

Abstract
Background.
Social aggression is a form of antisocial behavior in which social relationships and social status are used to damage reputations and inflict emotional harm on others. Despite extensive research examining the prevalence and consequences of social aggression, only a few studies have examined its genetic-environmental etiology, with markedly inconsistent results.
Method.
We estimated the etiology of social aggression using the nuclear twin family (NTF) model. Maternal-report, paternal-report, and teacher-report data were collected for twin social aggression (N=1,030 pairs). We also examined the data using the classical twin model (CT) to evaluate whether its strict assumptions may have biased previous heritability estimates.
Results.
The best-fitting NTF model for all informants was the ASFE model, indicating that additive genetic, sibling environmental, familial environmental, and non-shared environmental influences significantly contribute to the etiology of social aggression in middle childhood. However, the best-fitting CT model varied across informants, ranging from AE and ACE to CE. Specific heritability estimates for both NTF and CT models also varied across informants such that teacher reports indicated greater genetic influences and father reports indicated greater shared environmental influences.
Conclusions.
Although the specific NTF parameter estimates varied across informants, social aggression generally emerged as largely additive genetic (A=0.15–0.77) and sibling environmental (S=0.42–0.72) in origin. Such findings not only highlight an important role for individual genetic risk in the etiology of social aggression, but also raise important questions regarding the role of the environment.
Keywords: Social aggression, etiology, behavior genetics, nuclear twin family, relational aggression, twins
Social aggression is a form of antisocial behavior (Burt et al., 2012) in which social relationships and social status are used to damage reputations and inflict emotional harm on others, and centers on behaviors such as gossiping, ostracism, and threatening to end a friendship. Although socially aggressive behaviors are legal and relatively typical during particular developmental periods, they have been associated with pathological outcomes in both victims and aggressors. Victims of social aggression often experience as much emotional distress as victims of physical aggression, including emotional and social difficulties such as peer rejection, loneliness, and internalizing problems (Crick et al., 2002). The perpetrators of social aggression also exhibit a number of maladaptive outcomes, including poor quality friendships marked by conflict and instability, depression, externalizing behavior, and borderline personality disorder features. Not surprisingly then, there is substantial interest in developing prevention and intervention programs that target social aggression. To date, however, there is relatively little work available on its underlying etiology to inform these efforts.
Extant research regarding the origins of social aggression primarily focuses on social processes and psychosocial risk factors, and particularly on peer relationships (e.g. popularity, victimization, quality of dyadic relationships; Crick and Grotpeter, 1995, Sijtsema et al., 2010) and parenting factors (e.g. psychological control, coercive parenting; Kawabata et al., 2011; Kuppens et al., 2013). Other work has focused on marital conflict, sibling relationships, media exposure, and cultural values (e.g. Forbes et al., 2009; Gentile et al., 2011; Karriker-Jaffe et al., 2013). Relatively few of these studies, however, have been designed to examine both environmental and genetic risk factors simultaneously. This is an important omission in the literature, as it means that studies have thus far been unable to disentangle the relative contributions of genetic factors and different environmental influences.
Current theory and research has been largely silent on the role of genetic transmission and genetic risk for social aggression. Indeed, we know of no molecular genetic studies of social aggression. Fortunately, genetically-informed twin designs can also be used to estimate genetic influences on social aggression, with the added advantage that they are able to simultaneously estimate environmental influences as well. To date, three such studies have been conducted, with notably inconsistent results. The first of these assessed social aggression via teacher and peer ratings in a small sample of six year-old twins (N=234 pairs; Brendgen et al., 2005). Brendgen et al. (2005) employed a constrained ACE model in which additive genetic (A) and shared environmental (C) influences were constrained to be equal because an initial unconstrained ACE model suggested equivalent fits for AE and CE models. Based on these results, the researchers concluded that 60% of the variability in social aggression is due to non-shared environmental factors (E), with 20% due to A and C, respectively. However, a follow-up study of these same twins only a year later (N=203 pairs) failed to find evidence for significant shared environmental influences on social aggression (Brendgen et al., 2008). Instead, results indicated that an AE model provided the best fit to the data, such that 42.9% of the variance in social aggression was due to genetic factors and 57.1% of the variance was due to non-shared environmental factors. Although such findings could suggest a change in the heritability of social aggression from ages 6 to 7 year-old, we would argue against this interpretation, particularly given the very small sample sizes. Although heritability estimates for other antisocial behaviors do change throughout development (Burt and Neiderhiser, 2009), they rarely do so in such dramatic fashion over the course of single year. It is thus unclear what to make of these conflicting results.
Following up on these results, Tackett et al. (2009) made use of a multivariate psychometric model to test etiological influences on the variance common to both mother- and twin-reported child relational aggression in a large sample of 6–18 year-old twins (N=1,981 pairs). In this model, variance common across informant-reports is decomposed into genetic and environmental components, as is the variance unique to each informant. In contrast to Brendgen et al.’s (2005, 2008) studies, Tackett et al. (2009) found evidence for substantial genetic and shared environmental contributions to the latent relational aggression factor (63% and 37% of the variance, respectively). Non-shared environmental influences did not contribute to the latent factor. The variance specific to maternal informant-reports, by contrast, was entirely due to non-shared environmental influences and measurement error. The variance unique to twin-reports was due to unique genetic, shared environmental, and non-shared environmental influences.
Although Tackett et al. (2009) provided important new information on the etiology of social aggression, much remains unknown. This lack of clarity stems from two methodological issues. First, the sample used in Tackett et al., (2009) spans childhood, adolescence, and emerging adulthood, which is less than ideal should the relative contribution of etiological factors differ across those developmental periods (as in Burt and Neiderhiser, 2009). More importantly, however, a burgeoning body of work (Keller and Coventry 2005; Keller and Medland, 2008; Keller et al., 2010) has indicated that heritability estimates from classical twin (CT) studies, such as those outlined above, can be biased in important ways, sometimes severely so. The CT model exploits the comparison of twin similarity across zygosity to infer etiology. To do so, however, it relies on a number of strict assumptions (Keller and Coventry 2005; Keller et al., 2010). When these assumptions are met, the results should accurately reflect the proportions of genetic and environmental influences on the phenotype under study. However, when one or more of these assumptions are violated, the parameter estimates are likely to be biased in predictable ways.
