

Abstract
Many theoretical studies have proposed different causal mechanisms by which the structure of a host population could have important implications for life history traits of pathogens. However, little information is available from real systems to test these hypotheses. The domestic cat, Felis silvestris catus, whose populations exhibit a great variability in social and spatial structure, represent an ideal case study to assess this question. In the present article, we show how cat population structure may have influenced the evolution of feline viruses and, in return, how these viruses may have modified the genetic structure of cat populations. To cite this article: D. Pontier et al., C. R. Biologies 332 (2009).
Keywords: Domestic cats, Host population structures, Viral strategies, Vaccination, Host life history traits, Mathematical modelling
Résumé
Beaucoup d'études théoriques ont proposé différents mécanismes par lesquels la structure d'une population d'hôtes peut avoir des implications importantes pour les traits d'histoire de vie des agents infectieux. Cependant, peu d'études de systèmes hôtes-parasites évoluant en conditions naturelles ont pu tester ces hypothèses. Le chat domestique, Felis silvestris catus, dont les populations se caractérisent par une forte variabilité de leurs structures sociale et spatiale, représente un cas d'étude idéal pour évaluer cette question. Dans cet article, nous montrons comment la structure des populations de chats a pu influencer l'évolution des virus, et en retour, comment ces virus ont pu modifier la structure génétique des populations de chats. Pour citer cet article : D. Pontier et al., C. R. Biologies 332 (2009).
Mots-clés: Chats domestiques, Structures des populations hôtes, Stratégies virales, Vaccination, Traits d'histoire de vie des hôtes, Modélisation mathématique
1. Introduction
Despite the huge contribution of Darwin's work to biological science, diseases have long been excluded from evolution theory, with studies focusing more on proximal rather than evolutionary aspects. Only in recent decades have disease characteristics been considered as the product of selection through a co-evolutionary arms-race between parasites and host immune defences [1]. Many studies have highlighted the role of the host population structure as a selection pressure for parasites; arguing that it may have an impact on pathogen virulence [2], [3] or infectious period [4], as well as the impact of parasitism on the evolution of host life-history traits [5], [6]. Even if these studies have ‘opened a new chapter’ in the field of infectious diseases, they remain mainly theoretical and very few have dealt with real systems (but see [7], [8], [9]). The domestic cat (Felis silvestris catus) provides an interesting host model because this species carries a large diversity of well studied viruses, and ecological and genetic research has shown that the species exhibits great intra-specific variability in terms of population structures.
Cats and humans have a long history in common that began in the Middle East's Fertile Crescent around 9500 years B.C. [10], when cats were attracted to rodents infesting the grain stores of the first farming communities. Since then, the domestic cat has been particularly invasive and has colonised all continents and many islands. An extraordinary behavioural flexibility permits cats to adapt to a large ecological diversity and to exploit different resources. Depending on their location and the rural versus urban environment, cats form groups of varying size characterised by different spatial and social structures, different mating systems [11], [12], and also different genetic structures as exemplified by coat colours [13].
The domestic cat is the natural host of several viruses that infect the feline population worldwide as well as wild felids and sometimes other wild carnivores [14], [15]. Moreover, the genetic diversity among viral strains across their geographic range suggests local viral adaptations [16]. Interest in cat viruses has increased since recent events have shown that domestic cats may act as an interface between humans and wild animals in the propagation of some viruses, such as cowpox virus [17] or H5N1 avian influenza virus (AIV) [18], as a result of contact with infected rodents or birds, respectively, as was already described for rabies [19].
The great variability in cat population structures makes the domestic cat and its viruses a suitable model to analyse the evolution of both host and virus life history traits. More precisely, how do viruses respond to the constraint imposed by cat population features in a given environment? What may be the expected consequences of changes in cat population structures on the evolution of viruses? On the other hand, how do viruses alter host fitness traits? Interestingly, some feline infectious diseases share features with human infectious diseases, e.g., FIV is an animal model of HIV [20] and the strong antigenic and clinical diversity of feline calicivirus is similar to that of human noroviruses [21]. A better understanding of evolutionary mechanisms of host-parasite interactions acquired from the cat-virus system may then be extended, by analogy, to humans.
2. The domestic cat and its viruses
There are notable differences in the organisation of cat populations in terms of dispersal patterns, mating systems and individual behaviour according to resource distribution. Cats living on natural prey or on food distributed by human beings in rural areas live at low density (100–300 cats/km2, [11]), either alone or in small groups of 2 or 3 related females associated with human dwellings. Most males disperse after reaching sexual maturity whereas females are philopatric [11]. During reproduction, a small number of dominant males defend access to females and can roam over large areas competing aggressively to mate with receptive females, leading to a polygynous mating system [12], [22]. In contrast, in the urban environment, stray cats live in dense populations (up to 3000 cats/km2, [11]) and can form large multimale–multifemale social groups sharing food and shelters provided by humans. Habitat fragmentation is more pronounced in the urban environment since, in some locations, resources are highly aggregated [23]. Dispersal between social groups, called colonies, is very rare and mainly concerns adult females [24]. Because of the locally high density of females, males of the same colony are unable to monopolize females and share mating opportunities with their congeners without any aggressiveness. As a result a promiscuous mating system is observed [12], [25], [26].
