

Abstract
NLRP3 inflammasome is engaged in the inflammatory response during acute lung injury (ALI). Purinergic receptor P2X7 has been reported to be upstream of NLRP3 activation. However, the therapeutic implication of P2X7 in ALI remains to be explored. The present study used lipopolysaccharide (LPS)-induced mouse model to investigate the therapeutic potential of P2X7 blockage in ALI. Our results showed that P2X7/NLRP3 inflammasome pathway was significantly upregulated in the lungs of ALI mice as compared with control mice. P2X7 antagonist A438079 suppressed NLRP3/ASC/caspase 1 activation, production of IL-1β, IL-17A and IFN-γ and neutrophil infiltration but not the production of IL-10, resulting in a significant amelioration of lung injury. Moreover, blockage of P2X7 significantly inhibited NLRP3 inflammasome activation and IL-1β production in bone marrow derived macrophages. Similar results were obtained using another P2X7 inhibitor brilliant blue G (BBG) in vivo. Thus, pharmacological blockage of P2X7/NLRP3 pathway can be considered as a potential therapeutic strategy in patients with ALI.
Keywords: Acute lung injury, P2X7, Inflammasome, IL-1β, Neutrophils
Highlights
-
•
P2X7/NLRP3 pathway is activated in mice with acute lung injury (ALI).
-
•
Blockade of P2X7 suppressed NLRP3 inflammasome activation in mice with ALI.
-
•
Blockade of P2X7 suppressed cytokine production and neutrophil infiltration.
-
•
Pharmacological blockage of P2X7 has therapeutic potential on ALI.
1. Introduction
Acute lung injury (ALI) or a more severe ALI defined as acute respiratory distress syndrome (ARDS) is the major cause of hypoxemic respiratory failure in adults during hospitalization with high morbidity and mortality [1]. Many conditions, such as trauma, infection and sepsis could lead to ALI [2]. Despite the diversity in etiology, inflammation takes an indispensable part in the process of ALI. Although many medical therapies are used to treat ALI/ARDS, there is still an in-hospital mortality rate over 30% [1]. Thus, development of novel therapeutic strategies especially targeting the overwhelming inflammation in ALI remains urgent.
NLRP3 is the best characterized family member of NOD-like receptors (NLRs) that are involved in innate and adaptive immune responses by composing inflammasomes with other intracellular molecules such as inflammatory caspases and apoptosis-associated speck-like protein containing a CARD (ASC) [3], [4]. Briefly, upon activation, NLRP3 interacts with ASC which bridges NRLP3 to procaspase 1 and allows for activation of caspase 1. Once activated, caspase 1 can cleave the proforms of IL-1β and IL-18 into their mature and active forms [5]. In the lungs, NLRP3 inflammasome activation and IL-1β production are augmented with LPS challenge in the ALI mice [6].
P2X7 is a membrane protein which can be activated by extracellular ATP and act as upstream of NLRP3 inflammasome to mediate the production of IL-1β [7], [8]. LPS administration increased P2X7 expression in the lung parenchyma and P2X7−/− mice showed decreased polymorphonuclear cell infiltration, less inflammatory cytokine production and reduced collagen deposition [8]. However, it is still unknown how P2X7 contributes to the pathogenesis of ALI. In this study, we demonstrated that pharmacological blockade of P2X7 by using selective antagonists effectively ameliorated ALI in mice via inhibiting NLRP3 inflammasome pathway.
2. Materials and methods
2.1. Mice
Male C57BL/6 mice were purchased and housed under specific pathogen-free conditions in the Experimental Animal Center at Sun Yat-sen University, Guangzhou, China. Experiments were approved by the Ethics Committee of Sun Yat-sen University. Mice were kept in these pathogen-free facilities and the experiments were performed in accordance with the National Institutes of Health Guide for Care and Use of Animals.
2.2. LPS-induced acute lung injury (ALI) model
LPS-induced ALI model was created as described previously [9]. Male C57BL/6 mice (8–9 weeks of age) were anesthetized with intraperitoneal ketamine (80 mg/kg) and xylazine (15 mg/kg). 6 mg/kg of LPS (Sigma-Aldrich, St. Louis, MO, USA) was delivered in 50 μl of phosphate-buffered saline (PBS) to the lungs via a 20-gauge angiocath catheter used as an endotracheal tube. The animals were rolled from side to side after administration to ensure even distribution. The normal control mice were given intratracheal instillation of PBS only.
2.3. Mouse bone marrow-derived macrophages (BMDMs)
BMDMs were generated from C57BL/6 mice as previously described [10]. Briefly, femora and tibiae were flushed with cold RPMI 1640 and cells were suspended in RPMI 1640 medium containing 10% fetal calf serum (Hyclone, Logan, Utah, USA). Cells were then cultured in a humidified atmosphere containing 5% CO2 at 37 °C with 20 ng/ml macrophage colony stimulating factor (M-CSF, Life Technologies, Grand Island, NY, USA) at 5 × 105/ml in culture medium for 6 days. The medium was changed every 2 days. PE-conjugated mouse anti-CD11b and FITC-conjugated mouse anti-F4/80 (both from eBioscience, San Diego, CA, USA) were used to confirm > 95% purity of BMDMs by flow cytometric analysis.
