

Abstract
BACKGROUND: Severe acute respiratory syndrome coronavirus (SARS‐CoV) has been detected in the blood of infected individuals, which may have the potential to contaminate donated blood and plasma‐derived products in the event of a future outbreak. Effective methods for inactivating the SARS‐CoV in protein solutions are described in this report.
STUDY DESIGN AND METHODS: Heat, ultraviolet (UV) irradiation, octanoic acid, and solvent/detergent (S/D) methods were tested individually for their ability to inactivate SARS‐CoV in protein solutions appropriately mimicking blood‐derived products. Treated samples were tested for inactivation in a tissue culture growth assay.
RESULTS: Viral inactivation by heat treatment at 60°C required 15 to 30 minutes to inactivate the SARS‐CoV. UVC efficiently inactivated SARS‐CoV in 40 minutes, whereas UVA required the addition of psoralen to enhance inactivation of the virus. The presence of bovine serum albumin limited the ability of UVC and UVA to inactivate SARS‐CoV and octanoic acid treatment does not reduce the infectivity of SARS‐CoV–spiked protein solutions. S/D treatment required 2, 4, and up to 24 hours for Triton X‐100, Tween 80, and sodium cholate inactivation, respectively.
CONCLUSION: Heat, UVC irradiation, and S/D treatments effectively inactivate SARS‐CoV, whereas octanoic acid treatment is insufficient for inactivation of the virus.
ABBREVIATIONS
- CPE
cytopathic effect
- HSV‐1
herpes simplex virus‐1
- RSV
respiratory syncytial virus
- SARS‐CoV
severe acute respiratory syndrome coronavirus
- TCID50
50 percent tissue culture infectious dose assay
- TNBP
tri‐(n‐butyl) phosphate.
Coronaviruses (CoV) are the agents of respiratory and enteric disease in a variety of animals. The life‐threatening severe acute respiratory syndrome (SARS) is caused by the recently described coronavirus, SARS‐CoV. 1 , 2 , 3 According to the World Health Organization, 8098 people were diagnosed with SARS and 774 people died of this disease during the initial outbreak in 2002 to 2003.
SARS‐CoV was detected by quantitative reverse transcriptase‐polymerase chain reaction (RT‐PCR) assays in serum from infected patients. 4 Although most, if not all, viremic individuals would be expected to be symptomatic, there is a chance that some fraction of individuals early in infection is asymptomatic and could transmit the virus if they donated blood.
It has been shown that heat or ultraviolet (UV) irradiation of the SARS‐CoV in culture medium eliminates viral infectivity. 5 We previously determined that acidic and basic conditions, and formalin and glutaraldehyde, inactivate SARS‐CoV in culture medium. 6 Many of the viral inactivation conditions established for SARS‐CoV in culture medium, however, are not appropriate for human blood and blood products. Viral inactivation methods used for blood products need to be safe for injection and must not compromise the stability of the proteins.
MATERIALS AND METHODS
Virus and cells
The Urbani strain of SARS‐CoV was grown in African green monkey kidney cells (Vero E6) as described previously. 6 Briefly, Vero E6 cells SARS‐CoV (Urbani strain, a gift of L.J. Anderson and T.G. Ksiazek, Centers for Disease Control and Prevention, Atlanta, GA) at 106.33 50 percent tissue culture infectious dose assay (TCID50) per mL in Dulbecco’s modified Eagle’s medium (DMEM; Biosource International, Rockville, MD). After 1 hour at 25°C, DMEM with supplements (10% fetal bovine serum, 2 mmol/L/mL l‐glutamine, 100 U/mL penicillin, 100 µg/mL streptomycin, and 0.5 µg/mL fungizone; Biosource International) was added to the flasks, and the infected cells were incubated at 37°C for 3 days. Supernatants were collected, pooled, clarified by centrifugation, and stored at −70°C for use as the viral stock. The viral stock was filtered through a 0.22‐µm filter unit (Millex‐GS, Millipore Corp., Bedford, MA) and diluted 10‐fold in bovine serum albumin (BSA; Sigma‐Aldrich Inc., St. Louis, MO) solutions, phosphate‐buffered saline (PBS; Biosource International), or DMEM without supplements for inactivation. Respiratory syncytial virus (RSV) strain A2 was provided by J.A. Beeler (CBER, FDA). Herpes simplex virus‐1 (HSV‐1) strain 17+ was provided by A.S. Bertke of the FDA. Personnel wore powered air‐purifying respirators (3M, St. Paul, MN) and performed all work with infectious virus inside a biosafety cabinet, within a Biosafety Level 3 containment facility.
