

This genetic association study evaluates common single-nucleotide variations associated with spontaneous coronary artery dissection susceptibility.
Key Points
Question
Are there common genetic variations associated with spontaneous coronary artery dissection?
Findings
In this genome-wide association study of approximately 5 million genotyped and imputed single-nucleotide variants, 5 genetic risk loci associated with spontaneous coronary artery dissection were identified and replicated.
Meaning
This study identified positional candidate genes associated with spontaneous coronary artery dissection that have established associations with extracoronary arteriopathies.
Abstract
Importance
Spontaneous coronary artery dissection (SCAD), an idiopathic disorder that predominantly affects young to middle-aged women, has emerged as an important cause of acute coronary syndrome, myocardial infarction, and sudden cardiac death.
Objective
To identify common single-nucleotide variants (SNVs) associated with SCAD susceptibility.
Design, Setting, and Participants
This single-center genome-wide association study examined approximately 5 million genotyped and imputed SNVs and subsequent SNV-targeted replication analysis results in individuals enrolled in the Mayo Clinic SCAD registry from August 30, 2011, to August 2, 2018. Data analysis was performed from June 21, 2017, to December 30, 2019.
Main Outcomes and Measures
Genetic loci and positional candidate genes associated with SCAD.
Results
This study included 484 white women with SCAD (mean [SD] age, 46.6 [9.2] years) and 1477 white female controls in the discovery cohort (mean [SD] age, 64.0 [14.5] years) and 183 white women with SCAD (mean [SD] age, 47.1 [9.9] years) and 340 white female controls in the replication cohort (mean [SD] age, 51.0 [15.3] years). Associations with SCAD risk reached genome-wide significance at 3 loci (1q21.3 [OR, 1.78; 95% CI, 1.51-2.09; P = 2.63 × 10−12], 6p24.1 [OR, 1.77; 95% CI, 1.51-2.09; P = 7.09 × 10−12], and 12q13.3 [OR, 1.67; 95% CI, 1.42-1.97; P = 3.62 × 10−10]), and 7 loci had evidence suggestive of an association (1q24.2 [OR, 2.10; 95% CI, 1.58-2.79; P = 2.88 × 10−7], 3q22.3 [OR, 1.47; 95% CI, 1.26-1.71; P = 6.65 × 10−7], 4q34.3 [OR, 1.84; 95% CI, 1.44-2.35; P = 9.80 × 10−7], 8q24.3 [OR, 2.57; 95% CI, 1.76-3.75; P = 9.65 × 10−7], 15q21.1 [OR, 1.75; 95% CI, 1.40-2.18; P = 7.23 × 10−7], 16q24.1 [OR, 1.91; 95% CI, 1.49-2.44; P = 2.56 × 10−7], and 21q22.11 [OR, 2.11; 95% CI, 1.59-2.82; P = 3.12 × 10−7]) after adjusting for the top 5 principal components. Associations were validated for 5 of the 10 risk alleles in the replication cohort. In a meta-analysis of the discovery and replication cohorts, associations for the 5 SNVs were significant, with relatively large effect sizes (1q21.3 [OR, 1.77; 95% CI, 1.54-2.03; P = 3.26 × 10−16], 6p24.1 [OR, 1.71; 95% CI, 1.49-1.97; P = 4.59 × 10−14], 12q13.3 [OR, 1.69; 95% CI, 1.47-1.94; P = 1.42 × 10−13], 15q21.1 [OR, 1.79; 95% CI, 1.48-2.17; P = 2.12 × 10−9], and 21q22.11 [OR, 2.18; 95% CI, 1.70-2.81; P = 1.09 × 10−9]). Each index SNV was within or near a gene highly expressed in arterial tissue and previously linked to SCAD (PHACTR1) and/or other vascular disorders (LRP1, LINC00310, and FBN1).
Conclusions and Relevance
This study revealed 5 replicated risk loci and positional candidate genes for SCAD, most of which are associated with extracoronary arteriopathies. Moreover, the alternate alleles of 3 SNVs have been previously associated with atherosclerotic coronary artery disease, further implicating allelic susceptibility to coronary artery atherosclerosis vs dissection.
Introduction
Spontaneous coronary artery dissection (SCAD) is an increasingly recognized cause of acute coronary syndrome (ACS), myocardial infarction (MI), and sudden cardiac death in otherwise healthy, predominantly young to middle-aged individuals in whom atherosclerotic risk factors are uncommon.1 SCAD is characterized by disruption of the coronary artery wall by an intramural hematoma within the medial layer and/or an intimal tear, both of which can impair blood flow. SCAD is more common in women (>80% in most series) and may be the cause of ACS in as many as 35% of women 50 years or younger.1 The pathogenesis of SCAD is unknown, although it is thought to be multifactorial with environmental, hormonal, and genetic risk factors. Vascular imaging reveals noncoronary fibromuscular dysplasia (FMD) in 25% to 86% of patients, indicating that SCAD is the clinical manifestation of a systemic arteriopathy.1 SCAD occurs as a sporadic disorder in most individuals, yet rare familial cases composed of 2 or more affected relatives implicate a hereditary predisposition.2 Genetic factors associated with SCAD susceptibility may include rare sequence variants with major effect size and common risk-associated variants. Recent next-generation sequencing and targeted genotyping studies3,4 have identified missense variants in TLN1 (OMIM 186745) in association with familial and sporadic SCAD and a noncoding risk allele in PHACTR1 (OMIM 608723) in cohorts with SCAD. We performed a genome-wide association study (GWAS) to gain further insight into the genetic architecture of SCAD.
Methods
In this GWAS, participants for the discovery and replication cohorts were consecutively enrolled in the Mayo Clinic SCAD registry after diagnostic confirmation of SCAD by review of coronary angiograms and abstraction of demographic and clinical data from questionnaires and medical records from August 30, 2011, to August 2, 20181 (Table 1). Study participants provided written informed consent for genetic research under a protocol approved by the Mayo Clinic Institutional Review Board. All authors were approved for full access to clinical and genetic data under the institutional review board–approved study protocol, and no data were shared with investigators or uploaded into databases external to Mayo Clinic.
