

Abstract
Previous studies found that opening the cocaine economy by providing post-session access to cocaine had no effect on animals’ demand for cocaine, whereas post-session access to saccharin or food made demand for these non-drug reinforcers more elastic. It is possible that there was no effect of economy type on cocaine taking in these earlier studies because of the delay to the post-session cocaine in the open economy. The present experiment tested whether forming an open economy by providing additional within-session cocaine, rather than post-session cocaine, would make rats’ demand for cocaine more elastic. Saccharin was used as a non-drug comparison reinforcer. Three groups of rats pressed one lever for cocaine and one for saccharin on an ascending series of fixed ratio (FR) schedules where the number of responses required per reinforcer increased from 1 to 48 over sessions. In the open cocaine and open saccharin economy groups, rats had occasional access during the session to a third lever where cocaine or saccharin reinforcers, respectively, were always available on an FR-1 schedule. The main finding was that demand for cocaine was more elastic in the open cocaine economy group than in either of the other groups. Demand for saccharin was more elastic in the open saccharin economy group than in the open cocaine economy group. This study shows that cocaine taking is sensitive to economy type when the additional source of cocaine in an open economy is available close in time to when rats work for cocaine.
Keywords: open economy, closed economy, cocaine, demand elasticity, saccharin, rat
Introduction
Economy type can be an important determinant of the elasticity of demand for a reinforcer (Hursh, 1980). When there is only one source of the reinforcer, the economy is closed. When the reinforcer can be obtained from multiple sources, the economy is open. For example, if the only source of food is that which a subject earns during an experimental session, the food economy is closed. If, on the other hand, the subject earns food during the session and is also given supplemental food by the experimenter after the session, the food economy is open. The behavioral economic prediction is that subjects should work harder for the reinforcer in a closed economy than in an open economy. This is because the extra-session reinforcers available in an open economy can substitute for those earned during the session. In a closed economy, there are no such substitutes. This prediction has been confirmed in studies investigating the elasticity of demand for food in monkeys and mice (Hursh et al., 1989; Soto et al., 2016), saccharin in rats (Gunawan et al., 2019; Kim et al., 2018), and coffee or cigarettes in humans (Mitchell et al., 1994, 1995, 1998).
Kim et al. (2018) tested whether opening the cocaine economy would make demand for it more elastic. In that study, rats first lever pressed for cocaine during a “work” session wherein the fixed ratio (FR) delivering reinforcers increased over sessions up to FR 96. In the open economy, the work session was followed by a 3-h second session during which cocaine was available on an FR-1 schedule. Rats in the closed economy had no access to cocaine outside of the work session. The rats worked just as hard for cocaine as its price increased in the open cocaine economy as in the closed one. In contrast, opening the saccharin economy caused rats to work less hard for saccharin under the same conditions. That the availability of an additional, cheaper source of the reinforcer did not alter elasticity of demand for cocaine, but made demand for saccharin more elastic, might suggest that cocaine and saccharin qualitatively differ with respect to the effect of economy type.
Alternatively, the lack of an effect of economy type on cocaine demand in the study of Kim et al. (2018) could have been due to the temporal parameters under which the extra source of cocaine was presented. In the post-session access open economy arrangement used, the extra-session reinforcers were necessarily delayed. Perhaps for cocaine, but not saccharin, the delay to the extra post-session reinforcers was too great to influence animals’ work session behavior. This might occur if rats’ delay discounting rates were higher for cocaine than for saccharin. It would be expected, however, that a simultaneously available source of cheaper cocaine would substitute for a more expensive source of cocaine. The findings that animals given a choice between cocaine and cocaine on concurrent schedules prefer larger to smaller doses (Iglauer & Woods, 1974; Llewellyn et al., 1976) and prefer richer to leaner schedules of reinforcement (Woolverton, 1996) are consistent with this expectation. By more closely approximating simultaneous availability of more expensive cocaine that animals must work for and a cheaper source of cocaine, an effect of economy type on demand for cocaine might be observed.
The goal of the present experiment was to determine whether presenting the extra cocaine reinforcers available in an open economy nearer in time to when subjects work for cocaine would make demand for cocaine more elastic. The design was similar to that used by Imam (1993), who created an open food economy by occasionally presenting pigeons with food on a variable-time (VT) schedule while they worked for food on a FR-50 schedule. Additional “free” food during the session reduced pigeons’ work output on the FR-50 schedule. A similar strategy was used here to form open cocaine and saccharin economies by providing within-session, rather than post-session, access to cocaine or saccharin reinforcers. Rats pressed one lever for cocaine and a second lever for saccharin on an ascending series of FRs. In the closed economy, this was rats’ only opportunity to obtain these reinforcers. In the open cocaine economy, a third lever was occasionally inserted into the chamber according to a VT schedule and a single press on this lever resulted in a cocaine infusion. The open saccharin economy was formed similarly. It was hypothesized that rats would work harder for cocaine and for saccharin when available in the closed economy than when either of these were available within an open economy.
Methods
Subjects
Twenty-nine adult male Long Evans rats (Envigo, Frederick, MD), weighing approximately 350-450 g at the start of training, completed the experiment. (Other rats began the experiment but did not complete it due to failed catheters [n = 3], illness [n = 1], failure to meet lever press acquisition [n = 8] or stability [n = 4] criteria in a timely manner, or failure to habituate to being tethered/untethered [n = 1]). Rats were individually housed in plastic cages with wood-chip bedding and had unlimited access to rat chow and water in their home cages. The colony room where the rats were housed had a 12-h light:dark cycle with lights on at 08:00 h. Training sessions were conducted five days per week during the light phase of the light:dark cycle. Throughout the experiment, rats were treated in accordance with the Guide for the Care and Use of Laboratory Animals (National Academy of Sciences, 2011) and all procedures were approved by American University’s Institutional Animal Care and Use Committee.
Apparatus
Training took place in Med Associates (St. Albans, VT) operant test chambers. Each chamber measured 30.5 × 24 × 29 cm and had aluminum front and rear walls with clear polycarbonate side walls. Three Med-Associates retractable levers were located on the front wall of the chamber. A Med-Associates small cup liquid receptacle was located directly above the middle lever. Saccharin reinforcers (0.3 ml of 0.2% saccharin water solution) were delivered to this liquid receptacle. A 100-mA cuelight was located above each lever. A speaker located in the center of the front wall provided a tone stimulus (4500 Hz and 80 dB). A 100-mA houselight was located above this speaker. Cocaine (provided by the Drug Supply Program, National Institute on Drug Abuse, Bethesda, MD) in a saline solution at a concentration of 2.56 mg/ml was infused at a rate of 3.19 ml/min by 10-ml syringes driven by Med-Associates (St. Albans, VT) syringe pumps. Tygon tubing extended from the 10-ml syringes to a 22-gauge rodent single-channel fluid swivel (Instech Laboratories, Plymouth Meeting, PA) and tether apparatus (Plastics One, Roanoke, VA) that descended through the ceiling of the chamber. Cocaine was delivered to the subject through tubing that passed through the metal spring of the tether apparatus.
