

Abstract
Most of the microorganisms responsible for vector-borne diseases (VBD) have hematophagous arthropods as vector/reservoir. Recently, many new species of microorganisms phylogenetically related to agents of VBD were found in a variety of aquatic eukaryotic hosts; in particular, numerous new bacterial species related to the genus Rickettsia (Alphaproteobacteria, Rickettsiales) were discovered in protist ciliates and other unicellular eukaryotes. Although their pathogenicity for humans and terrestrial animals is not known, several indirect indications exist that these bacteria might act as etiological agents of possible VBD of aquatic organisms, with protists as vectors. In the present study, a novel strain of the Rickettsia-Like Organism (RLO) endosymbiont “Candidatus (Ca.) Trichorickettsia mobilis” was identified in the macronucleus of the ciliate Paramecium multimicronucleatum. We performed transfection experiments of this RLO to planarians (Dugesia japonica) per os. Indeed, the latter is a widely used model system for studying bacteria pathogenic to humans and other Metazoa. In transfection experiments, homogenized paramecia were added to food of antibiotic-treated planarians. Treated and non-treated (i.e. control) planarians were investigated at day 1, 3, and 7 after feeding for endosymbiont presence by means of PCR and ultrastructural analyses. Obtained results were fully concordant and suggest that this RLO endosymbiont can be transiently transferred from ciliates to metazoans, being detected up to day 7 in treated planarians’ enterocytes. Our findings might offer insights into the potential role of ciliates or other protists as putative vectors for diseases caused by Rickettsiales or other RLOs and occurring in fish farms or in the wild.
Keywords: Planarians, Molecular characterization, Rickettsiaceae, Paramecium multimicronucleatum, “Candidatus Trichorickettsia”, FISH, Vector-borne diseases, Rickettsia-like organism (RLO) endosymbiont, Ultrastructure, Transfection experiments
Introduction
Bacteria of order Rickettsiales (Alphaproteobacteria) live in an obligate association with a wide range of eukaryotes (Dumler et al., 2001; Taylor, Bandi & Hoerauf, 2005; Perlman, Hunter & Zchori-Fein, 2006; Weinert et al., 2009; Gillespie et al., 2012; Dumler & Walker, 2015; Castelli, Sassera & Petroni, 2016; Chan et al., 2018), and, for their vast majority, are localized intracellularly, although the case of an extracellular Rickettsiales bacterium was recently documented (Castelli et al., 2019a). Rickettsiales are widely studied for their involvement in medical and veterinary fields. Indeed, many of them (e.g., Rickettsia spp., Anaplasma spp., Ehrlichia spp., and Orientia tsutsugamushi) are vectored by lice, ticks and mites, and cause mild to severe disease (Dumler et al., 2001; Dumler & Walker, 2015), such as epidemic typhus, Rocky Mountain spotted fever (Raoult & Roux, 1997; Walker & Ismail, 2008; McQuiston & Paddock, 2012), anaplasmosis, ehrlichiosis (Dumler et al., 2007; Rikihisa, 2010), heartwater (Allsopp, 2015), and scrub typhus (Ge & Rikihisa, 2011).
In the last two decades, many new genera and species of Rickettsiales were found as symbionts (with “symbiosis” simply meaning the even-temporary association of organisms from different species) in a variety of other non-vector eukaryotic hosts, both from terrestrial and aquatic environments (reviewed in Perlman, Hunter & Zchori-Fein, 2006; Weinert et al., 2009; Gillespie et al., 2012; Castelli, Sassera & Petroni, 2016). Numerous such novel bacterial species were retrieved in aquatic protists (Kawafune et al., 2015; Schulz et al., 2016; Yang, Narechania & Kim, 2016; Hess, 2017; Yurchenko et al., 2018), including, notably, parasitic (Sun et al., 2009; Zaila et al., 2017) and various free-living ciliates (Ferrantini et al., 2009; Boscaro et al., 2013; Schrallhammer et al., 2013; Vannini et al., 2014; Senra et al., 2016; Szokoli et al., 2016a; Szokoli et al., 2016b; Castelli et al., 2019b; Lanzoni et al., 2019).
Such findings strongly suggest the ability of those “aquatic” Rickettsiales to perform horizontal transmission including host species shift, although mostly from indirect evidence, i.e., closely related bacteria in hosts as different as ciliates, hydra (Fraune & Bosch, 2007), corals (Sunagawa et al., 2009), ring worms (Murakami et al., 2017), ascidians (Kwan & Schmidt, 2013; Dishaw et al., 2014), placozoans (Driscoll et al., 2013), as well as incongruent host and symbionts phylogenies (Perlman, Hunter & Zchori-Fein, 2006; Epis et al., 2008; Driscoll et al., 2013). In few cases, experimental interspecific transfer between unicellular hosts, although no stable infection was documented (Schulz et al., 2016; Senra et al., 2016).
In most cases the relationships between Rickettsiales associated with aquatic eukaryotes and their hosts were not clarified in detail (Castelli, Sassera & Petroni, 2016), with the exception of “Candidatus (Ca.) Xenohaliotis californiensis”, which is considered the cause of the withering syndrome in its abalone hosts (Friedman et al., 2000; Crosson et al., 2014). It has been hypothesized by different authors that bacteria harbored by aquatic protists might constitute etiological agents of possible Vector Borne Diseases (VBD) of aquatic animals (Barker & Brown, 1994; Taylor-Mulneix et al., 2017). These include the numerous cases of epidemics caused by Rickettsia-Like Organisms (RLOs; i.e., intracellular bacteria morphologically similar to Rickettsia) determining an increasing number of massive deaths in intensive aquaculture facilities during the last years (Gollas-Galván et al., 2014), including mollusks (Friedman et al., 2000; Sun & Wu, 2004; Ross et al., 2018), crustaceans (Loy et al., 1996; Longshaw, 2011; Wang, 2011) and fishes (Athanassopoulou et al., 2004; Corbeil, Hyatt & Crane, 2005; Timur et al., 2013). Although many RLOs were actually shown to be phylogenetically unrelated to Rickettsiales (e.g., Gammaproteobacteria) (Fryer et al., 1992; Tan & Owens, 2000; Longshaw, 2011), at least in one case available data indicate a probable connection between a truly Rickettsiales bacterium and fish disease. In detail, a “Ca. Midichloria mitochondrii”-related bacterium was linked to the red-mark syndrome in rainbow trout (Lloyd et al., 2008; Cafiso et al., 2015). Most importantly, although a transmission route was not directly proven, the same bacterium was found in association with the ciliate Ichthyophthirius multifiliis (Zaila et al., 2017), which is indeed a fish parasite. Interestingly, even other Rickettsiales symbionts (“Ca. Megaira” genus) can be found in the same ciliate host species, which might suggest a potential transmission route for these symbionts as well (Sun et al., 2009; Zaila et al., 2017).
Taking into account such premises, in order to investigate the hypothesis that protists can act as natural reservoir for potentially pathogenic bacteria (Barker & Brown, 1994; Gao, Harb & Abu Kwaik, 1997; Görtz & Brigge, 1998; Harb, Gao & Abu Kwaik, 2000; Horn & Wagner, 2004; Molmeret et al., 2005; Ferrantini et al., 2009; Taylor-Mulneix et al., 2017), we experimentally tested the possibility that endosymbionts of ciliates could be per os transferred to aquatic Metazoa, such as planarians. The Rickettsiales “Ca. Trichorickettsia” was chosen as candidate for transfection experiments as it shows a broad ciliate host range, infecting different cell compartment (i.e., cytoplasm and nucleus) of multiple ciliates (Vannini et al., 2014; Sabaneyeva et al., 2018). Additionally, in many hosts, it is covered by long flagella and is able to actively move (Vannini et al., 2014; Sabaneyeva et al., 2018). Thus, for our research purpose, as donor host in transfection experiments, we employed the P. multimicronucleatum strain US_Bl 16I1, which was herein newly characterized and shown to bear a flagellated “Ca. Trichorickettsia mobilis” symbiont. On the other side, we selected the freshwater planarian Dugesia japonica because it is a benthic organism, living in mud and under rocks in ponds and streams. Planarians are zoophages or carnivorous animals, but ingest also detritus, fungi, and bacteria (González-Estévez, 2009). Thus, they may encounter in their environment a large variety of microbes (Petersen, 2014), including ciliates possibly hosting endosymbionts. Planarians have always been considered an important model for studying stem cells and regeneration (Rossi et al., 2008), but recently they became also important for studying the natural immunity system of Metazoa (Abnave et al., 2014; Conti, Abnave & Ghigo, 2014; Torre & Ghigo, 2015; Arnold et al., 2016; Torre et al., 2017; Tsoumtsa et al., 2017). Based on all these considerations, they appeared a suitable model for such experiments.
Bacterial endosymbiont transfer from P. multimicronucleatum to planarians was investigated by checking for the presence of “Ca. Trichorickettsia mobilis” in tissues of ciliate-fed planarians by means of PCR and Transmission Electron Microscopy (TEM) at day 1, 3, and 7 after feeding. The collected data are indicative of the ability of the endosymbiont to temporarily survive and divide in the planarian intestine, and to possibly escape from planarian phagosomes.
Materials & Methods
Ciliate host isolation, culturing, and identification
Paramecium multimicronucleatum monoclonal strain US_Bl 16I1 was established from a cell isolated from a freshwater sample collected from the Yellowwood Lake (39°11′29,0″N, 86°20′31,4″W), IN, USA and cultivated in San Benedetto mineral water (San Benedetto S. p. A., Italy).
The strain was maintained in the laboratory in an incubator at a temperature of 19 ± 1 °C and on a 12:12 h light/dark cycle (light source: NATURAL L36W/76 and FLORA L36W/77 neon tubes, OSRAM, Berlin, Germany). Instead of using bacteria as common food source for paramecia (Krenek, Berendonk & Petzoldt, 2011), cells were fed monoclonal cultures of flagellated green algae, i.e., Chlorogonium sp. (freshwater) or, alternatively, Dunaliella tertiolecta (brackish water, 1‰ of salinity) to minimize bacterial load in cell cultures. They were fed two to three times per week to obtain a mass culture (1.5 L) (Castelli et al., 2015) suitable for transfection experiments. Living ciliates were observed with a Leitz (Weitzlar, Germany) microscope equipped with Differential Interference Contrast (DIC) at a magnification of 100–1250× for general morphology and swimming behavior according to Modeo et al. (2013a) and Nitla et al., (2019). DIC observation of ciliates was also aimed at checking for macronuclear endosymbiont motility. Feulgen-stained ciliates were observed to obtain information on the nuclear apparatus. Microscope images were captured with a digital camera (Canon PowerShot S45). Cell measurements were performed using ImageJ (https://imagej.nih.gov/ij/). Morphological identification was conducted according to literature data (Fokin, 2010/2011).
