

Abstract
Ageing reduces the functional capacity of all organs, so does that of the nervous system; the latter is evident in the reduction of cognitive abilities, learning and memory. While the exact mechanisms of ageing of the nervous system remain elusive, it is without doubt that morpho-functional changes in a variety of neuroglial cells contribute to this process. The age-dependent changes in neuroglia are characterised by a progressive loss of function. This reduces glial ability to homeostatically nurture, protect and regenerate the nervous tissue. Such neuroglial paralysis also facilitates neurodegenerative processes. Ageing of neuroglia is variable and can be affected by environmental factors and comorbidities.
Keywords: Ageing, Astrocyte, Microglia, NG2 cells, Oligodendrocytes
8.1. Mechanisms of Ageing
Ageing reduces the functional capacity of all organs and systems ultimately weakening the whole organism, reducing its adaptability, wearing its defensive systems and bringing it to death through age-dependent diseases. The nervous system similarly undergoes senescence that often impairs upon cognitive abilities, affects learning capabilities and enfeebles memory. Nonetheless, brain sustains ageing with surprising tenacity; the cognitive functions attain the summit in middle age and often sustain into old age, while the decline in other systems (including skeleto-muscular, cardiovascular or endocrine) progresses much faster. What are the mechanisms of ageing and which molecular and cellular processes underlie the ageing of the brain remains a matter of intense polemics. Indeed, there are in excess of 300 theories of ageing, which highlight various pathways, many of which do contribute to this process [52].
Probably, the very first coherent theory of ageing was proposed by August Weismann [99], who considered ageing as a very natural process favoured by natural selection to prevent competition of species with their own progeny. According to this view, mechanisms of ageing could be many and they can be different in different organisms. The only common future is guaranteed termination of individual life after a presumed fulfilment of the reproductive duties. One of the widely considered pathways is the activation of endoplasmic reticulum stress/unfolded protein responses that positively correlate with longevity and negatively with fertility [86]. Another relatively old group of theories regards ageing as a result of mechanochemical deterioration of molecules and basic systems such as organelles or cell membranes; this was initially proposed as ‘hysteresis of colloids’ [72, 73]. The mitochondrial or free radical theory of ageing links the damage of biological systems to reactive oxygen species and regards mitochondria as the age-defining clock [29, 30]. By reducing caloric intake this may resist the rate of ageing [48]. The gene regulation theories assume that changes in gene expression define ageing process [39], while the telomere theory postulates that it is the telomere DNA localised at the end of chromosomes which determines the life span: the shortening of telomeres eventually brings to the arrest of cell replication and death [28], The inflammatory theory of ageing became popular in recent years leading to the concept of ‘inflammaging’ [21]. Additionally, several theories look into the role of signalling systems both at organism (e.g., neuroendocrine or immune theories of ageing [20]) and cellular (e.g., calcium theory of ageing [40, 45, 89]) levels.
8.2. Ageing of the Brain
The maintenance of cognitive capacity of the brain over most of the human’s lifespan results, most likely, from prominent neuroplasticity, remarkably long development and high degree of homeostatic and protective capabilities of neuroglia. The human brain is optimised for learning, with numerous mechanisms from adult neurogenesis (which supplies the hippocampus with new neurones [54]) and adult myelination (which lasts well into the fourth decade of human life [1, 98], while oligodendroglial progenitors are present throughout the brain across the whole lifespan and probably contribute to late-life regenerative myelination), to the highly sophisticated glymphatic system that purges the brain from toxic waste products [36], thus maintaining neural environment. Ageing affects cognition components in a rather distinct way. The age-dependent decline mainly affects the real-time processing and formation of new memories and behaviours, whereas the capacity to analyse semantic and long-term memories suffer much less [18, 35]. For example, a group of young adults were significantly better than the group of old people in recalling a list of words. However the ability of elders to use complex processing activities was indistinguishable from the youngsters [47]. This benign or physiological brain ageing is not granted to all, and age-dependent pathologies, most notably of neurodegenerative nature, affect a substantial part of population.
Age is the main risk factor for neurodegenerative diseases, which are often considered as a natural outcome of senescence process. However, there is a fundamental difference between physiological ageing and neurodegeneration. The latter reflects massive neuronal death and atrophy of the brain tissue, whereas the former is not associated with a substantial neuronal loss. The overall number of neurones is not significantly affected in physiological ageing in rodents, primates and humans [6, 12, 19, 100]. Likewise, the number and density of synapses are not significantly affected by ageing [23, 80], albeit synaptic size is reduced [56].
Factors which determine the fateful difference between physiological and pathological ageing are many. These are represented by genetic factors (the best example being familial Alzheimer’s disease or Huntington disease), the environment and life style (including diet, education, mental or physical activity) and the associated pathology (such as vascular disorders and ischaemic lesions). Another fundamental factor that defines the degree of cognitive deficit of ageing and age-dependent neuropathologies is known as the cognitive reserve. The cognitive reserve is an intrinsic quality of an individual brain that determines the neurological deficit when a similar brain damage results in very different cognitive outcomes in different subjects [82, 103]. The cognitive reserve in turn is defined by (i) neuronal reserve, which is the status of neuronal networks acquired during the life span through learning and cognitive load and (ii) neuronal compensation that reflects the defensive, plastic and regenerative capacities of the individual brain. To a large extent, the neuronal compensation is defined by neuroglia, which is responsible for neuroprotection, regeneration and post-lesion remodelling of the neural circuitry. The role of neuroglia is therefore fundamental in defining physiological versus pathological senescence; the failure of glial cells to protect and sustain the neuronal networks, the neural tissue and the CNS as an organ facilitates the progression from physiological to pathological brain ageing [93].
8.3. Astroglia in Physiological Ageing
8.3.1. Morphology and Gene Profiling
Age-dependent changes in astroglial morphology and gene expression are complex and region specific. Total number of astrocytes in physiologically aged human brain does not seem to change significantly, even in centenarians [19, 59]. When it comes to astroglial morphological profiles and expression of glial fibrillary acidic protein (GFAP), which are indicative of astroglial reactivity, the data remain quite controversial. Both a decrease [7] and an increase [14] in the number of GFAP-positive astrocytes, in particular in hypothalamic areas [27], as well as astroglial atrophy and astroglial hypertrophy were observed. The volume of astroglial territorial domains has been found to almost double in 21-month-old mice when compared to 5-month-old animals [26]. Increase of GFAP expression and hypertrophy of GFAP-positive astrocytes have been described in the hippocampus of aged rodents [7, 34, 49] and humans [10, 55]. Ageing had a distinct effect on different subpopulation of astrocytes in a region-dependent manner (Fig. 8.1, [69]). The densities of GFAP-positive astrocytes in the CA1 region and dentate gyrus of the hippocampus of old (24-month-old) mice demonstrated prominent hypertrophy when compared to young (3-month-old) or adult (9-month-old) controls. To the contrary, GFAP-positive profiles of astrocytes in the entorhinal cortex of old animals were atrophic when compared to the young or adult mice. Ageing results in a substantial decrease in the number and complexity of processes of astrocytes in the entorhinal cortex. The astrocytes immunoreactive to s100β protein were hypertrophic in the aged dentate gyrus but not in the CA1 region of the hippocampus as well as in the entorhinal cortex, whereas the profiles of a subpopulation of astrocytes labelled with glutamine synthetase were atrophic in the hippocampus with no changes in the entorhinal cortex [69]. Glutamine synthetase is a central enzyme necessary for operation of glutamine–glutamate/GABA shuttle, as well as for ammonium detoxification [70]. Suppression of expression of this enzyme may therefore affect neurotransmission and promote astroglial synthesis of GABA, an inhibitory neurotransmitter [22]. In parallel, hypertrophy of GFAP-positive astrocytes may be connected with environmental stimulation and plasticity representing the neural compensation. Exposure to the enriched environment is known for its positive effects on learning and memory, which occur in parallel with an increase in GFAP-positive astroglial profiles [68, 75].
