradiation-induced apoptosis and inflammation [176, 177]. Normally, Nrf2 together with Kelch ECH associating protein 1 (Keap1) forms a complex, which is degraded by the ubiquitin-proteasome system [178]. Upon oxidative stress (including UV irradiation-induced oxidative stress), Nrf2 is released from Nrf2/Keap1 complex and translocates into the nucleus, where Nrf2 binds to antioxidant response element (ARE) in a heterodimeric complex, consequently leading to increased production of phase 2 antioxidant enzymes. While UV irradiation can increase the production of reactive oxygen species and oxidative products [179, 180], resveratrol can attenuate UV-induced oxidative stress via upregulation and/or activation of Nrf2. For example, treatment of keratinocytes with resveratrol either before or after UVA irradiation induced >50% increase in Nrf2 content, while increasing content of Nrf2 in the nuclear fraction [86, 90]. Similarly, treatment of either normal mice or oxidative-stressed keratinocytes with resveratrol increases Nrf2 expression and activation [98, 99], leading to increased expression of phase 2 antioxidant enzymes and reductions in reactive oxygen species [85, 90, 100], ultimately protecting/alleviating cell damage induced by UV irradiation or other oxidative stressors.
With regard to how resveratrol upregulates Nrf2 expression and activity, at least three mechanisms probably are operative. One mechanism involves upregulation of SIRT1 expression. Resveratrol is a SIRT1 activator [83]. Treatment of adipocytes with resveratrol significantly increased expression levels of SIRT1 mRNA [181]. Upregulation of SIRT1 expressions, in turn, increases expression levels of Nrf2 and phase 2 antioxidant enzymes, while silencing SIRT1 with siRNA decreases Nrf2 protein as well as activity of ARE promotor [182]. Moreover, upregulation and activation of Nrf2 expression by SIRT1 were also observed [183]. The second mechanism comprises direct upregulation of Nrf2 expression because studies have shown that resveratrol increases Nrf2 expression in kidney, heart, and lung tissues [184, 185]. The third mechanism is direct downregulation of Keap1 expression. Treatment of keratinocytes with resveratrol either before or post-UVA irradiation lowers expression levels of Keap1 protein [86]. Resveratrol-induced reduction of Keap1 expression was also observed in the kidney and lung tissues of obese and asthmatic rats [185]. Reductions in Keap1 expression can slow Nrf2 degradation, resulting in an increase in Nrf2 expression. Other studies also showed that resveratrol stimulated Nrf2 expression and nuclear translocation without changing the expression levels of Keap1 [98, 184]. Interestingly, Nrf2 and SIRT1 can positively coregulate each other. For example, treatments of either renal tubular cells or glomerular mesangial cells with resveratrol parallelly increased Nrf2 and SIRT1 expression. Knockdown of Nrf2 with siRNA decreases the expression levels of SIRT1, and vice versa [184, 186]. Therefore, resveratrol can sequentially or separately upregulate SIRT1 and Nrf2, while downregulation of Keap1 expression, resulting in increased expression of phase 2 antioxidant enzymes, which in turn protect the skin from UV irradiation- and oxidative stress-induced damage. Finally, one study showed that inhibition of phosphatidylinositol-3-kinase prevented the activation of Nrf2 induced by pterostilbene, a resveratrol analog, suggesting that resveratrol can also activate Nrf2 via activation of phosphatidylinositol-3-kinase [90].
Still other mechanisms could also account for the actions of anti-UV irradiation and antioxidative stress. For example, pretreatment of keratinocytes with resveratrol almost completely prevents the activation of NFκB induced by UVB irradiation [89], suggesting that resveratrol-induced inhibition of NFκB activation could contribute to its anti-UV irradiation properties. Resveratrol-induced upregulation of heat-shock protein 27 (HSP27) and downregulation of caspase 3 could also contribute to its anti-UV irradiation property [187]. Thus, resveratrol protects skin against UV irradiation and oxidative stress via multiple mechanisms.
4.3. Anticancer Activity
Both in vitro and in vivo studies have shown that resveratrol also inhibits proliferation, while stimulating apoptosis of cancer cells via several mechanisms. First, resveratrol induces apoptosis and phosphorylation of MAPK/ERK and MAPK/p38 in addition to increasing expression levels of caspase 3 and p53, while conversely, inhibition of p38 abolished its apoptotic effects [81, 188]. It appears that resveratrol-induced phosphorylation of p53 and apoptosis is mediated by c-Jun NH2-terminal kinases because knockdown of c-Jun NH2-terminal kinase genes prevented both phosphorylation of p53 and apoptosis induced by resveratrol [189]. Therefore, resveratrol-induced activation of the MAPK/p38 signaling pathway likely accounts, at least in part, for its anticancer effects. Regarding antiproliferation of cancer cells, resveratrol inhibits expression of MEK1-P and ERK1/2-P, leading to reductions in cyclin D1 and cyclin-dependent kinase 6 expression, resulting in cell cycle at rest [109]. Moreover, resveratrol also decreased c-Jun levels and reduced DNA-binding and transcriptional activity of activator protein-1, which is required for initiation of DNA synthesis [190]. Other studies showed that inhibition of NF-κB, cyclooxygenase 2, phosphatidylinositol-3-kinase, and P450 isoenzyme CYP1A1 and induction of caspases 3 and 9 also could contribute to anticancer effects of resveratrol [191–196]. Thus, resveratrol-induced reductions in expression levels of MEK1-P and ERK1/2-P and decreased activator protein-1 activity could contribute to its inhibition of cancer cell proliferation.
4.4. Anti-Inflammatory Activity
The anti-inflammatory effects of resveratrol have been demonstrated in various in vivo and in vitro models, but the mechanisms of the actions of resveratrol are often unclear, depending on the inflammatory models employed in the studies. One possible mechanism is inhibition of NF-κB signaling pathways. Activation of NF-κB can upregulate transcription of cytokines while IκB inhibits NF-κB activity [102, 197]. Hence, degradation of phosphorylated IκB would increase NF-κB activity. Resveratrol-containing mixture inhibited IκB phosphorylation and decreased NF-κB, resulting in reductions in cytokine production in keratinocytes stimulated by TNF-α [102]. Another study suggests that inhibition of cytokine production by resveratrol seems linked to upregulation of miR-17 expression in keratinocytes stimulated with lipopolysaccharide because inhibition of miR-17 overcame the inhibitory effects of resveratrol on inflammation [112]. But one study showed that resveratrol increases IL-8 production in keratinocytes stimulated with a combination of TNF-α and IFNγ via upregulation of aryl hydrocarbon receptor expression [91]. Inhibition of allergic contact dermatitis by resveratrol could be attributable to the downregulation of interferon regulatory factor 1/STAT1 signaling pathway and inhibition of phosphorylation of MAPK/p38 and/or phospholipase Cγ [113, 116, 198]. Moreover, resveratrol inhibited proliferation and differentiation of CD+ T cells and proliferation of Th17 T cells via upregulation of phosphorylated MAPK and downregulation of phosphorylated mammalian target of rapamycin (p-mTOR) in Jurkat cells [199]. Furthermore, resveratrol-induced inhibition of TNF-α-induced cytokine production in fibroblasts is via activation of SIRT1 because knockdown of SIRT1 abolishes the inhibitory effect resveratrol on inflammation [200]. Thus, resveratrol can inhibit cutaneous inflammation via a variety of mechanisms, including inhibition of NF-κB, MAPK/p38, phospholipase Cγ, and p-mTOR, upregulation of miR-17, and activation of SIRT1.
4.5. Wound Healing
Cutaneous wound healing is a complex process that can be accelerated by resveratrol via stimulation of neovascularization, keratinocyte differentiation, permeability barrier maturation, and antimicrobial activity. One study showed that resveratrol accelerates cutaneous wound healing and vascularization in aged rats through upregulation of SIRT1 and adenosine monophosphate-activated protein kinase (AMPK) pathway [121]. The role of SIRT1 signaling in vascularization has also been demonstrated in cutaneous wounds of diabetic mice. Topical applications of resveratrol to the wounded area of diabetic mice stimulated proliferation and inhibited apoptosis of endothelial cells, leading to accelerated wound healing, while either SIRT1 inhibitor or knockout of SIRT1 abolished the benefits of resveratrol in wound healing [125]. SIRT1-mediated benefits of resveratrol in diabetic wound healing can also be attributable to protection of endothelial cells from oxidative stress [201]. In addition, studies in mice indicate that resveratrol accelerates cutaneous wound healing by upregulation of vascular endothelial growth factor mediated by activation of at least two antioxidant enzymes (thioredoxin-1 and heme oxygenase-1) [123]. Because wound infections are the major cause of delayed wound healing [202], the antimicrobial properties of resveratrol could be another mechanism whereby wound healing is accelerated. Lastly, the ultimate goal of cutaneous wound healing is the formation of intact permeability barrier, which requires both lipid production and keratinocyte differentiation. Thus, resveratrol could also accelerate cutaneous wound healing through its well-known ability to stimulate keratinocyte differentiation and lipid production [83, 84, 161]. Collectively, the resveratrol-induced acceleration of cutaneous wound healing can be attributable to activation of SIRT1 and AMPK signaling, antioxidative stress, and enhanced formation of epidermal permeability barrier.
4.6. Others
The mechanisms whereby resveratrol induces apoptosis and inhibition of fibroblasts include inhibition of hypoxia-inducible factor 1, in which activation stimulates fibroblast proliferation while inhibiting apoptosis [203], downregulation of transforming growth factor β1 [169], miR-17 [204], as well as expression levels of mRNA for collagen 1 and procollagen 3 [167], whereas resveratrol-induced upregulation of antimicrobial peptides is via enhancing expression of sphingosine-1-phosphate, leading to activation of N-FκB-C/EBPα signaling pathway [161].