Characterization of ciliate and endosymbiont
Characterization of the ciliate host and its bacterial endosymbiont was performed through the “full cycle rRNA approach” (Amann, Ludwig & Schleifer, 1995), i.e., through the nuclear 18S rRNA gene (generally considered the preferred marker to study molecular taxonomy and phylogeny of eukaryotic organisms—just as an example see Illa et al., 2019), ITS region and partial 28S gene (ciliate), and 16S rRNA gene (endosymbiont) sequencing, combined with Fluorescence In Situ Hybridization (FISH) experiments. Additionally, mitochondrial cytochrome c oxidase subunit I (COI) gene was sequenced as well, to display haplogroup variation in P. multimicronucleatum (Barth et al., 2006).
Molecular characterization
For molecular characterization of both ciliate host and bacterial endosymbiont, total DNA was extracted from approximately 50 Paramecium cells as described in Szokoli et al. (2016a) and Szokoli et al. (2016b). All PCR reactions were carried out with the Takara ExTaq (Takara, Kusatsu, Japan) reaction kit. COI gene was amplified using the forward primer F338dT and reverse primer R1184dT (Strüder-Kypke & Lynn, 2010), while for 18S rRNA gene amplification the forward 18S F9 Euk (Medlin et al., 1988) and the reverse 18S R1513 Hypo primers (Petroni et al., 2002) were used. The ITS region (including partial 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2, and partial 28S rRNA gene) sequence was obtained by PCR with the forward 18S F919 and the reverse 28S R671 primers (modified according to Boscaro et al., 2012). The 16S rRNA gene of the bacterial symbiont was amplified with the Alphaproteobacteria-specific forward primer 16S Alpha F19b and the universal bacterial reverse primer 16S R1522a (Vannini et al., 2004). PCR products were purified by NucleoSpin gel and PCR cleanup kit (Macherey-Nagel GmbH Düren, Germany) and directly sequenced at GATC Biotech AG (Constance, Germany). Internal primers were used to sequence 16S rRNA gene (Vannini et al., 2004), 18S rRNA gene (Rosati et al., 2004) and ITS regions (Boscaro et al., 2012). The latter two sequences were then joined together, exploiting the partial overlap on the 18S rRNA gene sequences. For the COI gene sequencing, amplification primers were employed from both directions.
FISH experiments
Preliminary FISH experiments were performed using the bacterial universal probe EUB338 (Amann et al., 1990) labeled with fluorescein-isothiocyanate (FITC) in combination with the specifically designed probe Rick_697 (5′-TGTTCCTCCTAATATCTAAGAA-3′) labeled with Cy3 to verify the presence of endosymbiotic bacteria belonging to the family Rickettsiaceae (Vannini et al., 2014). Based on the obtained results, i.e., the presence of a single, coincident positive signal in the ciliate macronucleus and the newly characterized 16S rRNA gene sequence corresponding to “Ca. Trichorickettsia mobilis”, a second FISH experiment was carried out using a species-specific probe, i.e., the probe Trichorick_142 (5′-GTTTCCAAATGTTATTCCATAC-3′) (Vannini et al., 2014) in combination with the bacterial universal probe EUB338 (Amann et al., 1990). The experiments followed the protocol by Fokin et al. (2019) employing a hybridization buffer containing no formamide, according to the recommendations for the used probes. Slides were mounted with SlowFade Gold Antifade with DAPI (Invitrogen, Carlsbad, USA) and viewed with a Zeiss AxioPlan fluorescence microscope (Carl Zeiss, Oberkochen, Germany) equipped with an HBO 100W/2 mercuric vapor lamp. Digital images were captured by means of a dedicated software (ACT2U, version 1.0).
Planarian culturing
Planarians used in this work belonged to the species Dugesia japonica, asexual strain GI (Rossi et al., 2018). Animals were kept in a solution of: CaCl2 2.5 mM, MgSO4 0.4 mM, NaHCO3 0.8 mM, and KCl 77µM, hereafter referred to as culturing water, at 18 °C in dim light conditions and fed with chicken liver (purchased from local food stores) prepared according to King & Newmark (2013), once a week. Non-regenerating specimens, within 5–8 mm of length, were used for all experimental procedures, after being starved for about two weeks.
Transmission electron microscopy (TEM)
TEM preparations were obtained both for P. multimicronucleatum cells and planarians to study the ultrastructure of the ciliate-associated endosymbiont “Ca. Trichorickettsia mobilis” and to verify the success of transfection experiments (i.e., to check the animals for the presence of transferred ciliate endosymbionts in their tissues with particular attention to bacteria showing possible signs of division) respectively.
TEM preparations of P. multimicronucleatum cells were obtained according to Modeo et al. (2013b). Briefly, cells were fixed in a 1:1 mixture of 2.5% (v/v) glutaraldehyde in 0.1 M cacodylate buffer (pH 7.4), and 2% (w/v) OsO4 in distilled water, ethanol and acetone dehydrated, and embedded in Epon-Araldite mixture. Ultrathin sections were stained with 4% (w/v) uranyl acetate followed by 0.2% (w/v) lead citrate.
TEM preparations of planarian specimens were obtained as previously described (Rossi et al., 2014; Cassella et al., 2017), with minor modifications. Planarians were fixed with 3% glutaraldehyde in 0.1 M cacodylate buffer, and post-fixed with 2% osmium tetroxide for 2 h. After ethanol dehydration, samples were embedded in Epon-Araldite mixture. Ultrathin sections were stained with uranyl acetate and lead citrate and observed with a Jeol 100 SX Transmission Electron Microscope.
Negative staining
For negative staining, several P. multimicronucleatum cells of the strain US_Bl 16I1 were washed in distilled water and squashed; a drop of the resulting suspension was placed on a grid. Bacteria were allowed to settle for 2–3 min, then a drop of 1% uranyl acetate in distilled water was added for no longer than 1 min. The liquid was then absorbed with filter paper, the grid was air-dried, and observed under TEM (adapted from Sabaneyeva et al., 2018).
Transfection experiments
Two transfection experiments with the same protocol were sequentially carried out over a period of four months. Each experiment was conducted by treating a fixed number of planarians with ciliate-enriched food, i.e., liver paste mixed with P. multimicronucleatum homogenate; from now on these animals will be referred to as “treated planarians”. As for experimental control, planarians fed plain liver paste were used; these animals will be referred to as “control planarians”. Each transfection experiment was performed according to the following protocol:
-
1.
96 planarians were selected by eye from culture (see above), washed in fresh culturing water and collected in a Petri dish with 50 µg/ml gentamicin sulfate (Sigma, Saint Luis, MO, USA) dissolved in their culturing water (see above). This preliminary antibiotic treatment was performed to minimize potential endogenous bacteria contaminants and was not harmful to the animals (Roberts-Galbraith, Brubacher & Newmark, 2016). Planarians were left in antibiotic treatment for 24 h under regular culturing conditions concerning temperature and light (see above) and kept under visual control during that period to verify their viability during the antibiotic treatment and by the time of transfer. Then, planarians were washed six times in their fresh culturing water, and split into two equal groups of 48 individuals each and accommodated in two different Petri dishes for the transfection procedure by means of feeding.
-
2.
In order to get a bacterial load consistent with previous experiments (Abnave et al., 2014), 1.5 L of P. multimicronucleatum mass culture (cell concentration: ∼4 × 105 cell/L) were selected. Indeed, considering that FISH experiments indicated roughly 100 endosymbionts/cell (see Results section), in total ciliates were estimated to contain ∼6 × 107 symbiont cells. P. multimicronucleatum cells were filtered with a nylon filter (pore size: 100 µm), washed twice in sterile San Benedetto mineral water to minimize potential bacterial contaminants, concentrated and harvested by means of centrifugation (400× g per 10 min), so to reduce the medium volume to 2 ml. Then, cells were mechanically squashed and homogenized by syringing (syringe needle: 22GA, 0.70 mm in diameter) for 20 min at room temperature (adapted from Vannini et al., 2017). Cell homogenate was checked under the stereomicroscope so to exclude the presence of intact Paramecium cells, centrifuged (10,000× g per 10 min), and the resulting pellet was resuspended in 50 µl of planarian food (homogenized liver paste) by direct resuspension.
-
3.
A group of 48 planarians was fed P. multimicronucleatum-enriched liver paste (treated planarians), while the other group was in parallel fed plain liver paste (control planarians). For feeding, food was spread over the bottom of the Petri dish. Animals were allowed to feed freely for a period of 2 h under regular culturing conditions (see above). Attention was paid to planarian survival and feeding behavior during this period. Two washing steps were then carried out removing the medium and adding fresh planarian culturing water. Finally, the two groups of animals were left in their fresh culturing water and in regular cultivation conditions in the two Petri dishes until the collection of specimens for the next TEM and PCR analyses at the three timepoints (see below) of the experiments. Animals were kept under visual control throughout the experiment to regularly check their viability.
-
4.
At day 1, 3, and 7 after feeding (experimental timepoints), from each of the two Petri dishes containing treated and control planarians a group consisting of 16 animals were sampled: 4 animals were fixed and processed for TEM, and 12 were immediately frozen and stored at −80 °C, and dedicated to DNA extraction (4 animals per each sample).