Procedure
Surgery.
Before beginning operant training, all rats were surgically prepared with chronic indwelling jugular vein catheters, using procedures described in detail elsewhere (Thomsen & Caine, 2005; Tunstall & Kearns, 2014). In brief, approximately 3.5 cm of Silastic tubing was inserted into the right jugular vein. From this insertion site, an additional 8 cm of Silastic tubing passed under the skin to the midscapular region where it connected to the 22-gauge stainless steel tubing of a backmount catheter port (Plastics One, Roanoke, VA) that was implanted subcutaneously. The spring tether in the chamber was attached to the threaded plastic cylindrical shaft of the port that protruded through an opening in the skin. All surgery was conducted under ketamine (60 mg/kg) and xylazine (10 mg/kg) anesthesia. Rats were given 7–10 days to recover from surgery. Catheters were flushed daily with 0.1 ml of a saline solution containing 1.25 U/ml heparin and 0.08 mg/ml gentamicin.
Lever press acquisition.
The left and right levers were designated the work levers. The assignment of cocaine or saccharin to the left or right levers was counterbalanced over rats and remained consistent throughout the experiment. All rats were first trained to press the work levers for cocaine and for saccharin in alternate sessions. There was one 3-h session per day. Cocaine sessions began with insertion of the designated lever and illumination of the houselight. A single lever press resulted in delivery of a 0.5 mg/kg cocaine infusion and the initiation of a 10-s timeout signaled by the tone stimulus and illumination of the cuelight above the lever. The houselight turned off during the 10-s timeout. Lever presses during the timeout were recorded but had no scheduled consequences. Saccharin sessions began with illumination of the houselight and insertion of the lever not used for cocaine self-administration. Presses on this lever resulted in delivery of saccharin solution, illumination of the bulb inside the liquid receptacle for 1.66 s (the time it took to fill the well), and initiation of a 10-s timeout signaled by illumination of the cuelight above the lever. The houselight turned off during the 10-s timeout.
During these lever press acquisition sessions, only one of the levers was inserted each session and it remained inserted for the duration of the session. Rats were trained on this procedure, where cocaine sessions alternated with saccharin sessions, for a minimum of 5 sessions with each reinforcer and until they met the acquisition criteria of either obtaining at least 20 cocaine infusions and 20 saccharin reinforcers per session for at least 3 sessions or until they obtained 10 cocaine infusions and 10 saccharin reinforcers per session for at least 5 sessions. Once subjects met the acquisition criteria for one of the reinforcers, they no longer had alternating sessions with the two reinforcer types, but instead they had sessions only with the other reinforcer until meeting the acquisition criteria for that reinforcer. For example, if a rat had met the acquisition criteria for pressing the saccharin lever in five sessions, but had not yet met the cocaine lever pressing acquisition criteria, saccharin sessions were discontinued and only cocaine sessions were given until the rat met the acquisition criteria on the cocaine lever. On average, rats required 5.3 lever press acquisition sessions with the saccharin lever and 11.7 such sessions with the cocaine lever (see Results for more detailed acquisition data). There were no formal criteria for deciding when to exclude a rat from the experiment for failure to meet the lever press acquisition criteria. This decision was made on a subject-by-subject basis and was made prior to group assignment. All of the rats that were excluded had at least 20 sessions without showing evidence of progress in acquiring the lever press response.
Once rats learned to press both work levers, they were assigned to one of three groups: the Closed Economy group (n = 9), the Open Cocaine Economy group (OpenCoc; n = 10), or the Open Saccharin Economy group (OpenSac; n = 10). Group assignment was made with the goal of matching groups as closely as possible on numbers of cocaine infusions and saccharin reinforcers obtained during the final three lever press acquisition sessions, while counterbalancing within each group the left vs. right assignments of the cocaine and saccharin work levers. Rats in the OpenCoc and OpenSac groups then learned to press the middle lever for cocaine or saccharin, respectively. In a subsequent stage, presses on the middle lever would result in the delivery of the extra reinforcers available in the open economy groups. For two 3-h sessions, only the middle lever was inserted. Presses on it were followed by cocaine infusions (OpenCoc group) or saccharin reinforcers (OpenSac group) plus initiation of a 10-s timeout signaled by the same cues previously associated with each reinforcer type. Rats in the Closed group skipped this step of acquisition because they would never receive reinforcers other than those obtained by pressing the work levers.
Demand Training.
All groups then began demand training. Now, the work levers were presented during alternating cocaine and saccharin components within the same session. These work components lasted 18 min each and were followed by 10-min inter-component intervals during which all levers were retracted. There were three work components of each reinforcer type per session, which meant that the total duration spent in work components for each reinforcer was 54 minutes per session. The order of presentation of cocaine and saccharin components was randomized within two-component blocks. Initially, an FR-1 schedule operated during work components.
The key manipulation of the experiment was the scheduling of extra reinforcers during work components for the two open economy groups. For the OpenCoc group, extra opportunities to obtain cocaine occurred according to a VT 2.5-min schedule (range: 1-4 min) that operated during cocaine work components. When the VT schedule timed out, the cocaine work lever retracted, the timer controlling the work component duration paused, and the middle lever inserted. A single press on the middle lever resulted in delivery of a cocaine infusion and presentation of the associated timeout cues. At the end of the 10-s timeout, the middle lever retracted, the cocaine work lever inserted, and the cocaine work component resumed. A limited hold operated such that if no response was made within two min, the middle lever retracted and the work component resumed. Because the work component timer paused while the middle lever inserted, rats still had 54 minutes (divided by 3 work components) of access to the cocaine work lever per session. Dividing 54 min by the 2.5-min average interval of the VT schedule meant that there were, on average, 21.6 insertions of the middle lever per session. The OpenSac group was treated similarly except the middle lever inserted according to the VT 2.5-min schedule during saccharin work components and a press on the middle lever resulted in delivery of saccharin plus associated cues. The Closed group never experienced insertions of the middle lever.
