

Abstract
There are substantial sex differences in renal structure and ammonia metabolism that correlate with differences in expression of proteins involved in ammonia generation and transport. This study determined the role of testis-derived testosterone in these differences. We studied 4-mo-old male C57BL/6 mice 4 and 8 wk after either bilateral orchiectomy (ORCH) or sham-operated control surgery and determined the effect of testosterone replacement to reverse the effects of ORCH. Finally, we determined the cellular expression of androgen receptor (AR), testosterone’s canonical target receptor. ORCH decreased kidney and proximal tubule size, and testosterone replacement reversed this effect. ORCH increased ammonia excretion in a testosterone-dependent fashion; this occurred despite similar food intake, which is the primary component of endogenous acid production. ORCH increased expression of both phosphoenolpyruvate, a major ammonia-generating protein, and Na+-K+-2Cl− cotransporter, which mediates thick ascending limb ammonia reabsorption; these changes were reversed with testosterone replacement. Orchiectomy also decreased expression of Na+/H+ exchanger isoform 3, which mediates proximal tubule ammonia secretion, in a testosterone-dependent pattern. Finally, ARs are expressed throughout the proximal tubule in both the male and female kidney. Testosterone, possibly acting through ARs, has dramatic effects on kidney and proximal tubule size and decreases ammonia excretion through its effects on several key proteins involved in ammonia metabolism.
Keywords: acid-base, ammonia, androgen receptor, sex differences, testosterone
INTRODUCTION
Sex differences are present in many aspects of mammalian biology and involve essentially all physiological processes (23a, 24, 47). These differences have been identified in the occurrence and development of various diseases such as hypertension, chronic kidney disease, and acute or chronic renal ischemia (11, 25, 43, 47, 49, 51). Abnormal acid-base homeostasis causes many disorders, including faster progression of chronic kidney disease, leading to increased progression to end-stage renal disease, decreased muscle mass and strength, increased risk of skeletal bone disease, worsened control of hyperglycemia, and increased risk of cardiac arrhythmias (9, 14, 39). Furthermore, abnormal acid-base homeostasis correlates with increased morality (29, 42, 45). We have recently shown that there are substantial sex differences in renal structure, ammonia excretion, and expression of proteins involved in ammonia metabolism and transport under basal conditions (23). We have also shown that there are substantial sex differences in the ammonia response to acid loading (22). However, as of yet, the mechanism of these sex differences has not been identified.
The purpose of the present study was to determine the role of testis-derived testosterone in renal ammonia metabolism and renal structure. First, we determined the effect of orchiectomy (ORCH) on kidney structure, proximal tubule volume density and cell height, ammonia excretion, and expression of key proteins involved in ammonia metabolism. Second, we determined whether testosterone replacement reversed the effects of ORCH. Finally, we identified the localization of the testosterone target receptor, androgen receptor (AR), in the kidney. Our results show that testis-derived testosterone regulates renal structure, proximal tubule size, ammonia excretion, and expression of key proteins involved in ammonia metabolism and that these effects may be mediated through activation of the AR.
METHODS
Animals.
C57BL/6 mice (4 mo old) for these experiments were obtained from the Jackson Laboratory (Bar Harbor, ME). Mice were maintained on standard rodent diet (18% protein, Harlan Teklad, Madison, WI) with free access to water. The University of Florida College of Medicine and the North Florida/South Georgia Veterans Health System Institutional Animal Care and Use Committees approved all animal protocols.
Orchiectomy.
Under isoflurane (2%) anesthesia, bilateral ORCH was performed through a single scrotal incision. Testicles were exposed and removed. The incision was closed with a single suture. Sham-operated control (SOC) mice, with scrotal incision and testicle exposure without removal, served as control mice. Postoperative pain control was provided using buprenorphine (0.05 mg/kg) every 12 h for 2 days.
Testosterone hormone replacement.
A subset of mice had a controlled-release pellet (60-day release, Innovative Research of America, Sarasota, FL) containing a 1.5 mg/pellet of testosterone subcutaneously implanted at the time of ORCH (ORCH + T). Control ORCH mice (ORCH + P) received placebo pellets that released no hormone.
Metabolic cage experiments.
Mice were placed in metabolic cages for 3 days and fed a powdered normal diet (18% protein; Harlan Teklad). Daily food consumption was measured, and urine samples were collected. Urine samples were collected in tubes containing water-equilibrated mineral oil to prevent evaporation. Daily urine volume and pH were recorded. Urine samples were stored at −20°C until analyzed further.
Blood analysis.
Blood was obtained by cannulation of the abdominal aorta in mice anesthetized with isoflurane, drawn into a heparinized syringe, and immediately analyzed for Na+, K+, and bicarbonate concentrations using a Siemens Microanalytic Blood Gas Analyzer (RAPIDLab 348 analyzer, Siemens, Munich Germany). Serum testosterone was measured in The University of Virginia School of Medicine Center for Research in Reproduction Ligand Assay and Analysis Core (IBL America mouse testosterone ELISA, Minneapolis, MN).
Urinary analysis.
Urine ammonia was measured using a modification of a commercially available kit (A7553, Pointe Scientific, Canton, MI), as previously described (23, 36). Urine pH was measured using a micro-pH electrode (ROSS semi-micro pH, Orion 8103BN, Thermo Scientific, Waltham, MA).
Antibodies.
Table 1 shows the antibodies used in these experiments, their suppliers, and their catalog/identification numbers.
Table 1.
Antibodies
| Target | Supplier | Location | Identification (catalog no.) | Dilution |
|---|---|---|---|---|
| PEPCK | Cayman Chemical | Ann Arbor, MI | 10004943 | W: 1:3,000 I: 1:5,000 |
| GS | Abcam | Cambridge, MA | ab73593 | W: 1:1,500 I: 1:60,000 |
| NBCe1 | Proteintech | Rosemont, IL | 885-1-AP | W: 1:2,000 |
| NHE3 | EMD Millipore | Temecula, CA | 2736700 | W: 1:500 I: 1:1,000 |
| NKCC2 | Dr. H. Moo Kwon | Ulsan National Institute of Science and Technology, Ulsan, South Korea | NA | W: 1:4,000 I: 1:50,000 |
| Rhbg | Weiner Laboratory | Gainesville, FL | 359 | W: 1:400 I: 1:50,000 |
| Rhcg | Weiner Laboratory | Gainesville, FL | 361 | I: 1:30,000 |
| AR | Dr. Gail Prins | University of Illinois at Chicago, Chicago, IL | AR21 | W: 1:500 I: 1:1,000 |
PEPCK, phosphoenolpyruvate carboxykinase; GS, glutamine synthetase; NBCe1, electrogenic Na+-bicarbonate cotransporter 1; NHE3, Na+/H+ exchanger isoform 3; NKCC2, Na+-K+-2Cl− cotransporter; Rhbg, Rhesus type B glycoprotein; Rhcg, Rhesus type C glycoprotein; AR, androgen receptor; W, Western blot; I, immunohistochemistry dilutions; NA, not applicable.
Protein preparation.
Animals were anesthetized with inhalant isoflurane, and kidneys were rinsed by in vivo cardiac perfusion with PBS (pH 7.4) containing 6,000 U/L sodium heparin and 120 mg/L lidocaine. The right kidney was removed rapidly, and the cortex, outer medulla, and inner medulla were isolated on a cold stage under a dissecting microscope. Samples were snap frozen in liquid nitrogen and stored frozen at −80°C until used. Tissues were homogenized in T-PER tissue protein extraction reagent (Pierce Biotechnology, Rockford, IL) using microtube pestles (USA Scientific, Ocala, FL), and protein was extracted according to the manufacturer’s recommendations. An aliquot was used for total protein quantification using a BCA assay, and the remainder was stored frozen at −80°C until use.
Immunoblot procedures.
Renal protein (10 μg) was electrophoresed on 10% PAGE ReadyGel (Bio-Rad, Hercules, CA). Gels were transferred electrophoretically to nitrocellulose membranes, blocked with 5 g/dL nonfat dry milk diluted in Blotto buffer (50 mM Tris, 150 mM NaCl, 5 mM Na2EDTA, and 0.05% Tween 20, pH 7.6), and incubated at 4°C overnight with primary antibody in nonfat dry milk. Loading and transfer equivalence were assessed with Ponceau S staining and by evaluating expression of the housekeeping protein β-actin. After being washed, membranes were exposed to secondary antibody (goat anti-rabbit IgG, Cell Signaling Technology, Beverly, MA) conjugated to horseradish peroxidase at a dilution of 1:5,000. Sites of antibody-antigen reaction were visualized using enhanced chemiluminescence (SuperSignal West Pico Substrate, Pierce) and a Kodak Image Station 440CF digital imaging system. In selected experiments, blots were stripped. Band density was normalized such that the mean density in a region (cortex or outer medulla) in SOC tissue was 100. The absence of saturation was confirmed by examining pixel intensity distribution in all immunoblots.