Quantitation of viral titers
Viral titers were determined by observing infected Vero E6 monolayers in 24‐ and 96‐well plates with a TCID50. Tenfold serial dilutions of viral samples were incubated at 37°C for 4 days, and then examined for cytopathic effect (CPE) in infected cells as described by Ksiazek and coworkers. 2 Briefly, CPE of SARS‐CoV–infected Vero E6 cells was determined by observing rounded, detached cells in close association to each other. An absence of CPE in the Vero E6 cells indicated a loss of infectivity and was evidence of inactivation of the SARS‐CoV samples. As the first dilution of viral sample inoculated onto the Vero cells was a 1‐in‐10 dilution, the limit of viral detection for this assay was 1 log TCID50 per mL. If the amount of viral particles was below the limit of detection, then a theoretical, yet low content of viruses may be considered.
Heat treatment of virus
Virus was diluted in BSA‐PBS solutions or PBS, and samples were incubated in a water bath at 56, 60, or 65°C and removed at time points up to 120 minutes. After heat treatment, samples were immediately frozen at −70°C for further analysis by TCID50 assay with CPE as an endpoint.
UV light treatment
UV light treatment was performed with SARS‐CoV diluted in BSA‐PBS, PBS, or DMEM. Psoralen (Sigma‐Aldrich Inc.) was dissolved in 100 percent ethanol to make a stock of 15 mmol per L. SARS‐CoV was incubated in a final concentration of 15 µmol per L psoralen for 30 minutes at room temperature and kept in the dark before exposure to UV. Virus samples were placed in 24‐well plates (Corning Inc., Corning, NY) and the volume depth at the center of the well was 1 cm. The UV light source (Spectronics Corporation, Westbury, NY) was placed above the plate such that the distance from the light to the bottom of the wells containing the virus samples was 3 cm. At a distance of 3 cm, our UVC light source (254 nm) emitted 4016 µW per cm2 (where µW = 10−6 J/sec) and the UVA light source (365 nm) emitted 2133 µW per cm2, as measured by radiometric analysis (Spectronics Corporation). After exposure to UV light, virus samples were immediately frozen at −70°C for further analysis by TCID50 assay once all of the sampling was completed.
Octanoic acid treatment
SARS‐CoV was diluted in PBS or BSA‐PBS. Some of the BSA solutions (pH 6.9) were adjusted with hydrochloric acid (Sigma‐Aldrich Inc.) to achieve a pH value of 4.5 or 5.5. RSV and HSV‐1 were diluted in DMEM that was similarly adjusted to pH 4.5. Sodium octanoate (stock concentration of 200 mmol/L or 2 mol/L) was added to the samples to achieve a final concentration of 20 or 40 mmol per L. Samples were treated at room temperature or in a 30°C water bath. After a timed incubation, SARS‐CoV samples were immediately frozen at −70°C for further analysis by TCID50 assay with CPE as the endpoint. RSV and HSV‐1 samples were immediately analyzed by TCID50 assay.
Solvent/detergent treatment
Virus was diluted in BSA‐PBS protein solutions or PBS. Solvent/detergent (S/D) solutions (20× stock) were added to each sample of virus in BSA‐PBS or in PBS to achieve a final concentration of nonvolatile solvent (0.3% v/v), specifically, tri‐(n‐butyl) phosphate (TNBP; Aeros Organics, Pittsburgh, PA) and either 1 percent Tween 80 (Sigma‐Aldrich Inc.), 1 percent Triton X‐100 (Sigma‐Aldrich Inc.), or 0.2 percent sodium cholate (Sigma‐Aldrich Inc.). Before the addition of the S/D solution and at the time points 2, 4, 6, and 24 hours after the addition of the S/D solution, an aliquot was removed from each of the samples and diluted 10‐fold (Tween 80 and sodium cholate) or 100‐fold (Triton X‐100) in DMEM. This dilution was necessary to stop the inactivation reaction and to negate the cytotoxic effects of the S/D on the Vero cells during the titration analysis. 7 The dilution step was considered during the final calculation. Immediately after this dilution in DMEM, the aliquots were frozen at −70°C and then analyzed by TCID50 assay with CPE as an endpoint.
Statistical analysis
Data are presented as the geometric mean of three or more samples ± standard deviation.
