

Abstract
Insertion sequences (ISs), arguably the smallest and most numerous autonomous transposable elements (TEs), are important players in shaping their host genomes. This review focuses on prokaryotic ISs. We discuss IS distribution and impact on genome evolution. We also examine their effects on gene expression, especially their role in activating neighbouring genes, a phenomenon of particular importance in the recent upsurge of bacterial antibiotic resistance. We explain how ISs are identified and classified into families by a combination of characteristics including their transposases (Tpases), their overall genetic organisation and the accessory genes which some ISs carry. We then describe the organisation of autonomous and nonautonomous IS‐related elements. This is used to illustrate the growing recognition that the boundaries between different types of mobile element are becoming increasingly difficult to define as more are being identified. We review the known Tpase types, their different catalytic activities used in cleaving and rejoining DNA strands during transposition, their organisation into functional domains and the role of this in regulation. Finally, we consider examples of prokaryotic IS domestication. In a more speculative section, we discuss the necessity of constructing more quantitative dynamic models to fully appreciate the continuing impact of TEs on prokaryotic populations.
Keywords: insertion sequence, genome, mechanism, diversity, evolution
We describe the diversity of prokaryotic insertion sequences, their effect on their host genomes, their relationship to other transposable elements and their transposition properties including the organisation of their transposases into functional domains.
Introduction
The idea that many prokaryotic genomes are mosaic, composed of a ‘central genome backbone’ of essential and house‐keeping genes (the core genome) interspersed with DNA segments constituting the ‘mobilome’ (a variety of accessory genes that form part of the pan genome), is now common currency (Medini et al., 2005; Tettelin et al., 2008). The mobilome embraces several types of genetic unit which, as their collective name indicates, can move from place to place in a particular genome or from cell to cell. These mobile genetic elements (MGEs) can be divided into two major groups: those, such as plasmids and bacteriophages, that are transmissible from cell to cell (the intercellular MGEs), and those that cannot themselves undergo transfer but which are transferred following integration into members of the first group (the intracellular MGEs). Intracellular MGE or transposable elements (TEs) include transposons (Tn) and insertion sequences (ISs) but can embrace integrons (In) and introns (Craig et al., 2002). Originally, Tn were distinguished from ISs because they carry passenger (also called cargo) genes not involved in catalysing or regulating TE movement. Most eukaryotic DNA transposons have relatives among the prokaryotic ISs (see (Hickman et al., 2010a, b)) and it is not surprising that a variety of these elements carrying passenger genes are now also being identified (Bao et al., 2009; Bao & Jurka, 2013). Prokaryotes harbour a host of such elements as well as several types of structure possessing characteristics of both groups (e.g. integrative conjugative elements, ICEs, originally called conjugative transposons, and other types of genomic island) (Burrus & Waldor, 2004) (Dobrindt et al., 2004; Guerillot et al., 2013).
TEs insert into many different sites within a genome using mechanisms, which do not involve large regions of DNA homology between the TE and its target. In contrast to MGEs in eukaryotes, which include a large proportion whose movement uses RNA intermediates, the vast majority of known prokaryotic TEs transpose using DNA intermediates.
This review will be limited to the simplest TEs: the ISs. ISs are classified into families (Fig. 1, Table 1) using a variety of characteristics (Mahillon & Chandler, 1998) (see ISfinder: www‐is.biotoul.fr, below). This includes (1) the length and sequence of the short imperfect terminal inverted repeat sequences (IRs) carried by many ISs at their ends (TIRs or ITRs in eukaryotes); (2) the length and sequence of the short flanking direct target DNA repeats (DRs) (TSD, target site duplication, in eukaryotes) often generated on insertion; (3) the organisation of their open reading frames; or (4) the target sequences into which they insert. However, the principal factor in IS classification is the similarity, at the primary sequence level, of the enzymes which catalyse their movement, their transposases (Tpases) (see ‘Major IS groups are defined by transposase type’ below).
Figure 1.
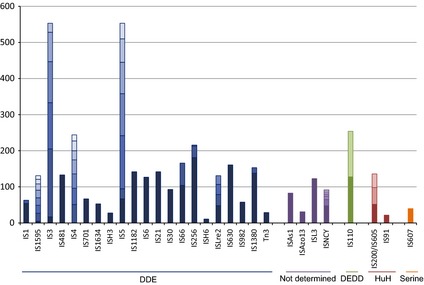
Distribution of IS families in the ISfinder database. The histogram shows the number of IS of a given family, as defined in the text, in the ISfinder database (June 2013). The horizontal boxes indicate the number and relative size of different subgroups (see Table 1 for the subgroups names) within the family. They are grouped by colour to indicate the type of Tpase used: DDE, blue; undetermined, purple; DEDD, green; HUH, red; Serine, orange.
Table 1.
General characteristics of IS families
| Families | Subgroups | Typical size‐range | DR (bp) | Ends | IRs | Nb ORF | Frameshift | Catalytic residus | Comments | Mechanism |
|---|---|---|---|---|---|---|---|---|---|---|
| IS1 | – | 740–1180 | 8–9 | GGnnnTG | Y | 2 | ORFAB | DDE | Copy‐and‐paste co‐integrate | |
| Single ORF | 800–1200 | 0–9 | 1 | |||||||
| ISMhu11 | 900–4600 | 0–10 | Y | 2 | ORFAB | |||||
| IS1595 | ISPna2 | 1000–1150 | 8 | GGCnnTG | Y | 1 | DDNK | Copy‐and‐paste ? | ||
| ISPna2 + pass | 1500–2600 | 8 | ||||||||
| ISH4 | 1000 | 8 | CGCTCTT | 1 | DDNK | |||||
| IS1016 | 700–745 | 7–9 | GGGgctg | 1 | DDEK | |||||
| IS1595 | 900–1100 | 8 | CcTGATT | 1 | DDNK + ER4R7 | |||||
| ISSod11 | 1000–1100 | 8 | nnnGcnTATC | 1 | DDHK + ER4R7 | |||||
| ISNwi1 | 1080–1200 | 8 | ggnnatTAT | 1 | DDEK + ER4 | |||||
| ISNwi1 + pass | 1750–4750 | 8 | 1 | |||||||
| ISNha5 | 3450–7900 | 8 | CGGnnTT | 1 | DDER/K | |||||
| IS3 | IS150 | 1200–1600 | 3–4 | TG | Y | 2 | ORFAB | DDE | ORFB coordinates defined between two STOP codons since there are no inherent initiation signals and the orf is not translated alone. | Copy‐paste |
| IS407 | 1100–1400 | 4 | TG | |||||||
| IS51 | 1000–1400 | 3–4 | TG | |||||||
| IS3 | 1150–1750 | 3–4 | TGa/g | |||||||
| IS2 | 1300–1400 | 5 | TG | |||||||
| IS481 | – | 950–1300 | 4–15 | TGT | Y | 1 | Copy‐paste ? | |||
| IS4 | IS10 | 1200–1350 | 9 | CT | Y | 1 | DDE | Hairpin intermediate | Cut‐and paste | |
| IS50 | 1350–1550 | 8–9 | C | Hairpin intermediate | ||||||
| ISPepr1 | 1500–1600 | 7–8 | ‐T‐AA | |||||||
| IS4 | 1400–1600 | 10–13 | ‐AAT | |||||||
| IS4Sa | 1150–1750 | 8–10 | CA | |||||||
| ISH8 | 1400–1800 | 10 | CAT | |||||||
| IS231 | 1450–5400 | 10–12 | CAT | 1 or +* | *Passenger genes | |||||
| IS701 | – | 1400–1550 | 4 | Y | 1 | DDE | ||||
| ISH3 | – | 1225–1500 | 4–5 | C‐GT | Y | 1 | DDE | |||
| IS1634 | – | 1500–2000 | 5–6 | C | Y | 1 | DDE | |||
| IS5 | IS903 | 950–1150 | 9 | GG | Y | 1 | DDE | |||
| ISL2 | 850–1200 | 2–3 | 1 | |||||||
| ISH1 | 900–1150 | 8 | ‐GC | 1 | ||||||
| IS5 | 1000–1500 | 4 | Ga/g | 1 | ||||||
| IS1031 | 850–1050 | 3 | GAa/g | 1 | ||||||
| IS427 | 800–1000 | 2–4 | Ga/g | 2 | ORFAB | |||||
| IS1182 | 1330–1950 | 0–60 | Y | 1 | DDE | |||||
| IS6 | – | 700–900 | 8 | GG | Y | 1 | DDE | Co‐integrate | ||
| IS21 | – | 1750–2600 | 4–8 | TG | Y | 2* | DDE | *istB : helper of transposition | ||
| IS30 | – | 1000–1700 | 2–3 | Y | 1 | DDE | Copy and paste | |||
| IS66 | – | 2000–3000 | 8–9 | GTAA | Y | 3* | DDE* | *TnpC has the DDE domain | ||
| ISBst12 | 1350–1900 | 8–9 | GTAA | Y | 1 | DDE | ||||
| IS91 | – | 1500–2000 | 0 | N | 1 | HUH/Y2 | Target site GAAC () CAAG | Rolling circle | ||
| IS110 | – | 1200–1550 | 0 | N | 1 | DEDD | ||||
| IS1111 | Y* | *IRs not at the termini of the IS | ||||||||
| IS200/IS605 | IS200 | 600–750 | 0 | N | 1* | HUH/Y1 | *TnpA | Peel and paste | ||
| – | 1300–2000 | 2* | HUH/Y1** | *TnpA + TnpB; **Y1 on TnpA | ||||||
| IS1341 | 1200–1500 | 1* | *TnpB | |||||||
| IS607 | – | 1700–2500 | 0 | N | 2* | Serine** | *TnpA + TnpB; **Y1 on TnpA | |||
| IS256 | – | 1200–1500 | 8–9 | Ga/g | Y | 1 | DDE | Copy‐paste | ||
| IS630 | – | 1000–1400 | 2* | Y | 1 or 2 | ORFAB | DDE | *Target site : often NTAN with duplication of the TA | Cut and paste | |
| IS982 | – | 1000 | 3–9 | AC | Y | 1 | DDE | |||
| IS1380 | – | 1550–2000 | 4–5 | CC | Y | 1 | DDE | |||
| ISAs1 | – | 1200–1500 | 8–10 | CAGGG | Y | 1 | ||||
| ISL3 | – | 1300–2300 | 8 | GG | Y | 1 | ||||
| Tn3 | – | > 3000 | 0 | GGGG | Y | > 1 | DDE | Co‐integrate | ||
| ISAzo13 | – | 1250–2200 | 0–4 | Ga/g | Y | 1 |
IS families as defined in ISfinder are listed in related groups. Column 1: IS families; column 2: IS subgroups; column 3: typical length in base pairs; column 4: length of direct target repeats generated on insertion in base pairs; column 5: typical sequence of the IS ends; column 6: presence of imperfect terminal inverted repeats (Y = yes); column 7: presence of a frameshift; column 8: catalytic residus (DDE = OH as nucleophile; Y1 = single nucleophilic tyrosine; Y2 = two nucleophilic tyrosines; Ser = nucleophilic serine); column 9: mechanism if known; column 10: general comments; ? clearly means we do not know.
This article is being made freely available through PubMed Central as part of the COVID-19 public health emergency response. It can be used for unrestricted research re-use and analysis in any form or by any means with acknowledgement of the original source, for the duration of the public health emergency.
As ISs have been reviewed a number of times over the past years [e.g. (Mahillon & Chandler, 1998; Chandler & Mahillon, 2002; Curcio & Derbyshire, 2003; Hickman et al., 2010a, b; Montano & Rice, 2011; Dyda et al., 2012)], we have not included a detailed in‐depth description of each different IS family. Instead, we first discuss IS distribution and impact on genome evolution and expression and explain how they are identified and classified into families by their Tpases and accessory genes. We then describe various mobile elements related to ISs to illustrate the growing recognition that the boundaries between different types of mobile element are becoming increasingly difficult to define as more are being identified. We also describe the different types of Tpase and their activities as well as their organisation into domains and its role in regulation. Finally, we consider the few known examples of prokaryotic IS domestication.
Distribution
ISs are widespread and can occur in very high numbers in prokaryotic genomes. A recent study concluded that proteins annotated as Tpases, or as proteins with related functions, are by far the most abundant functional class in both the prokaryotic and eukaryotic genomic and metagenomic public data bases (Aziz et al., 2010).
Since the last surveys [e.g. (Mahillon & Chandler, 1998; Chandler & Mahillon, 2002)], many new ISs have been identified largely as a result of the massive increase in available sequenced prokaryotic genomes. Careful analysis of a number of these has also revealed that some genomes contain significant levels of truncated and partial ISs devoid of Tpase genes. These genomic ‘scars’ represent traces of numerous ancestral transposition events. However, genome annotations are often based simply on the presence of Tpase genes and do not include the entire DNA sequence with the IS ends. Indeed, a significant number of solo IS‐related IRs have been identified in various genomes (www‐is.biotoul.fr; genome section). Small IS fragments are rarely taken into account even though they can provide important insights into the evolutionary history of the host genome. Not only can this seriously impair studies attempting to provide an overview of the evolutionary influence of TEs on bacterial and archeal genomes, but such fragments may encode truncated proteins and these could influence gene regulation [e.g. (Cordaux et al., 2006; Liu et al., 2007; Shaheen et al., 2010)]. In bacteria (Gueguen et al., 2006) (Stalder et al., 1990) (Salvatore et al., 2001) and eukaryotes (Vos et al., 1993) (Rio, 1991), truncated transposases have been shown to inhibit transposition. One example where annotation of IS fragments has provided important information is in the obligatory intracellular insect endosymbiont, Wolbachia, which also carries high numbers of full‐length ISs. The sequence divergence observed suggests that several waves of IS invasion and elimination have occurred over evolutionary time (Cerveau et al., 2011).
Impact of ISs on genome evolution
ISs have had an important impact on genome structure and function. Several of these effects are considered in the following sections. In this context, it is useful to understand the time scales involved in these processes because they are often confounded. Evolutionary time is used to compare species (106 years), historical time in comparisons within or between populations (102–104 years), variety time in selection experiments (1–102 years) and laboratory time for ongoing events such as experimental measurement of transposition frequencies or in biochemical analyses (10−3–1 years) (A. Schulman, pers. commun.). Thus, a ‘burst’ of transposition in evolutionary time is many orders of magnitude longer than a ‘burst’ of transposition in experimental biology.
IS expansion, elimination and genome streamlining
Perhaps, one of the most striking concerns IS expansion. ISs can undergo massive expansion and loss accompanied by gene inactivation and decay, genome rearrangement and genome reduction. Clearly, host lifestyle strongly influences these IS‐mediated effects on genome structure, presumably by determining the level of genetic isolation of the microbial population. Factors affecting this include: whether the bacteria are ectosymbionts, primary endosymbionts having long evolutionary histories with their hosts, or secondary endosymbionts with more recent associations; whether they are transmitted in a strictly vertical manner or pass through a step of horizontal transfer via reinfection or passage through a second host vector (Bordenstein & Reznikoff, 2005; Moya et al., 2008).
IS expansion has been commonly observed in bacteria with recently adopted fastidious, host‐restricted lifestyles. Those which may have more ancient host‐restricted lifestyles (e.g. Wigglesworthia in the Tsetse fly; Buchnera aphidicola in the aphid; Blochmannia floridanus in the ant) tend to possess small streamlined genomes with few pseudogenes or MGEs (see (Bordenstein & Reznikoff, 2005; Moya et al., 2008); Supporting Information, Table S1).