Resveratrol inhibits melanogenesis by at least four different mechanisms: (1) in human melanocyte cultures, resveratrol inhibited tyrosinase synthesis and activity along with accelerated transport of newly synthesized tyrosinase to proteasomal complex, without dramatic alterations in mRNA levels of either melanocytic microphthalmia-associated transcription factor (MIFT) or tyrosinase [205]; (2) in melan-A cells, inhibition of melanogenesis by resveratrol is via induction of autophagy, leading to reduction in α melanocyte-stimulating hormone levels [206]. The latter stimulates melanin production and release via activation of melanocortin-1 receptor. Deletion of autophagy-related genes could prevent resveratrol-induced reduction in melanogenesis; (3) in human melanocyte cultures, resveratrol activated c-Jun N-terminal kinase, resulting downregulation of MITF expression [207]; and (4) anti-inflammatory effects of resveratrol can be an additional mechanism contributing to decreasing pigmentation [171]. The antioxidant properties of resveratrol largely account for its antiaging effects [208].
5. Additional Points
Both in vitro and in vivo studies showed that resveratrol regulates multiple cutaneous functions, including UV protection, anti-inflammation, antioxidant defense, acceleration of wound healing, antimicrobe, anticancer, and inhibition of melanogenesis. Upregulation of SIRT1 largely accounts for the mechanisms of resveratrol action although various other mechanisms are also involved. Thus, resveratrol benefits multiple cutaneous functions through a variety of mechanisms. While clinical data have revealed the benefits of resveratrol for extracutaneous systems/organs, including improvements in bone density, osteoarthritis, renal function, and diabetes, evidence is still insufficient to conclude its benefits for cutaneous functions in clinical settings. Therefore, proper clinical trials are still required to validate the clinical significance of the utility of resveratrol in the skin.
Acknowledgments
This work was supported, in part, by the NIH grant AR061106, administered by the Northern California Institute for Research and Education, with resources from the Research Service, Department of Veterans Affairs.
Conflicts of Interest
All authors declare no conflicts of interest.
Authors' Contributions
MQM originated the concept and drafted the manuscript; SW and JZ performed literature search, drafted the manuscript, and contributed equally to this work; BY and PME critically reviewed the manuscript. All authors approved the final version.
Supplementary Materials
Supplemental Table 1: content of resveratrol in some plants and foods/beverages.
References
- 1.Wood L. C., Elias P. M., Calhoun C., Tsai J. C., Grunfeld C., Feingold K. R. Barrier disruption stimulates interleukin-1α expression and release from a pre-formed pool in murine epidermis. Journal of Investigative Dermatology. 1996;106(3):397–403. doi: 10.1111/1523-1747.ep12343392. [DOI] [PubMed] [Google Scholar]
- 2.Wood L. C., Jackson S. M., Elias P. M., Grunfeld C., Feingold K. R. Cutaneous barrier perturbation stimulates cytokine production in the epidermis of mice. Journal of Clinical Investigation. 1992;90(2):482–487. doi: 10.1172/jci115884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Tsai J. C., Feingold K. R., Crumrine D., Wood L. C., Grunfeld C., Elias P. M. Permeability barrier disruption alters the localization and expression of TNF?/protein in the epidermis. Archives of Dermatological Research. 1994;286(5):242–248. doi: 10.1007/bf00387595. [DOI] [PubMed] [Google Scholar]
- 4.Lin T.-K., Man M.-Q., Santiago J.-L., et al. Topical antihistamines display potent anti-inflammatory activity linked in part to enhanced permeability barrier function. Journal of Investigative Dermatology. 2013;133(2):469–478. doi: 10.1038/jid.2012.335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Proksch E., Brasch J., Sterry W. Integrity of the permeability barrier regulates epidermal Langerhans cell density. British Journal of Dermatology. 1996;134(4):630–638. doi: 10.1046/j.1365-2133.1996.66828.x. [DOI] [PubMed] [Google Scholar]
- 6.Proksch E., Brasch J. Influence of epidermal permeability barrier disruption and Langerhans’ cell density on allergic contact dermatitis. Acta Dermato-Venereologica. 1997;77(2):102–104. doi: 10.2340/0001555577102104. [DOI] [PubMed] [Google Scholar]
- 7.Katoh N., Hirano S., Kishimoto S., Yasuno H. Acute cutaneous barrier perturbation induces maturation of Langerhans’ cells in hairless mice. Acta Dermato-Venereologica. 1997;77(77):365–369. doi: 10.2340/0001555577365369. [DOI] [PubMed] [Google Scholar]
- 8.Elias P., Wood L. C., Feingold K. R. Epidermal pathogenesis of inflammatory dermatoses ∗1, ∗2. American Journal of Contact Dermatitis. 1999;10(3):119–126. doi: 10.1016/s1046-199x(99)90054-4. [DOI] [PubMed] [Google Scholar]
- 9.Nishijima T., Tokura Y., Imokawa G., Seo N., Furukawa F., Takigawa M. Altered permeability and disordered cutaneous immunoregulatory function in mice with acute barrier disruption. Journal of Investigative Dermatology. 1997;109(2):175–182. doi: 10.1111/1523-1747.ep12319282. [DOI] [PubMed] [Google Scholar]
- 10.Scharschmidt T. C., Man M.-Q., Hatano Y., et al. Filaggrin deficiency confers a paracellular barrier abnormality that reduces inflammatory thresholds to irritants and haptens. Journal of Allergy and Clinical Immunology. 2009;124(3):496–506. doi: 10.1016/j.jaci.2009.06.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ye L., Lv C., Man G., Song S., Elias P. M., Man M.-Q. Abnormal epidermal barrier recovery in uninvolved skin supports the notion of an epidermal pathogenesis of psoriasis. Journal of Investigative Dermatology. 2014;134(11):2843–2846. doi: 10.1038/jid.2014.205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Proksch E., Feingold K. R., Man M. Q., Elias P. M. Barrier function regulates epidermal DNA synthesis. Journal of Clinical Investigation. 1991;87(5):1668–1673. doi: 10.1172/jci115183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Proksch E., Jensen J.-M., Elias P. M. Skin lipids and epidermal differentiation in atopic dermatitis. Clinics in Dermatology. 2003;21(2):134–144. doi: 10.1016/s0738-081x(02)00370-x. [DOI] [PubMed] [Google Scholar]
- 14.Wanke I., Skabytska Y., Kraft B., Peschel A., Biedermann T., Schittek B. Staphylococcus aureus skin colonization is promoted by barrier disruption and leads to local inflammation. Experimental Dermatology. 2013;22(2):153–155. doi: 10.1111/exd.12083. [DOI] [PubMed] [Google Scholar]
- 15.Jinnestål C. L., Belfrage E., Bäck O., Schmidtchen A., Sonesson A. Skin barrier impairment correlates with cutaneous Staphylococcus aureus colonization and sensitization to skin-associated microbial antigens in adult patients with atopic dermatitis. International Journal of Dermatology. 2014;53(1):27–33. doi: 10.1111/ijd.12198. [DOI] [PubMed] [Google Scholar]