BMDMs were primed with 1 μg/ml LPS in the presence or absence of A438079 (5 mM) for 4 h followed by stimulation with ATP (5 mM) for 1 h. Following different interventions, cell supernatants were collected for detection of IL-1β by ELISA (R&D Systems, Minneapolis, USA) according to the manufacturer's instructions. Cells were subjected to Western blot analysis of NLRP3 inflammasome activation.
2.4. Pharmacological blockage of P2X7
A438079 (Abcam, Cambridge, UK) was diluted at 5 mg/ml in saline. As previously described [11], mice were treated intraperitoneally with A438079 (80 mg/kg) every 24 h while the first injection began at 1 h before LPS administration. Groups of 6 mice were sacrificed 48 h after LPS administration. The bronchoalveolar lavage (BAL) fluid was collected for cell counts, protein quantification and Enzyme-linked immunosorbent assay (ELISA). Lung tissues were collected for histology, immunohistochemistry, ELISA and western blot analysis.
Another P2X7 antagosist Brilliant blue G (BBG, Sigma-Aldrich, St. Louis, MO, USA) was also used in this study. BBG was diluted at 3 mg/ml in vehicle (saline) solution. Mice were treated intraperitoneally with BBG (45.5 mg/kg) every 48 h while the first injection began at 1 h before LPS administration, as previously described [12]. Mice were sacrificed 48 h after LPS administration.
2.5. Bronchoalveolar lavage
Bronchoalveolar lavage (BAL) was performed as previously described [13]. Briefly, mice were anesthetized and sacrificed by heart puncture after opening the thoracic cavity. The trachea was exposed and an 18G sterile needle with blunt end was inserted into the trachea through a small semi-excision. 0.8 ml PBS was injected and withdrawn for the first lavage. The lavage procedure was repeated twice. A total volume of 2.4 ml BAL fluid per mouse was collected. The BAL fluid was centrifuged at 300 g for 10 min to pellet cells. Supernatants were collected for ELISA and total protein analysis. The pelleted cells were resuspended in a small volume of serum-free DMEM culture medium (Life Technologies, Grand Island, NY, USA). Total cell counts were determined on a grid hemocytometer. Differential cell counts were enumerated on cytospin-prepared slides that were stained with Wright–Giemsa stain. A total of 500 cells were counted in cross-section per sample, and the number of neutrophil was calculated as the total cell count times the percentage of neutrophil in the BALF sample. Total protein level was determined by using BCA Protein Assay Kit (Thermo Fisher Scientific, Rockford, IL, USA) according to the manufacturers' instructions.
2.6. Enzyme-linked immunosorbent assay (ELISA)
IL-1β, IL-10, IL-17A and IFN-γ in lung tissues and BAL fluid were measured using ELISA kits for mouse IL-1β and IL-10 (R&D Systems, Minneapolis, USA) and ELISA Ready-Set-Go kit for mouse IL-17A and IFN-γ (eBioscience, San Diego, CA, USA), respectively, following the manufacturers' instructions.
2.7. Histology and immunohistochemistry
Paraformaldehyde-fixed, paraffin-embedded lung tissues were sectioned for hematoxylin and eosin staining and immunohistochemical staining of F4/80 (Abcam, Cambridge, UK) and IL-1β (Santa Cruz Biotechnology, Dallas, Texas, USA). Sections were counterstained with hematoxylin. The number of F4/80+ cells in the lung was counted in 20 consecutive fields under high power fields (hpf) (× 40) and expressed as cells/hpf.
2.8. Western blotting
Proteins from lung tissues were extracted with cell lysis buffer (Cell Signaling Technology, Beverly, MA, USA) and analyzed by western blotting as described previously [14]. The primary antibodies used in this study included: mouse anti-NLRP3 (Adipogen, San Diego, CA, USA), rabbit anti-P2X7 antibodies (Abcam, Cambridge, UK), mouse anti-caspase1-p20 (Adipogen), rabbit anti-ASC (Adipogen) and anti-GAPDH antibodies (Santa Cruz Biotechnology). HRP conjugated anti-mouse and anti-Rabbit IgG (both from Cell Signaling Technology) were used as secondary antibodies. Signals were detected with enhanced chemiluminescence (Cell Signaling Technology).
2.9. Statistical analysis
Data were presented as mean ± SEM. Statistical analyses were performed using one-way ANOVA. All data were analyzed using SPSS software (version 16.0). P values less than 0.05 were considered significant.