PCR verification of transfer success
Treated and control planarians were processed for genomic DNA extraction, purification, and PCR amplification with endosymbiont-specific primers. For each experimental condition (treated and control planarians) and each experimental timepoint (1, 3, and 7 days after feeding), genomic DNA was extracted from frozen samples by using the Wizard Genomics DNA purification kit (Promega, Madison, WI, USA). One microliter of purified DNA was analysed by gel electrophoresis to check for integrity. DNA was quantified using a Nanodrop spectrophotometer. For each experimental timepoint of both experimental conditions (i.e., treated and control planarians), similar amounts of genomic DNA were used for amplification using the ampli-Taq-gold DNA polymerase (Applied Biosystems, Foster City, CA, USA) and for a nested PCR assay. The first primer pair used, specific for “Ca. Trichorickettsia mobilis”, was RickFla_ F69 5′-GTTAACTTAGGGCTTGCTC-3′and Rick_R1455 5′-CCGTGGTTGGCTGCCT-3′ (Vannini et al., 2014); PCR conditions were: 3 min at 94 °C; 5 cycles consisting of 30 s at 94 °C, 30 s at 58 °C, 2 min at 72 °C each; 10 cycles consisting of 30 s at 94 °C, 30 s at 54 °C, 2 min at 72 °C each; 25 cycles consisting of 30 s at 94 °C, 30 s at 50 °C, 2 min at 72 °C each; ending with 7 min at 72 °C. The second, nested primer pair used was RickFla_F87 5′-CTCTAGGTTAATCAGTAGCAA-3′and Rick_R1270 5′-TTTTAGGGATTTGCTCCACG-3′ (Vannini et al., 2014). For nested PCR assay, one microliter of PCR product of the first PCR assay was used as a template; PCR conditions were as above.
For each experimental condition and each timepoint of the two transfection experiments, the DNA amplification was performed in duplicate. Samples were considered positive if a single band of the expected size was recorded after nested amplification. Sequencing of amplicons was carried out to confirm the presence of “Ca. Trichorickettsia mobilis” using the primer RickFla_F87 (see above) and Sanger sequencing (BMR Genomics, Padova, Italy).
Samples obtained from planarians fed with plain liver paste were used as control. As positive control, genomic DNA purified from P. multimicronucleatum monoclonal strain US_Bl 16I1 was processed.
Results
Host morphological and molecular identification
Ciliate strain US_Bl 16I1 (Figs. 1A, 1B; Figs. S1 and S2 in Article S1) was confirmed in morphological inspections as Paramecium multimicronucleatum Powers & Mitchell, 1910 considering features of key-characters such as cell size, number and features of micronuclei (mi), and number and features of contractile vacuoles (Figs. S1 and S2 in Article S1), as described in previous literature (Wichterman, 1986; Allen, 1988; Fokin, 1997; Fokin & Chivilev, 2000; Fokin, 2010/2011). The molecular analysis of the combined (partial) 18S rRNA-ITS1-5S rRNA gene-ITS2-(partial) 28S rRNA gene sequence (2,792 bp, GenBank accession number: MK595741) confirmed the species assignation by morphological identification with a 100% sequence identity with the sequences of other P. multimicronucleatum strains already present in GenBank presenting either 18S rRNA gene portion only (AB252006 and AF255361), or the ITS portion (AY833383, KF287719, JF741240 and JF741241). COI gene sequence identity of strain US_Bl 16I1 (760 bp, accession number: MK806287) is highest with another P. multimicronucleatum haplotype (FJ905144.1; 96.3%).
Figure 1. Results of FISH experiments on a P. multimicronucleatum US_Bl 16I1 cell.

(A) Bright field microscopy (Ma, macronucleus). (B) After DAPI staining. (C) Treated with the almost universal bacterial probe EUB338 double labelled with fluorescein (green) (probe labelling both at 5′ and 3′ ends). (D) Treated with species-specific probe Trichorick_142 labelled with Cy3 (red) targeting “Ca. Trichorickettsia mobilis”. (E) Merge of C and D. The number of endosymbionts targeted by the species-specific probe in the macronucleus reached ∼100 (D, E). Scale bars stand for 10 µm.
Endosymbiont identification and features
The 16S rRNA gene amplification was performed to identify endosymbionts associated with P. multimicronuclatum US_Bl 16I1 strain (sequence length 1,563 bp, accession number: MK598854), which allowed to assign it to “Ca. Trichorickettsia mobilis”. Specifically, among the three subspecies previously described, it was more similar to “Ca. Trichorickettsia mobilis subsp. hyperinfectiva”, an endosymbiont described in the cytoplasm of Paramecium calkinsi (99.8% identity of the novel sequence with type strain MF039744.1) (Sabaneyeva et al., 2018). FISH experiments (Figs. 1C–1E) performed by using bacterial universal probe EUB338 (Fig. 1C) and the species-specific probe Trichorick_142 (Fig. 1D) confirmed this result and indicated that, contrarily to the P. calkinsi symbiont, the bacterium was localized in the ciliate macronucleus (roughly with a presence of about 100 endosymbionts per cell). Additionally, the full overlapping of bacterial universal probe EUB338 and “Ca. Trichorickettsia”-specific probe signals indicated that this symbiont constituted the total set of intracellular bacteria in host P. multimicronucleatum US_Bl 16I1 cells (Fig. 1E, merge).
In TEM-processed ciliate cells, the endosymbionts were confirmed to be hosted in the macronucleus; they showed a two-membrane cell wall typical of Gram-negative bacteria and appeared rod-shaped, with rounded to narrower ends (Fig. 2). They were surrounded by a clear halo and not encircled by any symbiosomal vacuole, i.e., they appeared in direct contact with host nuclear material. Their size was ∼1.2–2.1 × 0.5–0.6 µm, and they were scattered throughout the macronucleus of the ciliate (Figs. 2A–2D). Sometimes in their cytoplasm several electron-lucid “holes” (diameter: ∼0.2 µm), were observed (Fig. 2A). Bacterial cytoplasm was electrondense; no other structures were visible except for abundant ribosomes (Fig. 2B). Endosymbionts displayed thin (diameter: ∼9 nm) and short flagella distributed all around the cell (Figs. 2A, 2C–2D) which sometimes formed a putative tail emerging from a cell end (Fig. 2D). The presence of longer flagella was evident after negative staining procedure (Fig. 3): besides some short flagella occurring all around the cell, at least a few longer and thicker flagella (∼2000 × 12 nm) arose from one of cell ends.
Figure 2. TEM pictures showing the morphological-ultrastructural details of “Ca. Trichorickettsia mobilis” in the macronucleus of P. multimicronucleatum US_Bl 16I1.

(A) Longitudinally and transversely sectioned endosymbionts inside macronucleus (Ma) with a double membrane and surrounded by a clear halo; electron-lucid roundish areas occur in the cytoplasm of some endosymbionts. (B) Numerous bacterial ribosomes are visible. (C, D) Bacterial flagella are short, located either around (arrow) the endosymbiont cell or at a cell pole, where they can form a putative tail (arrowhead). Scale bars stand for 0.5 µm.
Figure 3. Negative contrast at TEM of a specimen of “Ca. Trichorickettsia mobilis”.

Some short flagella are present and at least two longer flagella (arrow) arise from one of cell ends. Scale bar stands for 1 µm.
Some active bacterial motility, likely obtained by means of their flagella, was recorded inside the macronucleus of intact P. multimicronucleatum cells during observation under DIC microscope at 1,000×. Endosymbionts were seen to move through the chromatin bodies covering short distances; additionally, after ciliate cell squashing, the released bacteria were still capable of movement (L Modeo, pers. obs., 2017).
Transfection experiments
To verify RLO transfer to planarians, we fed animals either with ciliate-enriched liver paste (treated planarians) or plain liver paste (control planarians). 100% survival of the planarians (control and treated animals) was confirmed throughout the transfection experiments; in particular, the survival of treated planarians was not affected by the treatment and the animals did not show any morphological or behavioral alteration. The success of each of the two independent transfection experiments performed was investigated by means of PCR experiments and TEM observation on treated and control planarians at the different experimental timepoints, according to the described experimental procedure.
The DNA of “Ca. Trichorickettsia mobilis” was detected at all experimental timepoints (day 1, 3, and 7 after feeding) in genomic DNA preparations obtained from all investigated planarians fed with P. multimicronucleatum lysate-enriched liver paste. The size of the nested amplicon (1,360 bp) obtained from treated planarian samples matched the size of the amplicon obtained in DNA purified from ciliate monoclonal strain US_Bl 16I1 (positive control) (Fig. 4). The sequencing of the obtained amplicons confirmed the specificity for “Ca. Trichorickettsia mobilis” (100% sequence identity with the sequenced obtained from the symbiont). No amplification product was obtained in genomic DNA samples from control planarians fed with plain liver paste (negative control) (Fig. 4).
Figure 4. Detection of “Ca. Trichorickettsia mobilis” DNA in treated and control planarians by PCR.
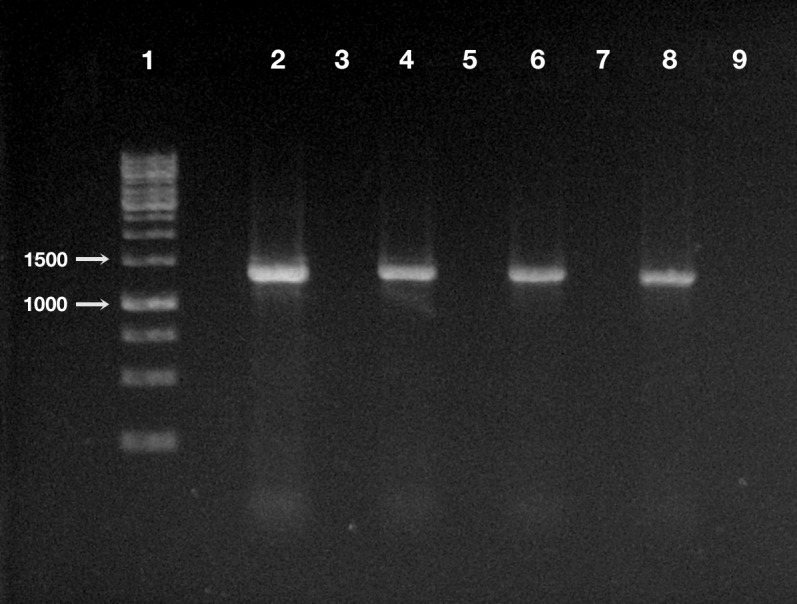
Line 1, ladder (Gene Ruler 1 Kb DNA ladder Fermentas, Waltham, Massachusetts, USA); line 2, positive control (i.e., DNA extracted from P. multimicronucleatum US_Bl 16I1); line 3, negative control (i.e., no DNA); line 4, treated planarian at day 1 after feeding (a.f.); line 5, control planarian at day 1 a.f.; line 6, treated planarian at day 3 a.f.; line 7, control planarian at day 3 a.f.; line 8, treated planarian at day 7 a.f.; line 9, control planarian at day 7 a.f.