Fig. 8.1.
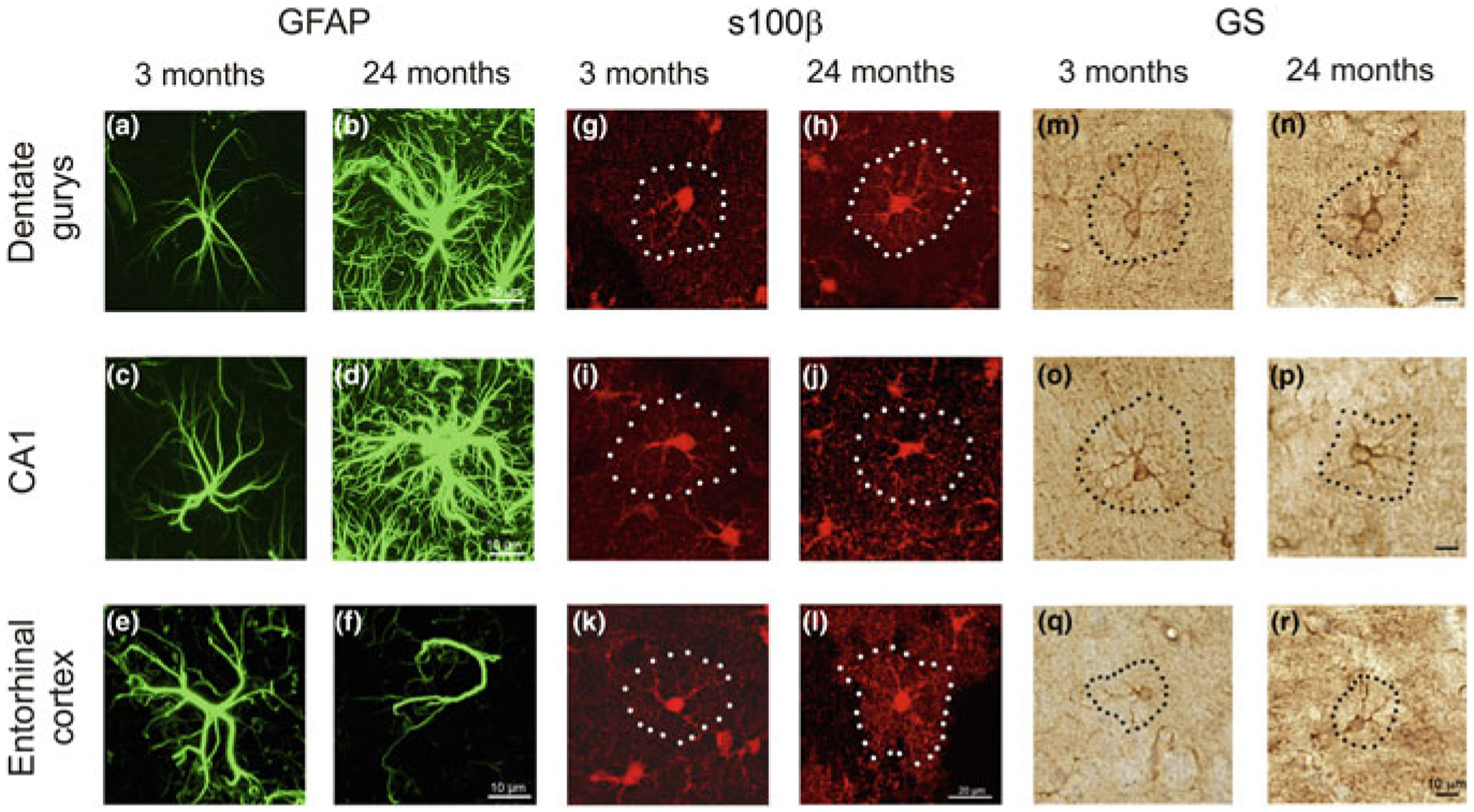
Age-dependent remodelling of astroglial profiles in different brain areas. Confocal images showing glial fibrillary acidic protein—GFAP (a to f), s100β (g to l) and glutamine synthetase—GS (m to r) immunolabelled astrocytes in the dentate gyrus and CA1 hippocampal areas as well as in the entorhinal cortex of mice at 3 and 24 months. Modified from [69]
The transcriptomic analysis of aged astrocytes similarly found a complex modification in genes expression. For example, astroglial cells from the cerebral cortex of aged mice demonstrated an increase in genes related to immune response with a decrease in expression of GFAP and genes related to neuroprotection and neuronal support [58]. Comparison of RNA-Seq from old and young astrocytes in the motor and visual cortices, hypothalamus and cerebellum revealed region specificity, with more significant changes in astroglia from the hippocampus and cerebellum [4], where astrocytes increased an expression of proinflammatory genes, genes encoding GFAP and Serpin3n, and genes linked to synaptic elimination such as complement component 3 and 4b [4]. Very similar results have been obtained in analysing the gene expression profiles of astrocytes from the hippocampus, cortex and striatum. Ageing affected hippocampal and striatal astrocytes the most with up-regulation of inflammatory genes and genes related to synaptic elimination [8]. Analysis of the gene expression profiles of different brain cells from ten brain regions of post-mortem tissue of humans, aged between 16 and 102 years, found that changes in astrocytes and oligodendrocytes were more prominent and complex compared to other cell types [81]; in particular, no age-dependent changes in neuronal gene expression pattern were identified. Again, these results indicate that functional preservation of neuroglia is critical for maintaining the ageing brain.
8.3.2. Astroglial Function
Although the data on physiology of aged astrocytes are rather limited, there are some hints for age-dependent remodelling of signalling and homeostatic profiles of astroglial cells. The resting membrane potential (around −80 mV) and membrane input resistance of astrocytes in cortical slices (from animals aged between 1 and 21 months) does not change much in ageing; if anything the input resistance is some-what smaller in young adult mice (3–6-month-old—Fig. 8.2, [44]). Astrocytes from older mice express major types of receptors and are capable to generate ionotropic receptor-driven glial “postsynaptic” currents in response to neuronal activity [24, 44]. The density of ionotropic glutamate (AMPA and NMDA) receptors and P2X purinoceptors, as well as the density of plasmalemmal glutamate transporter currents demonstrate bell-shaped age dependency (Fig. 8.3). Ionic currents generated by the above receptors and transporters are maximal in young adult (3- to 6-month-old) animals; at 9–21 months of age these currents are much smaller, although they are similar to currents recorded in 1-month-old animals [44].
Fig. 8.2.
