

Abstract
The Keap1/Nrf2/ARE system is a central defensive mechanism against oxidative stress which plays a key role in the pathogenesis and progression of many diseases. Nrf2 is a redox-sensitive transcription factor controlling a variety of downstream antioxidant and cytodefensive genes. Nrf2 has a powerful anti-inflammatory activity mediated via modulating NF-κB. Therefore, pharmacological activation of Nrf2 is a promising therapeutic strategy for the treatment/prevention of several diseases that are underlined by both oxidative stress and inflammation. Coumarins are natural products with promising pharmacological activities, including antioxidant, anticancer, antimicrobial, and anti-inflammatory efficacies. Coumarins are found in many plants, fungi, and bacteria and have been widely used as complementary and alternative medicines. Some coumarins have shown an ability to activate Nrf2 signaling in different cells and animal models. The present review compiles the research findings of seventeen coumarin derivatives of plant origin (imperatorin, visnagin, urolithin B, urolithin A, scopoletin, esculin, esculetin, umbelliferone, fraxetin, fraxin, daphnetin, anomalin, wedelolactone, glycycoumarin, osthole, hydrangenol, and isoimperatorin) as antioxidant and anti-inflammatory agents, emphasizing the role of Nrf2 activation in their pharmacological activities. Additionally, molecular docking simulations were utilized to investigate the potential binding mode of these coumarins with Keap1 as a strategy to disrupt Keap1/Nrf2 protein-protein interaction and activate Nrf2 signaling.
1. Introduction
Coumarins are organic compounds in the benzopyrone class. This group comprises a large number of compounds that are widely distributed in the plant kingdom and have the highest concentration in fruits, seeds, roots, and leaves [1]. The name coumarin has been derived from the French word “Coumarou,” the common name of tonka beans (Dipteryx odorata) [2, 3]. The number of coumarins identified as secondary metabolites in bacteria, fungi, and about 150 species of plants has been estimated to be more than 1300 compounds [4]. Coumarins are most abundant in those plants taxonomically assigned to the Apiaceae, Asteraceae, and Rutaceae families and play a significant role in human health. Coumarins have been widely used in complementary and alternative medicine and possess a diversity of pharmacological activities with low cost and few side effects [5–7]. Carrot, cherries, citrus fruits, apricots, celery, parsnip, and strawberries, as well as spices like cinnamon and fennels are some of the commonly consumed coumarin-containing phytofoods [8].
Natural coumarins are classified into 6 main types based on their chemical structure. These include simple coumarins, furanocoumarins, dihydrofuranocoumarins, phenylcoumarins, pyranocoumarins, and biscoumarins [6]. All have a coumarin core and are characterized by structural diversity which could be considered for drug discovery and development of therapeutic agents for multiple diseases [9–12]. In plants, coumarins have been suggested to function as growth regulators and bacterio- and fungistatic agents [13]. In addition, coumarins possess a broad range of pharmacological activities basically relying on the type of coumarin nucleus. The beneficial effects of coumarins include antimicrobial [14–17], antimutagenic [12, 18], anti-inflammatory [19, 20], anticoagulant [21], antithrombotic [22–24], vasodilatory [25, 26], and anticancer activities [27]. Inhibition of matrix metalloproteinases (MMPs) and cancer cell growth, migration, and invasion and induction of apoptosis have been demonstrated as the effects underlying the anticancer activity of coumarins [28, 29]. Coumarins have also shown antihyperglycemic, antifibrotic, antiadipogenic, and cytochrome P450 inhibitory activities [30–34]. In a mouse model of cerebral ischemia/reperfusion (I/R) injury, the coumarin esculetin showed a potent neuroprotective effect when administered intracerebroventricularly [35].
The antioxidant and anti-inflammatory activities of coumarins have been well-acknowledged in several in vitro and in vivo studies [36, 37]. Coumarins suppress oxidative stress through their ability to scavenge reactive oxygen species (ROS) and inhibit neutrophil-dependent superoxide anion generation and lipid peroxidation. Moreover, coumarins can effectively reduce tissue edema-associated inflammation through suppressing both lipoxygenase and cyclooxygenase enzymatic activities and prostaglandin synthesis and release [20, 38–40].
Oxidative stress is a state of imbalance between the production of free radicals and their degradation by antioxidants. This redox imbalance occurs as a result of increased ROS generation and diminished antioxidant defenses. Although produced normally through different metabolic processes, excess ROS can provoke inflammation and damage lipids, proteins, and other cellular macromolecules, leading to oxidative stress and cell death. Therefore, oxidative stress is implicated in the pathogenesis of a wide range of metabolic disorders and chronic diseases [41–43]. Given their ability to suppress excessive ROS generation and enhance antioxidants [30, 34, 44], the pharmacologic effects of coumarins could be mediated through their antioxidant efficacy. The present review presents an overview of the modulatory role of a number of plant-derived coumarins (Figure 1) on nuclear factor (erythroid-derived 2)-like 2 (Nrf2), a transcription factor which protects against oxidative injury and inflammation [45]. In addition, we investigated the potential binding mode of coumarins to Kelch-like ECH-associated protein 1 (Keap1) as a strategy to disrupt Keap1/Nrf2 protein-protein interaction (PPI) using molecular docking simulations.
Figure 1.
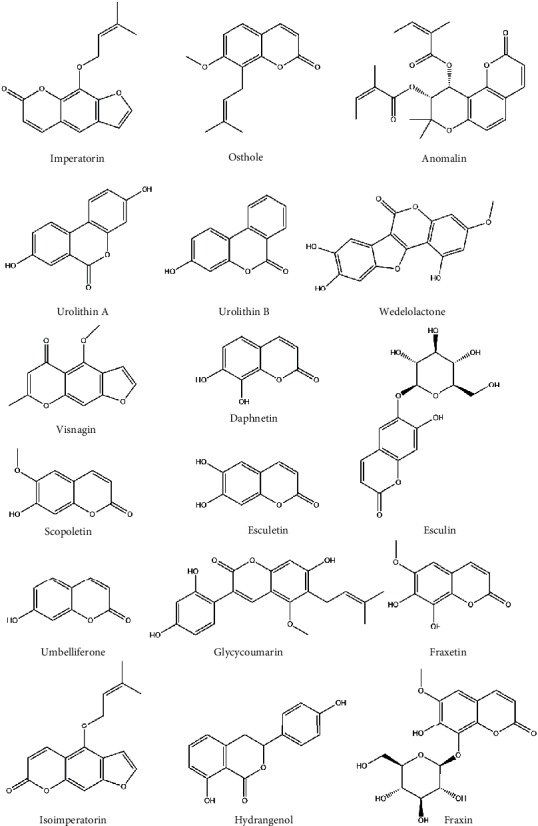
Chemical structure of selected coumarin-derived compounds.
2. Keap1/Nrf2/ARE Signaling Pathway
Nrf2 is a transcription factor generally known to enhance the cellular defense system to counteract oxidative injury and inflammation. In conditions without oxidative stimuli, Nrf2 exists in the cytoplasm sequestered by Keap1 [46]. Keap1 mediates Nrf2 ubiquitination and subsequent proteasomal degradation through acting as an adaptor molecule for the CUL-E3 ligase. The dissociation of Keap1 from the CUL-E3 ligase is elicited upon exposure to electrophilic/oxidative stress which modifies the cysteine residues of Keap1, in particular Cys151, leading to accumulation of Nrf2 [47]. As a result, Nrf2 liberates and translocates into the nucleus where it binds to the antioxidant response element (ARE) and promotes the transcription of antioxidant genes [48] (Figure 2). Nrf2-target antioxidant genes include heme oxygenase-1 (HO-1), NAD(P)H dehydrogenase quinone 1 (NQO1), γ-glutamyl cysteine ligase modulatory and catalytic subunits (GCLM and GCLC, respectively), and ferritin, which function to maintain the oxidant/antioxidant balance inside the cells [48]. Thus, activation of Nrf2 is an effective strategy to suppress oxidative stress.
Figure 2.
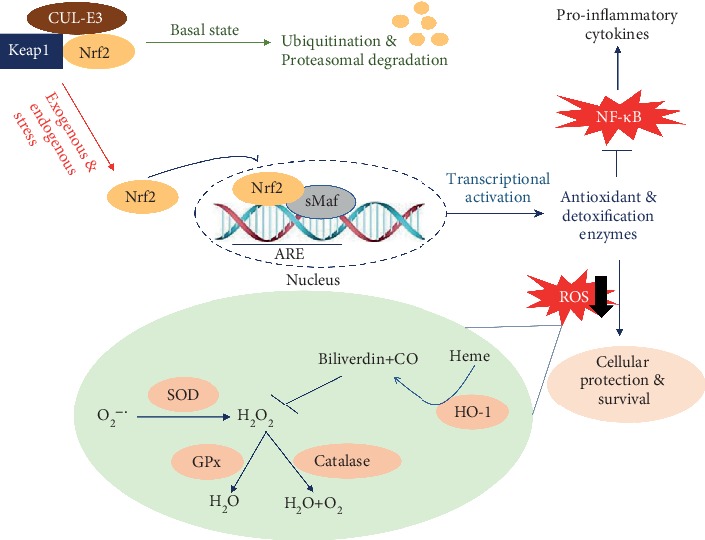
Nrf2 activity is augmented by exogenous and/or endogenous stressors. Under basal conditions, Keap1 mediates Nrf2 ubiquitination and subsequent proteasomal degradation through acting as an adaptor molecule for CUL-E3 ligase. Upon exposure to exogenous and/or endogenous stressors, such as xenobiotics and ROS, respectively, Nrf2 translocates into the nucleus and binds to the ARE to activate cytoprotective molecules, including antioxidant and detoxification enzymes. Superoxide dismutase (SOD) mediates the dismutation of superoxide radicals (O2-·) leading to the formation of hydrogen peroxide (H2O2). Catalase and glutathione peroxidase (GPx) catalyze the degradation of H2O2. HO-1 catalyzes degradation of heme to biliverdin and bilirubin which are potential antioxidants [49]. CO: carbon monoxide.
Besides attenuation of oxidative stress, Nrf2 activation can effectively prevent inflammation. Nrf2 signaling is regarded as the most sensitive redox pathway linked to oxidative injury and nuclear factor-κB (NF-κB), a master regulator of proinflammatory mediators. Both are redox-sensitive factors where NF-κB is activated in oxidative stress conditions, and the lack of Nrf2 resulted in increased oxidative/nitrosative stress and subsequently amplification of cytokine production [50]. The crosstalk between Nrf2 and NF-κB has been reviewed by Wardyn et al. [51]. The lack of Nrf2 can aggravate NF-κB activity leading to increased inflammatory cytokine release [52], whereas Nrf2 upregulation resulted in diminished inflammatory responses in rodent models of liver and kidney injury [53–60]. The Nrf2 target gene HO-1 has been demonstrated to inhibit NF-κB-mediated transcription of adhesion molecules possibly through decreasing free intracellular iron in endothelial cells [61]. NF-κB activity can regulate Nrf2-mediated antioxidant gene expression. In this context, Yu et al. [62] have demonstrated that overexpression of the canonical NF-κB subunit p65 increased nuclear Keap1 levels resulting in decreased Nrf2/ARE signaling. Given that Keap1 is mostly localized in the cytosol and proteins larger than 40 kDa cannot enter through the nuclear envelope, the nuclear translocation of Keap1 has been suggested to occur via interaction with karyopherin alpha 6 (KPNA6). In this context, overexpression of KPNA6 has been associated with decreased HO-1 and NQO1 [63]. Although different mechanisms of the inhibitory effect of NF-κB p65 on Nrf2 have been postulated, a competition for the transcriptional coactivator CBP-p300 complex is the best supported mechanism. CBP-p300 has an intrinsic acetyl transferase activity and acetylates Nrf2 and p65 [64, 65].
3. The Modulatory Effect of Coumarins on Nrf2
In this section, we reviewed studies showing the role of Nrf2 signaling in mediating the pharmacologic effects of coumarins. Only coumarin derivatives reported to modulate Nrf2 signaling were included.
3.1. Imperatorin (IMP)
IMP (9-(3-methylbut-2-enoxy)furo[3,2-g]chromen-7-one) is a furanocoumarin present in high concentration in plants of the genus Angelica such as Angelica archangelica and Angelica dahurica. The pharmacological properties of IMP make it a promising candidate for drug development. It has been documented to possess antioxidant [66, 67], anti-inflammatory [68, 69], anticancer [68, 70], antibacterial [71], and antiallergic activities [72]. Singh et al. reported that IMP treatment significantly reduced the nociceptive behavior provoked with formalin and acetic acid [73]. IMP effectively reduced the lipopolysaccharide- (LPS-) induced rise in IL-1β and TNF-α levels in mice in a dose-dependent manner [73]. In a mouse model of paw edema induced by carrageenan, a potent anti-inflammatory activity of IMP was evident 4 h after the injection of carrageenan [73]. In ovalbumin- (OA-) challenged mice and LPS-induced dendritic cells (DCs), the antiallergic and anti-inflammatory effects of IMP were demonstrated [72]. In asthmatic mice, IMP, in a dose-dependent manner, reduced IgE levels, airway hyperresponsiveness, and Th2 cytokines and increased IL-10-producing T cells. In LPS-stimulated DCs, IMP increased IL-10 and suppressed the release of proinflammatory cytokines [72]. Li et al. have reported that IMP effectively diminished COX-2, IL-6, TNF-α, and iNOS expression via downregulation of NF-κB and JAK/STAT signaling in alveolar macrophages [74]. In addition, IMP relieved zymosan-induced immune cell infiltration in mice, thereby attenuating lung inflammation, edema, and fibrosis [74]. Although these studies have documented the potent anti-inflammatory activity of IMP, none of them have pointed to the involvement of Nrf2 signaling. The ability of IMP to activate Nrf2 signaling has been evaluated in few studies. By using HepG2 stably transfected with ARE reporter and Nrf2 knockout mice, Prince et al. have studied whether IMP activates hepatic Nrf2. The results showed that IMP increased hepatic GST activity via the Nrf2/ARE mechanism [75]. Hu et al. reported that IMP-induced Nrf2 activation effectively upregulated its downstream antioxidants, possibly offering cellular protection to the heart from injury induced by arsenic trioxide (As2O3) [76]. Very recently, Xian et al. investigated the protective effect of IMP against excess ROS and chronic airway inflammation in OA-administered mice [77]. Treatment with IMP suppressed ROS, proinflammatory cytokines, inflammatory cell infiltration, collagen deposition, and goblet cell hyperplasia. These effects have been suggested to be mediated via modulation of Nrf2 signaling following IMP administration [77].
3.2. Visnagin
Visnagin (4-methoxy-7-methylfuro[3,2-g]chromen-5-one) is a bioactive compound extracted from Ammi visnaga fruits [78]. Visnagin possesses widespread pharmacological activities, including hypotensive and smooth muscle relaxation. In a dose-dependent manner, visnagin decreased blood pressure when administered intravenously. In isolated mesenteric arteries precontracted with noradrenaline, visnagin treatment resulted in a concentration-dependent relaxation [79]. In isolated rat aortic rings, visnagin inhibited the vascular smooth muscle contraction induced by different agents [80]. Visnagin has also shown protective effects against doxorubicin cardiotoxicity mediated via cytochrome P450 family 1 (CYP1) inhibition [81] and modulation of mitochondrial malate dehydrogenase [82]. In addition, visnagin prevented the deposition of renal crystals in hyperoxaluric rats [83]. Besides these activities, the antioxidant and anti-inflammatory activities of visnagin have been demonstrated. In this context, Lee et al. have examined the anti-inflammatory activity of visnagin in BV2 microglial cells challenged with LPS. The obtained results demonstrated that visnagin ameliorates LPS-induced TNF-α, IL-1β, iNOS, IFN-γ, and IL-6 expression via a NF-κB-dependent mechanism and increases the production of the anti-inflammatory cytokine IL-10 [84]. A recent study by Khalil et al. evaluated the anti-inflammatory activity of visnagin using molecular docking simulation and in vivo and in vitro assays [85]. Visnagin exhibited a strong anti-inflammatory activity with a high safety profile in vivo and showed an in vitro selective COX-2 inhibition [85]. The role of Nrf2 in mediating the anti-inflammatory and antioxidant activities of visnagin has been recently reported by Pasari et al. using a model of cerulein-induced acute pancreatitis in mice [86]. Visnagin decreased the expression of IL-1β, TNF-α, IL-6, and IL-17 in a dose-dependent manner. Importantly, visnagin enhanced the antioxidant defenses by effective upregulation of Nrf2 and mitigated pancreatic inflammation through suppression of the NF-κB expression in the acinar cells. Additionally, visnagin inhibited the release of inflammatory cytokines in pulmonary and intestinal tissues [86].
3.3. Urolithin B
Urolithin B (3-hydroxybenzo[c]chromen-6-one) is one of the gut microbial metabolites of ellagitannins, a class of compounds found in many plants, including medicinal herbs, pomegranates, and tropical fruits [87, 88]. Ellagitannin is hydrolyzed to ellagic acid which is then catabolized by microflora in the intestinal tract into urolithin derivatives (urolithin A-D) [89]. Urolithin B is the final product catabolized among the urolithin derivatives that possesses anticancer activity on prostate [90] and colon cancer [91]. Treatment with urolithin B induced apoptosis of HT-29 colon cancer cells in vitro [91]. The anticancer activity of urolithin B was mediated through disruption of the mitochondrial membrane potential and activation of caspases 8, 9, and 3 [91]. In addition, urolithin B exhibited potent anti-inflammatory [92] and antioxidant effects [93] and has been reported to attenuate neurotoxicity in Alzheimer's disease by preventing β-amyloid fibrillation in vitro [94]. The dual anti-inflammatory and antioxidant effect of urolithin B has been recently investigated by Lee et al. [92] in microglia challenged with LPS. Urolithin B reduced cytokine release and NO production, while it significantly increased the expression of IL-10. In addition, urolithin B significantly decreased TNF-α, IL-6, and NO production in stimulated BV2 cells. Additionally, the in vivo studies reported that urolithin B inhibited LPS-induced microglia activation in the mouse brain [92]. The study has also scrutinized the antioxidant activity of urolithin B. The results showed a strong antioxidant activity mediated by reducing intracellular ROS production and NADPH oxidase subunit expression through upregulation of Nrf2/ARE signaling and HO-1 expression. Interestingly, urolithin B suppressed the activity of NF-κB by reducing IκBα phosphorylation. In addition, urolithin B inhibited both ERK and JNK phosphorylation while it enhanced the AMPK phosphorylation which is associated with a dual anti-inflammatory and antioxidant activity [92].
3.4. Urolithin A
Urolithin A (3,8-dihydroxybenzo[c]chromen-6-one) is the one of the main microbiota metabolites of pomegranate ellagitannins. It is characterized by high safety as evidenced by a previous study carried out by Heilman et al. [95]. Urolithin A has a wide range of bioactivities, such as antioxidant [96], anti-inflammatory [97, 98], and anticancer effects [99]. In human colonic fibroblasts stimulated with IL-1β, urolithin A inhibited PGE2 production, downregulated COX-2, and suppressed NF-κB nuclear translocation [97]. In human aortic endothelial cells exposed to TNF-α, urolithin A showed a great ability to inhibit monocyte adhesion, chemokine expression, and cell migration [98]. Urolithin A improved the gut microbiota in high fat diet- (HFD-) fed rats and reversed the LPS-induced inflammatory response in Caco2 cells [100]. Fu et al. have conducted a study to examine the role of urolithin A on IL-1β-induced inflammatory response in human osteoarthritis through in vitro and in vivo approaches. In human chondrocytes in vitro, urolithin A prevented the IL-1β-induced overproduction of TNF-α, IL-6, PGE2, COX-2, iNOS, and NO in a concentration-dependent manner via suppressing NF-κB activation [101]. Another recent study carried out by Gong et al. concluded that urolithin A attenuates memory impairment and neuroinflammation in APP/PS1 mice via suppressing p38 MAPK and NF-κB p65 activation along with enhancing cerebral AMPK activation [102]. Two recent studies have reported the impact of Nrf2 activation by urolithin A. The first study was done by Liu et al. who demonstrated that urolithin A is a promising antiaging agent through potent inhibition of intracellular ROS promoted by activation of the Nrf2-mediated antioxidative response [103]. The second study done by Singh et al. concluded that urolithin A can be used for the treatment of colitis through remedying barrier dysfunction [104]. This study showed the anti-inflammatory effects of urolithin A and highlighted the role of the activation of Nrf2-dependent pathways [104].