Ultrastructural observation was conducted on all TEM-processed specimens per each timepoints of the experiments, for both experimental groups, i.e., treated and control planarians.
In tissues collected from all the investigated planarians fed with Paramecium US_Bl 16I1 lysate-enriched liver paste, the presence of bacteria with morphology and sizes (see below for details) fitting with those of ciliate endosymbionts within intestinal cells was detected at all timepoints of the two experiments (Figs. 5 and 6).
Figure 5. TEM pictures of the intestine of antibiotic-treated D. japonica fed on liver paste enriched with the homogenate of P. multimicronucleatum US_Bl 16I1 cells and fixed at day 1 after feeding.

(A) A view through planarian intestine where several bacteria are visible (arrows). (B) Enlargement of a particular of (A) to show a dividing bacterium with flagella arising from its pole (arrowhead). (C) A dividing bacterium (arrow) in planarian intestine (D) Extruded trichocysts (t) (extrusive organelles) of Paramecium detected in planarian intestine along with bacteria, also dividing (arrow). (E) A longitudinally sectioned bacterium (arrow) free in the cytoplasm of an intestinal cell of a treated planaria. (F) Enlargement of a particular of (E). Arrowhead, flagella. Scale bars stand for 1 µm.
Figure 6. TEM pictures of the intestine of antibiotic-treated D. japonica fed on liver paste enriched with the homogenate of P. multimicronucleatum US_Bl 16I1 cells and fixed at day 3 and 7 after feeding.

(A–D) day 3 after feeding, (E–F) day 7 after feeding. (A, B) A bacterium (arrow) in proximity with the border of a phagosome, in the cytoplasm of a planarian intestinal cell. Ld, lipidic droplet. (C) A dividing bacterium (arrow) next to a phagosome. (D) Enlargement of a particular of (C). (E) Bacteria (arrow) inside phagosome and (F) free in the cytoplasm. Scale bars stand for 1 µm.
At day 1 after feeding, several bacteria were recognizable in planarian enterocytes. Some of them appeared to be in division (Figs. 5A–5D). In several cases, the membrane of bacteria-including phagosomes was damaged and interrupted, and bacteria showing a two-membrane cell wall and a surrounding clear halo were detected free in the cytoplasm of planarian intestinal cells (i.e., in direct contact with their cytoplasm) (Figs. 5E, 5F). Flagella (diameter: ∼9 nm) could also be detected on bacterial surface (Figs. 5B, 5F). Additionally, extruded trichocysts (extrusive organelles typical of Paramecium), presumably included in Paramecium homogenate and ingested by treated planarians, were easily recognizable inside intestinal cell phagosomes (Fig. 5D).
A similar scenario was observed at day 3 after feeding (Figs. 6A–6D), with several bacteria in apparent division as well (Figs. 6C, 6D). At day 7 after feeding, a few bacteria (i.e., roughly less bacteria than the several observed after 1 and 3 days) were still well visible in planarian enterocytes (Fig. 6E), also free in the cytoplasm (Fig. 6F).
No bacteria were ever observed in tissues other than intestine in treated planarians; similarly, no bacteria were observed in tissues of TEM-processed control animals (Fig. S3).
Discussion
A “Ca. Trichorickettsia mobilis” endosymbiont, closely related to “Ca. Trichorickettsia mobilis subsp. hyperinfectiva” previously described in the cytoplasm of P. calkinsi (Sabaneyeva et al., 2018), was retrieved in the macronucleus of the ciliate P. multimicronucleatum strain US_Bl 16I1. “Ca. Trichorickettsia mobilis” up to now has been exclusively found as an endosymbiont of ciliates belonging to the genera Paramecium and Euplotes (Vannini et al., 2014; Sabaneyeva et al., 2018). In the light of the present findings, a comparison among all strains characterized to date suggesting a certain morphological plasticity of this bacterium, is presented in Article S1.
The aim of the present research was to obtain a first indication on the potential transfer of the Rickettsia-related macronuclear endosymbiont “Ca. Trichorickettsia mobilis” from the ciliate P. multimicronucleatum strain US_Bl 16I1 to the metazoan model D. japonica. There are several studies reporting on the host/symbiont relationships of different Paramecium species with different Rickettsiales; P. multimicronucleatum lies in this ciliate selection, and is a rather a common species, sharing the freshwater habitat with planarians. Thus, it was chosen as donor in transfection experimental context as, in our opinion, it can be considered a suitable candidate as putative environmental vector for RLOs. As we dealt with endosymbionts, which are present in limited numbers inside their ciliate host, we chose to treat the planarians with cell mass culture homogenate instead of adding living ciliates to planarians food. This allowed processing of as many ciliates as possible to maximize the probability of endosymbiont ingestion by the animals, thereby increasing the chance of detection of successful transfer via PCR and TEM-based approaches.
According to the present findings, the transfection experiments were successful, i.e., they showed the capability of “Ca. Trichorickettsia mobilis” to enter planarian tissues. Indeed, in the intestine of planarians previously antibiotic-treated to avoid bacterial contamination and fed with liver paste enriched with pellet of ciliate homogenate (rich in “Ca. Trichorickettsia” symbionts), “Ca. Trichorickettsia” bacterial cells in apparent division were observed up to day 3 after feeding. We observed several bacteria free from phagosomal membrane, i.e., in direct contact with enterocyte cytoplasm (Figs. 5 and 6). The presence of bacteria with a morphology and a size fully resembling those of the RLO endosymbiont of P. multimicronucleatum US_Bl 16I1 were observed up to day 7 after feeding inside and outside planarian phagosomes (Fig. 6). Based on our findings, we hypothesize that dividing bacteria might also be present in treated planarian parenchyma after day 3.
On the contrary, with respect to treated animals, in TEM preparations of controls (i.e., antibiotic-treated planarians fed with plain liver paste) no bacteria were found (Fig. S3).
These results were confirmed and supported by PCR analysis and sequencing of obtained amplicons: the DNA of “Ca. Trichorickettsia mobilis” was recovered in treated planarians up to day 7 after feeding while in control animals no RLO DNA was ever amplified and detected (Fig. 4). On the other side, survival of treated planarians was not affected by the treatment and animals did not manifest morphological or behavioral alterations.
Our findings are in line with those by Gong et al. (2016). These authors studied the phylogenetic identities of digestion-resistant bacteria that could survive starvation and form relatively stable associations with some marine and freshwater ciliate species, and demonstrated that the classes Alphaproteobacteria (which includes the order Rickettsiales) and Gammaproteobacteria are prevalent as digestion-resistant bacteria; from this study a putative significant role of secretion systems in promoting marine protist-bacteria associations resulted as well.
In our experiments, after being ingested by planarians, the bacteria were observed enclosed inside phagosomes of intestinal cells. This also occurred in previous experiments investigating the resistance of planarians to infection by bacterial strains pathogenic for humans and other metazoans (Abnave et al., 2014). In that research, planarians could eliminate most of the phagocytized bacterial strains within 3–6 days post-feeding thanks to 18 resistance genes, such as MORN2, so the authors suggested that planarians can be considered a model to identify innate resistance mechanisms. Under this perspective, the evidence we obtained that the “Ca. Trichorickettsia” endosymbionts of P. multimicronucleatum US_Bl 16I1 are still detectable in planarian intestine enterocytes inside and outside phagosomes up to day 7 after feeding is a good indication that the bacteria are able to survive, at least temporarily, within this time-span. This is reinforced also considering that the bacterial morphology in the planarians is comparable to that of endosymbionts in their original localization inside the macronucleus of P. multimicronucleatum (Figs. 2, 5 and 6), thus not evidencing any lethal alteration. Along this line of thought, we hypothesize that “Ca. Trichorickettsia mobilis” might be capable to avoid typical defense mechanisms exploited by planarians entering planarian tissues and transiently surviving within. Interestingly, similarly to some Gammaproteobacteria such as Rheinheimera sp. strain EpRS3 (Chromatiaceae), capable of escaping from phagosomes of the ciliate Euplotes aediculatus when fed the bacterium plus its culture medium (Chiellini et al., 2019), Rickettsiaceae are already known for their ability to escape the host vacuolar membrane, residing freely in the host cytoplasm, where they may exploit host cytoskeleton for movement (Renesto et al., 2003; Gouin et al., 2004; Walker & Ismail, 2008; Cardwell & Martinez, 2009; Ge & Rikihisa, 2011; Rennoll-Bankert et al., 2016).
In the past, efforts have been put to experimentally verify the transfer of morphologically RLOs among aquatic organisms. For example, Nunan et al. (2003) performed bioassays to verify the transfer of the infection between two species of commercially farmed shrimps, i.e., the infected Penaeus monodon and the specific pathogen-free Penaeus vannamei, with the aim of investigating the suspected causative agent of severe mortality in farms where those organisms are in co-culture (grow-out ponds). In that case, only injection bioassays were successful leading to an infection, while per os infection failed. Among different possible reasons for this negative result those authors cited the potential need for a vector to disseminate the disease. According to our findings, ciliates could be seen as suitable vectors in this kind of situation.
Conclusions
To the best of our knowledge, this is the first time that a set of experimental bioassays was performed to investigate the possible transfer of a “true” and ascertained Rickettsiales bacterium from an infected protist to an uninfected metazoan of the same aquatic environment (freshwater). We believe that our findings, indicating, at least transiently, an effective transfer of ciliate endosymbionts to the intestinal cells of planarian, can offer intriguing insights concerning the diseases caused by Rickettsiales or RLOs occurring in fish farms or in the wild. Indeed, our study reinforces the notion that these might have ciliates or other protists as putative vectors. Although further investigations on this topic are necessary to expand its implications, we think that our study may represent the basis for conceiving long-lasting experiments aiming to better understand whether “Ca. Trichorickettsia mobilis”, as well as other Rickettsiales symbionts of protists, can be able to survive longer in tissues of planarians and other aquatic Metazoa, and whether these RLOs may have some impact on the recipient host health.