Tissue preparation for immunohistochemistry.
Mice were anesthetized with inhalant isoflurane. The kidneys were preserved by in vivo cardiac perfusion with PBS (pH 7.4) containing 6,000 U/L of sodium heparin and 120 mg/L lidocaine followed by periodate-lysine-2% paraformaldehyde, cut transversely into several 2- to 3-mm-thick slices, and then immersed for 24–30 h at 4°C in the same fixative. Kidney samples from each animal were embedded in polyester wax made with of polyethylene glycol 400 distearate (Polysciences, Warrington, PA) with 10% 1-hexadecanol, and 2-µm-thick sections were cut and mounted on gelatin-coated slides.
Immunohistochemistry.
Immunolocalization for Rhesus type B glycoprotein (Rhbg), Rhesus type C glycoprotein (Rhcg), Na+/H+ exchanger isoform 3 (NHE3), glutamine synthetase (GS), Na+-K+-2Cl− cotransporter (NKCC2), and AR was accomplished using previously described immunoperoxidase procedures (3, 35, 37, 52). Briefly, sections were dewaxed in ethanol, rehydrated, heated in Trilogy (Cell Marque, Rocklin, CA) to 88°C for 30 min and then to 96°C for 30 min, cooled for 30 min, and rinsed in PBS. Endogenous peroxidase activity was blocked by incubating the sections in 3% H2O2 in distilled water for 45 min. Sections were blocked for 15 min with serum-free protein block (Dako Cytomation) and then incubated overnight at 4°C with primary antibody. Sections were washed in PBS and incubated for 30 min with polymer-linked peroxidase-conjugated horse anti-rabbit IgG (ImmPRESS, Vector Laboratories, Burlingame, CA), washed again with PBS, and then exposed to diaminobenzidine for 5 min. Sections were washed with distilled water, dehydrated with xylene, mounted, and observed by light microscopy. Comparisons of labeling were made between sections from the same immunohistochemistry experiment. Sections were examined on a Nikon E600 microscope equipped with DIC optics and photographed using a DXM1200F digital camera and ACT-1 software (Nikon).
Immunolocalization for phosphoenolpyruvate (PEPCK) was accomplished using modified previously described immunoperoxidase procedures (17, 18, 32, 33). Briefly, sections were dewaxed in ethanol, rehydrated, heated in Trilogy (Cell Marque) to 96°C for 60 min, cooled for 30 min, and rinsed in PBS. Endogenous peroxidase activity was blocked by incubation of the sections in 3% H2O2 in methanol for 45 min. Sections were treated with 0.5% Triton X-100 in PBS for 15 min. Sections then underwent several washes in PBS containing 1% BSA, 0.05% saponin, and 0.2% gelatin followed by being blocked for 15 min with serum-free protein block (DAKO Cytomation) and then incubated overnight at 4°C with primary antibody. Sections were washed in PBS containing 0.1% BSA, 0.05% saponin, and 0.2% gelatin followed by PBS, incubated for 60 min with polymer-linked peroxidase-conjugated goat anti-rabbit IgG (MACH2. Biocare Medical, Concord, CA), washed again in PBS, and then exposed to diaminobenzidine for 5 min.
Negative control.
Each immunohistochemistry experiment included a section that was exposed to the immunolabeling procedure without primary antibody to ensure that the label was the result of the primary antibody binding only.
Morphometric analysis.
The volume density of the proximal tubule in the cortex and outer stripe of the outer medulla (OMo) as well as collecting duct in the cortex and inner stripe of the outer medulla (OMi) was determined using standard point-counting techniques as previously described (2, 23). Briefly, we used PEPCK immunolabel to identify proximal tubule segments and Rhbg immunolabel to identify collecting duct segments. We used high-resolution digital micrographs of PEPCK and Rhbg immunolabel overlaid with a standardized grid using Adobe Photoshop software. The volume density of the specific tubule of interest (proximal tubule or collecting duct) in each photomicrograph was calculated by dividing the number of points in the grid overlying the epithelial segment of interest (including tubule lumen) by the total number of points over all structures in the micrograph, excluding vessels, and then multiplied by 100. At least four photomicrographs were analyzed for each kidney, and data from all photomicrographs examined for each mouse were averaged to yield a single data point per animal for statistical analysis.
Quantitative immunohistochemistry.
Quantitative immunohistochemistry was performed as we have previously described and validated (23, 26, 33, 52). Proximal tubule segments were studied in sections labeled under identical conditions in the same immunolabeling experiment by an observer blinded to the treatment group. The specific proximal tubule segments measured were the initial proximal convoluted tubule (PCT), defined as PCT segments continuous with Bowman’s capsule, the proximal straight tubule (PST) in the medullary ray, and the PST in the OMo. We used high-resolution digital micrographs taken of randomly selected fields of the renal cortex and OMo using a NIKON E600 microscope equipped with a DXM1200F digital camera and ACT-1 software (Nikon) using no imaging enhancement techniques. Using ImageJ software (version 1.34j, National Institutes of Health, Bethesda, MD), we measured pixel intensity across a straight line drawn from the tubule lumen through an individual cell. These data were then analyzed using custom-written software executed in Microsoft Excel 2016. Net intensity at each pixel on the line was then determined as the difference between absolute intensity and mean background pixel intensity measured outside the cell. Total cellular expression was determined by integrating net pixel intensity across the entire cell. Cell height was determined as the distance in pixels between the apical and basolateral edges of the cells and concerted to absolute length using calibrated determination of individual pixel size. A minimum of 15 individual cells from at least 4 photomicrographs from each kidney were analyzed. Data from all cells examined of a given proximal tubule segment type were averaged to yield a single data point per animal for statistical analysis.
We determined type A intercalated cell size and cell-specific protein expression as previously detailed (23). Briefly, we obtained high-power digital micrographs of the OMi of tissue sections that had undergone immunohistochemistry for the intercalated cell-specific marker, Rhbg or Rhcg. Individual intercalated cells were circumscribed using ImageJ software (version 1.34j, National Institutes of Health). Cell size was determined as the number of pixels within the outlined regions and converted to area using calibrated measurement of pixels per micrometer. Immunolabel intensity at each pixel was determined as the difference between absolute pixel intensity and mean background intensity, and single cell immunolabel intensity was determined by integrating net pixel intensity within the cell using custom-written software executed in Microsoft Excel 2016.
Statistical analysis.
Results are presented as means ± SE; n refers to the number of animals studied. For quantitative immunohistochemistry, all measurements of a specific cell type in a mouse kidney were averaged and used as a single measurement for statistical analysis. Statistical analyses were performed using an unpaired Student’s t test or one-way ANOVA. P < 0.05 was considered statistically significant. We performed statistical analysis using SPSS Software and Microsoft Excel.
RESULTS
Effect of ORCH on physiological parameters at 4 and 8 wk.
We began by determining the effect of ORCH at 4 and 8 wk. ORCH decreased plasma testosterone levels significantly (Table 2). Food intake and plasma Na+, K+, and bicarbonate did not differ significantly. Despite no difference in food intake, both body and kidney weights were decreased significantly.
Table 2.
Physiological parameters 4 and 8 wk after orchiectomy
| Parameter | 4 wk |
8 wk |
||||
|---|---|---|---|---|---|---|
| Parameter | SOC | ORCH | P value | SOC | ORCH | P value |
| Plasma testosterone, ng/dL | 252 ± 275 | 27 ± 9 | <0.05 | 685 ± 467 | 36 ± 12 | <0.01 |
| Body weight at the indicated time, g | 31.1 ± 1.8 | 26.8 ± 9 | <0.01 | 35.6 ± 3.0 | 31.0 ± 1.3 | <0.01 |
| Change in body weight, g | +1.3 ± 0.6 | −2.7 ± 0.4 | <0.01 | +4.4 ± 1.6 | −0.4 ± 0.2 | <0.01 |
| Kidney weight, mg | 236 ± 27 | 169 ± 14 | <0.01 | 314 ± 20 | 231 ± 37 | <0.001 |
| Food intake, g/day | 11.3 ± 1.0 | 11.8 ± 0.5 | NS | 11.4 ± 1.3 | 10.4 ± 1.0 | NS |
| Plasma Na+, mmol/L | 149 ± 1 | 148 ± 2 | NS | 158 ± 3 | 156 ± 3 | NS |
| Plasma K+, mmol/L | 4.3 ± 0.4 | 4.3 ± 0.3 | NS | 4.5 ± 0.5 | 4.6 ± 0. | NS |
| Plasma , mmol/L | 21.3 ± 1.2 | 20.5 ± 2.0 | NS | 21.6 ± 1.4 | 20.1 ± 1.0 | NS |
| Net acid excretion, µmol/day | 134 ± 43 | 245 ± 52 | <0.01 | 131 ± 24 | 231 ± 33 | <0.01 |
| Urinary ammonia, µmol/day | 63 ± 15 | 125 ± 22 | <0.01 | 55 ± 14 | 106 ± 22 | <0.001 |
| Titratable acid, µmol/day | 70 ± 26 | 120 ± 30 | <0.05 | 76 ± 15 | 125 ± 26 | <0.05 |
| Urine pH | 6.66 ± 0.17 | 6.29 ± 0.20 | <0.01 | 6.53 ± 0.10 | 6.21 ± 0.11 | <0.01 |
Results are reported as means ± SE; n = 6 animals/group. SOC, sham-operated control; ORCH, orchiectomized; NS, not significant.