Rats were trained with a FR-1 schedule operating during work components until for three consecutive sessions, the number of total cocaine infusions and total saccharin reinforcers obtained per session did not differ by more than 25% from the rolling three-session mean. Once rats met this stability criterion, the FR on the work levers incremented across three-session blocks according to the following sequence: 3, 6, 12, 24, 48. Extra reinforcers obtained by the open economy groups for pressing the middle lever were always available on a FR-1 schedule, regardless of the FR that was in effect on the work levers.
Data Analysis
For the demand data, the number of cocaine infusions self-administered and the number of saccharin reinforcers earned during work components were averaged over the training sessions at each FR (for FR 1, the mean of the last three sessions was used). Individual and group mean consumption data were fit by Hursh and Silberberg’s (2008) exponential-demand equation:
where Q is quantity consumed, Q0 is consumption as price approaches 0, k is a constant defining the consumption range in log units (k = 2.55 here), α determines the rate of decline in consumption, and C is cost (FR size).
The primary measures of interest were α, which reflects the elasticity of demand, and Q0, which reflects the intensity, or level, of demand. Statistical analyses were performed on these measures derived from the fits of the model to individual subjects’ data. For illustrative purposes, graphs of fits of the model to the group mean data are presented. The group mean demand curves are presented in terms of normalized consumption and normalized price (e.g., see Christensen et al., 2008). To normalize consumption, the numbers of cocaine infusions and saccharin reinforcers obtained were expressed as a percentage of Q0. Price was normalized by converting it to the number of responses required at a particular FR to obtain 1% of Q0.
For all statistical tests, the Type 1 error rate was set to 0.05. One-way or mixed ANOVAs were performed on sessions to meet acquisition criteria, consumption of cocaine and saccharin across FRs, α, Q0, and numbers of extra reinforcers obtained by rats in the open economies. Significant effects of Group were followed by Tukey posthoc tests.
Results
Of the eight rats that were dropped from the experiment before being assigned to groups due to failure to acquire the lever press operant, one rat never regularly pressed either lever after 34 sessions and seven rats learned to press the saccharin lever, but not the cocaine lever despite having over 20 cocaine self-administration sessions. The Closed, OpenCoc, and OpenSac groups required means of 9.4 (± 1.2 SEM), 13.6 (± 3.2), and 11.9 (± 1.9) cocaine self-administration sessions, respectively, to meet the lever press acquisition criteria on the cocaine lever during the acquisition period where only one lever was inserted into the chamber per session. These groups required means of 5.3 (± 0.4 SEM), 5.1 (± 0.1), and 5.6 (± 0.3) sessions, respectively, with the saccharin lever to meet the acquisition criteria. A 3 X 2 (Group X Reinforcer) mixed ANOVA performed on the numbers of acquisition sessions indicated that there was no effect of Group (F < 1) or Group X Reinforcer interaction (F < 1), but there was a significant effect of Reinforcer (F[1,26] = 22.9, p < 0.001). After meeting the lever-press acquisition criteria, the groups advanced to the procedure where cocaine work components and saccharin work components alternated within the same session and an FR-1 schedule was in effect in both types of components. The Closed, OpenCoc, and OpenSac groups required means of 4.9 (± 1.3), 4.7 (± 0.4), and 4.3 (± 0.8) sessions, respectively (no group differences, F < 1), to meet the stability criterion on this procedure before advancing to the procedure where the FR was increased over three-session blocks.
The upper panels of Figure 1 show mean cocaine and saccharin consumption during work components for all groups across the range of work FRs tested. Rats’ work-component cocaine consumption (upper left panel) decreased from about 10-15 infusions per session to about 1-2 as the FR increased from 1 to 48. The OpenSac group tended to self-administer the most cocaine during work components. Saccharin consumption during work components (upper right panel) decreased from about 55-75 reinforcers per session to about 3-4 as the FR increased to 48. At the low FRs, the OpenSac group earned about 20 fewer work component saccharin reinforcers than the OpenCoc group, but as the FR increased this difference disappeared.
Fig 1.
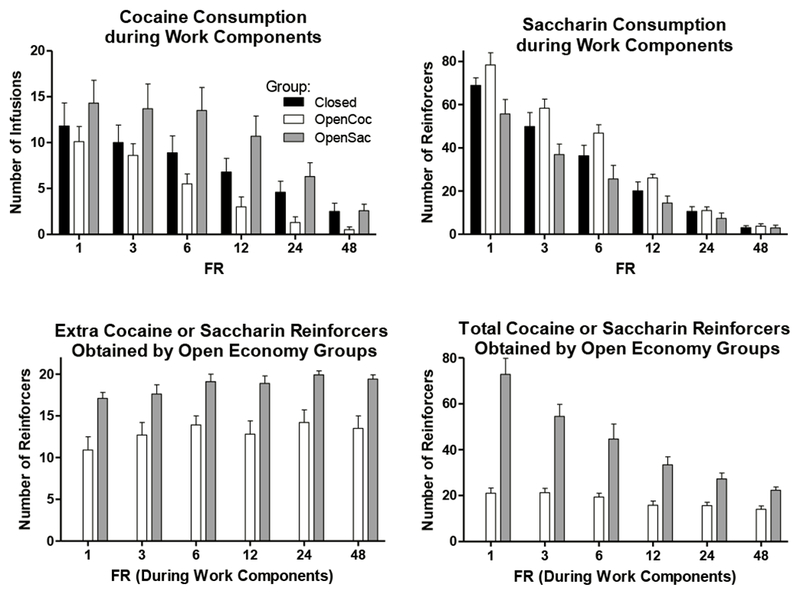
Upper left panel: Mean (± SEM) numbers of cocaine infusions obtained by pressing the cocaine work lever in the Closed (black bars), OpenCoc (white bars), and OpenSac (gray bars) groups across the range of FRs tested. Data are averaged over the three sessions at each FR. Upper right panel: Mean (± SEM) numbers of saccharin infusions obtained by pressing the saccharin work lever in the three groups. Lower left panel: Mean (± SEM) numbers of additional cocaine infusions or saccharin reinforcers obtained by the OpenCoc and OpenSac groups, respectively, by pressing the middle lever. Lower right panel: Mean (± SEM) numbers of total (work lever plus middle lever) cocaine infusions or saccharin reinforcers per session obtained by the OpenCoc and OpenSac groups, respectively.