- 16.Berents T. L., Carlsen K. C. L., Mowinckel P., et al. Skin barrier function and Staphylococcus aureus colonization in vestibulum nasi and fauces in healthy infants and infants with eczema: a population-based cohort study. PLoS One. 10(6) doi: 10.1371/journal.pone.0130145.e0130145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ye L., Mauro T. M., Dang E., et al. Topical applications of an emollient reduce circulating pro‐inflammatory cytokine levels in chronically aged humans: a pilot clinical study. Journal of the European Academy of Dermatology and Venereology. 2019;33(11):2197–2201. doi: 10.1111/jdv.15540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hu L., Mauro T. M., Dang E., et al. Epidermal dysfunction leads to an age-associated increase in levels of serum inflammatory cytokines. Journal of Investigative Dermatology. 2017;137(6):1277–1285. doi: 10.1016/j.jid.2017.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lee H.-J., Yoon N. Y., Lee N. R., Jung M., Kim D. H., Choi E. H. Topical acidic cream prevents the development of atopic dermatitis- and asthma-like lesions in murine model. Experimental Dermatology. 2014;23(10):736–741. doi: 10.1111/exd.12525. [DOI] [PubMed] [Google Scholar]
- 20.Wat M., Olicker A., Meyerson H., Nedorost S., Paller A. S., Cooper K. Topical hypochlorite and skin acidification improves erythroderma of omenn syndrome. Pediatrics. 2018;141(5):S408–S411. doi: 10.1542/peds.2017-1249. [DOI] [PubMed] [Google Scholar]
- 21.Lee H.-J., Lee N. R., Kim B.-K., et al. Acidification of stratum corneum prevents the progression from atopic dermatitis to respiratory allergy. Experimental Dermatology. 2017;26(1):66–72. doi: 10.1111/exd.13144. [DOI] [PubMed] [Google Scholar]
- 22.Choi M. Q., Elias P. M. Stratum corneum hydration regulates key epidermal function and serves as an indicator and contributor to other conditions. Journal of the European Academy of Dermatology and Venereology. 2019;33(1):15–16. doi: 10.1111/jdv.15374. [DOI] [PubMed] [Google Scholar]
- 23.Man M. Q., Ye L., Hu L., Jeong S., Elias P. M., Lv C. Improvements in epidermal function prevent relapse of psoriasis: a self-controlled study. Clinical and Experimental Dermatology. 2019;44(6):654–657. doi: 10.1111/ced.13888. [DOI] [PubMed] [Google Scholar]
- 24.Salam R. A., Darmstadt G. L., Bhutta Z. A. Effect of emollient therapy on clinical outcomes in preterm neonates in Pakistan: a randomised controlled trial. Archives of Disease in Childhood—Fetal and Neonatal Edition. 2015;100(3):F210–F215. doi: 10.1136/archdischild-2014-307157. [DOI] [PubMed] [Google Scholar]
- 25.Chamlin S. L., Kao J., Frieden I. J., et al. Ceramide-dominant barrier repair lipids alleviate childhood atopic dermatitis: changes in barrier function provide a sensitive indicator of disease activity. Journal of the American Academy of Dermatology. 2002;47(2):198–208. doi: 10.1067/mjd.2002.124617. [DOI] [PubMed] [Google Scholar]
- 26.Williams A. S., Estanqueiro M., Oliveira M. B., Sousa Lobo J. M. Main benefits and applicability of plant extracts in skin care products. Cosmetics. 2015;2(2):48–65. doi: 10.3390/cosmetics2020048. [DOI] [Google Scholar]
- 27.Hollinger J. C., Angra K., Halder R. M. Are natural ingredients effective in the management of hyperpigmentation? A systematic review. Journal of Clinical and Aesthetic Dermatology. 2018;11(2):28–37. [PMC free article] [PubMed] [Google Scholar]
- 28.Bowe W. P., Pugliese S. Cosmetic benefits of natural ingredients. Journal of Drugs in Dermatology. 2014;13(9):1021–1025. [PubMed] [Google Scholar]
- 29.Takaoka M. The phenolic substances of white hellebore (Veratrum grandiflorum hoes. Fil.) I. Nippon Kagaku Kaishi. 1939;60(11):1090–1100. doi: 10.1246/nikkashi1921.60.1090. [DOI] [Google Scholar]
- 30.Guiso M., Marra C., Farina A. A new efficient resveratrol synthesis. Tetrahedron Letters. 2002;43(4):597–598. doi: 10.1016/S0040-4039(01)02227-4. [DOI] [Google Scholar]
- 31.Lara-Ochoa F., Sandoval-Minero L. C., Espinosa-Pérez G. A new synthesis of resveratrol. Tetrahedron Letters. 2015;56(44):5977–5979. doi: 10.1016/j.tetlet.2015.09.005. [DOI] [Google Scholar]
- 32.Farina A., Ferranti C., Marra C. An improved synthesis of resveratrol. Natural Product Research. 2006;20(3):247–252. doi: 10.1080/14786410500059532. [DOI] [PubMed] [Google Scholar]
- 33.Sanders T. H., McMichael R. W., Jr., Hendrix K. W. Occurrence of resveratrol in edible peanuts. Journal of Agricultural and Food Chemistry. 2000;48(4):1243–1246. doi: 10.1021/jf990737b. [DOI] [PubMed] [Google Scholar]
- 34.Burns J., Yokota T., Ashihara H., Lean M. E. J., Crozier A. Plant foods and herbal sources of resveratrol. Journal of Agricultural and Food Chemistry. 2002;50(11):3337–3340. doi: 10.1021/jf0112973. [DOI] [PubMed] [Google Scholar]
- 35.Sato M., Suzuki Y., Okuda T., Yokotsuka K. Contents of resveratrol, piceid, and their isomers in commercially available wines made from grapes cultivated in Japan. Bioscience, Biotechnology, and Biochemistry. 1997;61(11):1800–1805. doi: 10.1271/bbb.61.1800. [DOI] [PubMed] [Google Scholar]
- 36.Nour V., Trandafir I., Muntean C. Ultraviolet irradiation of trans-resveratrol and HPLC determination of trans-resveratrol and cis-resveratrol in Romanian red wines. Journal of Chromatographic Science. 2012;50(10):920–927. doi: 10.1093/chromsci/bms091. [DOI] [PubMed] [Google Scholar]
- 37.Fang L., Hou Y., Wang L., Xin H., Wang N., Li S. Myb14, a direct activator of STS, is associated with resveratrol content variation in berry skin in two grape cultivars. Plant Cell Reports. 2014;33(10):1629–1640. doi: 10.1007/s00299-014-1642-3. [DOI] [PubMed] [Google Scholar]
- 38.Wang W., Tang K., Yang H.-R., et al. Distribution of resveratrol and stilbene synthase in young grape plants (Vitis vinifera L. cv. cabernet sauvignon) and the effect of UV-C on its accumulation. Plant Physiology and Biochemistry. 2010;48(2-3):142–152. doi: 10.1016/j.plaphy.2009.12.002. [DOI] [PubMed] [Google Scholar]
- 39.Huang R., Ge H., Howard S., Qiu W. Transcriptional expression of stilbene synthase genes are regulated developmentally and differentially in response to powdery mildew in norton and cabernet sauvignon grapevine. Plant Science. 2012;197:70–76. doi: 10.1016/j.plantsci.2012.09.004. [DOI] [PubMed] [Google Scholar]
- 40.Versari A., Parpinello G. P., Tornielli G. B., Ferrarini R., Giulivo C. Stilbene compounds and stilbene synthase expression during ripening, wilting, and UV treatment in grape cv. corvina. Journal of Agricultural and Food Chemistry. 2001;49(11):5531–5536. doi: 10.1021/jf010672o. [DOI] [PubMed] [Google Scholar]
- 41.Wang L., Ma L., Xi H., Duan W., Wang J., Li S. Individual and combined effects of CaCl2 and UV-C on the biosynthesis of resveratrols in grape leaves and berry skins. Journal of Agricultural and Food Chemistry. 2013;61(29):7135–7141. doi: 10.1021/jf401220m. [DOI] [PubMed] [Google Scholar]
- 42.Wang J.-F., Ma L., Xi H.-F., Wang L.-J., Li S.-H. Resveratrol synthesis under natural conditions and after UV-C irradiation in berry skin is associated with berry development stages in “Beihong” (V. vinifera×V. amurensis) Food Chemistry. 2015;168:430–438. doi: 10.1016/j.foodchem.2014.07.025. [DOI] [PubMed] [Google Scholar]
- 43.Ji M., Li Q., Ji H., Lou H. Investigation of the distribution and season regularity of resveratrol in Vitis amurensis via HPLC-DAD-MS/MS. Food Chemistry. 2014;142:61–65. doi: 10.1016/j.foodchem.2013.06.131. [DOI] [PubMed] [Google Scholar]
- 44.Magee J. B., Smith B. J., Rimando A. Resveratrol content of muscadine berries is affected by disease control spray program. Hortscience. 2002;37(2):358–361. doi: 10.21273/hortsci.37.2.358. [DOI] [Google Scholar]
- 45.Rimando S. Y., Chen C.-T., Wang C. Y., Chen P. Resveratrol content in strawberry fruit is affected by preharvest conditions. Journal of Agricultural and Food Chemistry. 2007;55(20):8269–8274. doi: 10.1021/jf071749x. [DOI] [PubMed] [Google Scholar]
- 46.Sears A. Resveratrol, an antiaging power house. 2019. http://www.holisticonline.com/herbal-Med/articles/hol_herb-resveratrol-antiaging.htm.