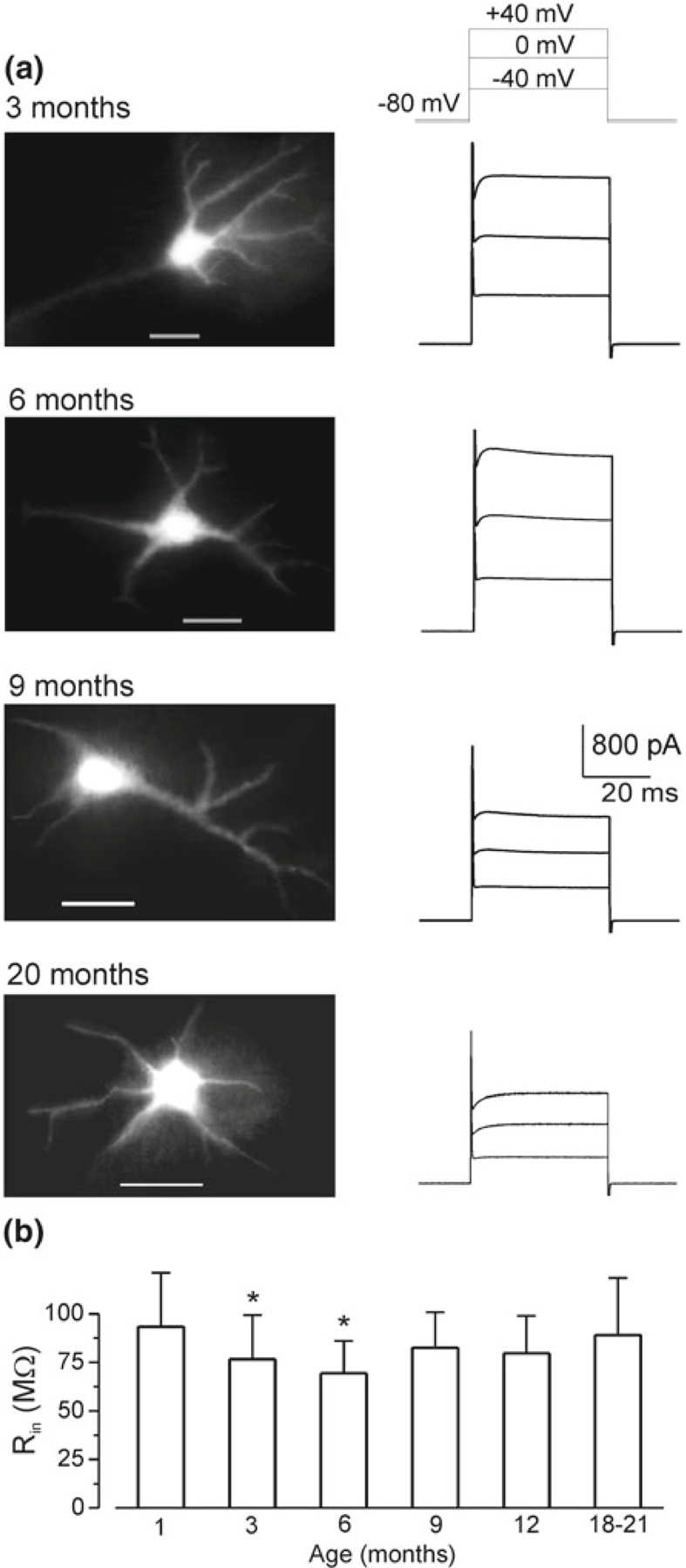
Basic electrophysiological properties of cortical astrocytes in brain slices from mice of different ages. a Experiments were performed on mice expressing EGFP under control of human GFAP promoter. Representative EGFP images (left column) of cortical layer II astrocytes of different age groups and corresponding whole-cell currents (right column) evoked by depolarizing steps from the holding potential of −80 mV; the voltage protocol is shown on the top. b Age-related changes in the input resistance of cortical astrocytes. Data are presented as mean ± SD for 15 cells in each age group; decrease in the astrocyte Rin measured in 3 and 6 months in comparison to 1 month was statistically significant with P < 0.05 (one-way ANOVA). Reproduced with permission from [44]
Fig. 8.3.
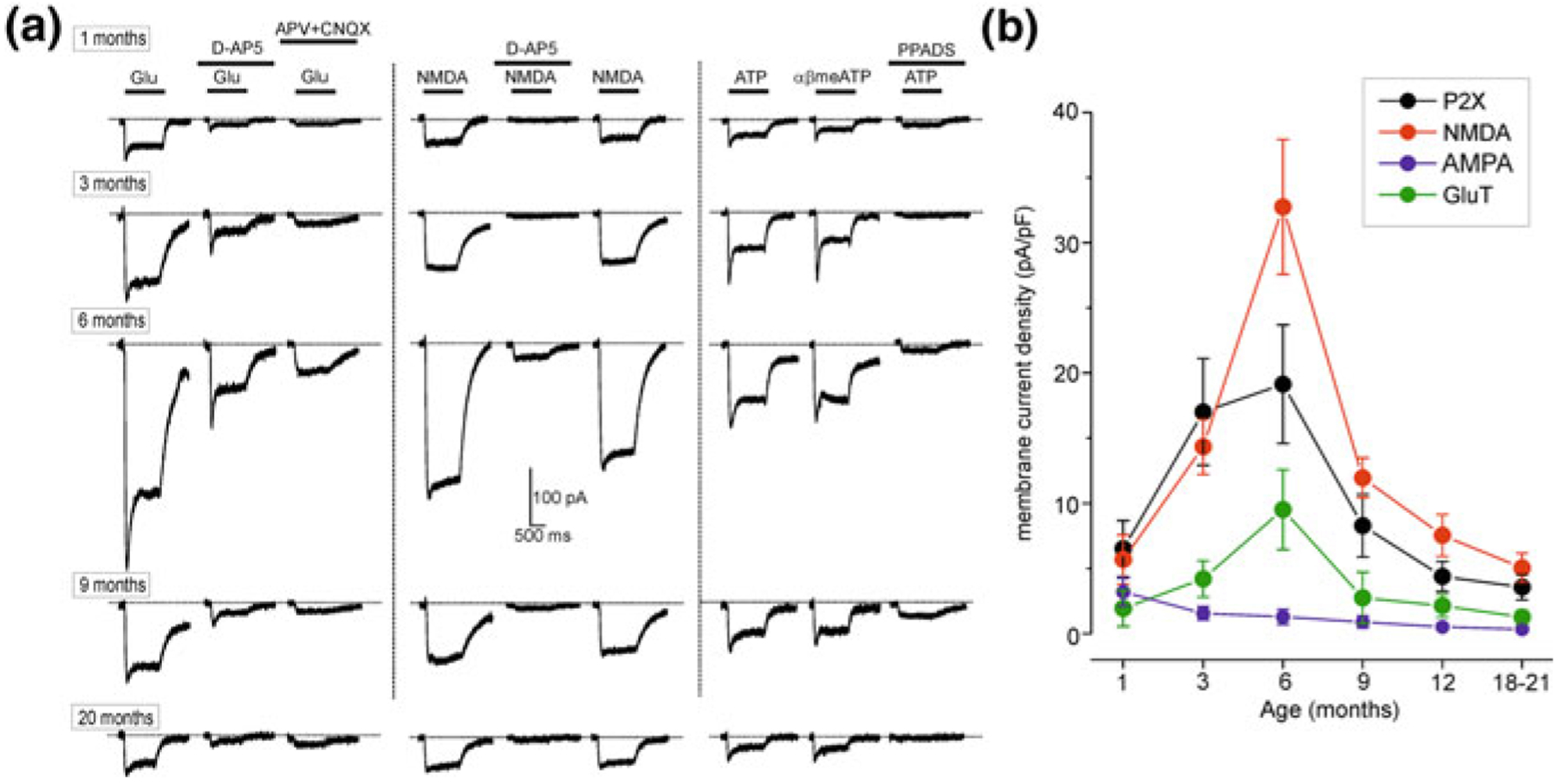
Ageing affects the density of plasmalemmal glutamate transporters and ionotropic receptor-mediated currents in acutely isolated single cortical astrocytes. a Representative whole-cell currents elicited in the acutely isolated astrocytes by application of 100 μM glutamate (left column), 10 μM NMDA (middle column) and 10 μM ATP/αβmeATP (a potent and stable agonist at P2X1, P2X3, P2X2/3, P2X1/5 and P2X4/6 receptors is also a weak partial agonist at human and mouse P2X4 receptors, but an antagonist at the rat P2X4 receptor; it has little or no effect at other P2X and P2Y receptors), at holding potential of −80 mV. Glutamate- and NMDA-evoked currents were inhibited by 10 μM D-AP5, an NMDA antagonist and 30 μM CNQX, an AMPA receptor antagonist; ATP-evoked currents were inhibited by 10 μM PPADS, a selective purinergic P2X antagonist. b The density of currents mediated by P2X, NMDA and AMPA receptors and plasmalemmal glutamate transporters (GluT) in cortical astrocytes (mean ± SD for 9–12 cells for each age group); statistical significance of difference between average value for 1 month and corresponding values for 3 and 6 months P < 0.02 (ANOVA) for all types of currents
Astrocytes are endowed with specific type of excitability, known as ionic excitability, which is associated with spatially and temporally organised fluctuations in the cytosolic concentration of several ions, including Ca2+, Na+, Cl− and possibly K+ and H+ [95, 97]. Intracellular Ca2+ and Na+ signalling are of particular importance [95, 96] being involved in regulation of numerous astroglial physiological processes such as secretion [94] or homeostatic transport [41, 71]. Neurotransmitter or synaptically induced astroglial Ca2+ signals are age dependent. For example, Ca2+ signals are the largest in young adult mice and are relatively small in old and very young animals (Fig. 8.4; [44]). This dependence may be reflected in the functional expression of astrocytic receptors. Most likely an increase in the density of receptors, as well as in the density of plasmalemmal glutamate transporters and in the amplitude of Ca2+ signals, occur in the period of maximal environmental stimulation associated with intense learning; in younger and older ages synaptic activity is lower, which is reflected in a decrease in receptors expression.