Table 1. Clinical Characteristics of the Study Participants.
| Characteristic | Participantsa | P value | |
|---|---|---|---|
| Discovery (n = 484) | Replication (n = 183) | ||
| Age at first event, mean (SD), y | 46.6 (9.2) | 47.1 (9.9) | .49 |
| BMI, mean (SD) | 26.0 (5.9) | 27.1 (6.8) | .05 |
| Coronary artery distribution | |||
| LM | 5 (1.0) | 3 (1.6) | .69 |
| LAD | 225 (46.5) | 71 (38.8) | .08 |
| RCA | 32 (6.6) | 11 (6.0) | .86 |
| LC | 32 (6.6) | 9 (4.9) | .48 |
| Multivessel | 87 (18.0) | 37 (20.2) | .51 |
| Other territoryb | 103 (21.3) | 52 (28.4) | .06 |
| Imaged for FMD | 315 (65.1) | 139 (76.0) | .01 |
| Diagnosis | |||
| FMD | 175 (55.6) | 91 (65.5) | .05 |
| Extracoronary vascular abnormality | 25 (5.2) | 9 (4.9) | >.99 |
| Peripartum status | 55 (11.4) | 20 (10.9) | >.99 |
| Before SCAD | |||
| Extreme exertion | 75 (15.5) | 31 (16.9) | .64 |
| Extreme emotion | 88 (18.2) | 45 (24.6) | .08 |
| Chronic hypertension | 157 (32.4) | 53 (29.0) | .40 |
| Hyperlipidemia | 168 (34.7) | 46 (25.1) | .02 |
| Diabetes | 14 (2.9) | 4 (2.2) | .79 |
| Tobacco use | |||
| Active | 12 (2.5) | 3 (1.6) | .77 |
| Prior | 129 (26.7) | 53 (29.0) | .56 |
| Migraine headaches | 175 (36.2) | 77 (42.1) | .18 |
| No. of recurrences | |||
| 0 | 402 (83.1) | 151 (82.5) | .91 |
| 1 | 64 (13.2) | 28 (15.3) | .53 |
| 2 | 17 (3.5) | 4 (2.2) | .47 |
| 3 | 1 (0.2) | 0 | >.99 |
Abbreviations: BMI, body mass index (calculated as weight in kilograms divided by height in meters squared); FMD, fibromuscular dysplasia; LAD, left anterior descending; LC, left circumflex; LM, left main; RCA, right coronary artery; SCAD, spontaneous coronary artery dissection.
Data are presented as number (percentage) of participants unless otherwise indicated.
Diagonal artery, left and right posterior lateral artery, left and right posterior descending artery, obtuse marginal artery, or ramus intermedius artery.
As previously described, criteria for diagnosis of SCAD were the presence of a noniatrogenic dissection plane and the absence of coronary atherosclerosis.5 Whole blood or saliva samples were obtained for genomic DNA extraction. The discovery cohort consisted of 484 white women with SCAD. Individuals diagnosed with a connective tissue disorder and/or aortopathy were excluded. Control individuals were from the Mayo Genome Consortia, Center for Individualized Medicine.6 These controls consisted of 1477 white women without known arteriopathy, excluding individuals with diagnostic codes for atherosclerotic coronary artery disease (CAD), acute MI, FMD, arterial aneurysm or dissection, cerebral infarction, Marfan syndrome, and Ehlers-Danlos syndrome. Associated single-nucleotide variants (SNVs) were tested for replication in an independent cohort of 183 white women with SCAD. The replication cohort was recruited after the discovery analysis. Replication controls were from the Mayo Clinic Biobank, Center for Individualized Medicine.7 These controls consisted of 340 white women who met the arteriopathy exclusion criteria.
Statistical Analysis
Demographic and clinical data from individuals comprising the discovery and replication cohorts were compared using the 2-tailed t test or 2-tailed Fisher exact test. Genome-wide association analyses of SNVs and SCAD risk were evaluated using logistic regression models assuming additive allele effects. To control for population stratification, the first 5 principal components were used as covariates in the models. Strength of association was estimated by calculating the odds ratio (OR) and corresponding 95% CI. An SNV with a 2-sided P < 5 × 10−8 was considered to achieve genome-wide statistical significance, and P < 1 × 10−6 was considered to achieve nominal significance. Association results from the discovery and replication studies were combined in a fixed-effects meta-analysis model using PLINK.8 Data analysis was performed from June 21, 2017, to December 30, 2019. Details for genetic and statistical analyses and candidate gene prioritization are described in the eMethods in the Supplement.