Concomitantly, a rural-urban effect is highlighted in coat colour frequencies, the orange allele being very rare in urban cat populations, whereas the reverse situation is observed for the non-agouti allele that is much more common in urban than in rural cat populations [13], [27]. Coat colour genes are suspected to have pleiotropic effects upon morphology and behaviour which could be either favourable or disadvantageous according to population characteristics [13] and could modulate the contact structure among cats.
2.1. Cat population structure modulates viral transmission efficiency
The most important viruses carried by cats are feline calicivirus (FCV), feline herpes virus (FHV), feline parvovirus (FPV), feline coronavirus (FCoV), feline leukemia virus (FeLV) and feline immunodeficiency virus (FIV).
Field observations reveal a high variability in virus prevalence according to cat social organisation and mating system, which can be explained by the mode of transmission of these viruses; through contact with oral, nasal and ocular secretions for FHV or FCV, contact with faeces for FCoV, contact with any bodily secretions such as urine and faeces for FPV, or through saliva and bite wounds for FeLV. Whatever their route of transmission, these viruses are all very efficiently transmitted in cat colonies owing to the close proximity and high social contact rates among individuals, and the communal rearing of kittens by females [28], [29], [30]. Accordingly, a high prevalence of FHV, FCV and FPV occur in urban cat populations because of their higher densities and much higher social contact rates compared to rural ones [31].
Contrary to these aforementioned viruses, the prevalence of FIV is lower in urban (0–14%, [32], [33]) than in rural cat populations (20%, [34]), which may be explained by its transmission mode. In natural cat populations, FIV is practically solely transmitted by biting [32], [34], [35], [36], which frequently occurs when males fight and during coitus when the males bite females at the neck. As a result, males are more often infected in rural populations where a polygynous mating system gives rise to more fights [34]. Surprisingly, prevalence in females is similar in both types of populations, suggesting that the higher number of mates may counterbalance the lower prevalence of males in urban populations [32].
3. Cat population structure influences virus evolution
In host populations of small size, viruses face a high risk of extinction due to stochastic fluctuations in the transmission process [37], [38]. According to recent theoretical studies, stochastic extinction risk exerts an important selection pressure [4], [39] on virus evolution, in order to maintain virus circulation through time. Cats, whose populations are fragmented and of small size (between 60 and 300 cats, [13]), represent an interesting case study to illustrate this theory.
3.1. “Hit and run” vs. “hit and stay” strategies
Basically, viruses fall into two categories: (i) those that quickly kill or are eradicated from the host; and (ii) those that remain for a long time within the host. Viruses from the first category follow the so-called “hit and run” strategy, while those of the second category apply the “hit and stay” strategy [40]. These categories are based on the length of the infectious period, which impacts on the probabilities of transmission and of persistence in the host population.
For most viruses, acute infection of domestic cats will be followed by a phase of intense viral shedding. For these “hit and run” viruses, excretion lasts a few weeks. For some viruses such as FPV [41] and FCV [42], infected cats mount an immune response and get rid of the virus. In such conditions, the susceptible pool is quickly depleted before the birth of new susceptible individuals and these viruses should disappear from the local cat population. This strategy favours quick disease transmission, but exposes viruses to a higher risk of extinction, so their persistence relies on additional strategies. FPV is excreted in large amounts in the faeces and the virus remains infectious for months in the contaminated environment [43], from where it can infect subsequent susceptible generations [44]. In addition, the DNA parvovirus of carnivores can cross the species barrier, specifically between cats and dogs, two species living in close proximity [45], [46], [47]. This is due to its especially high mutation rate, which is closer to that of RNA viruses than to that of DNA viruses [46]. Therefore, dogs may act as a reservoir that allows the virus to persist when the number of susceptible cats would be insufficient. The genetic variability of FCV, a RNA virus, is associated with a strong antigenic diversity. This could explain the ability of the virus to escape the host immune response, which contributes to the persistence of the virus in the cat population. The continuity of FCV transmission relies also on some chronically infected cats shedding the virus over prolonged periods of time (up to several months, [30]).
