

Abstract
Terpenes are the largest class of natural products with extensive structural diversity and are widely used as pharmaceuticals, herbicides, flavourings, fragrances, and biofuels. While they have mostly been isolated from plants and fungi, the availability and analysis of bacterial genome sequence data indicates that bacteria also possess many putative terpene synthase genes. In this study, we further explore this potential for terpene synthase activity in bacteria. Twenty two potential class I terpene synthase genes (TSs) were selected to represent the full sequence diversity of bacterial synthase candidates and recombinantly expressed in E. coli. Terpene synthase activity was detected for 15 of these enzymes, and included mono-, sesqui- and diterpene synthase activities. A number of confirmed sesquiterpene synthases also exhibited promiscuous monoterpene synthase activity, suggesting that bacteria are potentially a richer source of monoterpene synthase activity then previously assumed. Several terpenoid products not previously detected in bacteria were identified, including aromandendrene, acora-3,7(14)-diene and longiborneol. Overall, we have identified promiscuous terpene synthases in bacteria and demonstrated that terpene synthases with substrate promiscuity are widely distributed in nature, forming a rich resource for engineering terpene biosynthetic pathways for biotechnology.
Introduction
Terpenoids, or isoprenoids, are a large class of structurally diverse natural products, with more than 80,000 compounds described in the Dictionary of Natural Compounds (http://dnp.chemnetbase.com). The vast majority of terpenoids have been isolated from plants and fungi; however, bacteria are also known producers of volatile odoriferous metabolites. All terpenoids are synthesised from the universal C5 isoprenoid precursors isopentenyl diphosphate (IPP) and dimethylallyl diphosphate (DMAPP), which are joined by isoprenyl transferases to form isoprenyl diphosphate substrates of varying lengths, such as geranyl diphosphate (GPP, C10), farnesyl diphosphate (FPP, C15) and geranylgeranyl diphosphate (GGPP, C20). Terpene synthases (TSs) convert the linear isoprenyl diphosphate substrates into structurally diverse mono- (C10), sesqui- (C15) and diterpene (C20) scaffolds. Due to their structural diversity, terpenoids have a wide range of industrial applications as pharmaceuticals, flavourings, fragrances, antimicrobials, pesticides and alternative fuels [1]. Recovery of terpenoids from natural sources is hampered by the accumulation of low quantities of target compounds and the dependence on crop yields. Likewise, chemical synthesis is hampered by the structural complexity of terpenoids, which often contain multiple stereo-centres, making synthetic chemistry expensive and environmentally costly. Recent advances in synthetic biology offer new routes to the diverse terpenoid chemistry via the expression of TSs in the presence of heterologous isoprenoid production pathways in engineered microorganisms [2–7].
TSs are capable of creating chemical diversity by converting linear precursors, via diphosphate abstraction (class I enzymes) or protonation of an olefinic double bond (class II), resulting in highly reactive allylic cations. The allylic cations can then undergo wider chemical reactions such as intramolecular attack on olefinic double bonds, hydride or proton migration, and Wagner–Meerwein rearrangement. Ultimately, the reaction is terminated by deprotonation or nucleophilic attack, furthermore, the initial product may be re-ionised for a second round of processing [8]. Hydrophobic residues form a pocket which protects the cationic intermediates from the undirected attack of water, and aromatic residues stabilize charged intermediates by cation–π interactions [9,10]. Such relatively inert active sites result in very little overall sequence similarity between TSs. Despite this overall low sequence similarity, TSs from plants, fungi, and bacteria, share a common fold and contain highly conserved metal-binding motifs [11]. Class I enzymes exhibit a highly conserved aspartate-rich motif (DDXXD) at the active centre and a (N,D)D(L,I,V)X(S,T)XXXE consensus sequence (NSE/DTE triad) involved in Mg2+ cofactor binding that complexes the diphosphate for ionization in order to cyclise the linear precursors. Similarly, Class II enzymes share a highly conserved DXDD motif for protonation of the substrate (11). Interestingly, a newly discovered class of microbial-like TPSs (MTPSL) from plants, alongside the common class I DDXXD signature motif, contain additional DDXXXD and DDXXX motifs suggesting a divergent evolutionary origin [12–14].
Actinomycetes, including many Streptomyces species, are known producers of terpenoids, including the terpene derivatives responsible for the characteristic musty odours of moist soil, geosmin and 2-methylisoborneol [15,16]. Geosmin synthase is an unusual bi-domain TS which exhibits an atypical mechanism, the N-terminal domain catalyses the cyclisation of FPP to germacradienol, while the C-terminal domain catalyses the conversion of germacradienol to geosmin resulting in the elimination of acetone [17]. Pentalenolactone is a sesquiterpenoid antibiotic that has been isolated from over 30 different Streptomyces species [18–20], and pentalenene synthase, the enzyme responsible for the cyclisation of FPP into pentalenene (the pentalenolactone hydrocarbon precursor), which is the first characterised bacterial TS [21]. After geosmin and 2-methylisoborneol synthases, epi-isozizaene synthases are the most widespread TSs found in bacteria, invariably from Streptomyces species [22]. Most characterised bacterial TSs exhibit sesquiterpene synthase activity, and only a few true bacterial monoterpene synthases are known so far [6,23].
Recent genome mining studies suggest that bacteria are a potentially much richer source of TSs, including enzymes from Gram-negative bacteria such as Cyanobacteria and Proteobacteria [22,24]. Since bacterial TSs, in particular, show only weak sequence conservation, an amino acid sequence-based product prediction for TSs is not possible. Hidden Markov Model (HMM) profiles are more suitable for sensitive database searches as they use statistical descriptions of a consensus sequence from one family. This approach was first applied to bacterial TSs by Komatsu et al (2008) to identify 2-methylisoborneol synthases in Actinomycetes [25]. Using similar approaches, the number of uncharacterised bacterial terpene cyclases has recently increased to more than 600 regular class I terpene synthases, 400 geosmin synthases and over 120 2-methylisoborneol synthases, due to the ever-increasing availability of genome sequencing data [22,24,26–28].
In this study, we further explore the potential for terpene synthase activity in bacteria. We have constructed a neighbour-joining tree containing 2,167 putative bacterial terpene cyclases/synthases, including geosmin synthases, 2-methylisoborneol synthases, and regular TSs, by using the Pfam HMM motif containing the two signature domains of the class I terpene synthase for the analysis of publicly available bacterial genome sequences. We were especially interested in determining the biochemical function of presumptive TSs that are separated by the neighbour-joining analysis into isolated clusters that do not contain any other assigned function and belong to diverse bacterial species other than Streptomyces. We have selected 22 potential class I TSs from different clusters and a diverse range of bacterial species for e.g. thermophilic or thermotolerant Actinobacteria (Actinomycetes), Proteobacteria, Firmicutes, Flavobacteria and Myxobacteria for recombinant expression in Escherichia coli. We screened for potential TS activity with GPP, FPP, and GGPP substrates using a combination of in vitro assays on purified recombinant proteins, and in vivo assays employing an engineered E. coli strain containing a heterologous mevalonate (MVA) pathway [7,29]. TS activity was detected for 15 enzymes, and included mono-, sesqui- and di-terpene synthase activities. Active TSs was obtained from several Proteobacteria species, as well as radio- or thermo-tolerant Actinobacteria species. Interestingly, several enzymes were active on more than one prenyl-pyrophosphate substrate. In particular, a number of sesquiterpene synthases also exhibited monoterpene synthase activity, suggesting that bacteria are a potentially much richer source of monoterpene synthase activity then previously assumed. Several new terpenoid products not seen before in bacterial species were detected, including aromandendrene, acora-3,7(14)-diene and longiborneol.
Materials and methods
Bioinformatic screening and selection of terpene synthases
Based on previous work [30], the HMM motif of Terpene_synth_C, which contains the two signature domains of class I TSs (DDXXD and (N,D)D(L,I,V)X(S,T)XXXE), with the HMM score larger than 23 was searched against 73,714 total protein sequences from 8,509 complete genomes of bacteria in the NCBI database (https://www.ncbi.nlm.nih.gov/genome/, 20 June, 2016), using the HMM search module in HMMER ver. 3.1b2 (hmmer.org). From the hits, a neighbour-joining tree of 2,167 known and unknown mono-, sesqui- and di-TSs was generated using the MAFFT program, ver.7.299 [31] with the options of—tree out—global pair—reorder—distout, and visualized on the iTOL (https://itol.embl.de/) [32] (Fig 1).
Fig 1. Neighbor-joining analysis of bacterial TSs.

Twenty two TSs selected in this study are named. TSs in red are those whose functionality could not be characterized in this work. The functionality of the selected TSs characterized in this study is indicated in colour: they produce either sesquiterpenoids (purple), diterpenoids (blue), or a mixture of sesqui- and mono-terpenoids (green). Known TSs were annotated in the tree and branches were indicated in colour based on their functionality, monoterpenoids (light blue), 2- methyisoborneol (orange), sesquiterpenoids (purple) and diterpenoids (blue) and also listed in S4 Table.