Results
SCAD Risk Loci
This study included 484 white women with SCAD (mean [SD] age, 46.6 [9.2] years) and 1477 white female controls in the discovery cohort (mean [SD] age, 64.0 [14.5] years) and 183 white women with SCAD (mean [SD] age, 47.1 [9.9] years) and 340 white female controls in the replication cohort (mean [SD] age, 51.0 [15.3] years). The patients who comprised the discovery and replication cohorts shared similar clinical characteristics (Table 1). To detect chromosomal loci associated with SCAD susceptibility, a GWAS was performed after systematic quality control analysis, harmonization, and imputation of genotyping data. Approximately 5 million genotyped or imputed SNVs spanning the 22 autosomes and X chromosome were assessed in the 484 SCAD samples and in the 1477 control samples. The genomic inflation factor was λ = 1.045, implying a low possibility of false-positive associations attributable to population stratification (eFigure 1 in the Supplement). Associations with genome-wide significance were identified for noncoding SNVs at 3 loci (1q21.3 [OR, 1.78; 95% CI, 1.51-2.09; P = 2.63 × 10−12], 6p24.1 [OR, 1.77; 95% CI, 1.51-2.09; P = 7.09 × 10−12], and 12q13.3 [OR, 1.67; 95% CI, 1.42-1.97; P = 3.62 × 10−10]) after adjusting for the top 5 principal components (Figure 1 and Table 2). Noncoding SNVs at 7 additional loci (1q24.2 [OR, 2.10; 95% CI, 1.58-2.79; P = 2.88 × 10−7], 3q22.3 [OR, 1.47; 95% CI, 1.26-1.71; P = 6.65 × 10−7], 4q34.3 [OR, 1.84; 95% CI, 1.44-2.35; P = 9.80 × 10−7], 8q24.3 [OR, 2.57; 95% CI, 1.76-3.75; P = 9.65 × 10−7], 15q21.1 [OR, 1.75; 95% CI, 1.40-2.18; P = 7.23 × 10−7], 16q24.1 [OR, 1.91; 95% CI, 1.49-2.44; P = 2.56 × 10−7], and 21q22.11 [OR, 2.11; 95% CI, 1.59-2.82; P = 3.12 × 10−7]) had evidence suggestive of an association (Figure 1 and Table 2). The associations for all 10 SNVs remained statistically significant after false discovery rate testing (q values: 3.63 × 10−7 for 1q21.3, 8.73 × 10−7 for 6p24.1, 3.26 × 10−5 for 12q13.3, 5.21 × 10−3 for 16q24.1, 5.43 × 10−3 for 1q24.2, 5.75 × 10−3 for 21q22.11, 8.98 × 10−3 for 3q22.3, 9.71 × 10−3 for 15q21.1, 0.01 for 8q24.3, and 0.01 for 4q34.3).
Figure 1. Manhattan Plot of the Discovery Genome-Wide Association Analysis.

The x-axis designates chromosomal position (23 designates the X chromosome), and the y-axis designates the P value derived by logistic regression on a –log10 scale. The horizontal orange and blue lines indicate the threshold for genome-wide significance (P < 5 × 10−8) and suggestive associations (P < 1 × 10−06), respectively. Identification numbers indicate index SNPs included in the independent replication study, with red font designating those that replicated.
Table 2. Association Results for Index Markers.
| SNV | Locus | Risk allele | Discovery | Replication | Meta-analysis | Candidate genes | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RAF in cases/controls | OR (95% CI) | P value | q Value | RAF cases/controls | OR (95% CI) | P value | OR (95% CI) | P value | ||||
| rs4970935 (I) | 1q21.3 | C | 0.37/0.25 | 1.78 (1.51-2.09) | 2.63 × 10−12 | 3.63 × 10−7 | 0.40/0.26 | 1.74 (1.34-2.26) | 3.37 × 10−5 | 1.77 (1.54-2.03) | 3.26 × 10−16 | ECM1,a,c C1orf54,b,d ADAMTSL4,a,c and MRPS21b,c |
| rs9349379 (G) | 6p24.1 | A | 0.73/0.59 | 1.77 (1.51-2.09) | 7.09 × 10−12 | 8.73 × 10−7 | 0.70/0.60 | 1.57 (1.20-2.06) | 9.90 × 10−4 | 1.71 (1.49-1.97) | 4.59 × 10−14 | PHACTR1a,c,d |
| rs11172113 (G) | 12q13.3 | T | 0.71/0.59 | 1.67 (1.42-1.97) | 3.62 × 10−10 | 3.26 × 10−5 | 0.71/0.58 | 1.75 (1.33-2.31) | 6.82 × 10−5 | 1.69 (1.47-1.94) | 1.42 × 10−13 | LRP1a,c,d |
| rs67049921 (I) | 16q24.1 | G | 0.90/0.83 | 1.91 (1.49-2.44) | 2.56 × 10−7 | 5.21 × 10−3 | 0.84/0.84 | 0.96 (0.69-1.35) | 0.83 | 1.51 (1.23-1.84) | 5.40 × 10−5 | NA |
| rs6700122 (I) | 1q24.2 | T | 0.11/0.06 | 2.10 (1.58-2.79) | 2.88 × 10−7 | 5.43 × 10−3 | 0.11/0.09 | 1.25 (0.83-1.88) | 0.28 | 1.77 (1.41-2.24) | 1.40 × 10−6 | NA |
| rs28451064 (I) | 21q22.11 | G | 0.93/0.86 | 2.11 (1.59-2.82) | 3.12 × 10−7 | 5.75 × 10−3 | 0.95/0.88 | 2.44 (1.45-4.09) | 7.61 × 10−4 | 2.18 (1.70-2.81) | 1.09 × 10−9 | LINC00310a,d |
| rs189056 (I) | 3q22.3 | A | 0.45/0.36 | 1.47 (1.26-1.71) | 6.65 × 10−7 | 8.98 × 10−3 | 0.43/0.41 | 1.06 (0.82-1.37) | 0.65 | 1.35 (1.18-1.54) | 7.53 × 10−6 | NA |
| rs2015637 (I) | 15q21.1 | C | 0.16/0.10 | 1.75 (1.40-2.18) | 7.23 × 10−7 | 9.71 × 10−3 | 0.18/0.10 | 1.91 (1.31-2.78) | 7.02 × 10−4 | 1.79 (1.48-2.17) | 2.12 × 10−9 | FBN1a,d |
| rs10095937 (I) | 8q24.3 | G | 0.06/0.02 | 2.57 (1.76-3.75) | 9.65 × 10−7 | 0.01 | 0.04/0.04 | 0.87 (0.46-1.68) | 0.69 | 1.96 (1.41-2.71) | 5.74 × 10−5 | NA |
| rs79603310 (I) | 4q34.3 | C | 0.14/0.08 | 1.84 (1.44-2.35) | 9.8 × 10−7 | 0.01 | 0.12/0.13 | 0.94 (0.64-1.38) | 0.74 | 1.52 (1.24-1.87) | 7.32 × 10−5 | NA |
Abbreviations: G, genotyped; I, imputed; OR, odds ratio; RAF, risk allele frequency; SNV, single-nucleotide variation.
Gene nearest to the index SNV.