Statistical analyses confirmed the reliability of these observations. A 3 X 6 (Group X FR) ANOVA performed on cocaine consumption during work components revealed significant main effects of Group (F[2,26] = 3.5, p < 0.05) and FR (F[5,130] = 48.0, p < 0.001), but no significant interaction (F[10,130) = 1.5, NS). Subsequent Tukey tests indicated that, collapsed across FR, the OpenSac group self-administered significantly more cocaine than the OpenCoc group (p < 0.05), but the other two group comparisons were not significant (both ps > 0.35). A 3 X 6 (Group X FR) ANOVA performed on the saccharin work component consumption data revealed significant effects of Group (F[2,26] = 4.9, p < 0.05) and FR (F[5,130] = 224.3, p < 0.001) as well as a significant interaction (F[10,130]= 3.1, p < 0.005). To resolve this interaction, a set of three separate 2 X 6 (Group X FR) ANOVAs were performed comparing each of the possible pairs of groups. Only when the OpenSac group was compared with the OpenCoc group did the effect of Group and the Group X FR interaction remain significant (Group: F[1,18] = 9.7, p < 0.1; interaction: F[5,90] = 5.6, p < 0.001). For the other two comparisons, there was no significant effect of Group (both F[1,17]s ≤ 2.4, NS) and no interaction (both F[5,85]s ≤ 1.8, NS).
The lower left panel of Figure 1 shows the number of extra reinforcers that rats in the OpenCoc and OpenSac groups obtained by pressing the middle lever. The lower right panel shows the number of total (work plus extra) cocaine infusions or saccharin reinforcers earned by these groups. The OpenSac group obtained about 17 to 20 extra saccharin reinforcers per session throughout demand testing. This was near the maximum possible number (21-22) of extra reinforcers they could obtain. The OpenCoc group obtained about 11-14 extra cocaine infusions per session. This meant that OpenCoc rats did not press the middle lever and gave up the opportunity to obtain an extra infusion, on about half of the times it was inserted. A 2 X 6 (Group X FR) ANOVA performed on the numbers of extra reinforcers obtained by rats in the open economy groups indicated that there were significant main effects of Group (F[1,18] = 16.4, p =0.001) and FR (F[5,90] = 5.0, p < 0.001), but no significant interaction (F < 1).
Figure 2 presents normalized demand curves for work component consumption of cocaine and saccharin. Consumption at each price is expressed as a percentage of Q0, the expected number of reinforcers obtained as price approaches 0. Price is expressed as the number of lever presses required at each FR tested to obtain 1% of Q0. The left panel shows that rats in the OpenCoc group worked less hard to defend baseline consumption of cocaine as its price increased than either of the other two groups. The right panel shows that the three groups were generally more similar to each other with respect to how hard they worked to defend consumption of saccharin, though the OpenCoc group worked somewhat harder than the OpenSac group.
Figure 2.
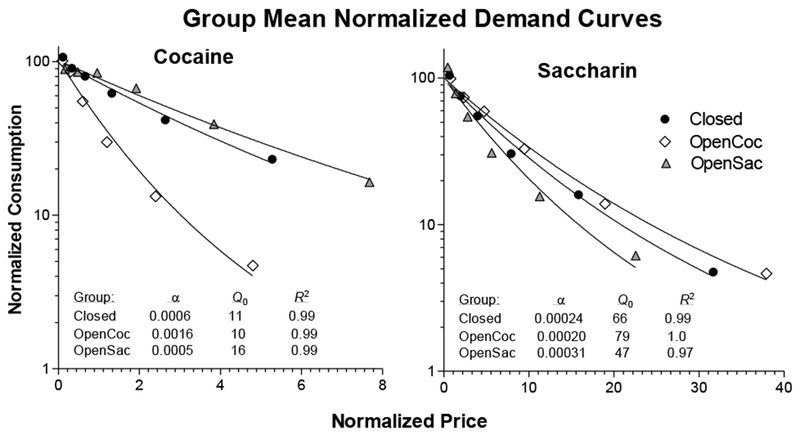
Normalized demand curves fit by Hursh and Silberberg’s (2008) exponential demand model to group mean cocaine (left panel) and saccharin (right panel) data. Consumption is expressed as a percentage of Q0, the expected amount consumed as price approaches 0. Price is expressed as the number of responses required at a particular FR value to produce 1% of Q0 [that is, normalized price = (FR x Q0)/100]. The inset tables present values of α, Q0, and R2 for fits of the model to the non-normalized data.
Figure 3 shows mean cocaine and saccharin α and Q0 derived from fits of the model to individual subjects’ consumption data. The pattern of results is largely consistent with Figure 1. Cocaine demand was more elastic (α was larger) in the OpenCoc group than in either of the other groups. The intensity of demand (Q0) for cocaine was comparable across groups. Elasticity of demand was overall much lower for saccharin than for cocaine and intensity of demand was overall much higher for saccharin than for cocaine (note differences in Y-axis scales for cocaine and saccharin data). Among groups, saccharin demand was most elastic in the OpenSac group. In contrast to the cocaine data where groups did not differ in intensity of demand, there was a group difference in intensity of demand for saccharin, with the OpenSac group having the lowest level of saccharin demand.
Figure 3.