RESULTS
To evaluate the efficacy of treatments used to inactivate viruses in human blood products, we examined SARS‐CoV survival under current protocols. To validate these techniques, BSA protein solutions were spiked with SARS‐CoV, whereas PBS or DMEM were spiked with the virus and used as a control. After treatment to inactivate SARS‐CoV, these virus‐spiked solutions were tested for infectivity in a TCID50 assay with a limit of viral detection of not greater than 1 log TCID50 per mL.
Heat inactivation of SARS‐CoV
Heat may inactivate viruses by denaturing the secondary structures of proteins and thereby may alter the conformation of virion proteins involved in attachment and replication within a host cell. 8 , 9 To test the ability of heat to inactivate SARS‐CoV, we incubated normal human serum spiked with virus at two temperatures (56 and 65°C) for increasing periods of time. After 20 minutes at 56°C, SARS‐CoV was inactivated to the level of the detection limit for the assay (Fig. 1A). Human serum treated at 65°C required only 10 minutes to achieve viral inactivation to the detection limit of the assay (Fig. 1A).
Figure 1.
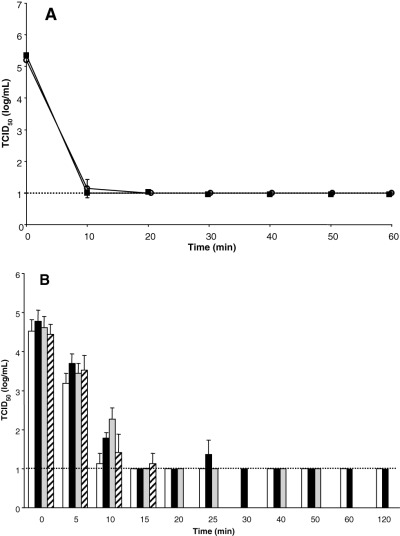
Inactivation of SARS‐CoV by heat treatment of (A ) human serum and (B ) BSA solutions. SARS‐CoV was diluted 1:10 in human serum, PBS, or BSA solutions. The virus/diluent mixtures were incubated in a water bath at (A) 56°C (○) and 65°C (▪) and (B) 60°C, and samples were removed at designated time points. Samples were frozen and analyzed simultaneously by TCID50 assay. The broken line shows the limit of detection for the assay. (□) PBS; (▪) 10 percent BSA; ( ) 16 percent BSA; ( ) 25 percent BSA.
To test the ability of heat to inactivate SARS‐CoV in solutions with higher protein concentrations, we incubated BSA solutions or PBS spiked with virus at 60°C for increasing periods of time. After 15 minutes at 60°C, inactivation of the virus in the PBS, 10 percent BSA, and 16 percent BSA solutions was observed (Fig. 1B). Although the virus in 10 percent BSA appeared to be inactivated in 15 minutes, there was a small, yet detectable amount of virus present in one of the triplicate 25‐minute samples (Fig. 1B). This outlier may have arisen due to experimental error or the virus may have aggregated thus making it more resistant to inactivation. By 30 minutes, SARS‐CoV was inactivated in the 10 percent BSA solution and was no longer detectable for the duration of the experiment (120 min). Fifteen minutes at 60°C significantly decreased, to near the limit of detection of the assay, the titer of SARS‐CoV in the 25 percent BSA protein solution (Fig. 1B). Unfortunately, at 20 minutes, the 25 percent BSA solutions were denatured and rendered them inappropriate for virus titer testing. These results suggest that SARS‐CoV inactivation by pasteurization may be effective; however, the rate of inactivation may be dependent upon protein concentration.
Effect of UV radiation on SARS‐CoV
The continuum of UV light is divided into three classifications: UVC (200‐280 nm), UVB (280‐320 nm), and UVA (320‐400 nm). UVC and UVB light causes pyrimidine dimers, 10 whereas UVA may cause RNA and DNA strand breaks. 11 The effect of UV irradiation on SARS‐CoV infectivity was examined by spiking solutions of BSA or DMEM with SARS‐CoV in 24‐well tissue culture plates on ice. The samples were exposed to UV light for varying lengths of time and tested for infectivity. As expected from our previous results, 6 UVC treatment significantly decreased the virus titer after 20 minutes and inactivated SARS‐CoV in PBS to the limit of detection by 40 minutes (Fig. 2A). UVC treatment, however, did not inactivate the virus in BSA protein solutions, and the virus was still infectious even after 60 minutes of treatment (Fig. 2A). Additionally, lower concentrations of protein resulted in some inactivation after exposure to UVC. We determined that some inactivation of SARS‐CoV occurred in 2 and 5 percent BSA protein solutions but to a lesser extent than when no protein was present (data not shown). This suggests that there is a concentration‐dependent protective effect of BSA not present in PBS alone.
