

Abstract
The National Institutes of Health (NIH) has long supported using nonhuman primate (NHP) models for research on kidney, pancreatic islet, heart, and lung transplantation. The primary purpose of this research has been to develop new treatments for down-modulating or preventing deleterious immune responses after transplantation in human patients. Here, we discuss NIH-funded NHP studies of immune cell depletion, costimulation blockade, regulatory cell therapy, desensitization, and mixed hematopoietic chimerism that either preceded clinical trials or prevented the human application of therapies that were toxic or ineffective.
INTRODUCTION
Despite excellent short-term outcomes after organ transplantation with current immunosuppressive drug regimens, long-term outcomes remain disappointing due to chronic rejection of allografts (organ transplants from genetically nonidentical donors of the same species) and the morbidities of lifelong immunosuppression. To address these challenges, the development of new, less toxic, targeted therapeutics to prevent rejection or to establish and maintain immune tolerance to allografts is required. Many immunomodulating therapies lead to allograft tolerance in rodent models, but few successfully translate to nonhuman primate (NHP) models or to humans (1). The NHP model represents a compelling means to rigorously evaluate candidate immunosuppressive strategies as an ethical precursor to human clinical trials. NHP experimentation meets a critical need in the translation of tolerance protocols to the clinic by rigorously evaluating candidate tolerance strategies aimed at the withdrawal of standard-of-care immunosuppression before they are applied in human clinical trials. Favorable NHP results are mandatory before initiating a human trial that includes complete immunosuppression withdrawal. Genetic similarities to humans, emerging molecular tools to assess NHP histocompatibility complex antigens, and cross-reactivity between drug targets of humans and NHPs confer on NHP models an ability to test drugs and therapeutic strategies intended for human patients undergoing transplantation (2).
In 1976, the National Heart, Lung, and Blood Institute of the National Institutes of Health (NIH) funded one of the first grants that included research on tolerance induction in NHP kidney and heart transplant recipients. It generated preliminary data that ultimately led to immunological tolerance in a small number of human kidney transplant recipients (3). In 1999, the National Institute of Allergy and Infectious Diseases (NIAID), with cofunding by the National Institute of Diabetes and Digestive and Kidney Diseases, established the Nonhuman Primate Transplantation Tolerance Cooperative Study Group to develop and evaluate new immunosuppressive drug regimens and to develop biomarkers for rejection and tolerance of kidney, pancreatic islet, and (since 2004) heart and lung transplants.
These models have substantially improved over the years with refinements in surgical techniques, postoperative care, colony management, and advances in NHP major histocompatibility complex (MHC) typing. In addition, filling a critical need in NHP research, the NIH’s NHP Reagent Resource (www.nhpreagents.org/NHP/default.aspx) works with investigators to develop and provide NHP-specific research reagents and immunotherapeutics for a range of research fields. Whereas no animal model perfectly mirrors clinical events in humans, here we present examples illustrating that NHP research is a critical and necessary step in the development of human therapeutics for transplantation (Fig. 1).
Fig. 1. The transplantation research cycle.
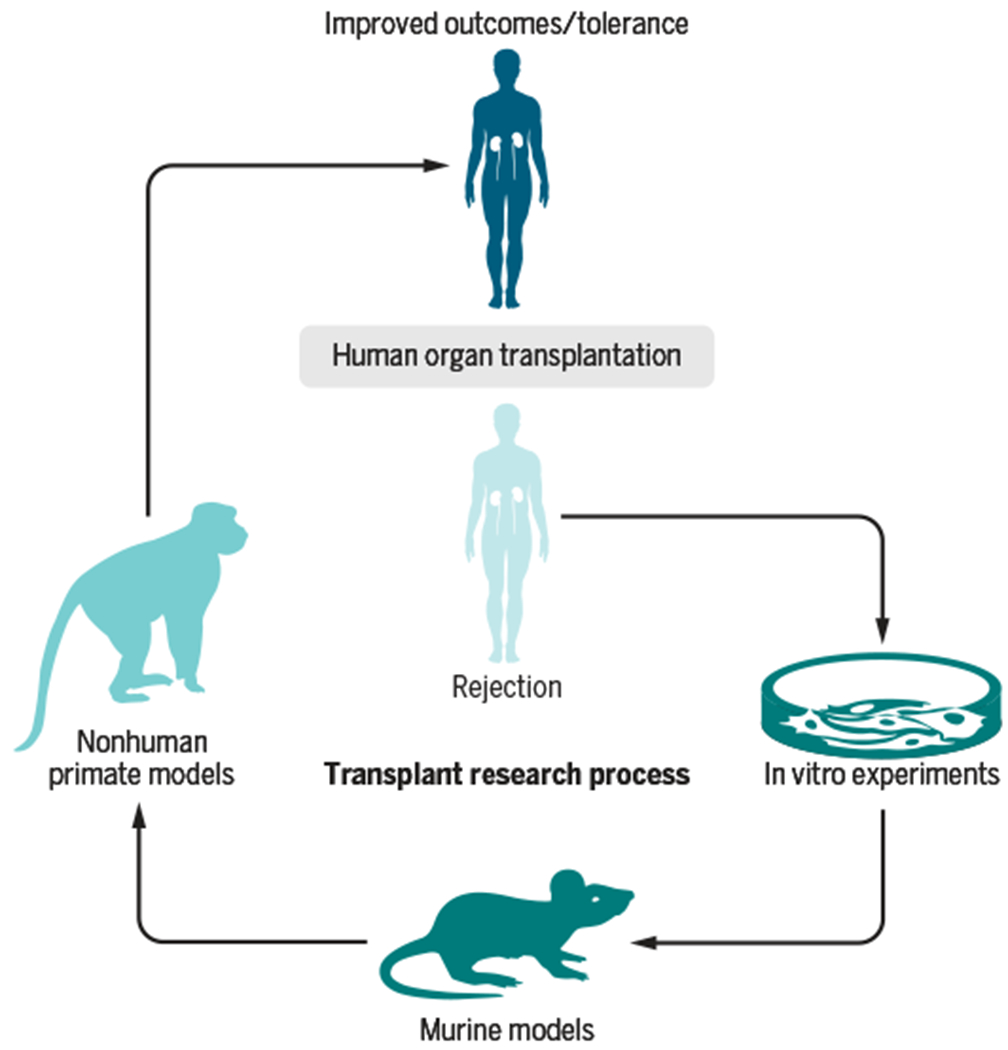
The cycle begins with mechanistic diagnosis of the clinical problem of tissue rejection in human transplant patients. It progresses to a hypothesis regarding potential solutions. The next step involves testing in vitro experimental solutions and selecting the most promising new therapies for in vivo evaluation in murine models of organ transplantation. This is followed by more discriminating experimental evaluation of the therapy in nonhuman primate (NHP) transplant models. Successful NHP therapies progress to controlled human clinical trials before FDA approval for general clinical use in transplant patients.