- 47.Nicoletti I., Bello C., De Rossi A., Corradini D. Identification and quantification of phenolic compounds in grapes by HPLC-PDA-ESI-MS on a semimicro separation scale. Journal of Agricultural and Food Chemistry. 2008;56(19):8801–8808. doi: 10.1021/jf801411m. [DOI] [PubMed] [Google Scholar]
- 48.López-Hernández J., Rodríguez-Bernaldo de Quirós A. Trans-stilbenes in commercial grape juices: quantification using HPLC approaches. International Journal of Molecular Sciences. 2016;17(10):p. E1769. doi: 10.3390/ijms17101769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wang K.-H., Lai Y.-H., Chang J.-C., Ko T.-F., Shyu S.-L., Chiou R. Y.-Y. Germination of peanut kernels to enhance resveratrol biosynthesis and prepare sprouts as a functional vegetable. Journal of Agricultural and Food Chemistry. 2005;53(2):242–246. doi: 10.1021/jf048804b. [DOI] [PubMed] [Google Scholar]
- 50.Vitrac X., Bornet A., Vanderlinde R., et al. Determination of stilbenes (δ-viniferin, trans-astringin, trans-piceid, cis-andtrans-resveratrol, ε-viniferin) in Brazilian wines. Journal of Agricultural and Food Chemistry. 2005;53(14):5664–5669. doi: 10.1021/jf050122g. [DOI] [PubMed] [Google Scholar]
- 51.Teissédre F., Coccioli F., Jasionowska R., Merolle M., Terracciano A. Analysis of some stilbenes in Italian wines by liquid chromatography/tandem mass spectrometry. Rapid Communications in Mass Spectrometry. 2007;21(18):2955–2964. doi: 10.1002/rcm.3174. [DOI] [PubMed] [Google Scholar]
- 52.Ribeiro de Lima M. T., Waffo-Téguo P., Teissedre P. L., et al. Determination of stilbenes (trans-astringin, cis-andtrans-piceid, and cis-and trans-resveratrol) in Portuguese wines. Journal of Agricultural and Food Chemistry. 1999;47(7):2666–2670. doi: 10.1021/jf9900884. [DOI] [PubMed] [Google Scholar]
- 53.Mérillon P., Chaudruc D., Robillard B., et al. Determination of the trans-resveratrol content of champagne wines by reversed-phase HPLC. OENO One. 2006;40(2):117–119. doi: 10.20870/oeno-one.2006.40.2.876. [DOI] [Google Scholar]
- 54.Duteurtre M., Chung I.-M., Rajakumar G., et al. Evaluation of polyphenolic compounds and pharmacological activities in hairy root cultures of Ligularia fischeri turcz. F. spiciformis (nakai) Molecules. 2019;24(8):p. 1586. doi: 10.3390/molecules24081586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Lyons M. M., Yu C., Toma R. B., et al. Resveratrol in raw and baked blueberries and bilberries. Journal of Agricultural and Food Chemistry. 2003;51(20):5867–5870. doi: 10.1021/jf034150f. [DOI] [PubMed] [Google Scholar]
- 56.Sales J. M., Resurreccion A. V. A. Resveratrol in peanuts. Critical Reviews in Food Science and Nutrition. 2014;54(6):734–770. doi: 10.1080/10408398.2011.606928. [DOI] [PubMed] [Google Scholar]
- 57.Wang L., Xu M., Liu C., et al. Resveratrols in grape berry skins and leaves in vitis germplasm. PLoS One. 2013;8(4) doi: 10.1371/journal.pone.0061642.e61642 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Fan X., Wu B., Wang L., Li S. Extractable amounts oftrans-resveratrol in seed and berry skin in vitis evaluated at the germplasm level. Journal of Agricultural and Food Chemistry. 2006;54(23):8804–8811. doi: 10.1021/jf061722y. [DOI] [PubMed] [Google Scholar]
- 59.Limmongkon A., Janhom P., Amthong A., et al. Antioxidant activity, total phenolic, and resveratrol content in five cultivars of peanut sprouts. Asian Pacific Journal of Tropical Biomedicine. 2017;7(4):332–338. doi: 10.1016/j.apjtb.2017.01.002. [DOI] [Google Scholar]
- 60.Surangkul J. M., Harmon R., Weston L. A., Bessis R., Breuil A. C., Jeandet P. Resveratrol content of two California table grape cultivars. Vitis. 2001;40(1):43–44. [Google Scholar]
- 61.Liu C., Wang L., Wang J., et al. Resveratrols in vitis berry skins and leaves: their extraction and analysis by HPLC. Food Chemistry. 2013;136(2):643–649. doi: 10.1016/j.foodchem.2012.08.017. [DOI] [PubMed] [Google Scholar]
- 62.Li A., Fang Y., Li X., et al. Occurrence and estimation of trans-resveratrol in one-year-old canes from seven major Chinese grape producing regions. Molecules. 2011;16(4):2846–2861. doi: 10.3390/molecules16042846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ragab A. S., Van Fleet J., Jankowski B., Park J.-H., Bobzin S. C. Detection and quantitation of resveratrol in tomato fruit (Lycopersicon esculentum Mill.) Journal of Agricultural and Food Chemistry. 2006;54(19):7175–7179. doi: 10.1021/jf0609633. [DOI] [PubMed] [Google Scholar]
- 64.Tříska J., Vrchotová N., Balík J., Soural I., Sotolář R. Variability in the content of trans-resveratrol, trans-ε-viniferin and R2-viniferin in grape cane of seven Vitis vinifera L. varieties during a three-year study. Molecules. 2017;22(6):p. E928. doi: 10.3390/molecules22060928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Chen H., Tuck T., Ji X., et al. Quality assessment of Japanese knotweed (Fallopia japonica) grown on Prince Edward Island as a source of resveratrol. Journal of Agricultural and Food Chemistry. 2013;61(26):6383–6392. doi: 10.1021/jf4019239. [DOI] [PubMed] [Google Scholar]
- 66.Kovarova M., Bartunkova K., Frantik T., Koblihova H., Prchalova K., Vosatka M. Factors influencing the production of stilbenes by the knotweed, Reynoutria xbohemica. BMC Plant Biology. 2010;10(1):p. 19. doi: 10.1186/1471-2229-10-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Chukwumah Y., Walker L., Vogler B., Verghese M. Changes in the phytochemical composition and profile of raw, boiled, and roasted peanuts. Journal of Agricultural and Food Chemistry. 2007;55(22):9266–9273. doi: 10.1021/jf071877l. [DOI] [PubMed] [Google Scholar]
- 68.Sobolev V. S., Cole R. J. Trans-resveratrol content in commercial peanuts and peanut products. Journal of Agricultural and Food Chemistry. 1999;47(4):1435–1439. doi: 10.1021/jf9809885. [DOI] [PubMed] [Google Scholar]
- 69.Hasan M. M., Yun H.-K., Kwak E.-J., Baek K.-H. Preparation of resveratrol-enriched grape juice from ultrasonication treated grape fruits. Ultrasonics Sonochemistry. 2014;21(2):729–734. doi: 10.1016/j.ultsonch.2013.08.008. [DOI] [PubMed] [Google Scholar]
- 70.Soural I., Vrchotová N., Tříska J., et al. Various extraction methods for obtaining stilbenes from grape cane of Vitis vinifera L. Molecules. 2015;20(4):6093–6112. doi: 10.3390/molecules20046093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Sung M. M., Byrne N. J., Robertson I. M., et al. Resveratrol improves exercise performance and skeletal muscle oxidative capacity in heart failure. American Journal of Physiology-Heart and Circulatory Physiology. 2017;312(4):H842–H853. doi: 10.1152/ajpheart.00455.2016. [DOI] [PubMed] [Google Scholar]
- 72.Riba A., Deres L., Sumegi B., Toth K., Szabados E., Halmosi R. Cardioprotective effect of resveratrol in a postinfarction heart failure model. Oxidative Medicine and Cellular Longevity. 2017;2017:10. doi: 10.1155/2017/6819281.6819281 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Öztürk E., Arslan A. K. K., Yerer M. B., Bishayee A. Resveratrol and diabetes: a critical review of clinical studies. Biomedicine & Pharmacotherapy. 2017;95:230–234. doi: 10.1016/j.biopha.2017.08.070. [DOI] [PubMed] [Google Scholar]
- 74.Malaguarnera L. Influence of resveratrol on the immune response. Nutrients. 2019;11(5):p. 946. doi: 10.3390/nu11050946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Xie Y. K., Zhou X., Yuan H. T., et al. Resveratrol reduces brain injury after subarachnoid hemorrhage by inhibiting oxidative stress and endoplasmic reticulum stress. Neural Regeneration Research. 2019;14(10):1734–1742. doi: 10.4103/1673-5374.257529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Chiang M.-C., Nicol C. J., Cheng Y.-C. Resveratrol activation of AMPK-dependent pathways is neuroprotective in human neural stem cells against amyloid-beta-induced inflammation and oxidative stress. Neurochemistry International. 2018;115:1–10. doi: 10.1016/j.neuint.2017.10.002. [DOI] [PubMed] [Google Scholar]
- 77.Holian O., Walter R. J. Resveratrol inhibits the proliferation of normal human keratinocytes in vitro. Journal of Cellular Biochemistry. 2001;81(S36):55–62. doi: 10.1002/jcb.1085. [DOI] [PubMed] [Google Scholar]
- 78.Wu Z., Uchi H., Morino-Koga S., Shi W., Furue M. Resveratrol inhibition of human keratinocyte proliferation via SIRT1/ARNT/ERK dependent downregulation of aquaporin 3. Journal of Dermatological Science. 2014;75(1):16–23. doi: 10.1016/j.jdermsci.2014.03.004. [DOI] [PubMed] [Google Scholar]
- 79.Pastore S., Lulli D., Maurelli R., Dellambra E., De Luca C., Korkina L. G. Resveratrol induces long-lasting IL-8 expression and peculiar EGFR activation/distribution in human keratinocytes: mechanisms and implications for skin administration. PLoS One. 2013;8(3) doi: 10.1371/journal.pone.0059632.e59632 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Redondo P., Jimenez E., Perez A., García-Foncillas J. N-acetylcysteine downregulates vascular endothelial growth factor production by human keratinocytes in vitro. Archives of Dermatological Research. 2000;292(12):621–628. doi: 10.1007/s004030000187. [DOI] [PubMed] [Google Scholar]