Figure 2.
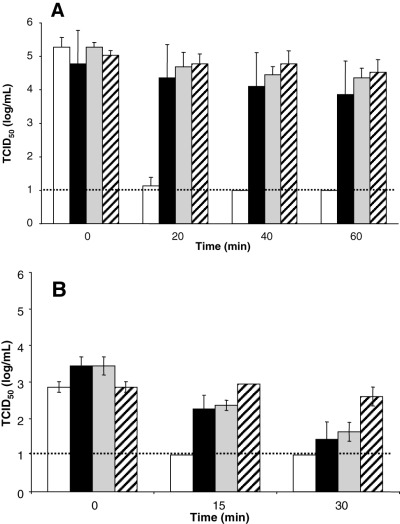
Inactivation of SARS‐CoV by ultraviolet light. (A ) Virus was incubated in PBS, with and without BSA, and exposed to UVC light for the indicated times. (□) PBS; (▪) 10 percent BSA; ( ) 16 percent BSA; ( ) 25 percent BSA. (B ) Virus was mixed into DMEM, with and without 10 percent BSA. (□) DMEM, psoralen; (▪) 10 percent BSA, psoralen; ( ) DMEM, no psoralen; ( ) 10 BSA, no psoralen. Psoralen was added to half of the viral samples. All viral samples were analyzed for TCID50 after exposure to UVA. The broken line shows the limit of detection for the assay.
UVA treatment to inactivate viruses in platelet (PLT) concentrates is combined with psoralen compounds to increase the effectiveness of the method. 12 We determined that UVA treatment of SARS‐CoV in DMEM decreased the infectivity by 100‐fold after 30 minutes, whereas the addition of psoralen inactivated the virus to the limit of detection after only 15 minutes (Fig. 2B). Although UVA treatment of SARS‐CoV in 10 percent BSA resulted in no inactivation after 30 minutes, the addition of psoralen brought about a 100‐fold decrease in infectivity during the same amount of time, yet did not result in complete inactivation of the virus (Fig. 2B). These data suggest that the addition of psoralen increases the effectiveness of UVA treatment regardless of the presence of BSA protein, and the presence of BSA protein limits the effectiveness of UVA inactivation of SARS‐CoV.
Effect of octanoic acid on SARS‐CoV
Octanoic acid is commonly used to inactivate lipid‐enveloped viruses during the production of human blood products. 13 To determine the effect of octanoic acid on SARS‐CoV, virus‐spiked BSA solutions were incubated with octanoic acid for 1 hour at pH 6.9 and at room temperature. Octanoic acid had no apparent effect on the infectivity of SARS‐CoV (Fig. 3A). We also determined that 1 hour incubation of SARS‐CoV in BSA solutions with octanoic acid at pH 4.5 or 5.5 did not inactivate the virus (data not shown). Changing the conditions by lengthening the time of incubation to 24 hours, increasing the concentration of octanoic acid from 20 to 40 mmol per L, and increasing the temperature to 30°C, while maintaining the acidic pH of 4.5, also did not inactivate SARS‐CoV (data not shown). When RSV was exposed to 40 mmol per L octanoic acid at 30°C with a pH value of 4.5, however, the titer was significantly decreased after 24 hours when compared to untreated virus (Fig. 3B). The octanoic acid treatment of HSV‐1 for 24 hours did not reduce the titer nearly as effectively as it did for RSV (Fig. 3B). Although this method is effective for the inactivation of many lipid enveloped viruses, such as RSV and to a lesser extent, HSV‐1, it does not appear to inactivate SARS‐CoV under any of the conditions described.
Figure 3.