COSTIMULATION BLOCKADE IMMUNOTHERAPEUTICS IN TRANSPLANTATION
The costimulatory pathways represent one of the most effective targets for new transplant immunotherapeutics to date. The application of costimulation blockade in transplant models began with the finding that combined CD28-CD80/CD86 and CD40-CD154 blockade prevented allogeneic skin graft rejection in mice (4, 5). The subsequent demonstration of the power of costimulation blockade in an NHP renal allograft model (6) spurred efforts to develop CTLA4-Ig (CD80/CD86 blockade) and anti-CD154/CD40 antibody for clinical use (Table 1). It also catalyzed the establishment of NIAID’s Immune Tolerance Network for clinical trials of new strategies to induce tolerance in allergy, asthma, autoimmune disease, and kidney and islet transplantation.
Table 1.
Select immunosuppressive agents and their targets.
| Generic name | Brand or other name | Primary target/action | FDA approved for use in human transplantation |
|---|---|---|---|
| Abatacept | CTLA4-Ig | Blocks CD80/CD86 on antigen-presenting cells; prevents CD28 T cell activation | Off label |
| Alemtuzumab | Campath1-H | Depletes T cells, some B cells (not plasma cells). Expression on other immune cells varies | Off label |
| Anti-CD52 | |||
| Lemtrada | |||
| Alefacept | LFA3-Ig | Blocks LFA3/CD2, results in T memory cell apoptosis | No |
| Anti-thymocyte globulin | ATG | Primarily T cell depletion | Yes |
| Thymoglobulin | |||
| Basiliximab | Anti-IL-2R antibody | Targets IL-2Rα (CD25) on activated T cells; blocks IL-2R | Yes |
| Belatacept | LEA29Y | Same as abatacept, but higher affinity | Yes |
| (CTLA4-Ig variant) | |||
| Nulojix | |||
| Bortezomib | Proteasome inhibitor | Plasma cell apoptosis | Off label |
| Cyclophosphamide | Cytoxan; CYP | Myeloablative drug | Off label |
| Cyclosporine | CSA | Calcineurin inhibitor; prevents IL-2 production | Yes |
| Efalizumab | Anti-LFA-1 | Targets T memory cells | No |
| Raptiva | |||
| Mycophenolate Mofetil | MMF | Inhibits T and B cell proliferation | Yes |
| Rituximab | Anti-CD20 antibody | Targets CD20 on B cells (not plasma cells) and induces their apoptosis | Off label |
| Rituxan | |||
| Sirolimus | Rapamycin/Rapamune | mTOR inhibitor; blocks T and B cell activation/proliferation | Yes |
| Tacrolimus | FK506 | Calcineurin inhibitor; prevents IL-2 production | Yes |
Clinical development of CD80/CD86 blockade was heavily informed by work in NHP models, including the adoption of extended rather than short-term costimulation blockade to prevent rejection (7). Reduced efficacy in NHPs as compared to rodent models led to the realization that a more potent form of CTLA4-Ig was needed, and ultimately, this led to the development of the biologic belatacept. Subsequent testing in NHP allograft models supported CTLA4-Ig’s clinical translation (8, 9). Compared to the immunosuppressive drug cyclosporine (a calcineurin inhibitor), belatacept demonstrated improved renal allograft function in two phase 3 clinical trials after 2 years (10). This led to U.S. Food and Drug Administration (FDA) approval of belatacept for use in human kidney transplantation (Fig. 2). Translation of NHP studies to a pancreatic islet transplantation clinical trial confirmed the efficacy of immunosuppression with belatacept in the absence of maintenance therapy with a calcineurin inhibitor. This trial relied on anti-thymocyte globulin (ATG) induction with belatacept plus rapamycin and mycophenolate maintenance therapy. This drug combination resulted in insulin independence for more than 1 year in four of five patients after allogeneic islet transplant from single donors (11, 12).
Fig. 2. Timeline for development of belatacept.

Blocking the CD28-CD80/CD86 costimulatory pathway has evolved from a mechanistic in vitro experimental tool to in vivo application for solid organ and islet transplantation, first in murine models and then in NHPs. In NHP transplant models, it was recognized that continuous therapy to prevent organ rejection would be more clinically useful than induction of tolerance and that a higher affinity molecule (belatacept) would be preferable to CTLA4-Ig. The FDA approved belatacept for kidney transplantation in 2010, 24 years after CTLA4-Ig was originally developed.
When similar efforts to block CD40 ligand with anti-CD154 antibody resulted in thrombotic complications in both NHPs and humans (13), the development focus shifted to direct blockade of CD40. After encouraging NHP transplant results (14–18), new antibodies against CD40 are in clinical trials, with results pending (ClinicalTrials.gov NCT01780844 and NCT02217410). As described below, immunotherapeutics administered in combination with belatacept or anti-CD40 antibody are currently under investigation in NHP models, reflecting the value of this preclinical system for optimization of therapeutics before clinical application.
INDUCTION OF MIXED CHIMERISM IN TRANSPLANTATION
In a murine model, nonmyeloablative conditioning followed by bone marrow transplantation (BMT) led to mixed chimerism and subsequent tolerance to an allogeneic skin graft (19). Mixed chimerism implies stable coexistence of donor and recipient lymphohematopoietic cells in a transplant recipient. Translating this approach to NHP kidney transplantation required successive modifications of the conditioning regimen, finally including total body irradiation, thymic irradiation, and ATG, followed by combined BMT and kidney transplantation, splenectomy, and short-term cyclosporine immunosuppressive treatment (20). Most (9 of 13) NHP transplant recipients had long-term normal renal function without maintenance immunosuppression despite losing chimerism within months of BMT, indicating that long-term tolerance relied upon peripheral rather than central tolerance mechanisms. The same regimen did not achieve tolerance of cardiac allografts (21), suggesting that organ-specific differences influence the effectiveness of transient mixed chimerism to induce tolerance. Murine studies demonstrating that costimulation blockade promoted durable chimerism (22) led to the successful use of anti-CD154 antibody without splenectomy in NHP studies, although chimerism remained transient (23). When this costimulation blockade mixed-chimerism approach was applied to islet allografts, monkeys failed to achieve tolerance (24) unless transplanted with a kidney as a composite allograft (25). Similarly, a kidney cotransplanted with a heart under the mixed-chimerism protocol conferred tolerance to both the kidney and heart (26). Although activation and expansion of host T regulatory cells (Tregs) occurs, understanding the underlying mechanisms whereby a renal allograft confers tolerance to a cotransplanted heart or to islets is an area of active research (26).
The early NHP results led to clinical application in human leukocyte antigen (HLA)–mismatched living-donor kidney transplantation. Using a human-adapted conditioning regimen that included cyclophosphamide, anti-CD2 antibody, and rituximab, 10 patients received kidney transplants simultaneously with donor BMT. Three patients lost their grafts due to antibody-mediated rejection at 10 days, thrombotic microangiopathy at 6 months, and cellular rejection at 3 years after transplantation (27). Immunosuppressive drugs were weaned from the remaining seven patients. Three of these seven required reinstitution of maintenance immunosuppression because of recurrent disease or chronic rejection at 6, 8, and 9 years after transplantation. The other four recipients remained off all immunosuppression for periods of up to 15 years. These results demonstrate that like NHPs, but unlike mice, some humans can develop long-lasting tolerance even after loss of chimerism (28). However, the morbidity associated with this conditioning regimen prevents widespread application. Current NHP research efforts are focused on minimizing toxicity of this regimen.
