

Abstract
Overconsumption of carbohydrates and lipids are well known to cause nonalcoholic fatty liver disease (NAFLD), while the role of nutritional protein intake is less clear. In Western diet, meat and other animal products are the main protein source, with varying concentrations of specific amino acids. Whether the amount or composition of protein intake is associated with a higher risk for disease severity has not yet been examined. In this study, we investigated associations of dietary components with histological disease activity by analyzing detailed 14‐day food records in a cohort of 61 patients with biopsy‐proven NAFLD. Furthermore, we used 16S ribosomal RNA gene sequencing to detect associations with different abundances of the gut microbiota with dietary patterns. Patients with definite nonalcoholic steatohepatitis (NAFLD activity score of 5‐8 on liver biopsy) had a significantly higher daily relative intake of protein compared with patients with a NAFLD activity score of 0‐4 (18.0% vs. 15.8% of daily protein‐based calories, P = 0.018). After adjustment for several potentially confounding factors, a higher protein intake (≥17.3% of daily protein‐based calories) remained associated with definite nonalcoholic steatohepatitis, with an odds ratio of 5.09 (95% confidence interval 1.22‐21.25, P = 0.026). This association was driven primarily by serine, glycine, arginine, proline, phenylalanine, and methionine. A higher protein intake correlated with a lower Bacteroides abundance and an altered abundance of several other bacterial taxa. Conclusion: A high protein intake was independently associated with more active and severe histological disease activity in patients with NAFLD. Further studies are needed to investigate the potential harmful role of dietary amino acids on NAFLD, with special attention to meat as their major source.
Abbreviations
- ALT
alanine aminotransferase
- AST
aspartate aminotransferase
- BMR
basal metabolic rate
- BMI
body mass index
- CI
confidence interval
- EI
energy intake
- GGT
gamma‐glutamyl‐transferase
- HbA1c
glycated hemoglobin
- HDL
high‐density lipoprotein
- mTOR
mammalian target of rapamycin
- NAFLD
non‐alcoholic fatty liver disease
- NAS
NAFLD activity score
- NASH
non‐alcoholic steatohepatitis
- OR
odds ratio
Nonalcoholic fatty liver disease (NAFLD) has evolved as the most common chronic liver disease in the world, affecting 17%‐46% of the general population in Western countries with growing incidence.( 1 ) NAFLD includes a variety of histopathological findings ranging from simple steatosis to steatohepatitis with inflammation to fibrosis and eventually cirrhosis.( 2 ) Nonalcoholic steatohepatitis (NASH) occurs in 7%‐30% of cases with NAFLD, and one‐third of these patients may develop liver cirrhosis.( 3 , 4 )
Risk factors such as older age, presence of type 2 diabetes, increased body mass index (BMI), and genetic variants like the common PNPLA3 (encoding patatin‐like phospholipase domain containing protein 3) p.I148M polymorphism are associated with disease activity and progression.( 5 , 6 , 7 , 8 )
Nutritional studies in patients with NAFLD have identified associations of fructose and other simple carbohydrates, fat, and saturated fatty acids with disease development and progression.( 9 ) Fructose intake stimulates de novo lipogenesis in both animals and humans, together with blocking of the intrahepatic β‐fatty acid oxidation, and has been suggested to play a key role in NAFLD.( 10 )
However, little is known about the role of dietary protein. Overconsumption of protein, and in particular animal protein, occurs worldwide; the global average protein consumption is approximately more than one‐third higher than the average daily adult requirement, with even higher amounts in the world’s wealthiest regions.( 11 ) There is emerging evidence for associations of high protein intake and NAFLD development. Two large recent studies have shown a higher prevalence of NAFLD among study participants consuming diets high in protein and red meat after adjustment for several potentially confounding factors.( 12 , 13 )
In Western diet, meat and other animal products are the main source of dietary amino acids.( 11 , 14 , 15 ) In parallel, concentrations of plasma amino acids were found to be increased in patients with more severe stages of NAFLD( 16 ); however, associations of dietary amino acids with histological disease activity and severity in NAFLD have not yet been investigated.
Protein is an important substrate for commensal gut bacteria. The dietary pattern influences the abundance of several bacteria in this ecosystem,( 17 ) and the bacteria themselves play a role in regulating host bioavailability of amino acids by fermentation and metabolic transformation.( 18 )
In this cross‐sectional study, we investigated associations of dietary macronutrients with histological disease activity in a well‐described cohort of patients with NAFLD. In addition, we analyzed associations of dietary compounds with alterations in the bacterial gut microbiota composition.
Methods
Study Population
This study was designed to detect associations among diet, the gut microbiota, as well as genetic factors and the activity and severity of NAFLD. Therefore, a total of 180 patients with NAFLD were prospectively consecutively enrolled between July 2015 and December 2018 during regular follow‐up visits in the outpatient liver department of the Clinic for Gastroenterology and Hepatology, University Hospital of Cologne, Germany. The study cohort has been described previously.( 19 )
A total of 61 patients with NAFLD with available liver biopsy results and dietary records were included in the analysis, with the aim of detecting associations between histological disease activity and dietary nutrient intake. A total of 99 patients with NAFLD with fecal samples and dietary record were included in the analysis that aimed to investigate associations of nutrients with different abundances in the gut microbiota, whereas 59 patients of the biopsy‐proven cohort were overlapping and included in the microbiota analysis.
Within this study, a detailed history, physical exam, anthropometric measurements, blood pressure measurement, ultrasound and/or magnetic resonance imaging, transient elastography and liver biopsy, if clinically indicated, as per standard of care, was performed. NAFLD was diagnosed if the following conditions were met: hepatic steatosis on liver imaging (ultrasound and/or magnet resonance imaging) and/or the presence of at least 5% fat in histological analysis of liver biopsy, daily alcohol consumption of less than 10 g in women and less than 20 g in men, absence of steatogenic drugs such as glucocorticoids, methotrexate, amiodarone and tamoxifen, absence of other diseases causing secondary steatosis such as human immunodeficiency virus infection, celiac disease or inflammatory bowel disease, and absence of other chronic liver diseases (e.g., viral hepatitis, autoimmune hepatitis, toxic liver injury, alcoholic steatohepatitis, cholestatic liver disease, Wilson’s disease, and hereditary hemochromatosis).
Exclusion criteria for all study subjects were oral or intravenous antibiotic treatment within the last 6 months prior to the study, known malignancy, pregnancy, and under 18 years of age. Any recommendations or treatment suggestions for study participants did not differ from usual patient care. Thus, patients with NAFLD received the same overall lifestyle recommendations as indicated in the current European guideline.( 20 ) Further exclusion criteria for patients with NAFLD were ongoing successful lifestyle modifications defined as more than 5% loss of body weight within the last 3 months prior to enrollment, changes of usual dietary habits within the last 6 months, or current or prior participation in an interventional NASH study.
Abdominal ultrasound was performed for all patients. All blood samples for laboratory analyses were collected under fasting conditions. Anthropometric measurements were carried out by trained physicians or research assistant nurses.
Type 2 diabetes was defined as glycated hemoglobin (HbA1c) ≥ 6.5% and/or fasting glucose ≥ 126 mg/dL and/or use of antidiabetic medications. Overweight was defined as BMI ≥ 25 kg/m2. Metabolic syndrome was defined following the International Diabetes Foundation criteria. Arterial hypertension was defined as office blood pressure ≥ 140/90 mmHg on two or more measurements during two or more occasions or antihypertensive drug treatment.
The protocol was approved by the Ethics Commission of Cologne University’s Faculty of Medicine, and written informed consent was obtained from each patient. The study was performed in accordance with the Declaration of Helsinki.
Liver Biopsies
If liver biopsy was performed, samples were evaluated by an experienced liver pathologist who was blinded for all clinical and laboratory patient data. The NASH Clinical Research Network histological scoring system( 21 ) was used to evaluate disease activity and severity. Accordingly, steatosis was graded 0‐3, ballooning was graded 0‐2, lobular inflammation was graded 0‐3. Fibrosis was staged from 0‐4. Stages 1a, 1b, and 1c were summarized as stage 1 (fibrosis stages: 0, none; 1, perisinusoidal or periportal; 2, perisinusoidal and portal/periportal; 3, bridging fibrosis; 4, cirrhosis). The NAFLD activity score (NAS) was obtained for each biopsy. This score is defined as the unweighted sum of the scores for steatosis (0‐3), lobular inflammation (0‐3), and ballooning (0‐2), thus ranging from 0 to 8.( 21 ) Definite NASH was defined as NAS 5‐8.