3. Results
3.1. A438079 inhibited the activation of NLRP3 inflammasome in mice with ALI
Expression of the P2X7/NLRP3 inflammasome pathway was examined in the mouse model of LPS-induced lung injury. Enhanced protein expression of P2X7, NLRP3, and ASC was observed in the lungs from LPS-induced lung injury mice compared with control mice treated by PBS (Fig. 1 ). The active caspase 1-p20 subunit represents the highly active p20/p10 tetrameric forms of processed caspase 1. Expression of the p20 subunit of caspase 1 was significantly elevated in the lungs from LPS-induced lung injury mice compared with normal control mice (Fig. 1).
Fig. 1.

A438079 suppresses activation of P2X7/NLRP3 pathway in LPS-induced lung injury mice. a, representative Western blot bands showed the protein expression of P2X7, NLRP3, ASC, pro-caspase 1, caspase 1p20 and GAPDH in the lung tissue at day 2 after LPS induction. b, Quantitative data showed respective protein expressions normalized to the values for GAPDH. Each bar represents mean ± SEM (n = 6). **P < 0.01 versus normal control (PBS) mice; #P < 0.05, ##P < 0.01 versus LPS-induced lung injury mice. NS = not significant.
The effects of the selective P2X7 inhibitor A438079 on activation of the NLRP3 inflammasome were tested. Expression of P2X7, NLRP3, ASC and caspase 1-p20 in lung tissue was significantly downregulated by A438079 treatment as demonstrated by western blot analysis (Fig. 1). However, expression of pro-caspase1 which is the precursor form of active caspase1p20 was not influenced by A438079.
3.2. A438079 ameliorated histopathologic injury in mice with ALI
The effects of A438079 on lung injury induced by LPS were investigated by histological examination with H&E staining and immunohistochemical staining of F4/80. As shown in Fig. 2a, inflammatory cell infiltration was observed in the pulmonary interstitium of LPS-induced lung injury mice and was significantly reduced by A438079 treatment. In addition, large number of F4/80+ macrophages was also detected in the LPS-induced injured lungs and was significantly reduced by A438079 treatment (Fig. 2b and c).
Fig. 2.
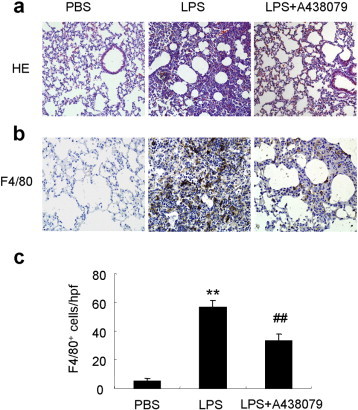
A438079 ameliorates lung injury and inflammatory cell infiltration in LPS-induced lung injury mice. a, HE staining of lung sections. Original magnification × 200. b, Immunohistochemistry staining of F4/80+ macrophages in lung tissues. c, Semiquantification of F4/80+ macrophages infiltration in the lungs. Results show that A438079 treatment attenuates pulmonary injury at day 2 after LPS induction. Each bar represents mean ± SEM (n = 6). **P < 0.01 versus normal control (PBS) mice; ##P < 0.01 versus LPS-induced lung injury mice.
3.3. A438079 decreased cell counts and total protein level in BAL fluid
To further investigate the effect of A438079 on lung inflammation, BAL fluids were collected for cell counts (Fig. 3b and c), Wright–Giemsa staining (Fig. 3a) and total protein analysis (Fig. 3d). In normal control mice, there were very few cells observed in the BAL fluid and almost 100% were alveolar macrophages. However, over 90% cells were neutrophils observed in the BLA fluids from the LPS-induced lung injury mice. In contrast, A438079 significantly reduces neutrophil and total cell number. Coincident with the cell counts, the total protein level was also elevated in the LPS-induced lung injury group, which was significantly reduced by A438079 treatment (Fig. 3d).
Fig. 3.

A438079 decreases cell counts and protein level in BAL fluid. a, Wright–Giemsa staining of cytospun cells from BAL fluids. b, Total cell counts of BAL fluids in PBS (normal control), LPS (LPS-induced lung injury) and LPS + A438079 (treatment) groups. c, Neutrophil counts in BAL fluid. d, Total protein level in BAL fluids. Each bar represents mean ± SEM (n = 6). **P < 0.01 versus normal control (PBS) mice; ##P < 0.01 versus LPS-induced lung injury mice.
3.4. A438079 reduced IL-1β level in both lung tissue and BAL fluid
IL-1β is an important downstream molecular modulated by P2X7/NLRP3 inflammasome pathway and acts as pivotal inflammatory factor in the LPS-induced lung injury. IL-1β expression in lung tissue and BAL fluid was apparently enhanced in the LPS-induced lung injury mice compared with normal control. The selective P2X7 inhibitor A438079 significantly inhibited IL-1β production as measured by ELISA (Fig. 4b and c). Similar results were confirmed by immunohistochemistry staining of IL-1β in lung sections as shown in Fig. 4a.
Fig. 4.