Fig. 8.4.
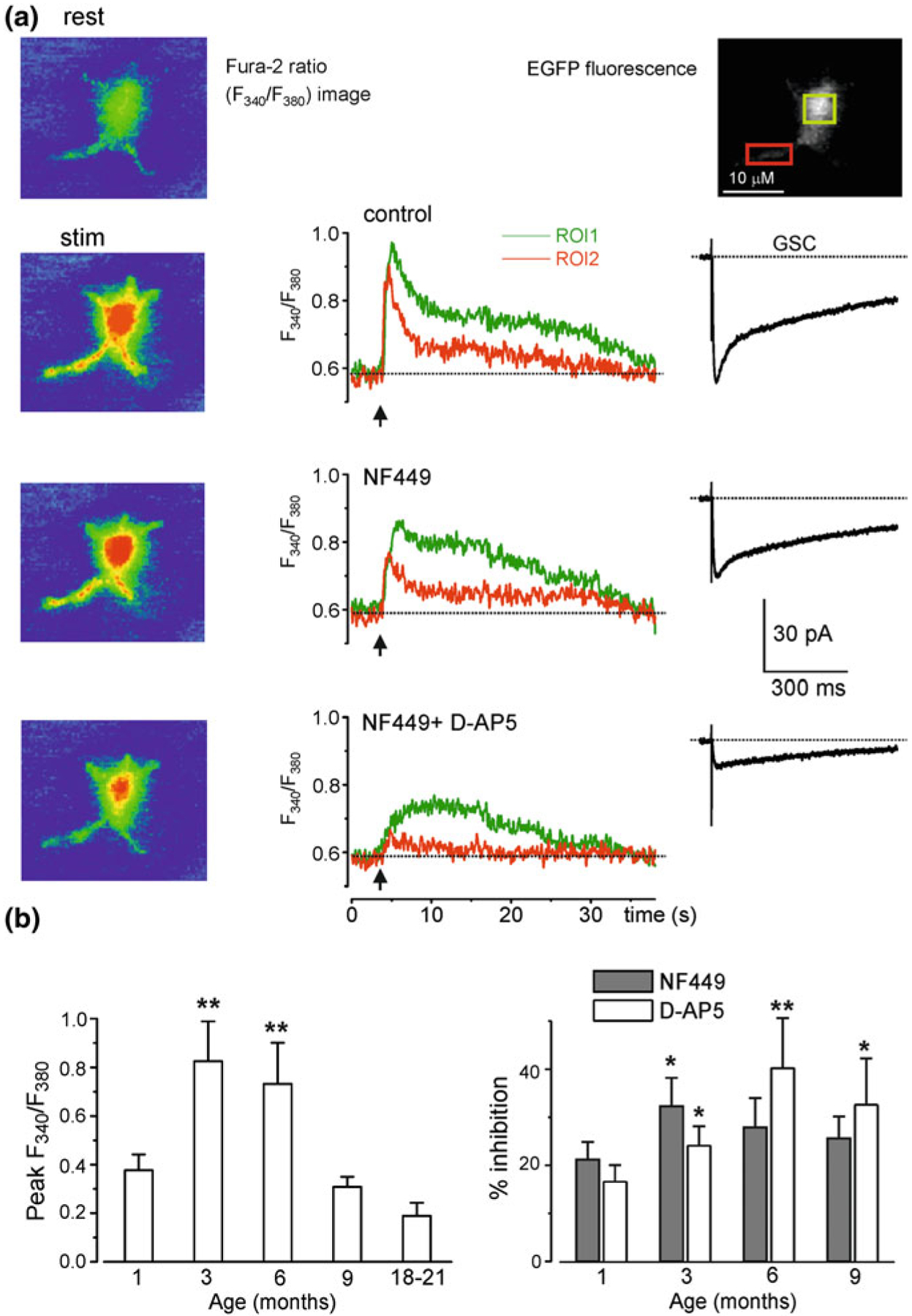
Age-dependent changes in synaptically induced ionotropic Ca2+ signals in protoplasmic astrocytes in situ in cortical slices. a Cortical layer II astrocyte of 9-month-old mouse was loaded with the Ca2+ indicator Fura-2 in situ via patch pipette. Fluorescence images were recorded simultaneously with glial currents evoked by neuronal afferent stimulation in presence of a mixture of TBOA (GluT blocker) and CNQX in control, and after consecutive application of 10 nM NF-449 (selective antagonist of P2X receptors) and at that in a company of 30 μM D-AP5. Representative images (pseudo-colour, pipette image subtracted) and glial synaptic currents (GSC, right column) were recorded before (rest) and after stimulation as indicated. Ca2+ transients (middle column) are expressed as F340/F380 ratio averaged over the corresponding regions of interest shown in the GFAP image of astrocyte (top right). b Age-related changes in the astrocytic Ca2+ signalling. Left panel, average peak amplitudes of [Ca2+]i increases, induced by stimulation of neuronal afferents in cortical astrocytes of different ages. Right panel, average inhibitory effect of antagonists of P2X (NF449) and NMDA (D-AP5) receptors on the amplitudes of [Ca2+]i increases responses in cortical astrocytes. Data are presented as mean ± SD for 3–4 cells for each age group; * P < 0.05, ** P < 0.01 one-way ANOVA compared to 1 month. Reproduced with permission from [44]
A decrease in astroglial gap junctional coupling was found in old (20–27-month-old mice) neocortical astroglial syncytia [63]; there were no changes at earlier ages (up to 14-month-old [9]). Astrocytes in older brains down-regulate expression of aquaporin 4 (AQP4). A decrease in the density of these channels in the perivascular endfeet affects clearance of the brain parenchyma through the glymphatic pathway [42]. This decrease in AQP4 in the endfeet may be linked to the deficits in vesicular trafficking, which is the key pathway in delivery of numerous molecules to specific locations at the plasmalemma [64]. Ageing affects astroglial metabolic pathways, as an age dependent increase in oxidative metabolism was reported in older astrocytes, which may limit their ability to supply neurones with metabolic substrates [37]. There is also evidence of age-dependent alterations in astroglial ability to produce lactate and hence to operate lactate shuttle [31]. Similarly, ageing is associated with an increase in the ratio of glutamate to glutamine in the brain that indicates some aberrations in the operation of the glutamate/GABA-glutamine shuttle [16, 32]. Ageing is also associated with a decrease in the brain levels of glutathione, mainly produced in astrocytes; this limits the ability of astroglia to resist the oxidative damage to the neural tissue [17, 50].