Net acid excretion involves two processes, ammonium () excretion and titratable acid excretion. ORCH increased net acid, ammonia, and titratable acid excretion significantly (Table 2 and Fig. 1, A and B). Urine pH was significantly lower in ORCH mice than SOC mice (P < 0.01). These differences were present at 4 wk and did not change further at 8 wk.
Fig. 1.

Ammonia excretion. A: ammonia excretion in sham-operated control (SOC) and orchiectomized (ORCH) mice at 4 wk postsurgery. Urinary ammonia excretion was significantly greater in ORCH mice than SOC mice at both time points. B: ammonia excretion in SOC and ORCH mice at 8 wk postsurgery. Urinary ammonia excretion was significantly greater in ORCH mice. C: ammonia excretion in SOC mice, control ORCH mice that received placebo pellets that released no hormone (ORCH + P), and mice that had a controlled-release pellet (60-day release) containing 1.5 mg/pellet of testosterone implanted subcutaneously at the time of ORCH (ORCH + T). ORCH + P mice had significantly greater ammonia excretion than did SOC mice or ORCH + T mice. n = 6 mice/group. NS, not significant.
Effect of testosterone replacement on physiological parameters.
We next determined whether the effects of ORCH were the result of loss of testosterone by evaluating the effect of testosterone replacement. Because findings in ORCH and SOC mice were almost identical at 4 and 8 wk, we compared ORCH + P, ORCH + T, and SOC mice at 4 wk. ORCH + P mice has significantly decreased plasma testosterone relative to both SOC and ORCH + T mice (Table 3). Neither body weight nor food intake was altered significantly [P = not significant (NS)]. ORCH + P mice had significantly decreased kidney weight compared with SOC and ORCH + T mice (P < 0.001). Neither plasma bicarbonate (P = NS) nor plasma electrolyte concentrations (P = NS) differed significantly.
Table 3.
Physiological parameters at 4 wk
| Parameter | SOC | ORCH + P | ORCH + T | P Value |
|---|---|---|---|---|
| Plasma testosterone, ng/dL | 120 ± 140 (8) | 24 ± 5 (7) | 159 ± 36 (7) | <0.05 |
| Body weight, g | 31.7 ± 2.4 (8) | 29.4 ± 2.0 (7) | 31.2 ± 2.4 (7) | NS |
| Change in body weight, g | +1.8 ± 0.8 (8) | −2.4 ± 0.7 (7) | +2.1 ± 1.3 (7) | <0.01 |
| Kidney weight, mg | 220 ± 16 (8) | 187 ± 18 (7) | 236 ± 19 (7) | <0.001 |
| Food intake, g/day | 12.3 ± 1.7 (8) | 12.6 ± 1.2 (7) | 13.6 ± 1.2 (7) | NS |
| Plasma Na+, mmol/L | 149 ± 2 (8) | 148 ± 2 (7) | 149 ± 2 (7) | NS |
| Plasma K+, mmol/L | 4.4 ± 0.2 (8) | 4.0 ± 0.5 (7) | 4.3 ± 0.3 (7) | NS |
| Plasma , mmol/L | 22.0 ± 2.1 (8) | 20.2 ± 1.9 (7) | 21.2 ± 3.0 (7) | NS |
| Net acid excretion, µmol/day | 126 ± 22 (8) | 246 ± 64 (6) | 105 ± 55 (6) | <0.01 |
| Ammonia excretion, µmol/day | 55 ± 13 (8) | 135 ± 32 (6) | 48 ± 28 (6) | <0.001 |
| Titratable acid excretion, µmol/day | 72 ± 19 (8) | 111 ± 56 (6) | 58 ± 28 (6) | NS |
| Urine pH | 6.61 ± 0.14 (8) | 6.32 ± 0.18 (6) | 6.51 ± 0.17 (6) | <0.01 |
| Urea nitrogen excretion, mg/day | 53 ± 12 (6) | 75 ± 15 (6) | 56 ± 11 (6) | NS |
Results are reported as means ± SE; numbers of mice are shown in parentheses. SOC, sham-operated control; ORCH + P, control orchiectomized (ORCH) mice that received placebo pellets that released no hormone; ORCH + T, mice that had a controlled-release pellet (60-day release) containing 1.5 mg/pellet of testosterone implanted subcutaneously subcutaneously at the time of ORCH.
ORCH + P mice had significantly increased net acid and ammonia excretion, which ORCH + T treatment reversed (P < 0.001; Fig. 1C). Titratable acid excretion and urea nitrogen excretion did not differ significantly (P = NS). Urine pH was significantly lower in ORCH + P mice than in SOC mice, which ORCH + T treatment reversed (P < 0.01). Thus, loss of testis-derived testosterone mediates the effect of ORCH on net acid and ammonia excretion.
Effect of ORCH on proximal tubule structure.
Because there are sex differences in male and female proximal tubule structure (23), we determined the effects of ORCH on the proximal tubule. In the cortex, ORCH appeared to decrease proximal tubule volume at 4 and 8 wk [see Supplemental Fig. S1A (Supplemental Data for this article are available online at https://doi.org/10.6084/m9.figshare.c.4771847)], and quantitative analysis confirmed this observation (Supplemental Fig. S1B). In the OMo, in contrast, ORCH did not alter proximal volume density (P = NS; Supplemental Fig. 1B). Thus, ORCH alters proximal tubule volume density in the cortex but not the outer medulla.
Effect of testosterone replacement on proximal tubule structure.
We next determined whether testosterone mediates the effects of ORCH on the proximal tubule. ORCH + P mice appeared to have decreased proximal tubule volume in the cortex, and this was reversed by ORCH + T treatment (Fig. 2A). Quantitative morphometric analysis confirmed this observation (P < 0.001; Fig. 2B). In the OMo, proximal tubule volume density did not differ significantly (P = NS; Fig. 2B). Thus, loss of testis-derived testosterone decreases cortical proximal tubule volume density.
Fig. 2.

Proximal tubule structure. A: low-power photomicrographs of phosphoenolpyruvate carboxykinase immunolabel in the cortex of sham-operated control (SOC; top) mice, control orchiectomized (ORCH) mice that received placebo pellets that released no hormone (ORCH + P; middle), and mice that had a controlled-release pellet (60-day release) containing 1.5 mg/pellet of testosterone implanted subcutaneously at the time of ORCH (ORCH + T; bottom). Proximal tubules appeared relatively sparse in the ORCH + P kidney than in SOC or ORCH + T kidneys. B: morphometric analysis of cortical and outer stripe of the outer medulla (OMo) proximal tubule volume density. ORCH + P mice had significantly lower cortical proximal tubule volume density than SOC or ORCH + T mice. OMo proximal tubule volume density did not differ significantly between the groups. n = 6 mice/group. C: quantitative analysis of proximal tubule cell height in the proximal convoluted tubule (PCT), cortical proximal straight tubule (PST), and outer medulla (OM) PST in SOC, ORCH + P, and ORCH + T mice. ORCH + P mice had significantly lower PCT and cortical PST cell height than did SOC or ORCH + T mice. OM PST cell height did not differ significantly between the groups. n = 6 mice/group. NS, not significant.
These changes in proximal tubule structure appear to involve changes in cell size. Proximal tubule cell height in the cortex (PCT and cortical PST) was decreased significantly by ORCH at both 4 and 8 wk (P < 0.01; Supplemental Fig. S1C). Testosterone appeared to mediate this difference. ORCH + P, but not ORCH + T, mice had decreased PCT and cortical PST cell height (P < 0.001; Fig. 2C). Thus, the loss of testis-derived testosterone results in smaller cortical proximal tubule cells and decreased cortical proximal tubule volume density.
In the outer medulla, in contrast, ORCH did not alter proximal tubule cell height significantly (P = NS; Supplemental Fig. S1C), nor did either ORCH + P or ORCH + T treatment (P = NS; Fig. 2C). These results parallel the lack of effect of ORCH on proximal tubule volume density in the outer medulla.
Effect of ORCH and testosterone replacement on collecting duct structure.