“Hit and stay” viruses have evolved diverse mechanisms for ensuring their survival within the infected hosts and, thus, their persistence in cat populations. FHV lies dormant in the host and reactivates when the animal suffers stressful conditions. Infected individuals can episodically transmit the virus to susceptible ones [48]. FIV induces a chronic infection, as is also the case for FCoV in some individuals. Chronically infected individuals shed the virus over long periods [28], ensuring its long-term persistence in cat populations. Like all retroviruses, FeLV and FIV integrate into the host genome as a provirus that will persist in the infected cell until it dies and, thereby, can establish “latent” infections [49], [50]. In addition, feline retroviruses have evolved various strategies to escape from the host specific immune response [51]. In the case of FIV, its high genetic variability explains its ability to persist in the face of a strong host immune response [52].
3.2. Targeting hosts that serve their interest
Targeting hosts is another virus strategy to maintain their circulation. In the rural environment Courchamp et al. [34] showed that dominant males represent a core-group [53] for FIV; that is, a group of individuals that contributes disproportionately to pathogen transmission within, but also between, populations. Actually, males can roam over very long distances during the reproductive season in search of receptive females and frequently get involved in fights [11]. These males are important targets for viruses that must spread between sub-populations in order to survive. This may explain why FIV is primarily transmitted through bites [39], out of all the possible modes of transmission (e.g., sexual, or vertical in-utero or in-lacto) reported experimentally [54], [55]. These results may clarify the apparent paradox of FIV in that it exhibits an endemic pattern in rural cat populations despite its low prevalence, relying on a small number of at-risk individuals. However, this strategy is ineffective in the urban environment where fights and overall male dispersion are almost non-existent [24], [56]. This urban social organisation is recent for the cat-FIV system in an evolutionary time-scale. So is FIV adapted to the urban context? If so, one would expect selection for a new mode of transmission and the emergence of more ‘fit’ viral strains adapted to the new social structure of cats generated by the urban environment.
4. Viruses may modulate the life history of cats
4.1. Viruses and the evolution of cat sexual size dimorphism
Due to their higher aggressiveness, both male and female orange cats (that are almost exclusively present in rural populations) are more heavily infected by FIV than other cats [57]. This is probably because orange males fight more and because there is a greater tendency for males to bite orange females during coitus because orange females struggle more. A curiosity is the greater sexual polymorphism of orange cats compared to non-orange individuals: orange males are heavier than non-orange males, whereas orange females are lighter than other females [57]. Due to their lighter body size, orange females reproduce earlier than non-orange ones but devote less energy resources to reproduction [23], [57], whereas orange males, because of their greater body size, monopolise receptive females earlier and more easily than other congeners in the polygynous mating system [12], [57]. A possible explanation of these different mating strategies between genotypes may be the differential impact of FIV on colour morphs. According to life history theory, early reproduction could balance the survival costs due to FIV. Since heavier males have an advantage over lighter males during conflicts, orange males with the highest growth rates may be selected for. In contrast, early reproduction in orange females may have arisen from selection for reduced size in order to attain sexual maturity earlier. This may explain the differing evolution of body mass in male and female orange cats. Overall, these results suggest that microparasites may play a role in the evolution of sexual size dimorphism in mammals [57].
4.2. Can FIV play a role in the maintenance of cat coat colour polymorphism?
The frequencies of orange cats are always approximately 20% in rural populations [13]. One potential cause for the long-term maintenance of such polymorphism is a parasite-driven frequency-dependence in natural selection [58]. This supposes a balance between a fitness advantage for disease-free orange cats, which compensates for their increased mortality risk due to FIV infection. If the proportion of orange cats in the population becomes too large, most orange cats will become infected sooner and other cats will have a selective advantage. In this model FIV prevents the orange genotype from replacing the alternative genotype in the cat population.
To test this hypothesis we built a highly simplified mathematical model (see Supplementary material). Our mathematical model assumes a small fitness benefit for the orange genotype, but a larger infection rate by FIV than in cats of other coat colours. The main point of the model is to see if, in such circumstances, cat coat polymorphism can be maintained.
The mathematical model and its parameter values are consistent with the scenario proposed above (see Supplementary material). For example, realistic values for cat polymorphism (∼20% orange cats) and FIV-prevalence in non-orange cats (∼7%) and in orange cats (∼30%) are obtained for realistic parameter values, i.e. assuming a 2.3% selective advantage of orange cats, a transmission rate of FIV between non-orange cats of 0.1 and that orange cats are 5.75 times more susceptible to FIV (Fig. 1 ). However, the model is quite sensitive to the values of some parameters and so, requires restrictive conditions for the long-term persistence of the polymorphism between orange and non-orange cats. Firstly, the fitness advantage of orange cats in populations free from FIV must be small. Typically it must be around half of the reduction in the life expectancy induced by FIV (which is lower than 10% in natural conditions, Avril, Devillard, Pontier, unpublished results). Secondly, orange cats must have around ten times more at-risk contacts than non-orange ones, and certainly not less than five times more. This last condition should be investigated further, but seems realistic since orange males seem to wander over wider areas and are more prone to fight than their non-orange counterparts.