One major assumption is that C and non-additive or dominant genetic effects (D) do not simultaneously influence the phenotype(s) under investigation. This assumption has no basis in biology, but is rather a statistical necessity of the model: because A and E must always be estimated, C and D cannot be simultaneously estimated or else the model will be under-identified. It is entirely conceivable, however, that social aggression is simultaneously influenced both by interactions between alleles (i.e., D) and common environmental factors that create similarities between siblings (i.e., C). Moreover, when C and D effects are both present, parameter estimates are biased, such that A estimates are inflated (sometimes grossly so) and C and D estimates are decreased. As an example, Keller and Medland (2008) simulated data in which A, D, and C variances were equal to .40, .15, and .15, respectively. Using the classical twin model, the ACE model (and not the ADE model) was chosen as the better fitting model, even though C and D effects were in fact equivalent in magnitude. Moreover, additive genetic influences were estimated at .60, whereas shared environmental influences were estimated at .02 (Keller and Medland, 2008).
Finally, the classical twin model assumes that there is no evidence of passive gene-environment correlations (passive rGE). Passive rGE refers to the association between the genes a child inherits from his or her parents and the rearing environment provided by those same parents. Put differently, it is possible that the genes parents pass on to their biological children also influence the home environment in which the child is reared. Within the classical twin model, passive rGE is thought to mimic shared environmental influences, serving to inflate C estimates and decrease A and D estimates. It is thus possible that the shared environmental influences on social aggression identified by Tackett et al. (2009) using the classical twin model may reflect passive rGE rather than actual environmental experiences.
Fortunately, there is a straightforward extension of the classical twin model, the nuclear twin family (NTF) model, which circumvents these assumptions by adding the twins’ biological parents to the model (Keller et al., 2010). In more precise terms, genetic and environmental parameter estimates in the nuclear twin family model (Figure 1) are obtained from four pieces of information (rather than two, as done in the classical twin model): the covariance between MZ twins, the covariance between DZ twins, the covariance between parents, and the covariance between parents and children. The additional information allows the nuclear twin family model to improve upon the classical twin model by simultaneously estimating C and D and differentiating passive rGE from true shared environmental influences. The latter is accomplished via the individuation of shared environmental influences shared only by siblings (S) from those shared by all family members (F). We are then able to explicitly model the extent to which A and F variance components are correlated with one another (w). Perhaps not surprisingly, given these additions, extant simulations studies have robustly demonstrated that the NTF model provides more etiologically precise estimates and is more robust to violations of assumptions than the CT model (Keller et al., 2010).
Figure 1:
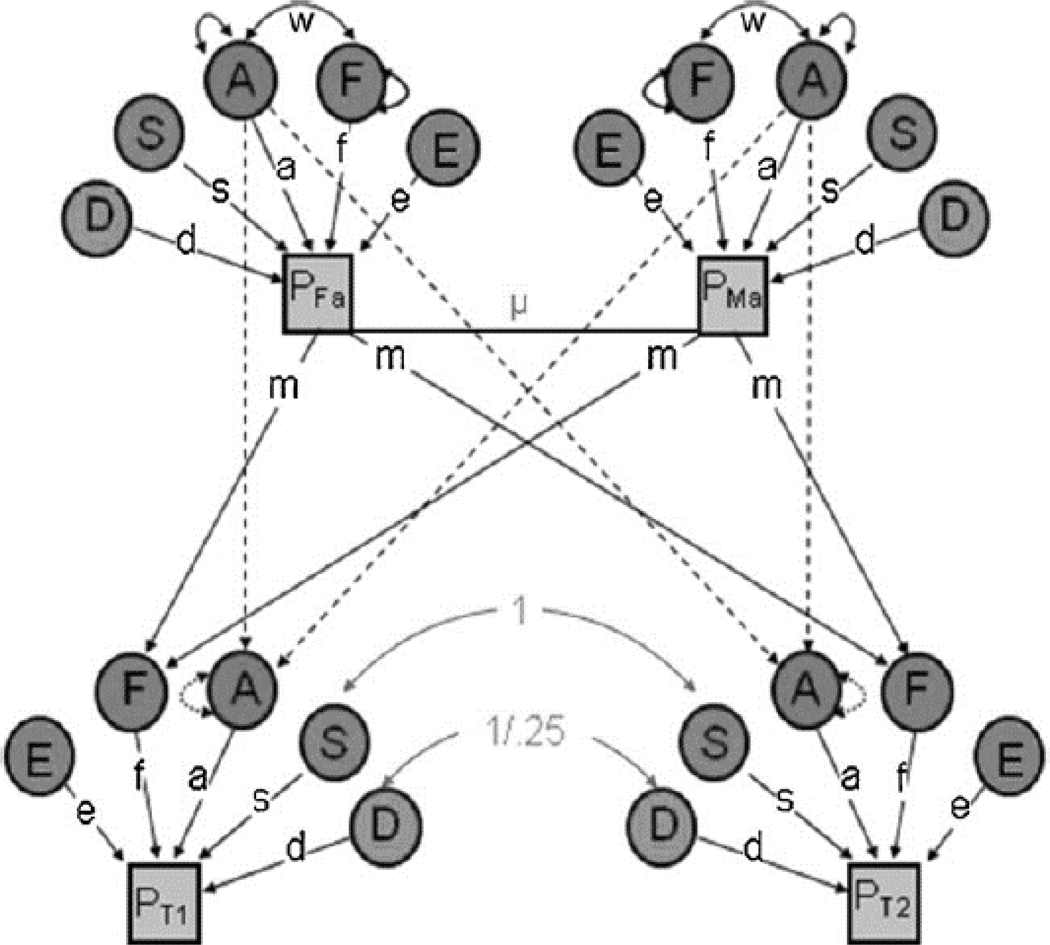
Path diagram of a Univariate Nuclear Twin Family model
Note: The variance in the phenotype (P) is parsed into that which is due to additive genetic effects (A), dominant genetic effects (D), sibling environmental influences (S), familial environmental influences (F), and non-shared environmental effects (E). See Table 1 for definitions. μ indexes primary phenotypic assortment (i.e., assortative mating) between the twin parents, and w indexes the covariance between A and F. Paths are squared to estimate the proportion of variance accounted for. D, S, and F effects cannot be estimated simultaneously (only two of the three can be estimated). Reprinted with permission from Burt & Klump (2012).