One view is that IS expansion is an early step in this genome reduction process (Moran & Plague, 2004; Touchon & Rocha, 2007; Gil et al., 2008; Plague et al., 2008) (Fig. 2). This results from a decrease in strength and efficacy of purifying selection due to the shift from free to intracellular lifestyles (Moran & Plague, 2004). It is reinforced by a phenomenon known as Muller's ratchet, which leads to the irreversible accumulation of mutations in a confined intracellular environment (Moran, 1996; Andersson & Kurland, 1998; Silva et al., 2003). In the nutritionally rich environment of the host, many genes of free‐living bacteria are inessential. Enhanced genetic drift would allow fixation of slightly deleterious mutations in the population, facilitated by the occurrence of successive population bottlenecks. The more genetically isolated the bacterial population, the more acute would be the effect. Indeed, many examples of this can be found among intracellular endosymbionts. This initial stage of transition from free‐living to host‐dependence would therefore result in the accumulation of pseudogenes, which will eventually be eliminated by so‐called deletional bias (Mira et al., 2001). Clearly, the activities of MGEs, and of ISs in particular, make them important instruments in these processes. IS expansion would contribute to pseudogenisation by IS‐mediated intrachromosomal recombination and genome reduction (Andersson & Andersson, 1999; Lawrence et al., 2001; Mira et al., 2001) by their capacities to generate deletions (Mahillon & Chandler, 1998). Such deletions would also eventually lead to complete or partial elimination of the ISs themselves. These processes are shown schematically in Fig. 2.
Figure 2.
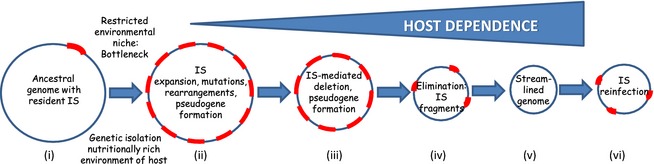
IS expansion, elimination and genome ‘streamlining’. The figure shows schematically from left to right events leading to the evolution of host‐dependence in bacteria. (i) The parental (ancestral) chromosome including a low number of resident IS (red arcs). Note that the entire genome might also include transmissible plasmids carrying their own IS load, which can in principle undergo transposition into the chromosome. (ii) IS expansion occurs as a result of isolation and the formation of population bottlenecks within a host organism. This is accompanied by mutation promoted by insertion of new IS copies and by their related transposition activities of deletion and rearrangement. These genome rearrangements can also occur by homologous recombination between identical IS copies. (iii) With time IS will have a tendency to undergo deletion with adjacent DNA sequences in the absence of direct selection. This leads to a reduction in genome size. (iv) Eventually, extensive deletion will lead to the generation of nonautonomous IS fragments and their elimination. (vi) This gives rise to streamlined, IS ‘free’ genomes which may become ‘reinfected’ by IS on rare contact with other, IS‐carrying strains or infection by IS‐carrying bacteriophage (v).
There are many striking examples of IS expansions in bacterial genomes (Table S1). The first to be identified was Shigella from the pregenomics era (Nyman et al., 1981; Ohtsubo et al., 1981). But IS expansion identified from sequenced genomes has been implicated in generating the present day Bordetella pertussis and B. parapertusis, Yersinia pestis, Enterococcus faecium, Mycobacterium ulcerans and many others. In at least some of these cases, it has been argued that large‐scale genome rearrangements and deletions associated with IS expansion have improved the ability of the bacterium to combat host defences for example by changing surface antigens and regulatory circuitry (Parkhill & Thomson, 2003). This has been particularly well documented in the Bordetellae (Parkhill et al., 2003; Preston et al., 2004).
The phenomenon is also common among endosymbionts such as Wolbachia sp. These are considered ancient endosymbionts, which might be expected to possess more streamlined genomes. However, evidence has been presented that they have been subjected to several waves of invasion and elimination of ISs (Cerveau et al., 2011). This may be related to the fact that they are not strictly transmitted vertically but may also undergo relatively low levels of horizontal transmission and coinfection. Other symbionts or host‐restricted bacteria also contain high IS loads. These include organisms such as Orientia tsutsugamushi, various Rickettsia, Sodalis glossinidius, Amoebophilus asiaticus 5a2, the γ1 symbiont of the marine oligochaete Olavius algarvensis, the Bacteroidete Cardinium hertigii, a symbiont of the parasitic wasp Encarsia pergandiella, and the primary symbionts of grain weevils. These obligate intracellular bacteria may carry intercellular MGEs such as phage (Hsia et al., 2000; Read et al., 2000) and conjugative elements (Blanc et al., 2007) capable of acting as IS vectors and motors of horizontal gene transfer. Similar arguments might be used for other niche‐restricted prokaryotes to explain increased IS loads found in some extremophiles (e.g. Sulfolobus solfataricus and certain cyanobacteria) (Brugger et al., 2004; Filee et al., 2007) (Papke et al., 2003; Allewalt et al., 2006).
Although IS expansion is generally assumed to occur stochastically over periods of evolutionary time, it has recently been observed that the Olavius algarvensis symbionts express significant levels of transposase (Kleiner et al., 2013). This raises the possibility that transposase expression is deregulated in this symbiont system. However, another symbiont, Amoebophilus asiaticus, with a high IS load, shows no evidence of recent transposition activity in spite of extensive IS transcription (Schmitz‐Esser et al., 2011). In view of the time scales involved, only a very small but sustained increase in transposition activity might be needed to give rise to the high loads observed. Further exploration of the relationship between IS gene expression and transposition activity is clearly essential to understanding the dynamics of ISs in these and other systems.
Of course, different ISs are involved in different expansions and it is therefore important to understand IS diversity and properties. This is clearly evident in studies concerning the behaviour of IS on storage of bacterial strains where certain ISs appear more active than others (Naas et al., 1994, 1995).Their detailed effects on the host genome will depend on their particular transposition mechanisms. For example, IS target specificity will have profound effects on the way the host genome is shaped.
Target choice
The choice of a target DNA can have an important influence on the impact of TE on their host genomes. Initially, it was often assumed that TE show no or perhaps only low sequence specificity in their choice of a target. While this remains approximately true, the enormous increase in available sequences has provided more statistically robust data which have revealed that several TE use rather subtle mechanisms in choosing a target. Inspection of the public databases suggests that IS density is significantly higher in bacterial plasmids than in their host chromosomes and it seems likely that plasmids are major vectors in IS transmission (as well as in transmission of accessory traits such as resistance to antibacterials). Indeed, plasmids, in particular those that use a rolling circle mechanism either for replication or in conjugation, appear to be preferential targets for certain TE. For example, transposon Tn7 has two modes of transposition: in one, a specific sequence within the highly conserved glmS is recognised and insertion occurs next to this essential gene (Craig, 2002); in the second, insertion occurs into replication forks directed by interactions with the β‐clamp (Parks et al., 2009). This results in a strong orientation bias of Tn7 insertions, consistent with insertion into the lagging strand of the replication fork formed during conjugative transfer. Although studies with IS are less advanced, a similar orientation bias was observed with IS903 (Hu & Derbyshire, 1998), suggesting that it too may use the β‐clamp in directing insertions. It seems probable that many other IS use this type of protein–protein interaction.
A second example of specialised target choice was observed in members of the IS200/IS605 family (see Transposases and mechanism: IS200/IS605‐family transposases below). These transpose using a strand‐specific single‐strand intermediate and insertion occurs 3′ to a tetra‐ or penta‐nucleotide on the lagging strand (Ton‐Hoang et al., 2010). Clear vestiges of this specificity can still be detected in a large number of bacterial genomes where the orientation of insertion is strongly correlated with the direction of replication. There are clearly incidences of insertion in the ‘wrong’ orientation, but many of these may be explained by postinsertion genome rearrangements involving inversions. This would place the ‘active’ strand of the IS on the lagging rather than on the leading strand. Interestingly, those IS that are not oriented in the ‘correct’ orientation with respect to replication are almost certainly inactive and unable to transpose further (Ton‐Hoang et al., 2010). Replication fork targeting was subsequently observed in eukaryotes (Spradling et al., 2011).
Other examples of sequence‐specific target choice have been described. IS1, for example, shows a preference for regions rich in AT, whereas the transposon TnGBS and members of the related ISLre2 family show a preference for insertion 15–17 bp upstream of σA promoters (Brochet et al., 2009; Guerillot et al., 2013). Targeting of upstream regions of transcription units has also been extensively documented for certain eukaryotic transposons [e.g.(Qi et al., 2012)].
There are also examples of IS (e.g. some members of the IS110, IS3 and IS4 families) which insert into potential secondary structures such as repeated extragenic palindromes (REP) (see The REP system below) (Clement et al., 1999); (Wilde et al., 2003), (Tobes & Pareja, 2006), integrons (Tetu & Holmes, 2008; Post & Hall, 2009) or even the ends of other TE) (Partridge & Hall, 2003) (Hallet et al., 1991).
These examples represent only a small part of the literature concerning factors influencing target choice but serve to illustrate the impact this can have on genomes. Further notable aspects of the effects of target choice are considered in the next section.
Impact of ISs on genome expression
While massive IS‐mediated genomic changes leading to streamlined genomes with increased pathogenicity and virulence are important and spectacular, they do not reflect the full impact of ISs. ISs can incorporate additional genes and subsequently act as vectors for these genes (see section ISs and relatives with passenger genes below). Areas of topical relevance in this respect are the transmission of antibacterial resistance and virulence.
ISs also play more subtle roles. They can insert upstream of a gene and activate its expression, a phenomenon known for some time (Glansdorff et al., 1981) and suspected for even longer (Reif & Saedler, 1974). This has recently received much attention due to the increase in resistance to various antibacterials (see (Aubert et al., 2006; Soki et al., 2013), which has become a worrying public health threat (Kieny, 2012; McKenna, 2013; Mole, 2013).
Activation of neighbouring gene expression can occur in two principal ways: either via promoters contained entirely within the IS driving transcripts that escape into neighbouring DNA (Glansdorff et al., 1981), or by the formation of hybrid promoters following insertion. Many ISs contain outward oriented ‐35 promoter components in their ends and insertion at the correct distance from a suitable ‐10 box can generate a strong promoter [e.g. (Prentki et al., 1986)]. This property has been noted for a very large number of ISs in a variety of bacterial strains. A preliminary survey of the literature, presented in Table S2, shows that the phenomenon is associated with over 30 different ISs and has occurred in at least 17 bacterial species (Depardieu et al., 2007). Indeed, specific vector plasmids have been designed to identify activating insertions [e.g. (Szeverenyi et al., 1996)].
IS activity can affect efflux mechanisms resulting in increased resistance: IS1 or IS10 insertion can up‐regulate the AcrAB‐TolC pump in Salmonella enterica (Olliver et al., 2005); IS1 or IS2 insertion upstream of AcrEF (Jellen‐Ritter & Kern, 2001; Kobayashi et al., 2001) and IS186 insertional inactivation of the AcrAB repressor, AcrR, in Escherichia coli (Jellen‐Ritter & Kern, 2001), all lead to increased resistance to fluoroquinolones. Insertional inactivation of specific porins can also play a significant role (Wolter et al., 2004).
The IS families
Diversity and classification
IS classification is needed to cope with the high numbers and diversity of ISs. It also permits identification of the many IS fragments present in numerous genomes, contributes to understanding their effects on their host genomes and can provide insights into their regulation and transposition mechanism. This role has been assumed by the ISfinder database (http://www-is.biotoul.fr) (Siguier et al., 2006) following the closure of the Stanford repository (Lederberg, 1981).
IS identification
The families in ISfinder are defined using an initial manual blast analysis often followed by reiterative blast analyses with the primary transposase sequence of representative elements used as a query in a blastp (Altschul et al., 1990) search of microbial genomes. Potential full‐length Tpases are retained and that with the lowest score then used as a query in a second blastp search. This is continued until no new potential candidates are detected. The clustalw multiple alignment algorithm (Thompson et al., 1994) is then used and the results displayed using the Jalview alignment editor (Clamp et al., 2004) for assessment. The corresponding DNA together with 1000 base pairs upstream and downstream is then extracted and examined manually for the IRs or other typical features such as secondary structures and flanking DRs. This, together with comparison of the DNA extremities of various elements, allows identification of both ends of the collected elements. In cases where more than a single IS copy is identified, blastn can be used to define the IS ends. Where only a single copy is found, the ends can often be defined by identifying and comparing with empty sites.
In a second step, we use the Markov cluster algorithm (MCL) (http://micans.org/mcl/) (Van Dongen, 2000; Enright et al., 2002) to weigh the relationships between clusters of ISs and to validate prior ISfinder classification of ISs into families and subgroups (Siguier et al., 2009). This is explained in detail in Siguier et al. (2009) and is based on the parameters used in the MCL in addition to characteristics, such as the specificity of target site duplications, the detailed sequence of the ends and genetic organisation. It should be understood that the distinction between families and subgroups can evolve as the number of ISs in the database increases.
The ISfinder repository contains over 4000 entries grouped into c. 26 families some of which can be conveniently divided into subgroups (Fig. 1, Table 1). This classification evolves continuously with the accumulation of additional ISs.
Several semi‐automatic IS annotation pipelines are now available. The interested reader is directed to three of these: ISsaga (which is integrated into the ISfinder platform (Siguier et al., 2006; Varani et al., 2011), ISScan (Wagner et al., 2007) and Oasis (Robinson et al., 2012). At present, de novo prediction of ISs is not efficient and these pipelines all employ the ISfinder database to function. While all three pipelines permit identification of IS fragments as well as full‐length ISs, a certain level of manual assessment is essential.
Major IS groups defined by transposase type
The primary difference between ISs is the nature of their transposases based on the type of chemistry they catalyse. These include DDE, DEDD, HUH and Ser transposases and are described in more detail in the following sections. Figure 1 shows the different families associated with each transposase type as of June 2013. For each family, the histogram is colour‐coded to indicate the different subgroups. The majority of these are classical ISs and encode Tpases of the DDE superfamily (see ‘Transposases’ below).
It is important to note that this is certainly not an unbiased sample. ISs are generally identified by their similarity to those already in the database and rarely, as was the case in the pregenomic era, by their transposition activity (i.e. by analysis of mutations they produce by transposition). Moreover, IS inclusion in ISfinder has not involved a systematic global search of the public databases. We therefore emphasise that this distribution should not be taken as a true representation of the relative abundance of different IS families in prokaryotes.
ISs with DDE transposases
Classical ISs with DDE transposases (named for a conserved amino acid triad, Asp, Asp, Glu, the active site) are small (c. 0.7–2.5 kb long) genetically compact DNA segments with one or two open reading frames (a Tpase and possibly a protein involved in regulation). They end with imperfect IRs and generate short flanking DRs on insertion (Table 1). The DR length is specific for each IS type. There are clearly several dominant families among the DDE IS group. These include IS3, whose Tpases are perhaps the most closely related to the retroviral integrase catalytic core (IN) by the spacing of the DDE triad and by the appearance of additional conserved residues [e.g. (Haren et al., 1999)]. The IS3 family contains several well‐defined clades delimiting subgroups. With subsequent accumulation of additional members, it has been necessary to redefine several large families, such as IS4, into a number of individual families (De Palmenaer et al., 2004). A similar situation has also occurred for the IS5 family (Table 1; Fig 1) whose original members are at present distributed over several families and subgroups. Some of these will certainly develop into separate families (P. Siguier, E. Gourbeyre and M. Chandler, unpublished).
In addition to the DDE encoding IS, three fundamentally different IS types have also been identified.
ISs with DEDD transposases
DEDD transposases (for Asp, Glu, Asp, Asp) are related to the Holiday junction resolvase, RuvC, itself related to DDE transposases. At present, only a single IS family, IS110, is known to encode this type of enzyme. The organisation of family members is quite different from that of the DDE ISs: they do not contain the typical terminal IRs of the DDE ISs and do not generate flanking target DRs on insertion. This implies that their transposition occurs using a different mechanism to the DDE ISs.