3.5. Scopoletin
Scopoletin (7-hydroxy-6-methoxychromen-2-one) is a natural coumarin found in many medicinal plants, including members of the genus Scopolia as well as species of the Artemisia, Brunfelsia, Solanum, and Mallotus and other genera. Scopoletin is present in many edible plants and foods, such as oats, garlic, lemon, grapefruit, celery, red pepper, chili pepper, carrots, chicory, and bael [105–107]. It possesses many pharmacological functions, including antimicrobial [108], antiaging [109], anti-inflammatory [110, 111], and antioxidant effects [112, 113]. Scopoletin is also known for its cytotoxic activity against different cancer cells [114, 115].
Scopoletin has also shown antihyperglycemic activity in diabetic mice and inhibited α-glucosidase in vitro [116]. The antidiabetic activity of scopoletin has been suggested to be exerted via inhibition of the carbohydrate digestive enzymes [116]. In the same context, scopoletin improved insulin sensitivity and enhanced glucose uptake through activation of GLUT4 translocation and PI3K and AMPK signaling in 3T3-L1 adipocytes in vitro [117]. The supplementation of scopoletin prevented hepatic steatosis in diabetic mice by inhibiting lipid biosynthesis and TLR4-MyD88 pathways [118]. The anti-inflammatory property of scopoletin was shown in vivo in various animal studies [110, 111, 118, 119]. The anti-inflammatory activity of scopoletin has been reported to involve suppression of myeloperoxidase (MPO), a neutrophil infiltration biomarker; adenosine-deaminase (ADA); TNF-α; IL-1β; and NO via inhibition of the NF-κB and p38 MAPK phosphorylation. In a rat model of osteoarthritis, scopoletin downregulated collagenases and reduced the proinflammatory mediators in a dose-dependent manner [119]. In human fibroblasts, scopoletin inhibited p38 phosphorylation, MMP-1, NF-κB, MAPK, and the mRNA abundance of IL-1α and TNF-α [120]. In a mouse model of cerulein-induced acute pancreatitis and lung injury, scopoletin suppressed pancreatic and pulmonary TNF-α, IL-1β, mast cell activation, and NF-κB signaling [111]. The ability of scopoletin to upregulate Nrf2/HO-1 signaling has been supported by few studies. Given the antidiabetic efficacy of scopoletin, Chang et al. have investigated its insulin sensitizing and antiglycation effects in diabetic rats, pointing to the role of Nrf2 signaling [121]. Scopoletin suppressed the formation of advanced glycation end products (AGEs), hyperglycemia, and insulin resistance and enhanced Nrf2, Akt, and GLUT2 in hepatocytes [121]. Very recently, Narasimhan et al. reported that scopoletin protected against oxidative stress and apoptosis induced by rotenone via Nrf2 activation and investigated its neuroprotective effects for Parkinson's disease in a rat model and in vitro using SH-SY5Y cells [122].
3.6. Daphnetin
Daphnetin (7,8-dihydroxychromen-2-one) is one of the coumarin derivatives extracted from Daphne Korean Nakai [123, 124]. It has been clinically used in the treatment of rheumatoid arthritis, lumbago, and coagulation disorders and as an antipyretic [125–127]. It possesses multiple pharmacological properties, including anti-inflammatory and oxidant activities [128, 129], and demonstrated a significant anticancer effect in vitro [130, 131]. In a rat model of collagen-induced arthritis, daphnetin suppressed joint destruction, synovial hyperplasia, and Th1/Th2/Th17-type cytokines in splenic lymphocytes and increased the expression of Foxp3 [125], demonstrating a potent anti-inflammatory activity. Treatment of arthritic rats with daphnetin resulted in reduced levels of TNF-α, IL-1β, and macrophage migration inhibitory factor (MIF) [126]. Shen et al. reported that daphnetin decreased LPS-induced inflammation and reduced endotoxin lethality in mice. In this study, daphnetin suppressed TNF-α, IL-1β, IL-6, NO, and PGE2 release along with the expression of iNOS and COX-2 and inhibited ROS production in Raw264.7 cells [129].
Zhang et al. have demonstrated the protective effect of daphnetin against cisplatin nephrotoxicity via suppressing oxidative injury and inflammation [132]. Daphnetin significantly inhibited cisplatin-induced ROS generation, lipid peroxidation, NF-κB activation, and the levels of IL-1β and TNF-α production in a dose-dependent manner. These effects have been accompanied with upregulation of Nrf2 and HO-1 expression [132]. Another study by Liu et al. reported that daphnetin protected against NAFLD in vitro in oleic acid-treated hepatocytes through Nrf2 activation while it effectively decreased CYP2E1 and CYP4A expression [133]. Daphnetin showed a potent inhibitory effect on oleic acid-induced ROS generation and promoted glucose uptake, insulin sensitivity, and PI3K/Akt signaling in hepatocytes [133]. In support of these findings, daphnetin mitigated oxidative stress in human lung epithelial cells exposed to arsenic through Keap1 protein downregulation and marked activation of the Nrf2-dependent antioxidant response with a dramatic upregulation of the ARE in a dose-dependent manner [134]. Daphnetin inhibited oxidative stress and inflammatory response in high glucose- (HG-) stimulated human glomerular mesangial cells (MCs) [135]. Daphnetin strikingly reduced ROS and decreased the production of IL-1β, IL-6, and TNF-α via suppression of the NF-κB pathway. Mechanistically, daphnetin positively upregulated Nrf2 while it inhibited the expression Keap1 in HG-stimulated MCs [135]. Zhi et al. reported that daphnetin protected hippocampal neurons exposed to oxygen-glucose deprivation-induced injury against I/R via marked enhancement of the nuclear translocation of Nrf2 and HO-1 expression [136]. Moreover, Nrf2 knockdown blocked the protective effect of daphnetin on I/R in hippocampal neurons, confirming the critical role of Nrf2/HO-1 signaling activation in the neuroprotective effect of daphnetin [136]. Lv and coworkers demonstrated that daphnetin effectively inhibited cytochrome c release and NLRP3 inflammasome activation through upregulation of the Nrf2 nuclear translocation along with Keap1 protein downregulation [137]. Additionally, daphnetin suppressed ROS generation induced by tert-butyl hydroperoxide (t-BHP) which is mostly blocked in Nrf2 knockout macrophages. Accordingly, daphnetin has a protective role against t-BHP-induced oxidative injury via the Nrf2/ARE signaling pathway [137]. Furthermore, daphnetin ameliorated carbon tetrachloride- (CCl4-) induced hepatotoxicity in rats through induction of the nuclear translocation of Nrf2, thus inducing HO-1 expression [138]. These studies supported the notion that Nrf2 activation is critical for the protective mechanism of daphnetin against oxidative injury and inflammation induced by several insults.
3.7. Esculin
Esculin (7-hydroxy-6-[(2S,3R,4S,5S,6R)-3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxychromen-2-one) is a coumarin derivative found in Aesculus hippocastanum L. (horse-chestnut). The anti-inflammatory [139] and antioxidant [140, 141] activities of esculin have been well-acknowledged. By using the in vitro ABTS, ORAC, and DPPH assays, Zhang et al. have demonstrated the potent antiradical activity of esculin [141]. In a rat model of colon carcinogenesis, esculin mitigated oxidative stress, DNA damage, and tumorigenesis [140]. Additionally, esculin showed wide pharmacological activities against different diseases, such as cognitive impairment in experimental diabetic nephropathy where it exhibited a strong anti-inflammatory activity marked by the suppressed p38 MAPK and JNK [142]. In models of streptozotocin-induced renal damage in diabetic mice [143], cold-restrained stress and pylorus ligation-induced ulcer [144], ethanol-induced gastric lesion [145], and LPS/D-galactosamine-induced acute liver injury [146], esculin significantly ameliorated inflammation evidenced by the suppressed TNF-α, IL-1β, and MPO. Li et al. reported the protective role of esculin against LPS-induced macrophages and endotoxin shock in mice and NO production in vitro. Esculin inhibited the LPS-induced increase in TNF-α and IL-6 and upregulated IL-10 via suppression of NF-κB [147]. Another study by Li et al. concluded that esculin markedly inhibited iNOS/NO levels, and NF-κB protein expression in gastric injury induced by alcohol [145]. Additionally, pretreatment with esculin suppressed TNF-α and IL-6 expression [145].
Pertaining to the impact of esculin on Nrf2 activation, Liu et al. have shown that esculin attenuated acute liver injury in mice induced by LPS/D-galactosamine and reduced pathological symptoms of acute hepatic injury via suppression of NF-κB expression as well as activation of Nrf2/HO-1 signaling [146]. Another study by Kim et al. reported that esculin activated Nrf2/ARE signaling in macrophages [148]. Additionally, esculin markedly inhibited neutrophilic lung inflammation which was not recapitulated in Nrf2 knockout mice, suggesting that the anti-inflammatory activity of esculin mainly acts via Nrf2 activation [148].
3.8. Esculetin
Esculetin (6,7-dihydroxychromen-2-one) is one of the main bioactive ingredients of Cortex Fraxini. Esculetin exhibits a potent antioxidant effect and showed a scavenging activity against DPPH radicals in a time- and concentration-dependent manner [149]. Esculetin has been widely used in antitussive aspects [150], and the study of Liang et al. has demonstrated its protective effect against oxidative stress-induced DNA damage [151]. In addition, esculetin possesses anti-inflammatory [152, 153], antibacterial [154, 155], and antitumor activities against different cancer cells in vitro [156–158] and in vivo [159, 160]. It has also been reported to enhance the inhibitory effect of 5-fluorouracil on the proliferation of colorectal cancer [161]. The anti-inflammatory activity of esculetin was effective in the inhibition of cartilage destruction in rheumatoid arthritis and osteoarthritis where it suppressed MMP-1 expression in cartilage and decreased NO and PGE2 levels in the synovium [162]. In the context of obesity, esculetin attenuated chronic inflammation by suppressing proinflammatory cytokine release during the interaction between adipocytes and macrophages [163]. In psoriatic mouse skin, esculetin attenuated the disease progression and dramatically decreased proinflammatory cytokines, including TNF-α, IL-6, IL-22, IL-23, IL-17A, and IFN-γ [152].
Several studies have supported the involvement of Nrf2 signaling in the pharmacological activities of esculetin. For instance, the study of Rubio et al. illustrated the different roles of NF-κB and Nrf2 in the antioxidant imbalance produced by esculetin on leukemia cells and concluded that esculetin resulted in a significant increase in the nuclear translocation of Nrf2 [164]. A recent study conducted by Xu et al. showed that esculetin attenuated neurological defects and alleviated cognitive impairments in transient bilateral common carotid artery occlusion in mice via Nrf2 activation and markedly ameliorated mitochondrial fragmentation and stress [165]. Sen et al. demonstrated the role of esculetin in attenuating the progression of diabetic nephropathy via Nrf2 activation and inhibition of HG-induced ROS production [166]. Han et al. reported that esculetin protected against H2O2-induced ROS accumulation in the myoblasts through the activation of the Nrf2/NQO1 pathway [167]. Treatment of the pancreatic cancer cell lines with esculetin resulted in significant inhibition of cell proliferation, intracellular ROS, and protein levels of NF-κB [168]. Additionally, esculetin increased the Nrf2 and NQO1 gene expression as well as Nrf2 nuclear accumulation and induced mitochondrial-dependent apoptosis [168]. The same study has shown the binding ability of esculetin to directly bind Keap1 as evidenced by molecular docking and in vitro assays [168]. Subramaniam and Ellis reported that esculetin protects HepG2 cells against H2O2-induced injury via activation of the Nrf2/NQO1 pathway [169]. Pretreatment of the HepG2 cells with esculetin preserved cell integrity following exposure to H2O2 and increased the nuclear accumulation of Nrf2 [169].
3.9. Umbelliferone (UMB)
UMB (7-hydroxychromen-2-one) is a coumarin widely spread in plants belonging to the family Umbelliferae. The Umbelliferae family is inclusive of economically important herbs, such as celery, cumin, fennel, parsley alexanders, angelica, asafoetida, and giant hogweed [9, 170]. UMB possesses a variety of bioactivities, and several investigators have previously reported its in vivo antioxidant and anti-inflammatory effects and studied the underlying mechanisms of action in several animal models [30, 171–175]. The anticancer activity of UMB in hepatocellular [176], colon [177], and oral [178] carcinomas has been well-acknowledged.
Several researchers have demonstrated the strong anti-inflammatory activity of UMB. The in vivo anti-inflammatory activity of UMB has recently been reported by Wang et al. who showed its protective effect against acute lung injury induced by LPS [179]. Another study by Yin et al. reported that UMB inhibited inflammation in diabetic mice through suppressing NF-κB and TLR-4. In addition, UMB mitigated hepatic oxidative injury via activating the Nrf2-mediated signal pathway [180]. Li et al. reported that UMB significantly attenuated ROS accumulation and cytotoxicity induced by methylglyoxal (MG) through the activation of Nrf2/ARE signaling [181]. Depletion of Nrf2 by siRNA markedly inhibited the protective effect of UMB against MG-induced alterations, suggesting the key role of Nrf2 in mediating UMB's activity [181]. The role of Nrf2 has been supported by the study of Sen et al. showing that UMB attenuated the progression of diabetic nephropathy via Nrf2 activation and inhibition of ROS production induced by HG [166]. Additionally, Mohamed et al. reported that UMB ameliorated CCl4-induced hepatotoxicity in rats through induction of the nuclear translocation of Nrf2, thereby ameliorating oxidative stress-related liver injury via enhancement of cellular antioxidant defenses [138]. Furthermore, UMB protected against renal injury induced by methotrexate (MTX) and attenuated oxidative injury via downregulation of Keap1 and upregulation of Nrf2. The results have also shown that UMB inhibited inflammatory responses via downregulation of both NF-κB and p38 MAPK in the kidney of MTX-intoxicated rats [174].
Previous work from our lab has demonstrated the antioxidant and anti-inflammatory activities of UMB in different animal models [30, 34, 44]. UMB administration mitigated cyclophosphamide-induced oxidative damage and inflammatory response through marked elevation of Nrf2, HO-1, and PPARγ expression. UMB attenuated lipid peroxidation, enhanced antioxidants, and suppressed serum proinflammatory mediators and hepatic iNOS and NF-κB expression. The results showed that coactivation of Nrf2 and PPARγ represents the main mechanism underlying the hepatoprotective effect of UMB [44]. In a rat model of CCl4-induced hepatic fibrosis, UMB mitigated inflammation, oxidative injury, and collagen deposition. In addition, UMB suppressed NF-κB p65 and TGF-β1/Smad3 and upregulated hepatic PPARγ [34]. Moreover, UMB attenuated lipid peroxidation, NO release, and cerebral inflammation and downregulated nNOS and soluble guanylate cyclase expression in the cerebrum of hyperammonemic rats [30].
3.10. Fraxetin
Fraxetin (7,8-dihydroxy-6-methoxychromen-2-one) is a simple coumarin compound extracted from the traditional medicinal plant Fraxinus rhynchophylla. Fraxetin is widely available and of relatively low cost and with few side effects. Fraxetin has received recent attention for its antitumor [182–184], radical-scavenging [67, 185–189], anti-inflammatory[190], and antibacterial activities [191]. Fraxetin has also been reported to protect against liver fibrosis induced by CCl4 via suppression of the NF-κB signaling pathway as well as phosphorylation of MAPK proteins [192].
The role of Nrf2 in mediating the pharmacologic effects of fraxetin has been demonstrated in few studies. In this context, the protective effect of fraxetin on oxidative stress induced by Plasmodium berghei infection in mice has been investigated by Singh et al. [193]. Postinfection treatment of the mice with fraxetin suppressed lipid peroxidation and boosted GSH and antioxidant enzymes. The authors have also reported a significant increase in the serum Nrf2-antioxidant response element level [193]. The effect of fraxetin on HO-1 expression in HaCaT human keratinocytes has been investigated by Kundu et al. [194]. Fraxetin activated the Nrf2/HO-1 pathway in HaCaT cells, induced the nuclear translocation of Nrf2, and increased the ARE-reporter gene activity [194]. Thuong et al. reported that fraxetin inhibited vascular proliferation and atherosclerosis and upregulated HO-1 in vascular smooth muscle cells (VSMCs) [195]. Subcellular fractionation and reporter gene analysis using an ARE construct demonstrated that fraxetin upregulated Nrf2 and reporter activity and concluded that fraxetin has direct protective properties against LDL oxidation via Nrf2/ARE activation [195].
3.11. Fraxin
Fraxin (7-hydroxy-6-methoxy-8-[(2S,3R,4S,5S,6R)-3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxychromen-2-one) is the main bioactive component of the Chinese traditional herb Cortex Fraxini [196]. This natural coumarin displayed inspiring biological activities, including anti-inflammatory [197], antioxidant [197, 198], and antihyperuricemic activities [199].
The dual antioxidant and anti-inflammatory activity of fraxin has been recently demonstrated in a mouse model of acute respiratory distress syndrome (ARDS) [200]. Fraxin inhibited the production of TNF-α, IL-1β, IL-6, ROS, and MDA; suppressed NF-κB, MAPK signaling, and MMP9; and increased SOD in the lung of mice with LPS-induced ARDS [200]. Niu et al. reported that fraxin exhibited hepatoprotective effects against CCl4-induced liver damage via mitigation of oxidative stress and inflammation [197]. Fraxin alleviated hepatic injury as indicated by the suppressed production of inflammatory mediators and enhancement of the antioxidant defense mechanisms. An in vitro study demonstrated that pretreatment of HepG2 with fraxin protected against the deleterious effects of CCl4. Fraxin inhibited CCl4-induced MAPK, NF-κB, and COX-2 protein expression [197]. In two different studies conducted by Li et al., the protective effect of fraxin against LPS-induced endotoxic shock [201] and acute lung injury in mice [202] has been investigated. In both studies, the ameliorative effect of fraxin was associated with decreased release of proinflammatory mediators, suppressed ROS generation and oxidative stress, and downregulated NF-κB and NLRP3 inflammasome signaling pathways. These findings highlighted the potent suppressive effect of fraxin on both inflammatory and oxidative responses [201, 202]. In an animal model of I/R-induced kidney injury, fraxin exhibited an ameliorative effect mediated through suppressing oxidative DNA damage and NF-κB [203]. Furthermore, Chang et al. have pointed to the role of Nrf2 in mediating the hepatoprotective efficacy of fraxin [198]. In their study, fraxin markedly inhibited the t-BHP-induced cytotoxicity and ROS generation in HepG2 through Nrf2 pathway-dependent HO-1 expression. In vivo studies showed that fraxin has potent hepatoprotective effects against CCl4-induced hepatotoxicity in rats via direct antioxidant activity and the Nrf2/ARE pathway [198].
3.12. Anomalin
Anomalin ([(9R,10R)-8,8-dimethyl-9-[(Z)-2-methylbut-2-enoyl]oxy-2-oxo-9,10-dihydropyrano[2,3-f]chromen-10-yl] (Z)-2-methylbut-2-enoate) is a pyranocoumarin constituent isolated from Saposhnikovia divaricata. Anomalin displayed numerous pharmacological effects including anti-inflammatory, antioxidant, and antitumor properties [204–206]. Anomalin protected against acute lung injury induced by LPS via inhibiting the production of TNF-α, IL-1β, IL-6, and NO [207]. Similarly, anomalin exerted a potent antioxidant activity through increasing enzymatic activities of GST and catalase. Additionally, an in vitro study reported that anomalin significantly downregulated the MAPK (p38, JNK, and ERK1/2) in the RAW264.7 cells [207]. In LPS-stimulated macrophages in vitro, anomalin inhibited inflammation and NF-κB DNA binding [206].
Anomalin has also exerted a neuroprotective effect in diabetic mice and sodium-nitroprusside- (SNP-) induced neuro-2a cells. Anomalin suppressed neuropathic pain in diabetic mice; abolished iNOS, COX-2, and NF-κB and MAPK signaling in SNP-stimulated cells; and inhibited proinflammatory cytokines in HG-induced primary neurons [208]. The modulatory effect of anomalin on Nrf2 signaling has been scarcely studied. In the study of Khan et al., anomalin reduced Nrf2 and HO-1 gene expression levels which were increased following stimulation of the neuro-2a cells with SNP [208]. However, the exact effect of anomalin on Nrf2 signaling needs to be investigated.