- 81.Zhang X., Liu X., Kang S., Liu C., Hao Y. Resveratrol enhances the effects of ALA-PDT on skin squamous cells A431 through p38/MAPK signaling pathway. Cancer Biomarkers. 2018;21(4):797–803. doi: 10.3233/cbm-170495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Elmore E., Siddiqui S., Navidi M., Steele V. E., Redpath J. L. Correlation of in vitro chemopreventive efficacy data from the human epidermal cell assay with animal efficacy data and clinical trial plasma levels. Journal of Cellular Biochemistry. 2005;95(3):571–588. doi: 10.1002/jcb.20426. [DOI] [PubMed] [Google Scholar]
- 83.Blander G., Bhimavarapu A., Mammone T., et al. SIRT1 promotes differentiation of normal human keratinocytes. Journal of Investigative Dermatology. 2009;129(1):41–49. doi: 10.1038/jid.2008.179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Arun S. N., Xie D., Dodd M. E., Zhong X., Bollag W. B. The potential use of protein kinase D inhibitors for prevention/treatment of epidermal tumors. Journal of Dermatological Science. 2010;60(1):29–39. doi: 10.1016/j.jdermsci.2010.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Chen M. L., Li J., Xiao W. R., et al. Protective effect of resveratrol against oxidative damage of UVA irradiated HaCaT cells. Zhong nan da xue xue bao. Yi xue ban = Journal of Central South University. Medical sciences. 2006;31(31):635–639. [PubMed] [Google Scholar]
- 86.Liu Y., Chan F., Sun H., et al. Resveratrol protects human keratinocytes HaCaT cells from UVA-induced oxidative stress damage by downregulating keap1 expression. European Journal of Pharmacology. 2011;650(1):130–137. doi: 10.1016/j.ejphar.2010.10.009. [DOI] [PubMed] [Google Scholar]
- 87.Cao C., Lu S., Kivlin R., et al. SIRT1 confers protection against UVB- and H2O2-induced cell death via modulation of p53 and JNK in cultured skin keratinocytes. Journal of Cellular and Molecular Medicine. 2009;13(9B):3632–3643. doi: 10.1111/j.1582-4934.2008.00453.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Juškaitė V., Ramanauskienė K., Briedis V. Testing of resveratrol microemulsion photostability and protective effect against UV induced oxidative stress. Acta Pharmaceutica. 2017;67(2):247–256. doi: 10.1515/acph-2017-0018. [DOI] [PubMed] [Google Scholar]
- 89.Adhami V. M., Afaq F., Ahmad N. Suppression of ultraviolet B exposure-mediated activation of NF-κB in normal human keratinocytes by resveratrol. Neoplasia. 2003;5(1):74–82. doi: 10.1016/s1476-5586(03)80019-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Li H., Jiang N., Liang B., et al. Pterostilbene protects against UVB-induced photo-damage through a phosphatidylinositol-3-kinase-dependent Nrf2/ARE pathway in human keratinocytes. Redox Report. 2017;22(6):501–507. doi: 10.1080/13510002.2017.1329917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Potapovich A. I., Lulli D., Fidanza P., et al. Plant polyphenols differentially modulate inflammatory responses of human keratinocytes by interfering with activation of transcription factors NFκB and AhR and EGFR-ERK pathway. Toxicology and Applied Pharmacology. 2011;255(2):138–149. doi: 10.1016/j.taap.2011.06.007. [DOI] [PubMed] [Google Scholar]
- 92.Vitale N., Kisslinger A., Paladino S., et al. Resveratrol couples apoptosis with autophagy in UVB-irradiated HaCaT cells. PLoS One. 2013;8(11) doi: 10.1371/journal.pone.0080728.e80728 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Potapovich A. I., Kostyuk V. A., Kostyuk T. V., de Luca C., Korkina L. G. Effects of pre- and post-treatment with plant polyphenols on human keratinocyte responses to solar UV. Inflammation Research. 2013;62(8):773–780. doi: 10.1007/s00011-013-0634-z. [DOI] [PubMed] [Google Scholar]
- 94.Subedi L., Lee T. H., Wahedi H. M., Baek S. H., Kim S. Y. Resveratrol-enriched rice attenuates UVB-ROS-Induced skin aging via downregulation of inflammatory cascades. Oxidative Medicine and Cellular Longevity. 2017;2017:15. doi: 10.1155/2017/8379539.8379539 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Afaq F., Adhami V. M., Ahmad N. Prevention of short-term ultraviolet B radiation-mediated damages by resveratrol in SKH-1 hairless mice. Toxicology and Applied Pharmacology. 2003;186(1):28–37. doi: 10.1016/s0041-008x(02)00014-5. [DOI] [PubMed] [Google Scholar]
- 96.Aziz M. H., Afaq F., Ahmad N. Prevention of ultraviolet-B radiation damage by resveratrol in mouse skin is mediated via modulation in survivin. Photochemistry and Photobiology. 2005;81(1):25–31. doi: 10.1562/2004-08-13-ra-274.1. [DOI] [PubMed] [Google Scholar]
- 97.Sirerol J. A., Feddi F., Mena S., et al. Topical treatment with pterostilbene, a natural phytoalexin, effectively protects hairless mice against UVB radiation-induced skin damage and carcinogenesis. Free Radical Biology and Medicine. 2015;85:1–11. doi: 10.1016/j.freeradbiomed.2015.03.027. [DOI] [PubMed] [Google Scholar]
- 98.Krajka-Kuźniak V., Szaefer H., Stefański T., Sobiak S., Cichocki M., Baer-Dubowska W. The effect of resveratrol and its methylthio-derivatives on the Nrf2-ARE pathway in mouse epidermis and HaCaT keratinocytes. Cellular and Molecular Biology Letters. 2014;19(3):500–516. doi: 10.2478/s11658-014-0209-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Soeur J., Eilstein J., Léreaux G., Jones C., Marrot L. Skin resistance to oxidative stress induced by resveratrol: from Nrf2 activation to GSH biosynthesis. Free Radical Biology and Medicine. 2015;78:213–223. doi: 10.1016/j.freeradbiomed.2014.10.510. [DOI] [PubMed] [Google Scholar]
- 100.Bastianetto S., Dumont Y., Duranton A., Vercauteren F., Breton L., Quirion R. Protective action of resveratrol in human skin: possible involvement of specific receptor binding sites. PLoS One. 2010;5(9) doi: 10.1371/journal.pone.0012935.e12935 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Berselli P. V. R., Zava S., Montorfano G., et al. A mint purified extract protects human keratinocytes from short-term, chemically induced oxidative stress. Journal of Agricultural and Food Chemistry. 2010;58(21):11428–11434. doi: 10.1021/jf1020285. [DOI] [PubMed] [Google Scholar]
- 102.Fasano E., Serini S., Mondella N., et al. Antioxidant and anti-inflammatory effects of selected natural compounds contained in a dietary supplement on two human immortalized keratinocyte lines. BioMed Research International. 2014;2014 doi: 10.1155/2014/327452.327452 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Sticozzi C., Belmonte G., Cervellati F., et al. Resveratrol protects SR-B1 levels in keratinocytes exposed to cigarette smoke. Free Radical Biology and Medicine. 2014;69:50–57. doi: 10.1016/j.freeradbiomed.2014.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Sticozzi C., Cervellati F., Muresan X. M., Cervellati C., Valacchi G. Resveratrol prevents cigarette smoke-induced keratinocytes damage. Food & Function. 2014;5(9):2348–2356. doi: 10.1039/c4fo00407h. [DOI] [PubMed] [Google Scholar]
- 105.Herbert K. J., Snow E. T. Modulation of arsenic-induced epidermal growth factor receptor pathway signalling by resveratrol. Chemico-Biological Interactions. 2012;198(1–3):38–48. doi: 10.1016/j.cbi.2012.05.004. [DOI] [PubMed] [Google Scholar]
- 106.Szaefer H., Cichocki M., Brauze D., Baer-Dubowska W. Alteration in phase I and II enzyme activities and polycyclic aromatic hydrocarbons-DNA adduct formation by plant phenolics in mouse epidermis. Nutrition and Cancer. 2004;48(1):70–77. doi: 10.1207/s15327914nc4801_10. [DOI] [PubMed] [Google Scholar]
- 107.Alonso C., Martí M., Barba C., Carrer V., Rubio L., Coderch L. Skin permeation and antioxidant efficacy of topically applied resveratrol. Archives of Dermatological Research. 2017;309(6):423–431. doi: 10.1007/s00403-017-1740-5. [DOI] [PubMed] [Google Scholar]
- 108.Fuggetta M. P., Datri S., Lanzilli G., et al. In vitro antitumour activity of resveratrol in human melanoma cells sensitive or resistant to temozolomide. Melanoma Research. 2004;14(3):189–196. doi: 10.1097/01.cmr.0000130007.54508.b2. [DOI] [PubMed] [Google Scholar]
- 109.Kim A. L., Zhu Y., Zhu H., et al. Resveratrol inhibits proliferation of human epidermoid carcinoma A431 cells by modulating MEK1 and AP-1 signalling pathways. Experimental Dermatology. 2006;15(7):538–546. doi: 10.1111/j.1600-0625.2006.00445.x. [DOI] [PubMed] [Google Scholar]
- 110.Tomikoshi Y., Nomura M., Okudaira N., Sakagami H., Wakabayashi H. Enhancement of cytotoxicity of three apoptosis-inducing agents against human oral squamous cell carcinoma cell line by benzoxazinotropone. In Vivo. 2016;30(5):645–650. [PubMed] [Google Scholar]