Gene overlapping SNV within strong linkage disequilibrium (r2>0.8) of index SNV.
Expression quantitative trait locus target gene for the index SNV.
Gene ranked in the top 5 for coronary artery, aorta, or tibial artery expression.
To test for replication of associations of SCAD and the 10 SNVs with the strongest association signal at each locus, targeted genotyping was conducted in independent cohorts composed of 183 SCAD samples and 340 control samples. Risk alleles of 5 SNVs within the 1q21.3 (OR, 1.74; 95% CI, 1.34-2.26; P = 3.37 × 10−5), 6p24.1 (OR, 1.57; 95% CI, 1.20-2.06; P = 9.90 × 10−4), 12q13.3 (OR, 1.75; 95% CI, 1.33-2.31; P = 6.82 × 10−5), 15q21.1 (OR, 1.91; 95% CI, 1.31-2.78; P = 7.02 × 10−4), and 21q22.11 (OR, 2.44; 95% CI, 1.45-4.09; P = 7.61 × 10−4) loci demonstrated statistically significant associations in the replication study (Figure 1 and Table 2). Associations between these 5 SNVs and SCAD were statistically significant in meta-analysis that combined discovery and replication results (1q21.3 [OR, 1.77; 95% CI, 1.54-2.03; P = 3.26 × 10−16], 6p24.1 [OR, 1.71; 95% CI, 1.49-1.97; P = 4.59 × 10−14], 12q13.3 [OR, 1.69; 95% CI, 1.47-1.94; P = 1.42 × 10−13], 21q22.11 [OR, 2.18; 95% CI, 1.70-2.81; P = 1.09 × 10−9], and 15q21.1 [OR, 1.79; 95% CI, 1.48-2.17; P = 2.12 × 10−9]) (Table 2).
Positional Candidate Genes
Seventy-four candidate genes within the 5 replicated SCAD-susceptibility loci, each located within 400 kb of the 5 index SNVs, were identified and prioritized (Figure 2). Supporting evidence for a gene candidate included (1) location within or nearest an index SNV or SNV within high linkage disequilibrium (r2>0.8) of the index SNV, (2) high arterial tissue expression ranking, and (3) a cis-acting expression quantitative trait locus (eQTL) target gene for the index SNV. Eight plausible candidate genes from the 5 SCAD risk loci were prioritized based on fulfilling 2 or more criteria (Table 2 and eTable 1 and eFigure 2 in the Supplement).
Figure 2. Regional LocusZoom Plots Demonstrating Positional Candidate Genes.
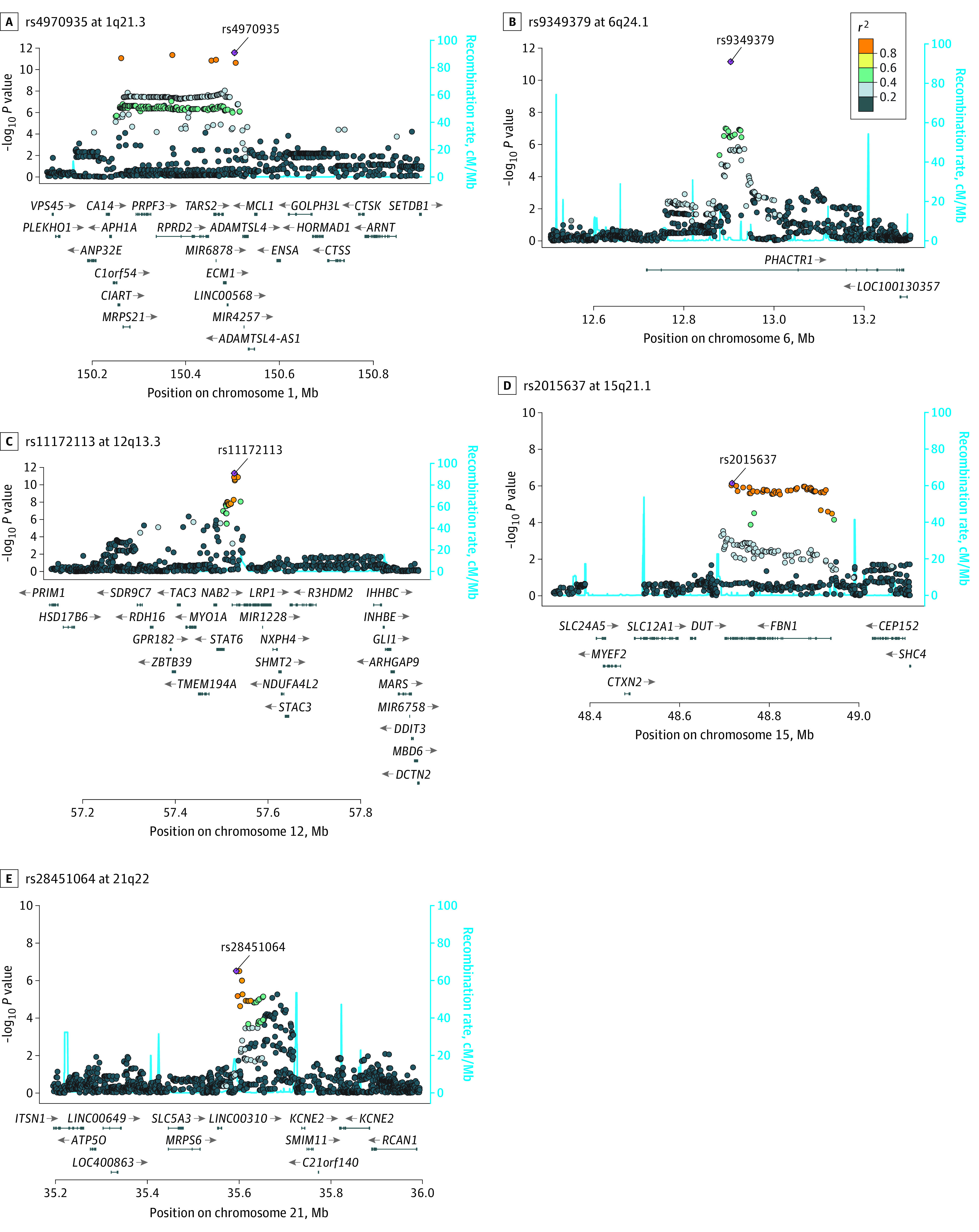
LocusZoom plots for rs4970935 at 1q21.3 (A), rs9349379 at 6p24.1 (B), rs11172113 at 12q13.3 (C), rs2015637 at 15q21.1 (D), and rs28451064 at 21q22 (E). The x-axis indicates chromosomal position and the y-axis the association significance (–log10 [P value]) for the 400 kilobase flanking the index SNP. Linkage disequilibrium estimates (r2) are color-coded. Recombination hotspots are indicated by blue lines.