Diet/Activity Records
The food intake and physical activity was recorded using an open, 14‐day self‐administered food/activity record. Patients were instructed to report each daily portion of consumed food and all beverages in as much detail as possible directly after ingestion, to weigh foods or to estimate doses in grams and not to change usual dietary and physical activity habits during the collection period. Patients were instructed to report physical habits for each day in as much detail as possible. EBISpro 2016 professional scientific software was used to analyze energy intake, basal metabolic rate, energy expenditure, and all macronutrients and micronutrients. The median absolute values over all days were calculated, and the total amount in grams as well as the relative intake regarding the overall energy intake using the nutrient density method( 22 ) was reported as described in a previous study. For example, 1 g of protein equals 4 kcal; hence, to calculate the energy percent of protein, (energy %) = (total protein intake [g] × 4/total kcal intake) × 100. Similarly, the energy percentage of carbohydrates (4 kcal/g), fats (9 kcal/g), fiber (2 kcal/g) and alcohol (6 kcal/g) were determined.( 12 ) The physical activity level was calculated by dividing the estimated energy expenditure by the basal metabolic rate.( 23 )
Energy misreporting is a very frequently observed issue in self‐reported dietary assessment and is considered to be unavoidable.( 24 , 25 ) However, excluding misreporters leads to a loss of statistical power and may bias estimates of associations.( 25 ) As an alternative approach, we calculated the ratio between energy intake (EI) and the basal metabolic rate (BMR) (EI:BMR ratio). Thirty‐six percent of the cohort were definite energy misreporters (EI:BMR < 1). To account for energy misreporting in our study, we adjusted our multivariate analyses additionally for the EI:BMR ratio in similarity to other publications.( 25 , 26 )
Gut Bacterial Sequencing
The DNA was isolated using the RNeasy Power Microbiome Kit (Qiagen, Hilden, Germany). Seven of the 9 variable bacterial 16S ribosomal RNA (rRNA) gene regions (pool 1: V2, V4, and V8; pool 2: V3, V6/7, and V9) were amplified with the Ion 16S Metagenomics Kit (Thermo Fisher Scientific, Waltham, MA) using two primer pools (an integrated research solution for bacterial identification using 16S rRNA sequencing on the Ion PGM System with Ion Reporter Software [https://www.thermofisher.com/content/ http://dam/LifeTech/Documents/PDFs/Ion-16S-Metagenomics-Kit-Software-Application-ote.pdf]). Amplicons were pooled and cleaned using the NucleoMag NGS Clean‐up (Macherey‐Nagel, Düren, Germany). The Qubit system was used to determine amplicon concentration, the library was prepared with the Ion Plus Fragment Library Kit (Thermo Fisher Scientific). For the template‐preparation amplicon, concentration was diluted to 30 ng/mL. The Ion Chef Kit and the Ion Chef system (Thermo Fisher Scientific) were used to enrich and prepare the template‐positive ion sphere particles. Amplicon library was sequenced using the Ion Torrent S5 system (pH‐dependent, Thermo Fisher Scientific). The amplicon sequences were clustered into operational taxonomic units before taxonomical alignment with the MicroSEQ 16S‐rDNA Reference Library v2013.1 (Thermo Fisher Scientific) and Greengenes v13.5 databases. Ninety‐seven percent similarity was used to genus‐level assignment, and 99% similarity for species‐level assignment. Data files were assigned by the Ion Reporter metagenomics 16S w1.1 workflow (Thermo Fisher Scientific). The raw data were processed using the programming language R version 3.5.1.
Accession Number Sequence Data
Sequence data were registered at NCBI under BioProject PRJNA540738. Sequence reads are available at NCBI under the following BioSample IDs: SAMN11554417 ‐SAMN11554484, SAMN13895359, SAMN13895361, SAMN13895362, SAMN13895369, SAMN13895370, SAMN13895372, SAMN13895374, SAMN13895377, SAMN13895378, SAMN13895381, SAMN13895382, SAMN13895384, SAMN13895388, SAMN13895391, SAMN13895392, SAMN13895394‐SAMN13895400, SAMN14219187‐SAMN14219190, SAMN14219205 and SAMN14219208‐SAMN14219211.
Statistical Analysis
Results are expressed as median and interquartile range (in parentheses) for each continuous variable and number and percentage for categorical variables. Two groups were compared using the Mann‐Whitney Wilcoxon test for continuous variables and chi‐squared tests for categorical variables.
Patients with NAFLD with available liver biopsy (n = 61) were categorized according to the histological staging into a group with a NAS of 0‐4 and definite NASH (NAS 5‐8), as well as no to mild fibrosis (F0‐F1) versus moderate fibrosis to cirrhosis (F2‐F4). We analyzed the nutrient intake as continuous variable (in energy percentage) using standardized values (increase per 1 SD, in analogy to a previous publication( 12 )), and patients were further grouped into a high and low intake group according to the median daily relative (%) intake of protein and the respective amino acids in the same cohort.
Multivariate logistic regression analyses were performed to detect associations of definite NASH on liver biopsy and liver fibrosis with dietary components. We adjusted our model for age, gender, BMI, type 2 diabetes, and arterial hypertension as risk factors for disease progression, relative cholesterol, relative disaccharide, and relative alcohol intake as potential confounding dietary factors, as a higher protein intake was accompanied by a significant different intake of these nutrients, as well as energy misreporting.
The bacterial operational taxonomic units were normalized to get the proportional, relative abundance of each bacterial taxa in each patient for further statistical analysis. Heat map analysis with partial Spearman’s correlation, adjusted for potentially confounding factors, was conducted to correlate the relative bacterial abundance at genus level with protein intake as well as the intake of each specific amino acid. Only taxa with an abundance of at least 2% in the overall cohort were included. To investigate associations of these selected bacteria with clinical variables within the NAFLD cohort, heatmap analysis with Spearman’s correlations was performed. Statistical analysis was performed using R statistical software (R version 3.5.1, 2018, the R Foundation for Statistical Computing). A P value of equal or less than 0.05 was considered as statistically significant.
Results
Increased Protein Intake in Patients With NASH
First, we compared the clinical characteristics and daily dietary macronutrient intake among patients with a NAS of 0‐4 or definitive NASH (NAS 5‐8) on liver biopsy. Sixty‐one patients with biopsy‐proven NAFLD were included. Patients with NASH had a significantly higher age and higher levels of aspartate aminotransferase (AST) and alanine aminotransferase (ALT). The median physical activity level was in the range of light activity( 23 ) and did not differ between the groups (Table 1).
Table 1.