ISs with HUH Y1 or Y2 transposases
Two of these families encode a tyrosine (Y) Tpase. Note that these Tpases are not related to the well‐characterised tyrosine site‐specific recombinases such as phage integrases. Neither carries terminal IRs nor do they generate DRs on insertion. One family includes IS91 (see below) (Zabala et al., 1982; Garcillan‐Barcia et al., 2002), and the other includes IS200/IS605 (Lam & Roth, 1983; Kersulyte et al., 2002). Members of these families transpose using an entirely different mechanism to ISs with DDE transposases (del Pilar Garcillan‐Barcia et al., 2001; Ton‐Hoang et al., 2005) (see below). These ISs carry subterminal sequences, which are able to forms hairpin secondary structures. This is particularly marked in the IS200/IS605 family elements.
These ISs are defined by the presence of a Tpase belonging to the HUH endonuclease superfamily (named for the conserved active site amino acid residues H = Histidine and U = large hydrophobic residue). There are two major HUH Tpase families: Y1 and Y2 enzymes (Chandler et al., 2013) (see Transposases and mechanism below) according to whether they carry one or two Y residues involved in catalysis. One (Y1) is associated with the IS200/IS605 family (Ton‐Hoang et al., 2005). The second (Y2) is associated with the IS91 insertion sequence family (Mendiola & de la Cruz, 1992), with a related and newly defined group, the ISCR (Toleman et al., 2006) (see IS91‐related ISCRs below) and with eukaryotic helitrons (Kapitonov & Jurka, 2007). Although these enzymes use the same Y‐mediated cleavage mechanism, they appear to carry out the transposition process in quite different ways (see Transposases and mechanism below).
While the IS91 family is fairly homogenous, the IS200/IS605 family is divided into three major subgroups, IS200, IS1341, and those that resemble IS605. This is based on the presence or absence of two reading frames tnpA and tnpB, which can occur individually or together in several configurations (Ton‐Hoang et al., 2005) (Fig. 3): tnpA encodes the Tpase and tnpB encodes a protein with a possible role in regulation (Pasternak et al., 2013) (see ‘IS with Accessory genes’ below).
Figure 3.
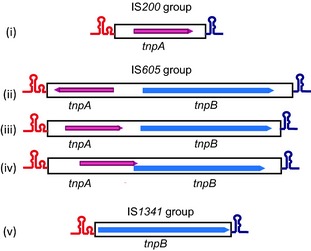
Organisation of the IS 200/IS 605 family. (i) IS 200 group with tnpA alone: examples include IS 200 (Salmonella typhimurium, Escherichia coli), IS 1541 (Yersinia pseudotuberculosis, Yersinia pestis), IS 1469 (Clostridium perifringens) and ISW 1 (Wolbachia sp.). (ii) IS 605/IS 606 type with tnpA and tnpB in a divergent orientation: IS 605, IS 606 (Helicobacter pylori); ISL jo5 (Lactobacillus johnsonii). (iii) IS 8301 type with nonoverlapping tnpA and tnpB orfs in the same direction: ISD ra2 (Deinococcus radiodurans), ISH 1‐8 (Halobacterium), ISE fa4 (Enterococcus faecium), IS 1253 (Dichelobacter nodosus). (iv) IS 608 with overlapping tnpA and tnpB: IS 608 (Helicobacter pylori). (v) IS 1341 (Thermophilic bacterium) group with tnpB alone. Left and right are represented as hairpin structures in red and blue, respectively. Orfs are indicated as boxes with arrowheads (showing the direction of translation). tnpA is shown in red and tnpB in blue.
ISs with serine transposases
The third family is represented by IS607, which carries a Tpase closely related to serine recombinases such as the resolvases of Tn3 family elements (see below). Little is known about their transposition mechanism. However, it appears likely, in view of the known activities of resolvases (Grindley, 2002), that IS607 transposition may involve a circular double‐strand DNA intermediate [N.D.F. Grindley cited as pers. commun. in (Filee et al., 2007)]. Members of this family also occur in a group of giant eukaryotic viruses, the nucleocytoplasmic large DNA viruses (NCLDV). These infect protists whose lifestyle involves grazing on various bacteria (Filée et al., 2007). They have presumably been incorporated into the viral genomes together with other bacterial genes by virtue of the highly promiscuous recombination properties of the virus. Interestingly, copies of IS607‐like elements have more recently been identified in several ‘lower’ eukaryote genomes (Rolland et al., 2009; Gilbert & Cordaux, 2013). Most are incomplete although complete copies were identified in the genomes of the protist Acanthamoeba castellani and the alga Ectocarpus siliculosus (Gilbert & Cordaux, 2013) (Boocock & Rice, 2013). These are closely related to the cyanobacterial ISArma1. In addition to the transposase, members of this family also sometimes include a tnpB gene similar to those carried by the IS200/IS605 family (Kersulyte et al., 2000).
Although there are isolated incidences of prokaryotic ISs identified in eukaryote genomes [e.g. IS5 in Rotifers; (Gladyshev & Arkhipova, 2009)], IS607 is at present the only example of a prokaryotic IS identified in multiple eukaryotic genomes. However, as is probable in the case of NCLDV, it is possible that these have been inherited as part of a larger DNA segment rather than by transposition (Filée et al., 2007).
Orphan ISs
In addition to these three major IS groupings, there are also several families for which the transposase signature is not clear. Although many have a potential DDE motif (e.g. ISL3, ISAs1 or the newly defined ISAzo13 family), experimental proof will be required to confirm the importance of these residues in catalysis. Another group, ISNCY (not classified yet), is composed of small numbers of unclassified ISs or orphans. Members of this group often emerge as families, or new distant groups of a known family, as more examples are added to the database.
ISs with accessory genes
Many ISs also include accessory genes involved in regulating their transposition. These are relatively specific for each IS family and thus also serve in definition of the family. However, in many cases, the exact role and activity of the gene product is unclear.
IS21
IS21 family members encode a ‘helper’ gene, istB (Berger & Haas, 2001) which exhibits some similarity to the DnaA replication initiator protein due to the presence of an ATP binding motif, and often appears in blast searches of complete genomes. The molecular details of IstB activity are not known.
IS200/IS605 and IS607
Although IS200/IS605 and IS607 family members carry very different types of transposase (see above), they often include a second orf, tnpB, in addition to their transposases (Fig. 2). TnpB is not required for transposition either in vitro or in vivo (Kersulyte et al., 1998; Ton‐Hoang et al., 2005). However, it has been observed to reduce ISDra2 (IS200/IS605 family) transposition activity both in its original host, Deinococcus radiodurans, and in E. coli (Pasternak et al., 2013). The molecular details of TnpB activity are not known.
Full‐length TnpB includes three domains, an N‐terminal HTH, a central domain and a C‐terminal zinc finger (ZF) domain (Pasternak et al., 2013). However, this TnpB configuration is quite variable and there are a large number which appear to be undergoing decay. TnpB analogues have been identified as part of IS607‐like elements in the Mimi virus and other NCLDV (Filée et al., 2007). They have recently been identified in a variety of eukaryotic genomes sometimes associated with other TEs (Bao & Jurka, 2013; Gilbert & Cordaux, 2013).
IS66
In addition to its DDE transposase gene, tnpC, IS66 can include two additional genes, tnpA and tnpB, whose function is as yet unknown (Han et al., 2001) (Fig. 4a). The three reading frames are disposed in a pattern suggesting translational coupling: tnpB is in general in translational reading frame ‐1 compared to tnpA and in most cases the termination codon of tnpA and the initiation codon of tnpB overlap (ATGA).
Figure 4.
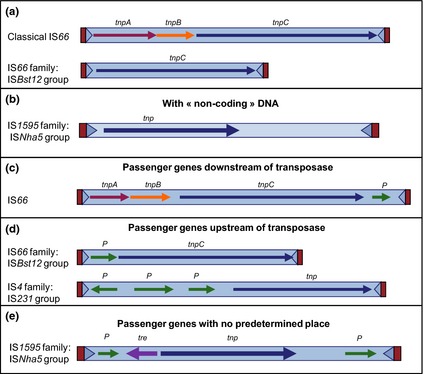
tIS: IS with passenger genes. The IS is represented as a rectangle with flanking direct repeats (DR) in red and terminal inverted repeats in blue (triangles). The Tpase orfs are shown in dark blue and passenger genes in green. (a) Organisation of a classical IS66 family member and of the ISBst12 group. The ‘accessory’ genes tnpA and tnpB are shown in red and orange, respectively. (b) An IS1595 family member with noncoding DNA. (c) A tIS 66 with a single orf downstream of the Tpase. (d) IS66 and IS4 tIS with passenger gene(s) upstream of the Tpase. (e) An IS1595 family tIS with upstream and downstream passenger genes.
IS91
While the canonical IS91 carries only a single orf, encoding an HUH Y2 transposase (Chandler et al., 2013), several other family members (e.g. ISAzo26; ISCARN110; ISMno23; ISSde12; ISShvi3; ISSod25 and ISWz1) include a second orf located upstream. This orf is a Y‐recombinase related to the phage integrase family whose role in transposition remains to be determined.
Tn3 family
The Tn3 family is an extensive group of transposons which encode large (> 900 aa) DDE transposases. They are included here because certain family members resemble ISs (e.g. IS1071) as they encode only the transposase. However, the replicative transposition mechanism of this family involves formation of a cointegrate in which donor and recipient replicons are fused and separated at each junction with a directly repeated transposon copy (Grindley et al., 2006). These structures must be ‘resolved’, by recombination between the two transposon copies, to generate the donor and target replicons each retaining a single transposon copy. This is accomplished by a ‘resolvase’, a site‐specific recombinase which acts at a unique DNA sequence in the transposon, the Res site. While many Tn3 family members encode a serine recombinase, several are now known to carry a tyrosine site‐specific recombinase (TnpI in Tn4655 and Tn4330) (Vanhooff et al., 2006) resembling phage integrases. Moreover, a third group of Tn3 family members include two genes, TnpS and TnpT (Yano et al., 2013). It is possible that, as for the bacteriophage λ which uses a Y site‐specific recombinase (Int) for integration together with a second protein, Xis for excision, TnpT is involved in assuring directionality in transposition. There is no evident difference in the Tpases of these two Tn3‐like groups.
IS derivatives
We have already indicated that the classical IS model (a single orf encoding a DDE transposase, two terminal IRs and flanking DRs) is not universal and that other models such as the IS200/IS605 and IS91 families exist and are also widespread (see ‘Major IS groups are defined by transposase type’ above). Below, we describe a variety of IS‐related TEs which share different levels of similarity with ISs. These include both autonomous (encoding a Tpase) and nonautonomous TEs (lacking a Tpase and whose transposition requires the Tpase of a related element in the same cell) and TEs with passenger genes not implicated in transposition or its regulation (Figs 4 and 5).
Figure 5.
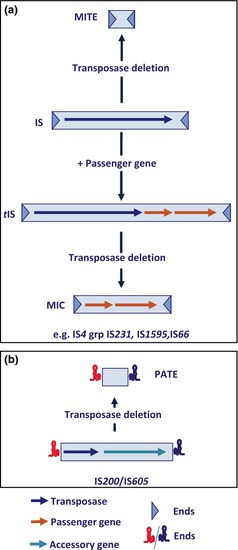
Relationship between IS, tIS and MITES. (a) The (hypothetical) relationship between different IS derivatives (shown as light blue boxes). Horizontal arrows indicate open reading frames encoding the Tpase (dark blue), passenger genes (orange). The terminal inverted repeats are shown as darker blue triangles. Examples of IS families which include such derivatives are indicated at the bottom of the panel. (b) The particular case of MITES derived from IS 200/IS 605 family members. The IS ends with their essential secondary structures are shown in red (left end) and blue (right end). The colour scheme is as described for (a). The TnpB accessory gene is shown as a green horizontal arrow.
ISs and relatives with passenger genes
An increasing number of ISs carrying passenger genes are being identified. These include genes for transcription regulators, methyltransferases and antibiotic resistance (see Fig. 4). They can be located upstream, downstream or on both sides of the transposase gene (Fig. 4c, d and e). These elements, simpler than known Tns, are called transporter ISs (tISs) (Siguier et al., 2009). They can be significantly longer than typical ISs (e.g. ISCausp2, 7915 bp). Several of these ‘extended’ ISs include a significant length of DNA with no clear coding capacity (e.g. ISBse1, ISSpo3, ISSpo8) (Fig. 4b). At least some incomplete ISs presently identified in sequenced genomes may be of this type because the second IS end would occur at an unexpectedly distant position and would not necessarily have been identified.
tISs are generally present in low copy number. Many occur only in single copy in a given genome raising the question of whether they are active. Moreover, more than one closely related but nonidentical derivative can be found in a single genome (e.g. ISSpo3, and ISSpo8 in Silicibacter pomeroyi DSS‐3 and ISNwi4 and ISNha5 in Nitrobacter winograski Nb‐255). Others are present in more than one copy. ISPre3, an IS66 relative from Pseudomonas resinovorans plasmid pCAR1 includes a hypothetical protein and is present as 2 copies with different insertion sites as judged by their typical 8 bp DRs (E. Gourbeyre, unpublished).
The mechanisms involved in acquisition of additional genes to generate tISs are at present unclear. They do not appear to carry programmed recombination systems as do some members of the Tn3 family (see ‘ISs with accessory genes: Tn3 family’ above). One possibility is that tISs are derived by deletion from ancestral compound transposons. These are composed of two ISs flanking any DNA segment either in direct or inverted orientation (Craig et al., 2002). The flanking ISs are able to mobilise the intervening DNA segment. They were among the first types of transposon described and include the models Tn5 (flanking IS50) and Tn10 (flanking IS10) as well as Tn9 (flanking IS1). As these early examples, many other such composite transposons have been identified either by experiment or from genome sequencing (Tn number registry: http://www.ucl.ac.uk/eastman/research/departments/microbial-diseases/tn).
Some early observations concerning Tn5 and Tn10 suggest that the flanking ISs can undergo mutation rendering them less autonomous [for example mutations within one IS which inactivate its transposase; see (Mahillon & Chandler, 1998)]. Furthermore, studies on IS101 from the pSC101 plasmid clearly indicated that transposition can occur using one established IS end and a second surrogate end located at some distance from the IS (Machida et al., 1982). If, as in this case, the intervening DNA includes a passenger gene, this creates a novel transposon. It has also been observed that other ISs such as IS911 can use surrogate ends during transposition (Polard et al., 1994). Moreover, isolated individual IS ends are often observed in sequenced genomes and could provide a source of surrogate ends.
The Tn3 transposon family derivatives
Tn3 family members are quite variable: several examples lack passenger genes and therefore do not fall into the strict definition of a transposon, while others lack both passenger and resolvase genes (e.g. ISVsa19, ISShfr9, ISBusp1, IS1071…) and therefore closely resemble ISs. Many family members carry a number of passenger genes. These can represent entire operons, notably mercury resistance, or individual genes involved in antibiotic resistance, breakdown of halogenated aromatics or virulence [e.g. (Liebert et al., 1999)]. They often carry integron recombination platforms enabling them to incorporate additional resistance genes by recruiting integron cassettes (Mazel, 2006).
IS‐related ICEs
Other structures which obscure the definition of an IS have been identified among various TEs. For example, ICEs (integrative conjugative elements) are found integrated into the host genome but can excise and transfer from cell to cell. Their insertion and excision are generally catalysed by enzymes related to site‐specific recombinases, whereas their transfer depends on a second set of proteins, which includes a ‘relaxase’, often a single‐strand endonuclease of the HUH superfamily (Burrus et al., 2002) (Chandler et al., 2013). However, ISSag10, a tIS member of the IS1595 family from Streptococcus agalactiae which includes an O‐lincosamide nucleotidyltransferase gene, encodes a DDE transposase and undergoes cell‐to‐cell transfer when complemented with an autonomous ICE, Tn916 (Achard & Leclercq, 2007). In this case, a cryptic origin of transfer is located within the 3′ end of the resistance gene. These nonautonomous ICEs have been called IMEs [integrative mobilisable elements; (Adams et al., 2002)] or cis‐mobilisable elements [CIMEs; (Pavlovic et al., 2004)].