1q21.3 Locus
At the 1q21.3 locus, 118 SNVs spanning a 250-kb region were significantly associated with SCAD risk. The strongest signal occurred at rs4970935, and risk association was confirmed in the replication study. The rs4970935-C allele had a higher prevalence among SCAD samples in the discovery and replication studies. The index SNV was intergenic, equidistant from ECM1 (OMIM 602210) and ADAMTSL4 (OMIM 610113), with a total of 25 genes located within the 400-kb flanking regions. Four plausible candidate genes were prioritized, each fulfilling 2 of the filtering criteria. A significant eQTL signal was identified for rs4970935 on ECM1 in the tibial artery and ADAMTSL4 in the aorta and tibial artery, with the SCAD risk allele associated with decreased expression. The C1orf54 (HGNC 26258) and MRPS21 (OMIM 611984) genes were nearest to an SNV (rs12740679), with genome-wide significance and high linkage disequilibrium with the index SNV. According to the Genotype Tissue Expression project, C1orf54 ranks highly in arterial tissue expression.9 In addition, a significant eQTL signal was identified for rs4970935 on MRPS21 in the aorta and tibial artery, with decreased expression associated with the risk allele. The index SNV overlapped a high-confidence enhancer region, with ADAMTSL4 ranked highest for gene-enhancer associations (eTable 2 in the Supplement).
6p24.1 Locus
At the 6p24.1 locus, rs9349379 was significantly associated with SCAD risk, and risk association was confirmed in the replication study. The rs9349379-A allele had a higher prevalence among SCAD samples in the discovery and replication studies. The index SNV was located within the third intron of PHACTR1 (OMIM 608723), the sole positional candidate at this locus. Significant eQTL signals were identified for rs9349379 on PHACTR1 in the aorta, tibial artery, and coronary artery, with the SCAD risk allele associated with increased expression. In addition, PHACTR1 ranked highly in tibial artery expression.
12q13.3 Locus
At the 12q13.3 locus, 10 SNVs spanning an 83-kb region were significantly associated with SCAD risk. The strongest signal occurred at rs11172113, and risk association was confirmed in the replication study. The rs11172113-T allele had a higher prevalence among SCAD samples in the discovery and replication studies. The index SNV was within the first intron of LRP1 (OMIM 107770), with a total of 27 genes located within the 400-kb flanking regions. LRP1 was the only gene to fulfill 2 of the filtering criteria. Significant eQTL signals were identified for rs11172113 on LRP1 in the aorta, coronary artery, and tibial artery, with increased expression associated with the SCAD risk allele. In addition, LRP1 ranked highly in aortic tissue expression. The SNV overlapped a high-confidence enhancer region, with LRP1 ranked highest for gene-enhancer associations (eTable 2 in the Supplement).
15q21.1 Locus
At the 15q21.1 locus, 2 SNVs had evidence suggestive of an association with SCAD risk. The strongest signal occurred at rs2015637, and risk association was confirmed in the replication study. The rs2015637-C allele had a higher prevalence among SCAD samples in the discovery and replication studies. The index SNV was located within intron 60 of FBN1 (OMIM 134797), with a total of 10 genes located within the 400-kb flanking regions. FBN1 was the only gene to fulfill 2 of the filtering criteria based on its proximity to the index SNV and ranking for aorta and coronary artery expression. The SNV overlapped a high-confidence enhancer region, with FBN1 ranked highest for gene-enhancer associations (eTable 2 in the Supplement).
21q22.11 Locus
At the 21q22.11 locus, 3 SNVs had evidence suggestive of an association with SCAD risk. The strongest signal occurred at rs28451064, and risk association was confirmed in the replication study. The rs28451064-G allele had a higher prevalence among SCAD samples in the discovery and replication studies. The index SNV was intergenic between LINC00310 (HGNC 16414) and KCNE2 (OMIM 603796), with a total of 10 genes located within the 400-kb flanking regions. LINC00310, nearest the index SNV and highly expressed in all 3 arterial tissues, was the only gene to fulfill 2 of the filtering criteria.
Potential Associations Between TLN1 and GWAS Candidate Genes
In a previous study,3 TLN1 was identified as a gene associated with familial and sporadic SCAD owing to rare missense variants. Talin 1 has multiple integrin- and actin-binding domains, reflecting its role as a major structural component that links the actin cytoskeleton to the extracellular matrix via integrins in coronary arteries. Accordingly, potential functional associations among talin 1, integrin and actin isoforms, and positional candidate genes identified by the GWAS were investigated by STRING software, version 11.10 Among the 8 prioritized GWAS candidate genes, 3 had high-confidence functional associations with integrin or actin genes that interact directly with TLN1 (Figure 3 and eTable 3 in the Supplement).
Figure 3. Network of Known and Predicted Functional Associations.

Talin 1 (red) interacts with 3 of the genome-wide association study candidate gene-encoding proteins (blue) via actin or integrin isoforms (white). STRING analysis was set at a high confidence interaction score (0.7).