Current Study
In sum, extant information on the etiology of social aggression is less certain than one would like. In an effort to meaningfully illuminate the relative magnitudes of genetic and environmental contributions to social aggression, the current study applied the NTF model to multi-informant social aggression data collected from a large sample of twins and their biological parents. As a point of comparison, we also fit the CT model to the data, thereby allowing us to explore the extent to which prior studies conducted using the CT model yielded unbiased results.
METHODOLOGY
Study Participants
The 1,030 families included in the current study were assessed as part of the Twin Study of Behavioral and Emotional Development in Children (TBED-C) within the Michigan State University Twin Registry (MSUTR) (Klump and Burt, 2006; Burt and Klump, 2013). The TBED-C consists of two independent sub-samples of twins in middle childhood. The first sample consists of a population-based epidemiologic sample of 528 families (1,056 twins and their parents). The second, ‘at-risk’ sample consists of 502 families (1,004 twins and their parents), who reside in modestly-to-severely disadvantaged neighborhoods. Recruitment procedures have been described previously (Burt and Klump, 2013; Burt et al., 2016). Children gave informed assent, while parents gave informed consent for themselves and their children.
Participating twins were 48.7% female and ranged in age from 6 to 10 years-old, although some (n = 59) were 11 (mean age (SD) = 8.02 years (1.49)). Rearing biological mothers ranged in age from 24.82 to 52.79 years (mean (SD) = 38.18 (5.21) years), and rearing biological fathers ranged in age from 24.12 to 64.22 years (mean (SD) = 40.12 (5.76) years). Twins’ racial and ethnic background was provided by their parents (81.7% non-Hispanic White, 9.5% African American, 1.1% Native American, 0.8% Asian, 0.7% Hispanic, 0.3% Pacific Islander, and 5.9% multiracial or other ethnic groups). Twin zygosity was determined using a standard 5-item, parent-report questionnaire that assesses within-pair physical similarity and is over 95% accurate (Peeters et al., 1998). Unclear zygosities were resolved by comparing twin sibling DNA markers (Klump and Burt, 2006). Monozygotic twins constituted 41.4% of the pairs (n=426 pairs, 202 female pairs), same-sex dizygotic twins constituted 40.4% (n=416 pairs, 208 female pairs), and opposite-sex dizygotic twins constituted 18.3% (n=188 pairs).
Measures
Social aggression was assessed using the Subtypes of Antisocial Behavior Questionnaire (STAB; Burt and Donnellan, 2009; Burt and Donnellan, 2010). The STAB is a 32-item measure assessing three major dimensions of antisocial behavior, one of which is social aggression. The Social Aggression Scale (SA) presents 11 behaviors and asks the informants to report on the frequency with which the child commits each behavior (Appendix I). This scale ranges from 1 (never) to 5 (nearly all the time).
Maternal-reported STAB data were available for 96.0% of the twins (α = .85) and paternal-reported STAB data were available for 80.9% of the twins (α = .84). The teachers of 115 participants were not available for assessment because the children were home-schooled or parental consents to contact the teachers were completed incorrectly. However, teacher-reported STAB data were available for 82.9% of the remaining twins (α = .91). To avoid shared informant variance with parent self-reports of his or her own behavior, teacher report of child social aggression served as our primary outcome variable. However, we also sought to confirm our primary findings by examining maternal- and paternal-reports of child social aggression. To adjust for positive skew, teacher ratings of twin social aggression were log-transformed prior to analysis to better approximate normality.
Parents each completed the self-report version of the STAB (α = .85 and .86 for mothers and fathers, respectively). In keeping with the assumptions of the nuclear twin family model, self-report data was omitted for parental figures who did not share 50% of their genes with the twins (e.g., grandparents and stepparents). Divorced or separated biological parents with joint custody arrangements or who were otherwise involved in their twins’ lives, however, were retained for analysis. Biological maternal and paternal self-report data were thus available for 93.9% and 76.6% of twin families, respectively.
Statistical analysis
The nuclear twin family model estimates the proportion of additive genetic (A), dominant genetic (D), shared sibling environment (S), shared familial environment (F), and non-shared environmental (E) influences on the variance in a phenotype. The classical twin model also estimates A, D, and E, but estimates broad shared environmental (C) influences rather than the more specific S and F. A and E are assumed to influence all phenotypes and are included in every model. Therefore, D, S, or F (in the NTF model) and C or D (in the CT model) must be fixed to zero, as there is not enough information in a given model to simultaneously estimate the effects of all parameters (Table 1).