More recent studies have identified a new ICE family, transposon of Group B Streptococcus (TnGBS), in which the enzyme catalysing their integration and excision belongs to another DDE‐group Tpase (Brochet et al., 2009; Guerillot et al., 2013). Indeed, this has led to the identification of an entirely new family of classic ISs carrying DDE Tpases, the ISLre2 family (Guerillot et al., 2014). In addition to the TnGBS family, other ICEs have been identified which include a DDE Tpase closely related to that of the IS30 family (Smyth & Robinson, 2009). It seems likely that examples of ICEs with other IS family Tpases are awaiting identification. Moreover, in addition to a variety of transfer functions, certain ICEs carry plasmid‐related replication genes important in ensuring sufficient stability of the transposition intermediates to enable their subsequent integration (Lee et al., 2010; Guerillot et al., 2013). Indeed, early examples of ICEs (Murphy & Pembroke, 1995) were initially thought to be resistance plasmids and assigned an incompatibility group, incJ. These are maintained as an integrated copy in the host chromosome but can nevertheless give rise to circular copies (Pembroke & Murphy, 2000). This is yet another example of the increasingly indistinct frontiers between phage, plasmids and transposons (Bi et al., 2012) (http://db-mml.sjtu.edu.cn/ICEberg/).
IS91‐related ISCRs
These MGE include a putative transposase of the HUH family similar to that of the IS91 family, the so‐called ‘common region’ (CR). Although it has yet to be demonstrated that such structures indeed transpose, ISCRs are associated with multiple flanking antibiotic resistance genes. It is thought that these genes are transmitted during the rolling circle type of transposition mechanism postulated to occur in IS91 transposition. This involves an initiation event at one IS end, polarised transfer of the IS strand into a target molecule and termination at the second end (Garcillan‐Barcia et al., 2002). Flanking gene acquisition is thought to occur when the termination mechanism fails and rolling circle transposition extends into neighbouring DNA where it may encounter a second surrogate end (Garcillan‐Barcia et al., 2002). This type of mobile element may prove to play an important role in the assembly and transmission of multiple antibiotic resistance (Toleman et al., 2006) (Toleman & Walsh, 2011).
Nonautonomous derivatives: Miniature Inverted repeat TEs (MITEs), Mobile Insertion Cassette (MICs), Palindrome‐associated TEs (PATEs)
Structures called MITEs (Feschotte et al., 2002) are related to ISs with DDE Tpases (Fig. 4a). They are composed of two appropriately oriented left and right ends similar to those of ISs. They are generally < 300 bp long and are probably derived from ISs by internal deletion. Some carry short noncoding sequences between these IRs which may or may not be IS‐derived. Others carry various coding sequences and have been called MICs (De Palmenaer et al., 2004). MITEs are considered to be nonautonomous TEs mobilisable in trans by Tpases of full‐length parental genomic copies. They were first identified in plants (Feschotte et al., 2002) and are related to Tc/mariner elements (distantly related to bacterial IS630 family). IS630‐related MITEs were also the first described bacterial examples (Correia et al., 1988; Oggioni & Claverys, 1999; Buisine et al., 2002). MITEs showing similarities to other IS families have now been observed in bacteria and archaea (Brugger et al., 2002; Filee et al., 2007). Among those derived from ISs with DDE transposases, representatives of the IS1, IS4, IS5, IS6, and even Tn3 family members such as ISRf1 from Sinorhizobium fredii have been identified.
MITE‐like structures related to elements with other types of transposase have also been identified. Among these are IS200/IS605 family derivatives (Filee et al., 2007); (P. Siguier, unpublished) now called PATEs (Dyall‐Smith et al., 2011) (Fig. 4b) reflecting the subterminal secondary structures which constitute the ends of these ISs. Although originally observed in the Archaea as derivatives of known IS200/IS605 family members, they have also been observed in certain cyanobacteria and Salmonella (P. Siguier, unpublished). They presumably represent decay products that appear quite frequently for this IS family.
Full‐length copies of the parental IS may not be available or may be so divergent as to escape detection in standard blast analysis. This is the case for certain MITEs from the archaea (Brugger et al., 2002) and is probably also true for the bacteria. Their detection and analysis are therefore arduous.
A good example of an IS group that includes examples of many of these IS derivatives (canonical ISs, MITEs, MICs and tISs) is the IS231 subgroup of the large IS4 family [Fig. 4; (De Palmenaer et al., 2004)].
Characteristics of family life
Having presented an overview of different characteristics that contribute to assignment of an IS to a specific family, we detail below some of the more specific features which are used to group ISs into families and which impact on their behaviour.
Transposases and mechanism
Transposition requires a set of DNA cleavages at the ends of a TE and a set of strand transfer reactions which move these ends into a suitable target DNA molecule catalysed by the TE‐encoded Tpase. Transposases are often multidomain proteins. In addition to catalysis, they must also recognise specific DNA sequences at the IS ends and engage with the DNA target to form multimeric nucleo‐protein assemblies, transpososomes. These nucleo‐protein assemblies provide a precise architecture within which the chemical steps of transposition are carried out. They are composed of two or more Tpase monomers and, in some cases, accessory proteins such as DNA‐architectural proteins (for example IHF, HU and HNS; reviewed in [Mizuuchi, 1992a, b; Haniford, 2006)] or possibly protein chaperones [e.g. GroEL in the case of IS1; (Ton‐Hoang et al., 2004)]. They are dynamic and undergo conformational changes to coordinate DNA cleavages and strand transfers and ensure that, once started, transposition goes to completion (Dyda et al., 2012; Montano et al., 2012).
DDE transposases
Many of the presently identified Tpases are members of the DDE family (Fig. 1; Table 1). These are structurally and catalytically related to RNaseH and other nucleic acid processing enzymes (Rice et al., 1996) and are said to have an RNaseH fold. The highly conserved DDE triad serves to coordinate one or two divalent cations such as Mg2+ which in turn assist polarisation of the phosphate group belonging to the target phosphodiester bond, facilitating cleavage. DDE enzymes use hydroxyl groups as nucleophiles for cleavage and for strand transfer: H2O for initial cleavage and a 3′OH, generated by cleavage of the TE ends, for the strand transfer reaction [reviewed in (1992a, b; Hickman et al., 2010a, b)].
The DDE triad is often followed by a basic amino acid residue, generally K or R, 7 amino acids downstream and on the same face of the α‐helix that also carries the final conserved E residue. Differences in the DDE spacing as well as the presence or absence of specific submotifs (for instance IS4 family transposases include the conserved N2 N3 C1 signatures which carry the D, D and E residues and a motif YREK [Y‐(2)‐R‐(3)‐E‐(6)‐(K)]; (Rezsohazy et al., 1993; De Palmenaer et al., 2008) are used in distinguishing different groups and families (Table 1). The earliest DDE motifs from transposases, those of the IS3 family, were identified by their similarities with retroviral integrases (IN) (Fayet et al., 1990) and, like IN, have a D35E spacing. Other DDE‐group enzymes exhibit a spacing of 33–35 residues between the conserved D and E. However, the DDE transposases of several families carry relatively long distinctive insertion domains of either α‐helical or β‐strand (Hickman et al., 2010a, b).
Certain IS groups appear to carry variants of the conserved DDE motif with N or H residues replacing the E. The role of these alternative residues in catalysis has yet to be tested. Some groups, for example the IS1595 family, include several of these variant motifs (Table 1) (Siguier et al., 2009) and the fact that these IS exist in several copies suggests that they are active.
DDE enzymes catalyse cleavage of only one DNA strand generally generating a 3′OH at the IS end. This is known as the transferred strand because the 3′OH is used as a nucleophile in the integration step to attack the target phophodiester bond and complete strand transfer. However, transposition of these ISs occurs via double‐strand DNA intermediates and therefore requires processing of the second strand (called the nontransferred strand) to liberate the IS from flanking DNA sequences in the donor molecule. This can occur in several different ways and is also IS family‐specific (Turlan & Chandler, 2000; Curcio & Derbyshire, 2003) (Table 1) and serves to reinforce groupings derived from bioinformatics comparison by providing a mechanistic coherence. Thus, for ISs of the IS4 family (IS50, IS10), the initial 3′OH is used to attack the opposite strand forming a transient hairpin bridge at the IS end. This is then cleaved using H2O as a nucleophile to liberate the IS and regenerate the 3′OH on the transferred strand ready for strand transfer and integration. This is known as a cut‐and‐paste mechanism. Other ISs such as the IS630 family, related to the eukaryote mariner/Tc superfamily, employ a mechanism in which the initial H2O‐catalysed cleavage of the nontransferred strand occurs with a small offset of 2 bases into the IS prior to cleavage of the transferred strand (Feng & Colloms, 2007) as do their eukaryotic cousins (Richardson et al., 2009).
An additional mechanism adopted by certain elements with DDE Tpases is cointegrate formation. Here, the transposon inserts into a target replicon in a process accompanied by TE replication. This results in fusion of the donor and target replicons with a directly repeated TE copy at each junction known as a Shapiro intermediate (Shapiro, 1979). Tn3 and IS6 family members transpose using this pathway as does bacteriophage Mu and its relatives (Chaconas & Harshey, 2002).
However, by far the most common mechanism is the so‐called copy–paste mechanism (Curcio & Derbyshire, 2003), which generates a transient double‐strand circular DNA intermediate. This has been adopted by a significant number of IS families including IS3, IS30, IS110, IS256, ISLre2 and possibly others. IS1 uses this as one of several transposition pathways (Turlan & Chandler, 1995). For IS3 family members, circle formation occurs in an asymmetric manner. One IS end is cleaved to generate the characteristic 3′OH of the transferred strand. This then serves to attack several nucleotides exterior to the second end to generate a single‐strand bridge leaving a free 3′OH on the IS flank. The 3′OH can act as a replication primer. IS replication would regenerate an intact copy reconstituting the donor plasmid and produce a double‐strand circular DNA intermediate. Due to low basal Tpase levels, this initial step may occur in a stochastic manner. However, formation of the circular intermediate results in the assembly of a transient strong promoter composed of a ‐35 promoter element in the right IS end oriented outwards and a ‐10 promoter element in the left end oriented inwards (Ton‐Hoang et al., 1997). This promoter serves to drive transposase synthesis and consequent integration and disassembly of the promoter. Thus, the circular intermediate once generated is committed to terminate transposition.
DEDD transposases
Another motif, DEDD (Buchner et al., 2005) is characteristic of the four‐way Holliday junction (HJ) resolvase, RuvC. RuvC also has an RNaseH fold (Ariyoshi et al., 1994). A DEDD motif has also been identified in transposases of the IS110 family (Tobiason et al., 2001), closely related to the Piv and MooV invertases from Moraxella lacunata/M. bovis (Fulks et al., 1990; Rozsa et al., 1997) and N. gonorrhoeae (Choi et al., 2003; Skaar et al., 2005), respectively. Piv catalyses inversion of a DNA segment permitting expression of a type IV pilin. However, the organisation of IS110 family members and the inversion systems are different. In the IS, the recombinase is located within the element, whereas in the inversion systems, it is located outside the invertible segment (Buchner et al., 2005).
Although it has proved difficult to determine the activity of these transposases in detail in vitro, transposition of ISs with DEDD Tpases may be unusual and involve HJ intermediates that must be resolved using a RuvC‐like mechanism. This type of recombination would be consistent with the close relationship between DEDD transposases and the Piv/MooV invertases that presumably resolve HJ structures during inversion (Tobiason et al., 1999).
HUH transposases
The second major group of transposases are the HUH enzymes [for a review see (Chandler et al., 2013)]. HUH refers to a pair of His residues (H) separated by a bulky hydrophobic residue (U) (Ilyina & Koonin, 1992). Together with a third residue, the two His coordinate a divalent metal ion cofactor required for catalysis. These enzymes also include one or two tyrosine (Y) residues, which act as nucleophiles and form transient 5′ phosphotyrosine enzyme–DNA transposition intermediates. Like DDE enzymes, HUH enzymes are widespread and assume other roles in the cell. In addition to transposition, they are involved in rolling circle plasmid and phage replication, in rolling hairpin replication of eukaryotic viruses (Rep proteins) and conjugative plasmid transfer (relaxases or Mob proteins).
IS200/IS605‐family transposases
These Y1 Tpases are among the smallest identified to date with c. 150 amino acids. These promote single‐strand transposition (Ronning et al., 2005; Ton‐Hoang et al., 2005) in contrast to DDE enzymes, which catalyse double‐strand transposition. They form obligatory dimers in which the catalytic site is composed of an HUH motif from one monomer and a single Y residue together with the third residue necessary for coordinating the essential divalent metal ion contributed by the second monomer (Ronning et al., 2005). Although the founding family member, IS200, was identified in Salmonella 30 years ago (Lam & Roth, 1983), their activities have only recently been unravelled (Ronning et al., 2005; Ton‐Hoang et al., 2005, 2010; Barabas et al., 2008; Guynet et al., 2008, 2009). Two model ISs, IS608 from Helicobacter pylori and ISDra2 (Hickman et al., 2010a, b; Pasternak et al., 2010; Ton‐Hoang et al., 2010) from D. radiodurans [IS8301 in (Islam et al., 2003)] have been studied in detail. Both use obligatory circular ssDNA intermediates for their mobility. They excise as ssDNA circles with abutted left and right ends and insert 3′ to a conserved element‐specific penta‐ or tetra‐nucleotide target (Guynet et al., 2009). As the transposon is ‘peeled’ from its donor site as a single strand, this mechanism might be called ‘peel‐and‐paste’ (B. Ton‐Hoang pers. commun.). The target sequence is essential for further transposition (Ton‐Hoang et al., 2005). The transposase, TnpA, recognises small, subterminal hairpin structures in a strand‐specific manner and catalyses single‐strand cleavage at both ends on the ‘top’ strand. Surprisingly, the left and right cleavage sites are not directly recognised by TnpA. Instead, they form a network of particular base interactions with short ‘guide’ sequences located 5′ to the TnpA‐bound subterminal secondary structures at each end. This network includes both canonical (Watson‐Crick) and noncanonical base interactions (Barabas et al., 2008; He et al., 2011) (Hickman et al., 2010a, b). These are also essential to stabilise the nucleoprotein complex, the transpososome, within which the DNA strand cleavages and transfers occur. Indeed, changes in the guide sequences result in predictable changes in insertion site specificity (Guynet et al., 2009).
Family members excise from, and insert preferentially into, the lagging strand template of chromosome and plasmid replication forks (Ton‐Hoang et al., 2010). Their lagging strand preference generates an insertion bias reflecting the mode of replication (uni‐ or bi‐directional) of the target replicon. Moreover, transposition of ISDra2 is strongly induced upon recovery of the highly radiation‐resistant D. radiodurans host from irradiation (Pasternak et al., 2010) as a result of the large amounts of ssDNA generated during the reassembly of the shattered D. radiodurans genome (Zahradka et al., 2006).
IS91/ISCR‐family transposases
The second major HUH Tpase group, those of the IS91 family, has been known for some time (Garcillan‐Barcia & de la Cruz, 2002; Garcillan‐Barcia et al., 2002). They were recognised as relatives of the rolling circle plasmid replication Rep proteins. They are larger than the simple Y1 transposases of the IS200/IS605 family, carry a pair of tyrosine residues that are both necessary for transposition (Garcillan‐Barcia et al., 2002) together with an N‐terminal ZF motif and are known as Y2 transposases. Tpases of the related ISCR elements are similar to IS91 transposases but include only a single Y residue. Although there is no information concerning the transposition of ISCRs, IS91 is thought to transpose by a rolling circle replication mechanism (RCR) initiating at one end (ori, 3′ to the transposase) and terminating at the other (ter, 5′ to the transposase) (Garcillan‐Barcia et al., 2002). Relatively, frequent failure of correct termination (1%) results in transposition of additional genes that flank the transposon in the donor molecule. Indeed, ori is essential for activity while removal of ter reduces but does not eliminate transposition.