Discussion
In this study of cohorts composed of 667 women with SCAD and 1817 women without SCAD, we identified replicated risk loci associated with SCAD, an enigmatic disorder that causes ACS, MI, and sudden death in an otherwise healthy, predominantly female population. Significant associations between 5 common variants and SCAD were identified with relatively large effect sizes compared with CAD GWASs, potentially because SCAD may be a less heterogeneous phenotype (eTable 4 in the Supplement). These findings reveal risk loci and positional candidate genes for which there is existing knowledge of associations with vascular biologic and pathophysiologic findings.
At the 1q21.3 locus, 4 genes had positional and expression evidence that supported their association with SCAD susceptibility. MRPS21 encodes a protein that is part of the small (28S) mammalian mitochondrial ribosomal subunit involved in the synthesis of polypeptides essential for oxidative phosphorylation.11,12 ECM1 encodes a secretory glycoprotein that interacts with most extracellular matrix proteins responsible for several processes, including angiogenesis by inducing endothelial cell proliferation.13 ADAMTSL4 encodes a secreted glycoprotein that is localized to the extracellular matrix and thought to play a role in cell-cell or cell-extracellular matrix adhesion.14,15 This class of proteins maintains cell polarity and positioning to secure and stabilize tissue architecture. Dysregulation of these processes may contribute to impaired integrity of blood vessel walls. C1orf54 encodes a predicted secreted protein of unknown function, but a previous study16 reported significant upregulation in a rabbit model of carotid artery aneurysms.
At the 6p24.1 locus, the rs9349379-A SCAD risk allele was previously reported as a risk allele in FMD,17 migraine,18 and cervical artery dissection19 (eTable 4 in the Supplement). Moreover, a targeted genotyping study4 that included patients in the discovery cohort of the current study established an association between SCAD and rs9349379-A. PHACTR1, the sole candidate at this locus, encodes a binding protein that interacts with both phosphatase 1 and actin and is responsible for regulation of the actin cytoskeleton.20 PHACTR1 depletion in human umbilical vein endothelial cells disrupts the fine-tuning of actin polymerization and lamellipodial dynamics, implicating PHACTR1 as a key component in angiogenesis.21 According to the Genotype Tissue Expression project, there are significant rs9349379-PHACTR1 eQTLs with the same directionality in 3 vascular tissues.9 Previous studies22,23 provide further supportive evidence of the biological association of rs9349379 with a vascular eQTL by demonstrating reduced expression of PHACTR1 with rs9349379-G in human coronary arteries. However, a subsequent report24 identified a transeffect of the rs9349379 SNV on differential expression of EDN1 (endothelin-1), a gene 600 kb from the index SNV that encodes a potent vasoconstrictor. Moreover, plasma levels of endothelin-1 were increased in 99 healthy individuals with rs9349379-G3 and decreased in 180 individuals with SCAD who carried the rs9349379-A,25 suggesting allele-specific regulation of EDN1 expression.
At the 12q13.3 locus, the rs11172113-T SCAD risk allele was previously reported as a risk allele in migraine,18 cervical artery dissection,19 and aortic aneurysms26 (eTable 4 in the Supplement). LRP1, with 3 lines of support for gene candidacy, encodes a large multifunctional endocytic and signaling receptor protein involved in a number of processes, including focal adhesion disassembly and downstream signaling for reorganization of focal adhesion.27 Inactivation of LRP1 in smooth muscle cells of mice leads to disruption of elastic fibers, aneurysm formation, and abnormally activated transforming growth factor β (TGFβ) signaling akin to the underlying mechanism of FBN1 aortopathy.28,29 Collectively, these studies highlight the importance of LRP1 in regulating and maintaining vascular integrity and suggest that it may play a protective role in vascular smooth muscle cells. SCAD occurs most frequently in women (>80% of cases), suggesting that estrogen and progesterone may be associated with susceptibility to the disorder.25 Estrogen has been implicated in the enhanced release of matrix metalloproteinases, responsible for degradation of the extracellular matrix and impaired structural integrity of the vessel wall.30 Matrix metalloproteinase 9 (MMP9), important for controlling angiogenesis and extracellular matrix remodeling, binds directly to LRP1, which mediates clearance of the matrix metalloproteinases.31 This finding suggests that the LRP1-MMP9 axis may be essential in the modulation of vascular extracellular matrix remodeling31 and may be associated with SCAD pathogenesis.
At the 15q21.1 locus, the C-allele of the triallelic rs2015637 SNV was associated with SCAD. The rarest G-allele was previously associated with thoracic aortic aneurysms and dissections32 (eTable 4 in the Supplement). FBN1, the candidate gene for this region, encodes a large extracellular matrix glycoprotein, a structural component of microfibrils in the extracellular matrix.33 FBN1 is important for maintenance of vascular integrity. Rare sequence variants within the gene that accentuate TGFβ signaling are a well-recognized monogenic cause of Marfan syndrome, a connective tissue disease associated with ascending aortic aneurysm and dissection.32,34 However, a case report described a de novo heterozygous missense variant in FBN1 in an individual with recurrent SCAD who lacked traits of Marfan syndrome.35 Of note, the protein encoded by ADAMTSL4, a SCAD candidate gene at the 1q21.3 locus, directly binds to FBN1 and promotes microfibril assembly.15
At the 21q22.11 locus, LINC00310 was identified as a top candidate. Once considered as transcriptional noise, long, noncoding RNAs, such as LINC00310, are now implicated in gene regulation and other cellular processes.36 Except for a study37 demonstrating an association with cell proliferation in breast cancer, the function and overall clinical significance of LINC00310 are unknown.