Plasmid construction
For in vivo diterpenoid synthase activity, 8 TSs listed in Table 1 were amplified by pETM11-fw and pETM11-rev using a pETM11-TS as a template containing homologous sequences at both ends for Gibson assembly [33]. IspAM22 (D2G, C155G) inserts were created by amplification of the ispA gene from E. coli DH5α with primers containing the desired base changes (S1 Table). A primer extension method was used to amplify the full-length gene which was inserted into pBbA2k-ispAM22-TS (for full list see S1 Table) by annealing at 50°C using Gibson assembly mix (NEB). For mono- or sesqui-terpene production pBbb2a-GPPS plasmid backbone was used for cloning and TSs (S1 Table) were amplified by pETM11-fw and pETM11-rev using a pETM11-TS as a template containing homologous sequences at both ends for Gibson assembly.
Table 1. List of 22 tested TSs and their activity.
| Gene ID | Source | Products | ||||
|---|---|---|---|---|---|---|
| GPP (C10) | FPP (C15) | GGPP(C20) | ||||
| 1 | AHY47823.1 | RrNerS | Rubrobacter radiotolerans | β-Myrcene‡, Ocimene†, R/S-Linalool†, Geraniol‡ | (±)-Nerolidol#, α/β-Farnesene‡, Farnesol‡ | NT |
| 2 | AHY45426.1 | RrBerS | Rubrobacter radiotolerans | β-Myrcene‡, Linalool#, trans-Geraniol‡ | α-Bergamotene#, (±)-trans Farnesene‡, (±)-trans Nerolidol‡, Farnesol‡ | NT |
| 3 | KYF56472.1 | ScAroS | Sorangium cellulosum | cis and trans- Ocimene‡, β-Pinene‡, R/S-Linalool‡ | Aromandendrene# | NT |
| 4 | KFG92939.1 | BpLonS | Burkholderia paludis | Geraniol‡ | (±)-Cadinene#, Longiborneol# | NT |
| 5 | AHH94051.1 | KaGerS | Kutzneria albida DSM 43870 | ND | Germacrene D# Germacradienol#, | ND |
| 6 | KPH97530.1 | AbAcoS | Actinobacteria bacterium OV450 | ND | Copaene†, Acora-3,7(14)-diene† | NT |
| 7 | ADU09376.1 | MiGerS | Micromonospora sp. L5 | ND | Germacrene A#, 1(10),4,11-Germacratriene# | NT |
| 8 | CAN96536.1 | ScEpiS | Sorangium cellulosum So ce56 [49] | cis and trans- Ocimene#, R/S Linalool#, β-Pinene | 10-epi-α/β-Cubinene#, β-Copaene #, Cadina-3,5-diene#, Cubebol#, α/γ/ δ-Cadinene#, 1,10-di-epi-Cubebol#, Cubenol# | ND |
| 9 | CAB41566.1 | ScGeoS | Streptomyces coelicolor A3(2) | β-Myrcene†, β-Ocimene†, Linalool†, Geraniol† | Geosmin†, Germacradienol#, Germacrene D# | NT |
| 10 | ACY98649.1 | TcCubS | Thermomonospora curvata DSM 43183 | ND | Cubebol‡ | ND |
| 11 | EXG82599.1 | CaCubS | Cryptosporangium arvum DSM 44712 | ND | Cubebol‡ | ND |
| 12 | ELQ82238.1 | SrGuaS | Streptomyces rimosus | ND | Guaia-1(10),11-diene‡, (+)-γ-Gurjunene‡ | ND |
| 13 | ABU58787.1 | RcDTPS | Roseiflexus castenholzii DSM 13941 | ND | ND | 2–3 unidentified compounds‡ |
| 14 | EJL71407.1 | CsDTPS | Chryseobacterium sp. CF314 | ND | ND | 2 unidentified compounds‡ |
| 15 | KFE96946.1 | ClDTPS | Chryseobacterium. Luteum | ND | ND | 2–3 unidentified compounds‡ |
| 16 | CFM47198.1 | BpTPS | Burkholderia pseudomallei | ND | ND | ND |
| 17 | KYF59190.1 | ScTPS1 | Sporangium cellulosum | ND | ND | ND |
| 18 | KFF23441.1 | CvTPS | Chryseobacterium vrystaatense | ND | ND | ND |
| 19 | KJK55818.1 | SsTPS | Saccharothrix sp. ST-888 | ND | ND | ND |
| 20 | KYF75876.1 | ScTPS2 | Sporangium cellulosum | ND | ND | ND |
| 21 | EST32145.1 | SniTPS | Streptomyces niveus NCIMB 11891 | ND | ND | ND |
| 22 | KIZ18902.1 | SnaTPS | Streptomyces natalensis ATCC 27448 | ND | ND | ND |
The products obtained by in vitro assay with purified TSs upon incubation with GPP and FPP or in vivo cultivation in E. coli are listed.
†–produced in in vitro conditions
‡–produced in in vivo conditions
#—produced in in vitro and in vivo conditions; ND–not detected, NT–not tested
Genes encoding selected TS from different bacteria were codon optimised for expression in E. coli (S1 Table), synthesized, and sub-cloned into pETM11 with N-terminal His-tag (GeneArt, Life Technologies). Protein sequence of the selected TSs are shown S2 Table.
Bacterial strains and growth conditions
For in vitro analysis, pre-cultures, LB medium (10 g/L tryptone, 5 g/L yeast extract, 5 g/L NaCl; pH 7.0) were used and cultures were incubated at 37°C for overnight (containing kanamycin at 50 mg/L) in 5 mL. For protein expression, precultures were diluted (1/1000) in 5 mL 2xYT medium (16 g/L Tryptone, 10 g/L Yeast Extract, 5.0 g/L NaCl) and 50 μL was inoculated into Auto Induction Medium Terrific Broth (AIMTB, Formedium) and incubated at 37°C until OD600 = 0.4–0.6 was reached. At this OD, they were cooled to 16°C and induced with 50 μM IPTG and incubated for 18–20 hours. Cultures were centrifuged at 10,000 rpm (JA10 rotor) for 10 minutes and the cell pellets were stored at -20°C until further use [6].
Heterologous expression and protein purification of selected bacterial TSs
One litre of grown cells of E. coli BL21 (DE3) or ArcticExpress (DE3) harbouring a pET-TS plasmid (S3 Table) were defrosted and re-suspended in 10 ml buffer A (binding buffer: 25 mM Tris-HCl, 0.5 M NaCl, 20 mM imidazole, 5 mM MgCl2 and 1 mM Tris (2-chloroethyl) phosphate (TCEP), pH 7.8) and sonicated on ice using a 50% duty cycle at 50% power for 5 mins. Cell debris was removed by centrifugation at 40,000 g for 30 mins at 4°C to separate the soluble protein fraction from the insoluble fraction. His-tagged recombinant proteins were purified by Ni-NTA affinity chromatography (Qiagen). Bound fractions were washed with binding buffer (2 × 10 mL/L culture) and eluted with buffer B (10 mL/L culture; 25 mM Tris-HCl, 0.5 M NaCl, 0.5 M imidazole, 1 mM MgCl2, and 1 mM TCEP, pH 7.8) as described [6]. The obtained fractions were analysed by SDS-PAGE to confirm the purity. The pure fractions were pooled and desalted using a PD-10 column according to the manufacturer’s instructions [34].
FPP and GPP synthesis
Farnesol or geraniol (150 mg) were dissolved in trichloroacetonitrile (0.6 mL) and stirred for 30 mins. Acetonitrile (20 mL) was then added followed by the ammonium phosphorylation salt (0.7 g) which was added in aliquots over 10 mins. The reaction was stirred for 4 hrs and ran a TLC using propanol/concentrated ammonia/water, 6/2/1 solvent system and stain with phosphomolybdic acid (PMA). The organic phase (ether layer) was washed with 1 M aqueous ammonia (3 x 30 mL). The ammonia washes were combined and washed 3 times with fresh diethyl ether (3 x 100 mL). The aqueous layer was reduced to a residue using a rotary evaporator and purified using silica gel. 50 mg of FPP and 50 mg of GPP was synthesized to use for the in vitro enzyme activity analysis.
In vitro enzyme assays, compound extraction
All enzyme reactions were performed with freshly prepared protein. For mono-terpenoid (C10) and sesqui-terpenoid (C15) enzyme assays: 10 μg of purified TS protein was incubated with 100 μM of GPP or 75 μM of FPP in 1 mL of 25 mM Tris-HCl (pH-7.8) with 5 mM MgCl2 (33). A 20% (v/v) organic layer (nonane for mono- and sesqui-terpenoids, n-hexane for di- and sesqui-terpenoids) was added to the reaction mixtures to trap the volatile terpene products, followed by incubation at 28°C for overnight with continuously shaking at 50 rpm. After incubation, the organic layer was removed and the reaction mixture extracted twice with 1 mL of hexane, dried over anhydrous MgSO4 and further concentrated to 100 μL before analysis by gas chromatography—Quadrupole Time-of-Flight Mass Spectrometry (GC-QToF).