Supplemental Information
COI gene and combined (partial) 18S rRNA-ITS1-5S rRNA gene-ITS2-(partial) 28S rRNA sequences of Paramecium multimicronucleatum strain US_Bl 16I1: GenBank accession numbers MK595741 and MK806287 respectively. 16S rRNA gene sequence of “Ca. Trichorickettsia mobilis”: Gen Bank accession number MK598854.
(A) Day 1 after feeding. (B) Day 7 after feeding. L, lumen of planarian intestine. No bacteria were observed in tissues of investigated control animals in none of the experimental timepoints. Scale bars stand for 1µm.
{kind=link}
Acknowledgments
The authors wish to thank Francesco Paolo Frontini, Fabrizio Erra, and Federica Vantaggio for ciliate culturing and in vivo processing; Claudio Ghezzani and Simone Gabrielli for assistance with TEM material; Simone Gabrielli for help with graphic artwork, Thomas Berendonk for the opportunity to collaborate on Paramecium. All authors critically read and approved the manuscript.
Funding Statement
This work received financial support from the following projects: Progetti di Ricerca di Ateneo (PRA), project PRA_ 2016_58, University of Pisa to Giulio Petroni; Russian Foundation for Basic Research (RFBR), grant No 18-04-00562-a to Elena Sabaneyeva. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Contributor Information
Letizia Modeo, Email: letizia.modeo@unipi.it.
Alessandra Salvetti, Email: alessandra.salvetti@unipi.it.
Additional Information and Declarations
Competing Interests
The authors declare there are no competing interests.
Author Contributions
Letizia Modeo, Alessandra Salvetti and Leonardo Rossi conceived and designed the experiments, performed the experiments, analyzed the data, prepared figures and/or tables, authored or reviewed drafts of the paper, and approved the final draft.
Michele Castelli and Franziska Szokoli performed the experiments, analyzed the data, authored or reviewed drafts of the paper, and approved the final draft.
Sascha Krenek, Valentina Serra, Elena Sabaneyeva, Graziano Di Giuseppe, Sergei I. Fokin and Franco Verni analyzed the data, authored or reviewed drafts of the paper, and approved the final draft.
Giulio Petroni conceived and designed the experiments, analyzed the data, authored or reviewed drafts of the paper, and approved the final draft.
DNA Deposition
The following information was supplied regarding the deposition of DNA sequences:
All the sequences are available in the Supplemental Files and at Genbank:
- Combined (partial) 18S rRNA-ITS1-5S rRNA gene-ITS2-(partial) 28S rRNA gene sequence of Paramecium multimicronucleatum strain US_Bl 16I1: MK595741;
- COI gene sequence identity of strain US_Bl 16I1: MK806287;
- 16S rRNA gene sequence of the Paramecium multimicronucleatum US_Bl 16I1 endosymbiont “Ca. Trichorickettsia mobilis” US_Bl 16I1 strain: MK598854.
Data Availability
The following information was supplied regarding data availability:
The raw data are available in the Supplemental Files.
References
- Abnave et al. (2014).Abnave P, Mottola G, Gimenez G, Boucherit N, Trouplin V, Torre C, Conti F, Ben Amara A, Lepolard C, Djian B, Hamaoui D, Mettouchi A, Kumar A, Pagnotta S, Bonatti S, Lepidi H, Salvetti A, Abi-Rached L, Lemichez E, Mege J-L, Ghigo E. Screening in planarians identifies MORN2 as a key component in LC3-associated phagocytosis and resistance to bacterial infection. Cell Host & Microbe. 2014;16:338–350. doi: 10.1016/j.chom.2014.08.002. [DOI] [PubMed] [Google Scholar]
- Allen (1988).Allen RD. Cytology. In: Görtz H-D, editor. Paramecium. Springer-Verlag; Berlin: 1988. pp. 4–40. [Google Scholar]
- Allsopp (2015).Allsopp BA. Heartwater—Ehrlichia ruminantium infection. Revue Scientifique et Technique (International Office of Epizootics) 2015;34:557–568. doi: 10.20506/rst.34.2.2379. [DOI] [PubMed] [Google Scholar]
- Amann et al. (1990).Amann RI, Binder BJ, Olson RJ, Chisholm SW, Devereux R, Stahl DA. Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Applied and Environmental Microbiology. 1990;56:1919–1925. doi: 10.1128/AEM.56.6.1919-1925.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amann, Ludwig & Schleifer (1995).Amann RI, Ludwig W, Schleifer KH. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiological Reviews. 1995;59:143–169. doi: 10.1128/MMBR.59.1.143-169.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnold et al. (2016).Arnold CP, Merryman MS, Harris-Arnold A, McKinney SA, Seidel CW, Loethen S, Proctor KN, Guo L, Sánchez Alvarado A. Pathogenic shifts in endogenous microbiota impede tissue regeneration via distinct activation of TAK1/MKK/p38. eLife. 2016;5:e16793. doi: 10.7554/eLife.16793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Athanassopoulou et al. (2004).Athanassopoulou F, Groman D, Prapas T, Sabatakou O. Pathological and epidemiological observations on rickettsiosis in cultured sea bass (Dicentrarchus labrax L.) from Greece. Journal of Applied Ichthyology. 2004;20:525–529. doi: 10.1111/j.1439-0426.2004.00571.x. [DOI] [Google Scholar]
- Barker & Brown (1994).Barker J, Brown MR. Trojan horses of the microbial world: protozoa and the survival of bacterial pathogens in the environment. Microbiology. 1994;140(Pt 6):1253–1259. doi: 10.1099/00221287-140-6-1253. [DOI] [PubMed] [Google Scholar]
- Barth et al. (2006).Barth D, Krenek S, Fokin S, Berendonk T. Intraspecific genetic variation in Paramecium revealed by mitochondrial cytochrome c oxidase I sequences. Journal of Eukaryotic Microbiology. 2006;53:20–25. doi: 10.1111/j.1550-7408.2005.00068.x. [DOI] [PubMed] [Google Scholar]
- Boscaro et al. (2013).Boscaro V, Fokin SI, Schrallhammer M, Schweikert M, Petroni G. Revised systematics of Holospora-like bacteria and characterization of Candidatus Gortzia infectiva, a novel macronuclear symbiont of Paramecium jenningsi. Microbial Ecology. 2013;65:255–267. doi: 10.1007/s00248-012-0110-2. [DOI] [PubMed] [Google Scholar]
- Boscaro et al. (2012).Boscaro V, Fokin SI, Verni F, Petroni G. Survey of Paramecium duboscqui using three markers and assessment of the molecular variability in the genus Paramecium. Molecular Phylogenetics and Evolution. 2012;65:1004–1013. doi: 10.1016/j.ympev.2012.09.001. [DOI] [PubMed] [Google Scholar]
- Cafiso et al. (2015).Cafiso A, Sassera D, Serra V, Bandi C, McCarthy U, Bazzocchi C. Molecular evidence for a bacterium of the family Midichloriaceae (order Rickettsiales) in skin and organs of the rainbow trout Oncorhynchus mykiss (Walbaum) affected by red mark syndrome. Journal of Fish Diseases. 2015;39:497–450. doi: 10.1111/jfd.12371. [DOI] [PubMed] [Google Scholar]
- Cardwell & Martinez (2009).Cardwell MM, Martinez JJ. The Sca2 autotransporter protein from Rickettsia conorii is sufficient to mediate adherence to and invasion of cultured mammalian cells. Infection and Immunity. 2009;77:5272–5280. doi: 10.1128/IAI.00201-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cassella et al. (2017).Cassella L, Salvetti A, Iacopetti P, Ippolito C, Ghezzani C, Gimenez G, Ghigo E, Rossi L. Putrescine independent wound response phenotype is produced by ODC-like RNAi in planarians. Scientific Reports. 2017;7:9736. doi: 10.1038/s41598-017-09567-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castelli et al. (2015).Castelli M, Lanzoni O, Fokin SI, Schrallhammer M, Petroni G. Response of the bacterial symbiont Holospora caryophila to different growth conditions of its host. European Journal of Protistology. 2015;51:98–108. doi: 10.1016/j.ejop.2014.11.006. [DOI] [PubMed] [Google Scholar]
- Castelli et al. (2019a).Castelli M, Sabaneyeva E, Lanzoni O, Lebedeva N, Floriano AM, Gaiarsa S, Benken K, Modeo L, Bandi C, Potekhin A, Sassera D, Petroni G. Deianiraea, an extracellular bacterium associated with the ciliate Paramecium, suggests an alternative scenario for the evolution of Rickettsiales. ISME Journal. 2019a;13:2280–2294. doi: 10.1038/s41396-019-0433-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castelli, Sassera & Petroni (2016).Castelli M, Sassera D, Petroni G. Biodiversity of “non-model” Rickettsiales and their association with aquatic organisms. In: Thomas S, editor. Rickettsiales—biology, molecular biology, epidemiology, and vaccine development. Springer International Publishing; Cham: 2016. pp. 59–91. [Google Scholar]
- Castelli et al. (2019b).Castelli M, Serra V, Senra MVX, Basuri CK, Soares CAG, Fokin SI, Modeo L, Petroni G. The hidden world of Rickettsiales symbionts: “Candidatus Spectririckettsia obscura”, a novel bacterium found in Brazilian and Indian Paramecium caudatum. Microbial Ecology. 2019b;77:748–758. doi: 10.1007/s00248-018-1243-8. [DOI] [PubMed] [Google Scholar]