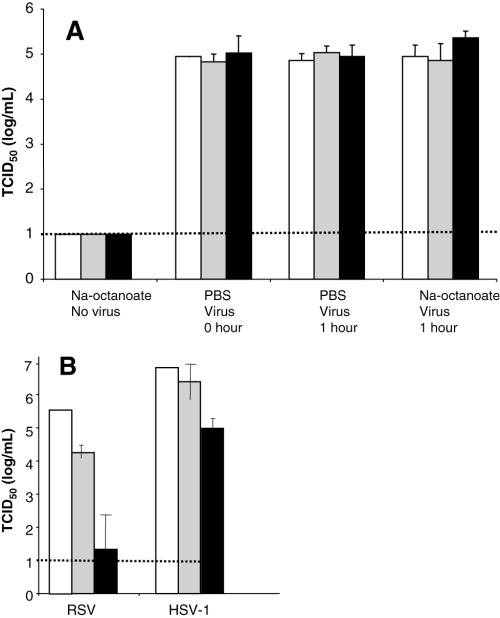
Octanoic acid treatment of virus. (A ) SARS‐CoV was diluted into 10 percent (□), 16 percent ( ), and 25 percent (▪) BSA solutions with octanoic acid (20 mmol/L) or PBS. (B ) Octanoic acid (40 mmol/L) was added to virus stocks (RSV or HSV‐1) in DMEM and then analyzed for infectivity by TCID50 assay. The broken line shows the limit of detection for the assay. (□) Time 0; ( ) 1 hour; (▪) 24 hours.
S/D treatment of human serum spiked with SARS‐CoV
S/D treatment is a standard method used to inactivate viruses in human blood products. 7 , 14 SARS‐CoV–spiked solutions of BSA and PBS were treated with S/D, incubated at room temperature, and sampled periodically over 24 hours. The SARS‐CoV–spiked samples treated with TNBP/Triton X‐100 were inactivated to the limit of detection by 2 hours, regardless of whether the virus was in PBS or in various BSA solutions (Fig. 4A). TNBP/Tween 80 treatments efficiently inactivated SARS‐CoV in PBS and in 10 percent BSA within 2 hours; however, 4 hours were required to inactivate the virus to the limit of detection in 16 and 25 percent BSA protein solutions (Fig. 4B). SARS‐CoV in PBS was inactivated by treatment with TNBP/sodium cholate after 2 hours; however, 24 hours were required to inactivate the virus to the limit of detection in 10 percent BSA (Fig. 4C). The virus titer in 16 and 25 percent BSA protein solutions was only decreased slightly after TNBP/sodium cholate treatment and was still detectable after 24 hours. These results suggest that the effectiveness of SARS‐CoV inactivation by S/D treatment is variable, depending upon the specific method used, duration of treatment, and the protein content of the solution.
Figure 4.
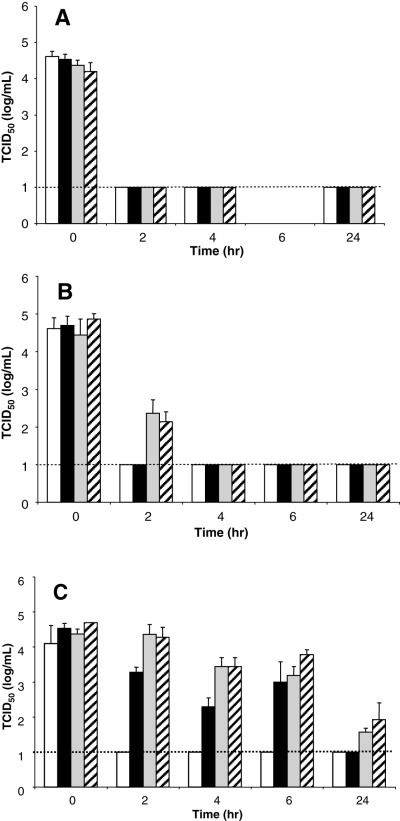
S/D treatment of SARS‐CoV. Virus was diluted in PBS or BSA solutions. The S/D concentrate (20×) was added and samples were removed at designated time points and frozen immediately. The detergents used were (A ) Triton X‐100, (B ) Tween 80, and (C ) sodium cholate. The broken line shows the limit of detection for the TCID50 assay. (□) PBS; (▪) 10 percent BSA; ( ) 16 percent BSA; ( ) 25 percent BSA.