Fig. 1.
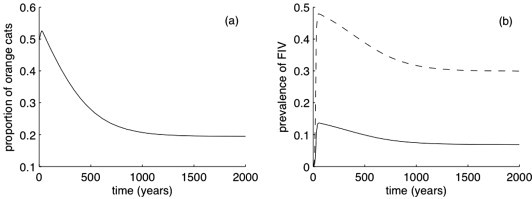
Potential role of FIV in the maintenance of cat coat colour polymorphism. The results have been derived from the model presented in Supplementary material, with the following parameters: transmission rate of FIV between non-orange cats of 0.1; life expectancy of non-orange cats is 4 years; a selective advantage of orange cats in a disease-free population of 2.3%; reduction in life expectancy in FIV infected individuals of 10%; and relative transmission rate of FIV in orange cats of 5.75. One FIV-infected cat is released in the fully susceptible cat population at time t = 0. (a) Evolution of the proportion of orange cats in the population and (b) prevalence of FIV in orange (dashed line) and non-orange (solid line) cats. See Supplementary material for a full description of the model.
In urban cat populations other selection pressures may explain the absence, or very low frequencies, of orange individuals independently of FIV, because aggressiveness is more of a handicap in these cat populations [13].
5. Future directions
Parasites induce an evolutionary pressure on their host and may drive cats to choose different strategies, promoting cat polymorphism. In return, the success of a parasite depends on the structure of its host population. The small size of cat groups and their dispersal behaviour have forced parasites to evolve accordingly and may explain some pathogen characteristics.
Taking this reasoning a step further, changes to cat population structure in the future may have consequences for pathogen expression. A typical example of such a change is vaccination. Vaccines are widely used in both human and animal medicine and are certainly one of the greatest medical successes of the last century. Efficient vaccines exist for many cat viruses, such as FeLV, FCV, FPV and rabies. It was estimated about fifteen years ago that at most 20% of cat populations were vaccinated in rural areas in France [23]. Moreover, some cat viruses pose a serious threat to other wild species like FPV and FeLV for free-ranging Florida panthers Puma concolor coryi [59]. Cross-species transmission may be restricted by reducing the number of infected cats by vaccination. While vaccines protect inoculated individuals from developing diseases, their effects go beyond that of the individual. Vaccinated individuals are less susceptible or not susceptible at all, to infection, which alters the structure of at-risk contacts in the host population. Vaccines have thus indirect consequences even in non-vaccinated individuals.
The most obvious effects for mass vaccination are expected to be positive: by removing vaccinated individuals from the transmission chain, perfect vaccines reduce the incidence of the disease, and even non-vaccinated individuals are less exposed. This mechanism is known as herd immunity [60] and is largely used to estimate the adequate proportion of a population requiring vaccination in order to stop disease spread. At the extreme, mass vaccination can, in theory, eradicate diseases [60], although in practice eradication has only been attained for smallpox in humans. In fragmented host populations, such as cat populations, vaccines make the persistence of parasites more difficult. As a result, we can expect that the widespread use of vaccines will force parasites to evolve persistence strategies (see above), e.g., by inducing more prolonged diseases. According to the trade-off hypothesis [61], the duration of a parasite-induced disease is inversely correlated with its virulence. Mass vaccination could select for less virulent strains in non-vaccinated individuals.
Not all theoretical studies share this optimistic view. With some vaccines the parasite can still spread in vaccinated hosts under the condition that it boosts its reproductive rate to overcome the host immune response. This could select for strains that replicate more rapidly within the host and, thus, are more virulent in non-vaccinated individuals [62]. As another example of a potential negative effect, Aguas et al. [63] argue that a reduction in exposure to pathogens may prevent the regular mild re-infections that are necessary to maintain efficient immune defences, leaving more individuals unprotected. Although infections are less frequent, since many individuals are not protected, they are also more severe.
It is almost impossible to know a priori the population effects of a mass vaccination campaign. Both positive and negative consequences have been predicted, but they remain mainly theoretical. Real time monitoring of the impact of the disease during vaccination campaigns, coupled with mathematical models predicting optimal vaccination strategies (such as those presented in [64], [65]) are necessary to understand and control the potential side effects of mass vaccination.
Finally, we would like to emphasise another important and often neglected factor, namely the parasitic community structure of the host population. Over the past decade there has been a growing body of evidence showing that parasites can interact where they occur in the same host [66]. For example, the symptoms of calicivirus are greatly aggravated in the presence of FIV [67]. These interactions can also be beneficial for the host, for example, the interaction between HIV and GB virus C [68]. Interactions between parasites occur in many species [66] and it would not be surprising that they also occur in cats. Improving our understanding of the impact of infectious diseases in the future may require integrative approaches that include more than one pathogen.