To extend mixed chimerism to recipients of deceased donor organs, researchers developed NHP delayed-tolerance protocols incorporating costimulation blockade wherein recipient conditioning and BMT occur months after organ transplant. This approach proved effective in inducing tolerance to kidney allografts after addition of CD8+ T cell–depleting monoclonal antibodies (29) and to lung allografts after addition of anti–interleukin-6 receptor (IL-6R) monoclonal antibodies (30).
In NHP hematopoietic stem cell transplantation models of mixed chimerism, costimulation blockade with both belatacept and anti-CD154 antibody (31) or nondepleting anti-CD40 antibody and CTLA4-Ig (32) prolonged chimerism and promoted engraftment in fully MHC disparate or one MHC haplotype-mismatched donors, respectively. These NHP results contributed to the application of CTLA4-Ig (abatacept) for the prevention of acute graft-versus-host disease (GVHD) during unrelated donor hematopoietic stem cell transplantation (Clinicaltrials.gov NCT01743131) (33). Thus, whereas mixed-chimerism models were essential to achieving tolerance in some human recipients, current results highlight the need for additional NHP research to improve the safety and consistency of this transplant tolerance approach.
IMMUNE CELL DEPLETION IN TRANSPLANTATION
NHP experiments suggested that T cell depletion in lymph nodes in addition to blood using NHP-specific anti-CD3 immunotoxin could induce peripheral tolerance to most renal and skin allografts (34). Long-term follow-up of these “tolerant” monkeys revealed a high incidence of alloantibody development more than 1 year after kidney transplantation, chronic allograft injury, and eventual graft failure (mean >600 days after transplant) (35). Human translation of this principle of profound T cell depletion required switching from the anti-CD3 immunotoxin to alemtuzumab, a monoclonal antibody that targets human CD52 (Table 1). Whereas alemtuzumab allowed for a reduction in maintenance immunosuppression (36), tolerance was not achieved in humans. Studies in both NHPs and humans revealed that T cell depletion was associated with homeostatic proliferation of inefficiently targeted memory T cells (37, 38) and increased expression of BAFF (the cytokine B cell activating factor) resulting in B cell activation (39, 40); this suggested that additional immunosuppression would be required. A clinical trial of alemtuzumab for tolerance induction followed by belatacept maintenance therapy with or without rapamycin led to rejection-free survival of renal allografts, although tolerance was not directly tested (41). This research increased our understanding of the role of T memory cells in allograft rejection and as a barrier to tolerance. NHP studies helped to expand clinical use of T cell–depleting reagents; today, T cell–depleting antibodies are used in 65% of kidney transplant patients in the United States (srtr.transplant.hrsa.gov) to reduce rejection and graft loss.
Induction therapy, that is, intense prophylactic immunosuppression delivered at the time of the transplant, with rituximab that depletes B cells, was beneficial in two NHP transplant models, but this success did not translate to humans. In an NHP cardiac transplant model, preemptive treatment with rituximab inhibited alloantibody production and reduced cardiac allograft vasculopathy (42). This encouraging finding led to the CTOT-11 clinical trial (ClinicalTrials.gov NCT011278745), which tested whether heart transplant patients receiving rituximab at the time of transplant would have less cardiac allograft vasculopathy, measured as the percent change in atheroma volume, compared to transplant recipients receiving placebo. Surprisingly, rituximab-treated patients had a greater percent change in atheroma volume compared to controls at 1 year after transplantation (43). NHP islet transplant recipients experienced long-term rejection-free survival when treated at the time of transplant with ATG and rituximab, followed by rapamycin maintenance immunosuppression (44). However, when islet transplant patients received the same therapy, the trial was terminated because of a lack of efficacy (ClinicalTrials.gov NCT00468442). In renal transplantation, recipients receiving rituximab and methylprednisolone followed by tacrolimus and mycophenolate mofetil maintenance therapy paradoxically experienced increased rejection, halting the clinical trial (45). Thus, whereas NHP B cell depletion experiments provided mechanistic insights to guide human transplant strategies, the benefits seen in NHP models did not translate to humans. Understanding the reasons behind these different outcomes will help to refine the translational potential of the NHP model.
TARGETING T MEMORY CELLS IN TRANSPLANTATION
Evidence supports an important role for T memory cells in transplant rejection, yet immunosuppressive drugs that have been evaluated or approved for transplantation do not adequately target this population (46). Leukocyte functional antigen-1 (LFA-1) is a protein preferentially expressed by T memory cells. Promising results were achieved by blocking LFA-1 in an NHP islet transplant model (47). In an islet transplantation clinical trial, all eight recipients treated with efalizumab instead of a calcineurin inhibitor achieved stable insulin independence after one or two islet transplants (48). However, in an NHP renal allograft model, graft survival was not prolonged and protective immunity was compromised (49). Despite encouraging outcomes in islet transplantation, long-term use of efalizumab in the clinic for treatment of psoriasis was associated with the development of progressive multifocal leukoencephalopathy, leading to its voluntary withdrawal from the market (50). Other approaches to reduce T memory cells in NHPs, such as LFA3-Ig, have shown limited success (51). T cell memory responses, in particular, are more effectively modeled in NHPs compared to rodent models (2). Nonetheless, achieving a clinically translatable approach to suppressing alloreactive T memory cells while maintaining protective immunity remains a challenge that requires a deeper mechanistic understanding of T memory cells and the complex effects of new immunosuppressive regimens on preexisting immunity.
IMMUNE REGULATORY CELL THERAPY IN TRANSPLANTATION
When donor-strain regulatory dendritic cells (DCregs) were administered to transiently immunodepleted rats 1 week before heart transplantation, permanent donor-specific tolerance was achieved in 50% of animals (52). In NHPs, addition of donor-derived DCregs to a regimen of short-term costimulation blockade and tapered rapamycin treatment improved renal allograft survival compared to animals treated similarly but without DCregs, and no evidence of sensitization was detected (53). A NIAID-sponsored phase 1 clinical trial using DCregs in living-donor kidney transplantation is currently enrolling participants (ClinicalTrials.gov NCT03726307).
Adoptive transfer of Tregs in rodent models of organ and islet transplantation or hematopoietic stem cell transplantation can prevent rejection or GVHD, respectively. The use of humanized mouse hematopoietic stem cell transplantation models provided sufficient data to support translation of Treg therapy to hematopoietic stem cell transplantation clinical trials (54). However, NHP data are crucial for clinical translation to organ transplantation that involves early immunosuppression withdrawal. Renal allograft studies from Japan and Canada provided the first NHP data on adoptive transfer of alloantigen-specific, recipient anergic T cells or Tregs, respectively. Although the immunosuppressive regimens differed, both protocols included splenectomy and reported prolonged renal allograft survival compared to control NHPs that did not receive anergic cells or Tregs (55, 56). More recently, NHP data demonstrated proof of principle that, absent early reactivation of cytomegalovirus, polyclonally expanded recipient Tregs can prolong nonmyeloablative mixed chimerism and promote long-term tolerance to a kidney allograft transplanted 4 months after BMT (57).