DISCUSSION
Transmission of SARS‐CoV occurs predominantly by respiratory droplets over a relatively close distance. 15 Although blood transfusions and administration of blood products have not been implicated in SARS‐CoV transmission, 16 low to moderate levels of viremia during the clinical phase of the illness have been shown. 1 Nie and associates 17 have shown that neutralizing antibodies to SARS‐CoV can be detected as early as 5 to 10 days after onset of symptoms. With an immunofluorescent antibody assay, Hsueh and associates 18 have detected immunoglobulin (Ig)G seroconversion by Day 10 and IgM and IgA seroconversion on Day 11. Wang and coworkers 19 determined that SARS‐CoV RNA was detectable by quantitative real‐time RT‐PCR assay as early as 2 days after onset of SARS symptoms in 50 percent of the patient plasma samples they examined. In 75 percent of SARS patients they found peak viral RNA titers detected at 5 to 7 days after symptom onset. These data do not exclude the possibility that (in the event of an outbreak) SARS may be present in the blood of asymptomatic but infected individuals at undetectable levels. Owing to the possibility for SARS‐CoV transmission via plasma‐derived components, current viral inactivation methods need to be evaluated for effectiveness against SARS‐CoV.
Pasteurization at 60°C for 10 hours is used to inactivate potential viruses in plasma‐derived products. 20 An advantage of this technique is that it is effective against both enveloped and nonenveloped viruses. 20 Stabilizers to protect the protein product, however, may also protect or mask the virus. Neoantigens may be formed from partial denaturation of human serum components that may appear foreign to the immune system. 20 This could be deleterious, because it may result in the onset of autoimmune disease. We determined that SARS‐CoV was inactivated by pasteurization at 60°C in less than 1 hour; however, protein concentration affected the time required for inactivation. Previously we examined the ability of heat to inactivate SARS‐CoV when in DMEM instead of human serum or BSA solutions. We found that 90 minutes at either 56 or 65°C was necessary to inactivate the virus. 6 Yunoki and associates 21 examined the sensitivity of SARS‐CoV to 60°C heat and found that the different protein compositions of the samples tested affected the time necessary, yet all samples were inactivated to the limit of detection by 1 hour. Additionally, Rabenau and coworkers 22 tested heat inactivation of SARS‐CoV at 56°C for 30 minutes and determined that the presence of 20 percent fetal calf serum protected the virus. We observed that incubation at 60°C for 30 minutes, however, was sufficient for complete inactivation to the limit of detection. Given that pasteurization of plasma‐derived products is usually performed at 60°C for 10 hours, 20 any SARS‐CoV present in such products, regardless of protein concentrations and in the presence of stabilizers, should be efficiently inactivated.
Both UVC and UVA, in conjunction with psoralen, can be used to inactivate pathogenic viruses in plasma‐derived products. Previously we determined that UVA light alone did not inactivate SARS‐CoV in DMEM after 20 minutes. 6 Lin and coworkers 12 added psoralen S‐59 to PLT concentrates and exposed them to UVA light. The authors determined that 3 J per cm2 UVA light for 3 to 4 minutes with the addition of 150 µmol per L psoralen was sufficient to inactivate human immunodeficiency virus, duck hepatitis B virus (a model for hepatitis B virus), and bovine viral diarrhea virus (a model for hepatitis C virus) and that neither psoralen, nor UVA light alone, was sufficient to inactivate these viruses. We determined that this also holds true for SARS. The addition of intercalating agents, such as psoralen, may be required to increase the efficiency of UVA viral inactivation and may be appropriate for treatment of PLTs or red cells.
We determined that UVC had no inactivating effect on the virus in 10 to 25 percent BSA solutions. Previously, however, we inactivated SARS‐CoV in DMEM 6 suggesting that some characteristic of the BSA protein protected the virus from inactivation. It is possible that BSA interfered with the light waves and prevented them from reaching the virus in the solution. Chin and colleagues 23 inactivated several virus types in plasma and Factor VIII concentrate by circulating the samples through a flat quartz cell that was 0.22 mm thick. Li and coworkers 24 described that UVC light was decreased by 105 at 1 mmol per L depth in a 20 percent (w/v) albumin solution. These data suggest that there is some factor in the BSA protein solutions, which has a protective effect on SARS‐CoV, and this effect might be overcome by exposing a very thin layer of the SARS‐CoV solution to the UVC light.