Characteristics of the Study Cohort Based on the NAS
| N/A, n | NAS 0‐4 | NAS 5‐8 | P Value | ||
|---|---|---|---|---|---|
| Total (n) | 35 | 26 | |||
| Demographics | |||||
| Age (years) | 47.5 (19.6) | 58.5 (11.0) | 0.028 | ||
| Gender (n [%]) | Female | 14 (40.0) | 12 (46.2) | 0.631 | |
| BMI (kg/m2) | 29.4 (5.3) | 31.0 (9.0) | 0.057 | ||
| Type 2 diabetes (n [%]) | 4 (11.8) | 8 (30.8) | 0.060 | ||
| Arterial hypertension (n [%]) | 17 (48.6) | 19 (73.1) | 0.054 | ||
| Metabolic syndrome (IDF criteria), n (%) | 10 (28.6) | 13 (52.0) | 0.066 | ||
| Waist circumference (cm) | 9 | 105.0 (17.5) | 111.0 (19.0) | 0.318 | |
| Liver Histology | |||||
| Total n | |||||
| Grade of steatosis (n [%]) | 0 | 0 (0.0) | 0 (0.0) | <0.001 | |
| 1 | 16 (45.7) | 2 (7.7) | |||
| 2 | 16 (45.7) | 10 (38.5) | |||
| 3 | 3 (8.6) | 14 (53.8) | |||
| Ballooning (n [%]) | 0 | 18 (51.4) | 0 (0.0) | <0.001 | |
| 1 | 17 (48.6) | 8 (30.8) | |||
| 2 | 0 (0.0) | 18 (69.2) | |||
| Grade of inflammation (n [%]) | 0 | 10 (28.6) | 0 (0.0) | <0.001 | |
| 1 | 21 (60.0) | 9 (34.6) | |||
| 2 | 4 (11.4) | 16 (61.5) | |||
| 3 | 0 (0.0) | 1 (3.8) | |||
| Stage of fibrosis (n [%]) | 0 | 14 (40.0) | 1 (3.8) | 0.002 | |
| 1 | 13 (37.1) | 7 (26.9) | |||
| 2 | 3 (8.6) | 10 (38.5) | |||
| 3 | 1 (2.9) | 3 (11.5) | |||
| 4 | 4 (11.4) | 5 (19.2) | |||
| Noninvasive Fibrosis Assessment | |||||
| Transient elastography (kPa) | 2 | 5.0 (2.4) | 7.5 (6.6) | 0.009 | |
| Laboratory Parameters | |||||
| Albumin (g/L) | 45.0 (3.5) | 44.5 (3.8) | 0.606 | ||
| Creatinine (mg/dL) | 0.9 (0.3) | 0.9 (0.2) | 0.614 | ||
| Urea (mg/dL) | 28.0 (11.5) | 29.0 (12.5) | 0.339 | ||
| Uric acid (mg/dL) | 6.1 (2.3) | 6.1 (1.9) | 0.834 | ||
| AST (U/L) | 30.0 (14.5) | 39.0 (34.5) | 0.015 | ||
| ALT (U/L) | 41.0 (27.5) | 71.0 (49.0) | 0.009 | ||
| GGT (U/L) | 89.0 (110.5) | 67.0 (48.2) | 0.703 | ||
| Alkaline phosphatase (U/L) | 75.0 (27.5) | 72.0 (18.0) | 0.624 | ||
| Bilirubin (mg/dL) | 1 | 0.5 (0.4) | 0.5 (0.2) | 0.887 | |
| Ferritin (µg/L) | 196.0 (232.5) | 229.5 (131.5) | 0.386 | ||
| Triglycerides (mg/dL) | 169.0 (111.5) | 150.5 (113.5) | 0.599 | ||
| Total cholesterol (mg/dL) | 182.0 (43.5) | 194.5 (61.2) | 0.470 | ||
| HDL cholesterol (mg/dL) | 2 | 50.0 (23.0) | 43.0 (13.8) | 0.120 | |
| LDL cholesterol (mg/dL) | 4 | 109.0 (47.8) | 126.0 (56.0) | 0.222 | |
| Platelet count (x1E9/L) | 215.0 (82.5) | 207.0 (58.5) | 0.197 | ||
| INR | 1 | 1.0 (0.1) | 1.0 (0.0) | 0.125 | |
| HbA1c (%) | 5 | 5.3 (0.6) | 5.5 (0.7) | 0.151 | |
| Fasting glucose (mg/dL) | 93.0 (17.5) | 97.0 (27.5) | 0.196 | ||
| Dietary Composition/Physical Activity | |||||
| Physical activity level | 1.5 (0.2) | 1.5 (0.2) | 0.989 | ||
| Daily energy intake (kcal) | 1679.8 (581.0) | 1739.1 (592.6) | 0.818 | ||
| EI:BMR ratio | 1.1 (0.4) | 1.1 (0.4) | 0.708 | ||
| Protein (%) | 15.8 (5.0) | 18.0 (2.7) | 0.018 | ||
| Protein (g) | 67.1 (29.3) | 79.0 (22.5) | 0.012 | ||
| Carbohydrates (%) | 46.0 (11.4) | 41.9 (8.8) | 0.083 | ||
| Carbohydrates (g) | 188.2 (80.5) | 185.7 (41.4) | 0.634 | ||
| Monosaccharides (%) | 6.0 (4.3) | 5.0 (3.0) | 0.134 | ||
| Monosaccharides (g) | 23.1 (21.6) | 18.1 (13.9) | 0.182 | ||
| Disaccharides (%) | 8.0 (6.0) | 8.9 (5.5) | 0.253 | ||
| Disaccharides (g) | 33.4 (30.9) | 38.0 (32.0) | 0.795 | ||
| Polysaccharides (%) | 24.2 (8.8) | 22.7 (5.9) | 0.347 | ||
| Polysaccharides (g) | 109.2 (41.0) | 104.5 (32.5) | 0.740 | ||
| Fiber (%) | 1.9 (0.7) | 1.9 (1.2) | 0.897 | ||
| Fiber (g) | 16.0 (8.4) | 17.3 (5.6) | 0.746 | ||
| Fructose (%) | 3.0 (2.0) | 2.6 (2.2) | 0.213 | ||
| Fructose (g) | 12.8 (9.8) | 10.9 (8.4) | 0.172 | ||
| Fat (%) | 33.5 (10.8) | 34.7 (7.9) | 0.325 | ||
| Fat (g) | 65.2 (34.7) | 69.6 (25.9) | 0.378 | ||
| PUFA (%) | 5.1 (2.4) | 5.1 (1.9) | 0.284 | ||
| PUFA (g) | 9.3 (6.1) | 10.1 (3.7) | 0.453 | ||
| MUFA (%) | 11.3 (3.7) | 11.8 (3.0) | 0.304 | ||
| MUFA (g) | 21.3 (10.7) | 24.4 (11.4) | 0.221 | ||
| SFA (%) | 15.1 (6.9) | 14.6 (4.1) | 0.829 | ||
| SFA (g) | 29.1 (14.6) | 30.8 (13.7) | 0.762 | ||
| Cholesterol (%) | 0.1 (0.0) | 0.1 (0.1) | 0.453 | ||
| Cholesterol (mg) | 252.5 (131.6) | 282.3 (145.0) | 0.247 | ||
| Alcohol (%) | 0.0 (0.1) | 0.0 (0.1) | 0.763 | ||
| Alcohol (g) | 0.0 (0.5) | 0.0 (0.4) | 0.814 |
Values are presented as the median and interquartile range (in parentheses) for continuous variables or number and percentage (in parentheses) for categorical variables. Wilcoxon Mann‐Whitney U test for continuous variables and chi‐squared tests for categorical variables. Bold font indicates significance (P value < 0.05). Patients with biopsy‐proven NAFLD with a NAS of 5‐8 were compared to patients with a NAS of 0‐4. The NAS is the unweighted sum of the scores for steatosis (0‐3), lobular inflammation (0‐3), and ballooning (0‐2), ranging from 0 to 8. Stage of fibrosis: 0, none; 1, perisinusoidal or periportal; 2, perisinusoidal and portal/periportal; 3, bridging fibrosis; and 4, cirrhosis. Grade of steatosis: 0, <5%; 1, 5%‐33%; 2, >33%‐66%; and 3, >66%. Grade of inflammation: 0, no foci; 1, <2 foci per ×200 field; 2, 2‐4 foci per ×200 field; and 3, >4 foci per ×200 field. Ballooning: 0, none; 1, few balloon cells; and 2, many cells/prominent ballooning. The number of missing values within the overall cohort is indicated in the third column (“N/A, n”).
Abbreviations: and SFA, saturated fatty acids; basal metabolic rate; BMR; EI, energy intake; IDF, International Diabetes Foundation; INR, international normalized ratio; MUFA, monounsaturated fatty acids, N/A, not applicable; PUFA, polyunsaturated fatty acids.
Among all dietary macronutrients, the main difference between these two groups was an increased protein intake in the NAS 5‐8 group (18.0% vs. 15.8% of total protein‐based calories per day; P = 0.018; Table 1). The detailed intake and univariate analysis regarding NASH for each specific amino acid is reported in Supporting Table S1.
We further analyzed clinical characteristics and dietary factors among patients with no to mild fibrosis (F0‐F1) and patients with significant fibrosis to cirrhosis (F2‐F4). Patients with at least significant fibrosis had a higher BMI, were more frequently affected from type 2 diabetes, arterial hypertension and the metabolic syndrome, and had significantly altered laboratory parameters (Supporting Table S2). Among dietary factors, patients in the F2‐F4 group had a significantly higher intake of cysteine, phenylalanine, tryptophan, arginine, glutamic acid, glycine, proline, and serine (Supporting Table S2).
We did not observe an increased intake of fructose, other simple carbohydrates, or saturated fatty acids among patients with NASH (Table 1) or liver fibrosis (Supporting Table S2).
When we compared dietary macronutrients between patients with NAFLD with low (<17.3%) versus high protein intake (≥17.3%) according to the daily median relative protein intake in the cohort with biopsy‐proven NAFLD, we found the gap of calories in the low‐protein group to be settled by carbohydrates. Furthermore, a higher protein intake was also associated with an increased cholesterol intake (Supporting Table S3).