Unlike these renal allograft studies, however, NHP heart transplant studies using polyclonally expanded autologous or third-party (nondonor allogeneic) Tregs in a protocol including ATG depletion, anti–IL-6R antibody, short-term tacrolimus, and tapering rapamycin treatment resulted in declining cardiac allograft function. Combined analysis of both Treg-infused groups compared to controls revealed elevated proinflammatory cytokines, T memory cells, and alloantibodies in treated recipients (58). This surprising result prevented untimely clinical application and underscores the need to optimize Treg products and protocols. In addition, we need a better understanding of Treg and T effector cell dynamics, particularly in the context of specific immunosuppressive regimens. As illustrated by several studies (59–61), the NHP model can be used to determine the influence of the immunosuppressive regimen and the phenotype, function, stability, timing, dosage, source, and antigen specificity of the Treg product on transplant outcomes. Numerous clinical trials of Treg therapy are in development or ongoing. Limited published data in human liver and renal transplant trials suggest that autologous Treg infusion is safe and not associated with increased rejection (62).
Encouraging results in rodent models of experimental autoimmune encephalomyelitis led transplant researchers to adopt the use of 1-ethyl-3(3-dimethylaminoproply)carbodiimide (ECDI)–treated apoptotic leukocytes for the prevention of allograft rejection. In mouse islet and cardiac allograft models, ECDI-treated apoptotic donor lymphocytes promoted the induction of tolerance through clonal T cell deletion and anergy or immunoregulation (63). In an NHP islet allograft model, addition of ECDI-treated apoptotic donor lymphocytes to a regimen of anti–IL-6R antibody, ATG, and rapamycin prolonged islet survival, associated with Treg expansion, but did not induce tolerance (64). Ongoing work with this NHP model may help to pave the way for clinical translation of apoptotic donor lymphocyte therapy in transplantation.
ALLOANTIBODY AS A BARRIER TO TRANSPLANTATION
Immunization to MHC antigens (HLA in humans) either before or after organ transplantation remains a formidable obstacle to long-term allograft function. The appearance of de novo donor-specific antibody to MHC antigens in human and NHP transplant recipients correlates with antibody-mediated rejection, chronic graft injury, and graft loss (65–67). Investigators use NHPs to model antibody-mediated rejection, to understand its mechanisms, and to test therapeutic approaches for preventing rejection in sensitized hosts. For example, proteasome inhibitors that deplete plasma cells (differentiated B cells producing antibodies) and thereby reduce alloantibody have been associated with germinal center activation and rebound of antibody production after cessation of therapy in NHPs (68), consistent with observations in humans (69, 70). However, the addition of costimulation blockade to proteasome inhibition in sensitized NHPs decreased bone marrow plasma cells and lymph node T follicular helper cells, inhibited B cell activation and proliferation, lowered alloantibody levels, and prolonged renal allograft rejection-free survival (71). This finding remains to be translated to humans. In contrast, in a nonsensitized NHP model of de novo alloantibody production, proteasome inhibition was not necessary as costimulation blockade (with either belatacept or anti-CD40 antibody) prevented development of alloantibody and rejection (15). Similarly, in the clinic, a belatacept-based regimen is associated with lower incidence of de novo alloantibody production (2 to 4%) compared to cyclosporine-based regimens (18%) (72).
SUPPRESSING INFLAMMATION TO PREVENT ALLOGRAFT INJURY
Preventing or reducing inflammation associated with ischemia-reperfusion injury, brain death, and the surgical procedure might limit rejection and improve organ viability (73). α1-Antitrypsin is a serine protease inhibitor that blunts expression of various proinflammatory cytokines and limits tissue damage caused by neutrophil elastase. Short-term, peritransplant treatment with α1-antitrypsin in monkeys undergoing autologous, marginal mass islet transplantation prevented islet loss and enabled long-term survival of islet grafts with improved rather than deteriorating graft function (74). These results contributed, in part, to the design of three ongoing clinical trials of α1-antitrypsin in islet transplantation (Clinicaltrials.gov NCT 02947087, NCT 02464878, and NCT 02713997).
IL-6 is a pleiotropic cytokine that amplifies the actions of proinflammatory mediators and facilitates the development of specific immunity. Blocking IL-6 in monkeys receiving orthotopic lung allografts was accompanied by a demonstrable increase in circulating Foxp3+ Tregs when administered with ATG (75). In a mixed-chimerism hematopoietic stem cell transplantation model, blocking IL-6 contributed to the first successful induction of long-term lung allograft tolerance in NHPs (30). On the basis of these NHP results and on human kidney transplant results with IL-6 blockade (76, 77), a NIAID-supported clinical trial testing the efficacy of IL-6 blockade to mitigate heart allograft rejection is in development.
RELEVANCE OF NHP TRANSPLANT STUDIES TO HUMANS
Over the past 40 years, NHP experiments have meaningfully informed immunosuppressive drug and regimen development for human transplantation and in many, but not all, cases have predicted efficacy, safety, and limitations of new therapies. New therapies introduced into human clinical transplantation practice after evaluation in NHP models include alemtuzumab, belatacept, and a regimen leading to transient mixed chimerism. With the support of encouraging NHP results, therapies such as anti-CD40 antibody, IL-6R blockade, and Treg/DCreg administration are currently in the planning stages for or are already in clinical trials. Other therapies, such as IL-15R blockade, showed promise in rodent studies but lacked efficacy in the NHP model and did not progress to transplant clinical trials (78). Compared to the rodent, similarities with respect to size, genetics, immunology, lifespan, and diverse immunological responses to their environment make the NHP a more relevant and predictive model for the development of transplant immunosuppression and tolerogenic therapies in humans.
Why NHP findings do not always predict human results is unclear, but may reflect seemingly minor differences in protocols, reagents, or interspecies immunobiology and pharmacokinetics (2). Advances in NHP MHC gene discovery and high-resolution MHC typing (79) improved viral immune management in immunosuppressed NHPs, and development of NHP-specific forms of immunotherapeutics has enhanced the scientific rigor and reproducibility of NHP transplant models. Determining preexisting, donor-specific alloreactive T memory cells in NHPs may enhance prediction and interpretation of outcomes. Moreover, similar determinations in human transplantation may allow for better modeling of clinical outcomes in NHPs (80). Recently developed methodologies like CYTOF (flow cytometry using antibodies labeled with metal ions and readout by time-of-flight mass spectrometry), plasma cell culture, RNA expression, and exosome analysis applied to biobanked NHP samples may allow retrospective interpretations of experimental outcomes and more discerning prospective study design. These advances, coupled with emerging technologies in imaging, cell tracking, and single-cell analytics, will enhance our understanding of immune-mediated rejection and tolerance and should serve to make NHP models more predictive of human responses after transplantation while reinforcing NIH mandates to reduce and refine experimental animal use. As technologies and NHP resources continue to improve, NHP models have the potential to address persistent knowledge gaps in our understanding of the mechanistic contributions of inflammation and innate, memory, and regulatory immune cell populations in organ- and cell-specific transplant tolerance. NIH support for NHP transplant tolerance research as a scientifically valid and ethical predecessor to clinical trials has contributed substantially to improvements in human health, and the benefits continue to accrue.
Acknowledgments:
We thank A. Kirk, B. Cosimi, L. Kean, D. Rotrosen, and N. D. Bridges for helpful suggestions. We also acknowledge non–NIH-funded investigators for their contributions to translational work in the NHP transplant model.
Footnotes
Competing interests: S.J.K. is a paid consultant to Novartis and VielaBio and owns stock in Bristol-Myers Squibb. B.J.H. has an equity interest in and serves as an executive officer of Diabetes-Free, an organization that may commercially benefit from the results of NHP research.