In vivo terpenoid production and extraction
Due to GGPP insolubility in the preferred assay buffer, identification of TS activity with GGPP was performed in in vivo conditions. For in vivo diterpenoid production, cells containing pBbA2k-EcispAM22 plasmid with the selected TS (S3 Table) were grown in 2xYT with 0.4% (w/v) glucose until OD600 = 0.6 and induced with 50 nM anhydrous tetracycline (aTc) overlaid with an 20% (v/v) nonane, incubated at 30°C for 36 hours. For in vivo production of mono- and sesqui-terpenoids the cells containing pBbB2a-GPPS-TS and pMVA plasmid were grown in TB media with 0.4% glucose (w/v) and cells were induced with 50 μM of IPTG and 50 nM anhydrous tetracycline (aTc) at OD = 0.6 overlaid with 20% (v/v) nonane, incubated at 30°C for 48 hrs. Nonane layers were harvested and clarified by centrifugation (14,000 rpm, 3 min and 4°C), dried over anhydrous MgSO4 and analysed by GC-QTOF.
Compound GC-MS analyses
The extracted terpenoids were subjected to GC–QToF (Agilent 7020) equipped with an Agilent Technologies 5977A MSD (Mass Selective Detector). The products were separated on HP5 capillary column (30 m, 0.25 mm i. d., 0.50 μm film, Agilent) using a temperature program of 50–280°C with a temperature gradient of 20°C/min and hold for 5 min at 280°C for sesquiterpenoids. For analysis of monoterpenoids, a temperature program of 50–230°C with a temperature gradient of 20°C/min and hold for 2 min at 230°C was used. For diterpenoids analysis a temperature program of 50–300°C with a temperature gradient of 20°C/min and hold for 5 min at 300°C was used. The injector temperature was set at 250°C with a split ratio of 10:1 (1 μL injection). The carrier gas was helium with a flow rate of 1 mL/min and a pressure of 5.1 psi. The ion source temperature of the MS was set to 250°C, and spectra were recorded from m/z 50 to m/z 450 with electron impact mode (70 eV). Compound identification was carried out using authentic standards and comparison to mass spectra to library spectra [35], and NIST (National Institute of Standards and Technology) library of MS spectra and fragmentation patterns, as described previously [36]. Authentic standards 3-carene (115576-25ML), limonene (183164-5ML), linalool (L2602-5G), ocimene isomers (CRM40748), nerolidol (H59605) and geosmin (G5908-1ML) purchased from Sigma-Aldrich were used to validate the compounds produced in in vitro and in vivo conditions.
Ethical statement
As per our knowledge we have provided all the data and followed the ethical guidelines.
Results and discussion
Identification of TS homologues
TSs from different origins show substantial differences in overall primary amino acid sequence, but possess a strongly conserved metal binding domain consisting of an acidic amino acid (AA)-rich motif (D/N) DXX (D/E) or DDXXXE located within 80–120 or 230–270 AA of the N-terminus and an Asn/Ser/Glu triad closer to the C-terminus, which are the signature domains of the class I TS. 2,167 protein sequences out of 73,714 proteins were identified to be TSs and their homologues based on the HMM search as described in the Material and Methods. In a previous study, 262 presumptive bacterial TSs were identified and 27 proteins were functionally characterised in in vivo conditions [37]. Several other bacterial TSs have been characterised and their activities identified with single or multiple substrates [24,28,36,38–41]. In this study, presumptive TS homologue protein sequences were clustered by pairwise similarity, and from the resulting neighbour-joining tree (Fig 1) we identified TSs from Gram-positive bacteria, mainly from the order Actinomycetales, as well as Gram-negative bacteria belonging to numerous orders. We annotated 2,167 TSs in the neighbour-joining tree including geosmin synthases, 2-methylisoborneol synthases and several presumptive TSs whose functionality could not be assigned based on their protein sequences (Fig 1). TSs already identified in the MIBiG database (125 TSs; https://mibig.secondarymetabolites.org) [42] were marked when they had e-values of 0 by BLASTp search against the 2,167 HMM hit proteins. Many bacterial strains in the Proteobacteria and Firmicutes phylum have TS-like genes, while more than 70% contain the phytoene synthase motif. Enzymes containing this motif are mostly involved in phytoene biosynthesis, a tetra-terpene (C40) precursor for lycopene biosynthesis. Geosmin synthases and 2-methylisoborneol synthases from bacteria that have been well studied were excluded and presumptive TSs from Actinobacteria and proteobacteria were selected for functional characterization in this study. In total 22 TSs were selected from different clades (Fig 1) based on one of the following criteria: (i) the sequences contain a single terpene synthase domain (330–350 AA) which belongs to the Isoprenoid Biosynthesis C1 superfamily; (ii) they are from Gram-positive, Gram-negative, thermophilic or thermo-tolerant bacteria; (iii) they are from bacteria that contain other known TSs and have at least 30–40% identity to known TSs but its functionality are not characterized. Functional characterisation of these TSs and identification of their products based on their acyclic allylic diphosphate substrate specificity requires experimental validation which is shown in this work. The 22 potential TSs are selected from the orders of Actinomycetales (Actinobacteria), Burkholderiales (Proteobacteria), Myxococcales (Myxobacteria), Flavobacteriales (Flavobacteria) and Chloroflexales (Terrabacteria). They are: two TSs (AHY47823.1 and AHY45426.1) from the radiation resistant and thermotolerant Rubrobacter radiotolerans [43]; Four TSs (KYF56472.1, CAN96536, KYF59190.1, KYF75876.1) from a prolific secondary metabolite producer Sorangium sps [44]; TS KFG92939.1 from Burkholderia paludis which was isolated from Southeast Pahang tropical peat swamp forest soil in Malaysia [45]; AHH94051.1 from Kutzneria albida DSM 43870; KPH97530 from Actinobacteria bacterium OV450; ADU09376.1 from Micromonospora sp. L5; geoS (CAB41566.1) from Streptomyces coelicolor A3(2); ACY98649.1 from an aerobic, cellulolytic, thermophilic Gram-positive bacterium Thermomonospora curvata [46]; EXG82599.1 from Cryptosporangium arvum DSM 44712; ELQ82238.1 from Streptomyces rimosus; ABU58787.1 from Roseiflexus castenholzii DSM 13941; three hypothetical proteins (KFE96946.1, KFF23441.1 and EJL71407.1) from Chryseobacterium sps [47]; two (EST32145.1 and KIZ18902.1) from soil bacteria belonging to Streptomyces sps; CFM47198.1 from C. vrystaatense; KJK55818.1 from Saccharothrix sp. ST-888 (Table 1, S1 Fig).
In vitro functional characterization of TSs
Codon optimized selected TSs were DNA synthesised and cloned into pETM11 with a TEV protease cleavable N-terminal His-tag by GeneArt (S3 Table). For the heterologous expression of these TSs, the plasmids were transformed into either E. coli Bl21 (DE3) or ArcticExpress (DE3) (S1 Table) and grown in 2xYT media with optimal inducer concentrations to obtain soluble protein. Recombinant proteins were purified using nickel affinity chromatography and subsequently salt and imidazole were removed by a desalting column prior to in vitro activity assays.
Terpenoids resulting from the incubation of the purified recombinant TSs from different bacteria with FPP were extracted using hexane (Fig 2; Table 1), while products formed with GPP were extracted using nonane (Fig 3; Table 1). Products were identified using authentic standards where possible. Purified recombinant geosmin synthase from Streptomyces coelicolor A3(2) [48] and crude extracts with overexpressed limonene synthase from Mentha spicata (LimS) [29] were used as a positive control to validate the in vitro assays for sesquiterpenoids and monoterpenoids, respectively. As expected, purified geosmin synthase yielded the sesquiterpenoids geosmin, germacradienol and germacrene D in the assay mixture upon incubation with FPP (S2 Fig and S3 Fig), whereas the monoterpene limonene was formed when GPP was added to limonene synthase containing crude extracts (S4 Fig and S5 Fig).
Fig 2. GC-QToF analysis of n-hexane extracts from in vitro assays obtained with selected TSs.

GC-MS chromatograms of extracts profile with FPP are shown for A. AHY47823.1 (RrNerS) from Rubrobacter radiotolerans; B. AHY45426 (RrBerS) from R. radiotolerans; C. KYF56472.1 (ScAroS) from Sorangium cellulosum; D. KFG92939 (BpLonS) from Burkholderia paludis; E. AHH94051.1 (KaGerS) from Kutzneria albida DSM 43870; F. KPH97530.1 (AbAcoS) from Actinobacteria bacterium OV450; G.ADU09376.1 (MiGerS) from Micromonospora sp. L5.; H. CAN96536 (ScEpiS) from Sorangium cellulosum. The name and structure of the produced terpenoids are indicated in each trace.
Fig 3. GC-QToF analysis of nonane extracts from in vitro assays obtained upon incubation with GPP for 16 hrs at 28°C.

A. AHY47823.1 (RrNerS) from Rubrobacter radiotolerans; B. AHY45426.1 (RrBerS) from R. radiotolerans and C. 10-epi-cubebol synthase (ScEpiS) from Sorangium cellulosum So ce56. Other small peaks could not be annotated using the NIST library search.