Insertion of IS91, like that of members of the IS200/IS605 family, is oriented with ori adjacent to the 3′ of a specific tetranucleotide target (5′‐CTTG or 5′‐GTTC) and, like that of the IS200/IS605 family, this target sequence is essential for further transposition (del Pilar Garcillan‐Barcia et al., 2001; Garcillan‐Barcia et al., 2002). In the proposed RCR mechanism, displacement of an IS91/ISCR active transposon strand would be driven by leading strand replication of the donor replicon from a 3′OH generated by cleavage at ori. Although the original model proposed that the cleaved IS end is transferred to the target DNA and the IS is replicated ‘into’ the target replicon, single‐strand and double‐strand IS91 circles have been observed and it is difficult to explain these as intermediates in the RCR model.
Serine transposases
IS607 family members encode a Tpase closely resembling serine site‐specific recombinases that use serine as a nucleophile for cleavage of the DNA strand (Grindley, 2002). At present, little is known about transposition of this IS family although it is thought that these elements generate circular intermediates [N.D.F. Grindley pers. commun. cited in (Filee et al., 2007)]. Presumably, the enzyme catalyses similar cleavages and strand transfers as its site‐specific serine recombinase cousins using a transitory 5′ phosphoserine covalent intermediate. Based on transposase structures from structural genomics studies and detailed knowledge of the general serine recombinase mechanism, (Boocock & Rice, 2013) have proposed a model for the transposition mechanism. This includes a synaptic transposase tetramer (as for classical serine recombinases). The model explains the lack of target specificity exhibited in IS607 transposition (Kersulyte et al., 2000), behaviour which is unusual for this type of recombinase.
Transposase organisation and its role in regulation: domain structure
Transposases are composed of a combination of domains with recognisable secondary structures. In many cases, the presence and order of these domains defines subgroups within an IS family.
Although there are only a limited number of Tpase structures available, these secondary structures can be predicted from the primary amino acid sequence and include ZF (Zn), helix‐turn‐helix (HTH), leucine zipper (LZ), RNaseH fold or HUH domains (see references in (Montano & Rice, 2011; Dyda et al., 2012).
DDE transposases
For DDE transposases, the domains of the protein which ensure sequence‐specific DNA binding (i.e. to the IS ends) are typically located towards the N‐terminal (N‐terminal) end and the catalytic domain more towards the C‐terminal (C‐terminal) end. From a functional point of view, this arrangement facilitates a phenomenon called cotranslational DNA binding in which the protein folds in the course of translation allowing the nascent polypeptide chain to initiate binding directly to the closest IS end. Indeed, for members of a number of IS families, including IS1 (Zerbib et al., 1987), IS3 (Haren et al., 1998), IS30 (Stalder et al., 1990), the isolated DNA‐binding domain bind more avidly than the entire protein, suggesting that the C‐terminal end may actually inhibit specific binding by the N‐terminal domain, possibly by steric masking. Once bound, the N‐terminal domain would be insensitive to C‐terminal domain masking. Thus, specific Tpase binding would only occur transitorily early during translation providing an attractive explanation for the preferential cis activity (the preference to act on the TE from which the Tpase is expressed) of many Tpases (Duval‐Valentin & Chandler, 2011). Both Helix‐turn‐Helix (HTH) and ZF domains have been implicated as DNA‐binding domains (e.g. IS911 (Rousseau et al., 2004); IS1 (Ohta et al., 2004; Ton‐Hoang et al., 2004). These can be found individually or together as illustrated in Fig. 6.
Figure 6.
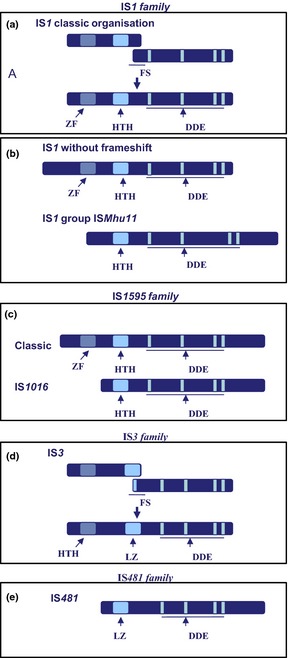
Domain organisation of transposases of the DDE family. The relative positions of the potential ZF, HTH, LZ and the ‘DDEK/R’ catalytic motif are indicated from left to right as light blue boxes. The figure illustrates the N‐terminal and C‐terminal extension of the different transposase examples. (a) Classical IS 1 with frameshift. The position of the frameshift window which is used to generate InsAB is indicated. (b) IS 1 without frameshift and the ISM hu11 group showing the deletion of the ZF, the C‐terminal extension and the increased spacing between the second (d) and (e) residues. (c) The IS 1595 family showing the classical IS 1595 group and the IS 1016 group which does not carry the N‐terminal ZF. (d) The IS 3 family including members with and without the translational frameshift. (e) The closely related IS 481 family which lack the N‐terminal HTH domain and exhibit an additional C‐terminal domain.
The ability to multimerise is another important Tpase property. This is because the TE ends are typically both involved simultaneously in the transposition process within the transpososome. Multimerisation can serve as an important regulatory mechanism because a Tpase monomer bound to one TE end often carries out its catalytic activities on the other end. This assures that catalysis only occurs once the transpososome complex has been correctly assembled and is primed to complete the transposition process.
Although multimerisation can be complex and can involve several different Tpase regions (Braam et al., 1999), in the case of IS3 family members, this is accomplished by a coiled‐coil LZ structure (Haren et al., 2000).
Several examples of variations in domain organisation found in related DDE transposases are shown in Fig. 6. For IS1, the transposase is generally expressed as a fusion protein by programmed translational frameshifting (Fig. 6a; see Recoding and domain organisation below). However, derivatives are found in which the transposase is produced from one continuous frame. In these cases, derivatives which have been truncated from the N‐terminal ZF or with additional N‐terminal extensions have been identified (Fig. 6b). Similar truncated derivatives are also found in the case of the IS1595 family distantly related to IS1 (Fig. 6c) and the large closely related IS3 and IS481 families (Fig. 6d and e).
DEDD Tpases
It is interesting to note that in the related DEDD Tpases that are related to the Piv and MooV invertases and limited at present to IS110 family members, the potential DNA‐binding domain is located downstream of the catalytic domain (Choi et al., 2003). This again reinforces the idea that the DEDD ISs possess a different transposition mechanism to those with DDE Tpases.
HUH transposases
The smaller Y1 HUH Tpases of the IS200/IS605 family are very compact with DNA binding, cleavage site and target recognition functions closely interwoven. The active Tpase is dimeric and, in the published protein structures, the active site is composed of the HUH motif of one monomer and the active site tyrosine of the other. The tyrosine residue is located downstream of the HUH on an α‐helix joined to the body of the protein by a flexible arm. The α‐helix also carries a third residue which, with the His pair, completes the amino acid triad essential for divalent metal ion binding (Ronning et al., 2005; Barabas et al., 2008), (Hickman et al., 2010a, b). Each Y residue in this ‘trans’ position cleaves one IS end generating a 5′ phosphotyrosine bond. Strand transfer is then thought to occur by rotation of the flexible arms so that the Y residue covalently linked to DNA engages with the HUH of the same monomer, a ‘cis’ configuration. The cleavage reaction is then reversed resulting in strand transfer and the enzyme then appears to be reset in the trans configuration (He et al., 2013).
The overall organisation of the much larger Y2 enzymes is less well determined although there is a ZF motif which possibly functions as a DNA recognition domain located in the N‐terminal region (Chandler et al., 2013).
Serine transposases
The family of serine recombinases is composed of three groups: the resolvase/integrase group; the large serine recombinases; and the serine transposases (Boocock & Rice, 2013). For the two former groups, the catalytic domain is invariably located at the N‐terminus and is followed by a sequence‐specific DNA‐binding domain (a simple HTH for the resolvase/invertase group, or a much larger domain of unknown structure in the large serine recombinases Rutherford [Van Duyne & Rutherford, 2013)].
It is interesting to note that the IS607 family serine transposases carry their DNA‐binding domain N‐terminal to the catalytic domain (Gilbert & Cordaux, 2013) (Boocock & Rice, 2013) in a similar way to other (DDE) transposases and may reflect a similar function.
Recoding and domain organisation
Certain TEs can express combinations of different functional protein domains, providing a way of encoding two proteins of different function in one DNA segment and resulting in highly compact genetic structures. These types of noncanonical readout of the genetic code are collectively known as ‘Recoding’ (Gesteland et al., 1992). Recoding is significantly more frequent and well documented in MGE‐encoded genes of both prokaryotes and eukaryotes than in any other gene family (Baranov et al., 2005; Sharma et al., 2011).
Programmed ribosomal frameshifting
The most commonly encountered recoding event is probably programmed ribosomal frameshifting (PRF) involving ribosome slippage of 1 nt. This often occurs in the 5′ direction to generate a ‐1 frameshift and requires specific signals in the mRNA such as an upstream RBS, a sequence facilitating slippage (slippery codons) and downstream secondary structures (hairpins and pseudoknots) (Sharma et al., 2011).
PRF occurs in retroviruses, coronaviruses and some plant viruses. It is also very common in prokaryotic ISs. Initially, PRF signals were identified in the transposase genes of IS1 and IS3 family members (Sekine & Ohtsubo, 1989; Escoubas et al., 1991; Polard et al., 1991; Sekine et al., 1994) where they serve to fuse the product of the upstream frame, generally a DNA‐binding protein which, on its own, acts as a regulator, to the downstream catalytic domain (Zerbib et al., 1990; Ton‐Hoang et al., 1998; Rousseau et al., 2007).
Programmed transcriptional realignment
A less well‐characterised phenomenon is programmed transcriptional realignment (PTR) (Sharma et al., 2011). This is mechanistically distinct from PRF and is due to insertion or deletion of a few nucleotides by transcription ‘slippage’ leading to different types of frameshift (+/−: 1, 2…). Although PTR needs signals such as a run of A, T or AnGn, these are not as distinct as those involved in PRF and the phenomenon is less well studied principally because it is more difficult to address experimentally.
Two additional families (IS5 sgr IS427 and certain IS630) whose transposases are also distributed over two consecutive reading phases have been identified although at present it is unclear whether transposase expression involves PRF or PTR.
Pyrrolysine‐ and selenocysteine‐mediated stop codon read‐through
Less common than PRF and PTR are recoding events involving stop codon (UAG) read‐through by insertion of the atypical amino acids pyrrolysine and selenocysteine (Söll, 2007). Insertion sites are characterised by a secondary structure element located downstream of the stop codon respectively called PYLIS (Namy et al., 2004) (Ibba & Soll, 2004) and SECIS (Liu et al., 1998). Certain ISs probably use stop codon read‐through to control transposase expression. These have been identified in organisms that encode these read‐through mechanisms such as Methanosarcina (ISMac17, ISMba9 and ISMma13 in the case of pyrrolysine) and Desulfovibrio vulgaris (ISDvu3, in the case of selenocysteine). As UAG read‐through has clearly evolved as a more general regulatory mechanism in these organisms, this implies that the IS in question have taken advantage of these host read‐through regulatory pathways and have evolved with their hosts to limit their transposition activity.
Domestication
One important aspect of the impact of ISs which has not been addressed in detail is the capacity of the host to harness their properties to perform various cellular functions. Such domestication has been described repeatedly in eukaryote systems (Sinzelle et al., 2009) but remains relatively rare in the prokaryotes. However, in addition to their activities in modulating gene expression (see Impact of ISs on genome expression above), there are two interesting examples of prokaryotic IS domestication. Both concern HUH enzymes related to those of the IS200/IS605 family (see “Major IS groups are defined by transposase type’ above).
The REP system
In one case, a TnpA‐like enzyme, TnpAREP (Ton‐Hoang et al., 2012) or RAYT (Rep Associated Tyrosine Transposase, (Nunvar et al., 2010) is associated with generally high copy number sequences for REP originally identified in the enterobacteria (Higgins et al., 1982). These resemble the ends of IS200/IS605 family members in structure and can occur in clusters localised around the chromosome. Some are grouped into pairs called bacterial interspersed mosaic elements (BIMES). They are present in many bacteria (Tobes & Ramos, 2005; Nunvar et al., 2010, 2013) and have several key roles in aspects of host physiology, including organisation of genome structure (Gilson et al., 1990; Boccard & Prentki, 1993; Espeli & Boccard, 1997), regulation of gene expression (Espeli et al., 2001; Khemici & Carpousis, 2004; Moulin et al., 2005; Aguena et al., 2009) and genome plasticity (Clement et al., 1999; Wilde et al., 2003; Tobes & Pareja, 2006). TnpAREP from E. coli cleaves and rejoins REP sequences (Ton‐Hoang et al., 2012) and is very similar in structure to TnpA of IS200/IS605 family members (Messing et al., 2012). Little is known at present about the mechanisms involved in their invasion of and spread throughout their host genomes.
Group I introns
The second example is the group I bacterial intron or IStron sometimes found in the Clostridia and Bacilli, which includes TnpA‐ and TnpB‐like genes presumably as homing endonucleases (Braun et al., 2000, Hasselmayer, et al., 2004a, b). However, in a large number of IStrons, these genes are clearly undergoing decay and exist as truncated derivatives as are many of their IS cousins (P. Siguier, unpublished). The IStron TnpA is quite similar to that of ISCpe2, itself from Clostridium perfringens and to that of ISDra2. Moreover, all IStron copies are inserted 3′ to a pentanucleotide, TTGAT, the target sequence for ISDra2. TTGAT is also complementary to the intron internal guide sequence presumably required at the RNA level in the splicing reaction. Little is known about IStron behaviour.
Conclusion
Diversity and the boundaries between different TE types
We have tried here to provide an overview of the contribution of prokaryotic IS to the life of their host. We have underlined their diversity in sequence, organisation and behaviour and have attempted to describe their importance in generating genomic modifications and their contribution to modifying gene expression. We have also highlighted the growing realisation that their classification as genetic objects such as Tn, ISs and ICEs is in some ways artificial because there appears to be a large degree of diversity in these MGEs such that the border between certain types is becoming increasingly ambiguous. This is clearly illustrated by the large spectrum of interrelated mobile elements which are vehicles for antibiotic resistance. In addition to the numerous ‘classical’ composite transposons with flanking ISs and unit transposons such as those of the large Tn3 family, we have described autonomous (tISs) and nonautonomous (MICs) IS derivatives, integrative conjugative elements (ICEs) with IS‐like Tpases (Brochet et al., 2009) and ISCR with IS91‐like HUH Tpases and multiple resistance genes both upstream and downstream of the Tpase (Toleman et al., 2006). It is tempting to suggest that ISs may be major building blocks used in assembling other more complex TEs. This clearly is the case in the domesticated IStron and widely spread TnpAREP systems.
Relationship with eukaryotic TE
Although often treated separately, it is worthwhile underlining that many prokaryotic IS families (at least those with DDE transposases) have relatives in TEs identified in eukaryotes (e.g. IS1595/merlin; IS256/mutator; IS630/Tc1/mariner; IS1380/piggyBac). They have similar sizes and, where known, similar transposition mechanisms (Hickman et al., 2010a, b). Although certain elements have received detailed attention (Hua‐Van & Capy, 2008), the phylogenetic relation between most has yet to be analysed.
Future directions
One view of the prokaryote genome is as an ecological niche in which TEs and, in particular ISs, form part of a genomic landscape continuously modified by their own activity. This involves both endogenous elements and those accumulated by horizontal transfer. Our present understanding of the way in which ISs shape genomes is based largely on descriptive studies which often employ inaccurately annotated genomes. To obtain a more dynamic picture of the forces involved and the processes shaping a genome, it is necessary to understand the detailed mechanism(s) of IS movement; their target sites, that is, whether they prefer replication forks, actively transcribed regions, plasmids, chromosomes, specific DNA sequences or secondary structures and in what combinations; the interactions (interference) between elements; involvement of host factors etc. There have been few attempts to understand these factors and even fewer attempts to integrate them.