Table 1:
Definitions of the parameters obtained via twin modeling
| Parameter | Model | Definition |
|---|---|---|
| A | CT NTF | Additive genetic variance: the effect of individual genes summed over loci; acts to increase familial correlations (either between twin siblings or between parents and their biological children) relative to the proportion of genes shared |
| D | CT NTF | Dominant genetic variance: non-additive interactions between alleles at a single genetic locus (e.g., the interaction between dominant and recessive alleles in the determination of eye color); because they involve interactions between alleles, D do not contribute to similarity between parents and their biological children (since each parent provides only one of the two alleles in question); also yield MZ correlations that more than twice as large as those seen for DZ twins |
| C | CT | Shared environmental variance: environmental influences common to family members that act to make them similar to each other regardless of the proportion of genes shared; C=S+F |
| S | NTF | Sibling environmental influences: those shared environmental influences that create similarity between siblings (e.g., exposure to common peers, school, and parenting style), but not between parents and their children |
| F | NTF | Familial environmental variance: shared environmental influences passed via vertical “cultural transmission” between parents and their offspring (e.g., socioeconomic status, social mores); create similarities between siblings and between parents and their children |
| E | CT NTF | Non-shared environmental variance: environmental influences that serve to differentiate family members regardless of the proportion of genes shared; measurement error is also included here |
| w | NTF | Covariance between additive genetic and familial environmental effects: reflects the extent to which the familial environment is a function of the genetically influenced preferences/ tendencies of the parent; also referred to as a passive gene-environment correlation (passive rGE) |
| μ | NTF | Assortative mating copath: spousal similarity on the trait in question; is assumed to be a function of primary phenotypic assortment, whereby mates choose each other based on phenotypic similarity |
Note: See also Keller et al. (2010). Reprinted with permission from Burt & Klump (2012). CT = Classical Twin Model. NTF = Nuclear Twin Family Model.
Mx, a structural-equation modeling program (Neale et al. 2003), was used to perform the model-fitting analyses. Because of missing data, we made use of Full-Information Maximum-Likelihood (FIML) raw data techniques, which produce less biased and more efficient and consistent estimates than pairwise or listwise deletion in the face of missing data. When fitting models to raw data, variances, covariances, and means are first freely estimated to get a baseline index of fit (minus twice the log-likelihood; −2lnL). Observed covariance matrices were equivalent between same- and opposite-sex DZ twins (Box’s M = 5.93, p = 0.436, ns), so all DZ twins were analyzed as one group. Model fit was evaluated using four information theoretic indices that balance overall fit with model parsimony: the Akaike’s Information Criterion (AIC; Akaike, 1987), the Bayesian Information Criteria (BIC; Raftery, 1995), the sample-size adjusted Bayesian Information Criterion (SABIC; Sclove, 1987), and the Deviance Information Criterion (DIC; Spiegelhalter et al., 2002). The lowest AIC, BIC, SABIC, and DIC among a series of nested models is considered best. As fit indices do not always agree, we reasoned that the best fitting model should yield lower or more negative values for at least three of the four fit indices.
RESULTS
Descriptive statistics are presented in Supplementary Table 1. Phenotypic and intraclass correlations are presented in Table 2. There was evidence of modest assortative mating, such that spouses were slightly more similar in their social aggression than would be expected by chance (r = 0.079, p < 0.01). Additionally, parents’ own social aggression was not correlated with their children’s behavior as measured via teacher report (Mother-Child r = 0.51 to 0.56, p = 0.16 to 0.20, ns; Father-Child r = 0.013 to 0.095, p = 0.27 to 0.76, ns), but was moderately correlated when child behavior was measured via parental report (Mother-Child r = .126 to .487, p > 0.001; Father-Child r = .079 to .489, p > 0.001 to p = 0.008). Parent reports of twin social aggression were moderately correlated with one another (r = .322; p > 0.001), but were only weakly correlated with teacher report of child behavior (Mother-Teacher r = .169, p > 0.001; Father-Teacher r = .114, p = 0.003).
Table 2.
Phenotypic and intraclass correlations
| Mother, Self-Report | Father, Self-Report | Twin, Teacher Report | Twin, Maternal Report | Twin, Paternal Report | Twin, Average Composite | ICC | |
|---|---|---|---|---|---|---|---|
| Mother, Self-Report | - | - | |||||
| Father, Self-Report | .079** | - | - | ||||
| Twin, Teacher Report | .056* | .046 | - | .630**/.372** | |||
| Twin, Maternal Report | .498** | .079** | .169** | - | .767**/.606** | ||
| Twin, Paternal Report | .143** | .489** | .114** | .322** | - | .742**/.735** | |
| Twin, Average Composite | .364** | .323** | .642** | .742** | .725** | - | .725**/.593** |
Note: ICCs are presented separately across zygosity (rMZ/rDZ).
p < 0.05
p < 0.01
Intraclass correlations within twin pairs, computed separately by degree of genetic relatedness, offer a preliminary indication of genetic and sibling environmental influences on social aggression. MZ intraclass correlations that are double those of DZ intraclass correlations are indicative of genetic effects, whereas MZ correlations that are less than double but still greater than DZ correlations suggest the importance of genetic and shared environmental effects. Teacher-reported MZ correlations of social aggression were almost double their corresponding DZ correlations, suggesting that genetic effects may be important for the etiology of social aggression. However, parent-reported MZ correlations were only marginally higher or equivalent to their corresponding DZ correlations, suggesting that, for those informant-reports of social aggression, genetic effects are less prominent and/or that shared environmental effects are also important.
Nuclear Twin Family Model Fitting Results
Model fitting results and parameter estimates are presented in Tables 3 and 4, respectively. According to all four fit indices, the best-fitting model for all informants was the ASFE model, indicating that additive genetic, sibling environmental, familial environmental, and non-shared environmental influences significantly contribute to the etiology of social aggression in middle childhood regardless of informant report, but that non-additive genetic influences do not. The ADSE model was also a good fit for teacher-reported social aggression, but this finding was not robust across alternative informants. In keeping with the familial environmental assumptions of the NTF model, a sensitivity analyses was conducted using only the families (N=749; 72.7%) in which the twins’ biological parents were married to one another. Model fitting results indicated that the ASFE model remained best-fitting model for all informants (Supplementary Table 2).