3.13. Wedelolactone
Wedelolactone (1,8,9-trihydroxy-3-methoxy-[1]benzofuro[3,2-c]chromen-6-one) is a natural coumarin isolated from Eclipta prostrata L. It exhibited an anticancer effect against different tumor cells, such as prostate [209], breast [210], and pituitary adenomas [211]. It has been reported to exert immunomodulatory, anti-inflammatory [212, 213], antimyotoxic, antihemorrhagic [214], and antioxidant activities [215], inhibit osteoclastogenesis, and enhance osteoblastogenesis [216, 217]. Wedelolactone can suppress LPS-induced inflammation in mouse embryo fibroblasts via suppression of NF-κB activity [218].
Wedelolactone protected against quinolinic acid-induced neurotoxicity and impaired motor function through marked inhibition of neuronal TNF-α, IL-6, and IL-β expression by suppressing NF-κB [212]. Cuong et al. reported that wedelolactone prevented zymosan-induced inflammatory responses in murine bone marrow-derived macrophages through downregulation of TNF-α and IL-6 [213]. Zhu et al. have shown that wedelolactone mitigated inflammation and oxidative injury induced by doxorubicin by suppressing the IκK/IκB/NF-κB signaling pathway [215].
Studies showing the modulatory effect of wedelolactone on Nrf2 signaling are very few. Lin et al. reported that wedelolactone inhibited t-BHP-induced damage in PC12 cells and D-galactose-induced neuronal cell loss in mice through improvement of the antioxidant defense capacity via Nrf2/ARE pathway activation [219]. On the other hand, Ding et al. demonstrated that wedelolactone protected human bronchial epithelial cells against cigarette smoke extract-induced oxidative stress and inflammation responses through Nrf2 inhibition [220].
3.14. Glycycoumarin
Glycycoumarin (3-(2,4-dihydroxyphenyl)-7-hydroxy-5-methoxy-6-(3-methylbut-2-enyl)chromen-2-one) is a major bioactive coumarin compound isolated from licorice. Given its favorable bioavailability features, glycycoumarin exhibited various pharmacological properties including antioxidant [221, 222], anti-inflammatory [223], and antimicrobial [222, 224] activities. Glycycoumarin dose-dependently inhibited LPS-induced ROS generation in macrophages and effectively suppressed NO, IL-6, and PGE2 expression [223]. Song et al. demonstrated that glycycoumarin attenuated hepatotoxicity induced by alcohol following either chronic or acute ethanol exposure via activation of Nrf2. p62 upregulation by a transcriptional mechanism has also been reported to contribute to Nrf2 activation via a positive feedback loop [225]. In contrast, Yan et al. have demonstrated that Nrf2 was not implicated in the protective effect of glycycoumarin on acetaminophen hepatotoxicity [226].
3.15. Osthole
Osthole (7-methoxy-8-(3-methylbut-2-enyl)chromen-2-one) is a coumarin found in a high content in the mature fruit of Cnidium monnieri which is commonly applied in the clinical practice of Chinese medicine. Osthole is also widely distributed in other medicinal plants of the genera Citrus, Clausena Angelica, and Archangelica. Fructus Cnidii improved male function and reinforced the immune system mainly due to its rich content of osthole [227, 228]. Osthole exhibited various pharmacological activities, including antioxidant [229–231], anticancer [232, 233], and anti-inflammatory properties [231, 234, 235].
Several reports of underlying molecular mechanisms reported that osthole displays a strong anti-inflammatory activity. Fan et al. reported that osthole effectively and safely protected against ulcerative colitis (UC) via marked inhibition of TNF-α expression in the colon and markedly reduced MPO activity via suppression of NF-κB p65 and p-IκBα [234]. In an in vitro study, osthole inhibited the production of TNF-α, NO, PGE2, and IL-6 in LPS-induced macrophages [234]. Osthole has a potent selective inhibitory effect on 5-lipoxygenase and COX-1 [236, 237]. It suppressed the immune response of LPS-stimulated macrophages by abolishing ROS generation, iNOS, MAPK, and COX-2 [238, 239]. Osthole also suppressed IL-4- and TNF-α-induced eotaxin expression in bronchial epithelial cells [240] and protected against carrageenan-induced hind paw edema in rats via suppression of PG and NO production [241]. Osthole protects against lumbar disc herniation-induced sciatica and relieved mechanical allodynia through decreasing the COX-2 and iNOS expression in the dorsal root ganglion in rats [242].
The modulatory effect of osthole on Nrf2 signaling has been well-acknowledged by different researchers. A recent study conducted by Chu et al. reported that the protective effect of osthole against glutamate-induced Alzheimer's disease in mice was mediated via Nrf2 activation and its downstream antioxidant proteins SOD-1 and HO-1 [243]. Osthole protected against Ang II-induced apoptosis of rat aortic endothelial cells through suppression of NF-κB and activation of Nrf2 and its downstream antioxidant genes and effectively inhibited Keap1, denoting its potential therapeutic effect against vascular injury [244]. In another study, osthole protected against LPS-induced inflammation in BV2 cells via NF-κB suppression and upregulation of the Nrf2/HO-1 pathway dose dependently [245]. Additionally, osthole exerted neuroprotective effects against global cerebral I/R injury by reducing oxidative stress via the upregulation of the Nrf2/HO-1 signaling pathway [246]. The renoprotective effect of osthole against accelerated focal segmental glomerulosclerosis was mediated via Nrf2 activation and subsequently downregulation of the NF-κB-mediated COX-2 expression [247]. Osthole has also shown renoprotective effects mediated through inhibition of ROS generation and NF-κB/NLRP3 signaling and increased Nrf2 nuclear translocation [248]. Osthole has protective effects on LPS-induced acute lung injury by upregulating the Nrf-2/Trx-1 pathway, whereas Nrf2 siRNA blocked its beneficial effects [249].
3.16. Hydrangenol
Hydrangenol (8-hydroxy-3-(4-hydroxyphenyl)-3,4-dihydroisochromen-1-one) is a natural dihydroisocoumarin mostly obtained from the Hydrangea species (Hydrangeaceae) leaves. Hydrangenol possesses anti-inflammatory [250], antidiabetic [251], antioxidant [250, 252], anticancer [253], and antiangiogenic activities [254]. It inhibited LPS-induced NO release and iNOS expression via suppression of NF-κB and consequently inhibiting NF-κB-DNA. Additionally, hydrangenol suppresses NO production by inducing HO-1 and promoting nuclear translocation of Nrf2. In contrast, transient knockdown of Nrf2 markedly inhibited hydrangenol-induced HO-1 expression, indicating that hydrangenol-induced Nrf2 is an upstream regulator of HO-1 [252]. Hydrangenol exerted antiphotoaging activity in vitro and in UVB-irradiated HR-1 hairless mice. Hydrangenol effectively reduced MMP-1/-3, COX-2, and IL-6 expression and attenuated the phosphorylation of MAPKs and STAT1. Interestingly, hydrangenol upregulated the expression of Nrf2, HO-1, NQO1, GCLM, and GCLC [252].
3.17. Isoimperatorin
Isoimperatorin (4-(3-methylbut-2-enoxy)furo[3,2-g]chromen-7-one) is a 6,7-furanocoumarin derivative. This compound is present in Angelica dahurica, Notopterygium incisum, Ferula lutea, Angelica pubescens, and Peucedanum praeruptorum. Isoimperatorin exhibits various pharmacological activities, including antioxidant [255, 256], anti-inflammatory [257], analgesic [258], antibacterial [259, 260], and anticancer properties [261, 262].
Wijerathne et al. reported that isoimperatorin protected against OA-induced asthma via mitigation of airway inflammation and mucus hypersecretion evidenced by the decreased IL-4, IL-5, and IL-13 production [257]. Mechanistically, isoimperatorin suppressed the activation of NF-κB, p38 MAPK, and ERK1/2 [257]. In addition, isoimperatorin inhibited TNF-α-induced vascular cell adhesion molecule-1 and ROS production and upregulated the PPARγ signaling pathway in human endothelial cells [263]. It has a strong hepatoprotective effect against cytotoxicity in H4IIE cells induced by aflatoxin B1 via Nrf2/ARE activation and induction of GST-α and suppression of CYP1A expression [256].
The effects of coumarins on Nrf2 signaling in in vivo and in vitro studies are summarized in Tables 1 and 2, respectively.
Table 1.
Effect of coumarins on Nrf2 in animal models of different diseases.
| Coumarin | Disease/model | Animal | Effects | Reference(s) |
|---|---|---|---|---|
| IMP | Nrf2 knockout | Mice | IMP induced hepatic antioxidant activities via the Nrf2/ARE mechanism. IMP induced hepatic GST and/or NQO1 activities. |
[75] |
| Allergic responses mediated by mast cells | Mice | IMP attenuated allergic responses. IMP inhibited mast cell degranulation, MAPK, NF-κB, and inflammatory mediators' expression. IMP activated PI3K/Akt and Nrf2/HO-1 pathways. |
[77] | |
|
| ||||
| Visnagin | Cerulein-induced acute pancreatitis | Mice | Visnagin upregulated Nrf2 and attenuated oxidative stress. Visnagin mitigated pancreatic inflammation and NF-κB p65 nuclear translocation. |
[86] |
|
| ||||
| Urolithin B | LPS-induced systemic inflammation | Mice | Urolithin B reduced intracellular ROS production and NADPH oxidase expression. Urolithin B upregulated AMPK phosphorylation and Nrf2/ARE signaling and HO-1 expression. |
[92] |
|
| ||||
| Urolithin A | Colitis | Mice | Urolithin A enhanced gut barrier function and inhibited inflammation through Nrf2-dependent pathways. | [104] |
| High cholesterol diet-fed rats | Rats | Urolithin A upregulated aortic scavenger receptor-class B type I expression and Nrf2 and inhibited ERK1/2 phosphorylation levels. | [264] | |
|
| ||||
| Scopoletin | MG-induced hyperglycemia and insulin resistance | Rats | Scopoletin increased insulin sensitivity, decreased AGEs, and activated Nrf2 by Ser40 phosphorylation. | [121] |
|
| ||||
| Daphnetin | Cisplatin-induced nephrotoxicity | Mice | Daphnetin inhibited ROS generation, lipid peroxidation, NF-κB activation, and proinflammatory cytokines. Daphnetin upregulated Nrf2 and HO-1 expression. |
[132] |
| CCl4-induced hepatotoxicity | Rats | Daphnetin improved liver function, inhibited histological alterations and lipid peroxidation, and increased Nrf2 and HO-1 gene expression. | [138] | |
| 7,12-Dimethylbenz[a]anthracene-induced mammary carcinogenesis | Rats | Daphnetin inhibited lipid peroxidation, enhanced GSH and antioxidant enzymes, decreased NF-κB expression, and activated Nrf2 pathway. | [265] | |
|
| ||||
| Esculin | LPS/D-galactosamine-induced liver injury | Mice | Esculin suppressed lipid peroxidation, MPO, TNF-α, IL-1β, and NF-κB and increased the expression of Nrf2 and HO-1. | [146] |
|
| ||||
| Esculetin | Cerebral I/R | Mice | Esculetin ameliorated mitochondrial oxidative stress, fragmentation, and stress and increased SOD and Nrf2 expression. | [165] |
|
| ||||
| UMB | Hepatic injury in diabetic db/db mice | Mice | UMB ameliorated liver function, serum lipids, and lipid peroxidation and suppressed NF-κB and TLR-4. UMB activated Nrf2 signaling pathway. |
[180] |
| Cyclophosphamide-induced hepatotoxicity | Rats | UMB ameliorated liver function and inhibited histological alterations, lipid peroxidation, and inflammation. UMB upregulated Nrf2, HO-1, PPARγ, and antioxidants and suppressed iNOS and NF-κB. |
[44] | |
| CCl4-induced hepatotoxicity | Rats | UMB improved liver function, inhibited histological alterations and lipid peroxidation, and increased Nrf2 and HO-1 gene expression. | [138] | |
| MTX-induced nephrotoxicity | Rats | UMB inhibited inflammatory response via downregulation of both NF-κB and p38 MAPK genes. UMB downregulated Keap1 and upregulated Nrf2. |
[174] | |
|
| ||||
| Fraxetin | Malaria infection | Mice | Fraxetin suppressed lipid peroxidation and boosted GSH and antioxidant enzymes via Nrf2-ARE activation. | [193] |
|
| ||||
| Fraxin | CCl4-induced hepatotoxicity | Rats | Fraxin ameliorated liver function and lipid peroxidation and increased GSH and Nrf2-mediated antioxidant enzyme system. | [198] |
|
| ||||
| Glycycoumarin | Acute alcoholic liver injury | Mice | Glycycoumarin prevented liver injury via induction of autophagy and activation of Nrf2 signaling. | [225] |
|
| ||||
| Osthole | Alzheimer's disease model | Mice | Osthole restored the mitochondrial membrane potential, ameliorated apoptosis markers, and activated Nrf2 and its downstream antioxidant proteins. | [243] |
| Transient global brain ischemia | Mice | Osthole improved the cognitive functions and upregulated Nrf2/HO-1 signaling pathway. | [246] | |
| Focal segmental glomerulosclerosis | Mice | Osthole suppressed NF-κB-mediated COX-2 expression, PGE2 production, apoptosis, and podocyte injury and activated Nrf2. | [247] | |
| IgA nephropathy | Mice | Osthole inhibited excessive ROS generation and NF-κB/NLRP3 signaling and increased Nrf2 nuclear translocation. | [248] | |
| LPS-induced acute lung injury | Mice | Osthole upregulated Nrf-2/thioredoxin 1 and prevented lung injury. | [249] | |
|
| ||||
| Hydrangenol | UVB-irradiated hairless mice | Mice | Hydrangenol downregulated MMP-1/-3, COX-2, IL-6, MAPKs, and STAT1 and upregulated Nrf2, HO-1, NQO1, GCLM, and GCLC. | [252] |
Table 2.
Effect of coumarins on Nrf2 in in vitro studies.
| Coumarin | Model/cells | Effects | Reference(s) |
|---|---|---|---|
| IMP | Arsenic trioxide-induced toxicity in H9c2 cells | IMP-attenuated ROS generation, cytotoxicity, and apoptosis triggered Nrf2 activation. | [76] |
| IgE-mediated allergic responses in RBL-2H3 cells | IMP inhibited mast cell degranulation; suppressed NF-κB, p38, JNK, and ERK MAPKs; and increased Nrf2 nuclear translocation. | [77] | |
|
| |||
| Urolithin B | LPS-induced BV2 microglial cells | Urolithin B reduced ROS production, NADPH oxidase expression, NF-κB, ERK, and JNK and increased AMPK phosphorylation, Nrf2, and HO-1. | [92] |
|
| |||
| Urolithin A | Senescent human skin fibroblasts | Urolithin A increased type I collagen expression, reduced intracellular ROS, abolished MMP-1 expression, and activated Nrf2/ARE signaling. | [103] |
| LPS-induced Caco2 and HT-29 cells | Urolithin A activated aryl hydrocarbon receptor- (AhR-) Nrf2-dependent pathways. | [104] | |
|
| |||
| Scopoletin | Rotenone-stimulated SH-SY5Y cells | Scopoletin prevented oxidative stress and apoptosis and activated Nrf2 signaling. | [122] |
|
| |||
| Daphnetin | Oleic acid-induced HepG2 cells | Daphnetin decreased CYP2E1 and CYP4A expression, promoted glucose uptake and insulin sensitivity, and enhanced PI3K/Akt and Nrf2 signaling. | [133] |
| Arsenic-induced human lung epithelial cells | Daphnetin reduced ROS, JNK, ERK, Keap1, and apoptosis and activated Nrf2/ARE pathway. | [134] | |
| HG-induced human glomerular mesangial cells | Daphnetin reduced ROS production; attenuated the release of IL-1β, IL-6, and TNF-α via suppression of NF-κB pathway; inhibited the expression Keap1; and upregulated Nrf2. | [135] | |
| Oxygen-glucose deprivation/reoxygenation-induced hippocampal neurons | Daphnetin inhibited oxidative stress and cell apoptosis and enhanced the nuclear translocation of Nrf2 and HO-1 expression. | [136] | |
| t-BHP-induced RAW264.7 cells | Daphnetin suppressed ROS, inhibited cytochrome c release and NLRP3 inflammasome activation, and upregulated Nrf2 nuclear translocation along with Keap1 protein downregulation. | [137] | |
|
| |||
| Esculin | EK 293 and RAW264.7 cells | Esculin suppressed ROS production and activated Nrf2/ARE signaling. | [148] |
|
| |||
| Esculetin | NB4 leukemia cells | Esculetin increased the nuclear translocation of Nrf2. | [164] |
| HG-induced rat mesangial cell line HBZY-1 | Esculetin suppressed ROS production and IL-6 expression and activated Nrf2. | [166] | |
| H2O2-induced C2C12 myoblasts | Esculetin suppressed ROS production and activated the Nrf2/NQO1 pathway. | [167] | |
| Pancreatic carcinoma cells (PANC-1) | Esculetin increased Nrf2 and NQO1 gene expression and Nrf2 nuclear accumulation. | [168] | |
|
| |||
| H2O2-induced HepG2 cells | Esculetin activated Nrf2/NQO1 pathway | [169] | |
| UMB | HG-induced mesangial cells | UMB suppressed ROS production and activated Nrf2. | [166] |
| MG-induced HepG2 cells | UMB abolished ROS generation and increased Nrf2 expression, effects inhibited by Nrf2 depletion. | [181] | |
|
| |||
| Fraxetin | Vascular smooth muscle cells | Fraxetin increased the expression of HO-1 and Nrf2. | [195] |
| HaCaT human keratinocytes | Fraxetin reduced ROS and upregulated Akt, AMPK, HO-1, and Nrf2. | [194] | |
|
| |||
| Fraxin | t-BHP-induced HepG2 cells | Fraxin inhibited t-BHP-induced cytotoxicity and ROS generation through Nrf2-dependent HO-1 expression. | [198] |
|
| |||
| Wedelolactone | t-BHP-induced adrenal pheochromocytoma cells | A wedelolactone-rich extract prevented apoptosis and activated Nrf2/ARE pathway. | [219] |
|
| |||
| Osthole | LPS-stimulated BV2 mouse microglia | Osthole suppressed NF-κB, IL-1β, IL-6, and TNF-α and upregulated Nrf2/HO-1 signaling | [245] |
| LPS-stimulated mesangial cells | Osthole inhibited ROS generation, MCP-1 secretion, and NF-κB activation and upregulated Nrf2. | [248] | |
|
| |||
| Hydrangenol | LPS-stimulated BV2 microglial cells | Hydrangenol attenuated NO production and iNOS expression by inhibiting NF-κB activation and stimulated Nrf2/HO-1 signaling pathway. | [250] |
|
| |||
| Isoimperatorin | Aflatoxin B1-inducible cytotoxicity in H4IIE | Isoimperatorin activated Nrf2/ARE and GST-α and suppressed CYP1A expression. | [256] |
4. In Silico Evidence for Binding of Coumarins to Keap1 Protein
Keap1 plays the key step in the ubiquitination and degradation of Nrf2. In this review, we aimed to provide in silico evidence that coumarins bind Keap1 and hence could be employed as promising Nrf2 activators. AutoDock Vina 1.5.6 was used to perform molecular docking of coumarin derivatives and Keap1 protein. The complex structure of Keap1 with (1S,2R)-2-[(1S)-1-[(1,3-dioxo-2,3-dihydro-1H-isoindol-2-yl)methyl]-1,2,3,4-tetrahydroisoquinolin-2-carbonyl]cyclohexane-1-carboxylic acid (compound (S,R,S)) with PDB ID: 4l7b was used as a model for the docking study (Figure 3). The binding pocket of Keap1 was used to identify the binding conformation of the different coumarin derivatives (Figure 4). The average of the lowest energy of docking was used to show the binding affinity for each coumarin derivative with Keap1. The best-scored conformation has been chosen and visually analyzed using the PyMOL 1.7.6 software. Briefly, 9 different orientations were generated, and the first pose with the lowest docking energy has been used. Each coumarin has a different conformation; however, all the conformations were allocated in the vicinity of the active site. The mean docking energy of all 9 generated models was calculated.