REFERENCES AND NOTES
- 1.Cosimi AB, Clinical application of tolerance induction in solid organ transplantation. Transplant. Proc 31, 1803–1805 (1999). [DOI] [PubMed] [Google Scholar]
- 2.Anderson DJ, Kirk AD, Primate models in organ transplantation. Cold Spring Harb. Perspect. Med 3, a015503 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kawai T, Cosimi AB, Spitzer TR, Tolkoff-Rubin N, Suthanthiran M, Saidman SL, Shaffer J, Preffer FI, Ding R, Sharma V, Fishman JA, Dey B, Ko DS, Hertl M, Goes NB, Wong W, Williams WW Jr., Colvin RB, Sykes M, Sachs DH, HLA-mismatched renal transplantation without maintenance immunosuppression. N. Engl. J. Med 358, 353–361 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Larsen CP, Elwood ET, Alexander DZ, Ritchie SC, Hendrix R, Tucker-Burden C, Cho HR, Aruffo A, Hollenbaugh D, Linsley PS, Winn KJ, Pearson TC, Long-term acceptance of skin and cardiac allografts after blocking CD40 and CD28 pathways. Nature 381,434–438 (1996). [DOI] [PubMed] [Google Scholar]
- 5.Preston EH, Xu H, Dhanireddy KK, Pearl JP, Leopardi FV, Starost MF, Hale DA, Kirk AD, IDEC-131 (anti-CD154), sirolimus and donor-specific transfusion facilitate operational tolerance in non-human primates. Am. J. Transplant 5, 1032–1041 (2005). [DOI] [PubMed] [Google Scholar]
- 6.Kirk AD, Harlan DM, Armstrong NN, Davis TA, Dong Y, Gray GS, Hong X, Thomas D, Fechner JH Jr., Knechtle SJ, CTLA4-Ig and anti-CD40 ligand prevent renal allograft rejection in primates. Proc. Natl. Acad. Sci. U.S.A 94, 8789–8794 (1997). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Larsen CP, Knechtle SJ, Adams A, Pearson T, Kirk AD, A new look at blockade of T-cell costimulation: A therapeutic strategy for long-term maintenance immunosuppression. Am. J. Transplant 6, 876–883 (2006). [DOI] [PubMed] [Google Scholar]
- 8.Larsen CP, Pearson TC, Adams AB, Tso P, Shirasugi N, Strobert E, Anderson D, Cowan S, Price K, Naemura J, Emswiler J, Greene J, Turk LA, Bajorath J, Townsend R, Hagerty D, Linsley PS, Peach RJ, Rational development of LEA29Y (belatacept), a high-affinity variant of CTLA4-Ig with potent immunosuppressive properties. Am. J. Transplant 5, 443–453 (2005). [DOI] [PubMed] [Google Scholar]
- 9.Adams AB, Shirasugi N, Durham MM, Strobert E, Anderson D, Rees P, Cowan S, Xu H, Blinder Y, Cheung M, Hollenbaugh D, Kenyon NS, Pearson TC, Larsen CP, Calcineurin inhibitor-free CD28 blockade-based protocol protects allogeneic islets in nonhuman primates. Diabetes 51, 265–270 (2002). [DOI] [PubMed] [Google Scholar]
- 10.Larsen CP, Grinyo J, Medina-Pestana J, Vanrenterghem Y, Vincenti F, Breshahan B, Campistol JM, Florman S, Rial Mdel C, Kamar N, Block A, Di Russo G, Lin CS, Garg P, Charpentier B, Belatacept-based regimens versus a cyclosporine A-based regimen in kidney transplant recipients: 2-year results from the BENEFIT and BENEFIT-EXT studies. Transplantation 90, 1528–1535 (2010). [DOI] [PubMed] [Google Scholar]
- 11.Posselt AM, Szot GL, Frassetto LA, Masharani U, Tavakol M, Amin R, McElroy J, Ramos MD, Kerlan RK, Fong L, Vincenti F, Bluestone JA, Stock PG, Islet transplantation in type 1 diabetic patients using calcineurin inhibitor-free immunosuppressive protocols based on T-cell adhesion or costimulation blockade. Transplantation 90, 1595–1601 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Moassesfar S, Masharani U, Frassetto LA, Szot GL, Tavakol M, Stock PG, Posselt AM, A comparative analysis of the safety, efficacy, and cost of islet versus pancreas transplantation in nonuremic patients with type 1 diabetes. Am. J. Transplant 16, 518–526 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kawai T, Andrews D, Colvin RB, Sachs DH, Cosimi AB, Thromboembolic complications after treatment with monoclonal antibody against CD40 ligand. Nat. Med 6, 114 (2000). [DOI] [PubMed] [Google Scholar]
- 14.Lowe M, Badell IR, Thompson P, Martin B, Leopardi F, Strobert E, Price AA, Abdulkerim HS, Wang R, Iwakoshi NN, Adams AB, Kirk AD, Larsen CP, Reimann KA, A novel monoclonal antibody to CD40 prolongs islet allograft survival. Am. J. Transplant 12, 2079–2087 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kim EJ, Kwun J, Gibby AC, Hong JJ, Farris AB III, Iwakoshi NN, Villinger F, Kirk AD, Knechtle SJ, Costimulation blockade alters germinal center responses and prevents antibody-mediated rejection. Am. J. Transplant 14, 59–69 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.O’Neill NA, Zhang T, Braileanu G, Sun W, Cheng X, Hershfeld A, Laird CT, Kronfli A, Hock LA, Dahi S, Kubicki N, Sievert E, Hassanein W, Cimeno A, Pierson RN III, Azimzadeh AM, Comparative Evaluation of αCD40 (2C10R4) and αCD154 (5C8H1 and IDEC-131) in a nonhuman primate cardiac allotransplant model. Transplantation 101,2038–2047 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Aoyagi T, Yamashita K, Suzuki T, Uno M, Goto R, Taniguchi M, Shimamura T, Takahashi N, Miura T, Okimura K, Itoh T, Shimizu A, Furukawa H, Todo S, A human anti-CD40 monoclonal antibody, 4D11, for kidney transplantation in cynomolgus monkeys: Induction and maintenance therapy. Am. J. Transplant 9, 1732–1741 (2009). [DOI] [PubMed] [Google Scholar]
- 18.Cordoba F, Wieczorek G, Audet M, Roth L, Schneider MA, Kunkler A, Stuber N, Erard M, Ceci M, Baumgartner R, Apolloni R, Cattini A, Robert G, Ristig D, Munz J, Haeberli L, Grau R, Sickert D, Heusser C, Espie P, Bruns C, Patel D, Rush JS, A novel, blocking, Fc-silent anti-CD40 monoclonal antibody prolongs nonhuman primate renal allograft survival in the absence of B cell depletion. Am. J. Transplant 15, 2825–2836 (2015). [DOI] [PubMed] [Google Scholar]
- 19.Sharabi Y, Sachs DH, Mixed chimerism and permanent specific transplantation tolerance induced by a nonlethal preparative regimen. J. Exp. Med 169, 493–502 (1989). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kawai T, Poncelet A, Sachs DH, Mauiyyedi S, Boskovic S, Wee SL, Ko DSC, Bartholomew A, Kimikawa M, Hong HZ, Abrahamian G, Colvin RB, Cosimi AB, Long-term outcome and alloantibody production in a non-myeloablative regimen for induction of renal allograft tolerance1. Transplantation 68, 1767–1775 (1999). [DOI] [PubMed] [Google Scholar]