- Chan et al. (2018).Chan LL, Mak JW, Ambu S, Chong PY. Identification and ultrastructural characterization of Acanthamoeba bacterial endocytobionts belonging to the Alphaproteobacteria class. PLOS ONE. 2018;13:e0204732. doi: 10.1371/journal.pone.0204732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiellini et al. (2019).Chiellini C, Pasqualetti C, Lanzoni O, Fagorzi C, Bazzocchi C, Fani R, Petroni G, Modeo L. Harmful effect of Rheinheimera sp. EpRS3 (Gammaproteobacteria) against the protist Euplotes aediculatus (Ciliophora, Spirotrichea): insights into the ecological role of antimicrobial compounds from environmental bacterial strains. Frontiers in Microbiology. 2019;10:510. doi: 10.3389/fmicb.2019.00510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conti, Abnave & Ghigo (2014).Conti F, Abnave P, Ghigo E. Unconventional animal models: a booster for new advances in host-pathogen interactions. Frontiers in Cellular and Infection Microbiology. 2014;4:142. doi: 10.3389/fcimb.2014.00142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corbeil, Hyatt & Crane (2005).Corbeil S, Hyatt AD, Crane MSTJ. Characterisation of an emerging rickettsia-like organism in Tasmanian farmed Atlantic salmon Salmo salar. Diseases of Aquatic Organisms. 2005;64:37–44. doi: 10.3354/dao064037. [DOI] [PubMed] [Google Scholar]
- Crosson et al. (2014).Crosson LM, Wight N, Van Blaricom GR, Kiryu I, Moore JD, Friedman CS. Abalone withering syndrome: distribution, impacts, current diagnostic methods and new findings. Diseases of Aquatic Organisms. 2014;108:261–270. doi: 10.3354/dao02713. [DOI] [PubMed] [Google Scholar]
- Dishaw et al. (2014).Dishaw LJ, Flores-Torres J, Lax S, Gemayel K, Leigh B, Melillo D, Mueller MG, Natale L, Zucchetti I, De Santis R, Pinto MR, Litman GW, Gilbet JA. The gut of geographically disparate Ciona intestinalis harbors a core microbiota. PLOS ONE. 2014;9:e93386. doi: 10.1371/journal.pone.0093386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Driscoll et al. (2013).Driscoll T, Gillespie JJ, Nordberg EK, Azad AF, Sobral BW. Bacterial DNA sifted from the Trichoplax adhaerens (Animalia: Placozoa) genome project reveals a putative rickettsial endosymbiont. Genome Biology and Evolution. 2013;5:621–645. doi: 10.1093/gbe/evt036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dumler et al. (2001).Dumler JS, Barbet AF, Bekker CP, Dasch GA, Palmer GH, Ray SC, Rikihisa Y, Rurangirwa FR. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and ‘HGE agent’ as subjective synonyms of Ehrlichia phagocytophila. International Journal of Systematic and Evolutionary Microbiology. 2001;51:2145–2165. doi: 10.1099/00207713-51-6-2145. [DOI] [PubMed] [Google Scholar]
- Dumler et al. (2007).Dumler JS, Madigan JE, Pusterla N, Bakken JS. Ehrlichioses in humans: epidemiology, clinical presentation, diagnosis, and treatment. Clinical Infectious Diseases. 2007;45:S45–S51. doi: 10.1086/518146. [DOI] [PubMed] [Google Scholar]
- Dumler & Walker (2015).Dumler JS, Walker DH. Rickettsiales. In: Whitman WB, Rainey F, Kämpfer P, Trujillo M, Chun J, De Vos P, Hedlund B, Dedyshed S, editors. Bergey’s manual of systematics of archaea and bacteria. John Wiley & Sons, Ltd.; 2015. [DOI] [Google Scholar]
- Epis et al. (2008).Epis S, Sassera D, Beninati T, Lo N, Beati L, Piesman J, Rinaldi L, McCoy KD, Torina A, Sacchi L, Clementi E, Genchi M, Magnino S, Bandi C. Midichloria mitochondrii is widespread in hard ticks (Ixodidae) and resides in the mitochondria of phylogenetically diverse species. Parasitology. 2008;13:485–494. doi: 10.1017/S0031182007004052. [DOI] [PubMed] [Google Scholar]
- Ferrantini et al. (2009).Ferrantini F, Fokin SI, Modeo L, Andreoli I, Dini F, Görtz HD, Verni F, Petroni G. “Candidatus Cryptoprodotis polytropus”, a novel Rickettsia-like organism in the ciliated protist Pseudomicrothorax dubius (Ciliophora, Nassophorea) Journal of Eukaryotic Microbiology. 2009;56:119–129. doi: 10.1111/j.1550-7408.2008.00377.x. [DOI] [PubMed] [Google Scholar]
- Fokin (1997).Fokin S. Morphological diversity in the micronuclei in Paramecium. Archiv für Protistenkunde. 1997;148:375–387. doi: 10.1016/S0003-9365(97)80017-6. [DOI] [Google Scholar]
- Fokin (2010/2011).Fokin S. Paramecium genus: biodiversity, some morphological features and the key to the main morphospecies discrimination. Protistology. 2010/2011;6:227–235. [Google Scholar]
- Fokin & Chivilev (2000).Fokin SI, Chivilev SM. Paramecium. Morphometric analysis and taxonomy. Acta Protozoologica. 2000;39:1–14. [Google Scholar]
- Fokin et al. (2019).Fokin SI, Serra V, Ferrantini F, Modeo L, Petroni G. “Candidatus Hafkinia simulans” gen. nov. sp. nov., a novel Holospora-like bacterium from the macronucleus of the rare brackish water ciliate Frontonia salmastra (Oligohymenophorea, Ciliophora): multidisciplinary characterization of the new endosymbiont and its host. Microbial Ecology. 2019;77:1092–1106. doi: 10.1007/s00248-018-1311-0. [DOI] [PubMed] [Google Scholar]
- Fraune & Bosch (2007).Fraune S, Bosch TC. Long-term maintenance of species-specific bacterial microbiota in the basal metazoan Hydra. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:13146–13151. doi: 10.1073/pnas.0703375104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedman et al. (2000).Friedman CS, Andree KB, Beauchamp KA, Moore JD, Robbins TT, Shields JD, Hedrick RP. “Candidatus Xenohaliotis californiensis”, a newly described pathogen of abalone, Haliotis spp. along the west coast of North America. International Journal of Systematic and Evolutionary Microbiology. 2000;50:847–855. doi: 10.1099/00207713-50-2-847. [DOI] [PubMed] [Google Scholar]
- Fryer et al. (1992).Fryer JL, Lannan CN, Giovannoni SJ, Wood ND. Piscirickettsia salmonis gen. nov. sp. nov., the causative agent of an epizootic disease in salmonid fishes. International Journal of Systematic Bacteriology. 1992;42:120–126. doi: 10.1099/00207713-42-1-120. [DOI] [PubMed] [Google Scholar]
- Gao, Harb & Abu Kwaik (1997).Gao LY, Harb OS, Abu Kwaik Y. Utilization of similar mechanisms by Legionella pneumophila to parasitize two evolutionarily distant host cells, mammalian macrophages and protozoa. Infection and Immunity. 1997;65:4738–4746. doi: 10.1128/IAI.65.11.4738-4746.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ge & Rikihisa (2011).Ge Y, Rikihisa Y. Subversion of host cell signaling by Orientia tsutsugamushi. Microbes and Infection. 2011;13:638–648. doi: 10.1016/j.micinf.2011.03.003. [DOI] [PubMed] [Google Scholar]
- Gillespie et al. (2012).Gillespie JJ, Nordberg EK, Azad AF, Sobral BWS. Phylogeny and comparative genomics: the shifting landscape in the genomics era. In: Palmer GH, Azad AF, editors. Intracellular pathogens II Rickettiales. ASM Press; Washington: 2012. pp. 84–141. [Google Scholar]
- Gollas-Galván et al. (2014).Gollas-Galván T, Avila-Villa LA, Martínez-Porchas M, Hernandez-Lopez J. Rickettsia-like organisms from cultured aquatic organisms, with emphasis on necrotizing hepatopancreatitis bacterium affecting penaeid shrimp: an overview on an emergent concern. Reviews in Aquaculture. 2014;6:256–269. doi: 10.1111/raq.12043. [DOI] [Google Scholar]
- Gong et al. (2016).Gong J, Qing Y, Fu R, Su L, Zhang X, Zhang Q. Protist-Bacteria associations: Gammaproteobacteria and Alphaproteobacteria are prevalent as digestion-resistant bacteria in ciliated protozoa. Frontiers in Microbiology. 2016;7:498. doi: 10.3389/fmicb.2016.00498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- González-Estévez (2009).González-Estévez C. Autophagy meets planarians. Autophagy. 2009;5:290–297. doi: 10.4161/auto.5.3.7665. [DOI] [PubMed] [Google Scholar]
- Görtz & Brigge (1998).Görtz H-D, Brigge T. Intracellular bacteria in protozoa. Naturwissenschaften. 1998;85:359–368. doi: 10.1007/s001140050517. [DOI] [PubMed] [Google Scholar]
- Gouin et al. (2004).Gouin E, Egile C, Dehoux P, Villiers V, Adams J, Gertler F, Li R, Cossart P. The RickA protein of Rickettsia conorii activates the Arp2/3 complex. Nature. 2004;427:457–461. doi: 10.1038/nature02318. [DOI] [PubMed] [Google Scholar]
- Harb, Gao & Abu Kwaik (2000).Harb OS, Gao LY, Abu Kwaik Y. From protozoa to mammalian cells: a new paradigm in the life cycle of intracellular bacterial pathogens. Environmental Microbiology. 2000;2:251–265. doi: 10.1046/j.1462-2920.2000.00112.x. [DOI] [PubMed] [Google Scholar]
- Hess (2017).Hess S. Description of Hyalodiscus flabellus sp. nov. (Vampyrellida, Rhizaria) and identification of its bacterial endosymbiont, “Candidatus Megaira polyxenophila” (Rickettsiales, Alphaproteobacteria) Protist. 2017;168:109–133. doi: 10.1016/j.protis.2016.11.003. [DOI] [PubMed] [Google Scholar]
- Horn & Wagner (2004).Horn M, Wagner M. Bacterial endosymbionts of free-living amoebae. Journal of Eukaryotic Microbiology. 2004;51:509–514. doi: 10.1111/j.1550-7408.2004.tb00278.x. [DOI] [PubMed] [Google Scholar]
- Illa et al. (2019).Illa K, Shameem U, Serra V, Melai M, Mangam S, Basuri CK, Petroni G, Modeo L. Multidisciplinary investigation on the catfish parasite Hamatopeduncularia Yamaguti, 1953 (Monogenoidea: Dactylogyridae): description of two new species from India and phylogenetic considerations. The European Zoological Journal. 2019;86:132–155. doi: 10.1080/24750263.2019.1597931. [DOI] [Google Scholar]