Figure 3.
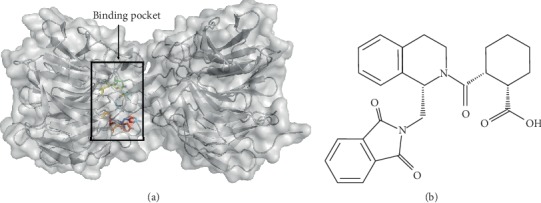
Surface map of Keap1 protein (a) and chemical structure of (S,R,S) (b). The colored residues represent the active site of Keap1 which is involved directly in the interaction with the inhibitor (S,R,S) (PDB ID: 4l7b) and include Y334, S363, R380, N414, R415, S508, S555, Y572, and S602.
Figure 4.
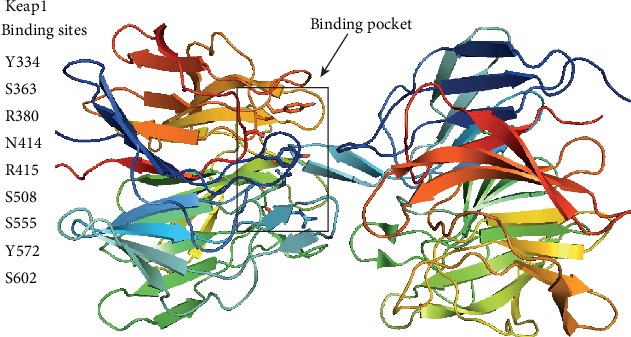
The binding pocket of Keap1 includes Y334, S363, R380, N414, R415, S508, S555, Y572, and S602. These residues were used for the site-specific docking of coumarin derivatives into Keap1.
There are very limited biophysical studies that include the experimental binding data of all listed coumarin derivatives and Keap1. Therefore, this review sheds light on the promising compounds which should be studied in the future. To perform an experimental study, the Keap1 protein needs to be expressed and purified, and the binding affinities with different coumarin derivatives could be estimated using isothermal titration calorimetry (ITC) and/or surface plasmon resonance (SPR). Based on the binding affinity, crystallization trials could be conducted to get an accurate binding mode between Keap1 and coumarins. This review highlights the promising target for biophysical studies.
We performed docking for (S,R,S) which naturally cocrystallizes and binds Keap1 with high affinity. This is to obtain the lowest energy score of docking as this energy represents the reference to judge the predicted binding affinity of different coumarin derivatives with Keap1. Most of the coumarin derivatives showed promising inhibitory effect on Keap1 based on the lowest energy score of docking. Eight derivatives (IMP, urolithin B, urolithin A, esculin, fraxin, wedelolactone, glycycoumarin, and hydrangenol) showed better binding with Keap1, and their affinities are quite close to the (S,R,S) compound as shown in Table 3. These eight compounds are well fitted in the vicinity of the binding pocket through forming hydrogen bonds with the side chains of the polar, positively, and negatively charged amino acids as illustrated in Figures 5 and 6. These results show that esculin and wedelolactone are the most promising coumarins for the development of pharmacological Keap1 inhibitors/Nrf2 activators.
Table 3.
Molecular docking of coumarin derivatives as potential inhibitors of Keap1.
| Compounds | Lowest energy of docking (kcal/mol) |
|---|---|
| IMP | −8.078 ± 0.28 |
| Visnagin | −7.33 ± 0.44 |
| Urolithin B | −8.02 ± 0.43 |
| Urolithin A | −8.01 ± 0.62 |
| Scopoletin | −6.72 ± 0.28 |
| Daphnetin | −6.50 ± 0.20 |
| Esculin | −9.31 ± 0.31 |
| Esculetin | −6.80 ± 0.18 |
| UMB | −6.51 ± 0.15 |
| Fraxetin | −7.02 ± 0.30 |
| Fraxin | −8.20 ± 0.47 |
| Anomalin | −7.21 ± 0.70 |
| Wedelolactone | −9.30 ± 0.33 |
| Glycycoumarin | −8.62 ± 0.53 |
| Osthole | −7.50 ± 0.38 |
| Hydrangenol | −8.41 ± 0.21 |
| Isoimperatorin | −7.60 ± 0.42 |
| Standard (S,R,S) | −10.71 ± 0.40 |
(S,R,S) is a synthetic compound that crystalized with Keap1 with high affinity (PDB ID: 4l7b) and is used as a standard control for comparison.
Figure 5.
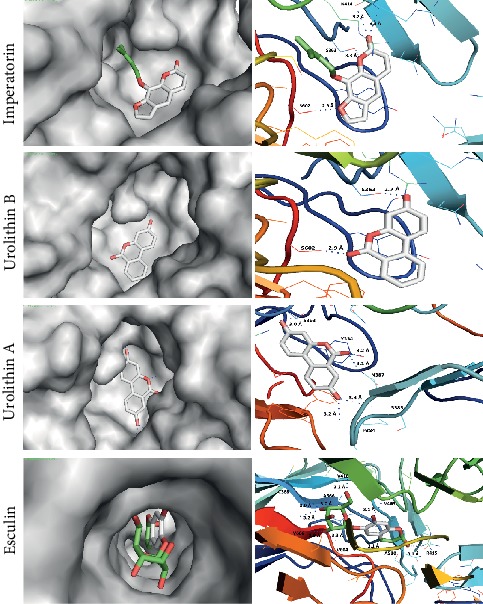
Docking models of imperatorin, urolithin B, urolithin A, and esculin with Keap1. All the compounds are rich with polar groups and form hydrogen bonds with the polar, negatively, and positively charged amino acids in the vicinity of the active site of Keap1.
Figure 6.
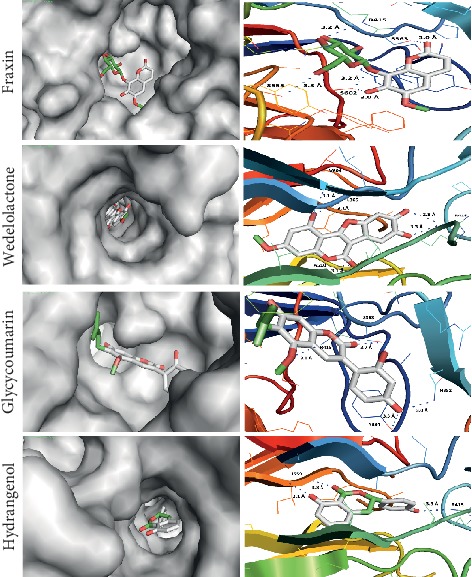
Docking models of fraxin, wedelolactone, glycycoumarin, and hydrangenol with Keap1. All the compounds are rich with polar groups and form hydrogen bonds with the polar, negatively and positively charged amino acids in the vicinity of the active site of Keap1.
IMP: the carbonyl group of a coumarin moiety forms a hydrogen bond and n⟶π∗ interaction with the amino group and carbonyl group of NH2 of the side chain of N414, respectively. Also, the oxygen atom of furan and pyran rings forms two hydrogen bonds with the -OH group of the side chain of S602 and S363, respectively.
Visnagin: the carbonyl group and the oxygen of the methoxy group of the coumarin moiety form two hydrogen bonds with the -OH group of the side chain of S602, while the oxygen of the furan ring forms a hydrogen bond with the amino group of the side chain of N387 at the dimeric interface of the Keap1 protein.
Urolithin B: the carbonyl and hydroxy groups of the coumarin moiety exhibited two hydrogen bonds with S602 and S363, respectively. These two hydrogen bonds have a remarkably short length approximately 2.9 Å and 2.7 Å which could enhance the interaction of urolithin B and binding pocket of Keap1.
Urolithin A: this forms five hydrogen bonds at the dimeric interface of the Keap1 protein. Both hydroxyl groups form hydrogen bonds with S363, P384, and S383. Also, the carbonyl groups form two hydrogen bonds with the -OH group of the side chain of Y334 and the amino group of the side chain of N387.
Daphnetin: the carbonyl oxygen forms a hydrogen bond with S602, and the two hydroxy groups form two hydrogen bonds with S363.
Esculin: this coumarin is rich in hydroxyl groups which serve as hydrogen bond donors to the residues in the active site of Keap1. It forms six hydrogen bonds with the carbonyl group of C368, A366, V418, V604, and V606. These hydroxyl groups of the oxan ring support the perfect fitting of esculin deep in the binding pocket of Keap1. In addition, the hydroxyl group of the coumarin moiety forms two hydrogen bonds with V463 and A510. The carbonyl group of the lactone ring forms a hydrogen bond with the amino group of the side chain of R415. Esculin makes hydrophobic interactions with the nonpolar amino acids in V418, V604, V606, V463, and A510.
UMB: this forms two hydrogen bonds with S363 and N387 at the dimeric interface of Keap1.
Fraxetin: the dihydroxy groups form bonds with the side chain of N382 and S363, and the oxygen of the methoxy group forms a hydrogen bond with S363. The carbonyl group and oxygen of the pyran ring form two extra hydrogen bonds with N387. These five hydrogen bonds are located at the dimeric interface of Keap1.
Fraxin: the hydroxyl groups of oxan rings and oxygen of the ether bond between the oxan ring and the coumarin nucleus form three hydrogen bonds with the side chain of R415, S555, and S602. Also, the hydroxyl and carbonyl groups of this coumarin form two hydrogen bonds with S602 and S363, respectively. The coumarin nucleus exhibits a different conformation in fraxin as compared to esculin which could prevent the perfect fit of fraxin in the binding pocket of Keap1. This explanation is supported by the lower energy of the docking of esculin than fraxin.
Esculetin: this forms six hydrogen bonds with S602, N382, N387, and Y334. Also, the benzene ring of the coumarin nucleus makes hydrophobic interaction with the aromatic ring of Y334.
Anomalin: this forms two hydrogen bonds with R415 and S602.
Wedelolactone: this fits properly in the active site of Keap1 due to its unique conformation. The hydroxyl group of the coumarin moiety forms two hydrogen bonds with the carbonyl groups of L365 and V604. The carbonyl group of coumarin exhibits n⟶π∗ interaction with the carbonyl group of A510. The two-hydroxyl group of the benzofuran ring forms two hydrogen bonds with the side chains of R415. Wedelolactone has hydrophobic interactions with L365, A366, V604, and A510.
Glycycoumarin: the oxygens of the methoxy and carbonyl groups of coumarin form two hydrogen bonds with R415 and S363. Another two hydrogen bonds are formed between the hydroxy group of the dihydroxyphenyl group and the side chain of N382 and Y334. The phenyl group of glycycoumarin has a hydrophobic interaction with the aromatic ring of the side chain of Y334.
Scopoletin: this forms six hydrogen bonds with the side chains of S363, N414, N382, and Y334 at the dimeric interface of Keap1. Also, there is n⟶π∗ interaction between the carbonyl group of coumarin and the carbonyl group of R336.
Osthole: this forms three hydrogen bonds with the side chains of N387 and S602 at the dimeric interface of Keap1.
Hydrangenol: this forms three hydrogen bonds with I559 and the side chain of R415. It has a hydrophobic interaction with A556, V604, I559, and L577.
Isoimperatorin: this forms two hydrogen bonds with the side chains of S602 and N387 at the dimeric interface of Keap1.
5. Conclusions
The Nrf2 signaling pathway plays a vital role in the protection against cell injury induced by oxidative stress and electrophiles. Excessive production of ROS and oxidative injury are associated with the pathogenesis of several diseases and disorders. There is crosstalk between the Nrf2/ARE signaling and other signaling pathways, including NF-κB, and this represents the main underlying mechanism by which Nrf2 exerts its anti-inflammatory activities. Given the dual role of Nrf2 activation in the prevention of oxidative stress and inflammation, pharmacological activation of this signaling pathway could represent a powerful strategy for treating diseases associated with excessive release of ROS and proinflammatory mediators. Coumarins represent a vast group of natural compounds with beneficial pharmacological properties. Several studies have demonstrated the role of Nrf2 activation in mediating the therapeutic effects of some coumarins both in vitro and in vivo. We employed molecular docking simulations to investigate the binding of coumarins with Keap1 and to verify whether their therapeutic effects are related to antioxidant properties mediated via Nrf2 modulation. IMP, visnagin, urolithin B, urolithin A, scopoletin, esculin, esculetin, UMB, fraxetin, fraxin, daphnetin, anomalin, wedelolactone, glycycoumarin, osthole, hydrangenol, and isoimperatorin showed binding affinities toward Keap1 through forming hydrogen bonds with the side chains of the polar, positively, and negatively charged amino acids. IMP, urolithin B, urolithin A, esculin, fraxin, wedelolactone, glycycoumarin, and hydrangenol exhibited strong binding with Keap1 with affinities quite close to a standard Keap1 inhibitor. Thus, activation of the Nrf2 signaling pathway can explain some of the pharmacological and biological effects of coumarins. Coumarins therefore represent promising leads in the development of effective Keap1 inhibitors/Nrf2 activators and a good resource for discovering drug candidates for the treatment/prevention of various diseases.
Conflicts of Interest
The authors report no conflict of interests.
References
- 1.Anand P., Singh B., Singh N. A review on coumarins as acetylcholinesterase inhibitors for Alzheimer’s disease. Bioorganic & medicinal chemistry. 2012;20(3):1175–1180. doi: 10.1016/j.bmc.2011.12.042. [DOI] [PubMed] [Google Scholar]
- 2.Razavi S. M. Plant coumarins as allelopathic agents. International Journal of Biological Chemistry. 2011;5(1):86–90. doi: 10.3923/ijbc.2011.86.90. [DOI] [Google Scholar]
- 3.Riveiro M., de Kimpe N., Moglioni A., et al. Coumarins: old compounds with novel promising therapeutic perspectives. Current Medicinal Chemistry. 2010;17(13):1325–1338. doi: 10.2174/092986710790936284. [DOI] [PubMed] [Google Scholar]
- 4.Iranshahi M., Askari M., Sahebkar A., Adjipavlou-Litina D. Evaluation of antioxidant, anti-inflammatory and lipoxygenase inhibitory activities of the prenylated coumarin umbelliprenin. DARU Journal of Pharmaceutical Sciences. 2009;17(2):99–103. [Google Scholar]
- 5.Gnonlonfin G. J. B., Sanni A., Brimer L. Review Scopoletin – A Coumarin Phytoalexin with Medicinal Properties. Critical Reviews in Plant Sciences. 2012;31(1):47–56. doi: 10.1080/07352689.2011.616039. [DOI] [Google Scholar]
- 6.Venkata Sairam K., Gurupadayya B. M., Chandan R. S., Nagesha D. K., Vishwanathan B. A review on chemical profile of coumarins and their therapeutic role in the treatment of cancer. Current Drug Delivery. 2016;13(2):186–201. doi: 10.2174/1567201812666150702102800. [DOI] [PubMed] [Google Scholar]
- 7.Kontogiorgis C., Detsi A., Hadjipavlou-Litina D. Coumarin-based drugs: a patent review (2008–present) Expert Opinion on Therapeutic Patents. 2012;22(4):437–454. doi: 10.1517/13543776.2012.678835. [DOI] [PubMed] [Google Scholar]
- 8.Ceska O., Chaudhary S. K., Warrington P. J., Ashwood-Smith M. J. Photoactive furocoumarins in fruits of some umbellifers. Phytochemistry. 1986;26(1):165–169. doi: 10.1016/s0031-9422(00)81503-4. [DOI] [Google Scholar]
- 9.Hoult J. R. S., Payá M. Pharmacological and biochemical actions of simple coumarins: natural products with therapeutic potential. General Pharmacology: The Vascular System. 1996;27(4):713–722. doi: 10.1016/0306-3623(95)02112-4. [DOI] [PubMed] [Google Scholar]
- 10.Sarker S. D., Nahar L. Dietary coumarins. Handbook of Dietary Phytochemicals. 2020:1–56. doi: 10.1007/978-981-13-1745-3_37-1. [DOI] [Google Scholar]
- 11.Lacy A., O'Kennedy R. Studies on coumarins and coumarin-related compounds to determine their therapeutic role in the treatment of cancer. Current Pharmaceutical Design. 2004;10(30):3797–3811. doi: 10.2174/1381612043382693. [DOI] [PubMed] [Google Scholar]
- 12.Venugopala K. N., Rashmi V., Odhav B. Review on natural coumarin lead compounds for their pharmacological activity. BioMed Research International. 2013;2013:14. doi: 10.1155/2013/963248.963248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.O'Kennedy R., Thornes R. D. Coumarins : Biology, Applications and Mode of Action. Chichester: Wiley; 1997. [Google Scholar]
- 14.Jain P. K., Joshi H. Coumarin: chemical and pharmacological profile. Journal of Applied Pharmaceutical Science. 2012;2(6):236–240. [Google Scholar]
- 15.Matos M. J., Vazquez-Rodriguez S., Santana L., et al. Looking for new targets: simple coumarins as antibacterial agents. Medicinal Chemistry. 2012;8(6):1140–1145. doi: 10.2174/1573406411208061140. [DOI] [PubMed] [Google Scholar]
- 16.Shi Y., Zhou C. H. Synthesis and evaluation of a class of new coumarin triazole derivatives as potential antimicrobial agents. Bioorganic & Medicinal Chemistry Letters. 2011;21(3):956–960. doi: 10.1016/j.bmcl.2010.12.059. [DOI] [PubMed] [Google Scholar]
- 17.Smyth T., Ramachandran V. N., Smyth W. F. A study of the antimicrobial activity of selected naturally occurring and synthetic coumarins. International Journal of Antimicrobial Agents. 2009;33(5):421–426. doi: 10.1016/j.ijantimicag.2008.10.022. [DOI] [PubMed] [Google Scholar]
- 18.Wall M. E., Wani M. C., Manikumar G., et al. Plant antimutagenic agents, 3. Coumarins. Journal of Natural Products. 1988;51(6):1148–1152. doi: 10.1021/np50060a016. [DOI] [PubMed] [Google Scholar]
- 19.Bansal Y., Sethi P., Bansal G. Coumarin: a potential nucleus for anti-inflammatory molecules. Medicinal Chemistry Research. 2013;22(7):3049–3060. doi: 10.1007/s00044-012-0321-6. [DOI] [Google Scholar]
- 20.Fylaktakidou K., Hadjipavlou-Litina D., Litinas K., Nicolaides D. Natural and synthetic coumarin derivatives with anti-inflammatory/antioxidant activities. Current pharmaceutical design. 2004;10(30):3813–3833. doi: 10.2174/1381612043382710. [DOI] [PubMed] [Google Scholar]
- 21.Lowenthal J., Birnbaum H. Vitamin K and coumarin anticoagulants: dependence of anticoagulant effect on inhibition of vitamin K transport. Science. 1969;164(3876):181–183. doi: 10.1126/science.164.3876.181. [DOI] [PubMed] [Google Scholar]
- 22.Jain M., Surin W. R., Misra A., et al. Antithrombotic activity of a newly synthesized coumarin derivative 3-(5-hydroxy-2, 2-dimethyl-chroman-6-yl)-N-{2-[3-(5-hydroxy-2, 2-dimethyl-chroman-6-yl)-propionylamino]-ethyl}-propionamide. Chemical Biology & Drug Design. 2013;81(4):499–508. doi: 10.1111/cbdd.12000. [DOI] [PubMed] [Google Scholar]
- 23.Hoak J. C., Connor W. E., Warner E. D., Carter J. R. The antithrombotic properties of coumarin drugs. Annals of Internal Medicine. 1961;54(1):73–81. doi: 10.7326/0003-4819-54-1-73. [DOI] [PubMed] [Google Scholar]
- 24.Kontogiorgis C., Nicolotti O., Mangiatordi G. F., et al. Studies on the antiplatelet and antithrombotic profile of anti-inflammatory coumarin derivatives. Journal of Enzyme Inhibition and Medicinal Chemistry. 2014;30(6):925–933. doi: 10.3109/14756366.2014.995180. [DOI] [PubMed] [Google Scholar]
- 25.Amin K. M., Awadalla F. M., Eissa A. A. M., Abou-Seri S. M., Hassan G. S. Design, synthesis and vasorelaxant evaluation of novel coumarin–pyrimidine hybrids. Bioorganic & Medicinal Chemistry. 2011;19(20):6087–6097. doi: 10.1016/j.bmc.2011.08.037. [DOI] [PubMed] [Google Scholar]
- 26.Bertin R., Chen Z., Martínez-Vázquez M., García-Argaéz A., Froldi G. Vasodilation and radical-scavenging activity of imperatorin and selected coumarinic and flavonoid compounds from genus Casimiroa. Phytomedicine. 2014;21(5):586–594. doi: 10.1016/j.phymed.2013.10.030. [DOI] [PubMed] [Google Scholar]
- 27.Menezes J. C., Diederich M. Translational role of natural coumarins and their derivatives as anticancer agents. Future Medicinal Chemistry. 2019;11(9):1057–1082. doi: 10.4155/fmc-2018-0375. [DOI] [PubMed] [Google Scholar]