Incubation of a recombinant TS (NCBI accession number AHY47823.1, RrNerS) from the radiation resistant, thermotolerant actinobacterium Rubrobacter radiotolerans [43] with FPP yielded a single product that was identified as trans-nerolidol by GC–QToF analysis (Fig 2A, Table 1) and this was further confirmed using standards (S6 Fig). Incubation with GPP yielded both R- and S- linalool isomers, which were confirmed using standards (Fig 3A and S7 Fig, Table 1). Therefore, this TS was annotated as linalool/nerolidol synthase.
Another terpene cyclase homologue, AHY45426 (RrBerS), from R. radiotolerans also showed activity with both FPP and GPP, yielding α-bergamotene (Fig 2B, S10A Fig, Table 1) and linalool (Fig 3B, S8 Fig, Table 1) respectively.
Therefore, these two TSs are the first to be identified in thermotolerant bacteria, R. radiotolerans, which have both mono- and sesqui-terpene synthases activities and are active up to 60°C (S9 Fig). These thermostable TSs can aid in engineering industrially important TSs to tolerate cultivation at higher temperatures [50].
The terpene cyclase (KYF56472.1, ScAroS) from the soil-dwelling Gram-negative bacterium Sorangium cellulosum converted FPP into only aromadendrene in in vitro analysis (Fig 2C and S10B Fig; Table 1). Aromadendrene is mostly observed in eucalyptus oil and is also produced by a citrus TS, CsSesquiTPS5 [51] and it has been shown to have antibacterial activity to multidrug resistant Gram-negative bacteria [52]. KYF56472.1 was therefore named aromandendrene synthase.
The sesquiterpene cyclase (KFG92939, BpLonS) from Burkholderia paludis was incubated with FPP to yield two sesquiterpenoids, (±)-cadinene and longiborneol (Fig 2D and S10C Fig and S10D Fig; Table 1) and was therefore assigned as longiborneol synthase. Incubation with GPP did not yield any observable products. Longiborneol synthases are mostly found in fungi [53] and Norway spruce [54]. Fusarium uses longiborneol as a precursor for producing the tricyclic mycotoxin culmorin [55]. This enzyme from B. paludis is the first bacterial longiborneol synthase identified.
AHH94051.1 (KaGerS) from Kutzneria albida DSM 43870 has 50% identity to germacradienol/ geosmin synthase (fragment) and we tested its functionality with FPP in in vitro conditions. This yielded germacradienol and germacrene D (Fig 2E and S10E Fig; Table 1) and did not yield any monoterpenes when incubated with GPP. Production of germacradienol was confirmed by comparison to spectra published by Agger et al (2008) [56].
The hypothetical protein 450_5823 (KPH97530.1, AbAcoS) from Actinobacteria bacterium OV450 converted FPP into copaene and acora-3,7(14)-diene (Fig 2F and S10F Fig and S10G Fig; Table 1), while incubation with GPP did not yield any products. KPH97530.1 was annotated as acoradiene synthase based on its product formation.
TS (ADU09376.1, MiGerS) from Micromonospora sp. L5 incubated with FPP yielded β-elemene, and 8-isopropenyl-1,5-dimethyl-1,5-cyclodecadiene (or also called 1(10),4,11-germacratriene) (Fig 2G and S10H Fig and S10I Fig; Table 1). The sesquiterpene hydrocarbon, β-elemene, found is most likely an artefact of GC-MS analysis arising from thermal cope rearrangement of germacrene A at elevated temperature [57–59]. Therefore we assumed this synthase produced germacrene A in addition to germacratriene, and was annotated as germacatriene synthase, as germacatriene was the major compound produced.
Incubation of the sesquiterpene synthase (10-epi-cubebol synthase; CAN96536.1, ScEpiS) from Sorangium cellulosum Soce56 with GPP yielded the monoterpene products cis- and trans-β-ocimene, which differ in the position of the isolated double bond, and linalool which was not shown in previous work (Fig 3C) [49]. All three monoterpene products were confirmed using commercial standards (S11 Fig). When ScEpiS was incubated with FPP the products obtained were 10-epi-α-cubinene, 10-epi-β-cubinene, β-copaene, cadina-3,5-diene, cubebol, γ-cadinene, δ-cadinene, sesquiterpene alcohol 10-epi-cubebol, α-cadinene, 1,10-di-epi-cubenol and cubenol (Fig 2H). These results confirmed previous observations [49] The other minor compounds previously observed: germacrene D, (+)-eremophilene, bicyclogermacrene, T-cadinol, and α-cadinol were not observed due to the solvent extraction method used in this work, instead of sensitive closed loop stripping analysis (CLSA) method used in the previous study. As we have shown in this study, others have observed that heterologous expression of the epi-cubebol synthase from Streptosporangium roseum in E. coli also resulted in the production of β-pinene, β-ocimene (cis- and trans-) and linalool isomers, demonstrating that many sesquiterpene synthases can also accept GPP as substrate [60].
Until now, only two bacterial monoterpene synthases linalool synthase (bLinS) and 1,8-cineole synthases (bCinS) from Streptomyces clavuligerus are well studied [61] and structurally characterized [6]. Broad substrate spectra have been observed previously for bacterial TSs: spata-13, 17-diene synthase is an enzyme with sesqui-, di-, and sester-terpene synthase activity [41], a bLinS from S. clavuligerus accepts GPP and FPP [23], and incubation of corvol ether B synthase (BAJ27126) from Kitasatospora setae with GPP yielded linalool as a major product and small amounts of several acyclic and cyclic monoterpenes [62].
CFM47198.1, ELQ82238.1, ACY98649.1, EXG82599.1, ABU58787.1, EJL71407.1, KFE96946.1 were tested for compound production with FPP and GPP but no terpenoids were detected. (S12 Fig)
In vivo production of mono- and sesqui-terpenoids by enhancing MVA pathway precursor supply
To quantify the in vivo product formation, all identified TSs were cloned into the pBbB2a-GPPS (as described in material and methods) backbone plasmid and co-expressed with pMVA plasmid in E. coli [7] and FPP is supplied via endogenous IspA activity [63]. Cultures were overlaid with nonane to extract the metabolites and analysed by GC-QToF. RrNerS produced monoterpenes β-myrcene, linalool, cis-geraniol; and sesquiterpenes: (±)-trans-nerolidol, α/β-farnesene and farnesol in in vivo conditions (Fig 4A) whereas its homologue RrBerS yielded monoterpenes β-myrcene, linalool, trans-geraniol and trans-α-bergamotene, α/β-farnesene, (±)-trans-nerolidol, farnesol (Fig 4B). MiGerS from Micromonospora sp. L5 expressed in E. coli produced germacrene A and 1,(10),4,11-germacratriene as seen in the in vitro assays (Fig 4C).
Fig 4. GC-QToF analysis of nonane extracts containing mono- and sesqui-terpenes by TSs produced in E. coli.
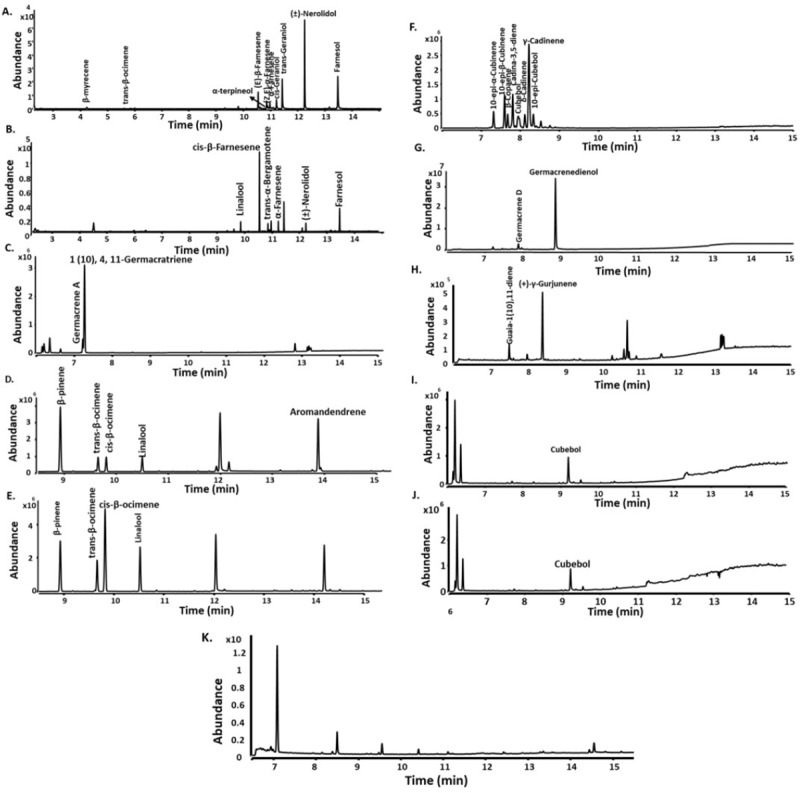
A. AHY47823.1 (RrNerS) from Rubrobacter radiotolerans; B. AHY45426 (RrBerS) from R. radiotolerans; C. ADU09376.1 (MiGerS) from Micromonospora sp. L5.; D. KYF56472.1 (ScAroS) from Sorangium cellulosum; E. monoterpenoids from CAN96536 (ScEpiS) from Sorangium sps.; F. sesquiterpenoids from CAN96536 (ScEpiS) from Sorangium sps.; G. AHH94051.1 (KaGerS) from Kutzneria albida DSM 43870; H. ELQ82238.1 (SrGuaS) from Streptomyces rimosus; I. ACY98649.1 (TcCubS) from Cryptosporangium arvum DSM 44712; J. EXG82599.1 (CaCubS) from Thermomonospora curvata DSM 43183; and K. Negative control (pMVA+pBbB2a-GFP). Peaks were annotated using NIST library spectra.