8.4. Oligodendroglia in Physiological Ageing
The human brain has a disproportionally large white matter when compared to other mammals and even high primates [76], as indeed the white matter occupies >50% of the human brain. Additionally, the level of myelination is well developed in the grey matter [43], further demonstrating the importance of connectome to the cognition and intelligence. The anatomical prevalence of the white matter in the human brain is also associated with very long development: myelination attains its peak at ~45–47 years of age, with a subsequent slow and yet progressive age-dependent decline [1]. Normal ageing causes rather substantial shrinkage of the white matter which diminishes by ~11%; in comparison, the volume of the grey matter is decreased by only ~3% [33]. The highest degree of age-dependent alterations of the white matter is detected in the prefrontal cortex and associative tracts [66], which suffer early in Alzheimer’s disease [15]. Incidentally, these brain regions emerge late in evolution and they are the slowest to develop, which instigated a ‘last in, first out’ hypothesis of the white matter ageing [66, 90]. Conceptually, changes in the white matter can be considered as a valuable marker of ageing [90], and moreover, accelerated degeneration of the white matter seems to indicate development of neurodegeneration and profound cognitive decline [65, 90].
Cells of the oligodendroglial lineage represented by oligodendrocytes and their precursors (also known as NG2 glia [13]) are, arguably, the most numerous glial cells in the human brain. Cells of the oligodendroglial lineage, in contrast to astrocytes, are highly vulnerable to excitotoxicity and to oxidative stress. The oligodendroglial precursors/NG2 cells, as well as more mature oligodendroglia, express several types of ionotropic glutamate receptors (including NMDA receptors) and P2X purinoceptors, which all can mediate excitotoxic Ca2+ overload and cause cell death [51, 53, 74, 92]. Furthermore, oligodendrocytes are highly vulnerable to oxidative damage, which is stipulated by a rather low content of antioxidants. In particular oligodendroglial cells contain two times less of glutathione compared to astrocytes, and yet they experience six times more of oxidative stress in physiological conditions [38, 87].
Ageing is associated with a significant decrease, by up to 30%, of the total number of oligodendrocytes [19, 59]. Rather surprisingly, in monkeys the number of oligodendroglial cells has been claimed to increase with age; for example, in the visual cortex of old monkeys the number of oligodendrocytes increased by 50% [62]. Notably, these oligodendrocytes also showed aberrant atrophic morphology and a deficiency in myelin production, which defined decreased CNS myelination in old primates [62]. The age-dependent myelin deficiencies are also associated with vasculature lesions in the white matter that add strain on oligodendrocytes and promote their degeneration [3, 101]. Ageing is also associated with a diminished capacity of remyelination supported by the NG2 glia. Notwithstanding the fact that the population of NG2-oligodendroglial precursors does not change numerically in the old brain, the capacity of NG2 cells to differentiate into mature oligodendrocytes is reduced. The NG2 cells in the old brain tend to retain their precursor status, so that the time of differentiation into mature myelinating phenotype is increased by almost two times [102]. All in all, age-dependent changes in the white matter are prominent and may be the leading cause of age-dependent cognitive decline.
8.5. Microglia in the Ageing Brain
Microglia in the ageing human brain undergoes rather idiosyncratic metamorphoses, which are not present in laboratory animals. Fundamentally, human microglia gradually degenerates, thus, reducing the defensive capabilities of the senescent nervous tissue.
In animals, the ageing process results in complex changes in microglial numbers and state. In old rats, microglial numbers decreased in the nigrostriatal system and cerebral cortex [77], and remained unchanged in the hippocampus [91]. In contrast, in old rhesus monkeys the densities of microglial cells increased, while these cells showed signs of increased phagocytosis [61]. In humans, ageing is associated with dystrophy and degeneration of microglia which resulted in deterioration of neuro-protective and defensive functions of these cells [84]. Morphological features of dystrophic aged microglia include deramification, spheroid formation, gnarling and fragmentation of processes [84]. The processes of aged microglial cells are shorter with less branching and reduced arborized area; the total number of microglia seems not to change with age [11]. Microglial dystrophy and a loss of function arguably increase the vulnerability of the old brain to neurodegeneration and may facilitate evolution of age-dependent cognitive disorders, including Alzheimer’s disease [83]. The age-dependent microglial dystrophy can be associated with cytoskeleton abnormalities that underlie the cytorrhexis, rupturing of cells [88]. Microglial cells can accumulate tau [5] and the aged microglia (in marmosets) were reported to contain hyperphosphorylated tau [67]; this microgliatauopathy can be a factor that initiates microglial degeneration and dystrophy [67]. The prevalence of dystrophic microglia limits the neuroinflammatory capabilities of the old brain tissue, questioning the concept of inflammaging.
There is also evidence for age-dependent microglial activation in normal ageing, especially in rodents [57, 60] and in Macaca nemestrina monkeys [78]. There is an overall trend of hyperreactivity of microglia in aged mice [25, 46, 79], which is strikingly different to the dystrophy and a loss of function of human aged microglia, questioning the validity of rodents as an experimental models for brain ageing.
Aged human microglial cells are represented by two morphologically distinct classes identified as dystrophic or senescent microglia and dark microglia. The dystrophic microglial cells [85] are characterised by spherical swellings of processes, dilatation of the endoplasmic reticulum and abundance of lipofuscin deposits (that emerge from incomplete lysosomal degradation and endolysosomal stress and overload). The dystrophic microglial cells have been identified both in old brains and in high densities around senile plaques of Alzheimer’s diseases patients [88]. Dystrophic microglial cells have fragmented processes and have a substantially diminished activation capacity [85, 88]. The dark microglia have been defined so because of the electron-dense cytoplasm and nucleoplasm, which in electron microscopy appear as dark as mitochondria [2]. The dark microglia are also characterised by ultrathin and highly ramified processes that frequently enwrap synaptic elements, axons and dendrites. This may indicate that dark microglial cells are involved in eliminating synapses [2]. In addition, dark microglia have altered expression of classical marker IBA1 and they do not express the P2Y12 purinoceptor, which is considered as a marker for healthy surveillance microglia. Dark microglial cells cumulate with ageing and even more so in age-dependent pathologies [2].
8.6. Conclusions
All types of neuroglial cells undergo age-dependent remodelling which seems to be critical to define a physiological or pathological outcome of ageing process. In general, the age-dependent changes in neuroglial cells are characterised by a progressive loss of function which limits neuroprotection and regenerative potential of the neural tissue. This process of neuroglial senescence, however, is variable and most likely individually tailored by the lifestyle, environmental stress and comorbidities. Neuroglial paralysis facilitates emergence of neurodegeneration and cognitive decline, and hence a neuroglial state represents a potential therapeutic target for age-associated neurological disorders.
Acknowledgments
VP’s work is supported by a grant from the National Institute of General Medical Sciences of the National Institutes of Health (R01GM123971). VP is an Honorary Professor at University of Rijeka, Croatia.
Contributor Information
Alexei Verkhratsky, Faculty of Biology, Medicine and Health, The University of Manchester, Manchester M13 9PT, UK; Faculty of Health and Medical Sciences, Center for Basic and Translational Neuroscience, University of Copenhagen, 2200 Copenhagen, Denmark; Achucarro Center for Neuroscience, IKERBASQUE, Basque Foundation for Science, 48011 Bilbao, Spain.
Robert Zorec, Laboratory of Neuroendocrinology-Molecular Cell Physiology, Faculty of Medicine, Institute of Pathophysiology, University of Ljubljana, Ljubljana, Slovenia; Celica BIOMEDICAL, Ljubljana, Slovenia.
Jose Julio Rodriguez-Arellano, Achucarro Center for Neuroscience, IKERBASQUE, Basque Foundation for Science, 48011 Bilbao, Spain.
Vladimir Parpura, Department of Neurobiology, The University of Alabama at Birmingham, Birmingham, AL, USA.