We additionally investigated whether patients with liver cirrhosis had a higher protein intake due to dietary recommendations often given to patients with liver cirrhosis. However, the dietary macronutrient composition did not differ between patients with NAFLD with and without liver cirrhosis (data not shown).
Independent Association of Protein and Specific Amino Acid Intake with Histological NAFLD Activity by Multivariate Analysis
Next, we performed multivariate logistic regression analyses to detect associations of protein intake and amino acids as break‐down products with disease activity and severity on liver histology. We analyzed the nutrient intake as a continuous variable (in energy percentage) using standardized values (increase per 1 SD), and patients were further grouped into a high and low intake group according to the median daily relative (%) intake of protein and the respective amino acids in the same cohort (n = 61). We adjusted our model for age, gender, BMI, type 2 diabetes, and arterial hypertension as risk factors for disease progression, relative cholesterol, relative disaccharide, and relative alcohol intake as potential confounding dietary factors, as a higher protein intake was accompanied by a significant different intake of these nutrients (Supporting Table S3) as well as energy misreporting.
After multivariate adjustment, protein intake analyzed as continuous variable and a high protein intake (≥17.3%) remained associated with the presence of definite NASH on liver biopsy with an odds ratio (OR) for the high intake group of 5.09 (95% confidence interval [CI] 1.22‐21.25; P = 0.026; Table 2 and Fig. 1). Serine, glycine, arginine, proline, phenylalanine, and methionine were the six amino acids with the highest standardized ORs for active NASH (Table 2, Fig 1).
Table 2.
Association of Protein and Amino Acid Intake With the Presence of Definite NASH on Liver Biopsy
| Outcome: NAFLD Activity Score 5‐8 | OR (95% CI, P Value) Continuous (per SD Increase) | OR (95% CI, P value) High vs. Low Intake (Based on Median Intake) |
|---|---|---|
| Protein | 3.02 (1.27‐7.18, P = 0.012) | 5.09 (1.22‐21.25, P = 0.026) |
| Lysine | 2.21 (1.01‐4.8, P = 0.046) | 2.7 (0.77‐9.53, P = 0.122) |
| Leucine | 1.96 (0.91‐4.22, P = 0.085) | 1.69 (0.5‐5.76, P = 0.398) |
| Isoleucine | 2.24 (1‐5.05, P = 0.05) | 1.62 (0.43‐6.18, P = 0.477) |
| Cysteine | 2.34 (1‐5.47, P = 0.05) | 2.48 (0.7‐8.72, P = 0.158) |
| Phenylalanine | 2.52 (1.06‐6, P = 0.037) | 2.25 (0.62‐8.14, P = 0.217) |
| Methionine | 2.51 (1.09‐5.78, P = 0.031) | 4.03 (1.03‐15.83, P = 0.046) |
| Tyrosine | 2.07 (0.94‐4.57, P = 0.07) | 1.7 (0.48‐5.96, P = 0.41) |
| Threonine | 2.25 (1.01‐5, P = 0.046) | 3.01 (0.81‐11.12, P = 0.099) |
| Tryptophane | 2.24 (0.96‐5.23, P = 0.061) | 3.73 (1.04‐13.4, P = 0.044) |
| Valine | 2.27 (1.01‐5.11, P = 0.048) | 2.41 (0.68‐8.55, P = 0.172) |
| Arginine | 2.81 (1.18‐6.73, P = 0.02) | 2.34 (0.64‐8.53, P = 0.199) |
| Histidine | 1.93 (0.9‐4.15, P = 0.093) | 1.93 (0.54‐6.91, P = 0.314) |
| Alanine | 2.36 (1.05‐5.3, P = 0.037) | 3.82 (0.98‐14.92, P = 0.054) |
| Aspartic acid | 2.34 (1.06‐5.2, P = 0.036) | 2.35 (0.65‐8.43, P = 0.19) |
| Glutamic acid | 2.11 (0.94‐4.7, P = 0.069) | 2.15 (0.62‐7.46, P = 0.227) |
| Glycine | 2.84 (1.23‐6.58, P = 0.015) | 4.01 (1.04‐15.43, P = 0.044) |
| Proline | 2.78 (1.22‐6.35, P = 0.015) | 4.01 (1.12‐14.39, P = 0.033) |
| Serine | 2.87 (1.14‐7.22, P = 0.025) | 6.33 (1.6‐25.05, P = 0.009) |
The multivariate logistic regression model was adjusted for age, gender, BMI, type 2 diabetes, and arterial hypertension as risk factors for disease progression, relative cholesterol, relative disaccharide, and relative alcohol intake as potential confounding dietary factors, as a higher protein intake was accompanied by a significant different intake of these nutrients, as well as energy misreporting. Shown are the ORs with 95% CIs per SD increase group regarding the presence of NASH in liver histology (NAS 5‐8 points). Furthermore, patients were categorized into high and low intake groups based on the median intake of the specific amino acid/ protein in the same cohort (n = 61). Accordingly, the following cutoffs separated the high‐intake group from the low‐intake group, corresponding to equal or higher protein: 17.3%, lysine: 1.05%, leucine: 1.22%, isoleucine: 0.75%, cysteine: 0.23%, phenylalanine: 0.71%, methionine: 0.34%, tyrosine: 0.56%, threonine 0.65%, tryptophan >0.18%, valine 0.88%, arginine: 0.86%, histidine: 0.46%, alanine: 0.81%, aspartic acid: 1.38%, glutamic acid: 3.39%, glycine: 0.70%, proline: 1.15%, and serine: 0.78%. The low‐intake group represents the reference level. The NAS is the unweighted sum of the scores for steatosis (0‐3), lobular inflammation (0‐3), and ballooning (0‐2). Of the 61 patients, 26 had a NAS of 5‐8. Bold font indicates significance (P value < 0.05).
Fig. 1.
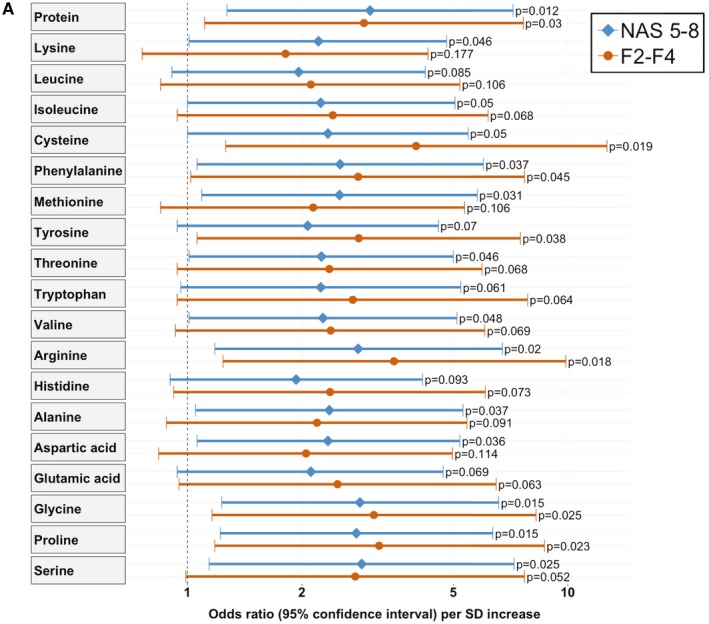
Multivariate analysis to detect associations of protein/amino acid intake with active NASH and fibrosis. (A) The multivariate logistic regression model was adjusted for age, gender, BMI, type 2 diabetes, and arterial hypertension as risk factors for disease progression, relative cholesterol, relative disaccharide, and relative alcohol intake as potential confounding dietary factors, as a higher protein intake was accompanied by a significant different intake of these nutrients, as well as energy misreporting. Shown are the ORs with 95% CIs per SD increase group regarding the presence of NASH in liver histology (NAS, 5‐8 points) and liver fibrosis stage F2‐F4. The NAS is the unweighted sum of the scores for steatosis (0‐3), lobular inflammation (0‐3), and ballooning (0‐2). Of the 61 patients, 26 had a NAS of 5‐8 and 26 were staged as fibrosis F2‐F4.
We further investigated independent associations of protein and amino acids with liver fibrosis. When analyzing protein as a continuous variable, a higher protein intake was associated with higher odds for having liver fibrosis (OR per SD increase: 2.91 (95% CI 1.11‐7.64, P = 0.03). Particularly, several specific amino acids were independently associated with at least moderate fibrosis on liver biopsy, and the intake of cysteine, arginine, proline, glycine, and tyrosine were independently associated with fibrosis when using these dietary variables both dichotomized and as continuous variables (Table 3 and Fig. 1).