ScAroS and ScEpiS both from Sorangium, produced similar mono-terpenoids, which are β-pinene, cis- and trans-ocimene, and linalool (Fig 4D and Fig 4E). However they produce different sesquiterpenes in both in vitro and in vivo conditions: aromandendrene is produced by ScAroS whereas 10-epi-α/β-cubinene, β-copaene, cadina-3,5-diene, cubebol, δ/γ-cadinene and 10-epi-cubebol are produced by CAN96536 (Fig 4D and Fig 4F). KaGerS from Kutzneria albida DSM 43870 produced germacrene D and germacrenedienol (Fig 4G).
Overexpression of hypothetical protein ELQ82238.1 (SrGuaS) from Streptomyces rimosus produced multiple sesquiterpenes: guaia-1(10)11-diene and (+)-γ-gurjunene when expressed in E. coli (Fig 4H, S14A Fig and S14B Fig). Whereas ACY98649.1 (TcCubS) from Thermomonospora curvata DSM 43183 (Fig 4I, S14C Fig) and EXG82599.1 (CaCubS) from Cryptosporangium arvum DSM 44712 (Fig 4J, S14D Fig) produced minor amounts of cubebol in E. coli. Terpene synthases may produce different terpene skeletons when expressed natively or in heterologous host compared to their product biosynthesis in in vitro conditions due to variations in protein folding and assay conditions [64].
Geosmin synthase from Streptomyces coelicolor A3(2) when expressed in E. coli produced various monoterpenes: β-myrcene, β-ocimene, linalool and geraniol (S13 Fig) which was unexpected and unexplored for this enzyme as well as the sesquiterpenes: germacradienol, germacrene D but geosmin was not detected, presumably due to inactivity of the C- terminal a-domain in in vivo (S13 Fig).
In vivo production of possible diterpenes
For diterpene synthase activity, the substrate GGPP was synthesized in house but due to its hydrophobic nature the substrate was insoluble in aqueous solution and could not be used in in vitro enzyme assays. For rapid identification of diterpene synthase activity it is essential to generate abundant amounts of GGPP for in vivo production in E. coli. For this purpose, the native farnesyl pyrophosphate synthase (ispA) from E. coli with two mutations D2G, C155G (ispAM22), which can function as geranylgeranyl diphosphate synthase [65], was employed to generate GGPP in vivo. GGPP synthase activity using the variant prenyltransferase was confirmed by overexpression of spata-13, 17-diene synthase from Streptomyces xinghaiensis [41] together with GGPP synthase (ispAM22). This yielded the diterpene spata-13,17-diene, where the produced compound was extracted using a nonane layer (S15 Fig). ABU58787.1 (RcDTPS) from Roseiflexus castenholzii DSM 13941 (Fig 5A, S16 Fig), EJL71407.1 (CsDTPS) from Chryseobacterium sp. CF314 (Fig 5B, S17 Fig, and KFE96946 (ClDTPS) from Chryseobacterium luteum (Fig 5C and S18 Fig) produced various possible diterpene products when expressed together with ispAM22 in E. coli DH5α. Due to low product yields, the detected compounds could not be annotated by a NIST library search alone. By co-expressing the heterologous MVA pathway, the precursor supply for diterpene, production was enhanced which led to a 10-fold increased production. However, this was still not enough to determine the compounds produced. Large scale production and purification of the diterpenes would be required for structural determination by NMR analysis.
Fig 5. GC-QToF analysis of nonane extracts containing possible diterpenes from E. coli in vivo production expressing the following TSs.

A. ABU58787.1 from Roseiflexus castenholzii DSM 13941 (RcDTPS), B. EJL71407.1 from Chryseobacterium sp. CF314 (CsDTPS), and C. KFE96946 from Chryseobacterium luteum (ClDTPS). Numbered peaks in each Fig correspond to compounds produced. The mass spectrum for each compound is in S16–S18 Figs.
In this study, we identified 15 putative bacterial TSs out of 22 that were tested, which produced structurally diverse mono-, sesqui- and di-terpenoids by in vitro and/or in vivo assays. We have shown TSs with new activity: 7 sesqui-terpenoid synthases, 3 di-terpenoid synthases and 5 mono-/sesqui-terpenoid synthases which were identified from Gram-positive, Gram-negative and thermophilic tolerant bacterial species. Many sesquiterpene synthases were shown to also have activity as a monoterpene synthase, which suggests that dual substrate specificity is very common for bacterial TSs.
In vivo production of all types of terpenoids by E. coli makes an attractive platform for rapid identification of enzymes as well as for better and cheaper production yields through metabolic engineering. Especially the newly discovered TSs from the thermophilic/tolerant bacterial species is very promising for further protein evolution studies to design the end terpene product, which we have started [50] and to exploit the enzyme stability at higher temperatures. In addition, production of diterpenoids in E. coli as an alternative screening method to in vitro assays is useful due to the difficulties with solubility of GGPP.
The results presented in this study suggest that further exploration of putative TSs from different bacteria along with Actinomycetes could expand the structural plethora of terpenes. The majority of the terpene products identified in this study are known to be produced by plant or fungi and reveals that TSs are widely distributed in bacteria. Given the number of uncharacterized bacterial enzymes that exist in nature, it is likely that there remains a wealth of chemistry to be discovered and exploited. Expanding the search for novel terpenoid biosynthesis will provide numerous structures with unexplored properties that could potentially help to develop novel compounds for pharmacological or industrial applications.
Supporting information
E. coli, DH5a cells were used for plasmid propagation and Bl21 (DE3) and Arctic Express (DE3) cells for recombinant protein expression.
(DOCX)
(DOCX)
(DOCX)
Known TSs annotated in the tree in Fig 1 where the branches were indicated in colour based on their functionality: monoterpenoids (light blue), 2- methyisoborneol (orange), sesquiterpenoids (purple) and diterpenoids (blue). The same colour scheme is adapted here.
(DOCX)
Neighbour-joining tree constructed from the amino acid sequences of characterized terpene synthases in this study (underlined) and in previous studies. Geosmin synthases (dark red), 2-methylisoborneol synthases (orange), monoterpene (light blue), diterpene (dark blue) and sesquiterpene synthases (purple) are shown and uncharacterized bacterial terpene synthases in this study (in red).
(DOCX)
GC-MS traces showing the separation of geosmin (0.1 mg mL-1) on a HP5 column and its retention time at 7.5 minutes. B. Mass spectra of geosmin.
(DOCX)
GC-MS chromatogram of geosmin synthase with 75 μM of FPP. Peak 1: geosmin (rt: 7.5), peak 2: Germacrene D (rt: 7.86), peak 3: germacradienol (rt: 8.84). B. Mass spectra of compounds observed in the extracts.
(DOCX)
A. GC-MS traces showing the separation of limonene (0.1 mg mL-1) produced in this study on a HP5 column. The internal standard, sec-butyl benzene (0.1%, v/v), has a retention time of 9.5 minutes. Peak 1: R- Limonene (rt: 9.41). B: Mass spectra of limonene.
(DOCX)
A. GC chromatogram of 10 μg of limonene synthase (pJBEI-6410) crude extracts with 75 μM of GPP. Peak 1: limonene (rt: 9.41). B. Mass spectra of limonene.
(DOCX)
A. GC-MS traces showing the separation of standard nerolidol mix (0.1 mg mL-1) on a HP5 column. B. MS spectra of -cis and -trans Nerolidol. C. GC-MS chromatogram of trans-nerolidol produced by AHY47823 with FPP. D. Mass spectra for trans-nerolidol produced by AHY47823.
(DOCX)
A. GC-MS traces showing the separation of standard linalool mix (0.1 mg mL-1) on a CP-Chirasil-DEX-CB column. B. GC-MS chromatogram of linalool isomers produced by AHY47823 with FPP. Samples were analyzed by GC on an Agilent Technologies 7890A GC system equipped with an FID detector, a 7693 auto sampler, and a CP-Chirasil-DEX-CB column (25 m × 0.25 mm i.d., 0.25 μm film thickness). For linalool, the program initiated at a temperature of 70°C which was then increased to 90°C at 8°C/min. This was followed by an increase in temperature to 150°C at a rate of 2°C/min and then to 190°C at 40°C/min (1 min hold). The FID detector was maintained at a temperature of 200°C with a flow of hydrogen at 30 mL/min.