References
- 1.Bartzokis G, Beckson M, Lu PH, Nuechterlein KH, Edwards N, Mintz J (2001) Age-related changes in frontal and temporal lobe volumes in men: a magnetic resonance imaging study. Arch Gen Psychiatry 58:461–465 [DOI] [PubMed] [Google Scholar]
- 2.Bisht K, Sharma KP, Lecours C, Sanchez MG, El Hajj H, Milior G, Olmos-Alonso A, Gomez-Nicola D, Luheshi G, Vallieres L, Branchi I, Maggi L, Limatola C, Butovsky O, Tremblay ME (2016) Dark microglia: a new phenotype predominantly associated with pathological states. Glia 64:826–839 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Black S, Gao F, Bilbao J (2009) Understanding white matter disease: imaging-pathological correlations in vascular cognitive impairment. Stroke 40:S48–52 [DOI] [PubMed] [Google Scholar]
- 4.Boisvert MM, Erikson GA, Shokhirev MN, Allen NJ (2018) The aging astrocyte transcriptome from multiple regions of the mouse brain. Cell Rep 22:269–285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bolos M, Llorens-Martin M, Jurado-Arjona J, Hernandez F, Rabano A, Avila J (2016) Direct evidence of internalization of tau by microglia in vitro and in vivo. J Alzheimer’s Dis 50:77–87 [DOI] [PubMed] [Google Scholar]
- 6.Calhoun ME, Kurth D, Phinney AL, Long JM, Hengemihle J, Mouton PR, Ingram DK, Jucker M (1998) Hippocampal neuron and synaptophysin-positive bouton number in aging C57BL/6 mice. Neurobiol Aging 19:599–606 [DOI] [PubMed] [Google Scholar]
- 7.Cerbai F, Lana D, Nosi D, Petkova-Kirova P, Zecchi S, Brothers HM, Wenk GL, Giovannini MG (2012) The neuron-astrocyte-microglia triad in normal brain ageing and in a model of neuroinflammation in the rat hippocampus. PLoS ONE 7:e45250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Clarke LE, Liddelow SA, Chakraborty C, Munch AE, Heiman M, Barres BA (2018) Normal aging induces A1-like astrocyte reactivity. Proc Natl Acad Sci USA 115:E1896–E1905 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cruz NF, Ball KK, Dienel GA (2010) Astrocytic gap junctional communication is reduced in amyloid-beta-treated cultured astrocytes, but not in Alzheimer’s disease transgenic mice. ASN Neuro 2:e00041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.David JP, Ghozali F, Fallet-Bianco C, Wattez A, Delaine S, Boniface B, Di Menza C, Dela-courte A (1997) Glial reaction in the hippocampal formation is highly correlated with aging in human brain. Neurosci Lett 235:53–56 [DOI] [PubMed] [Google Scholar]
- 11.Davies DS, Ma J, Jegathees T, Goldsbury C (2017) Microglia show altered morphology and reduced arborization in human brain during aging and Alzheimer’s disease. Brain Pathol 27:795–808 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Dickstein D, Kabaso D, Rocher A, Luebke J, Wearne S, Hof P (2006) Changes in the structural complexity of the aged brain. Aging Cell 6:275–284 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dimou L, Gallo V (2015) NG2-glia and their functions in the central nervous system. Glia 63:1429–1451 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Diniz DG, Foro CA, Rego CM, Gloria DA, de Oliveira FR, Paes JM, de Sousa AA, Tokuhashi TP, Trindade LS, Turiel MC, Vasconcelos EG, Torres JB, Cunnigham C, Perry VH, Vasconcelos PF, Diniz CW (2010) Environmental impoverishment and aging alter object recognition, spatial learning, and dentate gyrus astrocytes. Eur J Neurosci 32:509–519 [DOI] [PubMed] [Google Scholar]
- 15.Douaud G, Groves AR, Tamnes CK, Westlye LT, Duff EP, Engvig A, Walhovd KB, James A, Gass A, Monsch AU, Matthews PM, Fjell AM, Smith SM, Johansen-Berg H (2014) A common brain network links development, aging, and vulnerability to disease. Proc Natl Acad Sci USA 111:17648–17653 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Duarte JM, Do KQ, Gruetter R (2014) Longitudinal neurochemical modifications in the aging mouse brain measured in vivo by 1H magnetic resonance spectroscopy. Neurobiol Aging 35:1660–1668 [DOI] [PubMed] [Google Scholar]
- 17.Emir UE, Raatz S, McPherson S, Hodges JS, Torkelson C, Tawfik P, White T, Terpstra M (2011) Noninvasive quantification of ascorbate and glutathione concentration in the elderly human brain. NMR Biomed 24:888–894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Erickson CA, Barnes CA (2003) The neurobiology of memory changes in normal aging. Exp Gerontol 38:61–69 [DOI] [PubMed] [Google Scholar]
- 19.Fabricius K, Jacobsen JS, Pakkenberg B (2013) Effect of age on neocortical brain cells in 90+ year old human females—a cell counting study. Neurobiol Aging 34:91–99 [DOI] [PubMed] [Google Scholar]
- 20.Fabris N (1991) Neuroendocrine-immune interactions: a theoretical approach to aging. Arch Gerontol Geriatr 12:219–230 [DOI] [PubMed] [Google Scholar]
- 21.Franceschi C (2007) Inflammaging as a major characteristic of old people: can it be prevented or cured? Nutr Rev 65:S173–176 [DOI] [PubMed] [Google Scholar]
- 22.Garaschuk O, Verkhratsky A (2019) GABAergic astrocytes in Alzheimer’s disease. Aging (Albany NY) 11:1602–1604 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Geinisman Y, Ganeshina O, Yoshida R, Berry RW, Disterhoft JF, Gallagher M (2004) Aging, spatial learning, and total synapse number in the rat CA1 stratum radiatum. Neurobiol Aging 25:407–416 [DOI] [PubMed] [Google Scholar]
- 24.Gomez-Gonzalo M, Martin-Fernandez M, Martinez-Murillo R, Mederos S, Hernandez-Vivanco A, Jamison S, Fernandez AP, Serrano J, Calero P, Futch HS, Corpas R, Sanfeliu C, Perea G, Araque A (2017) Neuron-astrocyte signaling is preserved in the aging brain. Glia 65:569–580 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Griffin R, Nally R, Nolan Y, McCartney Y, Linden J, Lynch MA (2006) The age-related attenuation in long-term potentiation is associated with microglial activation. J Neurochem 99:1263–1272 [DOI] [PubMed] [Google Scholar]
- 26.Grosche A, Grosche J, Tackenberg M, Scheller D, Gerstner G, Gumprecht A, Pannicke T, Hirrlinger PG, Wilhelmsson U, Huttmann K, Hartig W, Steinhauser C, Pekny M, Reichenbach A (2013) Versatile and simple approach to determine astrocyte territories in mouse neocortex and hippocampus. PLoS ONE 8:e69143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hardy RN, Simsek ZD, Curry B, Core SL, Beltz T, Xue B, Johnson AK, Thunhorst RL, Curtis KS (2018) Aging affects isoproterenol-induced water drinking, astrocyte density, and central neuronal activation in female Brown Norway rats. Physiol Behav 192:90–97 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Harley CB, Vaziri H, Counter CM, Allsopp RC (1992) The telomere hypothesis of cellular aging. Exp Gerontol 27:375–382 [DOI] [PubMed] [Google Scholar]
- 29.Harman D (1965) The free radical theory of aging: effect of age on serum copper levels. J Gerontol 20:151–153 [DOI] [PubMed] [Google Scholar]
- 30.Harman D (1972) The biologic clock: the mitochondria? J Am Geriatr Soc 20:145–147 [DOI] [PubMed] [Google Scholar]