- 28.Luo K.-w., Sun J. G., Chan J. Y. W., et al. Anticancer effects of imperatorin isolated from Angelica dahurica: induction of apoptosis in HepG2 cells through both death-receptor- and mitochondria-mediated pathways. Chemotherapy. 2011;57(6, article 000331641):449–459. doi: 10.1159/000331641. [DOI] [PubMed] [Google Scholar]
- 29.YANG D., Tianwei G. U., WANG T., TANG Q., Changyan M. A. Effects of osthole on migration and invasion in breast cancer cells. Bioscience, Biotechnology, and Biochemistry. 2010;74(7):1430–1434. doi: 10.1271/bbb.100110. [DOI] [PubMed] [Google Scholar]
- 30.Germoush M. O., Othman S. I., al-Qaraawi M. A., et al. Umbelliferone prevents oxidative stress, inflammation and hematological alterations, and modulates glutamate-nitric oxide-cGMP signaling in hyperammonemic rats. Biomedicine & Pharmacotherapy. 2018;102:392–402. doi: 10.1016/j.biopha.2018.03.104. [DOI] [PubMed] [Google Scholar]
- 31.Fort D. M., Rao K., Jolad S. D., Luo J., Carlson T. J., King S. R. Antihyperglycemic activity of Teramnus labialis (Fabaceae) Phytomedicine. 2000;6(6):465–467. doi: 10.1016/S0944-7113(00)80075-6. [DOI] [PubMed] [Google Scholar]
- 32.Tinel M., Belghiti J., Descatoire V., et al. Inactivation of human liver cytochrome P-450 by the drug methoxsalen and other psoralen derivatives. Biochemical Pharmacology. 1987;36(6):951–955. doi: 10.1016/0006-2952(87)90190-0. [DOI] [PubMed] [Google Scholar]
- 33.Shin E., Choi K. M., Yoo H. S., Lee C. K., Hwang B. Y., Lee M. K. Inhibitory effects of coumarins from the stem barks of Fraxinus rhynchophylla on adipocyte differentiation in 3T3-L1 cells. Biological & Pharmaceutical Bulletin. 2010;33(9):1610–1614. doi: 10.1248/bpb.33.1610. [DOI] [PubMed] [Google Scholar]
- 34.Mahmoud A. M., Hozayen W. G., Hasan I. H., Shaban E., Bin-Jumah M. Umbelliferone ameliorates CCl4-induced liver fibrosis in rats by upregulating PPARγ and attenuating oxidative stress, inflammation, and TGF-β1/Smad3 signaling. Inflammation. 2019;42(3):1103–1116. doi: 10.1007/s10753-019-00973-8. [DOI] [PubMed] [Google Scholar]
- 35.Wang C., Pei A., Chen J., et al. A natural coumarin derivative esculetin offers neuroprotection on cerebral ischemia/reperfusion injury in mice. Journal of Neurochemistry. 2012;121(6):1007–1013. doi: 10.1111/j.1471-4159.2012.07744.x. [DOI] [PubMed] [Google Scholar]
- 36.Basile A., Sorbo S., Spadaro V., et al. Antimicrobial and antioxidant activities of coumarins from the roots of Ferulago campestris (Apiaceae) Molecules. 2009;14(3, article 14030939):939–952. doi: 10.3390/molecules14030939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kim S. H., Kang K. A., Zhang R., et al. Protective effect of esculetin against oxidative stress-induced cell damage via scavenging reactive oxygen species. Acta Pharmacologica Sinica. 2008;29(11):1319–1326. doi: 10.1111/j.1745-7254.2008.00878.x. [DOI] [PubMed] [Google Scholar]
- 38.Kostova I., Bhatia S., Grigorov P., et al. Coumarins as antioxidants. Current Medicinal Chemistry. 2011;18(25):3929–3951. doi: 10.2174/092986711803414395. [DOI] [PubMed] [Google Scholar]
- 39.Bubols G. B., da Rocha Vianna D., Medina-Remon A., et al. The antioxidant activity of coumarins and flavonoids. Mini Reviews in Medicinal Chemistry. 2013;13(3):318–334. doi: 10.2174/138955713804999775. [DOI] [PubMed] [Google Scholar]
- 40.Kontogiorgis C., Hadjipavlou-Litina D. Biological evaluation of several coumarin derivatives designed as possible anti-inflammatory/antioxidant agents. Journal of Enzyme Inhibition and Medicinal Chemistry. 2008;18(1):63–69. doi: 10.1080/1475636031000069291. [DOI] [PubMed] [Google Scholar]
- 41.Mahmoud A. M., Alexander M. Y., Tutar Y., Wilkinson F. L., Venditti A. Oxidative stress in metabolic disorders and drug-induced injury: the potential role of Nrf2 and PPARs activators. Oxidative Medicine and Cellular Longevity. 2017;2017:14. doi: 10.1155/2017/2508909.2508909 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Liguori I., Russo G., Curcio F., et al. Oxidative stress, aging, and diseases. Clinical Interventions in Aging. 2018;13:757–772. doi: 10.2147/CIA.S158513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Rani V., Deep G., Singh R. K., Palle K., Yadav U. C. S. Oxidative stress and metabolic disorders: pathogenesis and therapeutic strategies. Life Sciences. 2016;148:183–193. doi: 10.1016/j.lfs.2016.02.002. [DOI] [PubMed] [Google Scholar]
- 44.Mahmoud A. M., Germoush M. O., Alotaibi M. F., Hussein O. E. Possible involvement of Nrf2 and PPARγ up-regulation in the protective effect of umbelliferone against cyclophosphamide-induced hepatotoxicity. Biomedicine & Pharmacotherapy. 2017;86:297–306. doi: 10.1016/j.biopha.2016.12.047. [DOI] [PubMed] [Google Scholar]
- 45.Satta S., Mahmoud A. M., Wilkinson F. L., Alexander M. Y., White S. J. The role of Nrf2 in cardiovascular function and disease. Oxidative Medicine and Cellular Longevity. 2017;2017:18. doi: 10.1155/2017/9237263.9237263 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Itoh K., Wakabayashi N., Katoh Y., et al. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes & Development. 1999;13(1):76–86. doi: 10.1101/gad.13.1.76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Eggler A. L., Liu G., Pezzuto J. M., van Breemen R. B., Mesecar A. D. Modifying specific cysteines of the electrophile-sensing human Keap1 protein is insufficient to disrupt binding to the Nrf2 domain Neh2. Proceedings of the National Academy of Sciences. 2005;102(29):10070–10075. doi: 10.1073/pnas.0502402102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Venugopal R., Jaiswal A. K. Nrf2 and Nrf1 in association with Jun proteins regulate antioxidant response element-mediated expression and coordinated induction of genes encoding detoxifying enzymes. Oncogene. 1998;17(24, article BF1202237):3145–3156. doi: 10.1038/sj.onc.1202237. [DOI] [PubMed] [Google Scholar]
- 49.Stocker R., McDonagh A. F., Glazer A. N., Ames B. N. Oxygen Radicals in Biological Systems Part B: Oxygen Radicals and Antioxidants. 1990. [31] Antioxidant activities of bile pigments: Biliverdin and bilirubin; pp. 301–309. [DOI] [PubMed] [Google Scholar]
- 50.Ganesh Yerra V., Negi G., Sharma S. S., Kumar A. Potential therapeutic effects of the simultaneous targeting of the Nrf2 and NF-κB pathways in diabetic neuropathy. Redox Biology. 2013;1(1):394–397. doi: 10.1016/j.redox.2013.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Wardyn J. D., Ponsford A. H., Sanderson C. M. Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochemical Society transactions. 2015;43(4):621–626. doi: 10.1042/BST20150014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Pan H., Wang H., Wang X., Zhu L., Mao L. The Absence of Nrf2 Enhances NF- B-Dependent Inflammation following Scratch Injury in Mouse Primary Cultured Astrocytes. Mediators of Inflammation. 2012;2012:9. doi: 10.1155/2012/217580.217580 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Abd El-Twab S. M., Hussein O. E., Hozayen W. G., Bin-Jumah M., Mahmoud A. M. Chicoric acid prevents methotrexate-induced kidney injury by suppressing NF-κB/NLRP3 inflammasome activation and up-regulating Nrf2/ARE/HO-1 signaling. Inflammation Research. 2019;68(6, article 1241):511–523. doi: 10.1007/s00011-019-01241-z. [DOI] [PubMed] [Google Scholar]
- 54.Mahmoud A. M., Hussein O. E., Abd El-Twab S. M., Hozayen W. G. Ferulic acid protects against methotrexate nephrotoxicity via activation of Nrf2/ARE/HO-1 signaling and PPARγ, and suppression of NF-κB/NLRP3 inflammasome axis. Food & Function. 2019;10(8):4593–4607. doi: 10.1039/C9FO00114J. [DOI] [PubMed] [Google Scholar]
- 55.Aladaileh S. H., Abukhalil M. H., Saghir S. A. M., et al. Galangin activates Nrf2 signaling and attenuates oxidative damage, inflammation, and apoptosis in a rat model of cyclophosphamide-induced hepatotoxicity. Biomolecules. 2019;9(8):p. 346. doi: 10.3390/biom9080346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.ALHaithloul H. A. S., Alotaibi M. F., Bin-Jumah M., Elgebaly H., Mahmoud A. M. Olea europaea leaf extract up-regulates Nrf2/ARE/HO-1 signaling and attenuates cyclophosphamide-induced oxidative stress, inflammation and apoptosis in rat kidney. Biomedicine & Pharmacotherapy. 2019;111:676–685. doi: 10.1016/j.biopha.2018.12.112. [DOI] [PubMed] [Google Scholar]
- 57.Ranneh Y., Akim A. M., Hamid H. A., Khazaai H., Fadel A., Mahmoud A. M. Stingless bee honey protects against lipopolysaccharide induced-chronic subclinical systemic inflammation and oxidative stress by modulating Nrf2, NF-κB and p38 MAPK. Nutrition & Metabolism. 2019;16(1) doi: 10.1186/s12986-019-0341-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Mahmoud A. M., Germoush M. O., Al-Anazi K. M., Mahmoud A. H., Farah M. A., Allam A. A. Commiphora molmol protects against methotrexate-induced nephrotoxicity by up- regulating Nrf2/ARE/HO-1 signaling. Biomedicine & Pharmacotherapy. 2018;106:499–509. doi: 10.1016/j.biopha.2018.06.171. [DOI] [PubMed] [Google Scholar]
- 59.Mahmoud A. M., Hozayen W. G., Ramadan S. M. Berberine ameliorates methotrexate-induced liver injury by activating Nrf2/HO-1 pathway and PPARγ, and suppressing oxidative stress and apoptosis in rats. Biomedicine & Pharmacotherapy. 2017;94:280–291. doi: 10.1016/j.biopha.2017.07.101. [DOI] [PubMed] [Google Scholar]
- 60.Aly M. S., Galaly S. R., Moustafa N., Mohammed H. M., Khadrawy S. M., Mahmoud A. M. Hesperidin protects against diethylnitrosamine/carbon tetrachloride-induced renal repercussions via up-regulation of Nrf2/HO-1 signaling and attenuation of oxidative stress. Journal of Applied Pharmaceutical Science. 2017;7(11):7–14. doi: 10.7324/japs.2017.71102. [DOI] [Google Scholar]
- 61.Soares M. P., Seldon M. P., Gregoire I. P., et al. Heme oxygenase-1 modulates the expression of adhesion molecules associated with endothelial cell activation. The Journal of Immunology. 2004;172(6):3553–3563. doi: 10.4049/jimmunol.172.6.3553. [DOI] [PubMed] [Google Scholar]
- 62.Yu M., Li H., Liu Q., et al. Nuclear factor p65 interacts with Keap1 to repress the Nrf2-ARE pathway. Cellular Signalling. 2011;23(5):883–892. doi: 10.1016/j.cellsig.2011.01.014. [DOI] [PubMed] [Google Scholar]
- 63.Sun Z., Wu T., Zhao F., Lau A., Birch C. M., Zhang D. D. KPNA6 (importin {alpha}7)-mediated nuclear import of Keap1 represses the Nrf2-dependent antioxidant response. Molecular and Cellular Biology. 2011;31(9):1800–1811. doi: 10.1128/MCB.05036-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Sun Z., Chin Y. E., Zhang D. D. Acetylation of Nrf2 by p300/CBP augments promoter-specific DNA binding of Nrf2 during the antioxidant response. Molecular and Cellular Biology. 2009;29(10):2658–2672. doi: 10.1128/MCB.01639-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Liu G. H., Qu J., Shen X. NF-κB/p65 antagonizes Nrf2-ARE pathway by depriving CBP from Nrf2 and facilitating recruitment of HDAC3 to MafK. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 2008;1783(5):713–727. doi: 10.1016/j.bbamcr.2008.01.002. [DOI] [PubMed] [Google Scholar]
- 66.Patel Rajesh M., Patel Natvar J. In vitro antioxidant activity of coumarin compounds by DPPH, Super oxide and nitric oxide free radical scavenging methods. Journal of Advanced Pharmacy Education & Research. 2011;1:52–68. [Google Scholar]
- 67.WU C., HUANG M., LIN Y., JU H., CHING H. Antioxidant properties of Cortex Fraxini and its simple coumarins. Food Chemistry. 2007;104(4):1464–1471. doi: 10.1016/j.foodchem.2007.02.023. [DOI] [Google Scholar]
- 68.Nasser M. I., Zhu S., Hu H., Huang H., Guo M., Zhu P. Effects of imperatorin in the cardiovascular system and cancer. Biomedicine & Pharmacotherapy. 2019;120:p. 109401. doi: 10.1016/j.biopha.2019.109401. [DOI] [PubMed] [Google Scholar]
- 69.Chowdhury A. A., Gawali N. B., Shinde P., Munshi R., Juvekar A. R. Imperatorin ameliorates lipopolysaccharide induced memory deficit by mitigating proinflammatory cytokines, oxidative stress and modulating brain- derived neurotropic factor. Cytokine. 2018;110:78–86. doi: 10.1016/j.cyto.2018.04.018. [DOI] [PubMed] [Google Scholar]
- 70.Mi C., Ma J., Wang K. S., et al. Imperatorin suppresses proliferation and angiogenesis of human colon cancer cell by targeting HIF-1α via the mTOR/p70S6K/4E-BP1 and MAPK pathways. Journal of Ethnopharmacology. 2017;203:27–38. doi: 10.1016/j.jep.2017.03.033. [DOI] [PubMed] [Google Scholar]
- 71.Ouyang P., Chen J., Sun M., et al. Imperatorin inhibits the expression of alpha-hemolysin in Staphylococcus aureus strain BAA-1717 (USA300) Antonie van Leeuwenhoek. 2016;109(7):915–922. doi: 10.1007/s10482-016-0690-9. [DOI] [PubMed] [Google Scholar]
- 72.Lin C. L., Hsiao G., Wang C. C., Lee Y. L. Imperatorin exerts antiallergic effects in Th2-mediated allergic asthma via induction of IL-10-producing regulatory T cells by modulating the function of dendritic cells. Pharmacological Research. 2016;110:111–121. doi: 10.1016/j.phrs.2016.04.030. [DOI] [PubMed] [Google Scholar]
- 73.Singh G., Kaur J., Kaur M., Singh P., Bhatti R. Anti-nociceptive and anti-inflammatory effect of imperatorin: evidences for involvement of COX-2, iNOS, NFκB and inflammatory cytokines. International Journal of Neuroscience. 2020;130(2):176–185. doi: 10.1080/00207454.2019.1667789. [DOI] [PubMed] [Google Scholar]
- 74.Li Y. Z., Chen J. H., Tsai C. F., Yeh W. L. Anti-inflammatory property of imperatorin on alveolar macrophages and inflammatory lung injury. Journal of Natural Products. 2019;82(4):1002–1008. doi: 10.1021/acs.jnatprod.9b00145. [DOI] [PubMed] [Google Scholar]
- 75.Prince M., Li Y., Childers A., Itoh K., Yamamoto M., Kleiner H. E. Comparison of citrus coumarins on carcinogen-detoxifying enzymes in Nrf2 knockout mice. Toxicology Letters. 2009;185(3):180–186. doi: 10.1016/j.toxlet.2008.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Hu L., Sun J., Li H., et al. Differential mechanistic investigation of protective effects from imperatorin and sec-O-glucosylhamaudol against arsenic trioxide-induced cytotoxicity in vitro. Toxicology in Vitro. 2016;37:97–105. doi: 10.1016/j.tiv.2016.09.002. [DOI] [PubMed] [Google Scholar]
- 77.Xian Z., Jin G., Li H., et al. Imperatorin suppresses anaphylactic reaction and IgE-mediated allergic responses by inhibiting multiple steps of FceRI signaling in mast cells: IMP alleviates allergic responses in PCA. BioMed Research International. 2019;2019:12. doi: 10.1155/2019/7823761.7823761 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kaul B., Staba E. J. Visnagin: Biosynthesis and isolation from Ammi visnagi suspension cultures. Science. 1965;150(3704):1731–1732. doi: 10.1126/science.150.3704.1731. [DOI] [PubMed] [Google Scholar]
- 79.Duarte J., Torres A. I., Zarzuelo A. Cardiovascular effects of visnagin on rats. Planta Medica. 2000;66(1):35–39. doi: 10.1055/s-2000-11108. [DOI] [PubMed] [Google Scholar]
- 80.Duarte J., Pérez-Vizcaíno F., Torres A. I., Zarzuelo A., Jiménez J., Tamargo J. Vasodilator effects of visnagin in isolated rat vascular smooth muscle. European Journal of Pharmacology. 1995;286(2):115–122. doi: 10.1016/0014-2999(95)00418-k. [DOI] [PubMed] [Google Scholar]
- 81.Asnani A., Zheng B., Liu Y., et al. Highly potent visnagin derivatives inhibit Cyp1 and prevent doxorubicin cardiotoxicity. JCI Insight. 2018;3(1) doi: 10.1172/jci.insight.96753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Liu Y., Asnani A., Zou L., et al. Visnagin protects against doxorubicin-induced cardiomyopathy through modulation of mitochondrial malate dehydrogenase. Translational Medicine. 2014;6(266, article 266ra170) doi: 10.1126/scitranslmed.3010189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Vanachayangkul P., Byer K., Khan S., Butterweck V. An aqueous extract of Ammi visnaga fruits and its constituents khellin and visnagin prevent cell damage caused by oxalate in renal epithelial cells. Phytomedicine. 2010;17(8-9):653–658. doi: 10.1016/j.phymed.2009.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Lee J. K., Jung J. S., Park S. H., et al. Anti-inflammatory effect of visnagin in lipopolysaccharide-stimulated BV-2 microglial cells. Archives of Pharmacal Research. 2010;33(11):1843–1850. doi: 10.1007/s12272-010-1117-1. [DOI] [PubMed] [Google Scholar]
- 85.Khalil H. S., Sedky N. K., Amin K. M., Abd Elhafez O. M., Arafa R. K. Visnagin and benzofuran scaffold-based molecules as selective cyclooxygenase-2 inhibitors with anti-inflammatory and analgesic properties: design, synthesis and molecular docking. Future Medicinal Chemistry. 2019;11(7):659–676. doi: 10.4155/fmc-2018-0398. [DOI] [PubMed] [Google Scholar]
- 86.Pasari L. P., Khurana A., Anchi P., Aslam Saifi M., Annaldas S., Godugu C. Visnagin attenuates acute pancreatitis via Nrf2/NFκB pathway and abrogates associated multiple organ dysfunction. Biomedicine & Pharmacotherapy. 2019;112:p. 108629. doi: 10.1016/j.biopha.2019.108629. [DOI] [PubMed] [Google Scholar]
- 87.Okuda T., Yoshida T., Hatano T. Ellagitannins as active constituents of medicinal plants. Planta Medica. 1989;55(2):117–122. doi: 10.1055/s-2006-961902. [DOI] [PubMed] [Google Scholar]