Octanoic acid, or caprylate, is a treatment used to inactivate enveloped viruses in human blood products. The mechanism of action is due to the lipophilic nature of the nonionized form of the acid, which allows it to insert itself into the viral membrane and disrupt the lipid envelope and/or the proteins imbedded in the envelope. 25 Johnston and colleagues 26 have shown that caprylate treatment is effective against Sindbis virus and bovine viral diarrhea virus. We determined that SARS‐CoV was resistant to inactivation by octanoic acid treatment, even under the conditions of acidic pH (4.5), elevated temperature (30°C), a higher concentration (40 mmol/L) of octanoate, and a longer treatment time (24 hr). Based on the work of Lundblad and Seng, 25 we exposed both RSV and HSV‐1 to octanoate as a control. Octanoic acid treatment decreased the titer of RSV, suggesting that our conditions were correct for inactivation. In contrast, Lundblad and Seng 25 effectively inactivated HSV‐1 with octanoic acid at similar conditions, but the octanoic acid was removed with an ion‐exchange column, and this may account for differences in the end result. Under the conditions tested here, octanoic acid treatment was ineffective for inactivation of SARS‐CoV.
S/D methods are not used for inactivation of viruses in cellular blood components because S/D treatment damages cell membranes. 27 The S/D method causes a disruption of the structural integrity of lipid‐enveloped viruses. 20 The S/D treatment has the advantage of being highly efficient at inactivating enveloped viruses, while being nondenaturing on proteins, and yielding a high recovery of protein activity. 20 The disadvantages include requiring an extra manufacturing step to remove the S/D agents and the inability to inactivate nonenveloped viruses. 20 In our experiments, both Triton X‐100 and Tween 80 inactivated SARS‐CoV in BSA solutions. Sodium cholate was ineffective in high protein concentrations even after 24 hours of treatment. There may be a protective effect by some of the proteins in the BSA solutions, as the SARS‐CoV in the higher concentrations of BSA was not inactivated as quickly by Tween 80 or sodium cholate as it was with Triton X‐100. Horowitz and associates 28 have shown that the S/D method can very effectively inactivate cytomegalovirus, Sindbis virus, and other pathogenic viruses. The Tween 80 and Triton X‐100 methods should be sufficient to inactivate SARS‐CoV in plasma for the manufacture of plasma‐derived products.
Human plasma–derived components potentially contain transfusion‐transmissible viruses. To minimize the risk of infection, the manufacturing process includes two or more inactivation steps. Several methods show promise and should minimize the potential risk of transmission of SARS‐CoV from human plasma–derived components.
ACKNOWLEDGMENTS
We gratefully acknowledge Larry Anderson MD and Thomas Ksiazek DVM, PhD for the gift of SARS coronavirus, Mei‐Ying Yu PhD and Edward Tabor, MD for helpful discussions, and Kathryn Carbone MD and Jesse Goodman MD for institutional support. RSV was provided by Judy Beeler, MD CBER/FDA, and HSV‐1 was provided by Andrea Bertke, CBER/FDA. We thank Ewan Plant PhD and Chintamani D. Atreya PhD for critical reading of the manuscript. We acknowledge the Office of the Assistant Secretary for Public Health & Emergency Preparedness, Department of Health and Human Services, for financial support of this project.
REFERENCES
- 1. Drosten C, Gunther S, Preiser W, et al. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N Engl J Med 2003;348:1967‐76. [DOI] [PubMed] [Google Scholar]
- 2. Ksiazek TG, Erdman D, Goldsmith CS, et al. A novel coronavirus associated with severe acute respiratory syndrome. N Engl J Med 2003;348:1953‐66. [DOI] [PubMed] [Google Scholar]
- 3. Rota PA, Oberste MS, Monroe SS, et al. Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science 2003;300:1394‐9. [DOI] [PubMed] [Google Scholar]
- 4. Hung IF, Cheng VC, Wu AK, et al. Viral loads in clinical specimens and SARS manifestations. Emerg Infect Dis 2004;9:1550‐7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Duan SM, Zhao XS, Wen RF, et al. Stability of SARS coronavirus in human specimens and environment and its sensitivity to heating and UV irradiation. Biomed Environ Sci 2003;16:246‐55. [PubMed] [Google Scholar]