Table 3.
Association of Protein and Amino Acid Intake With the Presence of Liver Fibrosis on Liver Biopsy
| Outcome: Fibrosis Stage F2‐F4 | OR (95% CI, P Value) Continuous (per SD Increase) | OR (95% CI, P Value) High vs. Low Intake (Based on Median Intake) |
|---|---|---|
| Protein | 2.91 (1.11‐7.64, P = 0.03) | 3.75 (0.77‐18.26, P = 0.101) |
| Lysine | 1.81 (0.76‐4.29, P = 0.177) | 3.8 (0.9‐16.09, P = 0.07) |
| Leucine | 2.11 (0.85‐5.2, P = 0.106) | 3.68 (0.87‐15.59, P = 0.077) |
| Isoleucine | 2.41 (0.94‐6.17, P = 0.068) | 3.54 (0.77‐16.32, P = 0.105) |
| Cysteine | 3.99 (1.26‐12.67, P = 0.019) | 4.58 (1.04‐20.18, P = 0.044) |
| Phenylalanine | 2.81 (1.02‐7.69, P = 0.045) | 5.47 (1.13‐26.62, P = 0.035) |
| Methionine | 2.14 (0.85‐5.35, P = 0.106) | 7.17 (1.43‐35.81, P = 0.016) |
| Tyrosine | 2.82 (1.06‐7.5, P = 0.038) | 3.96 (0.89‐17.58, P = 0.071) |
| Threonine | 2.36 (0.94‐5.95, P = 0.068) | 5.67 (1.21‐26.68, P = 0.028) |
| Tryptophane | 2.72 (0.94‐7.84, P = 0.064) | 10.54 (2.07‐53.69, P = 0.005) |
| Valine | 2.38 (0.93‐6.05, P = 0.069) | 4.97 (1.1‐22.39, P = 0.037) |
| Arginine | 3.49 (1.24‐9.88, P = 0.018) | 10.05 (1.86‐54.39, P = 0.007) |
| Histidine | 2.37 (0.92‐6.07, P = 0.073) | 9.17 (1.69‐49.63, P = 0.01) |
| Alanine | 2.19 (0.88‐5.43, P = 0.091) | 5.94 (1.22‐28.94, P = 0.027) |
| Aspartic acid | 2.05 (0.84‐4.97, P = 0.114) | 3.53 (0.8‐15.67, P = 0.097) |
| Glutamic acid | 2.48 (0.95‐6.48, P = 0.063) | 6.33 (1.32‐30.4, P = 0.021) |
| Glycine | 3.09 (1.16‐8.25, P = 0.025) | 21.44 (3.1‐148.27, P = 0.002) |
| Proline | 3.19 (1.18‐8.68, P = 0.023) | 8.99 (1.86‐43.39, P = 0.006) |
| Serine | 2.76 (0.99‐7.69, P = 0.052) | 5.74 (1.27‐25.94, P = 0.023) |
The multivariate logistic regression model was adjusted for age, gender, BMI, type 2 diabetes, and arterial hypertension as risk factors for disease progression, relative cholesterol, relative disaccharide, and relative alcohol intake as potential confounding dietary factors, as a higher protein intake was accompanied by a significant different intake of these nutrients, as well as energy misreporting. Shown are the ORs with 95% CIs per SD increase group regarding the presence of at least moderate liver fibrosis (F2‐F4) on liver biopsy. Furthermore, patients were categorized into high‐intake and low‐intake groups based on the median intake of the specific amino acid/ protein in the same cohort (n = 61). Accordingly, the following cutoffs separated the high‐intake group from the low‐intake group, corresponding to equal or higher protein: 17.3%, lysine: 1.05%, leucine: 1.22%, isoleucine: 0.75%, cysteine: 0.23%, phenylalanine: 0.71%, methionine: 0.34%, tyrosine: 0.56%, threonine 0.65%, tryptophan >0.18%, valine 0.88%, arginine: 0.86%, histidine: 0.46%, alanine: 0.81%, aspartic acid: 1.38%, glutamic acid: 3.39%, glycine: 0.70%, proline: 1.15%, and serine: 0.78%. The low‐intake group represents the reference level. Shown are the ORs with 95% CIs regarding the presence of liver fibrosis stage F2‐F4 on liver biopsy. Bold font indicates significance (P value < 0.05).
A high relative intake of carbohydrates, fructose, overall fat, and cholesterol was not associated with disease activity and severity in the univariate analysis (Table 1) or after adjustment for age, gender, BMI, type 2 diabetes, and arterial hypertension (Supporting Table S4).
Gut Microbiota Alterations Associated With Protein and Specific Amino Acid Intake
Next, we investigated associations of dietary protein/amino acids with alterations in the gut bacterial microbiota. A total of 99 patients with NAFLD were included in this analysis, whereas 59 of these patients were overlapping patients with biopsy‐proven NAFLD who were also included in the regression analyses. Clinical characteristics for these patients are given in Supporting Table S5. We performed a heatmap analysis reporting partial Spearman’s correlations between the total dietary protein and break‐down amino acids with the relative abundance of intestinal bacteria with at least 2% relative abundance. This analysis was adjusted for total energy intake, age, gender, BMI, type 2 diabetes, arterial hypertension, cholesterol, alcohol, and disaccharide intake, as well as energy misreporting (in similarity to the multivariate regression analysis) as potentially confounding factors. Protein and several dietary amino acids were negatively or positively correlated with the abundance of specific bacteria (Fig. 2C). Most consistently, Bacteroidaceae (family) and Bacteroides (genus) were negatively correlated with the intake of protein and several amino acids (Fig. 2A‐C). In contrast, for example, Gemmiger (genus) was positively associated with an increased intake of protein and amino acids (Fig. 2A,D).
Fig. 2.
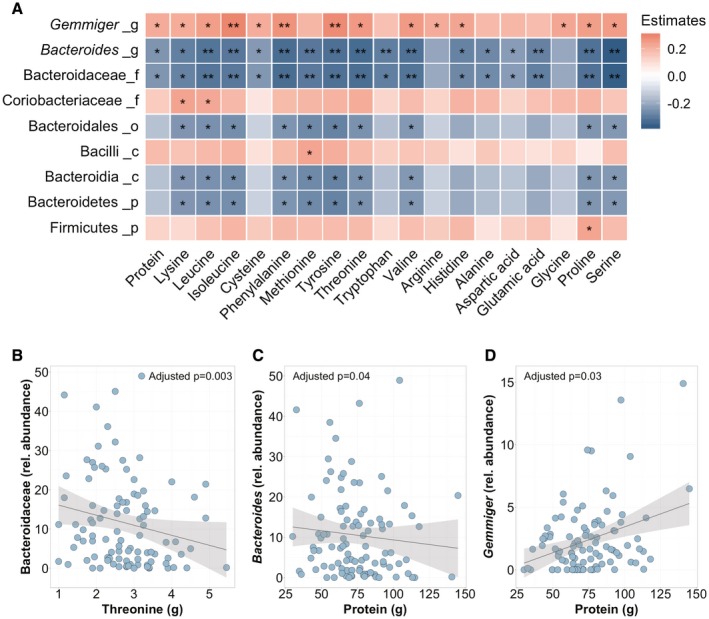
Alterations in gut bacterial microbiota associated with the intake of protein and specific amino acids. (A) Heatmap representing color‐coded partial Spearman’s correlations of dietary protein and amino acids with the abundance of intestinal bacteria. A total of 99 patients with NAFLD were included. The analysis was adjusted for total energy intake, age, gender, BMI, type 2 diabetes, arterial hypertension, cholesterol, alcohol, and disaccharide intake as well as energy misreporting (in similarity to the multivariate regression analysis) as potentially confounding factors. Red color indicates positive, and blue color negative, correlation. *P < 0.05, P > 0.01; **P < 0.01, P > 0.001. A total of 99 patients with NAFLD were included. Only taxa with a relative abundance of over 2% and significant associations were included in the heatmap. The level within the bacterial kingdom is indicated following the taxa name (_p, phylum; _c, class; _o, order; _f, family; and _g, genus). (B) Negative correlation of threonine intake with relative abundance of Bacteroidaceae. (C) Negative correlation of protein intake with relative abundance of Bacteroides. (D) Positive correlation of protein intake with relative abundance of Gemmiger.