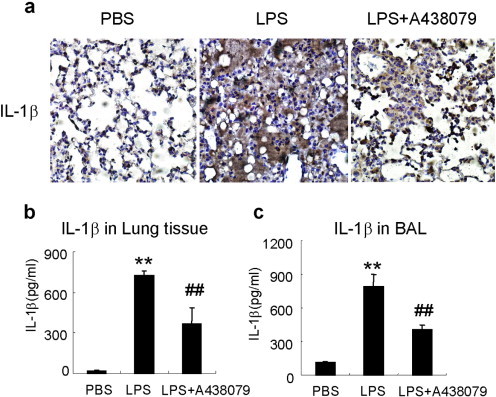
A438079 inhibits IL-1β production in lung tissues and BAL fluids. IL-1β in tissue homogenate or BAL fluid was measured by ELISA. a, Immunohistochemistry staining of IL-1β in lung tissues. b, IL-1β levels in lung tissues. c, IL-1β levels in BAL fluid. Each bar represents mean ± SEM (n = 6). **P < 0.01 versus normal control (PBS) mice; ##P < 0.01 versus control LPS-induced lung injury mice.
3.5. A438079 reduced IFN-γ and IL-17A level but not IL-10 level in both lung tissue and BAL fluid
IFN-γ and IL-17A were also examined by ELISA in this study. The concentrations of IFN-γ and IL-17A were remarkably enhanced in the lung tissue and BAL fluids of LPS-induced lung injury mice compared with normal control mice. Furthermore, A438079 treatment significantly suppressed IFN-γ (Fig. 5a and d) and IL-17A (Fig. 5b and e) expression.
Fig. 5.

A438079 inhibits IFN-γ and IL-17A but not IL-10 production in lung tissues and BAL fluids. Cytokine level in tissue homogenate or BAL fluid was measured by ELISA. a and d, IFN-γ levels in lung tissues (a) and BAL fluid (d). b and e, IL-17A levels in lung tissues (b) and BAL fluid (e). c and f, IL-10 levels in lung tissues (c) and BAL fluid (f). Each bar represents mean ± SEM (n = 6). **P < 0.01 versus normal control (PBS) mice; #P < 0.05, ##P < 0.01 versus LPS-induced lung injury mice. NS = not significant.
IL-10 considered as an anti-inflammatory cytokine was also remarkably upregulated in the lung tissue and BAL fluids of ALI mice. However, IL-10 level was not altered by A438079 treatment (Fig. 5c and f).
3.6. Inhibitory effect of A438079 on NLRP3 inflammasome activation and IL-1β production in mouse macrophages
In vitro, bone marrow derived macrophage was used to test the inhibitory effects of A438079 (Fig. 6 ). Expression of NLRP3 and ASC and activation of caspase 1 as evidenced by the increase of caspase1p20 as well as IL-1β production were enhanced with treatment of LPS and ATP. Enhanced expression of NLRP3 and ASC and activation of caspase 1 as well as IL-1β production were significantly inhibited by A438079. In contrast, expression of pro-caspase1 which is the precursor form of active caspase1p20 was not influenced by A438079.
Fig. 6.

A438079 inhibits NLRP3 inflammasome activation in mouse bone marrow derived macrophages (BMDMs). BMDMs were primed with 1 μg/ml LPS in the presence or absence of A438079 (5 mM) for 4 h followed by stimulation with ATP (5 mM) for 1 h. The NLRP3 inflammasome activation was evaluated by expression of NLRP3, ASC, pro-caspase 1 and caspase1p20. a, representative Western blot bands showed the protein expression of NLRP3, ASC, pro-caspase 1, caspase1p20 and GAPDH. b, Quantitative data showing respective protein expressions normalized to the values for GAPDH. c, IL-1β levels in cell supernatants from BMDMs determined by ELISA. Each bar represents mean ± SEM (n = 3). **P < 0.01 versus LPS only group; #P < 0.05, ##P < 0.01 versus LPS + ATP group. NS = not significant.
3.7. BBG inhibited NLRP3 inflammasome activation in ALI mice
To further confirm that the in vivo effect of P2X7 blockage is mediated through inhibition of NLRP3 pathway, we treated ALI mice with another selective P2X7 inhibitor, BBG. Similar inhibitory effects on NLRP3 inflammasome activation and IL-1β production were observed as shown in Fig. 7 . Inhibition of NLRP3 pathway resulted in reductions of inflammation (Fig. 8a) and macrophage infiltration (Fig. 8b) in the lungs of ALI mice. The total cell count (Fig. 8c) and protein level (Fig. 8d) in BAL fluids were also significantly reduced by BBG treatment.
Fig. 7.