There is a need to understand TE behaviour not only on a genomic scale but within populations and communities to explain their dynamics. The role of genome isolation in IS expansion is an excellent example. To understand TE behaviour, there is a need to build quantitative multiscale models (Dada & Mendes, 2011) rather than descriptive models. This is a challenging task because this type of systems biology approach will require incorporation of a large number of very diverse parameters. At one level, this involves considerations such as how the DNA spreads through communities, the regulatory circuitry including the signals involved in DNA transfer between cells, the nature and function of the acquired mobile DNA, the way in which cells deal with the ‘intrusion’ of newly introduced genes and how they accommodate the associated changes in fitness. On a second level, it requires knowledge of the fine regulatory circuits determining TE movement itself, both biochemical and temporal, how this depends on various aspects of host physiology (interaction with DNA structural proteins, with replication repair and the cell cycle, with different chromosomal domains) and to what extent there may be interaction/interference between different TEs in the same cell.
As a final word, there are clearly more potential TE types awaiting identification and characterisation [e.g. (Ricker et al., 2013)] and these will also need to be factored into the final picture.
Supporting information
Table S1. ISs in genomes.
Table S2. Gene activation by IS insertion.
Acknowledgements
We would like to thank Richard Cordeaux, Bernard Hallet, Alison Hickman, David Lane and members of the Mobile genetic element group (Isabelle Canal, Alix Corneloup, Olivier Fayet, Cathy Guynet, Susu He, Brigitte Marty, Marie‐Françoise Prère, Anne Sarcos, Bao Ton‐Hoang) for their comments on the manuscript. This work was supported by the intramural programme of the CNRS.
References
- Achard A & Leclercq R (2007) Characterization of a small mobilizable transposon, MTnSag1, in Streptococcus agalactiae . J Bacteriol 189: 4328–4331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adams V, Lyras D, Farrow KA & Rood JI (2002) The clostridial mobilisable transposons. Cell Mol Life Sci 59: 2033–2043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aguena M, Ferreira GM & Spira B (2009) Stability of the pstS transcript of Escherichia coli . Arch Microbiol 191: 105–112. [DOI] [PubMed] [Google Scholar]
- Allewalt JP, Bateson MM, Revsbech NP, Slack K & Ward DM (2006) Effect of temperature and light on growth of and photosynthesis by Synechococcus isolates typical of those predominating in the octopus spring microbial mat community of Yellowstone National Park. Appl Environ Microbiol 72: 544–550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altschul SF, Gish W, Miller W, Myers EW & Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215: 403–410. [DOI] [PubMed] [Google Scholar]
- Andersson JO & Andersson SG (1999) Insights into the evolutionary process of genome degradation. Curr Opin Genet Dev 9: 664–671. [DOI] [PubMed] [Google Scholar]
- Andersson SG & Kurland CG (1998) Reductive evolution of resident genomes. Trends Microbiol 6: 263–268. [DOI] [PubMed] [Google Scholar]
- Ariyoshi M, Vassylyev DG, Iwasaki H, Nakamura H, Shinagawa H & Morikawa K (1994) Atomic structure of the RuvC resolvase: a Holliday junction‐specific endonuclease from E. coli . Cell 78: 1063–1072. [DOI] [PubMed] [Google Scholar]
- Aubert D, Naas T, Heritier C, Poirel L & Nordmann P (2006) Functional characterization of IS1999, an IS4 family element involved in mobilization and expression of beta‐lactam resistance genes. J Bacteriol 188: 6506–6514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aziz RK, Breitbart M & Edwards RA (2010) Transposases are the most abundant, most ubiquitous genes in nature. Nucleic Acids Res 38: 4207–4217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bao W & Jurka J (2013) Homologues of bacterial TnpB_IS605 are widespread in diverse eukaryotic transposable elements. Mob DNA 4: 12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bao W, Jurka MG, Kapitonov VV & Jurka J (2009) New superfamilies of eukaryotic DNA transposons and their internal divisions. Mol Biol Evol 26: 983–993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barabas O, Ronning DR, Guynet C, Hickman AB, Ton‐Hoang B, Chandler M & Dyda F (2008) Mechanism of IS200/IS605 family DNA transposases: activation and transposon‐directed target site selection. Cell 132: 208–220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baranov PV, Hammer AW, Zhou J, Gesteland RF & Atkins JF (2005) Transcriptional slippage in bacteria: distribution in sequenced genomes and utilization in IS element gene expression. Genome Biol 6: R25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berger B & Haas D (2001) Transposase and cointegrase: specialized transposition proteins of the bacterial insertion sequence IS21 and related elements. Cell Mol Life Sci 58: 403–419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bi D, Xu Z, Harrison EM et al (2012) ICEberg: a web‐based resource for integrative and conjugative elements found in Bacteria. Nucleic Acids Res 40: D621–D626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanc G, Ogata H, Robert C, Audic S, Claverie J‐M & Raoult D (2007) Lateral gene transfer between obligate intracellular bacteria: evidence from the Rickettsia massiliae genome. Genome Res 17: 1657–1664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boccard F & Prentki P (1993) Specific interaction of IHF with RIBs, a class of bacterial repetitive DNA elements located at the 3′ end of transcription units. EMBO J 12: 5019–5027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boocock MR & Rice PA (2013) A proposed mechanism for IS607‐family serine transposases. Mob DNA 4: 24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bordenstein SR & Reznikoff WS (2005) Mobile DNA in obligate intracellular bacteria. Nat Rev Microbiol 3: 688–699. [DOI] [PubMed] [Google Scholar]
- Braam LM, Goryshin IY & Reznikoff WS (1999) A mechanism for Tn5 inhibition. Carboxyl‐terminal dimerization. J Biol Chem 274: 86–92. [DOI] [PubMed] [Google Scholar]
- Braun V, Mehlig M, Moos M, Rupnik M, Kalt B, Mahony DE & von Eichel‐Streiber C (2000) A chimeric ribozyme in Clostridium difficile combines features of group I introns and insertion elements. Mol Microbiol 36: 1447–1459. [DOI] [PubMed] [Google Scholar]
- Brochet M, Da Cunha V, Couve E, Rusniok C, Trieu‐Cuot P & Glaser P (2009) Atypical association of DDE transposition with conjugation specifies a new family of mobile elements. Mol Microbiol 71: 948–959. [DOI] [PubMed] [Google Scholar]
- Brugger K, Redder P, She Q, Confalonieri F, Zivanovic Y & Garrett RA (2002) Mobile elements in archaeal genomes. FEMS Microbiol Lett 206: 131–141. [DOI] [PubMed] [Google Scholar]
- Brugger K, Torarinsson E, Redder P, Chen L & Garrett RA (2004) Shuffling of Sulfolobus genomes by autonomous and non‐autonomous mobile elements. Biochem Soc Trans 32: 179–183. [DOI] [PubMed] [Google Scholar]
- Buchner JM, Robertson AE, Poynter DJ, Denniston SS & Karls AC (2005) Piv site‐specific invertase requires a DEDD motif analogous to the catalytic center of the RuvC Holliday junction resolvases. J Bacteriol 187: 3431–3437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buisine N, Tang CM & Chalmers R (2002) Transposon‐like Correia elements: structure, distribution and genetic exchange between pathogenic Neisseria sp. FEBS Lett 522: 52–58. [DOI] [PubMed] [Google Scholar]
- Burrus V & Waldor MK (2004) Shaping bacterial genomes with integrative and conjugative elements. Res Microbiol 155: 376–386. [DOI] [PubMed] [Google Scholar]
- Burrus V, Pavlovic G, Decaris B & Guedon G (2002) Conjugative transposons: the tip of the iceberg. Mol Microbiol 46: 601–610. [DOI] [PubMed] [Google Scholar]
- Cerveau N, Leclercq S, Leroy E, Bouchon D & Cordaux R (2011) Short‐ and long‐term evolutionary dynamics of bacterial insertion sequences: insights from Wolbachia endosymbionts . Genome Biol Evol 3: 1175–1186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaconas G & Harshey RM (2002) Transposition of phage Mu DNA Mobile DNA, Vol. II (Craig NL, Craigie R, Gellert M. & Lambowitz A, eds), pp. 384–402. ASM press, Washington, DC. [Google Scholar]
- Chandler M & Mahillon J (2002) Insertion sequences revisited Mobile DNA, Vol. II (Craig NL, Craigie R, Gellert M & Lambowitz A, eds), pp. 305–366. ASM press, Washington, DC. [Google Scholar]
- Chandler M, de la Cruz F, Dyda F, Hickman AB, Moncalian G & Ton‐Hoang B (2013) Breaking and joining single‐stranded DNA: the HUH endonuclease superfamily. Nat Rev Microbiol 11: 525–538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi S, Ohta S & Ohtsubo E (2003) A novel IS element, IS621, of the IS110/IS492 family transposes to a specific site in repetitive extragenic palindromic sequences in Escherichia coli . J Bacteriol 185: 4891–4900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clamp M, Cuff J, Searle SM & Barton GJ (2004) The Jalview Java alignment editor. Bioinformatics 20: 426–427. [DOI] [PubMed] [Google Scholar]
- Clement J‐M, Wilde C, Bachellier S, Lambert P & Hofnung M (1999) IS1397 is active for transposition into the chromosome of Escherichia coli K‐12 and inserts specifically into palindromic units of bacterial interspersed mosaic elements. J Bacteriol 181: 6929–6936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cordaux R, Udit S, Batzer MA & Feschotte C (2006) Birth of a chimeric primate gene by capture of the transposase gene from a mobile element. P Natl Acad Sci USA 103: 8101–8106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Correia FF, Inouye S & Inouye M (1988) A family of small repeated elements with some transposon‐like properties in the genome of Neisseria gonorrhoeae . J Biol Chem 263: 12194–12198. [PubMed] [Google Scholar]
- Craig NL (2002) Tn7 Mobile DNA, Vol. II (Craig NL, Craigie R, Gellert M. & Lambowitz A, eds), pp. 423–456. ASM press, Washington, DC. [Google Scholar]
- Craig NL, Craigie R, Gellert M & Lambowitz A (2002) Mobile DNA II. American Society of Microbiology, Washington, DC. [Google Scholar]
- Curcio MJ & Derbyshire KM (2003) The outs and ins of transposition: from Mu to Kangaroo. Nat Rev Mol Cell Biol 4: 865–877. [DOI] [PubMed] [Google Scholar]
- Dada JO & Mendes P (2011) Multi‐scale modelling and simulation in systems biology. Integr Biol (Camb) 3: 86–96. [DOI] [PubMed] [Google Scholar]
- De Palmenaer D, Vermeiren C & Mahillon J (2004) IS231‐MIC231 elements from Bacillus cereus sensu lato are modular. Mol Microbiol 53: 457–467. [DOI] [PubMed] [Google Scholar]
- De Palmenaer D, Siguier P & Mahillon J (2008) IS4 family goes genomic. BMC Evol Biol 8: 18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- del Pilar Garcillan‐Barcia M, Bernales I, Mendiola MV & de la Cruz F (2001) Single‐stranded DNA intermediates in IS91 rolling‐circle transposition. Mol Microbiol 39: 494–501. [DOI] [PubMed] [Google Scholar]
- Depardieu F, Podglajen I, Leclercq R, Collatz E & Courvalin P (2007) Modes and modulations of antibiotic resistance gene expression. Clin Microbiol Rev 20: 79–114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dobrindt U, Hochhut B, Hentschel U & Hacker J (2004) Genomic islands in pathogenic and environmental microorganisms. Nat Rev Microbiol 2: 414–424. [DOI] [PubMed] [Google Scholar]
- Duval‐Valentin G & Chandler M (2011) Cotranslational control of DNA transposition: a window of opportunity. Mol Cell 44: 989–996. [DOI] [PubMed] [Google Scholar]
- Dyall‐Smith ML, Pfeiffer F, Klee K et al (2011) Haloquadratum walsbyi: limited diversity in a global pond. PLoS One 6: e20968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dyda F, Chandler M & Hickman AB (2012) The emerging diversity of transpososome architectures. Q Rev Biophys 45: 493–521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Enright AJ, Van Dongen S & Ouzounis CA (2002) An efficient algorithm for large‐scale detection of protein families. Nucleic Acids Res 30: 1575–1584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Escoubas JM, Prere MF, Fayet O, Salvignol I, Galas D, Zerbib D & Chandler M (1991) Translational control of transposition activity of the bacterial insertion sequence IS1. EMBO J 10: 705–712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Espeli O & Boccard F (1997) In vivo cleavage of Escherichia coli BIME‐2 repeats by DNA gyrase: genetic characterization of the target and identification of the cut site. Mol Microbiol 26: 767–777. [DOI] [PubMed] [Google Scholar]
- Espeli O, Moulin L & Boccard F (2001) Transcription attenuation associated with bacterial repetitive extragenic BIME elements. J Mol Biol 314: 375–386. [DOI] [PubMed] [Google Scholar]
- Fayet O, Ramond P, Polard P, Prere MF & Chandler M (1990) Functional similarities between retroviruses and the IS3 family of bacterial insertion sequences? Mol Microbiol 4: 1771–1777. [DOI] [PubMed] [Google Scholar]
- Feng X & Colloms SD (2007) In vitro transposition of ISY100, a bacterial insertion sequence belonging to the Tc1/mariner family. Mol Microbiol 65: 1432–1443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feschotte C, Zhang X & Wessler S (2002) Miniature inverted repeat transposable elements and their relationship to established DNA transposons Mobile DNA, Vol. II (Craig NL, Craigie R, Gellert M. & Lambowitz A, eds), pp. 1147–1158. ASM press, Washington, DC. [Google Scholar]
- Filee J, Siguier P & Chandler M (2007) Insertion sequence diversity in archaea. Microbiol Mol Biol Rev 71: 121–157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Filée J, Siguier P & Chandler M (2007) I am what I eat and I eat what I am: acquisition of bacterial genes by giant viruses. Trends Genet 23: 10–15. [DOI] [PubMed] [Google Scholar]
- Fulks KA, Marrs CF, Stevens SP & Green MR (1990) Sequence analysis of the inversion region containing the pilin genes of Moraxella bovis . J Bacteriol 172: 310–316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcillan‐Barcia MP & de la Cruz F (2002) Distribution of IS91 family insertion sequences in bacterial genomes: evolutionary implications. FEMS Microbiol Ecol 42: 303–313. [DOI] [PubMed] [Google Scholar]
- Garcillan‐Barcia MP, Bernales I, Mendiola MV & De la Cruz F (2002) IS91 rolling circle transposition Mobile DNA, Vol. II (Craig NL, Craigie R, Gellert M. & Lambowitz A, eds), pp. 891–904. ASM press, Washington, DC. [Google Scholar]
- Gesteland RF, Weiss RB & Atkins JF (1992) Recoding: reprogrammed genetic decoding. Science 257: 1640–1641. [DOI] [PubMed] [Google Scholar]
- Gil R, Belda E, Gosalbes MJ et al (2008) Massive presence of insertion sequences in the genome of SOPE, the primary endosymbiont of the rice weevil Sitophilus oryzae . Int Microbiol 11: 41–48. [PubMed] [Google Scholar]
- Gilbert C & Cordaux R (2013) Horizontal transfer and evolution of prokaryote transposable elements in eukaryotes. Genome Biol Evol 5: 822–832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilson E, Perrin D & Hofnung M (1990) DNA polymerase I and a protein complex bind specifically to E. coli palindromic unit highly repetitive DNA: implications for bacterial chromosome organization. Nucleic Acids Res 18: 3941–3952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gladyshev EA & Arkhipova IR (2009) A single‐copy IS5‐like transposon in the genome of a bdelloid rotifer. Mol Biol Evol 26: 1921–1929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glansdorff N, Charlier D & Zafarullah M (1981) Activation of gene expression by IS2 and IS3. Cold Spring Harb Symp Quant Biol 45(Pt 1): 153–156. [DOI] [PubMed] [Google Scholar]