- 21.Kawai T, Cosimi AB, Wee SL, Houser S, Andrews D, Sogawa H, Phelan J, Boskovic S, Nadazdin O, Abrahamian G, Colvin RB, Sach DH, Madsen JC, Effect of mixed hematopoietic chimerism on cardiac allograft survival in cynomolgus monkeys1. Transplantation 73, 1757–1764 (2002). [DOI] [PubMed] [Google Scholar]
- 22.Wekerle T, Sayegh MH, Hill J, Zhao Y, Chandraker A, Swenson KG, Zhao G, Sykes M, Extrathymic T cell deletion and allogeneic stem cell engraftment induced with costimulatory blockade is followed by central T cell tolerance. J. Exp. Med 187, 2037–2044 (1998). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kawai T, Sogawa H, Boskovic S, Abrahamian G, Smith RN, Wee SL, Andrews D, Nadazdin O, Koyama I, Sykes M, Winn HJ, Colvin RB, Sachs DH, Cosimi AB, CD154 blockade for induction of mixed chimerism and prolonged renal allograft survival in nonhuman primates. Am. J. Transplant 4, 1391–1398 (2004). [DOI] [PubMed] [Google Scholar]
- 24.Oura T, Ko DS, Boskovic S, O’Neil JJ, Chipashvili V, Koulmanda M, Hotta K, Kawai K, Nadazdin O, Smith RN, Cosimi AB, Kawai T, Kidney versus islet allograft survival after induction of mixed chimerism with combined donor bone marrow transplantation. Cell Transplant. 25, 1331–1341 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Pathiraja V, Villani V, Tasaki M, Matar AJ, Duran-Struuck R, Yamada R, Moran SG, Clayman ES, Hanekamp J, Shimizu A, Sachs DH, Huang CA, Yamada K, Tolerance of vascularized islet-kidney transplants in rhesus monkeys. Am. J. Transplant 17, 91–102 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Madariaga ML, Kreisel D, Madsen JC, Organ-specific differences in achieving tolerance. Curr. Opin. Organ Transplant 20, 392–399 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kawai T, Sachs DH, Sprangers B, Spitzer TR, Saidman SL, Zorn E, Tolkoff-Rubin N, Preffer F, Crisalli K, Gao B, Wong W, Morris H, LoCascio SA, Sayre P, Shonts B, Long-term results in recipients of combined HLA-mismatched kidney and bone marrow transplantation without maintenance immunosuppression. Am. J. Transplant 14, 1599–1611 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Umemura A, Morita H, Li XC, Tahan S, Monaco AP, Maki T, Dissociation of hemopoietic chimerism and allograft tolerance after allogeneic bone marrow transplantation. J. Immunol 167, 3043–3048 (2001). [DOI] [PubMed] [Google Scholar]
- 29.Yamada Y, Boskovic S, Aoyama A, Murakami T, Putheti P, Smith RN, Ochiai T, Nadazdin O, Koyama I, Boenisch O, Najafian N, Bhasin MK, Colvin RB, Madsen JC, Strom TB, Sachs DH, Benichou G, Cosimi AB, Kawai T, Overcoming memory T-cell responses for induction of delayed tolerance in nonhuman primates. Am. J. Transplant 12, 330–340 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tonsho M, Lee S, Aoyama A, Boskovic S, Nadazdin O, Capetta K, Smith RN, Colvin RB, Sachs DH, Cosimi AB, Kawai T, Madsen JC, Benichou G, Allan JS, Tolerance of lung allografts achieved in nonhuman primates via mixed hematopoietic chimerism. Am. J. Transplant 15, 2231–2239 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kean LS, Adams AB, Strobert E, Hendrix R, Gangappa S, Jones TR, Shirasugi N, Rigby MR, Hamby K, Jiang J, Bello H, Anderson D, Cardona K, Durham MM, Pearson TC, Larsen CP, Induction of chimerism in rhesus macaques through stem cell transplant and costimulation blockade-based immunosuppression. Am. J. Transplant 7, 320–335 (2007). [DOI] [PubMed] [Google Scholar]
- 32.Page A, Srinivasan S, Singh K, Russell M, Hamby K, Deane T, Sen S, Stempora L, Leopardi F, Price AA, Strobert E, Reimann KA, Kirk AD, Larsen CP, Kean LS, CD40 blockade combines with CTLA4Ig and sirolimus to produce mixed chimerism in an MHC-defined rhesus macaque transplant model. Am. J. Transplant 12, 115–125 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Koura DT, Horan JT, Langston AA, Qayed M, Mehta A, Khoury HJ, Harvey RD, Suessmuth Y, Couture C, Carr J, Grizzle A, Johnson HR, Cheeseman JA, Conger JA, Robertson J, Stempora L, Johnson BE, Garrett A, Kirk AD, Larsen CP, Waller EK, Kean LS, In vivo T cell costimulation blockade with abatacept for acute graft-versus-host disease prevention: A first-in-disease trial. Biol. Blood Marrow Transplant 19, 1638–1649 (2013). [DOI] [PubMed] [Google Scholar]
- 34.Knechtle SJ, Vargo D, Fechner J, Zhai Y, Wang J, Hanaway MJ, Scharff J, Hu H, Knapp L, Watkins D, Neville DM, Fn18-Crm9 immunotoxin promotes tolerance in primate renal allografts1. Transplantation 63, 1–6 (1997). [DOI] [PubMed] [Google Scholar]
- 35.Torrealba JR, Fernandez LA, Kanmaz T, Oberley TD, Schultz JM, Brunner KG, Peters D, Fechner JH Jr., Dong Y, Hu H, Hamawy MM, Knechtle SJ, Immunotoxin-treated rhesus monkeys: A model for renal allograft chronic rejection. Transplantation 76, 524–530 (2003). [DOI] [PubMed] [Google Scholar]
- 36.Kirk AD, Hale DA, Mannon RB, Kleiner DE, Hoffmann SC, Kampen RL, Cendales LK, Tadaki DK, Harlan DM, Swanson SJ, Results from a human renal allograft tolerance trial evaluating the humanized CD52-specific monoclonal antibody alemtuzumab (CAMPATH-1H). Transplantation 76, 120–129 (2003). [DOI] [PubMed] [Google Scholar]
- 37.Pearl JP, Parris J, Hale DA, Hoffmann SC, Bernstein WB, McCoy KL, Swanson SJ, Mannon RB, Roederer M, Kirk AD, Immunocompetent T-cells with a memory-like phenotype are the dominant cell type following antibody-mediated T-cell depletion. Am. J. Transplant 5, 465–474 (2005). [DOI] [PubMed] [Google Scholar]
- 38.van der Windt DJ, Smetanka C, Macedo C, He J, Lakomy R, Bottino R, Ekser B, Echeverri GJ, Metes D, Ijzermans JN, Trucco M, Cooper DK, Lakkis FG, Investigation of lymphocyte depletion and repopulation using alemtuzumab (Campath-1H) in cynomolgus monkeys. Am. J. Transplant 10, 773–783 (2010). [DOI] [PubMed] [Google Scholar]