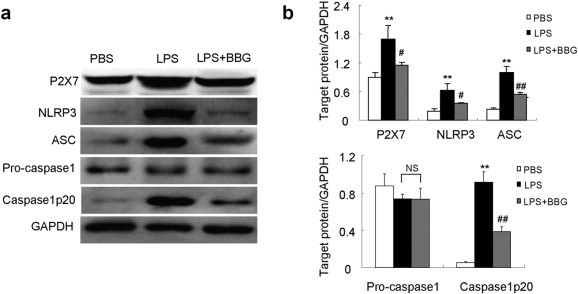
BBG suppresses activation of P2X7/NLRP3 pathway in LPS-induced lung injury mice. a, representative Western blot bands showed the protein expression of P2X7, NLRP3, ASC, pro-caspase 1, caspase 1p20 and GAPDH in the lung tissue at day 2 after LPS induction. b, Quantitative data showing respective protein expressions normalized to the values for GAPDH. Each bar represents mean ± SEM (n = 6). **P < 0.01 versus normal control (PBS) mice; #P < 0.05, ##P < 0.01 versus LPS-induced lung injury mice. NS = not significant.
Fig. 8.

BBG ameliorates lung injury and inflammatory cell infiltration in LPS-induced lung injury mice. a, HE staining of lung sections. Original magnification × 200. b, Semiquantification of F4/80+ macrophage infiltration in the lung tissues. c, Total cell counts of BAL fluids. d, Total protein level in BAL fluids. Each bar represents mean ± SEM (n = 6). **P < 0.01 versus normal control (PBS) mice; #P < 0.05, ##P < 0.01 versus LPS-induced lung injury mice.
4. Discussion
Inflammation is the hallmark in the development of ALI. However, the initiation and modification of this critical process are still beyond clear interpretation. LPS-induced lung injury mouse model was used in the present study which is manifested as damage of alveolar walls, neutrophil accumulation and release of inflammatory cytokines [9]. This study showed that blockade of P2X7 effectively ameliorated ALI via inhibiting NLRP3 inflammasome pathway.
P2X7 is expressed by many types of cells such as monocyte/macrophage [15], dendritic cells [16], lymphocytes [15], endothelial cells [17], mast cells [18], eosinophils [19] and alveolar type I epithelial cells [20]. Increasing data show P2X7 is correlated with inflammation. Activation of P2X7 in human macrophages triggers the release of IL-1β and IL-18 [21]. P2X7 null mice are protected from ALI induced by LPS [8]. In this study, P2X7 expression was significantly enhanced at the protein level in the lung tissues from ALI mice, paralleled with alveolar damage and inflammatory cytokine production. To dissect the functional role of P2X7 in ALI, a selective P2X7 antagonist A438079 was used to treat the ALI mice. We found that blockage of P2X7 inhibited the activation of NLRP3 inflammasome pathway, neutrophil accumulation and production of proinflammatory cytokines, resulting in reduction of lung damage.
IL-1β is an important mediator of the inflammatory response, which is proteolytically processed to its active from by caspase 1. The critical link between P2X7, IL-1β and NLRP3 has been brought to light recently. P2X7 has been reported as a trigger for NLRP3 activation [22]. Activation of NLRP3 results in assembly of the NLRP3/ASC/caspase 1 complex which facilitates caspase 1-mediated processing and release of the proinflammatory cytokine IL-1β [23]. Previous studies have shown that NLRP3 and caspase 1 regulate the inflammatory process in many lung diseases such as ALI [24], [25], Chlamydia pneumoniae infection [26], acute allergic airway inflammation [27], rituximab-induced interstitial lung disease (R-ILD) [28] and late lung fibrosis [29]. In our study, NLRP3 was activated in the lungs of ALI mice accompanied with enhanced expression of P2X7. Blockade of P2X7 resulted in inhibition of NLRP3/ASC/caspase 1 activation, suppression of IL-1β production and attenuation of lung injury, all of which indicates that the P2X7/NLRP3 pathway is involved in the pathogenesis of ALI.
IL-17 is another indispensable proinflammatory cytokine produced by T-helper cells (Th17) and is upregulated by IL-1β, IL-18, IL-6 and IL-23 [30]. IL-17 acts as a potent mediator in autoimmunity and neutrophil inflammation by increasing chemokine production in various tissues to recruit monocytes and neutrophils to the site of inflammation. IL-17A could regulate CXC chemokine and G-CSF production which is necessary for neutrophil differentiation, activation and recruitment [31], [32]. IL-17 producing γδ T and Th17 lymphocytes are reported to mediate lung inflammation in experimental silicosis while IL-17R-deficient animals or IL-17A antibody neutralization reduced neutrophil influx and lung injury [33]. IL-17 is also found to mediate ALI induced by influenza A (H1N1) virus [34]. In our study, IL-17A was markedly elevated in the lung tissues and BAL fluids of ALI mice as demonstrated by ELISA analysis. Blockade of P2X7 with A438079 significantly reduced IL-17 production as well as neutrophil accumulation in conjunction with inhibition of NLRP3 pathway. In addition, IFN-γ is a key contributor to human pulmonary injury and to the viral-storm during SARS coronavirus infection and in acute respiratory distress syndrome (ARDS) [35], [36], [37]. Our data showed that blockade of P2X7 also suppressed the production of IFN-γ.