Protein‐Related Gut Microbiota Alterations Are Associated With Clinical Characteristics in Patients With NAFLD
After identifying associations of nutritional protein and amino acid intake with distinct alterations of the gut microbiota, we further correlated these bacteria with clinical variables within the NAFLD cohort. Bacteroidaceae (family) and Bacteroides (genus) were associated with a higher high‐density lipoprotein (HDL) and lower levels of ALT, gamma‐glutamyl‐transferase (GGT), and ferritin levels (Fig. 3A‐D). In contrast, Gemmiger (genus), which correlates with a higher protein intake, was positively associated with obesity and an increased AST, ALT, and ferritin levels (Fig. 3A,E‐G).
Fig. 3.
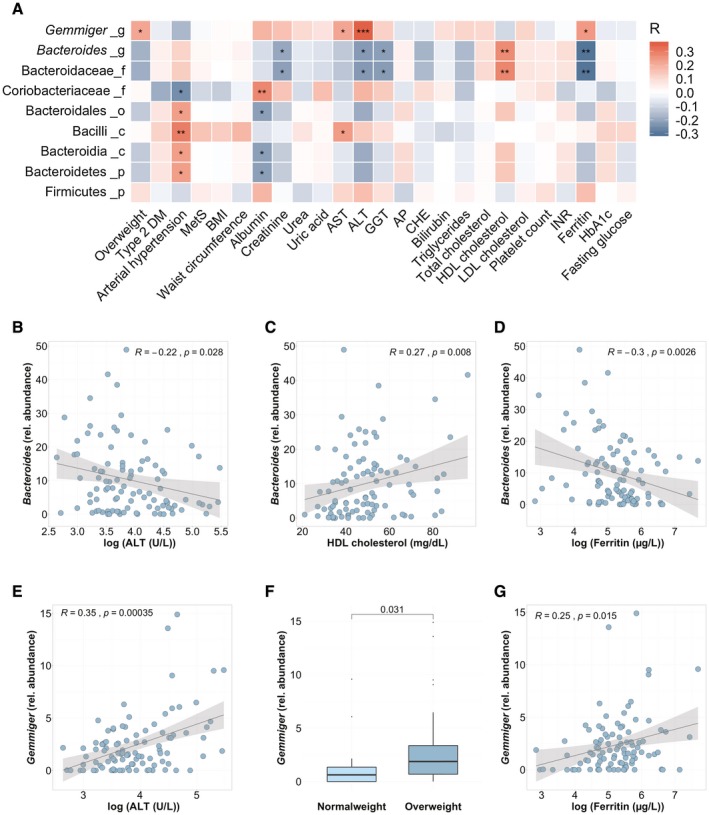
Correlations of altered bacterial taxa with clinical parameters in patients with NAFLD. Heatmap representing color‐coded Spearman’s correlations of bacterial taxa from Fig. 1, which are associated with an increased protein or amino acid intake, with clinical variables in patients with NAFLD. A total of 99 patients with NAFLD were included. Red color indicates positive, and blue color negative, correlation. *P < 0.05, P > 0.01; **P < 0.01, P > 0.001; ***P < 0.001. The level within the bacterial kingdom is indicated following the taxa name (_p, phylum; _c, class; _o, order; _f, family; and _g, genus). Correlations of the relative Bacteroides abundance with logarithmic ALT (B), HDL cholesterol (C), and logarithmic ferritin (D) levels. Correlations of the relative abundance of Gemmiger with logarithmic ALT (E), overweight (F), defined as BMI > 25 kg/m2 (n = 87) versus normal‐weight (BMI < 25 kg/m2) patients with NAFLD, and logarithmic ferritin levels (G). Abbreviations: AP, alkaline phosphatase; CHE, cholinesterase; DM, diabetes mellitus; INR, international normalized ratio; LDL, low‐density lipoprotein; MetS, metabolic syndrome.
A decreased Bacteroides abundance was associated with an increased protein intake. The abundance of several altered bacteria further correlates with clinical parameters in patients with NAFLD.
Discussion
The dietary pattern of patients with NAFLD with increased disease activity and severity on liver biopsy was characterized by a higher intake of protein compared to those with low histological activity and severity. These observations were independent from cofactors that also contribute to disease progression as well as potential dietary confounders. Similar observations could not be made for fat, carbohydrates, fructose, and cholesterol. A decreased Bacteroides abundance was independently associated with a higher dietary intake of protein and associated with clinical features of patients with NAFLD.
Our data thereby indicate that protein plays a role in NAFLD development and/or progression. In line with our observations, it has been shown previously that patients with NAFLD consume higher amounts of protein. In the recently published Rotterdam study including 3,882 patients, total protein intake was associated with the presence of NAFLD, and this association was particularly driven by the consumption of animal protein.( 12 ) High intake of meat, and in particular red meat, was further associated with the presence of NAFLD and insulin resistance in a study of 789 participants undergoing screening colonoscopy.( 13 ) A large population‐based study including 536,969 participants revealed an increased all‐cause, but in particular liver‐related, mortality for subjects ingesting higher amounts of unprocessed and processed red meat.( 27 ) Notably, meat and other animal products serve as the main source of protein in Western diet with varying concentrations of specific amino acids,( 11 , 14 , 15 ) whereas a vegetarian or vegan diet leads to a significantly lower intake of total and specific dietary amino acids.( 14 ) Not all amino acids were significantly associated with histological disease activity and severity in our study, but several amino acids were close to a P value equal or lower than 0.05. Future studies are required to clarify whether other amino acids will be significantly associated with histological activity when increasing the sample size and therefore increasing statistical power. Dichotomizing variables allows easier interpretation of results, but is also accompanied by a higher risk for false‐positive results and a possible loss of information,( 28 , 29 ) which might also be reflected in the fact that protein intake was only significantly associated with fibrosis when analyzing protein as a continuous variable. Another explanation as to why only specific amino acids were associated with increased disease severity might be that protein from different food sources substantially varies in the amino acid concentration and composition. Certain amino acids such as glycine, methionine, lysine, and tryptophan are predominantly contained in meat/animal products( 15 , 30 ) and might therefore inherit a higher risk for disease progression.
This study provides a description of associations between NAFLD severity according to liver histology criteria and protein/amino acids, which are compounds that are primarily components of meat and animal products in a Western diet. Definite NASH, the form of NAFLD with a high risk for disease progression, which is diagnosed when a NAS reaches ≥ 5 points,( 21 ) was independently associated with a higher protein and specific amino acid intake in our study.
Interestingly, dietary fructose consumption was not associated with disease activity or severity. In a global analysis including 43 countries, the United States has been found to ingest the highest per capita amount of high fructose corn syrup, whereas Germany was among the lowest ranked countries.( 31 ) With a median daily intake of 11.5 g/day (range 0.85‐50.9), corresponding to 3% of the daily calorie intake, patients with NAFLD in our study had a much lower fructose consumption compared with the mean dietary fructose intake of the U.S. population, which was reported to be 54.7 g/day (range, 38.4‐72.8), accounting for 10.2% of the total calorie intake.( 32 ) Thus, the critical values for a high fructose intake of more than 50 g/day might not apply for a Western European diet, and the pathogenetic influence of fructose on NAFLD in Western Europe might be smaller than previously assumed.
Unlike the average fructose intake, the protein consumption of populations living in the United States/Canada and the European Union is comparable, representing the overall highest ranks worldwide.( 11 ) We also could not detect a harmful association of carbohydrates and fat with histological disease activity.
The underlying mechanisms of how amino acids might contribute to disease progression in NAFLD are unclear. One potential way amino acids could interfere with metabolic pathways in hepatocytes is through induction of the mammalian target of rapamycin (mTOR) complex that is known to regulate cell growth.( 33 ) Beyond up‐regulation in hepatic malignancies,( 34 ) mTOR plays a role in hepatic lipogenesis,( 35 ) and chronic mTOR complex 1 (mTORC1) activation leads to insulin resistance in hepatic and adipose tissue,( 33 ) both of which play a key role in the pathogenesis of NAFLD. To what extend dietary amino acids might contribute to mTORC1 activation, however, requires further mechanistic investigation. The metabolic response to amino acids might be different depending on the prevailing anabolic or catabolic condition. In catabolic states, amino acids represent energy substrates, while in anabolic conditions, protein synthesis and cell growth is induced.( 36 )
Dietary amino acids further interact with the commensal gut microbiota. Gut bacteria regulate host bioavailability of amino acids by fermentation and transformation, while producing metabolites such as short‐chain fatty acids, hydrogen sulfide, phenolic and indolic compounds, ammonia, polyamines, biogenic amines, and nitrogen oxide.( 18 ) In addition, intestinal microbes play a role in de novo biosynthesis of amino acids.( 37 , 38 ) On the other hand, amino acids influence the composition of this microbial ecosystem itself. In line with our results, one study investigating the effect of a high protein diet on the gut microbiota found a decreased abundance of Bacteroides.( 39 ) The findings from our study that demonstrate a close association between specific amino acid intake with typical microbial changes in NAFLD suggests that the protein‐driven adaptation of the gut microbiota is a main mechanism linking nutritional proteins with NAFLD progression.