Beyond a relatively low proportion of traditional risk factors for CAD among patients with SCAD, coronary imaging demonstrates little or no evidence of typical atherosclerotic lesions.1,38 A previous report4 of the intronic rs9349379-A allele within PHACTR1 as a SCAD risk allele that included patients in the current discovery cohort contrasts a previous GWAS that identified rs9349379-G as a CAD risk allele.39,40 This dichotomy of opposing risk alleles in SCAD vs CAD is further exemplified in the intronic rs11172113-T within LRP1 and the LINC00310-associated rs28451064-G SCAD risk alleles identified in the current study as well as the rs11172113-C and rs28451064-A CAD and acute MI risk alleles reported in previous GWASs (eTable 4 and eFigure 3 in the Supplement).41,42 The differential associations of the risk alleles rs9349379 with PHACTR1 and EDN124 expression and rs11172113 with LRP1 expression (eFigure 2 in the Supplement) provide a plausible explanation of CAD vs SCAD susceptibility. For EDN1, contrasting vasoconstrictive and vasodilatory associations have been implicated.24 The functions of PHACTR1 and LRP1 are multifaceted and cell type specific. The risk alleles may be associated with disease susceptibility by differential perturbations of vascular inflammatory pathways, TGFβ signaling, and/or cytoskeletal integrity.28,43,44,45,46 Functional protein-protein interactions can elucidate unifying cellular processes associated with complex disease risk. The recently reported SCAD susceptibility gene, TLN1,3 has indirect functional associations with LRP1, PHACTR1, and FBN1 via actins and integrins. Talin 1 is responsible for linking actin in the cytosol to integrin in the extracellular matrix, forming the core of a focal adhesion complex.47 PHACTR1 and TLN1 interact directly with actin, and integrin-mediated interactions with FBN1, LRP1, and TLN1 regulate cell adhesion and migration and initiate signaling pathways that lead to reorganization of the actin cytoskeleton.47,48,49 Emerging data thus suggest that both rare and common genetic variants are associated with SCAD risk and that genetic perturbation of the actin cytoskeleton may be one of the pathways for disease.
Limitations
Our study design included strict inclusionary and exclusionary diagnostic criteria for patients and controls, a dense genotyping platform, and replication of associations. Notwithstanding, this single-center study was confined to white women, comprising 96% of patients enrolled in the SCAD genetics research cohort. To our knowledge, the current study is the only SCAD GWAS to date. Accordingly, our findings should be further investigated in additional, demographically diverse cohorts with SCAD to validate our results and determine whether SCAD risk alleles are specific to clinical subtypes, sex, and race/ethnicity. For example, a gene-based association study50 in a predominantly male Chinese Han population identified SCAD-associated variants in TSR1, a signal that was not identified within our study. The cases and controls in our study were genotyped or sequenced on multiple platforms with resultant gaps in coverage between genome-wide SNV platforms. However, genotype imputation and quality control metrics ensured harmonization across platforms. Beyond the identified common variants associated with SCAD risk, additional, unidentified gene and/or environmental substrates and triggers would be necessary to precipitate a SCAD event. The prioritized positional candidate genes identified to be associated with SCAD within GWAS loci have supportive evidence through bioinformatics. Notwithstanding, functional validation is required to demonstrate the association of noncoding SNVs with gene expression, including genes located beyond the 400-kb flanking regions, as demonstrated with rs9349379 and EDN1.24 Functional validation is also required to confirm protein-protein interactions between SCAD candidate genes and TLN1.
Conclusions
This study revealed risk loci and positional candidate genes associated with SCAD that have established associations with extracoronary arteriopathies, providing further insight into the genetic architecture of the disorder. Our data, together with findings in previous reports,4,39,40,41,42 further suggest allelic susceptibility to coronary artery atherosclerosis vs dissection.
eMethods. Supplementary Methods
eFigure 1. Quantile-Quantile Plot
eFigure 2. Allele-Specific Cis-eQTL Gene Expression in Arterial Tissues
eFigure 3. SNP Associations With SCAD Vs CAD
eTable 1. Prioritization of Genes Within Each Locus
eTable 2. Analysis of Regulatory Elements Overlapping Index SNPs
eTable 3. STRING Analysis of Candidate Gene Protein Interactions With TLN1
eTable 4. SNP-Trait Association and Genotyping Studies
eReferences
References
- 1.Hayes SN, Kim ESH, Saw J, et al. ; American Heart Association Council on Peripheral Vascular Disease; Council on Clinical Cardiology; Council on Cardiovascular and Stroke Nursing; Council on Genomic and Precision Medicine; and Stroke Council . Spontaneous coronary artery dissection: current state of the science: a scientific statement from the American Heart Association. Circulation. 2018;137(19):e523-e557. doi: 10.1161/CIR.0000000000000564 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Goel K, Tweet M, Olson TM, Maleszewski JJ, Gulati R, Hayes SN. Familial spontaneous coronary artery dissection: evidence for genetic susceptibility. JAMA Intern Med. 2015;175(5):821-826. doi: 10.1001/jamainternmed.2014.8307 [DOI] [PubMed] [Google Scholar]
- 3.Turley TN, Theis JL, Sundsbak RS, et al. Rare missense variants in TLN1 are associated with familial and sporadic spontaneous coronary artery dissection. Circ Genom Precis Med. 2019;12(4):e002437. doi: 10.1161/CIRCGEN.118.002437 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Adlam D, Olson TM, Combaret N, et al. ; DISCO Consortium; CARDIoGRAMPlusC4D Study Group . Association of the PHACTR1/EDN1 genetic locus with spontaneous coronary artery dissection. J Am Coll Cardiol. 2019;73(1):58-66. doi: 10.1016/j.jacc.2018.09.085 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Tweet MS, Hayes SN, Pitta SR, et al. Clinical features, management, and prognosis of spontaneous coronary artery dissection. Circulation. 2012;126(5):579-588. doi: 10.1161/CIRCULATIONAHA.112.105718 [DOI] [PubMed] [Google Scholar]