(DOCX)
A. GC-MS traces showing the separation of standard linalool mix (0.1 mg mL-1) on a CP-Chirasil-DEX-CB column. B. GC-MS chromatogram of linalool isomers produced by AHY45426 with FPP. Samples were analyzed by gas chromatography on an Agilent Technologies 7890A GC system equipped with an FID detector, a 7693 auto sampler, and a CP-Chirasil-DEX-CB column (25 m × 0.25 mm i.d., 0.25 μm film thickness). For linalool, the program initiated at a temperature of 70°C which was then increased to 90°C at 8°C/min. This was followed by an increase in temperature to 150°C at a rate of 2°C/min and then to 190°C at 40°C/min (1 min hold). The FID detector was maintained at a temperature of 200°C with a flow of hydrogen at 30 mL/min.
(DOCX)
Total ion chromatograms of products obtained from an incubation RrNerS (left) and RrBerS (right) with FPP for 3 hours at variable temperatures. Samples were analyzed by GC-QToF on HP5 column.
(DOCX)
The reference spectra from the NIST library shown in blue and the compound spectra shown in red.A. α–bergamotene produced by RrBerS, B. Aromandendrene produced by ScAroS, C and D. cadinene and longiborneol produced by BpLonS, E. germacrene D produced byKaGerS, F and G. copaene and acora-3,7(14)–diene produced byAbAcoS, and H and I. β-elemene and 1(10),4,7(11)-germacra-triene produced by MiGerS.
(DOCX)
A. GC-MS traces showing the separation of monoterpenoids (0.1 mg mL-1) standards used in this study on a HP5 column. The internal standard used, sec-butyl benzene (St; 0.1% v/v), has a retention time of 9 minutes. Peak 1: cis-ocimene (rt: 9.56), peak 2: trans-ocimene (rt: 9.72), 3: linalool (rt: 10.43). B. Comparison of obtained mass spectra with NIST Library spectra and standard as shown in A. of products yielded by CAN96536 with GPP in in vitro assay.
(DOCX)
GC-MS chromatograms of extracts profile with FPP are shown for A. BpTPS; B. ScTPS1; C. CvTPS; D. SsTPS; E. ScTPS2; F. SniTPS; G.SnaTPS. The peaks in the chromatogram do not correspond to any terpenoids.
(DOCX)
A. GC-MS chromatogram for geosmin synthase products, B. mass spectra for Germacradienol, C—G. Comparison of obtained mass spectra with NIST Library spectra of products yielded by GeoS.
(DOCX)
A–B. SrGuaS C. TcCubS D. CaCubS.
(DOCX)
A. spata-13,17-diene produced by overexpression of ispAM22 and WP_095757924 in E. coli. B. Mass spectra of produced spata-13,17-diene.
(DOCX)
(DOCX)
(DOCX)
(DOCX)
Acknowledgments
We thank Mrs. Rehana Sung and Dr. Katherine Hollywood for technical assistance with product analysis.
Data Availability
All relevant data are within the manuscript and its Supporting Information files.
Funding Statement
RB, NGHL, GKR, NS, ET was funded by the UK Biotechnology and Biological Sciences Research Council (BBSRC) BB/M000354/1. NGHL was funded as a Research Fellow of the EPSRC/BBSRC Future Biomanufacturing Research Hub (EP/S01778X/1). N.S.S. was funded as an EPSRC Established Career Fellow (EP/J020192/1). RB, NS, ET was funded BBSRC and Engineering and Physical Sciences Research Council (EPSRC) (BB/M017702/1).
References
- 1.Can Başer KH, Buchbauer G. Handbook of essential oils: Science, technology, and applications, second edition. Handbook of Essential Oils: Science, Technology, and Applications, Second Edition. 2015. 10.1201/b19393 [DOI]
- 2.Martin VJJ, Piteral DJ, Withers ST, Newman JD, Keasling JD. Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat Biotechnol. 2003;21: 796–802. 10.1038/nbt833 [DOI] [PubMed] [Google Scholar]
- 3.Peralta-Yahya PP, Ouellet M, Chan R, Mukhopadhyay A, Keasling JD, Lee TS. Identification and microbial production of a terpene-based advanced biofuel. Nat Commun. 2011;2: 483 10.1038/ncomms1494 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Paddon CJ, Westfall PJ, Pitera DJ, Benjamin K, Fisher K, McPhee D, et al. High-level semi-synthetic production of the potent antimalarial artemisinin. Nature. 2013;496: 528–532. 10.1038/nature12051 [DOI] [PubMed] [Google Scholar]
- 5.Currin A, Dunstan MS, Johannissen LO, Hollywood KA, Vinaixa M, Jervis AJ, et al. Engineering the “Missing Link” in Biosynthetic (−)-Menthol Production: Bacterial Isopulegone Isomerase. ACS Catal. 2018;8: 2012–2020. 10.1021/acscatal.7b04115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Karuppiah V, Ranaghan KE, Leferink NGH, Johannissen LO, Shanmugam M, Ní Cheallaigh A, et al. Structural Basis of Catalysis in the Bacterial Monoterpene Synthases Linalool Synthase and 1,8-Cineole Synthase. ACS Catal. 2017;7: 6268–6282. 10.1021/acscatal.7b01924 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Leferink NGH, Jervis AJ, Zebec Z, Toogood HS, Hay S, Takano E, et al. A ‘Plug and Play’ Platform for the Production of Diverse Monoterpene Hydrocarbon Scaffolds in Escherichia coli. ChemistrySelect. 2016;1: 1893–1896. 10.1002/slct.201600563 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cane DE. Enzymatic Formation of Sesquiterpenes. Chem Rev. 1990;90: 1089–1103. 10.1021/cr00105a002 [DOI] [Google Scholar]
- 9.Chow J-Y, Tian B-X, Ramamoorthy G, Hillerich BS, Seidel RD, Almo SC, et al. Computational-guided discovery and characterization of a sesquiterpene synthase from Streptomyces clavuligerus. Proc Natl Acad Sci. 2015;112: 5661–5666. 10.1073/pnas.1505127112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Tian B, Poulter CD, Jacobson MP. Defining the Product Chemical Space of Monoterpenoid Synthases. Fetrow JS, editor. PLoS Comput Biol. 2016;12: e1005053 10.1371/journal.pcbi.1005053 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Christianson DW. Structural and Chemical Biology of Terpenoid Cyclases. Chem Rev. 2017;117: 11570–11648. 10.1021/acs.chemrev.7b00287 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Karunanithi PS, Zerbe P. Terpene Synthases as Metabolic Gatekeepers in the Evolution of Plant Terpenoid Chemical Diversity. Front Plant Sci. 2019;10: 1166 10.3389/fpls.2019.01166 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Li G, Köllner TG, Yin Y, Jiang Y, Chen H, Xu Y, et al. Nonseed plant Selaginella moellendorffii has both seed plant and microbial types of terpene synthases. Proc Natl Acad Sci U S A. 2012. 10.1073/pnas.1204300109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jia Q, Li G, Köllner TG, Fu J, Chen X, Xiong W, et al. Microbial-type terpene synthase genes occur widely in nonseed land plants, but not in seed plants. Proc Natl Acad Sci U S A. 2016. 10.1073/pnas.1607973113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gerber NN, Lechevalier HA. Geosmin, an earthly-smelling substance isolated from actinomycetes. Appl Microbiol. 1965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.GERBER NN. A Volatile Metabolite of Actinomycetes, 2-Methylisoborneol. J Antibiot (Tokyo). 1969;22: 508–509. 10.7164/antibiotics.22.508 [DOI] [PubMed] [Google Scholar]
- 17.Jiang J, He X, Cane DE. Biosynthesis of the earthy odorant geosmin by a bifunctional Streptomyces coelicolor enzyme. Nat Chem Biol. 2007;3: 711–715. 10.1038/nchembio.2007.29 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Koe BK, SOBIN BA, CELMER WD. PA 132, a new antibiotic. I. Isolation and chemical properties. Antibiot Annu. 1957; 672–5. Available: http://www.ncbi.nlm.nih.gov/pubmed/13425447 [PubMed]
- 19.Martin DG, Slomp G, Mizsak S, Duchamp DJ, Chidester CG. The structure and absolute configuration of pentalenolactone (PA 132). Tetrahedron Lett. 1970;11: 4901–4904. 10.1016/S0040-4039(00)99739-9 [DOI] [PubMed] [Google Scholar]
- 20.Ogawa Y, Yonehara H. The structure of pentalenolactone (PA-132)Setsuo Takeuchi. Tetrahedron Lett. 1969;10: 2737–2740. 10.1016/S0040-4039(01)88256-3 [DOI] [PubMed] [Google Scholar]
- 21.Cane DE, Sohng J-K, Lamberson CR, Rudnicki SM, Wu Z, Lloyd MD, et al. Pentalenene Synthase. Purification, Molecular Cloning, Sequencing, and High-Level Expression in Escherichia coli of a Terpenoid Cyclase from Streptomyces UC5319. Biochemistry. 1994;33: 5846–5857. 10.1021/bi00185a024 [DOI] [PubMed] [Google Scholar]