Acknowledgements
We thank the FIV VAX Lyon-Biopôle project and the ANR SEST “Pathocénoses et émergence des maladies transmissibles : un concept unificateur mis à l'épreuve sur des pathologies exemplaires” for financial support. We thank Jean-Paul Gonzalez and John O'Brien for useful comments on an earlier version of the paper.
Footnotes
Supplementary material associated with this article can be found, in the online version, at DOI: 10.1016/j.crvi.2008.07.012.
Supplementary material
References
- 1.Combes C. Chicago University Press; 2001. Parasitism: Ecology and Evolution of Intimate Interactions. [Google Scholar]
- 2.Lipsitch M., Herre E.A., Nowak M.A. Host population structure and the evolution of virulence: a “low of diminishing returns”. Evolution. 1995;49:743–748. doi: 10.1111/j.1558-5646.1995.tb02310.x. [DOI] [PubMed] [Google Scholar]
- 3.van Baalen M. Contact networks and the evolution of virulence. In: Dieckmann U., Metz J.A., Sabelis M.W., Sigmund K., editors. Adaptive Dynamics of Infectious Diseases: In Pursuit of Virulence Management. Cambridge University Press; Cambridge: 2002. pp. 85–103. [Google Scholar]
- 4.Read J.M., Keeling M.J. Stochasticity generates an evolutionary instability for infectious disease. Ecology Letters. 2007;10:818–827. doi: 10.1111/j.1461-0248.2007.01078.x. [DOI] [PubMed] [Google Scholar]
- 5.Anderson R.M., May R.M. Coevolution of hosts and parasites. Parasitology. 1982;85:411–426. doi: 10.1017/s0031182000055360. [DOI] [PubMed] [Google Scholar]
- 6.Boots M., Knell R.J. The evolution of risky behaviour in the presence of a sexually transmitted disease. Proc. R. Soc. B. 2002;269:585–589. doi: 10.1098/rspb.2001.1932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ebert D. Experimental evolution of parasites. Science. 1998;282:1432–1435. doi: 10.1126/science.282.5393.1432. [DOI] [PubMed] [Google Scholar]
- 8.Levin S.A., Dushoff J., Plotkin J.B. Evolution and persistence of influenza A and other diseases. Math. Biosci. 2004;188:17–28. doi: 10.1016/j.mbs.2003.08.010. [DOI] [PubMed] [Google Scholar]
- 9.Read A.F., Mackinnon M.J., Anwar A., Taylor L.H. Kin-selection models as evolutionary explanations of Malaria. In: Dieckmann U., Metz J.A., Sabelis M.W., Sigmund K., editors. Adaptive Dynamics of Infectious Diseases: In Pursuit of Virulence Management. Cambridge University Press; Cambridge: 2002. pp. 165–178. [Google Scholar]
- 10.Driscoll C.A., Menotti-Raymond M., Roca A.L., Hupe K., Johnson W.E., Geffen E., Harley E.H., Delibes M., Pontier D., Kitchener A.C., Yamaguchi N., O‘Brien S.J., Macdonald D.W. The Near Eastern origin of cat domestication. Science. 2007;317:519–523. doi: 10.1126/science.1139518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Liberg O., Sandell M., Pontier D., Natoli E. Density, spatial organisation and reproductive tactics in the domestic cat and other felids. In: Turner D.C., Bateson P., editors. The Domestic Cat: the Biology of its Behaviour. Cambridge University Press; Cambridge: 2000. pp. 119–147. [Google Scholar]
- 12.Say L., Pontier D., Natoli E. High variation in multiple paternity of domestic cats (Felis catus L.) in relation to environmental conditions. Proc. R. Soc. B. 1999;266:2071–2074. doi: 10.1098/rspb.1999.0889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Pontier D., Rioux N., Heizmann A. Evidence of selection on the orange allele in the domestic cat Felis catus: the role of social structure. Oikos. 1995;73:299–308. [Google Scholar]
- 14.Driciru M., Siefert L., Prager K.C., Dubovi E., Sande R., Princee F., Friday T., Munson L. A serosurvey of viral infections in lions (Panthera leo), from Queen Elizabeth National Park, Uganda. J. Wildl. Dis. 2006;42:667–671. doi: 10.7589/0090-3558-42.3.667. [DOI] [PubMed] [Google Scholar]