- Kawafune et al. (2015).Kawafune K, Hongoh Y, Hamaji T, Sakamoto T, Kurata T, Hirooka S, Miyagishima S, Nozaki H. Two different rickettsial bacteria invading Volvox carteri. PLOS ONE. 2015;10:e0116192. doi: 10.1371/journal.pone.0116192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- King & Newmark (2013).King RS, Newmark PA. In situ hybridization protocol for enhanced detection of gene expression in the planarian Schmidtea mediterranea. BMC Developmental Biology. 2013;13:8. doi: 10.1186/1471-213X-13-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krenek, Berendonk & Petzoldt (2011).Krenek S, Berendonk TU, Petzoldt T. Thermal performance curves of Paramecium caudatum: a model selection approach. European Journal of Protistology. 2011;47:124–137. doi: 10.1016/j.ejop.2010.12.001. [DOI] [PubMed] [Google Scholar]
- Kwan & Schmidt (2013).Kwan JC, Schmidt W. Bacterial endosymbiosis in a chordate host: long-term co-evolution and conservation of secondary metabolism. PLOS ONE. 2013;8:e80822. doi: 10.1371/journal.pone.0080822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lanzoni et al. (2019).Lanzoni O, Sabaneyeva E, Modeo L, Castelli M, Lebedeva N, Verni F, Schrallhammer M, Potekhin A, Petroni G. Diversity and environmental distribution of the cosmopolitan endosymbiont “Candidatus Megaira”. Scientific Reports. 2019;9:1179. doi: 10.1038/s41598-018-37629-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lloyd et al. (2008).Lloyd SJ, La Patra SE, Snekvik KR, St-Hilaire S, Cain KD, Call DR. Strawberry disease lesions in rainbow trout from southern Idaho are associated with DNA from a Rickettsia-like organism. Diseases of Aquatic Organisms. 2008;82:111–118. doi: 10.3354/dao01969. [DOI] [PubMed] [Google Scholar]
- Longshaw (2011).Longshaw M. Diseases of crayfish: a review. Journal of Invertebrate Pathology. 2011;106:54–70. doi: 10.1016/j.jip.2010.09.013. [DOI] [PubMed] [Google Scholar]
- Loy et al. (1996).Loy JK, Dewhirst FE, Weber W, Frelier PF, Garbar TL, Tasca SI, Templeton JW. Molecular phylogeny and in situ detection of the etiologic agent of necrotizing hepatopancreatitis in shrimp. Applied and Environmental Microbiology. 1996;62:3439–3445. doi: 10.1128/AEM.62.9.3439-3445.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McQuiston & Paddock (2012).McQuiston JH, Paddock CD. Public health: Rickettsial infections and epidemiology. In: Palmer G, Azad A, editors. Intracellular pathogens II: Rickettsiales. American Society of Microbiology Press; Washington, D.C.: 2012. pp. 40–83. [DOI] [Google Scholar]
- Medlin et al. (1988).Medlin L, Elwood HJ, Stickel S, Sogin ML. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene. 1988;71:491–499. doi: 10.1016/0378-1119(88)90066-2. [DOI] [PubMed] [Google Scholar]
- Modeo et al. (2013a).Modeo L, Fokin SI, Boscaro V, Andreoli I, Ferrantini F, Rosati G, Verni F, Petroni G. Morphology ultrastructure and molecular phylogeny of the ciliate Sonderia vorax with insights into the systematics of order Plagiopylida. BMC Microbiology. 2013a;13:40. doi: 10.1186/1471-2180-13-40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Modeo et al. (2013b).Modeo L, Petroni G, Lobban CS, Verni F, Vannini C. Morphological, ultrastructural, and molecular characterization of Euplotidium rosati n. sp. (Ciliophora, Euplotida) from Guam. Journal of Eukaryotic Microbiology. 2013b;60:25–36. doi: 10.1111/jeu.12003. [DOI] [PubMed] [Google Scholar]
- Molmeret et al. (2005).Molmeret M, Horn M, Wagner M, Santic M, Abu Kwaik Y. Amoebae as training grounds for intracellular bacterial pathogens. Applied and Environmental Microbiology. 2005;71:20–28. doi: 10.1128/AEM.71.1.20-28.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murakami et al. (2017).Murakami T, Segawa T, Dial R, Takeuchi N, Kohshima S, Hongoh Y. Bacterial microbiota associated with the Glacier ice worm is dominated by both worm-specific and Glacier-derived facultative lineages. Microbes and Environments. 2017;32:32–39. doi: 10.1264/jsme2.ME16158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nitla et al. (2019).Nitla V, Serra V, Fokin SI, Modeo L, Verni F, Sandeep BV, Kalavati C, Petroni G. Critical revision of the family Plagiopylidae (Ciliophora: Plagiopylea), including the description of two novel species, Plagiopyla ramani and Plagiopyla narasimhamurthii, and redescription of Plagiopyla nasuta Stein, 1860 from India. Zoological Journal of the Linnean Society. 2019;186:1–45. doi: 10.1093/zoolinnean/zly041. [DOI] [Google Scholar]
- Nunan et al. (2003).Nunan LM, Noble B, Le Groumellec M, Lightner DV. Experimental infection of Penaeus vannamei by a rickettsia-like bacterium (RLB) originating from P. monodon. Diseases of Aquatic Organisms. 2003;54:43–48. doi: 10.3354/dao054043. [DOI] [PubMed] [Google Scholar]
- Perlman, Hunter & Zchori-Fein (2006).Perlman SJ, Hunter MS, Zchori-Fein E. The emerging diversity of Rickettsia. Proceedings of the Royal Society B: Biological Sciences. 2006;273:2097–2106. doi: 10.1098/rspb.2006.3541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petersen (2014).Petersen CH. Planarian resistance to blades and bugs. Cell Host & Microbe. 2014;16:271–272. doi: 10.1016/j.chom.2014.08.016. [DOI] [PubMed] [Google Scholar]
- Petroni et al. (2002).Petroni G, Dini F, Verni F, Rosati G. A molecular approach to the tangled intrageneric relationships underlying phylogeny in Euplotes (Ciliophora, Spirotrichea) Molecular Phylogenetics and Evolution. 2002;22:118–130. doi: 10.1006/mpev.2001.1030. [DOI] [PubMed] [Google Scholar]
- Powers & Mitchell (1910).Powers JH, Mitchell C. A new species of Paramecium (Paramecium multimicronucleata) experimentally determined. Biological Bulletin. 1910;19:324–332. doi: 10.2307/1535948. [DOI] [Google Scholar]
- Raoult & Roux (1997).Raoult V, Roux V. Rickettsioses as paradigms of new or emerging infectious diseases. Clinical Microbiology Reviews. 1997;10:694–719. doi: 10.1128/CMR.10.4.694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Renesto et al. (2003).Renesto P, Dehoux P, Gouin E, Touqui L, Cossart P, Raoult D. Identification and characterization of a phospholipase D-superfamily gene in rickettsiae. Journal of Infectious Diseases. 2003;188:1276–1283. doi: 10.1086/379080. [DOI] [PubMed] [Google Scholar]
- Rennoll-Bankert et al. (2016).Rennoll-Bankert KE, Rahman MS, Guillotte ML, Lehman SS, Beier-Sexton M, Gillespie JJ, Azad AF. RalF-mediated activation of Arf6 controls Rickettsia typhi invasion by co-opting phosphoinositol metabolism. Infection and Immunity. 2016;84:3496–3506. doi: 10.1128/IAI.00638-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rikihisa (2010).Rikihisa Y. Anaplasma phagocytophilum and Ehrlichia chaffeensis: subversive manipulators of host cells. Nature Reviews. Microbiology. 2010;8:328–339. doi: 10.1038/nrmicro2318. [DOI] [PubMed] [Google Scholar]
- Roberts-Galbraith, Brubacher & Newmark (2016).Roberts-Galbraith RH, Brubacher JL, Newmark PA. A functional genomics screen in planarians reveals regulators of whole-brain regeneration. eLife. 2016;2016:e17002. doi: 10.7554/eLife.17002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosati et al. (2004).Rosati G, Modeo L, Melai M, Petroni G, Verni F. A multidisciplinary approach to describe protists: a morphological, ultrastructural, and molecular study on Peritromus kahli Villeneuve-Brachon, 1940 (Ciliophora, Heterotrichea) Journal of Eukaryotic Microbiology. 2004;51:49–59. doi: 10.1111/j.1550-7408.2004.tb00160.x. [DOI] [PubMed] [Google Scholar]
- Ross et al. (2018).Ross PM, Pande A, Jones JB, Cope J, Flowers G. First detection of gas bubble disease and Rickettsia-like organisms in Paphies ventricosa, a New Zealand surf clam. Journal of Fish Diseases. 2018;41:187–190. doi: 10.1111/jfd.12684. [DOI] [PubMed] [Google Scholar]
- Rossi et al. (2014).Rossi L, Bonuccelli L, Iacopetti P, Evangelista M, Ghezzani C, Tana L, Salvetti A. Prohibitin 2 regulates cell proliferation and mitochondrial cristae morphogenesis in planarian stem cells. Stem Cell Reviews and Reports. 2014;10:871–887. doi: 10.1007/s12015-014-9540-1. [DOI] [PubMed] [Google Scholar]
- Rossi et al. (2018).Rossi L, Cassella L, Iacopetti P, Ghezzani C, Tana L, Gimenez G, Ghigo E, Salvetti A. Insight into stem cell regulation from sub-lethally irradiated worms. Gene. 2018;662:37–45. doi: 10.1016/j.gene.2018.04.009. [DOI] [PubMed] [Google Scholar]
- Rossi et al. (2008).Rossi L, Salvetti A, Batistoni R, Deri P, Gremigni V. Planarians, a tale of stem cells. Cellular and Molecular Life Science. 2008;65:16–23. doi: 10.1007/s00018-007-7426-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sabaneyeva et al. (2018).Sabaneyeva E, Castelli M, Szokoli F, Benken K, Lebedeva N, Salvetti A, Schweikert M, Fokin S, Petroni G. Host and symbiont intraspecific variability: the case of Paramecium calkinsi and “Candidatus Trichorickettsia mobilis”. European Journal of Protistology. 2018;62:79–94. doi: 10.1016/j.ejop.2017.12.002. [DOI] [PubMed] [Google Scholar]