- 22.Dickschat JS. Bacterial terpene cyclases. Nat Prod Rep. 2016;33: 87–110. 10.1039/c5np00102a [DOI] [PubMed] [Google Scholar]
- 23.Nakano C, Kim HK, Ohnishi Y. Identification of the first bacterial monoterpene cyclase, a 1,8-cineole synthase, that catalyzes the direct conversion of geranyl diphosphate. ChemBioChem. 2011;12: 1988–1991. 10.1002/cbic.201100330 [DOI] [PubMed] [Google Scholar]
- 24.Yamada Y, Kuzuyama T, Komatsu M, Shin-Ya K, Omura S, Cane DE, et al. Terpene synthases are widely distributed in bacteria. Proc Natl Acad Sci. 2015;112: 857–862. 10.1073/pnas.1422108112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Komatsu M, Tsuda M, Omura S, Oikawa H, Ikeda H. Identification and functional analysis of genes controlling biosynthesis of 2-methylisoborneol. Proc Natl Acad Sci. 2008/05/20. 2008;105: 7422–7427. 10.1073/pnas.0802312105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Cane DE, Ikeda H. Exploration and mining of the bacterial terpenome. Acc Chem Res. 2012;45: 463–472. 10.1021/ar200198d [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Minami A, Ozaki T, Liu C, Oikawa H. Cyclopentane-forming di/sesterterpene synthases: Widely distributed enzymes in bacteria, fungi, and plants. Nat Prod Rep. 2018;35: 1330–1346. 10.1039/c8np00026c [DOI] [PubMed] [Google Scholar]
- 28.Rinkel J, Lauterbach L, Dickschat JS. A Branched Diterpene Cascade: The Mechanism of Spinodiene Synthase from Saccharopolyspora spinosa. Angew Chemie—Int Ed. 2019;58: 452–455. 10.1002/anie.201812216 [DOI] [PubMed] [Google Scholar]
- 29.Alonso-gutierrez J, Chan R, Batth TS, Adams PD, Keasling JD, Petzold CJ, et al. Metabolic engineering of Escherichia coli for limonene and perillyl alcohol production. Metab Eng. 2013;19: 33–41. 10.1016/j.ymben.2013.05.004 [DOI] [PubMed] [Google Scholar]
- 30.Blin K, Medema MH, Kazempour D, Fischbach MA, Breitling R, Takano E, et al. antiSMASH 2.0—a versatile platform for genome mining of secondary metabolite producers. Nucleic Acids Res. 2013;41: W204–12. 10.1093/nar/gkt449 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. 2013;30: 772–80. 10.1093/molbev/mst010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Letunic I, Bork P. Interactive Tree Of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics. 2007;23: 127–128. 10.1093/bioinformatics/btl529 [DOI] [PubMed] [Google Scholar]
- 33.Gibson DG, Young L, Chuang RY, Venter JC, Hutchison CA, Smith HO. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat Methods. 2009;6: 343–345. 10.1038/nmeth.1318 [DOI] [PubMed] [Google Scholar]
- 34.GE Healthcare. Pd10 Desalting Column. GE Healthc Bio-Sciences. 2007.
- 35.Adams RP. Identification of Essential Components by Gas Chromatography/Mass Spectroscopy. Allured Corp Carol Stream, IL, USA. 2009. 10.1163/_q3_SIM_00374 [DOI]
- 36.Rabe P, Dickschat JS. Rapid chemical characterization of bacterial terpene synthases. Angew Chemie—Int Ed. 2013;52: 1810–1812. 10.1002/anie.201209103 [DOI] [PubMed] [Google Scholar]
- 37.Yamada Y, Arima S, Nagamitsu T, Johmoto K, Uekusa H, Eguchi T, et al. Novel terpenes generated by heterologous expression of bacterial terpene synthase genes in an engineered Streptomyces host. J Antibiot (Tokyo). 2015;68: 385–394. 10.1038/ja.2014.171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Baer P, Rabe P, Citron CA, de Oliveira Mann CC, Kaufmann N, Groll M, et al. Hedycaryol Synthase in Complex with Nerolidol Reveals Terpene Cyclase Mechanism. ChemBioChem. 2014;15: 213–216. 10.1002/cbic.201300708 [DOI] [PubMed] [Google Scholar]
- 39.Dickschat JS, Rinkel J, Rabe P, Kashkooli AB, Bouwmeester HJ. 18-Hydroxydolabella-3,7-diene synthase—A diterpene synthase from Chitinophaga pinensis. Beilstein J Org Chem. 2017;13: 1770–1780. 10.3762/bjoc.13.171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rabe P, Schmitz T, Dickschat JS. Mechanistic investigations on six bacterial terpene cyclases. Beilstein J Org Chem. 2016;12: 1839–1850. 10.3762/bjoc.12.173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Rinkel J, Lauterbach L, Dickschat JS. Spata-13,17-diene Synthase—An Enzyme with Sesqui-, Di-, and Sesterterpene Synthase Activity from Streptomyces xinghaiensis. Angew Chemie—Int Ed. 2017;56: 16385–16389. 10.1002/anie.201711142 [DOI] [PubMed] [Google Scholar]
- 42.Medema MH, Kottmann R, Yilmaz P, Cummings M, Biggins JB, Blin K, et al. Minimum Information about a Biosynthetic Gene cluster. Nature Chemical Biology. 2015. pp. 625–631. 10.1038/nchembio.1890 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Egas C, Barroso C, Froufe HJC, Pacheco J, Albuquerque L, da Costa MS. Complete genome sequence of the radiation-resistant bacterium Rubrobacter radiotolerans RSPS-4. Stand Genomic Sci. 2015;9: 1062–1075. 10.4056/sigs.5661021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Pradella S, Hans A, Spröer C, Reichenbach H, Gerth K, Beyer S. Characterisation, genome size and genetic manipulation of the myxobacterium Sorangium cellulosum So ce56. Arch Microbiol. 2002;178: 484–492. 10.1007/s00203-002-0479-2 [DOI] [PubMed] [Google Scholar]
- 45.Ong KS, Aw YK, Lee LH, Yule CM, Cheow YL, Lee SM. Burkholderia paludis sp. nov., an antibiotic-siderophore producing novel Burkholderia cepacia complex species, isolated from malaysian tropical peat swamp soil. Front Microbiol. 2016;7: 2046 10.3389/fmicb.2016.02046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Chertkov O, Sikorski J, Nolan M, Lapidus A, Lucas S, Del Rio TG, et al. Complete genome sequence of Thermomonospora curvata type strain (B9). Stand Genomic Sci. 2011;4: 13–22. 10.4056/sigs.1453580 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Behrendt U, Ulrich A, Spröer C, Schumann P. Chryseobacterium luteum sp. nov., associated with the phyllosphere of grasses. Int J Syst Evol Microbiol. 2007. 10.1099/ijs.0.65104-0 [DOI] [PubMed] [Google Scholar]
- 48.Harris GG, Lombardi PM, Pemberton TA, Matsui T, Weiss TM, Cole KE, et al. Structural Studies of Geosmin Synthase, a Bifunctional Sesquiterpene Synthase with αα Domain Architecture That Catalyzes a Unique Cyclization-Fragmentation Reaction Sequence. Biochemistry. 2015;54: 7142–55. 10.1021/acs.biochem.5b01143 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Schifrin A, Khatri Y, Kirsch P, Thiel V, Schulz S, Bernhardt R. A single terpene synthase is responsible for a wide variety of sesquiterpenes in Sorangium cellulosum Soce56. Org Biomol Chem. 2016;14: 3385–3393. 10.1039/c6ob00130k [DOI] [PubMed] [Google Scholar]
- 50.Styles MQ, Nesbitt EA, Marr S, Hutchby M, Leak DJ. Characterization of the first naturally thermostable terpene synthases and development of strategies to improve thermostability in this family of enzymes. FEBS J. 2017;284: 1700–1711. 10.1111/febs.14072 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Alquézar B, Rodríguez A, de la Peña M, Peña L. Genomic analysis of terpene synthase family and functional characterization of seven sesquiterpene synthases from citrus sinensis. Front Plant Sci. 2017;8 10.3389/fpls.2017.01481 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Mulyaningsih S, Sporer F, Zimmermann S, Reichling J, Wink M. Synergistic properties of the terpenoids aromadendrene and 1,8-cineole from the essential oil of eucalyptus globulus against antibiotic-susceptible and antibiotic-resistant pathogens. Phytomedicine. 2010;17: 1061–1066. 10.1016/j.phymed.2010.06.018 [DOI] [PubMed] [Google Scholar]
- 53.McCormick SP, Alexander NJ, Harris LJ. CLM1 of fusarium graminearum encodes a longiborneol synthase required for culmorin production. Appl Environ Microbiol. 2010;76: 136–141. 10.1128/AEM.02017-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Martin DM, Fäldt J, Bohlmann J. Functional characterization of nine Norway spruce TPS genes and evolution of gymnosperm terpene synthases of the TPS-d subfamily. Plant Physiol. 2004;135: 1908–1927. 10.1104/pp.104.042028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Yuan Y, Litzenburger M, Cheng S, Bian G, Hu B, Yan P, et al. Sesquiterpenoids Produced by Combining Two Sesquiterpene Cyclases with Promiscuous Myxobacterial CYP260B1. ChemBioChem. 2019;20: 677–682. 10.1002/cbic.201800670 [DOI] [PubMed] [Google Scholar]