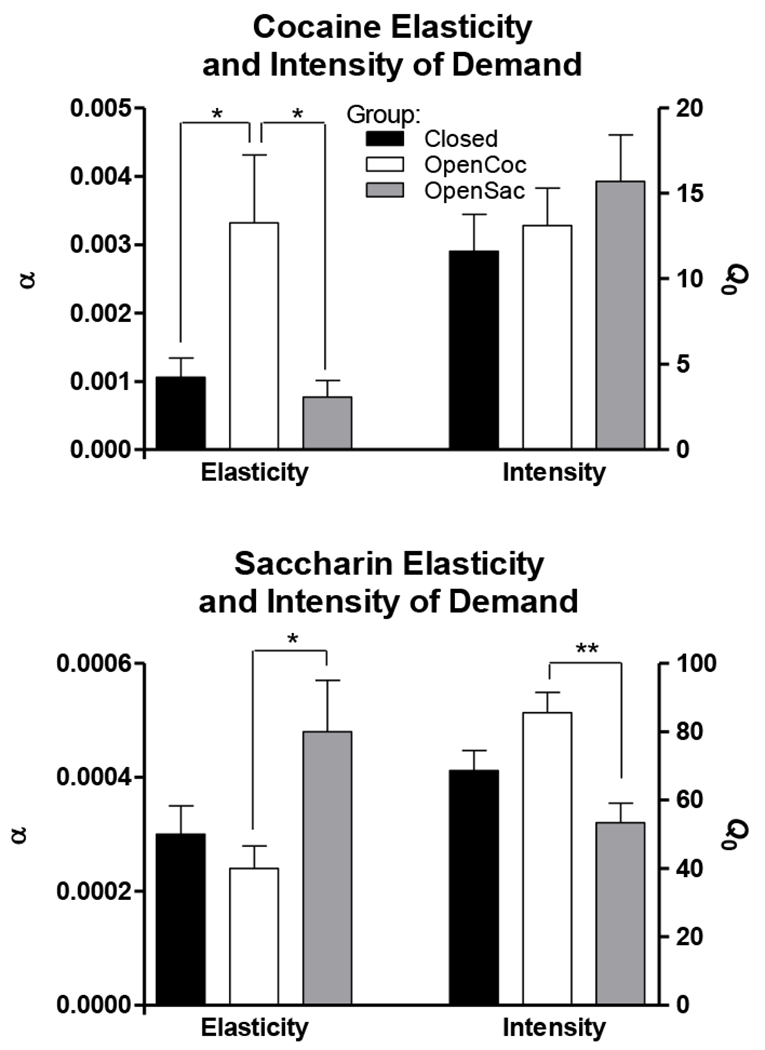
Mean (± SEM) values of α and Q0 derived from fits of the exponential demand model to individual subjects’ consumption data. The upper and lower panels present these estimates for cocaine and saccharin, respectively, for each of the three groups. * indicates p < 0.05 ** indicates p < 0.01
Statistical analyses confirmed these impressions. A 3 X 2 (Group X Reinforcer) ANOVA indicated that for α there were significant main effects of Group (F[2,26] = 4.3, p < 0.05) and Reinforcer (F[1,26] = 14.1, p < 0.005) as well as a significant interaction (F[2,26] = 5.7, p < 0.01). To resolve this interaction, one-way ANOVAs were performed on α for cocaine and saccharin separately. There was a significant effect of Group for cocaine α (F[2,26] = 5.0, p < 0.05) and subsequent Tukey posthoc tests confirmed that α in the OpenCoc group was significantly higher than α in the Closed or OpenSac groups (both ps < 0.05) but that these two latter groups did not differ from each other (p > 0.9). For saccharin α there was also a significant effect of Group (F[2,26] = 3.9, p < 0.05) and subsequent Tukey tests revealed that the OpenSac group had significantly higher α than the OpenCoc group (p < 0.05), but the other group comparisons were not significant (both ps > 0.15). A 3 X 2 (Group X Reinforcer) ANOVA performed on Q0 indicated that there were significant main effects of Group (F[2,26] = 4.9, p < 0.05) and Reinforcer (F[1,26] = 274.4, p < 0.001) as well as a significant interaction (F[2,26] = 9.3, p < 0.005). One-way ANOVAs performed on the cocaine and saccharin Q0 data separately revealed that there was no significant effect of Group for cocaine (F < 1), but there was a significant effect for saccharin (F[2,26] = 7.7, p < 0.005). Subsequent Tukey tests indicated that the OpenSac group had significantly lower Q0 than the OpenCoc group (p < 0.005), but there were no other significant group differences (both ps > 0.1).
Discussion
The main finding of the present experiment was that opening the cocaine economy by providing extra opportunities to obtain cocaine while rats worked for it caused demand for cocaine to become more elastic. The OpenCoc group, which had an additional source of cocaine, worked less hard to defend cocaine consumption during work components than the Closed or OpenSac groups, for which the cocaine economy was closed. This outcome is consistent with the behavioral economic prediction that an additional source of cheaper cocaine should substitute for cocaine available from a more expensive source.
The present results contrast with the previously reported findings that post-session access to cocaine had no effect on rats’ willingness to work for it during a work session (Carroll et al., 2000; Kim et al., 2018). That within-session, but not post-session, extra cocaine makes demand for it more elastic suggests that for cocaine, animals integrate the costs and benefits of current and future cocaine only over fairly short timeframes. In contrast, for non-drug reinforcers like food or saccharin, animals can anticipate future availability, and adjust their current behavior accordingly, over longer timeframes (e.g., Hursh et al., 1989; Kim et al., 2018; Morato et al., 1995). Timberlake et al. (1987) hypothesized that the time horizons over which animals are able to integrate different sources of reward would likely vary by behavior system and reinforcer type. The present results and those of the previous studies described above are consistent with this notion and they also suggest that rates of delay discounting may be steeper for cocaine than for saccharin in rats. Perhaps relatedly, cocaine-dependent humans have been found to have higher delay discount rates for cocaine than for money (Coffey et al., 2003).
A potential alternative to a behavioral economics substitution account of the reduced effort to obtain cocaine observed in the OpenCoc group is satiation. That is, the extra cocaine caused rats to become sated on cocaine and therefore less willing to work for it. Previous studies have shown that rats regulate their intake of cocaine within a limited range and satiation is thought to inhibit further intake as rats approach the upper limit of that range (Norman & Tsibulsky, 2006; Tsibulsky & Norman, 1999). There is evidence from the present study, however, indicating that satiation was not solely responsible for the effect reported here. When the FR was 1 during work components, the OpenCoc group self-administered a mean of 10.1 cocaine infusions during work components and 10.9 extra infusions during insertions of the middle lever, for a total of 21 total infusions per session. When the work component FR was 3, these rats self-administered a mean of 8.6 and 12.7 work and extra infusions, respectively, for a total of 21.3 cocaine infusions per session. Therefore, 21 infusions appears to reflect rats’ preferred, relatively unconstrained level of total cocaine intake per session. As the work component FR increased, OpenCoc group rats’ intake during work components dropped sharply (more so than in the other two groups) while their intake of extra infusions remained fairly constant. For example, at FR 12, they only self-administered an average of 3 infusions per session during work components while self-administering approximately 13 of the extra infusions available for a total of 16 infusions, which was less than their preferred total intake of 21 infusions per session. A similar effect can also be observed at FR 24 and FR 48. This suggests that, rather than satiation limiting intake at higher FRs, OpenCoc rats were very sensitive to cocaine’s price and less willing to work for it on high FRs.