- Grindley NDF (2002) The movement of Tn3‐like elements: transposition and cointegrate resolution Mobile DNA II, (Craig NL, Craigie R, Gellert M. & Lambowitz A, eds), pp. 230–271. ASM press, Washington, DC. [Google Scholar]
- Grindley ND, Whiteson KL & Rice PA (2006) Mechanisms of site‐specific recombination. Annu Rev Biochem 75: 567–605. [DOI] [PubMed] [Google Scholar]
- Gueguen E, Rousseau P, Duval‐Valentin G & Chandler M (2006) Truncated forms of IS911 transposase downregulate transposition. Mol Microbiol 62: 1102–1116. [DOI] [PubMed] [Google Scholar]
- Guerillot R, Da Cunha V, Sauvage E, Bouchier C & Glaser P (2013) Modular evolution of TnGBSs, a new family of integrative and conjugative elements associating insertion sequence transposition, plasmid replication, and conjugation for their spreading. J Bacteriol 195: 1979–1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guerillot R, Siguier P, Gourbeyre E, Chandler M & Glaser P (2014) The diversity of prokaryotic DDE transposases of the mutator superfamily, insertion specificity, and association with conjugation machineries. Genome Biol Evol 6: 260–272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guynet C, Hickman AB, Barabas O, Dyda F, Chandler M & Ton‐Hoang B (2008) In vitro reconstitution of a single‐stranded transposition mechanism of IS608. Mol Cell 29: 302–312. [DOI] [PubMed] [Google Scholar]
- Guynet C, Achard A, Hoang BT, Barabas O, Hickman AB, Dyda F & Chandler M (2009) Resetting the site: redirecting integration of an insertion sequence in a predictable way. Mol Cell 34: 612–619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hallet B, Rezsohazy R & Delcour J (1991) IS231A from Bacillus thuringiensis is functional in Escherichia coli: transposition and insertion specificity. J Bacteriol 173: 4526–4529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han CG, Shiga Y, Tobe T, Sasakawa C & Ohtsubo E (2001) Structural and functional characterization of IS679 and IS66‐family elements. J Bacteriol 183: 4296–4304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haniford DB (2006) Transpososome dynamics and regulation in Tn10 transposition. Crit Rev Biochem Mol Biol 41: 407–424. [DOI] [PubMed] [Google Scholar]
- Haren L, Polard P, Ton‐Hoang B & Chandler M (1998) Multiple oligomerisation domains in the IS911 transposase: a leucine zipper motif is essential for activity. J Mol Biol 283: 29–41. [DOI] [PubMed] [Google Scholar]
- Haren L, Ton‐Hoang B & Chandler M (1999) Integrating DNA: transposases and retroviral integrases. Annu Rev Microbiol 53: 245–281. [DOI] [PubMed] [Google Scholar]
- Haren L, Normand C, Polard P, Alazard R & Chandler M (2000) IS911 transposition is regulated by protein‐protein interactions via a leucine zipper motif. J Mol Biol 296: 757–768. [DOI] [PubMed] [Google Scholar]
- Hasselmayer O, Nitsche C, Braun V & von Eichel‐Streiber C (2004a) The IStron CdISt1 of Clostridium difficile: molecular symbiosis of a group I intron and an insertion element. Anaerobe 10: 85–92. [DOI] [PubMed] [Google Scholar]
- Hasselmayer O, Braun V, Nitsche C, Moos M, Rupnik M & von Eichel‐Streiber C (2004b) Clostridium difficile IStron CdISt1: discovery of a variant encoding two complete transposase‐like proteins. J Bacteriol 186: 2508–2510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He S, Hickman AB, Dyda F, Johnson NP, Chandler M & Ton‐Hoang B (2011) Reconstitution of a functional IS608 single‐strand transpososome: role of non‐canonical base pairing. Nucleic Acids Res 39: 8503–8512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He S, Guynet C, Siguier P, Hickman AB, Dyda F, Chandler M & Ton‐Hoang B (2013) IS200/IS605 family single‐strand transposition: mechanism of IS608 strand transfer. Nucleic Acids Res 41: 3302–3313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hickman AB, Chandler M & Dyda F (2010a) Integrating prokaryotes and eukaryotes: DNA transposases in light of structure. Crit Rev Biochem Mol Biol 45: 50–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hickman AB, James JA, Barabas O et al (2010b) DNA recognition and the precleavage state during single‐stranded DNA transposition in D. radiodurans . EMBO J 29: 3840–3852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Higgins CF, Ames GF, Barnes WM, Clement JM & Hofnung M (1982) A novel intercistronic regulatory element of prokaryotic operons. Nature 298: 760–762. [DOI] [PubMed] [Google Scholar]
- Hsia R, Ohayon H, Gounon P, Dautry‐Varsat A & Bavoil PM (2000) Phage infection of the obligate intracellular bacterium, Chlamydia psittaci strain guinea pig inclusion conjunctivitis. Microbes Infect 2: 761–772. [DOI] [PubMed] [Google Scholar]
- Hu WY & Derbyshire KM (1998) Target choice and orientation preference of the insertion sequence IS903. J Bacteriol 180: 3039–3048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hua‐Van Al & Capy P (2008) Analysis of the DDE motif in the Mutator superfamily. J Mol Evol 67: 670–681. [DOI] [PubMed] [Google Scholar]
- Ibba M & Soll D (2004) Aminoacyl‐tRNAs: setting the limits of the genetic code. Genes Dev 18: 731–738. [DOI] [PubMed] [Google Scholar]
- Ilyina TV & Koonin EV (1992) Conserved sequence motifs in the initiator proteins for rolling circle DNA replication encoded by diverse replicons from eubacteria, eucaryotes and archaebacteria. Nucleic Acids Res 20: 3279–3285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Islam SM, Hua Y, Ohba H, Satoh K, Kikuchi M, Yanagisawa T & Narumi I (2003) Characterization and distribution of IS8301 in the radioresistant bacterium Deinococcus radiodurans . Genes Genet Syst 78: 319–327. [DOI] [PubMed] [Google Scholar]
- Jellen‐Ritter AS & Kern WV (2001) Enhanced expression of the multidrug efflux pumps AcrAB and AcrEF associated with insertion element transposition in Escherichia coli mutants Selected with a fluoroquinolone. Antimicrob Agents Chemother 45: 1467–1472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kapitonov VV & Jurka J (2007) Helitrons on a roll: eukaryotic rolling‐circle transposons. Trends Genet 23: 521–529. [DOI] [PubMed] [Google Scholar]
- Kersulyte D, Akopyants NS, Clifton SW, Roe BA & Berg DE (1998) Novel sequence organization and insertion specificity of IS605 and IS606: chimaeric transposable elements of Helicobacter pylori . Gene 223: 175–186. [DOI] [PubMed] [Google Scholar]
- Kersulyte D, Mukhopadhyay AK, Shirai M, Nakazawa T & Berg DE (2000) Functional organization and insertion specificity of IS607, a chimeric element of Helicobacter pylori . J Bacteriol 182: 5300–5308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kersulyte D, Velapatino B, Dailide G et al (2002) Transposable element ISHp608 of Helicobacter pylori: nonrandom geographic distribution, functional organization, and insertion specificity. J Bacteriol 184: 992–1002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khemici V & Carpousis AJ (2004) The RNA degradosome and poly(A) polymerase of Escherichia coli are required in vivo for the degradation of small mRNA decay intermediates containing REP‐stabilizers. Mol Microbiol 51: 777–790. [DOI] [PubMed] [Google Scholar]
- Kieny M‐P (2012) The evolving threat of antimicrobial resistance: Options for action. World Health Organization.
- Kleiner M, Young JC, Shah M, VerBerkmoes NC & Dubilier N (2013) Metaproteomics reveals abundant transposase expression in mutualistic endosymbionts. MBio 4: e00223–00213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kobayashi K, Tsukagoshi N & Aono R (2001) Suppression of hypersensitivity of Escherichia coli acrB mutant to organic solvents by integrational activation of the acrEF operon with the IS1 or IS2 element. J Bacteriol 183: 2646–2653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lam S & Roth JR (1983) IS200: a Salmonella‐specific insertion sequence. Cell 34: 951–960. [DOI] [PubMed] [Google Scholar]
- Lawrence JG, Hendrix RW & Casjens S (2001) Where are the pseudogenes in bacterial genomes? Trends Microbiol 9: 535–540. [DOI] [PubMed] [Google Scholar]
- Lederberg EM (1981) Plasmid reference center registry of transposon (Tn) allocations through July 1981. Gene 16: 59–61. [DOI] [PubMed] [Google Scholar]
- Lee CA, Babic A & Grossman AD (2010) Autonomous plasmid‐like replication of a conjugative transposon. Mol Microbiol 75: 268–279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liebert CA, Hall RM & Summers AO (1999) Transposon Tn21, flagship of the floating genome. Microbiol Mol Biol Rev 63: 507–522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Z, Reches M, Groisman I & Engelberg‐Kulka H (1998) The nature of the minimal ‘selenocysteine insertion sequence’ (SECIS) in Escherichia coli . Nucleic Acids Res 26: 896–902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu D, Bischerour J, Siddique A, Buisine N, Bigot Y & Chalmers R (2007) The human SETMAR protein preserves most of the activities of the ancestral Hsmar1 transposase. Mol Cell Biol 27: 1125–1132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Machida Y, Machida C & Ohtsubo E (1982) A novel type of transposon generated by insertion element IS102 present in a pSC101 derivative. Cell 30: 29–36. [DOI] [PubMed] [Google Scholar]
- Mahillon J & Chandler M (1998) Insertion sequences. Microbiol Mol Biol Rev 62: 725–774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazel D (2006) Integrons: agents of bacterial evolution. Nat Rev Microbiol 4: 608–620. [DOI] [PubMed] [Google Scholar]
- McKenna M (2013) The last resort. Nature 499: 394–396. [DOI] [PubMed] [Google Scholar]
- Medini D, Donati C, Tettelin H, Masignani V & Rappuoli R (2005) The microbial pan‐genome. Curr Opin Genet Dev 15: 589–594. [DOI] [PubMed] [Google Scholar]
- Mendiola MV & de la Cruz F (1992) IS91 transposase is related to the rolling‐circle‐type replication proteins of the pUB110 family of plasmids. Nucleic Acids Res 20: 3521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Messing SA, Ton‐Hoang B, Hickman AB et al (2012) The processing of repetitive extragenic palindromes: the structure of a repetitive extragenic palindrome bound to its associated nuclease. Nucleic Acids Res 40: 9964–9979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mira A, Ochman H & Moran NA (2001) Deletional bias and the evolution of bacterial genomes. Trends Genet 17: 589–596. [DOI] [PubMed] [Google Scholar]
- Mizuuchi K (1992a) Transpositional recombination: mechanistic insights from studies of mu and other elements. Annu Rev Biochem 61: 1011–1051. [DOI] [PubMed] [Google Scholar]
- Mizuuchi K (1992b) Polynucleotidyl transfer reactions in transpositional DNA recombination. J Biol Chem 267: 21273–21276. [PubMed] [Google Scholar]
- Mole B (2013) Farming up trouble. Nature 499: 398–400. [DOI] [PubMed] [Google Scholar]
- Montano SP & Rice PA (2011) Moving DNA around: DNA transposition and retroviral integration. Curr Opin Struct Biol 21: 370–378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montano SP, Pigli YZ & Rice PA (2012) The mu transpososome structure sheds light on DDE recombinase evolution. Nature 491: 413–417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moran NA (1996) Accelerated evolution and Muller's rachet in endosymbiotic bacteria. P Natl Acad Sci USA 93: 2873–2878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moran NA & Plague GR (2004) Genomic changes following host restriction in bacteria. Curr Opin Genet Dev 14: 627–633. [DOI] [PubMed] [Google Scholar]
- Moulin L, Rahmouni AR & Boccard F (2005) Topological insulators inhibit diffusion of transcription‐induced positive supercoils in the chromosome of Escherichia coli . Mol Microbiol 55: 601–610. [DOI] [PubMed] [Google Scholar]
- Moya A, Pereto J, Gil R & Latorre A (2008) Learning how to live together: genomic insights into prokaryote‐animal symbioses. Nat Rev Genet 9: 218–229. [DOI] [PubMed] [Google Scholar]
- Murphy DB & Pembroke JT (1995) Transfer of the IncJ plasmid R391 to recombination deficient Escherichia coli K12: evidence that R391 behaves as a conjugal transposon. FEMS Microbiol Lett 134: 153–158. [DOI] [PubMed] [Google Scholar]
- Naas T, Blot M, Fitch WM & Arber W (1994) Insertion sequence‐related genetic variation in resting Escherichia coli K‐12. Genetics 136: 721–730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naas T, Blot M, Fitch WM & Arber W (1995) Dynamics of IS‐related genetic rearrangements in resting Escherichia coli K‐12. Mol Biol Evol 12: 198–207. [DOI] [PubMed] [Google Scholar]
- Namy O, Rousset JP, Napthine S & Brierley I (2004) Reprogrammed genetic decoding in cellular gene expression. Mol Cell 13: 157–168. [DOI] [PubMed] [Google Scholar]
- Nunvar J, Huckova T & Licha I (2010) Identification and characterization of repetitive extragenic palindromes (REP)‐associated tyrosine transposases: implications for REP evolution and dynamics in bacterial genomes. BMC Genomics 11: 44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nunvar J, Licha I & Schneider B (2013) Evolution of REP diversity: a comparative study. BMC Genomics 14: 385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nyman K, Nakamura K, Ohtsubo H & Ohtsubo E (1981) Distribution of the insertion sequence IS1 in gram‐negative bacteria. Nature 289: 609–612. [DOI] [PubMed] [Google Scholar]
- Oggioni MR & Claverys JP (1999) Repeated extragenic sequences in prokaryotic genomes: a proposal for the origin and dynamics of the RUP element in Streptococcus pneumoniae . Microbiology 145(Pt 10): 2647–2653. [DOI] [PubMed] [Google Scholar]
- Ohta S, Yoshimura E & Ohtsubo E (2004) Involvement of two domains with helix‐turn‐helix and zinc finger motifs in the binding of IS1 transposase to terminal inverted repeats. Mol Microbiol 53: 193–202. [DOI] [PubMed] [Google Scholar]
- Ohtsubo H, Nyman K, Doroszkiewicz W & Ohtsubo E (1981) Multiple copies of iso‐insertion sequences of IS1 in Shigella dysenteriae chromosome. Nature 292: 640–643. [DOI] [PubMed] [Google Scholar]
- Olliver A, Valle M, Chaslus‐Dancla E & Cloeckaert A (2005) Overexpression of the multidrug efflux operon acrEF by insertional activation with IS1 or IS10 elements in Salmonella enterica serovar typhimurium DT204 acrB mutants selected with fluoroquinolones. Antimicrob Agents Chemother 49: 289–301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papke RT, Ramsing NB, Bateson MM & Ward DM (2003) Geographical isolation in hot spring cyanobacteria. Environ Microbiol 5: 650–659. [DOI] [PubMed] [Google Scholar]
- Parkhill J & Thomson N (2003) Evolutionary strategies of human pathogens. Cold Spring Harb Symp Quant Biol 68: 151–158. [DOI] [PubMed] [Google Scholar]
- Parkhill J, Sebaihia M, Preston A et al (2003) Comparative analysis of the genome sequences of Bordetella pertussis, Bordetella parapertussis and Bordetella bronchiseptica . Nat Genet 35: 32–40. [DOI] [PubMed] [Google Scholar]
- Parks AR, Li Z, Shi Q, Owens RM, Jin MM & Peters JE (2009) Transposition into replicating DNA occurs through interaction with the processivity factor. Cell 138: 685–695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Partridge SR & Hall RM (2003) The IS1111 family members IS4321 and IS5075 have subterminal inverted repeats and target the terminal inverted repeats of Tn21 family transposons. J Bacteriol 185: 6371–6384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pasternak C, Ton‐Hoang B, Coste G, Bailone A, Chandler M & Sommer S (2010) Irradiation‐induced Deinococcus radiodurans genome fragmentation triggers transposition of a single resident insertion sequence. PLoS Genet 6: e1000799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pasternak C, Dulermo R, Ton‐Hoang B et al (2013) ISDra2 transposition in Deinococcus radiodurans is downregulated by TnpB. Mol Microbiol 88: 443–455. [DOI] [PubMed] [Google Scholar]