Gemmiger is a gram‐negative bacterium belonging to the ruminococcaceae family that has been associated with the presence of early hepatocellular carcinoma( 40 ) and relapse of Crohn’s disease after surgery.( 41 ) Gemminger was positively correlated with an increased intake in our cohort with NAFLD and was associated with increased liver enzymes, ferritin, and overweight. A decreased Bacteroides abundance in patients with NAFLD compared with healthy controls was reported in several other studies.( 42 , 43 ) In our study, patients with a higher intestinal Bacteroides abundance had lower liver enzymes and serum ferritin levels. Gut dysbiosis has been linked to NAFLD in several studies, but the results are inconsistent.( 44 ) The gut microbiota might be a potential mediator between protein intake and effects on clinical outcome. Although we could show correlations with only this type of study design, preclinical studies are needed to explore causal relationships. The gut microbiota is also a key for the production of trimethylamine (TMA) from meat and other dietary lecithin sources.( 45 ) TMA is oxidized by hepatic flavin‐containing monooxygenases to trimethylamine N‐oxide (TMAO), a potent driver of atherosclerosis.( 46 ) Dismissing red meat consumption leads to a sharp decrease in TMAO plasma levels within 4 weeks.( 47 ) In light of the increased cardiovascular mortality, a reduction in red meat consumption offers additional health benefits for patients with NAFLD.
The strength of our study is the very well‐characterized study cohort, which allowed us to adjust for major risk factors for disease progression. The 14‐day dietary report provides very detailed and precise information about food choices, with advantages compared with food frequency questionnaires.( 48 , 49 ) However, dietary assessment in general is vulnerable to distortion; even if the patients were instructed in detail on how to use the dietary report, high motivation is needed to follow the requirements. Thus, underreporting or miscalculation of food portions might have occurred, and even if we adjusted our multivariate analyses for energy misreporting, this might have biased our results. The analysis relies on honesty and accuracy of the patient’s disclosures. We cannot rule out the possibility that a higher protein intake is also associated with other confounding factors related to a more unhealthy lifestyle that we did not consider, such as a possibly underreported alcohol intake. For this reason, and because of the limited sample size, our data are not enough to draw final conclusions. With a cross‐sectional design, we further cannot conclude causality for the observed associations. Other limitations address the initial study design, which did not aim to investigate associations of dietary categories such as meat with disease activity. Thus, we could not take the proportion of animal or vegetable protein into account, which would have been interesting as well.
In conclusion, a higher intake of dietary protein, and in particular specific amino acids, is associated with disease activity and severity in patients with NAFLD. Similar observations could not be made for fructose consumption, which has been closely linked to NAFLD pathogenesis so far. The abundance of specific intestinal bacteria correlated with the intake of protein and specific amino acids. Mechanistic investigations are needed to assess potential causality behind the observed associations. Meat and other animal products that contain high concentrations of amino acids might be critical “parallel hits,” driving simple steatosis to NASH. If further studies confirm a potentially harmful role, these compounds should be taken into consideration when giving dietary recommendations to patients with NAFLD.
Supporting information
Table S1‐S5
Funding was provided by the Köln Fortune Research Pool, Faculty of Medicine, University of Cologne, Germany (160/2014) and Marga und Walter Boll‐Stiftung (210‐03‐2016). The study was supported by the NIDDK‐funded San Diego Digestive Diseases Research Center (P30 DK120515).
Potential conflict of interest: Dr. Schnabl consults for Ferring, Intercept Pharma, HOST Therabiomics, and Patara Pharma. He received grants from BiomX, NGM Bio, CymaBay Therapeutics, and Synglogic Operation Company. Dr. Vehreschild consults for, is on the speakers’ bureau, and received grants from Astella and MSD. He consults for and received grants from MaaT Pharma and DaVolterra. He is on the speakers’ bureau for and received grants from Gilead and Organobalance. He consults for Alb‐Fils Kliniken GmbH, Arderypharm, and Ferring. He is on the speakers’ bureau for Akademie fur Infektionsmedizin, Arztekammer Nordrhein, Basilea, and Pfizer. He received grants from 3M, Evoinik, Glycom, Morphochem, and Seres Therapeutics.
References
Author names in bold designate shared co‐first authorship.
- 1. Vernon G, Baranova A, Younossi ZM. Systematic review: the epidemiology and natural history of non‐alcoholic fatty liver disease and non‐alcoholic steatohepatitis in adults. Aliment Pharmacol Ther 2011;34:274‐285. [DOI] [PubMed] [Google Scholar]
- 2. Tiniakos DG, Vos MB, Brunt EM. Nonalcoholic fatty liver disease: pathology and pathogenesis. Annu Rev Pathol 2010;5:145‐171. [DOI] [PubMed] [Google Scholar]
- 3. Ekstedt M, Franzen LE, Mathiesen UL, Thorelius L, Holmqvist M, Bodemar G, et al. Long‐term follow‐up of patients with NAFLD and elevated liver enzymes. Hepatology 2006;44:865‐873. [DOI] [PubMed] [Google Scholar]
- 4. Wong VW, Wong GL, Choi PC, Chan AW, Li MK, Chan HY, et al. Disease progression of non‐alcoholic fatty liver disease: a prospective study with paired liver biopsies at 3 years. Gut 2010;59:969‐974. [DOI] [PubMed] [Google Scholar]
- 5. Harrison SA, Oliver D, Arnold HL, Gogia S, Neuschwander‐Tetri BA. Development and validation of a simple NAFLD clinical scoring system for identifying patients without advanced disease. Gut 2008;57:1441‐1447. [DOI] [PubMed] [Google Scholar]
- 6. Angulo P, Hui JM, Marchesini G, Bugianesi E, George J, Farrell GC, et al. The NAFLD fibrosis score: a noninvasive system that identifies liver fibrosis in patients with NAFLD. Hepatology 2007;45:846‐854. [DOI] [PubMed] [Google Scholar]
- 7. Valenti L, Al‐Serri A, Daly AK, Galmozzi E, Rametta R, Dongiovanni P, et al. Homozygosity for the patatin‐like phospholipase‐3/adiponutrin I148M polymorphism influences liver fibrosis in patients with nonalcoholic fatty liver disease. Hepatology 2010;51:1209‐1217. [DOI] [PubMed] [Google Scholar]
- 8. Loomba R, Abraham M, Unalp A, Wilson L, Lavine J, Doo E, et al. Association between diabetes, family history of diabetes, and risk of nonalcoholic steatohepatitis and fibrosis. Hepatology 2012;56:943‐951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. McCarthy EM, Rinella ME. The role of diet and nutrient composition in nonalcoholic Fatty liver disease. J Acad Nutr Diet 2012;112:401‐409. [DOI] [PubMed] [Google Scholar]
- 10. Jensen T, Abdelmalek MF, Sullivan S, Nadeau KJ, Green M, Roncal C, et al. Fructose and sugar: a major mediator of non‐alcoholic fatty liver disease. J Hepatol 2018;68:1063‐1075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Ranganathan J, Vennard D, Waite R, Dumas P, Lipinski B, Searchinger T. Shifting diets for a sustainable food future. Working Paper, Installment 11 of Creating a Sustainable Food Future. Washington, DC: World Resources Institute; 2016. [Google Scholar]
- 12. Alferink LJ, Kiefte‐de Jong JC, Erler NS, Veldt BJ, Schoufour JD, de Knegt RJ, et al. Association of dietary macronutrient composition and non‐alcoholic fatty liver disease in an ageing population: the Rotterdam Study. Gut 2019;68:1088‐1098. [DOI] [PubMed] [Google Scholar]
- 13. Zelber‐Sagi S, Ivancovsky‐Wajcman D, Fliss Isakov N, Webb M, Orenstein D, Shibolet O, et al. High red and processed meat consumption is associated with non‐alcoholic fatty liver disease and insulin resistance. J Hepatol 2018;68:1239‐1246. [DOI] [PubMed] [Google Scholar]