- 56.Agger SA, Lopez-Gallego F, Hoye TR, Schmidt-Dannert C. Identification of sesquiterpene synthases from Nostoc punctiforme PCC 73102 and Nostoc sp. strain PCC 7120. J Bacteriol. 2008;190: 6084–6096. 10.1128/JB.00759-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Bowers WS, Nishino C, Montgomery ME, Nault LR, Nielson MW. Sesquiterpene progenitor, germacrene a: An alarm pheromone in aphids. Science (80-). 1977;196: 680–681. 10.1126/science.558651 [DOI] [PubMed] [Google Scholar]
- 58.Rinkel J, Dickschat JS. Addressing the chemistry of germacrene A by isotope labeling experiments. Org Lett. 2019;21: 2426–2429. 10.1021/acs.orglett.9b00725 [DOI] [PubMed] [Google Scholar]
- 59.Weinheimer AJ, Youngblood WW, Washecheck PH, Karns TKB, Ciereszko LS. Isolation of the elusive (-)-germacrene-a from the gorgonian, eunicea mammosa chemistry of coelenterates. XVIII. Tetrahedron Lett. 1970;11: 497–500. 10.1016/0040-4039(70)89009-8 [DOI] [Google Scholar]
- 60.Dickschat JS, Pahirulzaman KAK, Rabe P, Klapschinski TA. An Improved technique for the rapid chemical characterisation of bacterial terpene cyclases. ChemBioChem. 2014;15: 810–814. 10.1002/cbic.201300763 [DOI] [PubMed] [Google Scholar]
- 61.Nakano C, Kim HK, Ohnishi Y. Identification and Characterization of the linalool/nerolidol Synthase from Streptomyces clavuligerus. ChemBioChem. 2011;12: 2403–2407. 10.1002/cbic.201100501 [DOI] [PubMed] [Google Scholar]
- 62.Rabe P, Pahirulzaman KAK, Dickschat JS. Structures and biosynthesis of corvol ethers—Sesquiterpenes from the actinomycete kitasatospora setae. Angew Chemie—Int Ed. 2015. 10.1002/anie.201501119 [DOI] [PubMed] [Google Scholar]
- 63.Fujisaki S, Hara H, Nishimura Y, Horiuchi K, Nishino T. Cloning and nucleotide sequence of the ispA gene responsible for farnesyl diphosphate synthase activity in Escherichia coli1. J Biochem. 1990. 10.1093/oxfordjournals.jbchem.a123327 [DOI] [PubMed] [Google Scholar]
- 64.Helfrich EJN, Lin G-M, Voigt CA, Clardy J. Bacterial terpene biosynthesis: challenges and opportunities for pathway engineering. Beilstein J Org Chem. 2019;15: 2889–2906. 10.3762/bjoc.15.283 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lee PC, Petri R, Mijts BN, Watts KT, Schmidt-Dannert C. Directed evolution of Escherichia coli farnesyl diphosphate synthase (IspA) reveals novel structural determinants of chain length specificity. Metab Eng. 2005;7: 18–26. 10.1016/j.ymben.2004.05.003 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
E. coli, DH5a cells were used for plasmid propagation and Bl21 (DE3) and Arctic Express (DE3) cells for recombinant protein expression.
(DOCX)
(DOCX)
(DOCX)
Known TSs annotated in the tree in Fig 1 where the branches were indicated in colour based on their functionality: monoterpenoids (light blue), 2- methyisoborneol (orange), sesquiterpenoids (purple) and diterpenoids (blue). The same colour scheme is adapted here.
(DOCX)
Neighbour-joining tree constructed from the amino acid sequences of characterized terpene synthases in this study (underlined) and in previous studies. Geosmin synthases (dark red), 2-methylisoborneol synthases (orange), monoterpene (light blue), diterpene (dark blue) and sesquiterpene synthases (purple) are shown and uncharacterized bacterial terpene synthases in this study (in red).
(DOCX)
GC-MS traces showing the separation of geosmin (0.1 mg mL-1) on a HP5 column and its retention time at 7.5 minutes. B. Mass spectra of geosmin.
(DOCX)
GC-MS chromatogram of geosmin synthase with 75 μM of FPP. Peak 1: geosmin (rt: 7.5), peak 2: Germacrene D (rt: 7.86), peak 3: germacradienol (rt: 8.84). B. Mass spectra of compounds observed in the extracts.
(DOCX)
A. GC-MS traces showing the separation of limonene (0.1 mg mL-1) produced in this study on a HP5 column. The internal standard, sec-butyl benzene (0.1%, v/v), has a retention time of 9.5 minutes. Peak 1: R- Limonene (rt: 9.41). B: Mass spectra of limonene.
(DOCX)
A. GC chromatogram of 10 μg of limonene synthase (pJBEI-6410) crude extracts with 75 μM of GPP. Peak 1: limonene (rt: 9.41). B. Mass spectra of limonene.
(DOCX)
A. GC-MS traces showing the separation of standard nerolidol mix (0.1 mg mL-1) on a HP5 column. B. MS spectra of -cis and -trans Nerolidol. C. GC-MS chromatogram of trans-nerolidol produced by AHY47823 with FPP. D. Mass spectra for trans-nerolidol produced by AHY47823.
(DOCX)
A. GC-MS traces showing the separation of standard linalool mix (0.1 mg mL-1) on a CP-Chirasil-DEX-CB column. B. GC-MS chromatogram of linalool isomers produced by AHY47823 with FPP. Samples were analyzed by GC on an Agilent Technologies 7890A GC system equipped with an FID detector, a 7693 auto sampler, and a CP-Chirasil-DEX-CB column (25 m × 0.25 mm i.d., 0.25 μm film thickness). For linalool, the program initiated at a temperature of 70°C which was then increased to 90°C at 8°C/min. This was followed by an increase in temperature to 150°C at a rate of 2°C/min and then to 190°C at 40°C/min (1 min hold). The FID detector was maintained at a temperature of 200°C with a flow of hydrogen at 30 mL/min.
(DOCX)
A. GC-MS traces showing the separation of standard linalool mix (0.1 mg mL-1) on a CP-Chirasil-DEX-CB column. B. GC-MS chromatogram of linalool isomers produced by AHY45426 with FPP. Samples were analyzed by gas chromatography on an Agilent Technologies 7890A GC system equipped with an FID detector, a 7693 auto sampler, and a CP-Chirasil-DEX-CB column (25 m × 0.25 mm i.d., 0.25 μm film thickness). For linalool, the program initiated at a temperature of 70°C which was then increased to 90°C at 8°C/min. This was followed by an increase in temperature to 150°C at a rate of 2°C/min and then to 190°C at 40°C/min (1 min hold). The FID detector was maintained at a temperature of 200°C with a flow of hydrogen at 30 mL/min.
(DOCX)
Total ion chromatograms of products obtained from an incubation RrNerS (left) and RrBerS (right) with FPP for 3 hours at variable temperatures. Samples were analyzed by GC-QToF on HP5 column.
(DOCX)
The reference spectra from the NIST library shown in blue and the compound spectra shown in red.A. α–bergamotene produced by RrBerS, B. Aromandendrene produced by ScAroS, C and D. cadinene and longiborneol produced by BpLonS, E. germacrene D produced byKaGerS, F and G. copaene and acora-3,7(14)–diene produced byAbAcoS, and H and I. β-elemene and 1(10),4,7(11)-germacra-triene produced by MiGerS.
(DOCX)
A. GC-MS traces showing the separation of monoterpenoids (0.1 mg mL-1) standards used in this study on a HP5 column. The internal standard used, sec-butyl benzene (St; 0.1% v/v), has a retention time of 9 minutes. Peak 1: cis-ocimene (rt: 9.56), peak 2: trans-ocimene (rt: 9.72), 3: linalool (rt: 10.43). B. Comparison of obtained mass spectra with NIST Library spectra and standard as shown in A. of products yielded by CAN96536 with GPP in in vitro assay.
(DOCX)
GC-MS chromatograms of extracts profile with FPP are shown for A. BpTPS; B. ScTPS1; C. CvTPS; D. SsTPS; E. ScTPS2; F. SniTPS; G.SnaTPS. The peaks in the chromatogram do not correspond to any terpenoids.
(DOCX)
A. GC-MS chromatogram for geosmin synthase products, B. mass spectra for Germacradienol, C—G. Comparison of obtained mass spectra with NIST Library spectra of products yielded by GeoS.
(DOCX)
A–B. SrGuaS C. TcCubS D. CaCubS.
(DOCX)
A. spata-13,17-diene produced by overexpression of ispAM22 and WP_095757924 in E. coli. B. Mass spectra of produced spata-13,17-diene.
(DOCX)
(DOCX)
(DOCX)
(DOCX)
Data Availability Statement
All relevant data are within the manuscript and its Supporting Information files.