Because there are sex differences in the collecting duct (23), we determined the effects of ORCH + P and ORCH + T treatment on the collecting duct. Low-power micrographs showed no detectable difference in the proportion of the cortex that was collecting ducts (Fig. 3A), and morphometric analysis showed that collecting duct volume density did not differ significantly among groups (P = NS; Fig. 3B). There are also sex differences in intercalated cell size in the OMi (23). However, ORCH + P and ORCH + T treatment did not alter OMi intercalated cell size significantly (P = NS; Fig. 3C). Thus, testis-derived testosterone does not alter collecting duct volume density or OMi intercalated cell size.
Fig. 3.

Collecting duct structure. A: low-power photomicrographs of Rhesus type B glycoprotein (Rhbg) immunolabel in the cortex of sham-operated control (SOC; top) mice, control orchiectomized (ORCH) mice that received placebo pellets that released no hormone (ORCH + P; middle), and mice that had a controlled-release pellet (60-day release) containing 1.5 mg/pellet of testosterone implanted subcutaneously at the time of ORCH (ORCH + T; bottom). Collecting duct abundance did not appear different between the groups. B: morphometric analysis of cortical and inner stripe of the outer medulla (OMi) collecting duct volume density. There was no significant difference in either the cortex or OMi between the groups. n = 6 mice/group. C: single cell digital quantitative analysis of intercalated cell area in the OMi. There were no significant differences in OMi intercalated cell area between the groups. n = 6 mice/group. NS, not significant.
Ammoniagenic enzyme expression.
The observation that ORCH-induced testosterone deficiency increases ammonia excretion while decreasing proximal tubule volume density and size suggests testosterone directly regulates ammonia metabolism. To examine this, we determined its effects on key proteins in ammonia generation.
PEPCK is a proximal tubule enzyme with a key role in ammoniagenesis (7, 56, 57). Cortical PEPCK protein expression was significantly greater in ORCH mice than in SOC mice at both 4 and 8 wk (P < 0.01; Supplemental Fig. S2). This appeared to be the result of testosterone deficiency. ORCH + P treatment increased PEPCK expression, and ORCH + T treatment reversed this effect (P < 0.01; Fig. 4A). Immunohistochemistry for PEPCK showed similar findings. Qualitatively, ORCH + P treatment increased PEPCK immunolabel intensity in cortical proximal tubule, whereas ORCH + T treatment did not (Fig. 4B). Quantitative analysis showed similar results (P < 0.01; Fig. 4C). In the OMo, neither ORCH + P nor ORCH + T treatment altered PEPCK immunolabel intensity (Fig. 4C). Thus, ORCH-induced testosterone deficiency increases cortical, but not outer medullary, PEPCK expression.
Fig. 4.

Phosphoenolpyruvate carboxykinase (PEPCK) expression. A: immunoblot analysis of PEPCK expression with quantification of sham-operated control (SOC) mice, control orchiectomized (ORCH) mice that received placebo pellets that released no hormone (ORCH + P), and mice that had a controlled-release pellet (60-day release) containing 1.5 mg/pellet of testosterone implanted subcutaneously at the time of ORCH (ORCH + T). ORCH + P mice had significantly greater PEPCK protein expression than SOC or ORCH + T mice. B: immunohistochemistry examining PEPCK immunolabel in proximal convoluted tubule (PCT; top), cortical proximal straight tubule (PST; middle), and PST outer medulla (bottom) in SOC (left), ORCH + P (middle), and ORCH + T (right) mice. PEPCK immunolabel intensity was greater in cortical proximal tubule segments in ORCH + P mice than in SOC or ORCH + T mice. C: quantitative immunohistochemistry demonstrated that ORCH + P mice had significantly greater PEPCK expression in PCT and cortical PST than did SOC or ORCH + T mice. There was no significant difference in PEPCK expression between the groups in the proximal tubule in the outer stripe of the outer medulla (OM). n = 6 SOC mice, 6 ORCH + P mice, and 5 ORCH + T mice. NS, not significant.
The proximal tubule has an ammonia recycling capability involving GS (6, 52) that shows greater expression in female mice than in male mice (23). However, neither ORCH (Supplemental Fig. S3), ORCH + P, nor ORCH + T (P = NS; Fig. 5A) treatment altered GS expression significantly. Thus, testis-derived testosterone does not appear to regulate GS expression.
Fig. 5.
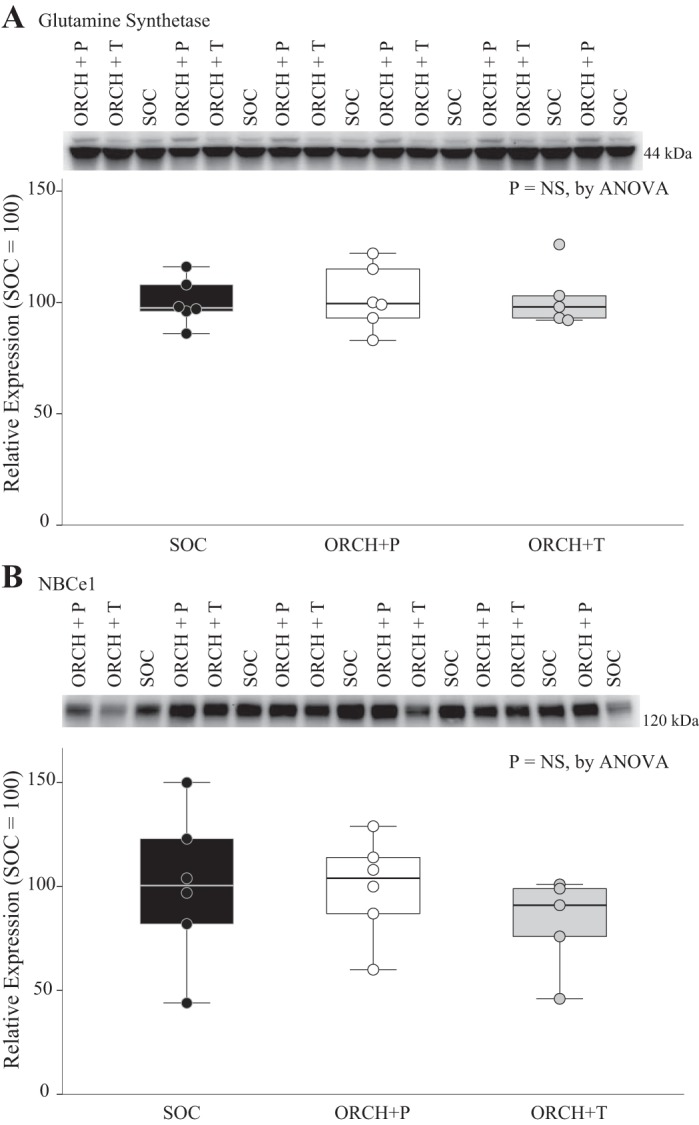
Glutamine synthetase (GS) and electrogenic Na+-bicarbonate cotransporter 1 (NBCe1) expression. A: immunoblot analysis of GS with quantification in homogenates of the renal cortex of sham-operated control (SOC) mice, orchiectomized (ORCH) mice that received placebo pellets that released no hormone (ORCH + P), and mice that had a controlled-release pellet (60-day release) containing 1.5 mg/pellet of testosterone implanted subcutaneously at the time of ORCH (ORCH + T). There was no significant difference in GS protein expression between groups. n = 6 SOC mice, 6 ORCH + P mice, and 5 ORCH + T mice. B: immunoblot analysis of NBCe1 with quantification in homogenates of the renal cortex of SOC, ORCH + P, and ORCH + T mice. There was no significant difference in NBCe1 protein expression between groups. n = 6 SOC mice, 6 ORCH + P mice, and 5 ORCH + T mice. NS, not significant.
Electrogenic Na+-bicarbonate cotransporter 1 expression.
Electrogenic Na+-bicarbonate cotransporter 1 (NBCe1) is a proximal tubule protein that regulates ammonia metabolism (21, 34). Using a pan-NBCe1 antibody that recognizes both NBCe1 variants expressed in the kidney, we found that neither ORCH + P nor ORCH + T treatment altered NBCe1 expression significantly (P = NS; Fig. 5B). Thus, testosterone-dependent effects on ammonia metabolism do not appear to result from changes in NBCe1 expression.
Effect of ORCH and testosterone replacement on ammonia transporters.
Ammonia excretion involves coordinated NH3/ transport by specific membrane proteins (53–56). This involves transporters expressed in the proximal tubule, thick ascending limb (TAL), and collecting duct, and sex differences in ammonia excretion correlate with differences in the expression of several key proteins (23).
NHE3 is thought generally to be the primary mechanism of proximal tubule ammonia secretion (31, 40, 41). ORCH decreased cortical NHE3 expression significantly, and this effect was reversed by testosterone replacement (P < 0.01; Fig. 6A). Immunohistochemistry showed parallel findings (Fig. 6B). Quantitative immunohistochemistry showed that ORCH + P treatment significantly decreased NHE3 immunolabel in the cortical proximal tubule (PCT and cortical PST, P < 0.05 vs. SOC) and that ORCH + T treatment abolished this difference (Fig. 6C). In the OMo, NHE3 immunolabel did not differ detectably among the three groups (P = NS; Fig. 6, B and C). Thus, ORCH-induced testosterone deficiency decreases cortical proximal tubule NHE3 expression.