- 39.Bloom D, Chang Z, Pauly K, Kwun J, Fechner J, Hayes C, Samaniego M, Knechtle S, BAFF is increased in renal transplant patients following treatment with alemtuzumab. Am. J. Transplant 9, 1835–1845 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kwun J, Page E, Hong JJ, Gibby A, Yoon J, Farris AB, Villinger F, Knechtle S, Neutralizing BAFF/APRIL with atacicept prevents early DSA formation and AMR development in T cell depletion induced nonhuman primate AMR model. Am. J. Transplant 15, 815–822 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kirk AD, Guasch A, Xu H, Cheeseman J, Mead SI, Ghali A, Mehta AK, Wu D, Gebel H, Bray R, Horan J, Kean LS, Larsen CP, Pearson TC, Renal transplantation using belatacept without maintenance steroids or calcineurin inhibitors. Am. J. Transplant 14, 1142–1151 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kelishadi SS, Azimzadeh AM, Zhang T, Stoddard T, Welty E, Avon C, Higuchi M, Laaris A, Cheng XF, McMahon C, Pierson III RN, Preemptive CD20+ B cell depletion attenuates cardiac allograft vasculopathy in cyclosporine-treated monkeys. J. Clin. Invest 120, 1275–1284 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chandraker A, Kobashigawa J, Stehlik J, Givertz M, Pierson R, Pinney S, Joren M, Nissen S, Guleria I, Morrison Y, Armstrong B, Bridges N, Sayegh M, Starling R, Rituximab induction in cardiac transplantation is associated with accelerated coronary artery vasculopathy: CTOT11. Am. J. Transplant 16 (suppl. 3) (2016). [Google Scholar]
- 44.Liu C, Noorchashm H, Sutter JA, Naji M, Prak EL, Boyer J, Green T, Rickels MR, Tomaszewski JE, Koeberlein B, Wang Z, Paessler ME, Velidedeoglu E, Rostami SY, Yu M, Barker CF, Naji A, B lymphocyte-directed immunotherapy promotes long-term islet allograft survival in nonhuman primates. Nat. Med 13, 1295–1298 (2007). [DOI] [PubMed] [Google Scholar]
- 45.Clatworthy MR, Watson CJ, Plotnek G, Bardsley V, Chaudhry AN, Bradley JA, Smith KG, B-cell-depleting induction therapy and acute cellular rejection. N. Engl. J. Med 360, 2683–2685 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Espinosa JR, Samy KP, Kirk AD, Memory T cells in organ transplantation: Progress and challenges. Nat. Rev. Nephrol 12, 339–347 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Badell IR, Russell MC, Thompson PW, Turner AP, Weaver TA, Robertson JM, Avila JG, Cano JA, Johnson BE, Song M, Leopardi FV, Swygert S, Strobert EA, Ford ML, Kirk AD, Larsen CP, LFA-1-specific therapy prolongs allograft survival in rhesus macaques. J. Clin. Invest 120, 4520–4531 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Posselt AM, Bellin MD, Tavakol M, Szot GL, Frassetto LA, Masharani U, Kerlan RK, Fong L, Vincenti FG, Hering BJ, Bluestone JA, Stock PG, Islet transplantation in type 1 diabetics using an immunosuppressive protocol based on the anti-LFA-1 antibody efalizumab. Am. J. Transplant 10, 1870–1880 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Anderson DJ, Lo DJ, Leopardi F, Song M, Turgeon NA, Strobert EA, Jenkins JB, Wang R, Reimann KA, Larsen CP, Kirk AD, Anti-leukocyte function-associated antigen 1 therapy in a nonhuman primate renal transplant model of costimulation blockade-resistant rejection. Am. J. Transplant 16, 1456–1464 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Carson KR, Focosi D, Major EO, Petrini M, Richey EA, West DP, Bennett CL, Monoclonal antibody-associated progressive multifocal leucoencephalopathy in patients treated with rituximab, natalizumab, and efalizumab: A review from the Research on Adverse Drug Events and Reports (RADAR) project. Lancet Oncol. 10, 816–824 (2009). [DOI] [PubMed] [Google Scholar]
- 51.Lowe MC, Badell IR, Turner AP, Thompson PW, Leopardi FV, Strobert EA, Larsen CP, Kirk AD, Belatacept and sirolimus prolong nonhuman primate islet allograft survival: Adverse consequences of concomitant alefacept therapy. Am. J. Transplant 13, 312–319 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.DePaz HA, Oluwole OO, Adeyeri AO, Witkowski P, Jin MX, Hardy MA, Oluwole SF, Immature rat myeloid dendritic cells generated in low-dose granulocyte macrophage-colony stimulating factor prolong donor-specific rat cardiac allograft survival. Transplantation 75, 521–528 (2003). [DOI] [PubMed] [Google Scholar]
- 53.Ezzelarab MB, Zahorchak AF, Lu L, Morelli AE, Chalasani G, Demetris AJ, Lakkis FG, Wijkstrom M, Murase N, Humar A, Shapiro R, Cooper DK, Thomson AW, Regulatory dendritic cell infusion prolongs kidney allograft survival in nonhuman primates. Am. J. Transplant 13, 1989–2005 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Wood KJ, Bushell A, Hester J, Regulatory immune cells in transplantation. Nat. Rev. Immunol 12, 417–430 (2012). [DOI] [PubMed] [Google Scholar]
- 55.Bashuda H, Kimikawa M, Seino K, Kato Y, Ono F, Shimizu A, Yagita H, Teraoka S, Okumura K, Renal allograft rejection is prevented by adoptive transfer of anergic T cells in nonhuman primates. J. Clin. Invest 115, 1896–1902 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ma A, Qi S, Song L, Hu Y, Dun H, Massicotte E, Dupuis M, Daloze P, Chen H, Adoptive transfer of CD4+CD25+ regulatory cells combined with low-dose sirolimus and anti-thymocyte globulin delays acute rejection of renal allografts in Cynomolgus monkeys. Int. Immunopharmacol 11 , 618–629 (2011). [DOI] [PubMed] [Google Scholar]
- 57.Duran-Struuck R, Sondermeijer HP, Buhler L, Alonso-Guallart P, Zitsman J, Kato Y, Wu A, McMurchy AN, Woodland D, Griesemer A, Martinez M, Boskovic S, Kawai T, Cosimi AB, Wuu CS, Slate A, Mapara M, Baker S, Tokarz R, D’Agati V, Hammer S, Pereira M, Lipkin WI, Wekerle T, Levings M, Sykes M, Effect of ex vivo-expanded recipient regulatory T cells on hematopoietic chimerism and kidney allograft tolerance across mhc barriers in cynomolgus macaques. Transplantation 101,274–283 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ezzelarab MB, Zhang H, Guo H, Lu L, Zahorchak AF, Wiseman RW, Nalesnik MA, Bhama JK, Cooper DK, Thomson AW, Regulatory t cell infusion can enhance memory t cell and alloantibody responses in lymphodepleted nonhuman primate heart allograft recipients. Am. J. Transplant 16, 1999–2015 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Singh K, Stempora L, Harvey RD, Kirk AD, Larsen CP, Blazar BR, Kean LS, Superiority of rapamycin over tacrolimus in preserving nonhuman primate Treg half-life and phenotype after adoptive transfer. Am. J. Transplant 14, 2691–2703 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zhang H, Guo H, Lu L, Zahorchak AF, Wiseman