Table 3:
Nuclear twin family design model fitting results
| Informant | Model | −2LnL | df | AIC | BIC | SABIC | DIC |
|---|---|---|---|---|---|---|---|
| Teacher | Baseline | 8635.428 | 3088 | - | - | - | - |
| ADSE | 8656.946 | 3110 | 2436.946 | −6437.769 | −1498.954 | −3579.871 | |
| ASFE | 8656.946 | 3110 | 2436.946 | −6437.769 | −1498.954 | −3579.871 | |
| ADFE | 8664.975 | 3110 | 2444.975 | −6433.755 | −1494.939 | −3575.856 | |
| ASE | 8674.114 | 3111 | 2452.114 | −6432.647 | −1492.243 | −3573.829 | |
| ADE | 8669.242 | 3111 | 2447.242 | −6435.083 | −1494.680 | −3576.266 | |
| AFE | 8665.907 | 3111 | 2443.907 | −6436.751 | −1496.347 | −3577.933 | |
| AE | 8759.569 | 3112 | 2535.569 | −6393.382 | −1451.390 | −3533.645 | |
| Mother | Baseline | 9597.672 | 3738 | - | - | - | - |
| ADSE | 9852.258 | 3760 | 2332.258 | −8105.038 | −2133.965 | −4649.829 | |
| ASFE | 9844.976 | 3760 | 2324.976 | −8108.679 | −2137.606 | −4653.470 | |
| ADFE | 9910.568 | 3760 | 2390.568 | −8075.883 | −2104.810 | −4620.674 | |
| ASE | 9852.258 | 3761 | 2330.258 | −8108.504 | −2135.843 | −4652.376 | |
| ADE | 9914.689 | 3761 | 2392.687 | −8077.289 | −2104.628 | −4621.161 | |
| AFE | 9938.126 | 3761 | 2416.126 | −8065.570 | −2091.909 | −4609.442 | |
| AE | 9978.983 | 3762 | 2454.983 | −8048.607 | −2074.358 | −4591.560 | |
| Father | Baseline | 8888.811 | 3427 | - | - | - | - |
| ADSE | 9079.581 | 3449 | 2181.581 | −7393.204 | −1916.055 | −4223.785 | |
| ASFE | 9021.367 | 3449 | 2123.367 | −7422.311 | −1945.162 | −4252.892 | |
| ADFE | 9161.829 | 3449 | 2263.829 | −7352.080 | −1874.931 | −4182.661 | |
| ASE | 9079.581 | 3450 | 2179.581 | −7396.664 | −1917.927 | −4226.326 | |
| ADE | 9167.985 | 3450 | 2267.985 | −7352.462 | −1873.725 | −4182.124 | |
| AFE | 9186.150 | 3450 | 2286.150 | −7343.380 | −1864.643 | −4173.042 | |
| AE | 9199.538 | 3451 | 2297.538 | −7340.145 | −1859.820 | −4168.889 |
Note: Additive genetic, dominant genetic, sibling environmental, familial environmental, and non-shared environmental influences are represented with A, D, S, F, and E, respectively. The best fitting model for each informant (as indicated by the lowest AIC, BIC, SABIC, and DIC values for at least 3 of the 4 fit indices) is highlighted in bold font.
Table 4:
Best fitting nuclear twin family design parameter estimates
| Informant | Model | A | S | F | E | Passive rGE | Assortative Mating |
|---|---|---|---|---|---|---|---|
| Teacher | ASFE |
0.770* [0.619, 0.899] |
0.422* [0.248, 0.543] |
−0.120 [−0.280, −0.111] |
0.589* [0.536, 0.643] |
−0.139 [−0.215, −0.069] |
0.075* [0.004, 0.146] |
| Mother | ASFE |
0.601* [0.505, 0.687] |
0.563* [0.502, 0.602] |
0.081* [0.022, 0.142] |
0.471* [0.440, 0.505] |
0.061* [0.018, 0.100] |
0.093* [0.023, 0.164] |
| Father | ASFE | .144 [−0.360, 0.360] |
.717* [.653, 736] |
.279* [.201, 325] |
.531* [.494, 560] |
0.062 [−0.118, 0.109] |
0.073* [0.003, 0.142] |
Note: Additive genetic, dominant genetic, sibling environmental, familial environmental, and non-shared environmental influences are represented with A, D, S, F, and E, respectively. 95% confidence intervals are presented below the point estimate in brackets.
p < 0.05.
Examinations of the parameter estimates revealed that social aggression is largely additive genetic (A = 0.14 – 0.77) and sibling environmental (S = 0.42 – 0.72) in origin. Familial environmental factors were estimated to be small but generally significant influences on social aggression (F = 0.08 – 0.28). Finally, the results suggest that the etiology of social aggression is not substantially influenced by assortative mating (μ = 0.07 – 0.09) or the covariance between additive genetic and familial environmental effects (e.g., passive rGE = −0.14 – 0.06). Although statistically significant according to some informants, the meager size of the passive rGE further implies that effects of the home environment are genuine and not instead a function of genetically influenced tendencies in disguise.
Classical Twin Model Results
We next fitted classical univariate models to visually compare to the above nuclear twin family model results and, perhaps more importantly, to previous CT model findings concerning the etiology of social aggression. Model fitting results and heritability estimates are presented in Supplementary Tables 3 and 4, respectively. These analyses are not nested within those of the nuclear twin family model, so the model fitting results from the two models cannot be directly compared. However, a comparison of the overall pattern of results is possible.
Unlike the nuclear twin family model, the best-fitting classical univariate twin model varied dramatically across informants. The ACE model was the best-fitting model for maternal-reported data, while, the CE model was the best-fitting model for paternal-reported data. The ADE and AE models resulted in equivalent best-fitting models for teacher-reported social aggression. These findings contrast with not only each other, but also some of those reported in previous research on the etiology of social aggression. When examining the informant reports separately, the only common source of variance across all was non-shared environmental influences (which includes measurement error).
DISCUSSION
The goal of the current study was to conduct a multi-informant analysis of the etiology of childhood social aggression using the more sophisticated and etiologically precise nuclear twin family model. Results from the nuclear twin family model revealed that social aggression is due to additive genetic (A), sibling environmental (S), familial environmental (F), and non-shared environmental (E) influences, although the relative magnitudes of each parameter differed across informants. Such results indicate that, in addition to genetic effects, sibling-level environmental influences, such as peer groups, school experiences, parenting practices, media exposure, and/or the neighborhood in which the twins are raised, make an important contribution to the etiology of social aggression. Environmental effects that are shared by both the parents and their children, such as socioeconomic status and/or cultural values, also contribute, although these contributions are smaller in magnitude. In sharp contrast, heritability estimates calculated using the classical twin model were notably inconsistent across informant. Moreover, classical twin analyses of only one informant-report (maternal) yielded estimates that were comparable across the NTF and CT models.