- 88.Tomás-Barberán F. A., González-Sarrías A., García-Villalba R., et al. Urolithins, the rescue of "old" metabolites to understand a "new" concept: metabotypes as a nexus among phenolic metabolism, microbiota dysbiosis, and host health status. Molecular Nutrition & Food Research. 2017;61(1):p. 1500901. doi: 10.1002/mnfr.201500901. [DOI] [PubMed] [Google Scholar]
- 89.Landete J. M. Ellagitannins, ellagic acid and their derived metabolites: a review about source, metabolism, functions and health. Food Research International. 2011;44(5):1150–1160. doi: 10.1016/j.foodres.2011.04.027. [DOI] [Google Scholar]
- 90.Stanisławska I. J., Piwowarski J. P., Granica S., Kiss A. K. The effects of urolithins on the response of prostate cancer cells to non- steroidal antiandrogen bicalutamide. Phytomedicine. 2018;46:176–183. doi: 10.1016/j.phymed.2018.03.054. [DOI] [PubMed] [Google Scholar]
- 91.Cho H., Jung H., Lee H., Yi H. C., Kwak H. K., Hwang K. T. Chemopreventive activity of ellagitannins and their derivatives from black raspberry seeds on HT-29 colon cancer cells. Food & Function. 2015;6(5):1675–1683. doi: 10.1039/C5FO00274E. [DOI] [PubMed] [Google Scholar]
- 92.Lee G., Park J. S., Lee E. J., Ahn J. H., Kim H. S. Anti-inflammatory and antioxidant mechanisms of urolithin B in activated microglia. Phytomedicine. 2019;55:50–57. doi: 10.1016/j.phymed.2018.06.032. [DOI] [PubMed] [Google Scholar]
- 93.Ito H. Metabolites of the ellagitannin geraniin and their antioxidant activities. Planta Medica. 2011;77(11):1110–1115. doi: 10.1055/s-0030-1270749. [DOI] [PubMed] [Google Scholar]
- 94.Yuan T., Ma H., Liu W., et al. Pomegranate's neuroprotective effects against Alzheimer's disease are mediated by urolithins, its ellagitannin-gut microbial derived metabolites. ACS Chemical Neuroscience. 2015;7(1):26–33. doi: 10.1021/acschemneuro.5b00260. [DOI] [PubMed] [Google Scholar]
- 95.Heilman J., Andreux P., Tran N., Rinsch C., Blanco-Bose W. Safety assessment of urolithin A, a metabolite produced by the human gut microbiota upon dietary intake of plant derived ellagitannins and ellagic acid. Food and Chemical Toxicology. 2017;108:289–297. doi: 10.1016/j.fct.2017.07.050. [DOI] [PubMed] [Google Scholar]
- 96.Bialonska D., Kasimsetty S. G., Khan S. I., Ferreira D. Urolithins, intestinal microbial metabolites of pomegranate ellagitannins, exhibit potent antioxidant activity in a cell-based assay. Journal of Agricultural and Food Chemistry. 2009;57(21):10181–10186. doi: 10.1021/jf9025794. [DOI] [PubMed] [Google Scholar]
- 97.González-Sarrías A., Larrosa M., Tomás-Barberán F. A., Dolara P., Espín J. C. NF-kappaB-dependent anti-inflammatory activity of urolithins, gut microbiota ellagic acid-derived metabolites, in human colonic fibroblasts. British Journal of Nutrition. 2010;104(4):503–512. doi: 10.1017/S0007114510000826. [DOI] [PubMed] [Google Scholar]
- 98.Giménez-Bastida J. A., González-Sarrías A., Larrosa M., Tomás-Barberán F., Espín J. C., García-Conesa M.-T. Ellagitannin metabolites, urolithin A glucuronide and its aglycone urolithin A, ameliorate TNF-α-induced inflammation and associated molecular markers in human aortic endothelial cells. Molecular Nutrition & Food Research. 2012;56(5):784–796. doi: 10.1002/mnfr.201100677. [DOI] [PubMed] [Google Scholar]
- 99.Kasimsetty S. G., Bialonska D., Reddy M. K., Ma G., Khan S. I., Ferreira D. Colon cancer chemopreventive activities of pomegranate ellagitannins and urolithins. Journal of Agricultural and Food Chemistry. 2010;58(4):2180–2187. doi: 10.1021/jf903762h. [DOI] [PubMed] [Google Scholar]
- 100.Zhao R., Long X., Yang J., et al. Pomegranate peel polyphenols reduce chronic low-grade inflammatory responses by modulating gut microbiota and decreasing colonic tissue damage in rats fed a high-fat diet. Food & Function. 2019;10(12):8273–8285. doi: 10.1039/C9FO02077B. [DOI] [PubMed] [Google Scholar]
- 101.Fu X., Gong L. F., Wu Y. F., et al. Urolithin A targets the PI3K/Akt/NF-κB pathways and prevents IL-1β-induced inflammatory response in human osteoarthritis:in vitroandin vivostudies. Food & Function. 2019;10(9):6135–6146. doi: 10.1039/C9FO01332F. [DOI] [PubMed] [Google Scholar]
- 102.Gong Z., Huang J., Xu B., et al. Urolithin A attenuates memory impairment and neuroinflammation in APP/PS1 mice. Journal of Neuroinflammation. 2019;16(1):p. 62. doi: 10.1186/s12974-019-1450-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Liu C. F., Li X. L., Zhang Z. L., et al. Antiaging effects of urolithin A on replicative senescent human skin fibroblasts. Rejuvenation Research. 2019;22(3):191–200. doi: 10.1089/rej.2018.2066. [DOI] [PubMed] [Google Scholar]
- 104.Singh R., Chandrashekharappa S., Bodduluri S. R., et al. Enhancement of the gut barrier integrity by a microbial metabolite through the Nrf2 pathway. Nature Communications. 2019;10(1):p. 89. doi: 10.1038/s41467-018-07859-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Basu M., Mayana K., Xavier S., Balachandran S., Mishra N. Effect of scopoletin on monoamine oxidases and brain amines. Neurochemistry International. 2016;93:113–117. doi: 10.1016/j.neuint.2016.01.001. [DOI] [PubMed] [Google Scholar]
- 106.Pandy V., Narasingam M., Kunasegaran T., Murugan D. D., Mohamed Z. Effect of Noni (Morinda citrifolia Linn.) Fruit and Its Bioactive Principles Scopoletin and Rutin on Rat Vas Deferens Contractility: An Ex Vivo Study. The Scientific World Journal. 2014;2014:11. doi: 10.1155/2014/909586.909586 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Shinde P. B., Katekhaye S. D., Mulik M. B., Laddha K. S. Rapid simultaneous determination of marmelosin, umbelliferone and scopoletin from Aegle marmelos fruit by RP-HPLC. Journal of Food Science and Technology. 2014;51(9, article 1270):2251–2255. doi: 10.1007/s13197-014-1270-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Napiroon T., Bacher M., Balslev H., Tawaitakham K., Santimaleeworagun W., Vajrodaya S. Scopoletin from Lasianthus lucidus Blume (Rubiaceae): a potential antimicrobial against multidrug-resistant Pseudomonas aeruginosa. Journal of Applied Pharmaceutical Science. 2018;8(9):1–6. doi: 10.7324/japs.2018.8901. [DOI] [Google Scholar]
- 109.Nam H., Kim M. M. Scopoletin has a potential activity for anti-aging via autophagy in human lung fibroblasts. Phytomedicine. 2015;22(3):362–368. doi: 10.1016/j.phymed.2015.01.004. [DOI] [PubMed] [Google Scholar]
- 110.Kim H. J., Jang S. I., Kim Y. J., et al. Scopoletin suppresses pro-inflammatory cytokines and PGE2 from LPS-stimulated cell line, RAW 264.7 cells. Fitoterapia. 2004;75(3-4):261–266. doi: 10.1016/j.fitote.2003.12.021. [DOI] [PubMed] [Google Scholar]
- 111.Leema G., Tamizhselvi R. Protective effect of scopoletin against cerulein-induced acute pancreatitis and associated lung injury in mice. Pancreas. 2018;47(5):577–585. doi: 10.1097/MPA.0000000000001034. [DOI] [PubMed] [Google Scholar]
- 112.Shaw C. Y., Chen C. H., Hsu C. C., Chen C. C., Tsai Y. C. Antioxidant properties of scopoletin isolated from Sinomonium acutum. Phytotherapy Research. 2003;17(7):823–825. doi: 10.1002/ptr.1170. [DOI] [PubMed] [Google Scholar]
- 113.Kalpana K., Sathiya Priya C., Dipti N., Vidhya R., Anuradha C. V. Supplementation of scopoletin improves insulin sensitivity by attenuating the derangements of insulin signaling through AMPK. Molecular and Cellular Biochemistry. 2019;453(1-2):65–78. doi: 10.1007/s11010-018-3432-7. [DOI] [PubMed] [Google Scholar]
- 114.Liu X. L., Zhang L., Fu X. L., Chen K., Qian B. C. Effect of scopoletin on PC3 cell proliferation and apoptosis. Acta Pharmacologica Sinica. 2001;22(10):929–933. [PubMed] [Google Scholar]
- 115.Adams M., Efferth T., Bauer R. Activity-Guided Isolation of Scopoletin and Isoscopoletin, the Inhibitory Active Principles towards CCRF-CEM Leukaemia Cells and Multi-Drug Resistant CEM/ADR5000 Cells, from Artemisia argyi. Planta Medica. 2006;72(9):862–864. doi: 10.1055/s-2006-947165. [DOI] [PubMed] [Google Scholar]
- 116.Jang J. H., Park J. E., Han J. S. Scopoletin inhibits α-glucosidase in vitro and alleviates postprandial hyperglycemia in mice with diabetes. European Journal of Pharmacology. 2018;834:152–156. doi: 10.1016/j.ejphar.2018.07.032. [DOI] [PubMed] [Google Scholar]
- 117.Jang J. H., Park J. E., Han J. S. Scopoletin increases glucose uptake through activation of PI3K and AMPK signaling pathway and improves insulin sensitivity in 3T3-L1 cells. Nutrition Research. 2020;74:52–61. doi: 10.1016/j.nutres.2019.12.003. [DOI] [PubMed] [Google Scholar]
- 118.Choi R. Y., Ham J. R., Lee H. I., et al. Scopoletin supplementation ameliorates steatosis and inflammation in diabetic mice. Phytotherapy Research. 2017;31(11):1795–1804. doi: 10.1002/ptr.5925. [DOI] [PubMed] [Google Scholar]
- 119.Wan Osman W. N., Lau S. F., Mohamed S. Scopoletin-standardized Morinda elliptica leaf extract suppressed inflammation and cartilage degradation to alleviate osteoarthritis: a preclinical study. Phytotherapy Research. 2017;31(12):1954–1961. doi: 10.1002/ptr.5949. [DOI] [PubMed] [Google Scholar]
- 120.Kim H. L., Woo S. M., Choi W. R., et al. Scopoletin downregulates MMP‑1 expression in human fibroblasts via inhibition of p38 phosphorylation. International Journal of Molecular Medicine. 2018;42(4):2285–2293. doi: 10.3892/ijmm.2018.3757. [DOI] [PubMed] [Google Scholar]
- 121.Chang W. C., Wu S. C., Xu K. D., Liao B. C., Wu J. F., Cheng A. S. Scopoletin protects against methylglyoxal-induced hyperglycemia and insulin resistance mediated by suppression of advanced glycation endproducts (AGEs) generation and anti-glycation. Molecules. 2015;20(2):2786–2801. doi: 10.3390/molecules20022786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Narasimhan K. K. S., Jayakumar D., Velusamy P., et al. Morinda citrifolia and its active principle scopoletin mitigate protein aggregation and neuronal apoptosis through augmenting the DJ-1/Nrf2/ARE signaling pathway. Oxidative Medicine and Cellular Longevity. 2019;2019:13. doi: 10.1155/2019/2761041.2761041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Liao M. J., Lin L. F., Zhou X., et al. Daphnetin prevents chronic unpredictable stress-induced cognitive deficits. Fundamental & Clinical Pharmacology. 2013;27(5):510–516. doi: 10.1111/j.1472-8206.2012.01049.x. [DOI] [PubMed] [Google Scholar]
- 124.NDong C., Anzellotti D., Ibrahim R. K., Huner N. P. A., Sarhan F. Daphnetin Methylation by a Novel O-Methyltransferase Is Associated with Cold Acclimation and Photosystem II Excitation Pressure in Rye. Journal of Biological Chemistry. 2003;278(9):6854–6861. doi: 10.1074/jbc.M209439200. [DOI] [PubMed] [Google Scholar]
- 125.Tu L., Li S., Fu Y., et al. The therapeutic effects of daphnetin in collagen-induced arthritis involve its regulation of Th17 cells. International Immunopharmacology. 2012;13(4):417–423. doi: 10.1016/j.intimp.2012.04.001. [DOI] [PubMed] [Google Scholar]
- 126.Gao Q., Shan J., Di L., Jiang L., Xu H. Therapeutic effects of daphnetin on adjuvant-induced arthritic rats. Journal of Ethnopharmacology. 2008;120(2):259–263. doi: 10.1016/j.jep.2008.08.031. [DOI] [PubMed] [Google Scholar]
- 127.Yeşilada E., Taninaka H., Takaishi Y., et al. In vitro inhibitory effects of Daphne oleoides ssp. oleoides on inflammatory cytokines and activity-guided isolation of active constituents. Cytokine. 2001;13(6):359–364. doi: 10.1006/cyto.2001.0838. [DOI] [PubMed] [Google Scholar]
- 128.Fukuda H., Nakamura S., Chisaki Y., et al. Daphnetin inhibits invasion and migration of LM8 murine osteosarcoma cells by decreasing RhoA and Cdc42 expression. Biochemical and Biophysical Research Communications. 2016;471(1):63–67. doi: 10.1016/j.bbrc.2016.01.179. [DOI] [PubMed] [Google Scholar]
- 129.Shen L., Zhou T., Wang J., et al. Daphnetin reduces endotoxin lethality in mice and decreases LPS-induced inflammation in Raw264.7 cells via suppressing JAK/STATs activation and ROS production. Inflammation Research. 2017;66(7):579–589. doi: 10.1007/s00011-017-1039-1. [DOI] [PubMed] [Google Scholar]
- 130.Finn G. J., Creaven B. S., Egan D. A. Daphnetin induced differentiation of human renal carcinoma cells and its mediation by p38 mitogen-activated protein kinase. Biochemical Pharmacology. 2004;67(9):1779–1788. doi: 10.1016/j.bcp.2004.01.014. [DOI] [PubMed] [Google Scholar]
- 131.Yang E. B., Zhao Y. N., Zhang K., Mack P. Daphnetin, one of coumarin derivatives, is a protein kinase inhibitor. Biochemical and Biophysical Research Communications. 1999;260(3):682–685. doi: 10.1006/bbrc.1999.0958. [DOI] [PubMed] [Google Scholar]
- 132.Zhang L., Gu Y., Li H., et al. Daphnetin protects against cisplatin-induced nephrotoxicity by inhibiting inflammatory and oxidative response. International Immunopharmacology. 2018;65:402–407. doi: 10.1016/j.intimp.2018.10.018. [DOI] [PubMed] [Google Scholar]
- 133.Liu Y., Liao L., Chen Y., Han F. Effects of daphnetin on lipid metabolism, insulin resistance and oxidative stress in OA-treated HepG2 cells. Molecular Medicine Reports. 2019;19(6):4673–4684. doi: 10.3892/mmr.2019.10139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Lv X., Li Y., Xiao Q., Li D. Daphnetin activates the Nrf2-dependent antioxidant response to prevent arsenic-induced oxidative insult in human lung epithelial cells. Chemico-Biological Interactions. 2019;302:93–100. doi: 10.1016/j.cbi.2019.02.001. [DOI] [PubMed] [Google Scholar]
- 135.Xu K., Guo L., Bu H., Wang H. Daphnetin inhibits high glucose-induced extracellular matrix accumulation, oxidative stress and inflammation in human glomerular mesangial cells. Journal of Pharmacological Sciences. 2019;139(2):91–97. doi: 10.1016/j.jphs.2018.11.013. [DOI] [PubMed] [Google Scholar]
- 136.Zhi J., Duan B., Pei J., Wu S., Wei J. Daphnetin protects hippocampal neurons from oxygen-glucose deprivation-induced injury. Journal of Cellular Biochemistry. 2018;120(3):4132–4139. doi: 10.1002/jcb.27698. [DOI] [PubMed] [Google Scholar]
- 137.Lv H., Liu Q., Zhou J., Tan G., Deng X., Ci X. Daphnetin-mediated Nrf2 antioxidant signaling pathways ameliorate tert-butyl hydroperoxide (t-BHP)-induced mitochondrial dysfunction and cell death. Free Radical Biology and Medicine. 2017;106:38–52. doi: 10.1016/j.freeradbiomed.2017.02.016. [DOI] [PubMed] [Google Scholar]
- 138.Mohamed M. R., Emam M. A., Hassan N. S., Mogadem A. I. Umbelliferone and daphnetin ameliorate carbon tetrachloride-induced hepatotoxicity in rats via nuclear factor erythroid 2-related factor 2-mediated heme oxygenase-1 expression. Environmental Toxicology and Pharmacology. 2014;38(2):531–541. doi: 10.1016/j.etap.2014.08.004. [DOI] [PubMed] [Google Scholar]
- 139.Stefanova Z., Neychev H., Ivanovska N., Kostova I. Effect of a total extract from Fraxinus ornus stem bark and esculin on zymosan- and carrageenan-induced paw oedema in mice. Journal of Ethnopharmacology. 1995;46(2):101–106. doi: 10.1016/0378-8741(95)01233-4. [DOI] [PubMed] [Google Scholar]
- 140.Kaneko T., Tahara S., Takabayashi F. Inhibitory effect of natural coumarin compounds, esculetin and esculin, on oxidative DNA damage and formation of aberrant crypt foci and tumors induced by 1, 2-dimethylhydrazine in rat colons. Biological & Pharmaceutical Bulletin. 2007;30(11):2052–2057. doi: 10.1248/bpb.30.2052. [DOI] [PubMed] [Google Scholar]
- 141.Zhang M., Xin X., Lai F., Zhang X., Li X., Wu H. Cellular transport of esculin and its acylated derivatives in Caco-2 cell monolayers and their antioxidant properties in vitro. Journal of Agricultural and Food Chemistry. 2017;65(34):7424–7432. doi: 10.1021/acs.jafc.7b02525. [DOI] [PubMed] [Google Scholar]
- 142.Song Y., Wang X., Qin S., Zhou S., Li J., Gao Y. Esculin ameliorates cognitive impairment in experimental diabetic nephropathy and induces anti-oxidative stress and anti-inflammatory effects via the MAPK pathway. Molecular Medicine Reports. 2018;17(5):7395–7402. doi: 10.3892/mmr.2018.8727. [DOI] [PubMed] [Google Scholar]
- 143.Kang K. S., Lee W., Jung Y., et al. Protective effect of esculin on streptozotocin-induced diabetic renal damage in mice. Journal of Agricultural and Food Chemistry. 2014;62(9):2069–2076. doi: 10.1021/jf403840c. [DOI] [PubMed] [Google Scholar]
- 144.Martin M. J., Marhuenda E., de la Lastra C. A. Esculine, ranitidine and carbenoxolone: different modes of action on gastric mucosa. General Pharmacology: The Vascular System. 1991;22(6):1001–1004. doi: 10.1016/0306-3623(91)90568-Q. [DOI] [PubMed] [Google Scholar]
- 145.Li W., Wang Y., Wang X., et al. Gastroprotective effect of esculin on ethanol-induced gastric lesion in mice. Fundamental & Clinical Pharmacology. 2017;31(2):174–184. doi: 10.1111/fcp.12255. [DOI] [PubMed] [Google Scholar]
- 146.Liu A., Shen Y., Du Y., et al. Esculin prevents lipopolysaccharide/D-galactosamine-induced acute liver injury in mice. Microbial Pathogenesis. 2018;125:418–422. doi: 10.1016/j.micpath.2018.10.003. [DOI] [PubMed] [Google Scholar]
- 147.Li W., Wang Y., Wang X., et al. Esculin attenuates endotoxin shock induced by lipopolysaccharide in mouse and NO production in vitro through inhibition of NF-κB activation. European Journal of Pharmacology. 2016;791:726–734. doi: 10.1016/j.ejphar.2016.10.013. [DOI] [PubMed] [Google Scholar]