- 15.Nishimura Y., Goto Y., Yoneda K., Endo Y., Mizuno T., Hamachi M., Maruyama H., Kinoshita H., Koga S., Kimori M., Fushuku S., Ushinohama K., Akuzawa M., Watari T., Hasegawa A., Tsujimoto H. Interspecies transmission of Feline Immunodeficiency Virus from the domestic cat to the Tsushima cat (Felis bengalensis euptilura) in the wild. J. Virol. 1999;73:7916–7921. doi: 10.1128/jvi.73.9.7916-7921.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Carpenter M.A., Brown E.W., Macdonald D.W., O'Brien S.J. Phylogeographic patterns of feline immunodeficiency virus genetic diversity in the domestic cat. Virology. 1998;251:234–243. doi: 10.1006/viro.1998.9402. [DOI] [PubMed] [Google Scholar]
- 17.Willemse A., Eqberink H.F. Transmission of cowpox virus infection from domestic cat to man. Lancet. 1985;1:1515. doi: 10.1016/s0140-6736(85)92299-8. [DOI] [PubMed] [Google Scholar]
- 18.Thiry E., Zicola A., Addie D., Egberink H., Hartmann K., Lutz H., Poulet H., Horzinek M.C. Highly pathogenic avian influenza H5N1 virus in cats and other carnivores. Vet. Microbiol. 2007;122:25–31. doi: 10.1016/j.vetmic.2006.12.021. [DOI] [PubMed] [Google Scholar]
- 19.Blanton J.D., Hanlon C.A., Rupprecht C.E. Rabies surveillance in the United States during 2006. J. Am. Vet. Med. Assoc. 2007;231:540–556. doi: 10.2460/javma.231.4.540. [DOI] [PubMed] [Google Scholar]
- 20.Burkhard M.J., Dean G.A. Transmission and immunopathogenesis of FIV in cats as a model for HIV. Curr. HIV Res. 2003;1:15–29. doi: 10.2174/1570162033352101. [DOI] [PubMed] [Google Scholar]
- 21.Radford A.D., Gaskell R.M., Hart C.A. Human norovirus infection and the lessons from animal caliciviruses. Curr. Opin. Infect. Dis. 2004;17:471–478. doi: 10.1097/00001432-200410000-00012. [DOI] [PubMed] [Google Scholar]
- 22.Pontier D., Natoli E. Male reproductive success in the domestic cat (Felis catus L.): a case history. Behav. Processes. 1996;37:85–88. doi: 10.1016/0376-6357(95)00070-4. [DOI] [PubMed] [Google Scholar]
- 23.D. Pontier, Analyse de la variabilité des traits d'histoire de vie chez les mammifères. Mémoire d'habilitation à diriger des recherches, Université Lyon 1, 1993
- 24.Devillard S., Say L., Pontier D. Dispersal pattern of domestic cats (Felis catus) in a promiscuous urban population: do females disperse or die? J. Anim. Ecol. 2003;72:203–211. [Google Scholar]
- 25.Natoli E., De Vito E. Agonistic behavior, dominance rank, and copulatory success in large multi-male feral cats Felis catus L. colony in central Rome. Anim. Behav. 1991;42:227–241. [Google Scholar]
- 26.Say L., Pontier D., Natoli E. Influence of oestrus synchronization on male reproductive success in the domestic cat (Felis catus L.) Proc. R. Soc. B. 2001;268:1049–1053. doi: 10.1098/rspb.2001.1583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Dreux P. Populations rurales de chats et effet urbain. Carn. Genet. Newsl. 1978;3:220–224. [Google Scholar]
- 28.Addie D.D., Schaap I.A., Nicolson L., Jarrett O. Persistence and transmission of natural type I feline coronavirus infection. J. Gen. Virol. 2003;84:2735–2744. doi: 10.1099/vir.0.19129-0. [DOI] [PubMed] [Google Scholar]
- 29.Bannasch M.J., Foley J.E. Epidemiologic evaluation of multiple respiratory pathogens in cats in animal shelters. J. Feline Med. Surg. 2005;7:109–119. doi: 10.1016/j.jfms.2004.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Coyne K.P., Dawson S., Radford A.D., Cripps P.J., Porter C.J., McCracken C.M., Gaskell R.M. Long-term analysis of feline calicivirus prevalence and viral shedding patterns in naturally infected colonies of domestic cats. Vet. Microbiol. 2006;118:12–25. doi: 10.1016/j.vetmic.2006.06.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Fromont E., Artois M., Pontier D. Cat population structure and circulation of feline viruses. Acta Oecol. 1996;17:609–620. [Google Scholar]
- 32.Courchamp F., Say L., Pontier D. Transmission of Feline Immunodeficiency Virus in a population of cats (Felis catus) Wildlife Research. 2000;27:603–611. [Google Scholar]
- 33.V. Xémar, Gestion sanitaire des populations de chats domestiques sur la ville de Nancy, Thèse vétérinaire, Université Lyon 1, 1997