Further evidence for the idea that satiation alone cannot account for the effect of opening the cocaine economy type comes from the fact that Q0, or the level of consumption expected at price 0, for cocaine did not differ across groups. Satiation would be expected to reduce Q0, or the intensity of demand (Hursh, 2014). For example, in a study with monkeys, opening the food economy allowed subjects to gain bodyweight which resulted in both reduced intensity of demand for food and increased elasticity of demand for food (Hursh et al., 1989). That in the present experiment, α, but not Q0, was altered in the OpenCoc group indicates that opening the cocaine economy affected the elasticity of demand, but not the intensity of demand. This all suggests that cocaine satiation was not the primary cause of the increased elasticity of demand for cocaine observed in the OpenCoc group.
The effects of opening the saccharin economy on saccharin demand were somewhat less clear in the present experiment. The OpenSac and Closed groups did not significantly differ from each other with respect to α, though it is worth noting that mean α was 60% higher in the OpenSac group than in the Closed group. Compared to the OpenCoc group, however, the OpenSac group had significantly more elastic demand for saccharin. Thus, the group with the open saccharin economy significantly differed in the predicted direction from one of the two groups that had closed saccharin economies (and these two latter groups were quite similar to each other in terms of α). In the studies of Kim et al. (2018) and Gunawan et al. (2019), where the saccharin economy was opened by providing post-session access to saccharin, the differences between the open and closed economy groups were significant. The more consistent effect observed in those earlier studies may have been due to the total amounts of extra saccharin available to subjects in the open saccharin economies. In both of those studies, rats had 3 h of unlimited access to saccharin on an FR-1 schedule after the work session. In the study of Kim et al. (2018) rats obtained approximately 40 post-session saccharin reinforcers and in the study of Gunawan et al. (2019 )they obtained about 40-80 extra saccharin reinforcers. In the present experiment, rats in the OpenSac group obtained only 17-20 extra saccharin reinforcers. A relatively low limit was imposed to minimize the possibility of the additional within-session saccharin reinforcers producing satiation. Perhaps giving a few additional free saccharin presentations here would have increased the size of the economy type effect such that the OpenSac group would have significantly differed from the Closed group.
Collapsed across FRs, the OpenSac group self-administered significantly more cocaine infusions than the OpenCoc group. Inspection of Figure 1 shows that the OpenSac group also tended to self-administer more cocaine than the Closed group, but this difference did not reach significance. That the OpenSac group self-administered the most cocaine is suggestive of a possible interaction between the reinforcers. Dynamics related to incentive contrast (Flaherty, 1982) could have been at work due to the alternating saccharin and cocaine components experienced by all groups in the present experiment. That rats in all groups self-administered more saccharin than cocaine and worked harder to defend saccharin intake than cocaine intake suggests that saccharin was the higher value reinforcer. Alternating saccharin and cocaine components could have produced a negative contrast effect, causing rats to self-administer less cocaine than they would have if saccharin were not available. Opening the saccharin economy in the OpenSac group might have reduced the value of saccharins enough to mitigate this effect, leading to increased cocaine self-administration in this group. It must be acknowledged, however, that the present experiment was not designed to manipulate or measure incentive contrast and therefore this account should be regarded cautiously.
An aspect of the procedure that should be considered when interpreting results is that rats in the open economy groups received their extra reinforcers by pressing the middle lever which was inserted on a VT schedule whereas the Closed group had no experience with insertions of the middle lever on a VT schedule. Experience with the VT schedule and middle lever itself (without regard to the reinforcer delivered) could have contributed to the results. Perhaps inserting the middle lever on a VT schedule for rats in the Closed group and allowing them to press it for an innocuous event like a saline infusion or stimulus flash would have made the groups more procedurally equivalent. This was not done here, however, because such a procedure might have caused the groups to differ behaviorally in unintended ways. For example, if the consequence for pressing the middle lever was so innocuous that it was not reinforcing, the middle lever might have served as a long timeout or extinction cue. Having periods where lever pressing was reinforced alternate with periods where lever pressing was not reinforced could have created conditions conducive to producing behavioral contrast (Reynolds, 1962) in the Closed group, but not the other groups.
The dose of cocaine used here was 0.5 mg/kg/infusion. Previous studies have found that this dose is on the descending limb of the dose-response function for low FR schedules in rats (e.g., Wee et al., 2007; Schenk et al., 1999; Mantsch et al., 2004). But this dose is on the ascending limb for progressive ratio (PR) schedules (e.g., Gancarz et al., 2012; Morgan et al., 2006; Roberts et al., 2002). That is, as the dose per infusion increases from 0 to as high as 3.0 mg/kg, rats’ PR breakpoints increase monotonically. Based on these findings, if a larger dose of cocaine were used in the present experiment, rats would be expected to self-administer fewer infusions at the low ratios, but more infusions at the higher ratios near rats’ breaking points. If a smaller dose were used here, rats would be expected to self-administer more infusions at the low FRs and fewer infusions at the high FRs. The 0.5 mg/kg/infusion dose was chosen with the goal of using a dose and set of FRs that would capture the complete demand curve. It is worth noting that previous studies have shown that elasticity of demand for cocaine is dose-independent if doses above a relatively low threshold dose are used (Hursh & Winger, 1995; Kearns & Silberberg, 2016; Winger et al., 2006). For example, Kearns & Silberberg (2016) found no difference in elasticity of demand for 0.33 mg/kg and 1.0 mg/kg infusions, but demand for 0.11 mg/kg infusions was more elastic than it was for the two higher doses.
In the OpenCoc group, the same 0.5 mg/kg dose was used for reinforcers earned during cocaine work components and for the extra reinforcers available for pressing the middle lever. If a dose smaller than 0.5 mg/kg/infusion were used for the extra reinforcers, OpenCoc rats likely would have self-administered more than 11-14 of them on average. If the dose were larger than 0.5 mg/kg, they would have likely self-administered fewer of them. The goal here was to use a dose that would result in rats self-administering the majority of the extra reinforcers to ensure sufficient contact with the scheduled contingencies. It appears that the 0.5 mg/kg dose was slightly on the high side. Nonetheless, rats had enough contact with the contingencies to learn that the extra, middle lever infusions were cheaper than the work component infusions as evidenced by the significant effect of economy type on cocaine’s demand elasticity.