- 111.Tyagi A., Gu M., Takahata T., et al. Resveratrol selectively induces DNA Damage, independent of Smad4 expression, in its efficacy against human head and neck squamous cell carcinoma. Clinical Cancer Research. 2011;17(16):5402–5411. doi: 10.1158/1078-0432.ccr-11-1072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Wang X., Zhang Y. Resveratrol alleviates LPS-induced injury in human keratinocyte cell line HaCaT by up-regulation of miR-17. Biochemical and Biophysical Research Communications. 2018;501(1):106–112. doi: 10.1016/j.bbrc.2018.04.184. [DOI] [PubMed] [Google Scholar]
- 113.Carbone M. L., Lulli D., Passarelli F., Pastore S. Topical plant polyphenols prevent type I interferon signaling in the skin and suppress contact hypersensitivity. International Journal of Molecular Sciences. 2018;19(9):p. E2652. doi: 10.3390/ijms19092652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Kang M. C., Cho K., Lee J. H., Subedi L., Yumnam S., Kim S. Y. Effect of resveratrol-enriched rice on skin inflammation and pruritus in the NC/nga mouse model of atopic dermatitis. International Journal of Molecular Sciences. 2019;20(6):p. E1428. doi: 10.3390/ijms20061428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Ravagnan G., De Filippis A., Cartenì M., et al. Polydatin, a natural precursor of resveratrol, induces β-defensin production and reduces inflammatory response. Inflammation. 2013;36(1):26–34. doi: 10.1007/s10753-012-9516-8. [DOI] [PubMed] [Google Scholar]
- 116.Han S.-Y., Choi Y.-J., Kang M.-K., Park J. H. Y., Kang Y.-H. Resveratrol suppresses cytokine production linked to FcεRI-MAPK activation in IgE-antigen complex-exposed basophilic mast cells and mice. The American Journal of Chinese Medicine. 2015;43(8):1605–1623. doi: 10.1142/s0192415x15500913. [DOI] [PubMed] [Google Scholar]
- 117.Caglayan Sozmen S., Karaman M., Cilaker Micili S., et al. Resveratrol ameliorates 2,4-dinitrofluorobenzene-induced atopic dermatitis-like lesions through effects on the epithelium. PeerJ. 2016;4:p. e1889. doi: 10.7717/peerj.1889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Karuppagounder V., Arumugam S., Thandavarayan R. A., et al. Resveratrol attenuates HMGB1 signaling and inflammation in house dust mite-induced atopic dermatitis in mice. International Immunopharmacology. 2014;23(2):617–623. doi: 10.1016/j.intimp.2014.10.014. [DOI] [PubMed] [Google Scholar]
- 119.Kjær T. N., Thorsen K., Jessen N., Stenderup K., Pedersen S. B. Resveratrol ameliorates imiquimod-induced psoriasis-like skin inflammation in mice. PLoS One. 2015;10(5) doi: 10.1371/journal.pone.0126599.e0126599 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Yaman I., Derici H., Kara C., et al. Effects of resveratrol on incisional wound healing in rats. Surgery Today. 2013;43(12):1433–1438. doi: 10.1007/s00595-012-0455-7. [DOI] [PubMed] [Google Scholar]
- 121.Zhao P., Sui B.-D., Liu N., et al. Anti-aging pharmacology in cutaneous wound healing: effects of metformin, resveratrol, and rapamycin by local application. Aging Cell. 2017;16(5):1083–1093. doi: 10.1111/acel.12635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Berce C., Muresan M.-S., Soritau O., et al. Cutaneous wound healing using polymeric surgical dressings based on chitosan, sodium hyaluronate and resveratrol. A preclinical experimental study. Colloids and Surfaces B: Biointerfaces. 2018;163:155–166. doi: 10.1016/j.colsurfb.2017.12.041. [DOI] [PubMed] [Google Scholar]
- 123.Lakshmanan R., Campbell J., Ukani G., et al. Evaluation of dermal tissue regeneration using resveratrol loaded fibrous matrix in a preclinical mouse model of full-thickness ischemic wound. International Journal of Pharmaceutics. 2019;558:177–186. doi: 10.1016/j.ijpharm.2019.01.001. [DOI] [PubMed] [Google Scholar]
- 124.Alasmari W. A., ElSawy N. A., Abourehab M. A. S., Faruk E. M., Alasmari A. A. Effect of topical resveratrol formulation on healing of experimental full thickness wound in diabetic male albino rats: (histological and immunohistochemical study) Life Science Journal. 2018;15(10):6–20. doi: 10.7537/marslsj151018.02. [DOI] [Google Scholar]
- 125.Huang X., Sun J., Chen G., et al. Resveratrol promotes diabetic wound healing via SIRT1-FOXO1-c-myc signaling pathway-mediated angiogenesis. Frontiers in Pharmacology. 2019;10:p. 421. doi: 10.3389/fphar.2019.00421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Gokce E. H., Tuncay Tanrıverdi S., Eroglu I., et al. Wound healing effects of collagen-laminin dermal matrix impregnated with resveratrol loaded hyaluronic acid-DPPC microparticles in diabetic rats. European Journal of Pharmaceutics and Biopharmaceutics. 2017;119:17–27. doi: 10.1016/j.ejpb.2017.04.027. [DOI] [PubMed] [Google Scholar]
- 127.Eckert R. L., Sturniolo M. T., Broome A.-M., Ruse M., Rorke E. A. Transglutaminase function in epidermis. Journal of Investigative Dermatology. 2005;124(3):481–492. doi: 10.1111/j.0022-202x.2005.23627.x. [DOI] [PubMed] [Google Scholar]
- 128.Hong S. P., Kim M. J., Jung M.-y., et al. Biopositive effects of low-dose UVB on epidermis: coordinate upregulation of antimicrobial peptides and permeability barrier reinforcement. Journal of Investigative Dermatology. 2008;128(12):2880–2887. doi: 10.1038/jid.2008.169. [DOI] [PubMed] [Google Scholar]
- 129.Pastore S., Lulli D., Pascarella A., et al. Resveratrol enhances solar UV-induced responses in normal human epidermal keratinocytes. Photochemistry and Photobiology. 2012;88(6):1522–1530. doi: 10.1111/j.1751-1097.2012.01195.x. [DOI] [PubMed] [Google Scholar]
- 130.Seve M., Chimienti F., Devergnas S., et al. Resveratrol enhances UVA-induced DNA damage in HaCaT human keratinocytes. Medicinal Chemistry. 2005;1(6):629–633. doi: 10.2174/157340605774598144. [DOI] [PubMed] [Google Scholar]
- 131.Bickers D. R., Athar M. Oxidative stress in the pathogenesis of skin disease. Journal of Investigative Dermatology. 2006;126(12):2565–2575. doi: 10.1038/sj.jid.5700340. [DOI] [PubMed] [Google Scholar]
- 132.Liguori I., Russo G., Curcio F., et al. Oxidative stress, aging, and diseases. Clinical Interventions in Aging. 2018;13:757–772. doi: 10.2147/CIA.S158513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Gupta R. K., Patel A. K., Shah N., et al. Oxidative stress and antioxidants in disease and cancer: a review. Asian Pacific Journal of Cancer Prevention. 2014;15(11):4405–4409. doi: 10.7314/apjcp.2014.15.11.4405. [DOI] [PubMed] [Google Scholar]
- 134.Pham-Huy L. A., He H., Pham-Huy C. Free radicals, antioxidants in disease and health. International Journal of Biomedical Science. 2008;4(2):89–96. [PMC free article] [PubMed] [Google Scholar]
- 135.Rajendran P., Nandakumar N., Rengarajan T., et al. Antioxidants and human diseases. Clinica Chimica Acta. 2014;436:332–347. doi: 10.1016/j.cca.2014.06.004. [DOI] [PubMed] [Google Scholar]
- 136.Wei T., Chen C., Hou J., Xin W., Mori A. Nitric oxide induces oxidative stress and apoptosis in neuronal cells. Biochimica et Biophysica Acta (BBA)—Molecular Cell Research. 2000;1498(1):72–79. doi: 10.1016/s0167-4889(00)00078-1. [DOI] [PubMed] [Google Scholar]
- 137.Ormerod A. D., Copeland P., Hay I., Husain A., Ewen S. W. B. The inflammatory and cytotoxic effects of a nitric oxide releasing cream on normal skin. Journal of Investigative Dermatology. 1999;113(3):392–397. doi: 10.1046/j.1523-1747.1999.00692.x. [DOI] [PubMed] [Google Scholar]
- 138.Silva S. A. M. E., Michniak-Kohn B., Leonardi G. R. An overview about oxidation in clinical practice of skin aging. Anais Brasileiros de Dermatologia. 2017;92(3):367–374. doi: 10.1590/abd1806-4841.20175481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Kruk J., Duchnik E. Oxidative stress and skin diseases: possible role of physical activity. Asian Pacific Journal of Cancer Prevention. 2014;15(2):561–568. doi: 10.7314/apjcp.2014.15.2.561. [DOI] [PubMed] [Google Scholar]
- 140.Bala A., Mondal C., Haldar P. K., Khandelwal B. Oxidative stress in inflammatory cells of patient with rheumatoid arthritis: clinical efficacy of dietary antioxidants. Inflammopharmacology. 2017;25(6):595–607. doi: 10.1007/s10787-017-0397-1. [DOI] [PubMed] [Google Scholar]
- 141.Yang F., Wolk A., Håkansson N., Pedersen N. L., Wirdefeldt K. Dietary antioxidants and risk of Parkinson’s disease in two population-based cohorts. Movement Disorders. 2017;32(11):1631–1636. doi: 10.1002/mds.27120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Gosselin L. E., Chrapowitzky L., Rideout T. C. Metabolic effects of α-lipoic acid supplementation in pre-diabetics: a randomized, placebo-controlled pilot study. Food & Function. 2019;10(9):5732–5738. doi: 10.1039/c9fo00390h. [DOI] [PubMed] [Google Scholar]