- 6. Darnell ME, Subbarao K, Feinstone SM, Taylor DR. Inactivation of the coronavirus that induces severe acute respiratory syndrome, SARS‐CoV. J Virol Methods 2004; 121:85‐91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Roberts PL, Dunkerley C. Effect of manufacturing process parameters on virus inactivation by solvent‐detergent treatment in a high‐purity factor IX concentrate. Vox Sang 2003;84:170‐5. [DOI] [PubMed] [Google Scholar]
- 8. Lelie PN, Reesink HW, Lucas CJ. Inactivation of 12 viruses by heating steps applied during manufacture of a hepatitis B vaccine. J Med Virol 1987;23:297‐301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Schlegel A, Immelmann A, Kempf C. Virus inactivation of plasma‐derived proteins by pasteurization in the presence of guanidine hydrochloride. Transfusion 2001; 41:382‐9. [DOI] [PubMed] [Google Scholar]
- 10. Perdiz D, Grof P, Mezzina M, et al. Distribution and repair of bipyrimidine photoproducts in solar UV‐irradiated mammalian cells. J Biol Chem 2000;275:26732‐42. [DOI] [PubMed] [Google Scholar]
- 11. Ravanat JL, Douki T, Cadet J. Direct and indirect effects of UV radiation on DNA and its components. J Photochem Photobiol B 2001;63:88‐102. [DOI] [PubMed] [Google Scholar]
- 12. Lin L, Cook DN, Wiesehahn GP, et al. Photochemical inactivation of viruses and bacteria in platelet concentrates by use of a novel psoralen and long‐wavelength ultraviolet light. Transfusion 1997;37:423‐35. [DOI] [PubMed] [Google Scholar]
- 13. Dichtelmuller H, Rudnick D, Kloft M. Inactivation of lipid enveloped viruses by octanoic acid treatment of immunoglobulin solution. Biologicals 2002;30:135‐42. [DOI] [PubMed] [Google Scholar]
- 14. Hellstern P, Haubelt H. Manufacture and composition of fresh frozen plasma and virus‐inactivated therapeutic plasma preparations: correlation between composition and therapeutic efficacy. Thromb Res 2002;107:S3‐S8. [DOI] [PubMed] [Google Scholar]
- 15. Dwosh HA, Hong HH, Austgarden D, et al. Identification and containment of an outbreak of SARS in a community hospital. CMAJ 2003;168:1415‐20. [PMC free article] [PubMed] [Google Scholar]
- 16. Berger A, Drosten C, Doerr HW, et al. Severe acute respiratory syndrome (SARS)—paradigm of an emerging viral infection. J Clin Virol 2004;29:13‐22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Nie Y, Wang G, Shi X, et al. Neutralizing antibodies in patients with severe acute respiratory syndrome‐associated coronavirus infection. J Infect Dis 2004;190:1119‐26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Hsueh PR, Huang LM, Chen PJ, et al. Chronological evolution of IgM, IgA, IgG and neutralisation antibodies after infection with SARS‐associated coronavirus. Clin Microbiol Infect 2004;10:1062‐6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Wang WK, Fang CT, Chen HL, et al. Detection of severe acute respiratory syndrome coronavirus RNA in plasma during the course of infection. J Clin Microbiol 2005;43:962‐5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Burnouf T, Radosevich M. Reducing the risk of infection from plasma products: specific preventative strategies. Blood Rev 2000;14:94‐110. [DOI] [PubMed] [Google Scholar]
- 21. Yunoki M, Urayama T, Yamamoto I, et al. Heat sensitivity of a SARS‐associated coronavirus introduced into plasma products. Vox Sang 2004;87:302‐3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Rabenau HF, Cinatl J, Morgenstern B, et al. Stability and inactivation of SARS coronavirus. Med Microbiol Immunol 2005;194:1‐6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Chin S, Williams B, Gottlieb P, et al. Virucidal short wavelength ultraviolet light treatment of plasma and factor VIII concentrate: protection of proteins by antioxidants. Blood 1995;86:4331‐6. [PubMed] [Google Scholar]
- 24. Li Q, MacDonald S, Bienek C, et al. Design of a UV‐C irradiation process for the inactivation of viruses in protein solutions. Biologicals 2005;33:101‐10. [DOI] [PubMed] [Google Scholar]
- 25. Lundblad JL, Seng RL. Inactivation of lipid‐enveloped viruses in proteins by caprylate. Vox Sang 1991;60:75‐81. [DOI] [PubMed] [Google Scholar]
- 26. Johnston A, Uren E, Johnstone D, Wu J. Low pH, caprylate incubation as a second viral inactivation step in the manufacture of albumin. Parametric and validation studies. Biologicals 2003;31:213‐21. [DOI] [PubMed] [Google Scholar]
- 27. Klein HG. Pathogen inactivation technology: cleansing the blood supply. J Int Med 2005;257:224‐37. [DOI] [PubMed] [Google Scholar]
- 28. Horowitz B, Prince AM, Horowitz MS, Watklevicz C. Viral safety of solvent‐detergent treated blood products. Dev Biol Stand 1993;81:147‐61. [PubMed] [Google Scholar]