Fig. 6.

Na+/H+ exchanger isoform 3 (NHE3) and Na+-K+-2Cl− cotransporter (NKCC2) expression. A: immunoblot analysis of cortical NHE3 expression with quantification of sham-operated control (SOC) mice, orchiectomized (ORCH) mice that received placebo pellets that released no hormone (ORCH + P), and mice that had a controlled-release pellet (60-day release) containing 1.5 mg/pellet of testosterone implanted subcutaneously at the time of ORCH (ORCH + T). ORCH + P mice had significantly less NHE3 expression than SOC or ORCH + T mice. B: immunohistochemistry examining NHE3 immunolabel in the proximal convoluted tubule (PCT; top), cortical proximal straight tubule (PST; middle), and PST outer medulla (OM; bottom) in SOC (left), ORCH + P (middle), and ORCH + T (right) mice. NHE3 immunolabel intensity was less in cortical proximal tubule segments in ORCH + P mice than SOC or ORCH + T mice. NHE3 immunolabel was not detectably different across groups in the PST OM. C: quantitative immunohistochemistry showing that ORCH + P mice had significantly less NHE3 expression in the PCT and cortical PST than did SOC or ORCH + T mice. There was no significant difference in NHE3 expression across groups in the PST OM. D: immunoblot analysis of cortical NKCC2 expression with quantification of SOC, ORCH + P, and ORCH + T mice. ORCH + P mice had significantly greater NKCC2 protein expression than SOC or ORCH + T mice. E: immunohistochemistry examining NKCC2 immunolabel in the cortex (CTX; top) and OM (bottom) in SOC (left), ORCH + P (middle), and ORCH + T (right) mice. NKCC2 immunolabel intensity was greater throughout the thick ascending limb (TAL) in ORCH + P mice than SOC or ORCH + T mice. n = 6 SOC mice, 6 ORCH + P mice, and 5 ORCH + T mice. NS, not significant.
NKCC2 mediates TAL ammonia reabsorption (13, 28). ORCH increased NKCC2 expression significantly at both 4 and 8 wk (P < 0.01; Supplemental Fig. S4). This effect of ORCH appeared to result from testosterone deficiency: ORCH + P treatment significantly increased cortical NKCC2 expression, and ORCH + T treatment abolished this difference (P < 0.01; Fig. 6D). Immunohistochemistry showed similar findings (Fig. 6E). Thus, physiological levels of testosterone suppresses NKCC2 expression in male mice.
The collecting duct secretes the majority of urinary ammonia, and the Rhesus glycoproteins Rhbg and Rhcg are the primary collecting duct ammonia-transporting proteins (4, 5, 20, 27). ORCH did not alter cortical Rhbg protein expression significantly at either 4 or 8 wk (Supplemental Fig. S5). Additional experiments at 4 wk showed no significant difference between ORCH + T, ORCH + P, or SOC Rhbg expression in the cortex (n = 6 SOC mice, 6 ORCH + P mice, and 5 ORCH + T mice, P = NS; Fig. 7A). Quantitative immunohistochemistry in the OMi showed similar findings (P = NS; Fig. 7B). Thus, testosterone does not alter Rhbg expression in male mice.
Fig. 7.

Rhesus protein expression. A: immunoblot analysis of cortical Rhesus type B glycoprotein (Rhbg) expression with quantification of sham-operated control (SOC) mice, control orchiectomized (ORCH) mice that received placebo pellets that released no hormone (ORCH + P), and mice that had a controlled-release pellet (60-day release) containing 1.5 mg/pellet of testosterone implanted subcutaneously at the time of ORCH (ORCH + T). There was no significant difference in protein expression in the cortex (CTX) between the groups. B, top: immunohistochemistry of Rhbg immunolabel in the inner stripe of the outer medulla (OMi) collecting duct in SOC (left), ORCH + P (middle), and ORCH + T (right) mice. There was no detectable difference in Rhbg immunolabel between groups. B, bottom: single cell digital quantitative analysis of OMi intercalated cells. There was no significant difference between groups. C, top: immunohistochemistry of Rhesus type C glycoprotein (Rhcg) immunolabel in the OMi collecting duct in SOC (left), ORCH + P (middle), ORCH + T (right). There was no detectable difference in Rhcg immunolabel between groups. C, bottom: single cell digital quantitative analysis of OMi intercalated cells. There was no significant difference between groups. n = 6 SOC mice, 6 ORCH + P mice, and 5 ORCH + T mice. NS, not significant.
Similar findings were observed when we examined Rhcg. Quantitative immunohistochemistry showed that Rhcg immunolabel intensity in OMi intercalated cells did not differ significantly between ORCH + P, ORCH + T, or SOC mice (P = NS; Fig. 7C). Thus, testosterone does not alter renal Rhbg or outer medullary collecting duct in the inner stripe intercalated cell Rhcg expression in male mice.
Renal AR localization.
The effects of testosterone are mediated primarily through activation of the AR (8, 48). Our next experiments determined AR expression in the mouse kidney. Because both sexes exhibit detectable serum testosterone levels, which might have biological effects in both sexes, we determined AR expression in both male and female kidneys (19). Real-time RT-PCR, using Taqman probes to confirm specificity of the amplified product, demonstrated AR mRNA expression in the cortex and outer medulla of both male and female kidneys. Expression did not differ significantly between male and female kidneys (P = NS by ANOVA; Table 4). Immunoblot analysis demonstrated AR protein expression by both sexes (Fig. 8A); expression was significantly greater in male mice than in female mice (P < 0.05; Fig. 8A).
Table 4.
Renal androgen receptor expression
| Sex | Cortex | Outer Medulla |
|---|---|---|
| Male | 29.8 ± 0.2 | 33.9 ± 2.3 |
| Female | 34.1 ± 7.1 | 26.8 ± 2.9 |
Results are reported as means ± SE and were normalized to the testes = 100; n = 3 mice/group.
Fig. 8.

Renal androgen receptor (AR) localization. A: immunoblot analysis of cortical AR expression with quantification of male and female mice. Male mice had significantly greater AR expression than female mice. CTX, cortex. B: immunohistochemistry of AR immunolabel in the proximal convoluted tubule (PCT; top), cortical proximal straight tubule (PST; middle), and outer medulla (OM) PST (bottom) in male (left) and female (right) mice. C: immunohistochemistry of the AR in parietal epithelial cells in male mice. D: double-label immunohistochemistry showed nuclear AR immunolabel throughout proximal tubule segments in both the cortex and OM. The nuclear AR immunolabel was present only in phosphoenolpyruvate carboxykinase (PEPCK)-positive proximal tubule cells in both the cortex and outer stripe of the OM in the mouse kidney. No AR immunolabel was detectable in PEPCK-negative cells. E: quantitative immunohistochemistry (IHC) demonstrated that male mice had significantly greater nuclear AR expression throughout the proximal tubule than did female mice. n = 6 mice/group.
Our next set of experiments used immunohistochemistry to determine the renal epithelial cell localization of the AR in male and female kidneys. Immunohistochemistry showed nuclear AR immunolabel throughout the proximal tubule in both male and female kidneys (Fig. 8B). Double-label immunohistochemistry, with the use of the proximal tubule-specific marker PEPCK, showed AR immunolabel in PEPCK-positive cells and no detectable immunolabel in PEPCK-negative epithelial cells (Fig. 8D). Immunohistochemistry also showed AR immunolabel in glomerular parietal epithelial cells, but only in the male kidney (Fig. 8C). There are axial variations along the proximal tubule function and in the expression of many proteins. Thus, we quantified axial AR immunolabel expression. Expression was lowest in the PCT and greatest in the PST in the outer medulla (P < 0.05; Fig. 8E). In all sites, expression was significantly greater in male mice (P < 0.001; Fig. 8E). Thus, there are both axial and sex differences in proximal tubule AR expression.
DISCUSSION
The present study provides important new information regarding sex differences in the kidney. ORCH increased ammonia excretion yet decreased total kidney size, proximal tubule volume density, and proximal tubule height, and testosterone replacement reversed these changes. The expression of multiple key proteins (PEPCK, NHE3, and NKCC2) involved in ammonia metabolism was altered by ORCH, and these were reversed with testosterone replacement. Finally, the AR is expressed in both male and female kidneys. In the renal tubule, the AR is present exclusively in the proximal tubule, where its expression exhibits axial heterogeneity. In the glomerulus, the AR is present in parietal epithelial cells, but only in male mice. These findings suggest that testis-derived testosterone, possibly acting through its receptor, AR, critically regulates renal ammonia excretion, proximal tubule structure, and expression of several key proteins required for ammonia metabolism.