- 14. Schmidt JA, Rinaldi S, Scalbert A, Ferrari P, Achaintre D, Gunter MJ, et al. Plasma concentrations and intakes of amino acids in male meat‐eaters, fish‐eaters, vegetarians and vegans: a cross‐sectional analysis in the EPIC‐Oxford cohort. Eur J Clin Nutr 2016;70:306‐312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Gorska‐Warsewicz H, Laskowski W, Kulykovets O, Kudlinska‐Chylak A, Czeczotko M, Rejman K. Food products as sources of protein and amino acids—the Case of Poland. Nutrients 2018;10:1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Gaggini M, Carli F, Rosso C, Buzzigoli E, Marietti M, Della Latta V, et al. Altered amino acid concentrations in NAFLD: impact of obesity and insulin resistance. Hepatology 2018;67:145‐158. [DOI] [PubMed] [Google Scholar]
- 17. Graf D, Di Cagno R, Fak F, Flint HJ, Nyman M, Saarela M, et al. Contribution of diet to the composition of the human gut microbiota. Microb Ecol Health Dis 2015;26:26164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Ma N, Ma X. Dietary amino acids and the gut‐microbiome‐immune axis: physiological metabolism and therapeutic prospects. J Compr Rev Food Sci Food Saf 2019;18:221‐242. [DOI] [PubMed] [Google Scholar]
- 19. Lang S, Demir M, Duan Y, Martin A, Schnabl B. Cytolysin‐positive Enterococcus faecalis is not increased in patients with non‐alcoholic steatohepatitis. Liver Int 2020. Jan 13. doi: 10.1111/liv.14377. [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. EASL‐EASD‐EASO . Clinical Practice Guidelines for the management of non‐alcoholic fatty liver disease. J Hepatol 2016;64:1388‐1402. [DOI] [PubMed] [Google Scholar]
- 21. Kleiner DE, Brunt EM, Van Natta M, Behling C, Contos MJ, Cummings OW, et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005;41:1313‐1321. [DOI] [PubMed] [Google Scholar]
- 22. Freedman LS, Schatzkin A, Midthune D, Kipnis V. Dealing with dietary measurement error in nutritional cohort studies. J Natl Cancer Inst 2011;103:1086‐1092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Westerterp KR. Physical activity and physical activity induced energy expenditure in humans: measurement, determinants, and effects. Front Physiol 2013;4:90‐90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Hill RJ, Davies PS. The validity of self‐reported energy intake as determined using the doubly labelled water technique. Br J Nutr 2001;85:415‐430. [DOI] [PubMed] [Google Scholar]
- 25. Banna JC, McCrory MA, Fialkowski MK, Boushey C. Examining plausibility of self‐reported energy intake data: considerations for method selection. Front Nutr 2017;4:45‐45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. McNaughton SA, Mishra GD, Brunner EJ. Food patterns associated with blood lipids are predictive of coronary heart disease: the Whitehall II study. Br J Nutr 2009;102:619‐624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Etemadi A, Sinha R, Ward MH, Graubard BI, Inoue‐Choi M, Dawsey SM, et al. Mortality from different causes associated with meat, heme iron, nitrates, and nitrites in the NIH‐AARP Diet and Health Study: population based cohort study. BMJ 2017;357:j1957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Austin PC, Brunner LJ. Inflation of the type I error rate when a continuous confounding variable is categorized in logistic regression analyses. Stat Med 2004;23:1159‐1178. [DOI] [PubMed] [Google Scholar]
- 29. Altman DG, Royston P. The cost of dichotomising continuous variables. BMJ 2006;332:1080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Gorissen SHM, Crombag JJR, Senden JMG, Waterval WAH, Bierau J, Verdijk LB, et al. Protein content and amino acid composition of commercially available plant‐based protein isolates. Amino Acids 2018;50:1685‐1695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Goran MI, Ulijaszek SJ, Ventura EE. High fructose corn syrup and diabetes prevalence: a global perspective. Glob Public Health 2013;8:55‐64. [DOI] [PubMed] [Google Scholar]
- 32. Vos MB, Kimmons JE, Gillespie C, Welsh J, Blanck HM. Dietary fructose consumption among US children and adults: the Third National Health and Nutrition Examination Survey. Medscape J Med 2008;10:160. [PMC free article] [PubMed] [Google Scholar]
- 33. Saxton RA, Sabatini DM. mTOR signaling in growth, metabolism, and disease. Cell 2017;168:960‐976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Bhat M, Sonenberg N, Gores GJ. The mTOR pathway in hepatic malignancies. Hepatology 2013;58:810‐818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Li Z, Xu G, Qin Y, Zhang C, Tang H, Yin Y, et al. Ghrelin promotes hepatic lipogenesis by activation of mTOR‐PPARgamma signaling pathway. Proc Natl Acad Sci USA 2014;111:13163‐13168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Bifari F, Nisoli E. Branched‐chain amino acids differently modulate catabolic and anabolic states in mammals: a pharmacological point of view. Br J Pharmacol 2017;174:1366‐1377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Davila AM, Blachier F, Gotteland M, Andriamihaja M, Benetti PH, Sanz Y, et al. Intestinal luminal nitrogen metabolism: role of the gut microbiota and consequences for the host. Pharmacol Res 2013;68:95‐107. [DOI] [PubMed] [Google Scholar]
- 38. Dai ZL, Wu G, Zhu WY. Amino acid metabolism in intestinal bacteria: links between gut ecology and host health. Front Biosci (Landmark Ed) 2011;16:1768‐1786. [DOI] [PubMed] [Google Scholar]
- 39. Russell WR, Gratz SW, Duncan SH, Holtrop G, Ince J, Scobbie L, et al. High‐protein, reduced‐carbohydrate weight‐loss diets promote metabolite profiles likely to be detrimental to colonic health. Am J Clin Nutr 2011;93:1062‐1072. [DOI] [PubMed] [Google Scholar]
- 40. Ren Z, Li A, Jiang J, Zhou L, Yu Z, Lu H, et al. Gut microbiome analysis as a tool towards targeted non‐invasive biomarkers for early hepatocellular carcinoma. Gut 2019;68:1014‐1023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Mondot S, Lepage P, Seksik P, Allez M, Treton X, Bouhnik Y, et al. Structural robustness of the gut mucosal microbiota is associated with Crohn's disease remission after surgery. Gut 2016;65:954‐962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Del Chierico F, Nobili V, Vernocchi P, Russo A, Stefanis C, Gnani D, et al. Gut microbiota profiling of pediatric nonalcoholic fatty liver disease and obese patients unveiled by an integrated meta‐omics‐based approach. Hepatology 2017;65:451‐464. [DOI] [PubMed] [Google Scholar]
- 43. Da Silva HE, Teterina A, Comelli EM, Taibi A, Arendt BM, Fischer SE, et al. Nonalcoholic fatty liver disease is associated with dysbiosis independent of body mass index and insulin resistance. Sci Rep 2018;8:1466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Chu H, Williams B, Schnabl B. Gut microbiota, fatty liver disease, and hepatocellular carcinoma. Liver Res 2018;2:43‐51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Tang WH, Wang Z, Levison BS, Koeth RA, Britt EB, Fu X, et al. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N Engl J Med 2013;368:1575‐1584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Koeth RA, Wang Z, Levison BS, Buffa JA, Org E, Sheehy BT, et al. Intestinal microbiota metabolism of L‐carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med 2013;19:576‐585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Wang Z, Bergeron N, Levison BS, Li XS, Chiu S, Jia X, et al. Impact of chronic dietary red meat, white meat, or non‐meat protein on trimethylamine N‐oxide metabolism and renal excretion in healthy men and women. Eur Heart J 2019;40:583‐594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Kowalkowska J, Slowinska MA, Slowinski D, Dlugosz A, Niedzwiedzka E, Wadolowska L. Comparison of a full food‐frequency questionnaire with the three‐day unweighted food records in young Polish adult women: implications for dietary assessment. Nutrients 2013;5:2747‐2776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Barrett‐Connor E. Nutrition epidemiology: how do we know what they ate? Am J Clin Nutr 1991;54:182s‐187s. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1‐S5