- 148.Kim K. H., Park H., Park H. J., et al. Glycosylation enables aesculin to activate Nrf2. Scientific Reports. 2016;6(1) doi: 10.1038/srep29956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Lee B. C., Lee S. Y., Lee H. J., et al. Anti-oxidative and photo-protective effects of coumarins isolated from Fraxinus chinensis. Archives of Pharmacal Research. 2007;30(10):1293–1301. doi: 10.1007/BF02980270. [DOI] [PubMed] [Google Scholar]
- 150.Pan Y., Zhu J., Wang H., et al. Antioxidant activity of ethanolic extract of Cortex fraxini and use in peanut oil. Food Chemistry. 2007;103(3):913–918. doi: 10.1016/j.foodchem.2006.09.044. [DOI] [Google Scholar]
- 151.Liang C., Ju W., Pei S., Tang Y., Xiao Y. Pharmacological activities and synthesis of esculetin and its derivatives: a mini-review. Molecules. 2017;22(3):p. 387. doi: 10.3390/molecules22030387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Chen Y., Zhang Q., Liu H., et al. Esculetin ameliorates psoriasis-like skin disease in mice by inducing CD4(+)Foxp3(+) regulatory T cells. Frontiers in Immunology. 2018;9:p. 2092. doi: 10.3389/fimmu.2018.02092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Hongyan L. Esculetin attenuates Th2 and Th17 responses in an ovalbumin-induced asthmatic mouse model. Inflammation. 2016;39(2):735–743. doi: 10.1007/s10753-015-0300-4. [DOI] [PubMed] [Google Scholar]
- 154.Issa Y. M., el-Hawary W. F., Moustafa M. E., Refaat M. ChemInform Abstract: Spectroscopic Studies on Some New Azo Dyes Derived from 4‐Methylesculetin and Their Biological Activity. ChemInform. 1998;29(38) doi: 10.1002/chin.199838212. [DOI] [Google Scholar]
- 155.Yang L., Wu L., Yao X., et al. Hydroxycoumarins: New, effective plant-derived compounds reduce Ralstonia pseudosolanacearum populations and control tobacco bacterial wilt. Microbiological Research. 2018;215:15–21. doi: 10.1016/j.micres.2018.05.011. [DOI] [PubMed] [Google Scholar]
- 156.Wang X., Yang C., Zhang Q., et al. In vitro anticancer effects of esculetin against human leukemia cell lines involves apoptotic cell death, autophagy, G0/G1 cell cycle arrest and modulation of Raf/MEK/ERK signalling pathway. Journal of B.U.ON. : official journal of the Balkan Union of Oncology. 2019;24(4):1686–1691. [PubMed] [Google Scholar]
- 157.Li J., Li S., Wang X., Wang H. Esculetin induces apoptosis of SMMC-7721 cells through IGF-1/PI3K/Akt-mediated mitochondrial pathways. Canadian Journal of Physiology and Pharmacology. 2017;95(7):787–794. doi: 10.1139/cjpp-2016-0548. [DOI] [PubMed] [Google Scholar]
- 158.Cho J. H., Shin J. C., Cho J. J., Choi Y. H., Shim J. H., Chae J. I. Esculetin (6,7-dihydroxycoumarin): a potential cancer chemopreventive agent through suppression of Sp1 in oral squamous cancer cells. International Journal of Oncology. 2015;46(1):265–271. doi: 10.3892/ijo.2014.2700. [DOI] [PubMed] [Google Scholar]
- 159.Zhang G., Xu Y., Zhou H. F. Esculetin inhibits proliferation, invasion, and migration of laryngeal cancer in vitro and in vivo by inhibiting Janus kinas (JAK)-signal transducer and activator of transcription-3 (STAT3) activation. Medical Science Monitor. 2019;25, article 916246:7853–7863. doi: 10.12659/MSM.916246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Sharma S. H., Thulasingam S., Chellappan D. R., Chinnaswamy P., Nagarajan S. Morin and Esculetin supplementation modulates c-myc induced energy metabolism and attenuates neoplastic changes in rats challenged with the procarcinogen 1,2 - dimethylhydrazine. European Journal of Pharmacology. 2017;796:20–31. doi: 10.1016/j.ejphar.2016.12.019. [DOI] [PubMed] [Google Scholar]
- 161.Yan L., Yu H. H., Liu Y. S., Wang Y. S., Zhao W. H. Esculetin enhances the inhibitory effect of 5-fluorouracil on the proliferation, migration and epithelial-mesenchymal transition of colorectal cancer. Cancer Biomarkers. 2019;24(2):231–240. doi: 10.3233/CBM-181764. [DOI] [PubMed] [Google Scholar]
- 162.Liu S. Q., He L., Peng H. Effect of esculetin on osteoarthritis in rabbit. 2004;9:567–570. [Google Scholar]
- 163.Kim Y., Park Y., Namkoong S., Lee J. Esculetin inhibits the inflammatory response by inducing heme oxygenase-1 in cocultured macrophages and adipocytes. Food & Function. 2014;5(9):2371–2377. doi: 10.1039/C4FO00351A. [DOI] [PubMed] [Google Scholar]
- 164.Rubio V., García-Pérez A. I., Herráez A., Diez J. C. Different roles of Nrf2 and NFKB in the antioxidant imbalance produced by esculetin or quercetin on NB4 leukemia cells. Chemico-Biological Interactions. 2018;294, article S0009279718306100:158–166. doi: 10.1016/j.cbi.2018.08.015. [DOI] [PubMed] [Google Scholar]
- 165.Xu B., Zhu L., Chu J., et al. Esculetin improves cognitive impairments induced by transient cerebral ischaemia and reperfusion in mice via regulation of mitochondrial fragmentation and mitophagy. Behavioural Brain Research. 2019;372:p. 112007. doi: 10.1016/j.bbr.2019.112007. [DOI] [PubMed] [Google Scholar]
- 166.Sen Z., Weida W., Jie M., Li S., Dongming Z., Xiaoguang C. Coumarin glycosides from Hydrangea paniculata slow down the progression of diabetic nephropathy by targeting Nrf2 anti-oxidation and smad2/3-mediated profibrosis. Phytomedicine. 2019;57:385–395. doi: 10.1016/j.phymed.2018.12.045. [DOI] [PubMed] [Google Scholar]
- 167.Han M. H., Park C., Lee D. S., et al. Cytoprotective effects of esculetin against oxidative stress are associated with the upregulation of Nrf2-mediated NQO1 expression via the activation of the ERK pathway. International Journal of Molecular Medicine. 2017;39(2):380–386. doi: 10.3892/ijmm.2016.2834. [DOI] [PubMed] [Google Scholar]
- 168.Arora R., Sawney S., Saini V., Steffi C., Tiwari M., Saluja D. Esculetin induces antiproliferative and apoptotic response in pancreatic cancer cells by directly binding to KEAP1. Molecular Cancer. 2016;15(1):p. 64. doi: 10.1186/s12943-016-0550-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Subramaniam S. R., Ellis E. M. Esculetin-induced protection of human hepatoma HepG2 cells against hydrogen peroxide is associated with the Nrf2-dependent induction of the NAD(P)H: quinone oxidoreductase 1 gene. Toxicology and Applied Pharmacology. 2011;250(2):130–136. doi: 10.1016/j.taap.2010.09.025. [DOI] [PubMed] [Google Scholar]
- 170.Mazimba O. Umbelliferone: sources, chemistry and bioactivities review. Bulletin of Faculty of Pharmacy, Cairo University. 2017;55(2):223–232. doi: 10.1016/j.bfopcu.2017.05.001. [DOI] [Google Scholar]
- 171.Rauf A., Khan R., Khan H., Pervez S., Pirzada A. S. In vivo antinociceptive and anti-inflammatory activities of umbelliferone isolated from Potentilla evestita. Natural Product Research. 2014;28(17):1371–1374. doi: 10.1080/14786419.2014.901317. [DOI] [PubMed] [Google Scholar]
- 172.Anwar F., al-Abbasi F. A., Bhatt P. C., Ahmad A., Sethi N., Kumar V. Umbelliferone β-d-galactopyranoside inhibits chemically induced renal carcinogenesis via alteration of oxidative stress, hyperproliferation and inflammation: possible role of NF-κB. Toxicology Research. 2015;4(5):1308–1323. doi: 10.1039/c5tx00146c. [DOI] [Google Scholar]
- 173.Sim M. O., Lee H. I., Ham J. R., Seo K. I., Kim M. J., Lee M. K. Anti-inflammatory and antioxidant effects of umbelliferone in chronic alcohol-fed rats. Nutrition Research and Practice. 2015;9(4):364–369. doi: 10.4162/nrp.2015.9.4.364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Hassanein E. H. M., Mohamed W. R., Shalkami A.-G. S., Khalaf M. M., Hemeida R. A. M. Renoprotective effects of umbelliferone on methotrexate-induced renal injury through regulation of Nrf-2/Keap-1, P38MAPK/NF-κB, and apoptosis signaling pathways. Food and Chemical Toxicology. 2018;116:152–160. doi: 10.1016/j.fct.2018.03.041. [DOI] [PubMed] [Google Scholar]
- 175.Ouyang L., Dan Y., Shao Z., et al. Effect of umbelliferone on adjuvant-induced arthritis in rats by MAPK/NF-κB pathway. Drug Design, Development and Therapy. 2019;Volume 13:1163–1170. doi: 10.2147/DDDT.S190155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Yu S. M., Hu D. H., Zhang J. J. Umbelliferone exhibits anticancer activity via the induction of apoptosis and cell cycle arrest in HepG2 hepatocellular carcinoma cells. Molecular Medicine Reports. 2015;12(3):3869–3873. doi: 10.3892/mmr.2015.3797. [DOI] [PubMed] [Google Scholar]
- 177.Muthu R., Selvaraj N., Vaiyapuri M. Anti-inflammatory and proapoptotic effects of umbelliferone in colon carcinogenesis. Human & Experimental Toxicology. 2016;35(10):1041–1054. doi: 10.1177/0960327115618245. [DOI] [PubMed] [Google Scholar]
- 178.Vijayalakshmi A., Sindhu G. Umbelliferone arrest cell cycle at G0/G1 phase and induces apoptosis in human oral carcinoma (KB) cells possibly via oxidative DNA damage. Biomedicine & Pharmacotherapy. 2017;92:661–671. doi: 10.1016/j.biopha.2017.05.128. [DOI] [PubMed] [Google Scholar]
- 179.Wang D., Wang X., Tong W., Cui Y., Li X., Sun H. Umbelliferone alleviates lipopolysaccharide-induced inflammatory responses in acute lung injury by down-regulating TLR4/MyD88/NF-κB signaling. Inflammation. 2019;42(2):440–448. doi: 10.1007/s10753-018-00953-4. [DOI] [PubMed] [Google Scholar]
- 180.Yin J., Wang H., Lu G. Umbelliferone alleviates hepatic injury in diabetic db/db mice via inhibiting inflammatory response and activating Nrf2-mediated antioxidant. Bioscience Reports. 2018;38(4) doi: 10.1042/BSR20180444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Li D., Wang N., Zhang J., Ma S., Zhao Z., Ellis E. M. Hepatoprotective effect of 7-hydroxycoumarin against methyl glyoxal toxicity via activation of Nrf2. Chemico-Biological Interactions. 2017;276:203–209. doi: 10.1016/j.cbi.2017.02.020. [DOI] [PubMed] [Google Scholar]
- 182.Kimura Y., Sumiyoshi M. Antitumor and antimetastatic actions of dihydroxycoumarins (esculetin or fraxetin) through the inhibition of M2 macrophage differentiation in tumor- associated macrophages and/or G1 arrest in tumor cells. European Journal of Pharmacology. 2015;746:115–125. doi: 10.1016/j.ejphar.2014.10.048. [DOI] [PubMed] [Google Scholar]
- 183.Zhang Y., Wang L., Deng Y., et al. Fraxetin suppresses proliferation of non-small-cell lung cancer cells via preventing activation of signal transducer and activator of transcription 3. The Tohoku Journal of Experimental Medicine. 2019;248(1):3–12. doi: 10.1620/tjem.248.3. [DOI] [PubMed] [Google Scholar]
- 184.Sánchez-Reus M. I., Peinado I. I., Molina-Jiménez M. F., Benedí J. Fraxetin prevents rotenone-induced apoptosis by induction of endogenous glutathione in human neuroblastoma cells. Neuroscience Research. 2005;53(1):48–56. doi: 10.1016/j.neures.2005.05.009. [DOI] [PubMed] [Google Scholar]
- 185.Martin-Aragon S., Benedi J. M., Villar A. M. Modifications on antioxidant capacity and lipid peroxidation in mice under fraxetin treatment. Journal of Pharmacy and Pharmacology. 1997;49(1):49–52. doi: 10.1111/j.2042-7158.1997.tb06751.x. [DOI] [PubMed] [Google Scholar]
- 186.FERNÁNDEZ-PUNTERO B., BARROSO I., IGLESIAS I., BENEDÍ J., VILLAR A. Antioxidant activity of fraxetin: in vivo and ex vivo parameters in normal situation versus induced stress. Biological & Pharmaceutical Bulletin. 2001;24(7):777–784. doi: 10.1248/bpb.24.777. [DOI] [PubMed] [Google Scholar]
- 187.Paya M., Goodwin P. A., de Las Heras B., Hoult J. R. S. Superoxide scavenging activity in leukocytes and absence of cellular toxicity of a series of coumarins. Biochemical Pharmacology. 1994;48(3):445–451. doi: 10.1016/0006-2952(94)90273-9. [DOI] [PubMed] [Google Scholar]
- 188.Molina-Jiménez M. F., Sánchez-Reus M. I., Cascales M., Andrés D., Benedí J. Effect of fraxetin on antioxidant defense and stress proteins in human neuroblastoma cell model of rotenone neurotoxicity. Comparative study with myricetin and N-acetylcysteine. Toxicology and Applied Pharmacology. 2005;209(3):214–225. doi: 10.1016/j.taap.2005.04.009. [DOI] [PubMed] [Google Scholar]
- 189.Chen X., Ying X., Sun W., Zhu H., Jiang X., Chen B. The therapeutic effect of fraxetin on ethanol-induced hepatic fibrosis by enhancing ethanol metabolism, inhibiting oxidative stress and modulating inflammatory mediators in rats. International Immunopharmacology. 2018;56:98–104. doi: 10.1016/j.intimp.2018.01.027. [DOI] [PubMed] [Google Scholar]
- 190.Yoshiyuki K., Hiromichi O., Shigeru A., Kimiye B., Mitsugi K. Inhibition of the formation of 5-hydroxy-6,8,11,14-eicosatetraenoic acid from arachidonic acid in polymorphonuclear leukocytes by various coumarins. Biochimica et Biophysica Acta (BBA) - Lipids and Lipid Metabolism. 1985;834(2):224–229. doi: 10.1016/0005-2760(85)90159-6. [DOI] [PubMed] [Google Scholar]
- 191.Wang H., Zou D., Xie K., Xie M. Antibacterial mechanism of fraxetin against Staphylococcus aureus. Molecular Medicine Reports. 2014;10(5):2341–2345. doi: 10.3892/mmr.2014.2529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Wu B., Wang R., Li S., et al. Antifibrotic effects of Fraxetin on carbon tetrachloride-induced liver fibrosis by targeting NF-κB/IκBα, MAPKs and Bcl-2/Bax pathways. Pharmacological Reports. 2019;71(3):409–416. doi: 10.1016/j.pharep.2019.01.008. [DOI] [PubMed] [Google Scholar]
- 193.Singh D. K., Cheema H. S., Saxena A., et al. Fraxetin and ethyl acetate extract from Lawsonia inermis L. ameliorate oxidative stress in P. berghei infected mice by augmenting antioxidant defence system. Phytomedicine. 2017;36:262–272. doi: 10.1016/j.phymed.2017.09.012. [DOI] [PubMed] [Google Scholar]
- 194.Kundu J., Chae I. G., Chun K. S. Fraxetin induces heme oxygenase-1 expression by activation of Akt/Nrf2 or AMP-activated protein kinase α/Nrf2 pathway in HaCaT cells. Journal of Cancer Prevention. 2016;21(3):135–143. doi: 10.15430/JCP.2016.21.3.135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Thuong P. T., Pokharel Y. R., Lee M. Y., et al. Dual anti-oxidative effects of fraxetin isolated from Fraxinus rhinchophylla. Biological & Pharmaceutical Bulletin. 2009;32(9):1527–1532. doi: 10.1248/bpb.32.1527. [DOI] [PubMed] [Google Scholar]
- 196.Wang H., Xiao B., Hao Z., Sun Z. Simultaneous determination of fraxin and its metabolite, fraxetin, in rat plasma by liquid chromatography-tandem mass spectrometry and its application in a pharmacokinetic study. Journal of Chromatography B. 2016;1017-1018:70–74. doi: 10.1016/j.jchromb.2016.02.030. [DOI] [PubMed] [Google Scholar]
- 197.Niu X., Liu F., Li W., et al. Hepatoprotective effect of fraxin against carbon tetrachloride-induced hepatotoxicity in vitro and in vivo through regulating hepatic antioxidant, inflammation response and the MAPK-NF-κB signaling pathway. Biomedicine & Pharmacotherapy. 2017;95:1091–1102. doi: 10.1016/j.biopha.2017.09.029. [DOI] [PubMed] [Google Scholar]
- 198.Chang B., Jung Y., Yoon C.-S., et al. Fraxin prevents chemically induced hepatotoxicity by reducing oxidative stress. Molecules. 2017;22(4):p. 587. doi: 10.3390/molecules22040587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Li J. M., Zhang X., Wang X., Xie Y. C., Kong L. D. Protective effects of Cortex Fraxini coumarines against oxonate-induced hyperuricemia and renal dysfunction in mice. European Journal of Pharmacology. 2011;666(1-3):196–204. doi: 10.1016/j.ejphar.2011.05.021. [DOI] [PubMed] [Google Scholar]
- 200.Ma X., Liu X., Feng J., et al. Fraxin alleviates LPS-induced ARDS by downregulating inflammatory responses and oxidative damages and reducing pulmonary vascular permeability. Inflammation. 2019;42(5):1901–1912. doi: 10.1007/s10753-019-01052-8. [DOI] [PubMed] [Google Scholar]
- 201.Li W., Li W., Yu J. J., et al. Fraxin inhibits lipopolysaccharide‐induced inflammatory cytokines and protects against endotoxic shock in mice. Fundamental & Clinical Pharmacology. 2019;34(1):91–101. doi: 10.1111/fcp.12500. [DOI] [PubMed] [Google Scholar]
- 202.Li W., Li W., Zang L., et al. Fraxin ameliorates lipopolysaccharide-induced acute lung injury in mice by inhibiting the NF-κB and NLRP3 signalling pathways. International Immunopharmacology. 2019;67:1–12. doi: 10.1016/j.intimp.2018.12.003. [DOI] [PubMed] [Google Scholar]
- 203.Topdağı Ö., Tanyeli A., Akdemir F. N. E., Eraslan E., Güler M. C., Çomaklı S. Preventive effects of fraxin on ischemia/reperfusion-induced acute kidney injury in rats. Life Sciences. 2020;242:p. 117217. doi: 10.1016/j.lfs.2019.117217. [DOI] [PubMed] [Google Scholar]
- 204.Matsuda H., Murakami T., Kageura T., et al. Hepatoprotective and nitric oxide production inhibitory activities of coumarin and polyacetylene constituents from the roots of Angelica furcijuga. Bioorganic & Medicinal Chemistry Letters. 1998;8(16):2191–2196. doi: 10.1016/S0960-894X(98)00391-6. [DOI] [PubMed] [Google Scholar]
- 205.Yoshikawa M., Nishida N., Ninomiya K., et al. Inhibitory effects of coumarin and acetylene constituents from the roots of Angelica furcijuga on D-galactosamine/lipopolysaccharide-induced liver injury in mice and on nitric oxide production in lipopolysaccharide-activated mouse peritoneal macrophages. Bioorganic & Medicinal Chemistry. 2006;14(2):456–463. doi: 10.1016/j.bmc.2005.08.038. [DOI] [PubMed] [Google Scholar]