In this study, the expression of the NLRP3 inflammasome components is upregulated in ALI mice. Such an increase in inflammasome components might be simply due to the increased population of innate immune cells in the lung tissues during ALI. Therefore, the suppressed expression of NLRP3 inflammmasome components by P2X7 treatment might reflect the reduced infiltration of innate immune cells. However, the activation of NLRP3 inflammasome was also significantly inhibited by P2X7 treatment, as evidenced by reduced cleavage of caspase 1, suggesting that therapeutic effects of P2X7 blockage is mediated through inhibition of NLRP3 activation rather than non-specific reduction of inflammatory infiltration. This was confirmed by our in vitro study using BMDMs, which showed over 50% decline in caspase 1 activation with the treatment of P2X7 antagonist.
Activation of the NLRP3 pathway is proposed to require two signals. Signal 1 for priming is a cytokine or Toll-like receptor ligand such as LPS, which trigger the synthesis of NLRP3. Signal 2 is the activation signal such as ATP, nigericin or monosodium urate, which acts through the P2X7 receptor [38], [39]. In our study, BMDMs were primed with LPS in the presence or absence of A438079 for 4 h followed by stimulation with ATP for 1 h. It is surprising to note that A438079 not only inhibited NLRP3 activation but also decreased the expression of NLRP3 and ASC. This finding suggests that A438079 also has other unspecified effects during the priming. Indeed, a recent study has shown that treatment with A438079 inhibits ERK1/2 phosphorylation [40]. Since NF-κB is required for the synthesis of NLRP3 induced by LPS [38], inhibition of ERK (upstream of NF-κB [41]) may reduce the expression of NLRP3.
In conclusion, the results of this study identify a new target for ALI therapy. Our data have shown that blockage of P2X7 signaling effectively ameliorated ALI by inhibiting NLRP3 inflammasome activation. Therefore, targeting the P2X7/NLRP3 signaling pathway would be a potential therapeutic strategy for human ALI.
Acknowledgments
This project was supported by grants from the Science and Technology Bureau of Guangdong Province (Grant No. 2013B021800113), the National Natural Science Foundation of China (81273278 and 81471598), the PhD Program Foundation of Ministry of Education of China (20120171110064), the Guangdong Natural Science Foundation (S2012010008780, and S2011010004578), and the Guangzhou Science and Technology Planning Program (2012J4100085).
All authors declare no conflicts of interest in this study.
References
- 1.Rubenfeld G.D., Caldwell E., Peabody E., Weaver J., Martin D.P., Neff M. Incidence and outcomes of acute lung injury. N. Engl. J. Med. 2005;353:1685–1693. doi: 10.1056/NEJMoa050333. [DOI] [PubMed] [Google Scholar]
- 2.Gullo A. Spectrum of acute lung injury, ARDS, sepsis. Minerva Anestesiol. 2002;68:45–53. [PubMed] [Google Scholar]
- 3.Schenten D., Medzhitov R. The control of adaptive immune responses by the innate immune system. Adv. Immunol. 2011;109:87–124. doi: 10.1016/B978-0-12-387664-5.00003-0. [DOI] [PubMed] [Google Scholar]
- 4.Gross O., Thomas C.J., Guarda G., Tschopp J. The inflammasome: an integrated view. Immunol. Rev. 2011;243:136–151. doi: 10.1111/j.1600-065X.2011.01046.x. [DOI] [PubMed] [Google Scholar]
- 5.Saleh M. The machinery of Nod-like receptors: refining the paths to immunity and cell death. Immunol. Rev. 2011;243:235–246. doi: 10.1111/j.1600-065X.2011.01045.x. [DOI] [PubMed] [Google Scholar]
- 6.Xu P., Wen Z., Shi X., Li Y., Fan L., Xiang M. Hemorrhagic shock augments Nlrp3 inflammasome activation in the lung through impaired pyrin induction. J. Immunol. 2013;190:5247–5255. doi: 10.4049/jimmunol.1203182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ferrari D., Pizzirani C., Adinolfi E., Lemoli R.M., Curti A., Idzko M. The P2X7 receptor: a key player in IL-1 processing and release. J. Immunol. 2006;176:3877–3883. doi: 10.4049/jimmunol.176.7.3877. [DOI] [PubMed] [Google Scholar]
- 8.Moncao-Ribeiro L.C., Cagido V.R., Lima-Murad G., Santana P.T., Riva D.R., Borojevic R. Lipopolysaccharide-induced lung injury: role of P2X7 receptor. Respir. Physiol. Neurobiol. 2011;179:314–325. doi: 10.1016/j.resp.2011.09.015. [DOI] [PubMed] [Google Scholar]