- 34.Courchamp F., Yoccoz N.G., Artois M., Pontier D. At-risk individuals in feline immunodeficiency virus epidemiology: evidence from a multivariate approach in a natural population of domestic cats (Felis catus) Epidemiol. Infect. 1998;121:227–236. doi: 10.1017/s0950268898008875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Bendinelli M., Pistello M., Lombardi S., Poli A., Garzelli C., Matteucci D., Ceccherini-Nelli L., Malvaldi G., Tozzini F. Feline Immunodeficiency Virus: an interesting model for AIDS studies and an important cat pathogen. Clin. Microbiol. Rev. 1995;8:87–112. doi: 10.1128/cmr.8.1.87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Courchamp F., Pontier D. Feline immunodeficiency virus: an epidemiological review. C. R. Acad. Sci. III. 1994;317:1123–1134. [PubMed] [Google Scholar]
- 37.Diekmann O., Heesterbeek J.A.P. J. Wiley; Chichester, New York: 2000. Mathematical Epidemiology of Infectious Diseases: Model Building, Analysis, and Interpretation. (Wiley Series in Mathematical and Computational Biology). [Google Scholar]
- 38.Lloyd-Smith J.O., Cross P.C., Briggs C.J., Daugherty M., Getz W.M., Latto J., Sanchez M.S., Smith A.B., Swei A. Should we expect population thresholds for wildlife disease? Trends Ecol. Evol. 2005;20:511–519. doi: 10.1016/j.tree.2005.07.004. [DOI] [PubMed] [Google Scholar]
- 39.Bahi-Jaber N., Fouchet D., Pontier D. Stochastic extinction and the selection of the transmission mode in microparasites. J. Roy. Soc. Interface. 2008;5:1031–1039. doi: 10.1098/rsif.2007.1326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hilleman M.R. Strategies and mechanisms for host and pathogen survival in acute and persistent viral infections. Proc. Natl. Acad. Sci. USA. 2004;101:14560–14566. doi: 10.1073/pnas.0404758101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Scott F.W., Geissinger C. Duration of immunity in cats vaccinated with an inactivated feline panleucopenia, herpes-virus and calici virus vaccine. Feline Pract. 1997;25:12–19. [Google Scholar]
- 42.Radford A.D., Coyne K.P., Dawson S., Porter C.J., Gaskell R.M. Feline calicivirus. Vet. Res. 2007;38:319–335. doi: 10.1051/vetres:2006056. [DOI] [PubMed] [Google Scholar]
- 43.Csiza C.K., Scott F.W., De Lahunta A., Gillepsie J.H. Immune carrier state of feline panleucopenia virus-infected cats. Am. J. Vet. Res. 1971;32:419–426. [PubMed] [Google Scholar]
- 44.Berthier K., Langlais M., Auger P., Pontier D. Dynamics of a feline virus with two transmission modes within exponentially growing host populations. Proc. R. Soc. B. 2000;267:2049–2056. doi: 10.1098/rspb.2000.1248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Ikeda Y., Nakamura K., Miyazawa T., Takahashi E., Mochizuki M. Feline host range of canine parvovirus: recent emergence of new antigenic types in cats. Emerg. Infect. Dis. 2002;8:341–346. doi: 10.3201/eid0804.010228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Shackelton L.A., Parrish C.R., Truyen U., Holmes E.C. High rate of viral evolution associated with the emergence of carnivore parvovirus. Proc. Natl. Acad. Sci. USA. 2005;102:379–384. doi: 10.1073/pnas.0406765102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Truyen U., Parrish C.R. The evolution and control of parvovirus host ranges. Seminars in Virology. 1995;6:311–317. [Google Scholar]
- 48.Cohen C., Artois M., Pontier D. A discrete-event computer model of feline herpes virus within cat populations. Prev. Vet. Med. 2000;45:163–181. doi: 10.1016/s0167-5877(00)00132-x. [DOI] [PubMed] [Google Scholar]
- 49.Assogba B.D., Leavell S., Porter K., Burkhard M.J. Mucosal administration of low-dose cell-associated feline immunodeficiency virus promotes viral latency. J. Infect. Dis. 2007;195:1184–1188. doi: 10.1086/512861. [DOI] [PubMed] [Google Scholar]
- 50.Pepin A.C., Tandon R., Cattori V., Niederer E., Riond B., Willi B., Lutz H., Hofmann-Lehmann R. Cellular segregation of feline leukemia provirus and viral RNA in leukocyte subsets of long-term experimentally infected cats. Virus Res. 2007;127:9–16. doi: 10.1016/j.virusres.2007.03.008. [DOI] [PubMed] [Google Scholar]
- 51.Jarrett O. Strategies of retrovirus survival in the cat. Vet. Microbiol. 1999;69:99–107. doi: 10.1016/s0378-1135(99)00095-4. [DOI] [PubMed] [Google Scholar]