There are several limitations to be considered in the present study. First, the heritability estimates found here are specific to middle childhood and should not be generalized to other developmental periods. Second, the age variation in both the twins and their parents may mask etiological changes across development. Additionally, we made use of Full-Information Maximum-Likelihood (FIML) raw data techniques to address missing data. FIML raw data analyses assume that missing data are missing at random (MAR; Allison, 2003; Croy and Novins, 2005). Although the missing mother and twin data did appear to be MAR, the missing father did not appear to be fully MAR. Maternal-reports of paternal felony convictions varied with father missingness (4.3% and 26.6% in participating versus non-participating fathers, respectively; p < 0.001). However, controlling for the other variables in our analyses (i.e., maternal and twin antisocial behavior, twin ethnicity, twin age, twin sex) appeared to significantly reduce this effect. In a regression of father missingness, the Beta for paternal felony convictions dropped from 0.32 (p < 0.001) when analyzed alone to 0.15 (p = 0.023) when analyzed with the other variables.
Although the examination of teacher-, mother-, and father-reports was a strength of the present study, the lack of twin self-reports and/or peers-ratings of the twins limited our ability to fully investigate the etiology of social aggression. These could be especially useful since social aggression is characterized by both overt behaviors (that adults are likely to witness) and covert behaviors (of which only the child and his or her peers may be aware). Peer ratings are commonly used with older children and adolescents and have been suggested to be more valid than teacher, parent, or self-ratings of social aggression (Archer and Coyne, 2005). Future research should incorporate self-, peer-, and adult-reports into the assessment of social aggression in adolescence and young adulthood.
One important limitation of the NTF model is its reliance on phenotypic measures across generations. Cognitive, social, and biological changes across development, especially those associated with puberty, may moderate the prevalence, manifestation, and/or etiology of a given phenotype. This is potentially problematic for social aggression because there is limited work on its developmental trajectories across the lifespan. Socially and relationally aggressive behaviors have been observed across the lifespan, from toddlers to the elderly but are thought to peak around adolescence (Crick et al., 2006; Trompetter et al., 2011), although there is little-to-no data examining changes in prevalence into adulthood (Vaillancourt et al., 2007; Underwood et al., 2009). Data also suggest that behavioral manifestations of social aggression may become more covert and sophisticated as individuals mature (Crick et al., 2007), although this literature is somewhat complicated by methodological differences in the assessment of social aggression across development.
In the present study, social aggression was assessed using the same questionnaire in both the parents and twins. Importantly, prior work has confirmed the factor structure of the STAB and provided consistent support for its criterion-related validity in multiple samples across many (but not all) developmental stages (Burt and Donnellan, 2009; 2010). In the current sample, parents exhibited higher levels of overall social aggression than did their children, but the magnitude of these differences were small. Item-level analyses of mean differences in specific socially aggressive behaviors revealed that there were also small differences in its manifestation in childhood versus adulthood, but these differences were not systematic (e.g., covert vs. overt). We thus think it is unlikely that these differences in frequency or manifestation compromise the validity of our findings. However, future studies should explore this possibility in more depth.
Conclusion
Relative to other behavior genetic studies of childhood social aggression, there are several significant findings, the most notable of which is that the NTF model provided model fitting results that were broadly consistent across various informants and data pooling methods. In contrast to this, prior multi-informant studies utilizing the classical twin model found notably inconsistent evidence for the etiology of twin social aggression, even when rating the same sample of twins. The latter results were replicated here, with various informant-reports yielding evidence of the ACE, CE, and ADE/AE models, respectively. Such findings raise the provocative hypothesis that the nuclear twin family model may yield results that are more robust to informant effects than does the CT, for which informant effects are widely reported (see Burt, 2009). Future work should more directly explore this possibility, and do so across a variety of phenotypes and informants.
Although the NTF model appeared more robust to informant bias when estimating etiology (relative to the CT model), we nevertheless observed informant effects in the NTF models as well. Specifically, paternal-reports yield different results relative to maternal- and teacher-reports. Both mothers and teachers rated monozygotic twins as more similar than dizygotic twins, while fathers rated both types of twins as equally similar, leading to higher estimates of shared environmental factors and lower estimates of genetic and unique environmental influences relative to other informants. Such findings could reflect bias on the part of fathers, or limited information/exposure to their twin’s socially aggressive behavior (and thus an inability to distinguish between them; Izquierdo-Sotorrío et al., 2016). Alternately, they could reflect bias on the part of mothers and teachers. Indeed, maternal reports have been criticized for producing biased estimates of social aggression, especially in regards to sex differences (Card et al., 2008). That said, it is not clear how this might shape zygosity differences in maternal-reports of twin similarity. Finally, these results could also reflect situational specificity, whereby the twins engage in different levels of and/or perpetrate different types of socially aggressive behavior with their fathers than with mothers or teachers. Although possible, the latter explanation is slightly undercut by the patterns of phenotypic correlations, in which both mother and father reports (rather than father only) were only weakly correlated with teacher reports.
Another important finding is that aspects of the shared sibling environment were found to substantially influence variability in the twins’ social aggression, whereas aspects of the shared family environment made more modest contributions. Such results suggest that social aggression in middle childhood is meaningfully shaped by experiences shared by the twins (rather than the twins and their parents), such as peer groups, parenting practices, marital conflict, media exposure, or neighborhood. Indeed, deviant peer groups have long been linked to the development of other antisocial behaviors, and peer influences have recently shown to be predictive of later socially aggressive behavior (Werner and Crick, 2004; Yeung and Leadbeater, 2007). Another sibling-level environmental mechanism potentially involved in the development of social aggression is parenting behaviors, such as psychological control and negative parenting (Bregden et al., 2005). For instance, hostile/inconsistent parenting at age two has been shown to be a significant predictor of increasing levels of social aggression across childhood (Vaillancourt et al., 2007). Future research should investigate which of these sibling environmental influences incrementally account(s) for the prominent shared environmental influences on social aggression.