Our first major observation is that testis-derived testosterone regulates kidney and proximal tubule size. The present study clearly demonstrates that testis-derived testosterone critically regulates both kidney and proximal tubule size, that this effect occurs in adult mice, occurs within 4 wk, is persistent at 8 wk, and is testosterone dependent. These results are consistent with excess androgen administration to male rats qualitatively increasing proximal tubule size (47). This effect of testis-derived testosterone is also consistent with the greater kidney size in males in mice, rats, and humans (38, 47, 48a). Moreover, the relative magnitude of these testosterone-dependent differences is almost identical to the male-female differences we have recently reported (23). It is also possible that the loss of testis-derived testosterone leads to a reduction in glomerular filtration rate that could also contribute to the observed changes in proximal tubule size. Previous studies, however, have not reported a consistent effect of male castration on glomerular filtration rate (1, 12). Thus, testosterone appears to be a major factor in sex differences in kidney and proximal tubule size.
The second major finding in the present study is that testis-derived testosterone regulates ammonia excretion. These effects likely involve direct effects of testosterone in the kidney. Although dietary fixed acid loads can alter ammonia excretion, the testosterone-dependent changes in ammonia excretion do not appear to be attributable to this mechanism, since neither ORCH nor testosterone replacement altered food intake. This effect of ORCH and testosterone on ammonia metabolism differs from most other conditions that regulate this critical process. Specifically, most changes in ammonia metabolism, such as occur with metabolic acidosis or hypokalemia, result in changes in kidney and proximal tubule size that parallel ammonia excretion, whereas ORCH-induced testosterone deficiency decreased total kidney and proximal tubule size, yet increased ammonia excretion. Thus, the effects of testosterone on ammonia excretion do not appear to be attributable to secondary mechanisms involving food intake or electrolyte differences.
Whether the skeletal muscle atrophy that occurs with ORCH contributes to or results partially from the increase in ammonia excretion is unclear. Acute increases in skeletal muscle protein catabolism, which might result from ORCH, can result in an additional endogenous acid load, which could increase ammonia excretion. Against this possibility is that the pattern of responses observed in orchiectomized male mice differed from the pattern we have observed previously in response to acid loading (22, 34). Also, the lack of significant difference in urinary urea excretion does not support ongoing acute protein breakdown. In contrast, conditions that stimulate ammonia metabolism, such as metabolic acidosis, increase skeletal muscle-derived glutamine release that is necessary for the increased ammonia generation and likely contributes to skeletal muscle atrophy observed in these conditions (30). Thus, it is possible that the increased skeletal muscle-derived glutamine release that is necessary for the increase in ammonia generation and excretion contributes to ORCH-induced skeletal muscle atrophy.
The proximal tubule generates the majority of urinary ammonia. Our previous work identified sexual dimorphism in proximal tubule ammonia generation (23). The results of the present study further extend these findings by showing that testis-derived testosterone modulates expression of the ammonia-generating enzyme PEPCK but not the ammonia-recycling enzyme GS. Importantly, the testosterone-dependent regulation of PEPCK expression is not a nonspecific effect related to proximal tubule size, since the effects of testosterone on proximal tubule size and volume density are the opposite of its effects on PEPCK expression. Moreover, the observation that testosterone regulates PEPCK but not GS expression identifies that GS is regulated though mechanisms distinct from that of PEPCK.
Ammonia produced in the proximal tubule is secreted preferentially in the luminal fluid. Apical NHE3 appears to be the primary mechanism of proximal tubule ammonia secretion (31, 40, 41), and the present study shows the loss of testis-derived testosterone decreases NHE3 expression. These findings are consistent with a previous report in hormonally manipulated rats (44). However, the effects of testosterone on NHE3 do not parallel either ammonia excretion or PEPCK expression. This suggests either that effects on other components of ammonia metabolism counterbalance changes in NHE3 expression. Finally, our previous findings, where proximal tubule NHE3 expression did not differ between male and female mice (23), in combination with the present findings suggest that testosterone-independent factors increase NHE3 expression in female mice.
TAL ammonia reabsorption establishes the interstitial ammonia gradient necessary for collecting duct ammonia secretion (13, 28). We have previously reported that NKCC2 expression is greater in female mice than in male mice (23). The present study shows that loss of testis-derived testosterone increased expression. Moreover, the magnitude of difference in NKCC2 expression in response to loss of testis-derived testosterone was almost identical to the difference between female and male mice (23). This suggests that testosterone has either direct or indirect effects on TAL NKCC2 expression. One possible indirect mechanism is that the observed proximal tubule atrophy in combination with decreased proximal tubule NHE3 expression could increase solute delivery to the TAL, which could lead to enhanced NKCC2 expression.
Collecting duct ammonia secretion is a major determinant of urinary ammonia excretion (54–56). The Rhesus glycoproteins Rhbg and Rhcg are the primary collecting duct ammonia transporters (4, 5, 20, 27). We have shown previous sexual dimorphism in the expression of both Rhbg and Rhcg in the collecting duct (23). However, the present study found no detectable change in Rhbg or Rhcg expression in response to ORCH or testosterone replacement. Testis-derived testosterone does not appear to mediate the sex differences in collecting duct Rhesus glycoprotein expression.
A third major finding in the present study is the identification of the cellular expression of testosterone’s canonical receptor, AR, in the kidney. The present study shows, for the first time, that AR protein is expressed in the proximal tubule in both male and female kidneys and not in other nephron/collecting duct cells and that there is axial heterogeneity in its proximal tubule expression. These observations substantially extend previous observations, which, although demonstrating AR protein and/or mRNA are expressed in both male and female kidneys, did not identify AR protein’s specific cellular localization (15, 46, 50, 58).
Proximal tubule AR expression likely mediates key aspects of some, but not all, sex differences in renal structure and ammonia metabolism. The presence of AR in proximal tubule cells likely explains the effects of testosterone on proximal tubule size and volume density, NHE3, and PEPCK expression. Moreover, the lack of collecting duct AR expression, a site where there are sex differences in Rhbg and Rhcg expression (20), suggests that mechanisms other than testosterone mediate sex differences in key components of ammonia metabolism. Finally, the sex- and testosterone-dependent differences in NKCC2 expression despite the lack of TAL AR expression suggest that NKCC2 expression involves AR-independent testosterone-dependent mechanisms. Whether this involves paracrine effects involving the proximal tubule cannot be determined at present.
The AR was present in outer medullary proximal tubule segments, but ORCH did not alter proximal tubule size, volume density, or ammoniagenic enzyme expression in these cells. One possible explanation is that nontestis-derived testosterone is sufficient to activate ARs in this segment. Testosterone levels were decreased, but still detectable, in orchiectomized mice in the present study, and female mice are known to have low but detectable testosterone levels (19). Consistent with this interpretation is that AR immunolabel was primarily nuclear in female mice and that AR activation results in nuclear translocation.
Finally, the present study demonstrates AR protein in glomerular parietal epithelial cells in the male, but not in the female, mouse. Another report found that AR expression in rat parietal epithelial cells but did not specify whether there were sex differences (50). If present in humans, this sex-dependent difference may contribute to sex differences in glomerular disorders that involve parietal epithelial cells, such as crescentic glomerulonephritis.
In summary, our study substantially advances our understanding of the role of androgens in modulating sex differences in renal ammonia metabolism. Testosterone regulates ammonia metabolism and excretion possibly through direct AR-mediated effects on proximal tubule structure and protein expression and indirect effects on TAL NKCC2 expression. Thus, testosterone, likely acting through the AR, modulates some of the identified sex differences in renal ammonia metabolism and transport.
GRANTS
This work was supported by National Institute of Diabetes and Digestive and Kidney Disease Grants R01-DK-045788 (to I. D. Weiner), R01-DK-107798 (to I. D. Weiner and J. W. Verlander), 5-T32-DK-104721, and K08-DK-120873 (to A. N. Harris).
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
A.N.H., J.W.V., and I.D.W. conceived and designed research; A.N.H., H.-W.L., and J.W.V. performed experiments; A.N.H., H.-W.L., J.W.V., and I.D.W. analyzed data; A.N.H., H.-W.L., J.W.V., and I.D.W. interpreted results of experiments; A.N.H. prepared figures; A.N.H. drafted manuscript; A.N.H., H.-W.L., J.W.V., and I.D.W. edited and revised manuscript; A.N.H., H.-W.L., J.W.V., and I.D.W. approved final version of manuscript.
ACKNOWLEDGMENTS
We thank Dr. Sharon W. Matthews and Chao Chen (University of Florida College of Medicine Electron Microscopy Core Laboratory) for the tissue processing for immunohistochemical experiments. We thank The University of Virginia School of Medicine Center for Research in Reproduction Ligand Assay and Analysis Core for measurement of serum testosterone levels. We thank Dr. Gail Prins for anti-androgen antibodies and expertise in the androgen receptor and Dr. Paul S. Cooke for expertise in male sex steroid hormones.