- 143.Wagener F., Carels C., Lundvig D. Targeting the redox balance in inflammatory skin conditions. International Journal of Molecular Sciences. 2013;14(5):9126–9167. doi: 10.3390/ijms14059126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Garcia-Peterson L. M., Wilking-Busch M. J., Ndiaye M. A., Philippe C. G. A., Ahmad V. Sirtuins in skin and skin cancers. Skin Pharmacology and Physiology. 2017;30(4):216–224. doi: 10.1159/000477417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Venza M., Visalli M., Beninati C., De Gaetano G. V., Teti D., Venza I. Cellular mechanisms of oxidative stress and action in melanoma. Oxidative Medicine and Cellular Longevity. 2015:11. doi: 10.1155/2015/481782.481782 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Obrador E., Liu-Smith F., Dellinger R. W., Salvador R., Meyskens F. L., Estrela J. M. Oxidative stress and antioxidants in the pathophysiology of malignant melanoma. Biological Chemistry. 2019;400(5):589–612. doi: 10.1515/hsz-2018-0327. [DOI] [PubMed] [Google Scholar]
- 147.Kruk J., Aboul-Enein H. Y. Reactive oxygen and nitrogen species in carcinogenesis: implications of oxidative stress on the progression and development of several cancer types. Mini-Reviews in Medicinal Chemistry. 2017;17(11):904–919. doi: 10.2174/1389557517666170228115324. [DOI] [PubMed] [Google Scholar]
- 148.Godic A., Poljšak B., Adamic M., Dahmane R. The role of antioxidants in skin cancer prevention and treatment. Oxidative Medicine and Cellular Longevity. 2014:6. doi: 10.1155/2014/860479.860479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Ko J.-H., Sethi G., Um J.-Y., et al. The role of resveratrol in cancer therapy. International Journal of Molecular Sciences. 2017;18(12):p. 2589. doi: 10.3390/ijms18122589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Sayin V. I., Ibrahim M. X., Larsson E., Nilsson J. A., Lindahl P., Bergo M. O. Antioxidants accelerate lung cancer progression in mice. Science Translational Medicine. 2014;6(221) doi: 10.1126/scitranslmed.3007653.221ra15 [DOI] [PubMed] [Google Scholar]
- 151.Le Gal K., Ibrahim M. X., Wiel C., et al. Antioxidants can increase melanoma metastasis in mice. Science Translational Medicine. 2015;7(308) doi: 10.1126/scitranslmed.aad3740.308re8 [DOI] [PubMed] [Google Scholar]
- 152.Glazer A. M., Winkelmann R. R., Farberg A. S., Rigel D. S. Analysis of trends in US melanoma incidence and mortality. JAMA Dermatology. 2017;153(2):225–226. doi: 10.1001/jamadermatol.2016.4512. [DOI] [PubMed] [Google Scholar]
- 153.Boniol M., Autier P., Gandini S. Melanoma mortality following skin cancer screening in Germany. BMJ Open. 2015;5(9) doi: 10.1136/bmjopen-2015-008158.e008158 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Crowell J. A., Korytko P. J., Morrissey R. L., Booth T. D., Levine B. S. Resveratrol-associated renal toxicity. Toxicological Sciences. 2004;82(2):614–619. doi: 10.1093/toxsci/kfh263. [DOI] [PubMed] [Google Scholar]
- 155.Boocock D. J., Faust G. E. S., Patel K. R., et al. Phase I dose escalation pharmacokinetic study in healthy volunteers of resveratrol, a potential cancer chemopreventive agent. Cancer Epidemiology Biomarkers & Prevention. 2007;16(6):1246–1252. doi: 10.1158/1055-9965.epi-07-0022. [DOI] [PubMed] [Google Scholar]
- 156.Ido Y., Duranton A., Lan F., et al. Acute activation of AMP-activated protein kinase prevents H2O2-induced premature senescence in primary human keratinocytes. PLoS One. 2012;7(4) doi: 10.1371/journal.pone.0035092.e35092 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Ido Y., Duranton A., Lan F., Weikel K. A., Breton L., Ruderman N. B. Resveratrol prevents oxidative stress-induced senescence and proliferative dysfunction by activating the AMPK-FOXO3 cascade in cultured primary human keratinocytes. PLoS One. 2015;10(2) doi: 10.1371/journal.pone.0115341.e0115341 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Buonocore D., Nobile A., Cestone P., et al. Resveratrol-procyanidin blend: nutraceutical and antiaging efficacy evaluated in a placebo-controlled, double-blind study. Clinical, Cosmetic and Investigational Dermatology. 2012;5:159–165. doi: 10.2147/ccid.s36102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Farris P., Zeichner J., Berson D. Efficacy and tolerability of a skin brightening/anti-aging cosmeceutical containing retinol 0.5%, niacinamide, hexylresorcinol, and resveratrol. Journal of Drugs in Dermatology: JDD. 2016;15(15):863–868. [PubMed] [Google Scholar]
- 160.Bertuccelli G., Zerbinati N., Marcellino M., et al. Effect of a quality-controlled fermented nutraceutical on skin aging markers: an antioxidant-control, double-blind study. Experimental and Therapeutic Medicine. 2016;11(3):909–916. doi: 10.3892/etm.2016.3011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Park K., Elias P. M., Hupe M., et al. Resveratrol stimulates sphingosine-1-phosphate signaling of cathelicidin production. Journal of Investigative Dermatology. 2013;133(8):1942–1949. doi: 10.1038/jid.2013.133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Taylor E. J. M., Yu Y., Champer J., Kim J. Resveratrol demonstrates antimicrobial effects against Propionibacterium acnes in vitro. Dermatology and Therapy. 2014;4(2):249–257. doi: 10.1007/s13555-014-0063-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Vitonyte J., Manca M. L., Caddeo C., et al. Bifunctional viscous nanovesicles co-loaded with resveratrol and gallic acid for skin protection against microbial and oxidative injuries. European Journal of Pharmaceutics and Biopharmaceutics. 2017;114:278–287. doi: 10.1016/j.ejpb.2017.02.004. [DOI] [PubMed] [Google Scholar]
- 164.Docherty J., Smith J., Fu M., Stoner T., Booth T. Effect of topically applied resveratrol on cutaneous herpes simplex virus infections in hairless mice. Antiviral Research. 2004;61(1):19–26. doi: 10.1016/j.antiviral.2003.07.001. [DOI] [PubMed] [Google Scholar]
- 165.Lipipun V., Sasivimolphan P., Yoshida Y., et al. Topical cream-based oxyresveratrol in the treatment of cutaneous HSV-1 infection in mice. Antiviral Research. 2011;91(2):154–160. doi: 10.1016/j.antiviral.2011.05.013. [DOI] [PubMed] [Google Scholar]
- 166.Sasivimolphan P., Lipipun V., Ritthidej G., et al. Microemulsion-based oxyresveratrol for topical treatment of herpes simplex virus (HSV) infection: physicochemical properties and efficacy in cutaneous HSV-1 infection in mice. AAPS PharmSciTech. 2012;13(4):1266–1275. doi: 10.1208/s12249-012-9828-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Zeng G., Zhong F., Li J., Luo S., Zhang P. Resveratrol-mediated reduction of collagen by inhibiting proliferation and producing apoptosis in human hypertrophic scar fibroblasts. Bioscience, Biotechnology, and Biochemistry. 2013;77(12):2389–2396. doi: 10.1271/bbb.130502. [DOI] [PubMed] [Google Scholar]
- 168.Bai X.-Z., Liu J.-Q., Yang L.-L., et al. Identification of sirtuin 1 as a promising therapeutic target for hypertrophic scars. British Journal of Pharmacology. 2016;173(10):1589–1601. doi: 10.1111/bph.13460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Ikeda K., Torigoe T., Matsumoto Y., Fujita T., Sato N., Yotsuyanagi T. Resveratrol inhibits fibrogenesis and induces apoptosis in keloid fibroblasts. Wound Repair and Regeneration. 2013;21(4):616–623. doi: 10.1111/wrr.12062. [DOI] [PubMed] [Google Scholar]
- 170.Wagemaker T. A. L., Maia Campos P. M. B. G., Shimizu K., Kyotani D., Yoshida D. Antioxidant-based topical formulations influence on the inflammatory response of Japanese skin: a clinical study using non-invasive techniques. European Journal of Pharmaceutics and Biopharmaceutics. 2017;117:195–202. doi: 10.1016/j.ejpb.2017.03.025. [DOI] [PubMed] [Google Scholar]
- 171.Na J. I., Shin J. W., Choi H. R., Kwon S. H., Park K. C. Resveratrol as a multifunctional topical hypopigmenting agent. International Journal of Molecular Sciences. 2019;20(4):p. E956. doi: 10.3390/ijms20040956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Lee J.-H., Kim J.-S., Park S.-Y., Lee Y.-J. Resveratrol induces human keratinocyte damage via the activation of class III histone deacetylase, Sirt1. Oncology Reports. 2016;35(1):524–529. doi: 10.3892/or.2015.4332. [DOI] [PubMed] [Google Scholar]
- 173.Borra M. T., Smith B. C., Denu J. M. Mechanism of human SIRT1 activation by resveratrol. Journal of Biological Chemistry. 2005;280(17):17187–17195. doi: 10.1074/jbc.m501250200. [DOI] [PubMed] [Google Scholar]
- 174.Hirota A., Kawachi Y., Yamamoto M., Koga T., Hamada K., Otsuka F. Acceleration of UVB-induced photoageing in nrf2 gene-deficient mice. Experimental Dermatology. 2011;20(8):664–668. doi: 10.1111/j.1600-0625.2011.01292.x. [DOI] [PubMed] [Google Scholar]
- 175.Saw C. L., Yang A., Huang M.-T., et al. Nrf2 null enhances UVB-induced skin inflammation and extracellular matrix damages. Cell & Bioscience. 2014;4(1):p. 39. doi: 10.1186/2045-3701-4-39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Hirota A., Kawachi Y., Itoh K., et al. Ultraviolet A irradiation induces NF-E2-related factor 2 activation in dermal fibroblasts: protective role in UVA-induced apoptosis. Journal of Investigative Dermatology. 2005;124(4):825–832. doi: 10.1111/j.0022-202x.2005.23670.x. [DOI] [PubMed] [Google Scholar]
- 177.Saw C. L., Huang M. T., Liu Y., Khor T. O., Conney A. H., Kong A. N. Impact of Nrf2 on UVB-induced skin inflammation/photoprotection and photoprotective effect of sulforaphane. Molecular Carcinogenesis. 2011;50:479–486. doi: 10.1002/mc.20725. [DOI] [PubMed] [Google Scholar]