- 31.Harris JL, Choi IY, Brooks WM (2015) Probing astrocyte metabolism in vivo: proton magnetic resonance spectroscopy in the injured and aging brain. Front Aging Neurosci 7:202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Harris JL, Yeh HW, Swerdlow RH, Choi IY, Lee P, Brooks WM (2014) High-field proton magnetic resonance spectroscopy reveals metabolic effects of normal brain aging. Neurobiol Aging 35:1686–1694 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Haug H, Eggers R (1991) Morphometry of the human cortex cerebri and corpus striatum during aging. Neurobiol Aging 12:336–338; discussion 352–335 [DOI] [PubMed] [Google Scholar]
- 34.Hayakawa N, Kato H, Araki T (2007) Age-related changes of astorocytes, oligodendrocytes and microglia in the mouse hippocampal CA1 sector. Mech Ageing Dev 128:311–316 [DOI] [PubMed] [Google Scholar]
- 35.Hedden T, Gabrieli JD (2004) Insights into the ageing mind: a view from cognitive neuroscience. Nat Rev Neurosci 5:87–96 [DOI] [PubMed] [Google Scholar]
- 36.Jessen NA, Munk AS, Lundgaard I, Nedergaard M (2015) The glymphatic system: a beginner’s guide. Neurochem Res 40:2583–2599 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Jiang T, Cadenas E (2014) Astrocytic metabolic and inflammatory changes as a function of age. Aging Cell [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Juurlink BH, Thorburne SK, Hertz L (1998) Peroxide-scavenging deficit underlies oligodendrocyte susceptibility to oxidative stress. Glia 22:371–378 [DOI] [PubMed] [Google Scholar]
- 39.Kanungo MS (1975) A model for ageing. J Theoret Biol 53:253–261 [DOI] [PubMed] [Google Scholar]
- 40.Khachaturian ZS (1987) Hypothesis on the regulation of cytosol calcium concentration and the aging brain. Neurobiol Aging 8:345–346 [DOI] [PubMed] [Google Scholar]
- 41.Kirischuk S, Parpura V, Verkhratsky A (2012) Sodium dynamics: another key to astroglial excitability? Trends Neurosci 35:497–506 [DOI] [PubMed] [Google Scholar]
- 42.Kress BT, Iliff JJ, Xia M, Wang M, Wei H, Zeppenfeld D, Xie L, Kang H, Xu Q, Liew J, Plog BA, Ding F, Deane R, Nedergaard M (2014) Impairment of paravascular clearance pathways in the aging brain. Ann Neurol [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Krimer LS, Hyde TM, Herman MM, Saunders RC (1997) The entorhinal cortex: an examination of cyto- and myeloarchitectonic organization in humans. Cereb Cortex 7:722–731 [DOI] [PubMed] [Google Scholar]
- 44.Lalo U, Palygin O, North RA, Verkhratsky A, Pankratov Y (2011) Age-dependent remodelling of ionotropic signalling in cortical astroglia. Aging Cell 10:392–402 [DOI] [PubMed] [Google Scholar]
- 45.Landfield PW (1987) ‘Increased calcium-current’ hypothesis of brain aging. Neurobiol Aging 8:346–347 [DOI] [PubMed] [Google Scholar]
- 46.Lee DC, Ruiz CR, Lebson L, Selenica ML, Rizer J, Hunt JB Jr, Rojiani R, Reid P, Kammath S, Nash K, Dickey CA, Gordon M, Morgan D (2013) Aging enhances classical activation but mitigates alternative activation in the central nervous system. Neurobiol Aging 34:1610–1620 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Logan JM, Sanders AL, Snyder AZ, Morris JC, Buckner RL (2002) Under-recruitment and nonselective recruitment: dissociable neural mechanisms associated with aging. Neuron 33:827–840 [DOI] [PubMed] [Google Scholar]
- 48.Lopez-Otin C, Galluzzi L, Freije JMP, Madeo F, Kroemer G (2016) Metabolic control of longevity. Cell 166:802–821 [DOI] [PubMed] [Google Scholar]
- 49.Lynch AM, Murphy KJ, Deighan BF, O’Reilly JA, Gun’ko YK, Cowley TR, Gonzalez-Reyes RE, Lynch MA (2010) The impact of glial activation in the aging brain. Aging Dis 1:262–278 [PMC free article] [PubMed] [Google Scholar]
- 50.Maher P (2005) The effects of stress and aging on glutathione metabolism. Ageing Res Rev 4:288–314 [DOI] [PubMed] [Google Scholar]
- 51.Matute C, Alberdi E, Domercq M, Sanchez-Gomez MV, Perez-Samartin A, Rodriguez-Antiguedad A, Perez-Cerda F (2007) Excitotoxic damage to white matter. J Anat 210:693–702 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Medvedev ZA (1990) An attempt at a rational classification of theories of ageing. Biol Rev Camb Philos Soc 65:375–398 [DOI] [PubMed] [Google Scholar]
- 53.Micu I, Jiang Q, Coderre E, Ridsdale A, Zhang L, Woulfe J, Yin X, Trapp BD, McRory JE, Rehak R, Zamponi GW, Wang W, Stys PK (2006) NMDA receptors mediate calcium accumulation in myelin during chemical ischaemia. Nature 439:988–992 [DOI] [PubMed] [Google Scholar]
- 54.Ming GL, Song H (2011) Adult neurogenesis in the mammalian brain: significant answers and significant questions. Neuron 70:687–702 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Nichols NR, Day JR, Laping NJ, Johnson SA, Finch CE (1993) GFAP mRNA increases with age in rat and human brain. Neurobiol Aging 14:421–429 [DOI] [PubMed] [Google Scholar]
- 56.Nicholson DA, Yoshida R, Berry RW, Gallagher M, Geinisman Y (2004) Reduction in size of perforated postsynaptic densities in hippocampal axospinous synapses and age-related spatial learning impairments. J Neurosci 24:7648–7653 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ogura K, Ogawa M, Yoshida M (1994) Effects of ageing on microglia in the normal rat brain: immunohistochemical observations. NeuroReport 5:1224–1226 [DOI] [PubMed] [Google Scholar]
- 58.Orre M, Kamphuis W, Osborn LM, Jansen AH, Kooijman L, Bossers K, Hol EM (2014) Isolation of glia from Alzheimer’s mice reveals inflammation and dysfunction. Neurobiol Aging [DOI] [PubMed] [Google Scholar]
- 59.Pelvig DP, Pakkenberg H, Stark AK, Pakkenberg B (2008) Neocortical glial cell numbers in human brains. Neurobiol Aging 29:1754–1762 [DOI] [PubMed] [Google Scholar]
- 60.Perry VH, Matyszak MK, Fearn S (1993) Altered antigen expression of microglia in the aged rodent CNS. Glia 7:60–67 [DOI] [PubMed] [Google Scholar]
- 61.Peters A, Josephson K, Vincent SL (1991) Effects of aging on the neuroglial cells and pericytes within area 17 of the rhesus monkey cerebral cortex. Anat Rec 229:384–398 [DOI] [PubMed] [Google Scholar]
- 62.Peters A, Sethares C (2004) Oligodendrocytes, their progenitors and other neuroglial cells in the aging primate cerebral cortex. Cereb Cortex 14:995–1007 [DOI] [PubMed] [Google Scholar]
- 63.Peters O, Schipke CG, Philipps A, Haas B, Pannasch U, Wang LP, Benedetti B, Kingston AE, Kettenmann H (2009) Astrocyte function is modified by Alzheimer’s disease-like pathology in aged mice. J Alzheimers Dis 18:177–189 [DOI] [PubMed] [Google Scholar]
- 64.Potokar M, Stenovec M, Jorgacevski J, Holen T, Kreft M, Ottersen OP, Zorec R (2013) Regulation of AQP4 surface expression via vesicle mobility in astrocytes. Glia 61:917–928 [DOI] [PubMed] [Google Scholar]
- 65.Provenzano FA, Muraskin J, Tosto G, Narkhede A, Wasserman BT, Griffith EY, Guzman VA, Meier IB, Zimmerman ME, Brickman AM, Alzheimer’s Disease Neuroimaging I (2013) White matter hyperintensities and cerebral amyloidosis: necessary and sufficient for clinical expression of Alzheimer disease? JAMA Neurol 70, 455–461 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Raz N, Lindenberger U, Rodrigue KM, Kennedy KM, Head D, Williamson A, Dahle C, Gerstorf D, Acker JD (2005) Regional brain changes in aging healthy adults: general trends, individual differences and modifiers. Cereb Cortex 15:1676–1689 [DOI] [PubMed] [Google Scholar]