REFERENCES
- 1.Baylis C. Age-dependent glomerular damage in the rat. Dissociation between glomerular injury and both glomerular hypertension and hypertrophy. Male gender as a primary risk factor. J Clin Invest 94: 1823–1829, 1994. doi: 10.1172/JCI117531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bellhouse DR. Area estimation by point-counting techniques. Biometrics 37: 303–312, 1981. doi: 10.2307/2530419. [DOI] [Google Scholar]
- 3.Bishop JM, Verlander JW, Lee HW, Nelson RD, Weiner AJ, Handlogten ME, Weiner ID. Role of the Rhesus glycoprotein, Rh B glycoprotein, in renal ammonia excretion. Am J Physiol Renal Physiol 299: F1065–F1077, 2010. doi: 10.1152/ajprenal.00277.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Biver S, Belge H, Bourgeois S, Van Vooren P, Nowik M, Scohy S, Houillier P, Szpirer J, Szpirer C, Wagner CA, Devuyst O, Marini AM. A role for Rhesus factor Rhcg in renal ammonium excretion and male fertility. Nature 456: 339–343, 2008. doi: 10.1038/nature07518. [DOI] [PubMed] [Google Scholar]
- 5.Bourgeois S, Bounoure L, Christensen EI, Ramakrishnan SK, Houillier P, Devuyst O, Wagner CA. Haploinsufficiency of the ammonia transporter Rhcg predisposes to chronic acidosis: Rhcg is critical for apical and basolateral ammonia transport in the mouse collecting duct. J Biol Chem 288: 5518–5529, 2013. doi: 10.1074/jbc.M112.441782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Conjard A, Komaty O, Delage H, Boghossian M, Martin M, Ferrier B, Baverel G. Inhibition of glutamine synthetase in the mouse kidney: a novel mechanism of adaptation to metabolic acidosis. J Biol Chem 278: 38159–38166, 2003. doi: 10.1074/jbc.M302885200. [DOI] [PubMed] [Google Scholar]
- 7.Curthoys NP, Moe OW. Proximal tubule function and response to acidosis. Clin J Am Soc Nephrol 9: 1627–1638, 2014. doi: 10.2215/CJN.10391012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Davey RA, Grossmann M. Androgen receptor structure, function and biology: from bench to bedside. Clin Biochem Rev 37: 3–15, 2016. [PMC free article] [PubMed] [Google Scholar]
- 9.de Brito-Ashurst I, Varagunam M, Raftery MJ, Yaqoob MM. Bicarbonate supplementation slows progression of CKD and improves nutritional status. J Am Soc Nephrol 20: 2075–2084, 2009. doi: 10.1681/ASN.2008111205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Fekete A, Vannay A, Vér A, Vásárhelyi B, Müller V, Ouyang N, Reusz G, Tulassay T, Szabó AJ. Sex differences in the alterations of Na(+), K(+)-ATPase following ischaemia-reperfusion injury in the rat kidney. J Physiol 555: 471–480, 2004. doi: 10.1113/jphysiol.2003.054825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Gafter U, Ben-Bassat M, Levi J. Castration inhibits glomerular hypertrophy and proteinuria in uninephrectomized male rats. Eur J Clin Invest 20: 360–365, 1990. doi: 10.1111/j.1365-2362.1990.tb01871.x. [DOI] [PubMed] [Google Scholar]
- 13.Good DW. Ammonium transport by the thick ascending limb of Henle’s loop. Annu Rev Physiol 56: 623–647, 1994. doi: 10.1146/annurev.ph.56.030194.003203. [DOI] [PubMed] [Google Scholar]
- 14.Goraya N, Simoni J, Jo CH, Wesson DE. Treatment of metabolic acidosis in patients with stage 3 chronic kidney disease with fruits and vegetables or oral bicarbonate reduces urine angiotensinogen and preserves glomerular filtration rate. Kidney Int 86: 1031–1038, 2014. doi: 10.1038/ki.2014.83. [DOI] [PubMed] [Google Scholar]
- 15.Grimont A, Bloch-Faure M, El Abida B, Crambert G. Mapping of sex hormone receptors and their modulators along the nephron of male and female mice. FEBS Lett 583: 1644–1648, 2009. doi: 10.1016/j.febslet.2009.04.032. [DOI] [PubMed] [Google Scholar]
- 17.Han K-H, Croker BP, Clapp WL, Werner D, Sahni M, Kim J, Kim H-Y, Handlogten ME, Weiner ID. Expression of the ammonia transporter, rh C glycoprotein, in normal and neoplastic human kidney. J Am Soc Nephrol 17: 2670–2679, 2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Han K-H, Lee H-W, Handlogten ME, Whitehill F, Osis G, Croker BP, Clapp WL, Verlander JW, Weiner ID. Expression of the ammonia transporter family member, Rh B glycoprotein, in the human kidney. Am J Physiol Renal Physiol 304: F972–F981, 2013. doi: 10.1152/ajprenal.00550.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Handelsman DJ, Jimenez M, Singh GK, Spaliviero J, Desai R, Walters KA. Measurement of testosterone by immunoassays and mass spectrometry in mouse serum, testicular, and ovarian extracts. Endocrinology 156: 400–405, 2015. doi: 10.1210/en.2014-1664. [DOI] [PubMed] [Google Scholar]
- 20.Handlogten ME, Hong SP, Westhoff CM, Weiner ID. Apical ammonia transport by the mouse inner medullary collecting duct cell (mIMCD-3). Am J Physiol Renal Physiol 289: F347–F358, 2005. doi: 10.1152/ajprenal.00253.2004. [DOI] [PubMed] [Google Scholar]
- 21.Handlogten ME, Osis G, Lee HW, Romero MF, Verlander JW, Weiner ID. NBCe1 expression is required for normal renal ammonia metabolism. Am J Physiol Renal Physiol 309: F658–F666, 2015. doi: 10.1152/ajprenal.00219.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Harris AN, Lee HW, Fang L, Verlander JW, Weiner ID. Differences in acidosis-stimulated renal ammonia metabolism in the male and female kidney. Am J Physiol Renal Physiol 317: F890–F905, 2019. doi: 10.1152/ajprenal.00244.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Harris AN, Lee HW, Osis G, Fang L, Webster KL, Verlander JW, Weiner ID. Differences in renal ammonia metabolism in male and female kidney. Am J Physiol Renal Physiol 315: F211–F222, 2018. doi: 10.1152/ajprenal.00084.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23a.Institute of Medicine (US) Committee on Understanding the Biology of Sex and Gender Differences Exploring the Biological Contributions to Human Health: Does Sex Matter?, edited by Wizemann TM, Pardue M-L. Washington, DC: National Academies, 2001. [PubMed] [Google Scholar]
- 24.Karp NA, Mason J, Beaudet AL, Benjamini Y, Bower L, Braun RE, Brown SDM, Chesler EJ, Dickinson ME, Flenniken AM, Fuchs H, Angelis MH, Gao X, Guo S, Greenaway S, Heller R, Herault Y, Justice MJ, Kurbatova N, Lelliott CJ, Lloyd KCK, Mallon AM, Mank JE, Masuya H, McKerlie C, Meehan TF, Mott RF, Murray SA, Parkinson H, Ramirez-Solis R, Santos L, Seavitt JR, Smedley D, Sorg T, Speak AO, Steel KP, Svenson KL, Wakana S, West D, Wells S, Westerberg H, Yaacoby S, White JK; International Mouse Phenotyping Consortium . Prevalence of sexual dimorphism in mammalian phenotypic traits. Nat Commun 8: 15475, 2017. doi: 10.1038/ncomms15475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kher A, Meldrum KK, Wang M, Tsai BM, Pitcher JM, Meldrum DR. Cellular and molecular mechanisms of sex differences in renal ischemia-reperfusion injury. Cardiovasc Res 67: 594–603, 2005. doi: 10.1016/j.cardiores.2005.05.005. [DOI] [PubMed] [Google Scholar]
- 26.Kim HY, Baylis C, Verlander JW, Han KH, Reungjui S, Handlogten ME, Weiner ID. Effect of reduced renal mass on renal ammonia transporter family, Rh C glycoprotein and Rh B glycoprotein, expression. Am J Physiol Renal Physiol 293: F1238–F1247, 2007. doi: 10.1152/ajprenal.00151.2007. [DOI] [PubMed] [Google Scholar]
- 27.Kim HY, Verlander JW, Bishop JM, Cain BD, Han KH, Igarashi P, Lee HW, Handlogten ME, Weiner ID. Basolateral expression of the ammonia transporter family member Rh C glycoprotein in the mouse kidney. Am J Physiol Renal Physiol 296: F543–F555, 2009. doi: 10.1152/ajprenal.90637.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kinne R, Kinne-Saffran E, Schütz H, Schölermann B. Ammonium transport in medullary thick ascending limb of rabbit kidney: involvement of the Na+,K+,Cl−-cotransporter. J Membr Biol 94: 279–284, 1986. doi: 10.1007/BF01869723. [DOI] [PubMed] [Google Scholar]