- 178.Villeneuve N. F., Lau A., Zhang D. D. Regulation of the Nrf2-Keap1 antioxidant response by the ubiquitin proteasome system: an insight into cullin-ring ubiquitin ligases. Antioxidants & Redox Signaling. 2010;13(11):1699–1712. doi: 10.1089/ars.2010.3211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Yarosh D., Dong K., Smiles K. UV-induced degradation of collagen I is mediated by soluble factors released from keratinocytes. Photochemistry and Photobiology. 2008;84(1):67–68. doi: 10.1111/j.1751-1097.2007.00230.x. [DOI] [PubMed] [Google Scholar]
- 180.González S., Pathak M. A. Inhibition of ultraviolet-induced formation of reactive oxygen species, lipid peroxidation, erythema and skin photosensitization by polypodium leucotomos. Photodermatology, Photoimmunology & Photomedicine. 1996;12(2):45–56. doi: 10.1111/j.1600-0781.1996.tb00175.x. [DOI] [PubMed] [Google Scholar]
- 181.Bai L., Pang W. J., Yang Y. J., Yang G. S. Modulation of Sirt1 by resveratrol and nicotinamide alters proliferation and differentiation of pig preadipocytes. Molecular and Cellular Biochemistry. 2008;307(307):129–140. doi: 10.1007/s11010-007-9592-5. [DOI] [PubMed] [Google Scholar]
- 182.Huang K., Chen C., Hao J., et al. Polydatin promotes Nrf2-ARE anti-oxidative pathway through activating Sirt1 to resist AGEs-induced upregulation of fibronetin and transforming growth factor-β1 in rat glomerular messangial cells. Molecular and Cellular Endocrinology. 2015;399:178–189. doi: 10.1016/j.mce.2014.08.014. [DOI] [PubMed] [Google Scholar]
- 183.Chai D., Zhang L., Xi S., Cheng Y., Jiang H., Hu R. Nrf2 activation induced by Sirt1 ameliorates acute lung injury after intestinal ischemia/reperfusion through NOX4-mediated gene regulation. Cellular Physiology and Biochemistry. 2018;46(2):781–792. doi: 10.1159/000488736. [DOI] [PubMed] [Google Scholar]
- 184.Kim E. N., Lim J. H., Kim M. Y., et al. Resveratrol, an Nrf2 activator, ameliorates aging-related progressive renal injury. Aging. 2018;10(1):83–99. doi: 10.18632/aging.101361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Li X. N., Ma L. Y., Ji H., et al. Resveratrol protects against oxidative stress by activating the Keap‑1/Nrf2 antioxidant defense system in obese-asthmatic rats. Experimental and Therapeutic Medicine. 2018;16(6):4339–4348. doi: 10.3892/etm.2018.6747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Huang K., Gao X., Wei W. The crosstalk between Sirt1 and Keap1/Nrf2/ARE anti-oxidative pathway forms a positive feedback loop to inhibit FN and TGF-β1 expressions in rat glomerular mesangial cells. Experimental Cell Research. 2017;361(1):63–72. doi: 10.1016/j.yexcr.2017.09.042. [DOI] [PubMed] [Google Scholar]
- 187.Zhou F., Huang X., Pan Y., et al. Resveratrol protects HaCaT cells from ultraviolet B-induced photoaging via upregulation of HSP27 and modulation of mitochondrial caspase-dependent apoptotic pathway. Biochemical and Biophysical Research Communications. 2018;499(3):662–668. doi: 10.1016/j.bbrc.2018.03.207. [DOI] [PubMed] [Google Scholar]
- 188.She Q. B., Bode A. M., Ma W. Y., Chen N. Y., Dong Z. Resveratrol-induced activation of p53 and apoptosis is mediated by extracellular-signal-regulated protein kinases and p38 kinase. Cancer Research. 2001;61(61):1604–1610. [PubMed] [Google Scholar]
- 189.Fernàndez-Busquets X., Fadda A. M., Manconi M., et al. Involvement of c-jun NH(2)-terminal kinases in resveratrol-induced activation of p53 and apoptosis. Mol Carcinog. 2002;33(4):244–250. doi: 10.1002/mc.10041. [DOI] [PubMed] [Google Scholar]
- 190.Riabowol K., Schiff J., Gilman M. Z. Transcription factor AP-1 activity is required for initiation of DNA synthesis and is lost during cellular aging. Proceedings of the National Academy of Sciences. 1992;89(1):157–161. doi: 10.1073/pnas.89.1.157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Leiro J., Arranz J. A., Fraiz N., Sanmartín M. L., Quezada E., Orallo F. Effect of cis-resveratrol on genes involved in nuclear factor kappa B signaling. International Immunopharmacology. 2005;5(2):393–406. doi: 10.1016/j.intimp.2004.10.006. [DOI] [PubMed] [Google Scholar]
- 192.Chun Y. J., Kim M. Y., Guengerich F. P. Resveratrol is a selective human cytochrome P450 1A1 inhibitor. Biochemical and Biophysical Research Communications. 1999;262(1):20–24. doi: 10.1006/bbrc.1999.1152. [DOI] [PubMed] [Google Scholar]
- 193.Schwarz D., Roots I. In vitro assessment of inhibition by natural polyphenols of metabolic activation of procarcinogens by human CYP1A1. Biochemical and Biophysical Research Communications. 2003;303(3):902–907. doi: 10.1016/s0006-291x(03)00435-2. [DOI] [PubMed] [Google Scholar]
- 194.Benitez D. A., Pozo-Guisado E., Alvarez-Barrientos A., Fernandez-Salguero P. M., Castellón E. A. Mechanisms involved in resveratrol-induced apoptosis and cell cycle arrest in prostate cancer-derived cell lines. Journal of Andrology. 2007;28(2):282–293. doi: 10.2164/jandrol.106.000968. [DOI] [PubMed] [Google Scholar]
- 195.Kundu J. K., Shin Y. K., Kim S. H., Surh Y.-J. Resveratrol inhibits phorbol ester-induced expression of COX-2 and activation of NF-κB in mouse skin by blocking IκB kinase activity. Carcinogenesis. 2006;27(7):1465–1474. doi: 10.1093/carcin/bgi349. [DOI] [PubMed] [Google Scholar]
- 196.Aziz M. H., Nihal M., Fu V. X., Jarrard D. F., Ahmad N. Resveratrol-caused apoptosis of human prostate carcinoma LNCaP cells is mediated via modulation of phosphatidylinositol 3′-kinase/Akt pathway and Bcl-2 family proteins. Molecular Cancer Therapeutics. 2006;5(5):1335–1341. doi: 10.1158/1535-7163.mct-05-0526. [DOI] [PubMed] [Google Scholar]
- 197.Vancurova I., Vancura A. Regulation and function of nuclear IκBα in inflammation and cancer. American Journal of Clinical and Experimental Immunology. 2012;1(1):56–66. [PMC free article] [PubMed] [Google Scholar]
- 198.Han S.-Y., Bae J.-Y., Park S.-H., Kim Y.-H., Park J. H. Y., Kang Y.-H. Resveratrol inhibits IgE-mediated basophilic mast cell degranulation and passive cutaneous anaphylaxis in mice. The Journal of Nutrition. 2013;143(5):632–639. doi: 10.3945/jn.112.173302. [DOI] [PubMed] [Google Scholar]
- 199.Doh K. C., Kim B.-M., Kim K. W., Chung B. H., Yang C. W. Effects of resveratrol on Th17 cell-related immune responses under tacrolimus-based immunosuppression. BMC Complementary and Alternative Medicine. 2019;19(1):p. 54. doi: 10.1186/s12906-019-2464-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Zhu X., Liu Q., Wang M., et al. Activation of sirt1 by resveratrol inhibits TNF-α induced inflammation in fibroblasts. PLoS One. 2011;6(11) doi: 10.1371/journal.pone.0027081.e27081 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Li X., Wu G., Han F., et al. SIRT1 activation promotes angiogenesis in diabetic wounds by protecting endothelial cells against oxidative stress. Archives of Biochemistry and Biophysics. 2019;661:117–124. doi: 10.1016/j.abb.2018.11.016. [DOI] [PubMed] [Google Scholar]
- 202.Thomas Hess C. Checklist for factors affecting wound healing. Advances in Skin & Wound Care. 2011;24(4):p. 192. doi: 10.1097/01.ASW.0000396300.04173.ec. [DOI] [PubMed] [Google Scholar]
- 203.Si L., Han Q., Long X., et al. Resveratrol inhibits proliferation and promotes apoptosis of keloid fibroblasts by targeting HIF-1α. Journal of Clinical & Experimental Dermatology Research. 2017;8(6):p. 6. doi: 10.4172/2155-9554.1000436. [DOI] [Google Scholar]
- 204.Wang Y., Lu Y., Ong’achwa M. J., et al. Resveratrol inhibits the TGF-β1-induced proliferation of cardiac fibroblasts and collagen secretion by downregulating miR-17 in rat. BioMed Research International. 2018:10. doi: 10.1155/2018/8730593.8730593 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Newton R. A., Cook A. L., Roberts D. W., Helen Leonard J., Sturm R. A. Post-transcriptional regulation of melanin biosynthetic enzymes by cAMP and resveratrol in human melanocytes. Journal of Investigative Dermatology. 2007;127(9):2216–2227. doi: 10.1038/sj.jid.5700840. [DOI] [PubMed] [Google Scholar]
- 206.Kim E. S., Chang H., Choi H., et al. Autophagy induced by resveratrol suppressesα-MSH-induced melanogenesis. Experimental Dermatology. 2014;23(3):204–206. doi: 10.1111/exd.12337. [DOI] [PubMed] [Google Scholar]
- 207.Kwon S. H., Choi H. R., Kang Y. A., Park K. C. Depigmenting effect of resveratrol is dependent on FOXO3a activation without SIRT1 activation. International Journal of Molecular Sciences. 2017;18(18):p. E1213. doi: 10.3390/ijms18061213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Kim J., Oh J., Averilla J. N., Kim H. J., Kim J. S., Kim J. S. Grape peel extract and resveratrol inhibit wrinkle formation in mice model through activation of Nrf2/HO-1 signaling pathway. Journal of Food Science. 2019;84(6):1600–1608. doi: 10.1111/1750-3841.14643. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental Table 1: content of resveratrol in some plants and foods/beverages.