- 206.Khan S., Shin E. M., Choi R. J., et al. Suppression of LPS-induced inflammatory and NF-κB responses by anomalin in RAW 264.7 macrophages. Journal of Cellular Biochemistry. 2011;112(8):2179–2188. doi: 10.1002/jcb.23137. [DOI] [PubMed] [Google Scholar]
- 207.Khan A., Khan S., Ali H., et al. Anomalin attenuates LPS-induced acute lungs injury through inhibition of AP-1 signaling. International Immunopharmacology. 2019;73:451–460. doi: 10.1016/j.intimp.2019.05.032. [DOI] [PubMed] [Google Scholar]
- 208.Khan S., Choi R. J., Lee J., Kim Y. S. Attenuation of neuropathic pain and neuroinflammatory responses by a pyranocoumarin derivative, anomalin in animal and cellular models. European Journal of Pharmacology. 2016;774:95–104. doi: 10.1016/j.ejphar.2016.02.008. [DOI] [PubMed] [Google Scholar]
- 209.Sarveswaran S., Gautam S. C., Ghosh J. Wedelolactone, a medicinal plant-derived coumestan, induces caspase-dependent apoptosis in prostate cancer cells via downregulation of PKCε without inhibiting Akt. International Journal of Oncology. 2012;41(6):2191–2199. doi: 10.3892/ijo.2012.1664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Lee Y. J., Lin W. L., Chen N. F., Chuang S. K., Tseng T. H. Demethylwedelolactone derivatives inhibit invasive growth in vitro and lung metastasis of MDA-MB-231 breast cancer cells in nude mice. European Journal of Medicinal Chemistry. 2012;56:361–367. doi: 10.1016/j.ejmech.2012.07.041. [DOI] [PubMed] [Google Scholar]
- 211.Vender J. R., Laird M. D., Dhandapani K. M. Inhibition Of Nfκb Reduces Cellular Viability In Gh3 Pituitary Adenoma Cells. Neurosurgery. 2008;62(5):1122–1128. doi: 10.1227/01.neu.0000325874.82999.75. [DOI] [PubMed] [Google Scholar]
- 212.Maya S., Prakash T., Goli D. Effect of wedelolactone and gallic acid on quinolinic acid-induced neurotoxicity and impaired motor function: significance to sporadic amyotrophic lateral sclerosis. NeuroToxicology. 2018;68:1–12. doi: 10.1016/j.neuro.2018.06.015. [DOI] [PubMed] [Google Scholar]
- 213.Cuong T. T., Diem G. H., Doan T. T., Huy N. Q., Phuong N., Hung H. T. Wedelolactone from Vietnamese Eclipta prostrata (L) L. protected zymosan-induced shock in mice. Iranian Journal of Pharmaceutical Research. 2018;17(2):653–660. [PMC free article] [PubMed] [Google Scholar]
- 214.Melo P. A., Do Nascimento M. C., Mors W. B., Suarez-Kurtz G. Inhibition of the myotoxic and hemorrhagic activities of crotalid venoms by Eclipta prostrata (Asteraceae) extracts and constituents. Toxicon. 1994;32(5):595–603. doi: 10.1016/0041-0101(94)90207-0. [DOI] [PubMed] [Google Scholar]
- 215.Zhu M. M., Wang L., Yang D., et al. Wedelolactone alleviates doxorubicin-induced inflammation and oxidative stress damage of podocytes by IκK/IκB/NF-κB pathway. Biomedicine & Pharmacotherapy. 2019;117:p. 109088. doi: 10.1016/j.biopha.2019.109088. [DOI] [PubMed] [Google Scholar]
- 216.Deng X., Liang L. N., Zhu D., et al. Wedelolactone inhibits osteoclastogenesis but enhances osteoblastogenesis through altering different semaphorins production. International Immunopharmacology. 2018;60:41–49. doi: 10.1016/j.intimp.2018.04.037. [DOI] [PubMed] [Google Scholar]
- 217.Zhu D., Deng X., Han X. F., et al. Wedelolactone enhances osteoblastogenesis through ERK- and JNK-mediated BMP2 expression and Smad/1/5/8 phosphorylation. Molecules. 2018;23(3):p. 561. doi: 10.3390/molecules23030561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Kobori M., Yang Z., Gong D., et al. Wedelolactone suppresses LPS-induced caspase-11 expression by directly inhibiting the IKK complex. Cell Death & Differentiation. 2004;11(1):123–130. doi: 10.1038/sj.cdd.4401325. [DOI] [PubMed] [Google Scholar]
- 219.Lin W. L., Wang S. M., Ho Y. J., Kuo H. C., Lee Y. J., Tseng T. H. Ethyl acetate extract of Wedelia chinensis inhibits tert-butyl hydroperoxide-induced damage in PC12 cells and D-galactose-induced neuronal cell loss in mice. BMC Complementary and Alternative Medicine. 2014;14(1) doi: 10.1186/1472-6882-14-491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Ding S., Hou X., Yuan J., et al. Wedelolactone protects human bronchial epithelial cell injury against cigarette smoke extract-induced oxidant stress and inflammation responses through Nrf2 pathway. International Immunopharmacology. 2015;29(2):648–655. doi: 10.1016/j.intimp.2015.09.015. [DOI] [PubMed] [Google Scholar]
- 221.Zhang E., Yin S., Zhao S., et al. Protective effects of glycycoumarin on liver diseases. Phytotherapy Research. 2019 doi: 10.1002/ptr.6598. [DOI] [PubMed] [Google Scholar]
- 222.Kırmızıbekmez H., Uysal G. B., Masullo M., et al. Prenylated polyphenolic compounds from Glycyrrhiza iconica and their antimicrobial and antioxidant activities. Fitoterapia. 2015;103:289–293. doi: 10.1016/j.fitote.2015.05.003. [DOI] [PubMed] [Google Scholar]
- 223.Fu Y., Chen J., Li Y. J., Zheng Y. F., Li P. Antioxidant and anti-inflammatory activities of six flavonoids separated from licorice. Food Chemistry. 2013;141(2):1063–1071. doi: 10.1016/j.foodchem.2013.03.089. [DOI] [PubMed] [Google Scholar]
- 224.Tanaka Y., Kikuzaki H., Fukuda S., Nakatani N. Antibacterial compounds of licorice against upper airway respiratory tract pathogens. Journal of Nutritional Science and Vitaminology. 2001;47(3):270–273. doi: 10.3177/jnsv.47.270. [DOI] [PubMed] [Google Scholar]
- 225.Song X., Yin S., Huo Y., et al. Glycycoumarin ameliorates alcohol-induced hepatotoxicity via activation of Nrf2 and autophagy. Free Radical Biology and Medicine. 2015;89:135–146. doi: 10.1016/j.freeradbiomed.2015.07.006. [DOI] [PubMed] [Google Scholar]
- 226.Yan M., Ye L., Yin S., et al. Glycycoumarin protects mice against acetaminophen-induced liver injury predominantly via activating sustained autophagy. British Journal of Pharmacology. 2018;175(19):3747–3757. doi: 10.1111/bph.14444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.You L., Feng S., An R., Wang X. Osthole: a promising lead compound for drug discovery from a traditional Chinese medicine (TCM) Natural product communications. 2009;4(2):297–302. [PubMed] [Google Scholar]
- 228.Zhou Z. W., Liu P. X. Progress in study of chemical constituents and anti-tumor activities of Cnidium monnieri. Zhongguo Zhong yao za zhi = Zhongguo zhongyao zazhi = China journal of Chinese materia medica. 2005;30(17):1309–1313. [PubMed] [Google Scholar]
- 229.Waqas M., Wang Y., Li A., et al. Osthole: a coumarin derivative assuage thiram-induced tibial dyschondroplasia by regulating BMP-2 and RUNX-2 expressions in chickens. Antioxidants. 2019;8(9):p. 330. doi: 10.3390/antiox8090330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Zhou W. B., Zhang X. X., Cai Y., Sun W., Li H. Osthole prevents tamoxifen-induced liver injury in mice. Acta Pharmacologica Sinica. 2019;40(5):608–619. doi: 10.1038/s41401-018-0171-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Khairy H., Saleh H., Badr A. M., Marie M. A. S. Therapeutic efficacy of osthole against dinitrobenzene sulphonic acid induced- colitis in rats. Biomedicine & Pharmacotherapy. 2018;100:42–51. doi: 10.1016/j.biopha.2018.01.104. [DOI] [PubMed] [Google Scholar]
- 232.Jiang G., Liu J., Ren B., et al. Anti-tumor effects of osthole on ovarian cancer cells in vitro. Journal of Ethnopharmacology. 2016;193:368–376. doi: 10.1016/j.jep.2016.08.045. [DOI] [PubMed] [Google Scholar]
- 233.Ye Y., Han X., Guo B., Sun Z., Liu S. Combination treatment with platycodin D and osthole inhibits cell proliferation and invasion in mammary carcinoma cell lines. Environmental Toxicology and Pharmacology. 2013;36(1):115–124. doi: 10.1016/j.etap.2013.03.012. [DOI] [PubMed] [Google Scholar]
- 234.Fan H., Gao Z., Ji K., et al. The in vitro and in vivo anti-inflammatory effect of osthole, the major natural coumarin from Cnidium monnieri (L.) Cuss, via the blocking of the activation of the NF-κB and MAPK/p38 pathways. Phytomedicine. 2019;58:p. 152864. doi: 10.1016/j.phymed.2019.152864. [DOI] [PubMed] [Google Scholar]
- 235.Jarząb A., Grabarska A., Skalicka-Woźniak K., Stepulak A. Pharmacological features of osthole. Postępy Higieny i Medycyny Doświadczalnej. 2017;71(1):p. 0. doi: 10.5604/01.3001.0010.3824. [DOI] [PubMed] [Google Scholar]
- 236.Liu J., Zschocke S., Reininger E., Bauer R. Inhibitory Effects ofAngelica pubescensf.biserrataon 5-Lipoxygenase and Cyclooxygenase. Planta Medica. 1998;64(6):525–529. doi: 10.1055/s-2006-957507. [DOI] [PubMed] [Google Scholar]
- 237.Resch M., Steigel A., Chen Z. L., Bauer R. 5-Lipoxygenase and cyclooxygenase-1 inhibitory active compounds from Atractylodes lancea. Journal of Natural Products. 1998;61(3):347–350. doi: 10.1021/np970430b. [DOI] [PubMed] [Google Scholar]
- 238.Nakamura T., Kodama N., Arai Y., et al. Inhibitory effect of oxycoumarins isolated from the Thai medicinal plant Clausena guillauminii on the inflammation mediators, iNOS, TNF-alpha, and COX-2 expression in mouse macrophage RAW 264.7. Journal of Natural Medicines. 2009;63(1):21–27. doi: 10.1007/s11418-008-0277-5. [DOI] [PubMed] [Google Scholar]
- 239.Liao P. C., Chien S. C., Ho C. L., et al. Osthole regulates inflammatory mediator expression through modulating NF-κB, mitogen-activated protein kinases, protein kinase C, and reactive oxygen species. Journal of Agricultural and Food Chemistry. 2010;58(19):10445–10451. doi: 10.1021/jf102812t. [DOI] [PubMed] [Google Scholar]
- 240.Chiu P. R., Lee W. T., Chu Y. T., Lee M. S., Jong Y. J., Hung C. H. Effect of the Chinese herb extract osthol on IL-4-induced eotaxin expression in BEAS-2B cells. Pediatrics & Neonatology. 2008;49(4):135–140. doi: 10.1016/S1875-9572(08)60028-5. [DOI] [PubMed] [Google Scholar]
- 241.Liu J., Zhang W., Zhou L., Wang X., Lian Q. Anti-inflammatory effect and mechanism of osthole in rats. Zhong yao cai = Zhongyaocai = Journal of Chinese medicinal materials. 2005;28(11):1002–1006. [PubMed] [Google Scholar]
- 242.Wei M., Zhang J. J., He Q. L., et al. Effects and mechanisms of osthole on sciatica induced by lumber disc herniation. Zhong yao cai = Zhongyaocai = Journal of Chinese medicinal materials. 2011;34(5):746–750. [PubMed] [Google Scholar]
- 243.Chu Q., Zhu Y., Cao T., et al. Studies on the neuroprotection of osthole on glutamate-induced apoptotic cells and an Alzheimer's disease mouse model via modulation oxidative stress. Applied Biochemistry and Biotechnology. 2020;190(2):634–644. doi: 10.1007/s12010-019-03101-2. [DOI] [PubMed] [Google Scholar]
- 244.Tao L., Gu X., Xu E., et al. Osthole protects against Ang II-induced endotheliocyte death by targeting NF-κB pathway and Keap-1/Nrf2 pathway. American Journal of Translational Research. 2019;11(1):142–159. [PMC free article] [PubMed] [Google Scholar]
- 245.Bao Y., Meng X., Liu F., et al. Protective effects of osthole against inflammation induced by lipopolysaccharide in BV2 cells. Molecular Medicine Reports. 2018;17(3):4561–4566. doi: 10.3892/mmr.2018.8447. [DOI] [PubMed] [Google Scholar]
- 246.Chen Z., Mao X., Liu A., et al. Osthole, a natural coumarin improves cognitive impairments and BBB dysfunction after transient global brain ischemia in C57 BL/6J mice: involvement of Nrf2 pathway. Neurochemical Research. 2015;40(1):186–194. doi: 10.1007/s11064-014-1483-z. [DOI] [PubMed] [Google Scholar]
- 247.Yang S. M., Chan Y. L., Hua K. F., et al. Osthole improves an accelerated focal segmental glomerulosclerosis model in the early stage by activating the Nrf2 antioxidant pathway and subsequently inhibiting NF-κB-mediated COX-2 expression and apoptosis. Free Radical Biology and Medicine. 2014;73:260–269. doi: 10.1016/j.freeradbiomed.2014.05.009. [DOI] [PubMed] [Google Scholar]
- 248.Hua K. F., Yang S. M., Kao T. Y., et al. Osthole mitigates progressive IgA nephropathy by inhibiting reactive oxygen species generation and NF-κB/NLRP3 pathway. PloS one. 2013;8(10, article e77794) doi: 10.1371/journal.pone.0077794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Chen X. J., Zhang B., Hou S. J., et al. Osthole improves acute lung injury in mice by up-regulating Nrf-2/thioredoxin 1. Respiratory physiology & neurobiology. 2013;188(2):214–222. doi: 10.1016/j.resp.2013.04.014. [DOI] [PubMed] [Google Scholar]
- 250.Kim H.-J., Kang C.-H., Jayasooriya R. G. P. T., et al. Hydrangenol inhibits lipopolysaccharide-induced nitric oxide production in BV2 microglial cells by suppressing the NF-κB pathway and activating the Nrf2-mediated HO-1 pathway. International Immunopharmacology. 2016;35:61–69. doi: 10.1016/j.intimp.2016.03.022. [DOI] [PubMed] [Google Scholar]
- 251.Zhang H., Matsuda H., Kumahara A., Ito Y., Nakamura S., Yoshikawa M. New type of anti-diabetic compounds from the processed leaves of Hydrangea macrophylla var. thunbergii (Hydrangeae Dulcis Folium) Bioorganic & Medicinal Chemistry Letters. 2007;17(17):4972–4976. doi: 10.1016/j.bmcl.2007.06.027. [DOI] [PubMed] [Google Scholar]
- 252.Myung D. B., Han H. S., Shin J. S., et al. Hydrangenol isolated from the leaves of Hydrangea serrata attenuates wrinkle formation and repairs skin moisture in UVB-irradiated hairless mice. Nutrients. 2019;11(10):p. 2354. doi: 10.3390/nu11102354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Shin S. S., Ko M. C., Park Y. J., et al. Hydrangenol inhibits the proliferation, migration, and invasion of EJ bladder cancer cells via p21WAF1-mediated G1-phase cell cycle arrest, p38 MAPK activation, and reduction in Sp-1-induced MMP-9 expression. EXCLI journal. 2018;17:531–543. doi: 10.17179/excli2018-1361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Gho Y., Shin S. S., Choi Y. H., Ko K., Kim W. J., Moon S. K. Hydrangenol suppresses VEGF-stimulated angiogenesis by targeting p27KIP1-dependent G1-cell cycle arrest, VEGFR-2-mediated signaling, and MMP-2 expression. Animal Cells and Systems. 2019;23(2):72–81. doi: 10.1080/19768354.2019.1578262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Znati M., Ben Jannet H., Cazaux S., Souchard J., Harzallah Skhiri F., Bouajila J. Antioxidant, 5-lipoxygenase inhibitory and cytotoxic activities of compounds isolated from the Ferula lutea flowers. Molecules. 2014;19(10):16959–16975. doi: 10.3390/molecules191016959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256.Pokharel Y. R., Han E. H., Kim J. Y., et al. Potent protective effect of isoimperatorin against aflatoxin B1-inducible cytotoxicity in H4IIE cells: bifunctional effects on glutathione S-transferase and CYP1A. Carcinogenesis. 2006;27(12):2483–2490. doi: 10.1093/carcin/bgl118. [DOI] [PubMed] [Google Scholar]
- 257.Wijerathne C. U. B., Seo C. S., Song J. W., et al. Isoimperatorin attenuates airway inflammation and mucus hypersecretion in an ovalbumin-induced murine model of asthma. International Immunopharmacology. 2017;49:67–76. doi: 10.1016/j.intimp.2017.05.012. [DOI] [PubMed] [Google Scholar]
- 258.Meng-yue W. A. N. G., Min-ru J. I. A., Yu-ying M. A., Xiao-bo L. I. Pharmacological effect of four linear furocoumarins in Radix Angelicae dahuricae. 2010;22(3):485–489. [Google Scholar]
- 259.Tan N., Yazıcı-Tütüniş S., Bilgin M., Tan E., Miski M. Antibacterial activities of pyrenylated coumarins from the roots of Prangos hulusii. Molecules. 2017;22(7):p. 1098. doi: 10.3390/molecules22071098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Guo N., Wu J., Fan J., et al. In vitro activity of isoimperatorin, alone and in combination, against Mycobacterium tuberculosis. Letters in Applied Microbiology. 2014;58(4):344–349. doi: 10.1111/lam.12195. [DOI] [PubMed] [Google Scholar]
- 261.Tong K., Xin C., Chen W. Isoimperatorin induces apoptosis of the SGC-7901 human gastric cancer cell line via the mitochondria-mediated pathway. Oncology Letters. 2017;13(1):518–524. doi: 10.3892/ol.2016.5387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Yang H. B., Gao H. R., Ren Y. J., et al. Effects of isoimperatorin on proliferation and apoptosis of human gastric carcinoma cells. Oncology Letters. 2018;15(5):7993–7998. doi: 10.3892/ol.2018.8303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Moon L., Ha Y. M., Jang H. J., et al. Isoimperatorin, cimiside E and 23-O-acetylshengmanol-3-xyloside from Cimicifugae Rhizome inhibit TNF-α-induced VCAM-1 expression in human endothelial cells: Involvement of PPAR-γ upregulation and PI3K, ERK1/2, and PKC signal pathways. Journal of Ethnopharmacology. 2011;133(2):336–344. doi: 10.1016/j.jep.2010.10.004. [DOI] [PubMed] [Google Scholar]
- 264.Cui G. H., Chen W. Q., Shen Z. Y. Urolithin A shows anti-atherosclerotic activity via activation of class B scavenger receptor and activation of Nef2 signaling pathway. Pharmacological Reports. 2018;70(3):519–524. doi: 10.1016/j.pharep.2017.04.020. [DOI] [PubMed] [Google Scholar]
- 265.Kumar A., Jha S., Pattanayak S. P. Daphnetin ameliorates 7,12-dimethylbenz[a]anthracene-induced mammary carcinogenesis through Nrf-2-Keap1 and NF-κB pathways. Biomedicine & Pharmacotherapy. 2016;82:439–448. doi: 10.1016/j.biopha.2016.05.028. [DOI] [PubMed] [Google Scholar]