- 67.Rodriguez-Callejas JD, Fuchs E, Perez-Cruz C (2016) Evidence of Tau hyperphosphorylation and dystrophic microglia in the common marmoset. Front Aging Neurosci 8:315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Rodriguez JJ, Terzieva S, Olabarria M, Lanza RG, Verkhratsky A (2013) Enriched environment and physical activity reverse astrogliodegeneration in the hippocampus of AD transgenic mice. Cell Death Dis 4:e678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Rodriguez JJ, Yeh CY, Terzieva S, Olabarria M, Kulijewicz-Nawrot M, Verkhratsky A (2014) Complex and region-specific changes in astroglial markers in the aging brain. Neurobiol Aging 35:15–23 [DOI] [PubMed] [Google Scholar]
- 70.Rose CF, Verkhratsky A, Parpura V (2013) Astrocyte glutamine synthetase: pivotal in health and disease. Biochem Soc Trans 41:1518–1524 [DOI] [PubMed] [Google Scholar]
- 71.Rose CR, Verkhratsky A (2016) Principles of sodium homeostasis and sodium signalling in astroglia. Glia [DOI] [PubMed] [Google Scholar]
- 72.Ruzicka V (1924) Beitrage zum Stadium der Protoplasmahysteretischen Vorgange (Zur Kausalitat der Alterns). Archiv fur mikroskopische Anatomie und Entwicklungsmechanik 101:459–482 [Google Scholar]
- 73.Ruzicka V (1926) Altern und Verjungung won Standpunkt der allegemeinen Biologie. Praha, Praha [Google Scholar]
- 74.Salter MG, Fern R (2005) NMDA receptors are expressed in developing oligodendrocyte processes and mediate injury. Nature 438:1167–1171 [DOI] [PubMed] [Google Scholar]
- 75.Sampedro-Piquero P, De Bartolo P, Petrosini L, Zancada-Menendez C, Arias JL, Begega A (2014) Astrocytic plasticity as a possible mediator of the cognitive improvements after environmental enrichment in aged rats. Neurobiol Learn Mem 114:16–25 [DOI] [PubMed] [Google Scholar]
- 76.Schoenemann PT, Sheehan MJ, Glotzer LD (2005) Prefrontal white matter volume is disproportionately larger in humans than in other primates. Nat Neurosci 8:242–252 [DOI] [PubMed] [Google Scholar]
- 77.Sharaf A, Krieglstein K, Spittau B (2013) Distribution of microglia in the postnatal murine nigrostriatal system. Cell Tissue Res 351:373–382 [DOI] [PubMed] [Google Scholar]
- 78.Sheffield LG, Berman NE (1998) Microglial expression of MHC class II increases in normal aging of nonhuman primates. Neurobiol Aging 19:47–55 [DOI] [PubMed] [Google Scholar]
- 79.Sierra A, Gottfried-Blackmore AC, McEwen BS, Bulloch K (2007) Microglia derived from aging mice exhibit an altered inflammatory profile. Glia 55:412–424 [DOI] [PubMed] [Google Scholar]
- 80.Smith TD, Adams MM, Gallagher M, Morrison JH, Rapp PR (2000) Circuit-specific alterations in hippocampal synaptophysin immunoreactivity predict spatial learning impairment in aged rats. J Neurosci 20:6587–6593 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Soreq L, Consortium UKBE, North American Brain Expression C, Rose J, Soreq E, Hardy J, Trabzuni D, Cookson MR, Smith C, Ryten M, Patani R, Ule J (2017) Major shifts in glial regional identity are a transcriptional hallmark of human brain aging. Cell Rep 18:557–570 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Stern Y (2009) Cognitive reserve. Neuropsychologia 47:2015–2028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Streit WJ, Braak H, Xue QS, Bechmann I (2009) Dystrophic (senescent) rather than activated microglial cells are associated with tau pathology and likely precede neurodegeneration in Alzheimer’s disease. Acta Neuropathol 118:475–485 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Streit WJ, Sammons NW, Kuhns AJ, Sparks DL (2004) Dystrophic microglia in the aging human brain. Glia 45:208–212 [DOI] [PubMed] [Google Scholar]
- 85.Streit WJ, Xue QS, Tischer J, Bechmann I (2014) Microglial pathology. Acta Neuropathol Commun 2:142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Taylor RC (2016) Aging and the UPR(ER). Brain Res 1648:588–593 [DOI] [PubMed] [Google Scholar]
- 87.Thorburne SK, Juurlink BH (1996) Low glutathione and high iron govern the susceptibility of oligodendroglial precursors to oxidative stress. J Neurochem 67:1014–1022 [DOI] [PubMed] [Google Scholar]
- 88.Tischer J, Krueger M, Mueller W, Staszewski O, Prinz M, Streit WJ, Bechmann I (2016) Inhomogeneous distribution of Iba-1 characterizes microglial pathology in Alzheimer’s disease. Glia 64:1562–1572 [DOI] [PubMed] [Google Scholar]
- 89.Toescu EC, Verkhratsky A (2007) The importance of being subtle: small changes in calcium homeostasis control cognitive decline in normal aging. Aging Cell 6:267–273 [DOI] [PubMed] [Google Scholar]
- 90.Tse KH, Herrup K (2017) DNA damage in the oligodendrocyte lineage and its role in brain aging. Mech Ageing Dev 161:37–50 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.VanGuilder HD, Bixler GV, Brucklacher RM, Farley JA, Yan H, Warrington JP, Sonntag WE, Freeman WM (2011) Concurrent hippocampal induction of MHC II pathway components and glial activation with advanced aging is not correlated with cognitive impairment. J Neuroinflammation 8:138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Verkhratsky A, Kirchhoff F (2007) NMDA Receptors in glia. Neuroscientist 13:28–37 [DOI] [PubMed] [Google Scholar]
- 93.Verkhratsky A, Marutle A, Rodriguez-Arellano JJ, Nordberg A (2015) Glial asthenia and functional paralysis: a new perspective on neurodegeneration and Alzheimer’s disease. Neuroscientist 21:552–568 [DOI] [PubMed] [Google Scholar]
- 94.Verkhratsky A, Matteoli M, Parpura V, Mothet JP, Zorec R (2016) Astrocytes as secretory cells of the central nervous system: idiosyncrasies of vesicular secretion. EMBO J 35:239–257 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Verkhratsky A, Nedergaard M (2018) Physiology of Astroglia. Physiol Rev 98:239–389 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Verkhratsky A, Rodriguez JJ, Parpura V (2012) Calcium signalling in astroglia. Mol Cell Endocrinol 353:45–56 [DOI] [PubMed] [Google Scholar]
- 97.Verkhratsky A, Untiet V, Rose CR (2019) Ionic signalling in astroglia beyond calcium. J Physiol [DOI] [PubMed] [Google Scholar]
- 98.Walhovd KB, Johansen-Berg H, Karadottir RT (2014) Unraveling the secrets of white matter—bridging the gap between cellular, animal and human imaging studies. Neuroscience 276C:2–13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Weismann A (1881) Ueber die Dauer des Lebens. Vortrag, in der 2. allgemeinen Sitzung d. 54. Versammlung Deutscher Naturforscher u. Aerzte in Salzburg, am 21, Sept 1881
- 100.West MJ (1993) Regionally specific loss of neurons in the aging human hippocampus. Neurobiol Aging 14:287–293 [DOI] [PubMed] [Google Scholar]
- 101.Young VG, Halliday GM, Kril JJ (2008) Neuropathologic correlates of white matter hyperintensities. Neurology 71:804–811 [DOI] [PubMed] [Google Scholar]
- 102.Zhu X, Hill RA, Dietrich D, Komitova M, Suzuki R, Nishiyama A (2011) Age-dependent fate and lineage restriction of single NG2 cells. Development 138:745–753 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Zorec R, Parpura V, Verkhratsky A (2018) Preventing neurodegeneration by adrenergic astroglial excitation. FEBS J [DOI] [PubMed] [Google Scholar]