- 29.Kovesdy CP, Anderson JE, Kalantar-Zadeh K. Association of serum bicarbonate levels with mortality in patients with non-dialysis-dependent CKD. Nephrol Dial Transplant 24: 1232–1237, 2009. doi: 10.1093/ndt/gfn633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kraut JA, Madias NE. Adverse effects of the metabolic acidosis of chronic kidney disease. Adv Chronic Kidney Dis 24: 289–297, 2017. doi: 10.1053/j.ackd.2017.06.005. [DOI] [PubMed] [Google Scholar]
- 31.Laghmani K, Preisig PA, Moe OW, Yanagisawa M, Alpern RJ. Endothelin-1/endothelin-B receptor-mediated increases in NHE3 activity in chronic metabolic acidosis. J Clin Invest 107: 1563–1569, 2001. doi: 10.1172/JCI11234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lee HW, Handlogten ME, Osis G, Clapp WL, Wakefield DN, Verlander JW, Weiner ID. Expression of sodium-dependent dicarboxylate transporter 1 (NaDC1/SLC13A2) in normal and neoplastic human kidney. Am J Physiol Renal Physiol 312: F427–F435, 2017. doi: 10.1152/ajprenal.00559.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lee HW, Osis G, Handlogten ME, Lamers WH, Chaudhry FA, Verlander JW, Weiner ID. Proximal tubule-specific glutamine synthetase deletion alters basal and acidosis-stimulated ammonia metabolism. Am J Physiol Renal Physiol 310: F1229–F1242, 2016. doi: 10.1152/ajprenal.00547.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lee HW, Osis G, Harris AN, Fang L, Romero MF, Handlogten ME, Verlander JW, Weiner ID. NBCe1-A regulates proximal tubule ammonia metabolism under basal conditions and in response to metabolic acidosis. J Am Soc Nephrol 29: 1182–1197, 2018. doi: 10.1681/ASN.2017080935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lee HW, Verlander JW, Bishop JM, Handlogten ME, Han KH, Weiner ID. Renal ammonia excretion in response to hypokalemia: effect of collecting duct-specific Rh C glycoprotein deletion. Am J Physiol Renal Physiol 304: F410–F421, 2013. doi: 10.1152/ajprenal.00300.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lee HW, Verlander JW, Bishop JM, Igarashi P, Handlogten ME, Weiner ID. Collecting duct-specific Rh C glycoprotein deletion alters basal and acidosis-stimulated renal ammonia excretion. Am J Physiol Renal Physiol 296: F1364–F1375, 2009. doi: 10.1152/ajprenal.90667.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lee HW, Verlander JW, Handlogten ME, Han KH, Weiner ID. Effect of collecting duct-specific deletion of both Rh B glycoprotein (Rhbg) and Rh C glycoprotein (Rhcg) on renal response to metabolic acidosis. Am J Physiol Renal Physiol 306: F389–F400, 2014. doi: 10.1152/ajprenal.00176.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.MacKay EM, Raulston BO. Factors which determine renal weight: XI. Renal function. J Exp Med 53: 109–113, 1931. doi: 10.1084/jem.53.1.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mitch WE. Metabolic and clinical consequences of metabolic acidosis. J Nephrol 19, Suppl 9: S70–S75, 2006. [PubMed] [Google Scholar]
- 40.Nagami GT. Luminal secretion of ammonia in the mouse proximal tubule perfused in vitro. J Clin Invest 81: 159–164, 1988. doi: 10.1172/JCI113287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Nagami GT. Role of angiotensin II in the enhancement of ammonia production and secretion by the proximal tubule in metabolic acidosis. Am J Physiol Renal Physiol 294: F874–F880, 2008. doi: 10.1152/ajprenal.00286.2007. [DOI] [PubMed] [Google Scholar]
- 42.Navaneethan SD, Schold JD, Arrigain S, Jolly SE, Wehbe E, Raina R, Simon JF, Srinivas TR, Jain A, Schreiber MJ Jr, Nally JV Jr. Serum bicarbonate and mortality in stage 3 and stage 4 chronic kidney disease. Clin J Am Soc Nephrol 6: 2395–2402, 2011. doi: 10.2215/CJN.03730411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Neugarten J, Acharya A, Silbiger SR. Effect of gender on the progression of nondiabetic renal disease: a meta-analysis. J Am Soc Nephrol 11: 319–329, 2000. [DOI] [PubMed] [Google Scholar]
- 44.Quan A, Chakravarty S, Chen JK, Chen JC, Loleh S, Saini N, Harris RC, Capdevila J, Quigley R. Androgens augment proximal tubule transport. Am J Physiol Renal Physiol 287: F452–F459, 2004. doi: 10.1152/ajprenal.00188.2003. [DOI] [PubMed] [Google Scholar]
- 45.Raphael KL, Murphy RA, Shlipak MG, Satterfield S, Huston HK, Sebastian A, Sellmeyer DE, Patel KV, Newman AB, Sarnak MJ, Ix JH, Fried LF; Health ABC Study . Bicarbonate concentration, acid-base status, and mortality in the health, aging, and body composition study. Clin J Am Soc Nephrol 11: 308–316, 2016. doi: 10.2215/CJN.06200615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ruizeveld de Winter JA. Immunocytochemical localization of androgen receptor with polyclonal antibody in paraffin-embedded human tissues. J Histochem Cytochem 42: 125–126, 1994. doi: 10.1177/42.1.8263324. [DOI] [PubMed] [Google Scholar]
- 47.Sabolić I, Asif AR, Budach WE, Wanke C, Bahn A, Burckhardt G. Gender differences in kidney function. Pflugers Arch 455: 397–429, 2007. doi: 10.1007/s00424-007-0308-1. [DOI] [PubMed] [Google Scholar]
- 48.Schiffer L, Arlt W, Storbeck KH. Intracrine androgen biosynthesis, metabolism and action revisited. Mol Cell Endocrinol 465: 4–26, 2018. doi: 10.1016/j.mce.2017.08.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48a.Selye H; H S . The effect of testosterone on the kidney. J Urol 42: 637–641, 1939. doi: 10.1016/S0022-5347(17)71560-1. [DOI] [Google Scholar]
- 49.Takaoka M, Yuba M, Fujii T, Ohkita M, Matsumura Y. Oestrogen protects against ischaemic acute renal failure in rats by suppressing renal endothelin-1 overproduction. Clin Sci (Lond) 103, Suppl 48: 434S–437S, 2002. doi: 10.1042/CS103S434S. [DOI] [PubMed] [Google Scholar]
- 50.Takeda H, Chodak G, Mutchnik S, Nakamoto T, Chang C. Immunohistochemical localization of androgen receptors with mono- and polyclonal antibodies to androgen receptor. J Endocrinol 126: 17–25, 1990. doi: 10.1677/joe.0.1260017. [DOI] [PubMed] [Google Scholar]
- 51.Tarry-Adkins JL, Ozanne SE, Norden A, Cherif H, Hales CN. Lower antioxidant capacity and elevated p53 and p21 may be a link between gender disparity in renal telomere shortening, albuminuria, and longevity. Am J Physiol Renal Physiol 290: F509–F516, 2006. doi: 10.1152/ajprenal.00215.2005. [DOI] [PubMed] [Google Scholar]
- 52.Verlander JW, Chu D, Lee HW, Handlogten ME, Weiner ID. Expression of glutamine synthetase in the mouse kidney: localization in multiple epithelial cell types and differential regulation by hypokalemia. Am J Physiol Renal Physiol 305: F701–F713, 2013. doi: 10.1152/ajprenal.00030.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Weiner ID. Roles of renal ammonia metabolism other than in acid-base homeostasis. Pediatr Nephrol 32: 933–942, 2017. doi: 10.1007/s00467-016-3401-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Weiner ID, Hamm LL. Molecular mechanisms of renal ammonia transport. Annu Rev Physiol 69: 317–340, 2007. doi: 10.1146/annurev.physiol.69.040705.142215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Weiner ID, Verlander JW. Ammonia transport in the kidney by Rhesus glycoproteins. Am J Physiol Renal Physiol 306: F1107–F1120, 2014. doi: 10.1152/ajprenal.00013.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Weiner ID, Verlander JW. Ammonia transporters and their role in acid-base balance. Physiol Rev 97: 465–494, 2017. doi: 10.1152/physrev.00011.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Weiner ID, Verlander JW. Renal ammonia metabolism and transport. Compr Physiol 3: 201–220, 2013. doi: 10.1002/cphy.c120010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Wilson CM, McPhaul MJ. A and B forms of the androgen receptor are expressed in a variety of human tissues. Mol Cell Endocrinol 120: 51–57, 1996. doi: 10.1016/0303-7207(96)03819-1. [DOI] [PubMed] [Google Scholar]