- 9.Mutlu G.M., Machado-Aranda D., Norton J.E., Bellmeyer A., Urich D., Zhou R. Electroporation-mediated gene transfer of the Na +, K +-ATPase rescues endotoxin-induced lung injury. Am. J. Respir. Crit. Care Med. 2007;176:582–590. doi: 10.1164/rccm.200608-1246OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhao J., Wang H., Huang Y., Zhang H., Wang S., Gaskin F. Lupus nephritis: glycogen synthase kinase 3beta promotes renal damage through activation of NLRP3 inflammasome in lupus-prone mice. Arthritis Rheum. 2015;67:1036–1044. doi: 10.1002/art.38993. (Hoboken, NJ) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Xie Y., Williams C.D., McGill M.R., Lebofsky M., Ramachandran A., Jaeschke H. Purinergic receptor antagonist A438079 protects against acetaminophen-induced liver injury by inhibiting p450 isoenzymes, not by inflammasome activation. Toxicol. Sci. 2012;131:325–335. doi: 10.1093/toxsci/kfs283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Diaz-Hernandez M., Diez-Zaera M., Sanchez-Nogueiro J., Gomez-Villafuertes R., Canals J.M., Alberch J. Altered P2X7-receptor level and function in mouse models of Huntington's disease and therapeutic efficacy of antagonist administration. FASEB J. 2009;23:1893–1906. doi: 10.1096/fj.08-122275. [DOI] [PubMed] [Google Scholar]
- 13.Fan J., Marshall J.C., Jimenez M., Shek P.N., Zagorski J., Rotstein O.D. Hemorrhagic shock primes for increased expression of cytokine-induced neutrophil chemoattractant in the lung: role in pulmonary inflammation following lipopolysaccharide. J. Immunol. 1998;161:440–447. [PubMed] [Google Scholar]
- 14.Zhao J., Wang H., Dai C., Wang H., Zhang H., Huang Y. P2X7 blockade attenuates murine lupus nephritis by inhibiting activation of the NLRP3/ASC/caspase 1 pathway. Arthritis Rheum. 2013;65:3176–3185. doi: 10.1002/art.38174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Di Virgilio F., Chiozzi P., Ferrari D., Falzoni S., Sanz J.M., Morelli A. Nucleotide receptors: an emerging family of regulatory molecules in blood cells. Blood. 2001;97:587–600. doi: 10.1182/blood.v97.3.587. [DOI] [PubMed] [Google Scholar]
- 16.Nihei O.K., de Carvalho A.C., Savino W., Alves L.A. Pharmacologic properties of P(2Z)/P2X(7)receptor characterized in murine dendritic cells: role on the induction of apoptosis. Blood. 2000;96:996–1005. [PubMed] [Google Scholar]
- 17.Ray F.R., Huang W., Slater M., Barden J.A. Purinergic receptor distribution in endothelial cells in blood vessels: a basis for selection of coronary artery grafts. Atherosclerosis. 2002;162:55–61. doi: 10.1016/s0021-9150(01)00681-5. [DOI] [PubMed] [Google Scholar]
- 18.Tatham P.E., Lindau M. ATP-induced pore formation in the plasma membrane of rat peritoneal mast cells. J. Gen. Physiol. 1990;95:459–476. doi: 10.1085/jgp.95.3.459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ferrari D., Idzko M., Dichmann S., Purlis D., Virchow C., Norgauer J. P2 purinergic receptors of human eosinophils: characterization and coupling to oxygen radical production. FEBS Lett. 2000;486:217–224. doi: 10.1016/s0014-5793(00)02306-1. [DOI] [PubMed] [Google Scholar]
- 20.Chen Z., Jin N., Narasaraju T., Chen J., McFarland L.R., Scott M. Identification of two novel markers for alveolar epithelial type I and II cells. Biochem. Biophys. Res. Commun. 2004;319:774–780. doi: 10.1016/j.bbrc.2004.05.048. [DOI] [PubMed] [Google Scholar]
- 21.Perregaux D.G., McNiff P., Laliberte R., Conklyn M., Gabel C.A. ATP acts as an agonist to promote stimulus-induced secretion of IL-1 beta and IL-18 in human blood. J. Immunol. 2000;165:4615–4623. doi: 10.4049/jimmunol.165.8.4615. [DOI] [PubMed] [Google Scholar]
- 22.Di Virgilio F. Liaisons dangereuses: P2X(7) and the inflammasome. Trends Pharmacol. Sci. 2007;28:465–472. doi: 10.1016/j.tips.2007.07.002. [DOI] [PubMed] [Google Scholar]
- 23.Qu Y., Franchi L., Nunez G., Dubyak G.R. Nonclassical IL-1 beta secretion stimulated by P2X7 receptors is dependent on inflammasome activation and correlated with exosome release in murine macrophages. J. Immunol. 2007;179:1913–1925. doi: 10.4049/jimmunol.179.3.1913. [DOI] [PubMed] [Google Scholar]