- Pavlovic G, Burrus V, Gintz B, Decaris B & Guedon G (2004) Evolution of genomic islands by deletion and tandem accretion by site‐specific recombination: ICESt1‐related elements from Streptococcus thermophilus . Microbiology 150: 759–774. [DOI] [PubMed] [Google Scholar]
- Pembroke JT & Murphy DB (2000) Isolation and analysis of a circular form of the IncJ conjugative transposon‐like elements, R391 and R997: implications for IncJ incompatibility. FEMS Microbiol Lett 187: 133–138. [DOI] [PubMed] [Google Scholar]
- Plague GR, Dunbar HE, Tran PL & Moran NA (2008) Extensive proliferation of transposable elements in heritable bacterial symbionts. J Bacteriol 190: 777–779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polard P, Prere MF, Chandler M & Fayet O (1991) Programmed translational frameshifting and initiation at an AUU codon in gene expression of bacterial insertion sequence IS911. J Mol Biol 222: 465–477. [DOI] [PubMed] [Google Scholar]
- Polard P, Seroude L, Fayet O, Prere MF & Chandler M (1994) One‐ended insertion of IS911. J Bacteriol 176: 1192–1196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Post V & Hall RM (2009) Insertion sequences in the IS1111 family that target the attC recombination sites of integron‐associated gene cassettes. FEMS Microbiol Lett 290: 182–187. [DOI] [PubMed] [Google Scholar]
- Prentki P, Teter B, Chandler M & Galas DJ (1986) Functional promoters created by the insertion of transposable element IS1. J Mol Biol 191: 383–393. [DOI] [PubMed] [Google Scholar]
- Preston A, Parkhill J & Maskell DJ (2004) The Bordetellae: lessons from genomics. Nat Rev Micro 2: 379–390. [DOI] [PubMed] [Google Scholar]
- Qi X, Daily K, Nguyen K et al (2012) Retrotransposon profiling of RNA polymerase III initiation sites. Genome Res 22: 681–692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Read TD, Fraser CM, Hsia RC & Bavoil PM (2000) Comparative analysis of Chlamydia bacteriophages reveals variation localized to a putative receptor binding domain. Microb Comp Genomics 5: 223–231. [DOI] [PubMed] [Google Scholar]
- Reif HJ & Saedler H (1974) IS1 is involved in deletion formation in the gal region of E.coli K12. Mol Gen Genet 137: 17–28. [DOI] [PubMed] [Google Scholar]
- Rezsohazy R, Hallet B, Delcour J & Mahillon J (1993) The IS4 family of insertion sequences: evidence for a conserved transposase motif. Mol Microbiol 9: 1283–1295. [DOI] [PubMed] [Google Scholar]
- Rice P, Craigie R & Davies DR (1996) Retroviral integrases and their cousins. Curr Opin Struct Biol 6: 76–83. [DOI] [PubMed] [Google Scholar]
- Richardson JM, Colloms SD, Finnegan DJ & Walkinshaw MD (2009) Molecular architecture of the Mos1 paired‐end complex: the structural basis of DNA transposition in a eukaryote. Cell 138: 1096–1108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ricker N, Qian H & Fulthorpe RR (2013) Phylogeny and organization of recombinase in trio (RIT) elements. Plasmid 70: 226–239. [DOI] [PubMed] [Google Scholar]
- Rio DC (1991) Regulation of Drosophila P element transposition. Trends Genet 7: 282–287. [DOI] [PubMed] [Google Scholar]
- Robinson DG, Lee MC & Marx CJ (2012) OASIS: an automated program for global investigation of bacterial and archaeal insertion sequences. Nucleic Acids Res 40: e174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rolland T, Neuveglise C, Sacerdot C & Dujon B (2009) Insertion of horizontally transferred genes within conserved syntenic regions of yeast genomes. PLoS One 4: e6515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ronning DR, Guynet C, Ton‐Hoang B, Perez ZN, Ghirlando R, Chandler M & Dyda F (2005) Active site sharing and subterminal hairpin recognition in a new class of DNA transposases. Mol Cell 20: 143–154. [DOI] [PubMed] [Google Scholar]
- Rousseau P, Gueguen E, Duval‐Valentin G & Chandler M (2004) The helix‐turn‐helix motif of bacterial insertion sequence IS911 transposase is required for DNA binding. Nucleic Acids Res 32: 1335–1344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rousseau P, Loot C, Guynet C, Ah‐Seng Y, Ton‐Hoang B & Chandler M (2007) Control of IS911 target selection: how OrfA may ensure IS dispersion. Mol Microbiol 63: 1701–1709. [DOI] [PubMed] [Google Scholar]
- Rozsa FW, Meyer TF & Fussenegger M (1997) Inversion of Moraxella lacunata type 4 pilin gene sequences by a Neisseria gonorrhoeae site‐specific recombinase. J Bacteriol 179: 2382–2388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salvatore P, Pagliarulo C, Colicchio R et al (2001) Identification, characterization, and variable expression of a naturally occurring inhibitor protein of IS1106 transposase in clinical isolates of Neisseria meningitidis . Infect Immun 69: 7425–7436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmitz‐Esser S, Penz T, Spang A & Horn M (2011) A bacterial genome in transition–an exceptional enrichment of IS elements but lack of evidence for recent transposition in the symbiont Amoebophilus asiaticus . BMC Evol Biol 11: 270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sekine Y & Ohtsubo E (1989) Frameshifting is required for production of the transposase encoded by insertion sequence 1. P Natl Acad Sci USA 86: 4609–4613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sekine Y, Eisaki N & Ohtsubo E (1994) Translational control in production of transposase and in transposition of insertion sequence IS3. J Mol Biol 235: 1406–1420. [DOI] [PubMed] [Google Scholar]
- Shaheen M, Williamson E, Nickoloff J, Lee SH & Hromas R (2010) Metnase/SETMAR: a domesticated primate transposase that enhances DNA repair, replication, and decatenation. Genetica 138: 559–566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shapiro JA (1979) Molecular model for the transposition and replication of bacteriophage Mu and other transposable elements. P Natl Acad Sci USA 76: 1933–1937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma V, Firth AE, Antonov I, Fayet O, Atkins JF, Borodovsky M & Baranov PV (2011) A pilot study of bacterial genes with disrupted ORFs reveals a surprising profusion of protein sequence recoding mediated by ribosomal frameshifting and transcriptional realignment. Mol Biol Evol 28: 3195–3211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siguier P, Perochon J, Lestrade L, Mahillon J & Chandler M (2006) ISfinder: the reference centre for bacterial insertion sequences. Nucleic Acids Res 34: D32–D36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siguier P, Gagnevin L & Chandler M (2009) The new IS1595 family, its relation to IS1 and the frontier between insertion sequences and transposons. Res Microbiol 160: 232–241. [DOI] [PubMed] [Google Scholar]
- Silva FJ, Latorre A & Moya A (2003) Why are the genomes of endosymbiotic bacteria so stable? Trends Genet 19: 176–180. [DOI] [PubMed] [Google Scholar]
- Sinzelle L, Izsvak Z & Ivics Z (2009) Molecular domestication of transposable elements: from detrimental parasites to useful host genes. Cell Mol Life Sci 66: 1073–1093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skaar EP, Lecuyer B, Lenich AG, Lazio MP, Perkins‐Balding D, Seifert HS & Karls AC (2005) Analysis of the Piv recombinase‐related gene family of Neisseria gonorrhoeae . J Bacteriol 187: 1276–1286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smyth DS & Robinson DA (2009) Integrative and sequence characteristics of a novel genetic element, ICE6013, in Staphylococcus aureus . J Bacteriol 191: 5964–5975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soki J, Eitel Z, Urban E & Nagy E (2013) Molecular analysis of the carbapenem and metronidazole resistance mechanisms of Bacteroides strains reported in a Europe‐wide antibiotic resistance survey. Int J Antimicrob Agents 41: 122–125. [DOI] [PubMed] [Google Scholar]
- Söll D (2007) The genetic code revisited — four decades after Francis Crick. Nucleic Acids Symp Ser 51: 13–14. [DOI] [PubMed] [Google Scholar]
- Spradling AC, Bellen HJ & Hoskins RA (2011) Drosophila P elements preferentially transpose to replication origins. P Natl Acad Sci USA 108: 15948–15953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stalder R, Caspers P, Olasz F & Arber W (1990) The N‐terminal domain of the insertion sequence 30 transposase interacts specifically with the terminal inverted repeats of the element. J Biol Chem 265: 3757–3762. [PubMed] [Google Scholar]
- Szeverenyi I, Hodel A, Arber W & Olasz F (1996) Vector for IS element entrapment and functional characterization based on turning on expression of distal promoterless genes. Gene 174: 103–110. [DOI] [PubMed] [Google Scholar]
- Tettelin H, Riley D, Cattuto C & Medini D (2008) Comparative genomics: the bacterial pan‐genome. Curr Opin Microbiol 11: 472–477. [DOI] [PubMed] [Google Scholar]
- Tetu SG & Holmes AJ (2008) A family of insertion sequences that impacts integrons by specific targeting of gene cassette recombination sites, the IS1111‐attC group. J Bacteriol 190: 4959–4970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson JD, Higgins DG & Gibson TJ (1994) clustal w: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position‐specific gap penalties and weight matrix choice. Nucleic Acids Res 22: 4673–4680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tobes R & Pareja E (2006) Bacterial repetitive extragenic palindromic sequences are DNA targets for Insertion Sequence elements. BMC Genomics 7: 62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tobes R & Ramos J‐L (2005) REP code: defining bacterial identity in extragenic space. Environ Microbiol 7: 225–228. [DOI] [PubMed] [Google Scholar]
- Tobiason DM, Lenich AG & Glasgow AC (1999) Multiple DNA binding activities of the novel site‐specific recombinase, Piv, from Moraxella lacunata . J Biol Chem 274: 9698–9706. [DOI] [PubMed] [Google Scholar]
- Tobiason DM, Buchner JM, Thiel WH, Gernert KM & Karls AC (2001) Conserved amino acid motifs from the novel Piv/MooV family of transposases and site‐specific recombinases are required for catalysis of DNA inversion by Piv. Mol Microbiol 39: 641–651. [DOI] [PubMed] [Google Scholar]
- Toleman MA & Walsh TR (2011) Combinatorial events of insertion sequences and ICE in Gram‐negative bacteria. FEMS Microbiol Rev 35: 912–935. [DOI] [PubMed] [Google Scholar]
- Toleman MA, Bennett PM & Walsh TR (2006) ISCR elements: novel gene‐capturing systems of the 21st century? Microbiol Mol Biol Rev 70: 296–316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ton‐Hoang B, Betermier M, Polard P & Chandler M (1997) Assembly of a strong promoter following IS911 circularization and the role of circles in transposition. EMBO J 16: 3357–3371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ton‐Hoang B, Polard P & Chandler M (1998) Efficient transposition of IS911 circles in vitro . EMBO J 17: 1169–1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ton‐Hoang B, Turlan C & Chandler M (2004) Functional domains of the IS1 transposase: analysis in vivo and in vitro . Mol Microbiol 53: 1529–1543. [DOI] [PubMed] [Google Scholar]
- Ton‐Hoang B, Guynet C, Ronning DR, Cointin‐Marty B, Dyda F & Chandler M (2005) Transposition of ISHp608, member of an unusual family of bacterial insertion sequences. EMBO J 24: 3325–3338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ton‐Hoang B, Pasternak C, Siguier P et al (2010) Single‐stranded DNA transposition is coupled to host replication. Cell 142: 398–408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ton‐Hoang B, Siguier P, Quentin Y, Onillon S, Marty B, Fichant G & Chandler M (2012) Structuring the bacterial genome: Y1‐transposases associated with REP‐BIME sequences. Nucleic Acids Res 40: 3596–3609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Touchon M & Rocha EP (2007) Causes of insertion sequences abundance in prokaryotic genomes. Mol Biol Evol 24: 969–981. [DOI] [PubMed] [Google Scholar]
- Turlan C & Chandler M (1995) IS1‐mediated intramolecular rearrangements: formation of excised transposon circles and replicative deletions. EMBO J 14: 5410–5421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turlan C & Chandler M (2000) Playing second fiddle: second‐strand processing and liberation of transposable elements from donor DNA. Trends Microbiol 8: 268–274. [DOI] [PubMed] [Google Scholar]
- Van Dongen S (2000) A cluster algorithm for graphs. Technical Report INS‐R0010, National Research Institute for Mathematics and Computer Science in the Netherlands, Amsterdam.
- Van Duyne GD & Rutherford K (2013) Large serine recombinase domain structure and attachment site binding. Crit Rev Biochem Mol Biol 48: 476–491. [DOI] [PubMed] [Google Scholar]
- Vanhooff V, Galloy C, Agaisse H, Lereclus D, Revet B & Hallet B (2006) Self‐control in DNA site‐specific recombination mediated by the tyrosine recombinase TnpI. Mol Microbiol 60: 617–629. [DOI] [PubMed] [Google Scholar]
- Varani A, Siguier P, Gourbeyre E, Charneau V, & Chandler M (2011) ISsaga is an ensemble of web‐based methods for high throughput identification and semi‐automatic annotation of insertion sequences in prokaryotic genomes. Genome Biol 12: R30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vos JC, van Luenen HG & Plasterk RH (1993) Characterization of the Caenorhabditis elegans Tc1 transposase in vivo and in vitro . Genes Dev 7: 1244–1253. [DOI] [PubMed] [Google Scholar]
- Wagner A, Lewis C & Bichsel M (2007) A survey of bacterial insertion sequences using IScan. Nucleic Acids Res 35: 5284–5293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilde C, Escartin F, Kokeguchi S, Latour‐Lambert P, Lectard A & Clement JM (2003) Transposases are responsible for the target specificity of IS1397 and ISKpn1 for two different types of palindromic units (PUs). Nucleic Acids Res 31: 4345–4353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolter DJ, Smith‐Moland E, Goering RV, Hanson ND & Lister PD (2004) Multidrug resistance associated with mexXY expression in clinical isolates of Pseudomonas aeruginosa from a Texas hospital. Diagn Microbiol Infect Dis 50: 43–50. [DOI] [PubMed] [Google Scholar]
- Yano H, Genka H, Ohtsubo Y, Nagata Y, Top EM & Tsuda M (2013) Cointegrate‐resolution of toluene‐catabolic transposon Tn4651: determination of crossover site and the segment required for full resolution activity. Plasmid 69: 24–35. [DOI] [PubMed] [Google Scholar]
- Zabala JC, de la Cruz F & Ortiz JM (1982) Several copies of the same insertion sequence are present in alpha‐hemolytic plasmids belonging to four different incompatibility groups. J Bacteriol 151: 472–476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zahradka K, Slade D, Bailone A et al (2006) Reassembly of shattered chromosomes in Deinococcus radiodurans . Nature 443: 569–573. [DOI] [PubMed] [Google Scholar]
- Zerbib D, Jakowec M, Prentki P, Galas DJ & Chandler M (1987) Expression of proteins essential for IS1 transposition: specific binding of InsA to the ends of IS1. EMBO J 6: 3163–3169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zerbib D, Polard P, Escoubas JM, Galas D & Chandler M (1990) The regulatory role of the IS1‐encoded InsA protein in transposition. Mol Microbiol 4: 471–477. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. ISs in genomes.
Table S2. Gene activation by IS insertion.