RW, Raimondi G, Cooper DK, Ezzelarab MB, Thomson AW, Sequential monitoring and stability of ex vivo-expanded autologous and nonautologous regulatory T cells following infusion in nonhuman primates. Am. J. Transplant 15, 1253–1266 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Anderson A, Martens CL, Hendrix R, Stempora LL, Miller WP, Hamby K, Russell M, Strobert E, Blazar BR, Pearson TC, Larsen CP, Kean LS, Expanded nonhuman primate tregs exhibit a unique gene expression signature and potently downregulate alloimmune responses. Am. J. Transplant 8, 2252–2264 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Tang Q, Vincenti F, Transplant trials with Tregs: Perils and promises. J. Clin. Invest 127, 2505–2512 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.McCarthy DP, Bryant J, Galvin JP, Miller SD, Luo X, Tempering allorecognition to induce transplant tolerance with chemically modified apoptotic donor cells. Am. J. Transplant 15, 1475–1483 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Lei J, Kim JI, Shi S, Zhang X, Machaidze Z, Lee S, Schuetz C, Martins PN, Oura T, Farkash EA, Rosales IA, Smith RN, Stott R, Lee KM, Soohoo J, Boskovic S, Cappetta K, Nadazdin OM, Yamada Y, Yeh H, Kawai T, Sachs DH, Benichou G, Markmann JF, Pilot study evaluating regulatory T cell-promoting immunosuppression and nonimmunogenic donor antigen delivery in a nonhuman primate islet allotransplantation model. Am. J. Transplant 15, 2739–2749 (2015). [DOI] [PubMed] [Google Scholar]
- 65.Smith RN, Kawai T, Boskovic S, Nadazdin O, Sachs DH, Cosimi AB, Colvin RB, Four stages and lack of stable accommodation in chronic alloantibody-mediated renal allograft rejection in Cynomolgus monkeys. Am. J. Transplant 8, 1662–1672 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Terasaki PI, Ozawa M, Predicting kidney graft failure by HLA antibodies: A prospective trial. Am. J. Transplant 4, 438–443 (2004). [DOI] [PubMed] [Google Scholar]
- 67.Terasaki PI, Cai J, Human leukocyte antigen antibodies and chronic rejection: From association to causation. Transplantation 86, 377–383 (2008). [DOI] [PubMed] [Google Scholar]
- 68.Kwun J, Burghuber C, Manook M, Iwakoshi N, Gibby A, Hong JJ, Knechtle S, Humoral compensation after bortezomib treatment of allosensitized recipients. J. Am. Soc. Nephrol 28, 1991–1996 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Woodle ES, Shields AR, Ejaz NS, Sadaka B, Girnita A, Walsh RC, Alloway RR, Brailey P, Cardi MA, Abu Jawdeh BG, Roy-Chaudhury P, Govil A, Mogilishetty G, Prospective iterative trial of proteasome inhibitor-based desensitization. Am. J. Transplant 15, 101–118 (2015). [DOI] [PubMed] [Google Scholar]
- 70.Moreno Gonzales MA, Gandhi MJ, Schinstock CA, Moore NA, Smith BH, Braaten NY, Stegall MD, 32 doses of bortezomib for desensitization is not well tolerated and is associated with only modest reductions in anti-HLA antibody. Transplantation 101, 1222–1227 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Kwun J, Burghuber C, Manook M, Ezekian B, Park J, Yoon J, Yi JS, Iwakoshi N, Gibby A, Hong JJ, Farris AB, Kirk AD, Knechtle SJ, Successful desensitization with proteasome inhibition and costimulation blockade in sensitized nonhuman primates. Blood Adv. 1,2115–2119 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Vincenti F, Charpentier B, Vanrenterghem Y, Rostaing L, Bresnahan B, Darji P, Massari P, Mondragon-Ramirez GA, Agarwal M, Di Russo G, Lin CS, Garg P, Larsen CP, A phase III study of belatacept-based immunosuppression regimens versus cyclosporine in renal transplant recipients (BENEFIT study). Am. J. Transplant 10, 535–546 (2010). [DOI] [PubMed] [Google Scholar]
- 73.Millington T, Koulmanda M, Ng C, Boskovic S, Nadazdin OM, Benichou G, Zheng XX, Strom TB, Madsen JC, Effects of an agonist interleukin-2/Fc fusion protein, a mutant antagonist interleukin-15/Fc fusion protein, and sirolimus on cardiac allograft survival in non-human primates. J. Heart Lung Transplant 31,427–435 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Koulmanda M, Sampathkumar RS, Bhasin M, Qipo A, Fan Z, Singh G, Movahedi B, Duggan M, Chipashvili V, Strom TB, Prevention of nonimmunologic loss of transplanted islets in monkeys. Am. J. Transplant 14, 1543–1551 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Aoyama A, Tonsho M, Ng CY, Lee S, Millington T, Nadazdin O, Wain JC, Cosimi AB, Sachs DH, Smith RN, Colvin RB, Kawai T, Madsen JC, Benichou G, Allan JS, Long-term lung transplantation in nonhuman primates. Am. J. Transplant 15, 1415–1420 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Vo AA, Choi J, Kim I, Louie S, Cisneros K, Kahwaji J, Toyoda M, Ge S, Haas M, Puliyanda D, Reinsmoen N, Peng A, Villicana R, Jordan SC, A phase I/II trial of the interleukin-6 receptor-specific humanized monoclonal (Tocilizumab) + intravenous immunoglobulin in difficult to desensitize patients. Transplantation 99, 2356–2363 (2015). [DOI] [PubMed] [Google Scholar]
- 77.Choi J, Aubert O, Vo A, Loupy A, Haas M, Puliyanda D, Kim I, Louie S, Kang A, Peng A, Kahwaji J, Reinsmoen N, Toyoda M, Jordan SC, Assessment of tocilizumab (anti-interleukin-6 receptor monoclonal) as a potential treatment for chronic antibody-mediated rejection and transplant glomerulopathy in HLA-sensitized renal allograft recipients. Am. J. Transplant 17, 2381–2389 (2017). [DOI] [PubMed] [Google Scholar]
- 78.Haustein S, Kwun J, Fechner J, Kayaoglu A, Faure JP, Roenneburg D, Torrealba J, Knechtle SJ, Interleukin-15 receptor blockade in non-human primate kidney transplantation. Transplantation 89, 937–944 (2010). [DOI] [PubMed] [Google Scholar]
- 79.Kean LS, Singh K, Blazar BR, Larsen CP, Nonhuman primate transplant models finally evolve: Detailed immunogenetic analysis creates new models and strengthens the old. Am. J. Transplant 12, 812–819 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Nadazdin O, Boskovic S, Murakami T, Tocco G, Smith RN, Colvin RB, Sachs DH, Allan J, Madsen JC, Kawai T, Cosimi AB, Benichou G, Host alloreactive memory T cells influence tolerance to kidney allografts in nonhuman primates. Sci. Transl. Med 3, 86ra51 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]