- Schrallhammer et al. (2013).Schrallhammer M, Ferrantini F, Vannini C, Galati S, Schweikert M, Görtz HD, Verni F, Petroni G. “Candidatus Megaira polyxenophila” gen. nov. sp. nov.: considerations on evolutionary history, host range and shift of early divergent rickettsiae. PLOS ONE. 2013;8:e72581. doi: 10.1371/journal.pone.0072581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schulz et al. (2016).Schulz F, Martijn J, Wascher F, Lagkouvardos I, Kostanjšek R, Ettema TJG, Horn M. A Rickettsiales symbiont of amoebae with ancient features. Environmental Microbiology. 2016;18:2326–2342. doi: 10.1111/1462-2920.12881. [DOI] [PubMed] [Google Scholar]
- Senra et al. (2016).Senra MVX, Dias RPJ, Castelli M, Silva-Neto I-D, Verni F, Soares CAG. A house for two-double bacterial infection in Euplotes woodruffi Sq1 (Ciliophora, Euplotia) sampled in southeastern Brazil. Microbial Ecology. 2016;71:505–517. doi: 10.1007/s00248-015-0668-6. [DOI] [PubMed] [Google Scholar]
- Strüder-Kypke & Lynn (2010).Strüder-Kypke MC, Lynn DH. Comparative analysis of the mitochondrial cytochrome c oxidase subunit I COI gene in ciliates Alveolata, Ciliophora and evaluation of its suitability as a biodiversity marker. Systematics and Biodiversity. 2010;8:131–148. doi: 10.1080/14772000903507744. [DOI] [Google Scholar]
- Sun et al. (2009).Sun HY, Noe J, Barber J, Coyne RS, Cassidy-Hanley D, Clark TG, Findly RC, Dickerson HW. Endosymbiotic bacteria in the parasitic ciliate Ichthyophthirius multifiliis. Applied and Environmental Microbiology. 2009;75:7445–7452. doi: 10.1128/AEM.00850-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun & Wu (2004).Sun J, Wu X. Histology, ultrastructure, and morphogenesis of a rickettsia-like organism causing disease in the oyster, Crassostrea ariakensis Gould. Journal of Invertebrate Pathology. 2004;86:77–86. doi: 10.1016/j.jip.2004.04.004. [DOI] [PubMed] [Google Scholar]
- Sunagawa et al. (2009).Sunagawa S, DeSantis TZ, Piceno YM, Brodie EL, DeSalvo MK, Voolstra CR, Weil E, Andersen GL, Medina M. Bacterial diversity and White Plague Disease-associated community changes in the Caribbean coral Montastraea faveolata. ISME Journal. 2009;3:512–521. doi: 10.1038/ismej.2008.131. [DOI] [PubMed] [Google Scholar]
- Szokoli et al. (2016a).Szokoli F, Castelli M, Sabaneyeva E, Schrallhammer M, Krenek S, Doak T, Berendonk TU, Petroni G. Disentangling the taxonomy of Rickettsiales and description of two novel symbionts (“Candidatus Bealeia paramacronuclearis” and “Candidatus Fokinia cryptica”) sharing the cytoplasm of the ciliate protist Paramecium biaurelia. Applied and Environmental Microbiology. 2016a;82:7236–7247. doi: 10.1128/AEM.02284-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szokoli et al. (2016b).Szokoli F, Sabaneyeva E, Castelli M, Krenek S, Schrallhammer M, Soares C, Da Silva-Neto ID, Berendonk TU, Petroni G. “Candidatus Fokinia solitaria”, a novel “stand-alone” symbiotic lineage of Midichloriaceae (Rickettsiales) PLOS ONE. 2016b;11:e0145743. doi: 10.1371/journal.pone.0145743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan & Owens (2000).Tan CK, Owens L. Infectivity, transmission and 16S rRNA sequencing of a rickettsia, Coxiella cheraxi sp. nov. from the freshwater crayfish Cherax quadricarinatus. Diseases of Aquatic Organisms. 2000;41:115–122. doi: 10.3354/dao041115. [DOI] [PubMed] [Google Scholar]
- Taylor, Bandi & Hoerauf (2005).Taylor MJ, Bandi C, Hoerauf A. Wolbachia endosymbionts of filarial nematodes. Advances in Parasitology. 2005;60:245–284. doi: 10.1016/S0065-308X(05)60004-8. [DOI] [PubMed] [Google Scholar]
- Taylor-Mulneix et al. (2017).Taylor-Mulneix DL, Bendor L, Linz B, Rivera I, Ryman VE, Dewan KK, Wagner SM, Wilson EF, Hilburger LJ, Cuff LE, West CM, Harvill ET. Bordetella bronchiseptica exploits the complex life cycle of Dictyostelium discoideum as an amplifying transmission vector. PLOS Biology. 2017;15:e2000420. doi: 10.1371/journal.pbio.2000420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Timur et al. (2013).Timur G, Erkan M, Yardimci RE, Ercan MD, Çanak O, Ürkü Ç. Light and electron microscopic study of rickettsia-like organisms causing systemic granulomas in farmed sea bass (Dicentrarchus labrax) The Israeli Journal of Aquaculture = Bamidgeh. 2013;65:874–880. [Google Scholar]
- Torre et al. (2017).Torre C, Abnave P, Tsoumtsa LL, Mottola G, Lepolard C, Trouplin V, Gimenez G, Desrousseaux J, Gempp S, Levasseur A, Padovani L, Lemichez E, Ghigo E. Staphylococcus aureus promotes Smed-PGRP-2/Smed-setd8-1 methyltransferase signalling in planarian neoblasts to sensitize anti-bacterial gene responses during re-infection. EBioMedicine. 2017;20:150–160. doi: 10.1016/j.ebiom.2017.04.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torre & Ghigo (2015).Torre C, Ghigo E. La planaire: un ver immortel pour élucider la réponse immunitaire de l’homme (Planaria: an immortal worm to clarify human immune response) Medical Sciences. 2015;31:20–22. doi: 10.1051/medsci/20153101006. (Article in French) [DOI] [PubMed] [Google Scholar]
- Tsoumtsa et al. (2017).Tsoumtsa LL, Torre C, Trouplin V, Coiffard B, Gimenez G, Mege JL, Ghigo E. Antimicrobial capacity of the freshwater planarians against S. aureus is under the control of timeless. Virulence. 2017;4:1–10. doi: 10.1080/21505594.2016.1276689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vannini et al. (2014).Vannini C, Boscaro V, Ferrantini F, Benken K, Mironov T, Schweikert M, Görtz HD, Fokin S, Sabaneyeva EV, Petroni G. Flagellar movement in two bacteria of the family Rickettsiaceae: a re-evaluation of motility in the evolutionary perspective. PLOS ONE. 2014;9(2):e87718. doi: 10.1371/journal.pone.0087718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vannini et al. (2004).Vannini C, Rosati G, Verni F, Petroni G. Identification of the bacterial endosymbionts of the marine ciliate Euplotes magnicirratus (Ciliophora, Hypotrichia) and proposal of “Candidatus Devosia euplotis”. International Journal of Systematic and Evolutionary Microbiology. 2004;54:1151–1156. doi: 10.1099/ijs.0.02759-0. [DOI] [PubMed] [Google Scholar]
- Vannini et al. (2017).Vannini C, Sigona C, Hahn M, Petroni G, Fujishima M. High degree of specificity in the association between symbiotic betaproteobacteria and the host Euplotes (Ciliophora, Euplotia) European Journal of Protistology. 2017;59:124–132. doi: 10.1016/j.ejop.2017.04.003. [DOI] [PubMed] [Google Scholar]
- Walker & Ismail (2008).Walker DH, Ismail N. Emerging and re-emerging rickettsioses: endothelial cell infection and early disease events. Nature Reviews. Microbiology. 2008;6:375–386. doi: 10.1038/nrmicro1866. [DOI] [PubMed] [Google Scholar]
- Wang (2011).Wang W. Bacterial diseases of crabs: a review. Journal of Invertebrate Pathology. 2011;106:18–26. doi: 10.1016/j.jip.2010.09.018. [DOI] [PubMed] [Google Scholar]
- Weinert et al. (2009).Weinert LA, Aebi A, Stone GN, Jiggins FM. Evolution and diversity of Rickettsia bacteria. BMC Biology. 2009;7:6. doi: 10.1186/1741-7007-7-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wichterman (1986).Wichterman R. The biology of Paramecium. 2nd edition Plenum Press; New York and London: 1986. [Google Scholar]
- Yang, Narechania & Kim (2016).Yang A, Narechania A, Kim E. Rickettsial endosymbiont in the early-diverging streptophyte green alga Mesostigma viride. Journal of Phycology. 2016;52:219–229. doi: 10.1111/jpy.12385. [DOI] [PubMed] [Google Scholar]
- Yurchenko et al. (2018).Yurchenko T, Ševčíková T, Přibyl P, El Karkouri K, Klimeš V, Amaral R, Zbránková V, Kim E, Raoult D, Santos LMA, Eliáš M. A gene transfer event suggests a long-term partnership between eustigmatophyte algae and a novel lineage of endosymbiotic bacteria. ISME Journal. 2018;12:2163–2175. doi: 10.1038/s41396-018-0177-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaila et al. (2017).Zaila KE, Doak TG, Ellerbrock H, Tung C-H, Martins ML, Kolbin D, Yao MC, Cassidy-Hanley DM, Clark TG, Chang WJ. Diversity and universality of endosymbiotic Rickettsia in the fish parasite Ichthyophthirius multifiliis. Frontiers in Microbiology. 2017;8:189. doi: 10.3389/fmicb.2017.00189. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
COI gene and combined (partial) 18S rRNA-ITS1-5S rRNA gene-ITS2-(partial) 28S rRNA sequences of Paramecium multimicronucleatum strain US_Bl 16I1: GenBank accession numbers MK595741 and MK806287 respectively. 16S rRNA gene sequence of “Ca. Trichorickettsia mobilis”: Gen Bank accession number MK598854.
(A) Day 1 after feeding. (B) Day 7 after feeding. L, lumen of planarian intestine. No bacteria were observed in tissues of investigated control animals in none of the experimental timepoints. Scale bars stand for 1µm.
Data Availability Statement
The following information was supplied regarding data availability:
The raw data are available in the Supplemental Files.