- 52.Rigby M.A., Holmes E.C., Pistello M., Mackay A., Brown A.J., Neil J.C. Evolution of structural proteins of feline immunodeficiency virus: molecular epidemiology and evidence of selection for change. J. Gen. Virol. 1993;74:425–436. doi: 10.1099/0022-1317-74-3-425. [DOI] [PubMed] [Google Scholar]
- 53.Hethcote H.W., Yorke J.A. vol. 56. Springer-Verlag; New York: 1984. Gonorrhea Transmission and Control. (Lecture Notes in Biomathematics). [Google Scholar]
- 54.O'Neil L.L., Burkhard M.J., Diehl L.J., Hoover E.A. Vertical transmission of feline immunodeficiency virus. AIDS Res. Hum. Retroviruses. 1995;11:171–182. doi: 10.1089/aid.1995.11.171. [DOI] [PubMed] [Google Scholar]
- 55.Jordan H.L., Howard J.G., Bucci J.G., Butterworth J.L., English R., Kennedy-Stoskopf S., Tompkins M.B., Tompkins W.A. Horizontal transmission of Feline Immunodeficiency Virus with semen from seropositive cats. Journal of Reproductive Immunology. 1998;41:341–357. doi: 10.1016/s0165-0378(98)00070-9. [DOI] [PubMed] [Google Scholar]
- 56.S. Devillard, T. Jombart, D. Pontier, Revealing cryptic genetic structures in a meta-population of stray cats Felis silvestris catus in a urban habitat, Mamm. Biol. (2008), in press, doi: 10.1016/j.mambio.2008.01.001
- 57.Pontier D., Fromont E., Courchamp F., Artois M., Yoccoz N.G. Retroviruses and sexual size dimorphism in domestic cats (Felis catus L.) Proc. R. Soc. B. 1998;265:167–173. doi: 10.1098/rspb.1998.0278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Takahata N., Nei M. Allelic genealogy under overdominant and frequency-dependent selection and polymorphism of Major Histocompatibility Complex Loci. Genetics. 1990;124:967–978. doi: 10.1093/genetics/124.4.967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Roelke M.E., Forester D.J., Jacobson E.R., Kollias G.V., Scott F.W., Barr M.C., Evermann J.F., Pirtel E.C. Seroprevalence of infectious disease agents in free ranging Florida panthers (Felis concolor coryi) J. Wildl. Dis. 1993;29:36–49. doi: 10.7589/0090-3558-29.1.36. [DOI] [PubMed] [Google Scholar]
- 60.Anderson R.M., May R.M. Vaccination and herd immunity to infectious diseases. Nature. 1985;318:323–329. doi: 10.1038/318323a0. [DOI] [PubMed] [Google Scholar]
- 61.Levin S.A., Pimentel D. Selection of intermediate rates of increase in parasite-host systems. Am. Nat. 1981;117:308–315. [Google Scholar]
- 62.Gandon S., Mackinnon M.J., Nee S., Read A.F. Imperfect vaccines and the evolution of pathogen virulence. Nature. 2001;414:751–756. doi: 10.1038/414751a. [DOI] [PubMed] [Google Scholar]
- 63.Aguas R., Goncalves G., Gomes M.G. Pertussis: increasing disease as a consequence of reducing transmission. Lancet Infect Dis. 2006;6:112–117. doi: 10.1016/S1473-3099(06)70384-X. [DOI] [PubMed] [Google Scholar]
- 64.Brisson M., Gay N.J., Edmunds W.J., Andrews N.J. Exposure to varicella boosts immunity to herpes-zooster: implications for mass vaccination against chickenpox. Vaccine. 2002;20:2500–2507. doi: 10.1016/s0264-410x(02)00180-9. [DOI] [PubMed] [Google Scholar]
- 65.Katzmann W., Dietz K. Evaluation of age-specific vaccination strategies. Theor. Pop. Biol. 1984;25:125–137. doi: 10.1016/0040-5809(84)90016-9. [DOI] [PubMed] [Google Scholar]
- 66.Graham A.L. Ecological rules governing helminth-microparasite coinfection. Proc. Natl. Acad. Sci. USA. 2008;105:566–570. doi: 10.1073/pnas.0707221105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Reubel G.H., George J.W., Higgins J., Pedersen N.C. Effect of chronic feline immunodeficiency virus infection on experimental feline calicivirus-induced disease. Vet. Microbiol. 1994;39:335–351. doi: 10.1016/0378-1135(94)90169-4. [DOI] [PubMed] [Google Scholar]
- 68.Tillmann H.L., Heiken H., Knapik-Botor A., Heringlake S., Ockenga J., Wilber J.C., Georgen B., Detmer J., McMorrow M., Stoll M., Schmidt R.E., Manns M.P. Infection with GB virus C and reduced mortality among HIV-infected patients. New England J. Med. 2001;345:715–724. doi: 10.1056/NEJMoa010398. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.