For the open economy groups, the FR was always 1 on the VT lever. The goal was to make the extra reinforcers cheaper than the work component reinforcers to test whether the extra reinforcers would substitute for the work component reinforcers. Ideally, the extra reinforcers would have been free. For example, cocaine or saccharin reinforcers in the open economy groups might have been given non-contingently. That was not done here because results of previous studies suggest that the experience of non-contingent cocaine infusions may qualitatively differ from that of self-administered infusions (Mark, et al., 1999; Palamarchouk et al., 2009; Ploense et al., 2018; Markou et al., 1999). Therefore, extra reinforcers were available on an FR-1 schedule. In theory, similar results might have been obtained if the FR on the middle lever were greater than 1 but less than the FR on the work levers. But making the ratios available on the middle and work levers more similar to each other would likely have made it more difficult for rats to learn that the middle lever signaled the availability of cheaper reinforcers. Relatedly, the price of the extra reinforcers could have been manipulated by varying the dose of cocaine or the magnitude of the saccharin reinforcer provided for pressing the middle lever. In the present experiment, the same reinforcer magnitude was used for the extra reinforcers and the work component reinforcers for the sake of simplicity.
The present finding that provision of an additional source of cheap cocaine made demand for cocaine more elastic may model the effects of replacement therapies. Indeed, Hursh (1991) has noted that, fundamentally, replacement therapies open the drug economy by providing a cheaper source of one drug that can substitute for the more expensive abused drug. For example, cheaper methadone available from a clinic substitutes for more expensive heroin available on the street. While opioid replacement therapies (methadone and buprenorphine) have been quite effective in treating opioid use disorder (Mattick et al., 2009; Shuckit, 2016; Sigmon et al., 2016), psychostimulant replacement therapy is not yet widely used to treat cocaine use disorder. The results of the OpenCoc group here suggest that an additional, cheaper source of cocaine itself can act to reduce cocaine’s value. Obviously, treating cocaine use disorder with cocaine is unlikely to be acceptable. A different psychostimulant, with an abuse potential lower than cocaine’s, might be able to open the psychostimulant economy enough to make demand for cocaine more elastic. Stoops and Rush (2013) reviewed research on psychostimulant replacement therapies and concluded that dopamine releasers, like d-amphetamine, and the atypical psychostimulant modafinil show some promise for treating cocaine use disorder. For example, Grabowski et al. (2001) found in a clinical trial that d-amphetamine maintenance significantly reduced cocaine use in cocaine-dependent individuals (see Grabowski et al., 2004, Levin et al., 2015, and Mooney et al., 2009 for similar results). If part of the effectiveness of replacement therapies is due to an opening of the drug economy, the present results along with those of Carroll et al. (2000) and Kim et al. (2018) suggest that the timing of the availability of the replacement psychostimulant could be an important determinant of its potential success in acting as a substitute for cocaine.
Acknowledgments
Funding Source: This research was supported by award number R01DA037269 from the National Institute on Drug Abuse. The National institute on Drug Abuse had no role other than financial support and as such the content is solely the responsibility of the authors.
Footnotes
Conflicts of interest: None
References
- Carroll ME, Cosgrove KP, Campbell UC, Morgan AD, Mickelberg JL (2000). Reductions in ethanol, phencyclidine, and food-maintained behavior by naltrexone pretreatment in monkeys is enhanced by open economic conditions. Psychopharmacology 148:412–422. [DOI] [PubMed] [Google Scholar]
- Christensen CJ, Silberberg A, Hursh SR, Huntsberry ME, Riley AL (2008). Essential value of cocaine and food in rats: tests of the exponential model of demand. Psychopharmacology 198:221–229. [DOI] [PubMed] [Google Scholar]
- Coffey SF, Gudleski GD, Saladin ME, Brady KT (2003). Impulsivity and rapid discounting of delayed hypothetical rewards in cocaine-dependent individuals. Exp Clin Psychopharmacology 11:18–25. [DOI] [PubMed] [Google Scholar]
- Gancarz AM, Kausch MA, Lloyd DR, Richards JB (2012). Between-session progressive ratio performance in rats responding for cocaine and water reinforcers. Psychopharmacology 222:215–223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grabowski J, Rhoades H, Schmitz J, Stotts A, Daruzska LA, Creson D, Moeller FG (2001). Dextroamphetamine for cocaine-dependence treatment: a double-blind randomized clinical trial. J Clin Psychopharmacology 21:522–526. [DOI] [PubMed] [Google Scholar]
- Grabowski J, Rhoades H, Stotts A, Cowan K, Kopecky C, Dougherty A, Moeller FG, Hassan S, Schmitz J. (2004). Agonist-like or antagonist-like treatment for cocaine dependence with methadone for heroin dependence: two double-blind randomized clinical trials. Neuropsychopharmacology 29:969–981. [DOI] [PubMed] [Google Scholar]
- Gunawan T, Tripoli CS, Silberberg A, Kearns DN (2019). The effect of economy type on heroin and saccharin essential value. Exp Clin Psychopharmacol, in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hursh SR (1991). Behavioral economics of drug self-administration and drug abuse policy. J Exp Anal Behav 56:377–393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hursh SR (2014). Behavioral economics and the analysis of consumption and choice In McSweeney FK, Murphy ES (Eds.), The Wiley Blackwell Handbook of Operant and Classical Conditioning (pp. 275–305). Hoboken, NJ: Wiley. [Google Scholar]
- Hursh SR, Silberberg A (2008). Economic demand and essential value. Psych Rev 115:186–198. [DOI] [PubMed] [Google Scholar]
- Hursh SR, Winger G (1995). Normalized demand for drugs and other reinforcers. J Exp Anal Behav 64:373–384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hursh SR, Raslear TG, Bauman R, Black H (1989). The quantitative analysis of economic behavior with laboratory animals In Grunert KG, Olander F (Eds.), Understanding Economic Behavior (pp. 393–407). Dordrecht, Netherlands: Springer. [Google Scholar]