Supplementary Material
Acknowledgements:
The authors thank all participating twins and their families for making this work possible.
Financial Support: This project was supported by R01-MH081813 from the National Institute of Mental Health (NIMH) and by R01-HD066040 from the Eunice Kennedy Shriver National Institute for Child Health and Human Development (NICHD). The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIMH, the NICHD, or the National Institutes of Health.
Footnotes
Required Statements
Ethical Standards: The authors assert that all procedures contributing to this work comply with the ethical standards of the relevant national and institutional committees on human experimentation and with the Helsinki Declaration of 1975, as revised in 2008.
Conflict of Interest: None
References
- Akaike H (1987). Factor analysis and AIC. Psychometrika, 52, 317–332. [Google Scholar]
- Allison PD (2003). Missing Data Techniques for Structural Equation Modeling. Journal of Abnormal Psychology, 112, 545–557. [DOI] [PubMed] [Google Scholar]
- Archer J, Coyne SM (2005). An integrated review of indirect, relational, and social aggression. Personality and Social Psychology Review, 101, 212–230. [DOI] [PubMed] [Google Scholar]
- Baker LA, Barton M, Lozano DI, Raine A, Fowler JH (2006). The Southern California Twin Register at the University of Southern California: II. Twin Research and Human Genetics, 9, 933–940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker LA, Jacobson KC, Raine A, Lozano DI, Bezdjian S (2007). Genetic and environmental bases of childhood antisocial behavior: A multi-informant twin study, Journal of Abnormal Psychology,116, 219–235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brendgen M, Dionne G, Girard A, Boivin M, Vitaro F, Pérusse D (2005). Examining Genetic and Environmental Effects on Social Aggression: A Study of 6-Year-Old Twins. Child Development, 76, 930–946. [DOI] [PubMed] [Google Scholar]
- Brendgen M, Boivin M, Vitaro F, Bukowski WM, Dionne G, Tremblay RE, Pérusse D (2008). Linkages between children’s and their friends’ social and physical aggression: Evidence for a gene-environment interaction? Child Development, 79, 13–29. [DOI] [PubMed] [Google Scholar]
- Burt SA (2009). Rethinking environmental contributions to child and adolescent psychopathology: A meta-analysis of shared environmental influences. Psychological Bulletin, 135, 608–637. [DOI] [PubMed] [Google Scholar]
- Burt SA, Donnellan MB (2009). Development and validation of the subtypes of antisocial behavior questionnaire. Aggressive Behavior, 35, 376–398. [DOI] [PubMed] [Google Scholar]
- Burt SA, Donnellan MB (2010). Evidence that the Sub-Types of Antisocial Behavior questionnaire (STAB) predicts momentary reports of acting-out behaviors. Personality and Individual Differences, 48, 917–920. [Google Scholar]
- Burt SA, Donnellan MB, Tackett JL (2012). Should social aggression be considered “antisocial”? Journal of Psychopathology and Behavioral Assessment, 34, 153–163. [Google Scholar]
- Burt SA, Klump KL (2013). The Michigan State University Twin Registry (MSUTR): An update. Twin Research and Human Genetics, 16, 344–350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burt SA, Klump KL, Gorman-Smith D, Neiderhiser JM (2016). Neighborhood disadvantage alters the origins of children’s non-aggressive conduct problems. Clinical Psychological Science, 4, 511–526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burt SA, McGue M, Krueger RF, Iacono WG (2005). Sources of covariation among the child-externalizing disorders: Informant effects and the shared environment. Psychological Medicine, 35, 1133–1144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burt SA, Neiderhiser JM (2009). Aggressive versus non-aggressive antisocial behavior: Distinctive etiological moderation by age. Developmental Psychology, 45, 1164–1176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Card NA, Stucky BD, Sawalani GM, Little TD (2008). Direct and indirect aggression during childhood and adolescence: A meta-analytic review of gender differences, intercorrelations, and relations to maladjustment. Child Development, 79, 1185–1229. [DOI] [PubMed] [Google Scholar]
- Crick NR, Casas JF, Nelson DA (2002). Toward a more comprehensive understanding of peer maltreatment: Studies of relational victimization. Current Directions in Psychological Science, 11, 98–101. [Google Scholar]
- Crick NR, Grotpeter JK (1995). Relational aggression, gender, and social-psychological adjustment. Child Development, 66, 710–722. [DOI] [PubMed] [Google Scholar]
- Crick NR, Ostrov JM, Burr JE, Cullerton-Sen C, Jansen-Yeh E, Ralston P (2006). A longitudinal study of relational and physical aggression in preschool. Journal of Applied Developmental Psychology, 27, 254–268. [Google Scholar]
- Crick NR, Ostrov JM, & Kawabata Y (2007). Relational aggression and gender: An overview In The Cambridge handbook of violent behavior and aggression (ed. Flannery DJ, Vazsonyi AT, and Waldman ID), pp. 245–259. Cambridge University: New York. [Google Scholar]
- Coventry WL, Keller MC (2005). Estimating the Extent of Parameter Bias in the Classical Twin Design: A Comparison of Parameter Estimates From Extended Twin-Family and Classical Twin Designs. Twin Research and Human Genetics, 8, 214–223. [DOI] [PubMed] [Google Scholar]
- Croy CD, Novins DK (2005). Methods for addressing missing data in psychiatric and developmental research. Journal of the American Academy of Child & Adolescent Psychiatry, 44, 1230–1240. [DOI] [PubMed] [Google Scholar]
- Forbes G, Zhang X, Doroszewicz K, Haas K (2009). Relationships between individualism-collectivism, gender, and direct or indirect aggression: A study in China, Poland, and the U.S. Aggressive Behavior, 35, 24–30. [DOI] [PubMed] [Google Scholar]