- 24.Liu Z., Zhao H., Liu W., Li T., Wang Y., Zhao M. NLRP3 inflammasome activation is essential for paraquat-induced acute lung injury. Inflammation. 2015;38:433–444. doi: 10.1007/s10753-014-0048-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Grailer J.J., Canning B.A., Kalbitz M., Haggadone M.D., Dhond R.M., Andjelkovic A.V. Critical role for the NLRP3 inflammasome during acute lung injury. J. Immunol. 2014;192:5974–5983. doi: 10.4049/jimmunol.1400368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.He X., Mekasha S., Mavrogiorgos N., Fitzgerald K.A., Lien E., Ingalls R.R. Inflammation and fibrosis during Chlamydia pneumoniae infection is regulated by IL-1 and the NLRP3/ASC inflammasome. J. Immunol. 2010;184:5743–5754. doi: 10.4049/jimmunol.0903937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ritter M., Straubinger K., Schmidt S., Busch D.H., Hagner S., Garn H. Functional relevance of NLRP3 inflammasome-mediated interleukin (IL)-1beta during acute allergic airway inflammation. Clin. Exp. Immunol. 2014;178:212–223. doi: 10.1111/cei.12400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kong H., Wang Y., Zeng X., Zhu Q., Xie W., Dai S. Involvement of NLRP3 inflammasome in rituximab-induced interstitial lung disease: a case report. J. Clin. Pharm. Ther. 2014;39:691–694. doi: 10.1111/jcpt.12198. [DOI] [PubMed] [Google Scholar]
- 29.Artlett C.M. The role of the NLRP3 inflammasome in fibrosis. Open Rheumatol. J. 2012;6:80–86. doi: 10.2174/1874312901206010080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.McAleer J.P., Kolls J.K. Directing traffic: IL-17 and IL-22 coordinate pulmonary immune defense. Immunol. Rev. 2014;260:129–144. doi: 10.1111/imr.12183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ouyang W., Kolls J.K., Zheng Y. The biological functions of T helper 17 cell effector cytokines in inflammation. Immunity. 2008;28:454–467. doi: 10.1016/j.immuni.2008.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.McGeachy M.J., Cua D.J. Th17 cell differentiation: the long and winding road. Immunity. 2008;28:445–453. doi: 10.1016/j.immuni.2008.03.001. [DOI] [PubMed] [Google Scholar]
- 33.Lo Re S., Dumoutier L., Couillin I., Van Vyve C., Yakoub Y., Uwambayinema F. IL-17A-producing gammadelta T and Th17 lymphocytes mediate lung inflammation but not fibrosis in experimental silicosis. J. Immunol. 2010;184:6367–6377. doi: 10.4049/jimmunol.0900459. [DOI] [PubMed] [Google Scholar]
- 34.Li C., Yang P., Sun Y., Li T., Wang C., Wang Z. IL-17 response mediates acute lung injury induced by the 2009 pandemic influenza A (H1N1) virus. Cell Res. 2012;22:528–538. doi: 10.1038/cr.2011.165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wong C.K., Lam C.W., Wu A.K., Ip W.K., Lee N.L., Chan I.H. Plasma inflammatory cytokines and chemokines in severe acute respiratory syndrome. Clin. Exp. Immunol. 2004;136:95–103. doi: 10.1111/j.1365-2249.2004.02415.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Huang K.J., Su I.J., Theron M., Wu Y.C., Lai S.K., Liu C.C. An interferon-gamma-related cytokine storm in SARS patients. J. Med. Virol. 2005;75:185–194. doi: 10.1002/jmv.20255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Theron M., Huang K.J., Chen Y.W., Liu C.C., Lei H.Y. A probable role for IFN-gamma in the development of a lung immunopathology in SARS. Cytokine. 2005;32:30–38. doi: 10.1016/j.cyto.2005.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bauernfeind F.G., Horvath G., Stutz A., Alnemri E.S., MacDonald K., Speert D. Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J. Immunol. 2009;183:787–791. doi: 10.4049/jimmunol.0901363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Franchi L., Eigenbrod T., Nunez G. Cutting edge: TNF-alpha mediates sensitization to ATP and silica via the NLRP3 inflammasome in the absence of microbial stimulation. J. Immunol. 2009;183:792–796. doi: 10.4049/jimmunol.0900173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Yan Y., Bai J., Zhou X., Tang J., Jiang C., Tolbert E. P2X7 receptor inhibition protects against ischemic acute kidney injury in mice. Am. J. Physiol. 2015;308:C463–C472. doi: 10.1152/ajpcell.00245.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tuyt L.M., Dokter W.H., Birkenkamp K., Koopmans S.B., Lummen C., Kruijer W. Extracellular-regulated kinase 1/2, Jun N-terminal kinase, and c-Jun are involved in NF-kappa B-dependent IL-6 expression in human monocytes. J. Immunol. 1999;162:4893–4902. [PubMed] [Google Scholar]