- Iglauer C, Woods JH (1974). Concurrent performances: reinforcement by different doses of intravenous cocaine in rhesus monkeys. J Exp Anal Behav 22:179–196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imam AA (1993). Response-reinforcer independence and the economic continuum: A preliminary analysis. J Exp Anal Behav 59:231–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kearns DN, & Silberberg A (2016). Dose and elasticity of demand for self-administered cocaine in rats. Behav Pharmacol 27:289–292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim JS, Gunawan T, Tripoli CS, Silberberg A, Kearns DN (2018) The effect of economy type on demand and preference for cocaine and saccharin in rats. Drug Alcohol Depend 192:150–157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levin FR, Mariani JJ, Specker S, Mooney M, Mahony A, Brooks DJ, Babb D, Bai Y, Eberly LE, Nunes EV, Grabowski J. (2015). Extended-release mixed amphetamine salts vs placebo for comorbid adult attention-deficit/hyperactivity disorder and cocaine use disorder: a randomized clinical trial. JAMA Psychiatry 72:593–602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Llewellyn ME, Iglauer C, & Woods JH (1976). Relative reinforcer magnitude under a nonindependent concurrent schedule of cocaine reinforcement in rhesus monkeys. J Exp Anal Behav 25:81–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mantsch JR, Yuferov V, Mathieu-Kia AM, Ho A, Kreek MJ (2004). Effects of extended access to high versus low cocaine doses on self-administration, cocaine-induced reinstatement and brain mRNA levels in rats. Psychopharmacology 175:26–36. [DOI] [PubMed] [Google Scholar]
- Mark GP, Hajnal A, Kinney AE, Keys AS (1999). Self-administration of cocaine increases the release of acetylcholine to a greater extent than response-independent cocaine in the nucleus accumbens of rats. Psychopharmacology 143:47–53. [DOI] [PubMed] [Google Scholar]
- Markou A, Arroyo M, Everitt BJ (1999). Effects of contingent and non-contingent cocaine on drug-seeking behavior measured using a second-order schedule of cocaine reinforcement in rats. Neuropsychopharmacology 20:542–555. [DOI] [PubMed] [Google Scholar]
- Mattick RP, Breen C, Kimber J, Davoli M (2009). Methadone maintenance therapy versus no opioid replacement therapy for opioid dependence. Cochrane Database Syst Rev 3. [DOI] [PubMed] [Google Scholar]
- Mitchell SH, De Wit H, Zacny JP (1994). Effects of varying the” openness” of an economy on responding for cigarettes. Behav Pharmacol 5:159–166. [DOI] [PubMed] [Google Scholar]
- Mitchell SH, Laurent CL, De Wit H, Zacny JP (1995). Effects of price,‘openness’ of the economy and magnitude of the alternative reinforcer on responding for caffeinated coffee. Hum Psychopharm Clin 10:39–46. [Google Scholar]
- Mitchell SH, De Wit H, Zacny JP (1998). The impact of three economic factors on cigarette procurement and consumption. Hum Psychopharm Clin 13:259–66. [Google Scholar]
- Mooney ME., Herin DV, Specker S, Babb D, Levin FR, Grabowski J. (2015). Pilot study of the effects of lisdexamfetamine on cocaine use: A randomized, double-blind, placebo-controlled trial. Drug Alc Depend 153:94–103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgan D, Liu Y, Roberts DC (2006). Rapid and persistent sensitization to the reinforcing effects of cocaine. Neuropsychopharmacology 31:121–128. [DOI] [PubMed] [Google Scholar]
- Morato S, Johnson DF, Collier G (1995). Feeding patterns of rats when food-access cost is alternately low and high. Physiol Behav 57:21–26. [DOI] [PubMed] [Google Scholar]
- National Academy of Sciences (2011). Guide for the Care and Use of Laboratory Animals. Washington, DC: National Academy Press. [Google Scholar]
- Norman AB, Tsibulsky VL (2006). The compulsion zone: a pharmacological theory of acquired cocaine self-administration. Brain Res 1116:143–152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palamarchouk V, Smagin G, Goeders NE (2009). Self-administered and passive cocaine infusions produce different effects on corticosterone concentrations in the medial prefrontal cortex (MPC) of rats. Pharm Biochem Behav 94:163–168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ploense KL, Vieira P, Bubalo L, Olivarria G, Carr AE, Szumlinski KK, Kippin TE (2018). Contributions of prolonged contingent and non-contingent cocaine exposure to escalation of cocaine intake and glutamatergic gene expression. Psychopharmacology 235:1347–1359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reynolds GS (1961). Behavioral contrast. J Exp Anal Behav 4:57–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts DC, Brebner K, Vincler M, Lynch WJ (2002). Patterns of cocaine self-administration in rats produced by various access conditions under a discrete trials procedure. Drug Alc Depend 67:291–299. [DOI] [PubMed] [Google Scholar]
- Schenk S, Partridge B, Shippenberg TS (1999). U69593, a kappa-opioid agonist, decreases cocaine self-administration and decreases cocaine-produced drug-seeking. Psychopharmacology 144: 339–346. [DOI] [PubMed] [Google Scholar]
- Schuckit MA (2016). Treatment of opioid-use disorders. N Engl J Med 375:357–68. [DOI] [PubMed] [Google Scholar]
- Sigmon SC, Ochalek TA, Meyer AC, Hruska B, Heil SH, Badger GJ, Rose G, Brooklyn JR, Schwartz RP, Moore BA, Higgins ST (2016). Interim buprenorphine vs. waiting list for opioid dependence. N Engl J Med 375:2504–2505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soto PL, Hiranita T, Xu M, Hursh SR, Grandy DK, Katz JL (2016). Dopamine D 2-like receptors and behavioral economics of food reinforcement. Neuropsychopharmacology 41:971–978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoops WW, Rush CR (2013). Agonist replacement for stimulant dependence: a review of clinical research. Curr Pharm Des 19:7026–7035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomsen M, Caine SB (2005). Chronic intravenous drug self-administration in rats and mice. Curr Prot Neurosci 32:9–20. [DOI] [PubMed] [Google Scholar]
- Timberlake W, Gawley DJ, Lucas GA (1987). Time horizons in rats foraging for food in temporally separated patches. J Exp Psychol: Anim Behav Proc 13:302–309. [PubMed] [Google Scholar]
- Tunstall BJ, Kearns DN (2014). Reinstatement in a cocaine versus food choice situation: reversal of preference between drug and non-drug rewards. Add Biol 19:838–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsibulsky VL, Norman AB (1999). Satiety threshold: a quantitative model of maintained cocaine self-administration. Brain Res 839:85–93. [DOI] [PubMed] [Google Scholar]
- Wee S, Specio SE, Koob GF (2007). Effects of dose and session duration on cocaine self-administration in rats. J Pharmacol Exp Ther 320: 1134–1143. [DOI] [PubMed] [Google Scholar]
- Winger G, Galuska CM, Hursh SR,Woods JH (2006). Relative reinforcing effects of cocaine, remifentanil, and their combination in rhesus monkeys. J Pharmacol Exp Ther 318:223–229. [DOI] [PubMed] [Google Scholar]
- Woolverton WL (1996). Intravenous self-administration of cocaine under concurrent VI schedules of reinforcement. Psychopharmacology 127:195–203. [PubMed] [Google Scholar]