- Gentile DA, Coyne S, Walsh DA (2011). Media violence, physical aggression, and relational aggression in school-age children: A short-term longitudinal study. Aggressive Behavior, 37, 193–206. [DOI] [PubMed] [Google Scholar]
- Hay DA, McStephen M, Levy F, Pearsall-Jones J. (2002). Recruitment and attrition in twin register studies of childhood behavior: The example of the Australian twin ADHD project. Twin Research, 5, 324–328. [DOI] [PubMed] [Google Scholar]
- Humbad MN, Donnellan MB, Iacono WG, Burt SA (2010). The association of marital quality with personality and psychopathology: Results from a very large sample of married couples. Journal of Abnormal Psychology, 119, 151–162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Izquierdo-Sotorrío E, Holgado-Tello FP, Carrasco MA (2016). Incremental Validity and Informant Effect from a Multi-Method Perspective: Assessing Relations between Parental Acceptance and Children’s Behavioral Problems. Frontiers in Psychology, 7, 664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karriker-Jaffe KJ, Foshee VA, Ennett ST, Suchindran C (2008). Associations of neighborhood and family factors with trajectories of physical and social aggression during adolescence. Journal of Youth and Adolescence, 42, 861–877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawabata Y, Alink LRA, Tseng WL, van Ijzendoorn MH, Crick NR (2011). Maternal and parental parenting styles associated with relational aggression in children and adolescents: A conceptual analysis and meta-analytic review. Developmental Review, 31, 240–273. [Google Scholar]
- Keller MC, Coventry WL (2005). Quantifying and addressing parameter indeterminacy in the classical twin design. Twin Research and Human Genetics, 8, 201–213. [DOI] [PubMed] [Google Scholar]
- Keller MC, Medland SE (2008). Evaluation of the cascade model - a new extended twin family model. Paper presented at the Behavioral Genetics Association: Louisville, KY. [Google Scholar]
- Keller MC, Medland SE, Duncan LE (2010). Are extended twin family designs worth the trouble? A comparison of the bias, precision, and accuracy of parameters estimated in four twin family models. Behavior Genetics, 40, 377–393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klump KL, Burt SA (2006). The Michigan State University Twin Registry (MSUTR): Genetic, environmental, and neurobiological influences on behavior across development. Twin Research and Human Genetics, 9, 971–977. [DOI] [PubMed] [Google Scholar]
- Kraemer HC, Measelle JR, Ablow JC, Essex MJ, Boyce WT, Kupfer DJ (2003). A new approach to integrating data from multiple informants in psychiatric assessment and research: Mixing and matching contexts and perspectives. American Journal of Psychiatry, 160, 1566–1577. [DOI] [PubMed] [Google Scholar]
- Krueger RF, Moffitt TE, Caspi A, Bleske A, Silva PA (1998). Assortative mating for antisocial behavior: Developmental and methodological implications. Behavior Genetics, 28, 173–186. [DOI] [PubMed] [Google Scholar]
- Kuppens S, Grietens H, Onghena P, Michiels D, Subramanian SV (2013). Relations between parental psychological control and childhood relational aggression: Reciprocal in nature? Journal of Clinical Child and Adolescent Psychology, 38, 117–131. [DOI] [PubMed] [Google Scholar]
- McGue M, Bouchard TJJ (1984). Adjustment of twin data for the effects of age and sex. Behavior Genetics, 14, 325–343. [DOI] [PubMed] [Google Scholar]
- Neale MC, Boker SM, Xie G, Maes HH (2003). Mx: Statistical Modeling. VCU Box 900126, Richmond, VA: 23298: Department of Psychiatry, 6th Edition. [Google Scholar]
- Peeters H, Gestel SV, Vlietinck R, Derom C, Derom R (1998). Validation of a telephone zygosity questionnaire in twins of known zygosity. Behavior Genetics, 28, 159–163. [DOI] [PubMed] [Google Scholar]
- Raftery AE (1995). Bayesian model selection in social research. Sociological Methodology, 25, 111–163. [Google Scholar]
- Sclove LS (1987). Application of model-selection criteria to some problems in multivariate analysis. Psychometrika, 53, 333–343. [Google Scholar]
- Sijtsema JJ, Ojanen T, Veenstra R, Lindenberg S, Hawley PH, Little TD (2010). Forms and functions of aggression in adolescent friendship selection and influence: A longitudinal social network analysis. Social Development, 19, 514–534. [Google Scholar]
- Spiegelhalter DJ, Best NG, Carlin BP, Van Der Linde A (2002). Bayesian measures of model complexity and fit. Journal of the Royal Statistical Society: Series B, 64, 583–639. [Google Scholar]
- Tackett JL, Waldman ID, Lahey BB (2009). Etiology and measurement of relational aggression: A multi-informant behavior genetic investigation. Journal of Abnormal Psychology, 118, 722–733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trompetter H, Scholte R, Westerhof G (2011). Resident-to-resident relational aggression and subjective well-being in assisted living facilities. Aging & Mental Health, 15, 59–67. [DOI] [PubMed] [Google Scholar]
- Underwood MK, Beron KJ, Rosen LH (2009). Continuity and change in social and physical aggression from middle childhood through early adolescence. Aggressive Behavior, 35, 357–375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vaillancourt T, Miller JL, Fagbemi J, Côte S, Tremblay RE (2007). Trajectories and predictors of indirect aggression: Results from a nationally representative longitudinal study of Canadian children aged 2–10. Aggressive Behavior, 33, 314–326. [DOI] [PubMed] [Google Scholar]
- Werner NE, Crick NR (2004). Maladaptive peer relationships and the development of relational and physical aggression during middle childhood. Social Development, 13, 495–514. [Google Scholar]
- Yeung RS, Leadbeater BJ (2007). Does hostile attributional bias for relational provocations mediate the short-term association between relational victimization and aggression in preadolescence? Journal of Youth and Adolescence, 36, 973–983. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.