- 6.Bielinski SJ, Chai HS, Pathak J, et al. ; Mayo Genome Consortia . Mayo Genome Consortia: a genotype-phenotype resource for genome-wide association studies with an application to the analysis of circulating bilirubin levels. Mayo Clin Proc. 2011;86(7):606-614. doi: 10.4065/mcp.2011.0178 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Olson JE, Ryu E, Johnson KJ, et al. The Mayo Clinic Biobank: a building block for individualized medicine. Mayo Clin Proc. 2013;88(9):952-962. doi: 10.1016/j.mayocp.2013.06.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Purcell S, Neale B, Todd-Brown K, et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet. 2007;81(3):559-575. doi: 10.1086/519795 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.GTEx Consortium Human genomics. the Genotype-Tissue Expression (GTEx) pilot analysis: multitissue gene regulation in humans. Science. 2015;348(6235):648-660. doi: 10.1126/science.1262110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Szklarczyk D, Gable AL, Lyon D, et al. STRING v11: protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019;47(D1):D607-D613. doi: 10.1093/nar/gky1131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cavdar Koc E, Burkhart W, Blackburn K, Moseley A, Spremulli LL. The small subunit of the mammalian mitochondrial ribosome: identification of the full complement of ribosomal proteins present. J Biol Chem. 2001;276(22):19363-19374. doi: 10.1074/jbc.M100727200 [DOI] [PubMed] [Google Scholar]
- 12.Gopisetty G, Thangarajan R. Mammalian mitochondrial ribosomal small subunit (MRPS) genes: a putative role in human disease. Gene. 2016;589(1):27-35. doi: 10.1016/j.gene.2016.05.008 [DOI] [PubMed] [Google Scholar]
- 13.Han Z, Ni J, Smits P, et al. Extracellular matrix protein 1 (ECM1) has angiogenic properties and is expressed by breast tumor cells. FASEB J. 2001;15(6):988-994. doi: 10.1096/fsb2fj990934com [DOI] [PubMed] [Google Scholar]
- 14.Porter S, Clark IM, Kevorkian L, Edwards DR. The ADAMTS metalloproteinases. Biochem J. 2005;386(pt 1):15-27. doi: 10.1042/BJ20040424 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hubmacher D, Apte SS. ADAMTS proteins as modulators of microfibril formation and function. Matrix Biol. 2015;47:34-43. doi: 10.1016/j.matbio.2015.05.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Holcomb M, Ding YH, Dai D, et al. RNA-sequencing analysis of messenger RNA/microRNA in a rabbit aneurysm model identifies pathways and genes of interest. AJNR Am J Neuroradiol. 2015;36(9):1710-1715. doi: 10.3174/ajnr.A4390 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kiando SR, Tucker NR, Castro-Vega LJ, et al. PHACTR1 is a genetic susceptibility locus for fibromuscular dysplasia supporting its complex genetic pattern of inheritance. PLoS Genet. 2016;12(10):e1006367. doi: 10.1371/journal.pgen.1006367 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Anttila V, Winsvold BS, Gormley P, et al. ; North American Brain Expression Consortium; UK Brain Expression Consortium . Genome-wide meta-analysis identifies new susceptibility loci for migraine. Nat Genet. 2013;45(8):912-917. doi: 10.1038/ng.2676 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Debette S, Kamatani Y, Metso TM, et al. ; International Stroke Genetics Consortium; CADISP Group . Common variation in PHACTR1 is associated with susceptibility to cervical artery dissection. Nat Genet. 2015;47(1):78-83. doi: 10.1038/ng.3154 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Allen PB, Greenfield AT, Svenningsson P, Haspeslagh DC, Greengard P. Phactrs 1-4: a family of protein phosphatase 1 and actin regulatory proteins. Proc Natl Acad Sci U S A. 2004;101(18):7187-7192. doi: 10.1073/pnas.0401673101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Allain B, Jarray R, Borriello L, et al. Neuropilin-1 regulates a new VEGF-induced gene, Phactr-1, which controls tubulogenesis and modulates lamellipodial dynamics in human endothelial cells. Cell Signal. 2012;24(1):214-223. doi: 10.1016/j.cellsig.2011.09.003 [DOI] [PubMed] [Google Scholar]
- 22.Beaudoin M, Gupta RM, Won HH, et al. Myocardial infarction-associated SNP at 6p24 interferes with MEF2 binding and associates with PHACTR1 expression levels in human coronary arteries. Arterioscler Thromb Vasc Biol. 2015;35(6):1472-1479. doi: 10.1161/ATVBAHA.115.305534 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wang X, Musunuru K. Confirmation of causal rs9349379-PHACTR1 expression quantitative trait locus in human-induced pluripotent stem cell endothelial cells. Circ Genom Precis Med. 2018;11(10):e002327. doi: 10.1161/CIRCGEN.118.002327 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gupta RM, Hadaya J, Trehan A, et al. A genetic variant associated with five vascular diseases is a distal regulator of endothelin-1 gene expression. Cell. 2017;170(3):522-533.e15. doi: 10.1016/j.cell.2017.06.049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Tweet MS, Miller VM, Hayes SN. The evidence on estrogen, progesterone, and spontaneous coronary artery dissection. JAMA Cardiol. 2019;4(5):403-404. doi: 10.1001/jamacardio.2019.0774 [DOI] [PubMed] [Google Scholar]
- 26.Guo DC, Grove ML, Prakash SK, et al. ; GenTAC Investigators; BAVCon Investigators . Genetic variants in LRP1 and ULK4 are associated with acute aortic dissections. Am J Hum Genet. 2016;99(3):762-769. doi: 10.1016/j.ajhg.2016.06.034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Orr AW, Pedraza CE, Pallero MA, et al. Low density lipoprotein receptor-related protein is a calreticulin coreceptor that signals focal adhesion disassembly. J Cell Biol. 2003;161(6):1179-1189. doi: 10.1083/jcb.200302069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Strickland DK, Au DT, Cunfer P, Muratoglu SC. Low-density lipoprotein receptor-related protein-1: role in the regulation of vascular integrity. Arterioscler Thromb Vasc Biol. 2014;34(3):487-498. doi: 10.1161/ATVBAHA.113.301924 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zhou L, Takayama Y, Boucher P, Tallquist MD, Herz J. LRP1 regulates architecture of the vascular wall by controlling PDGFRbeta-dependent phosphatidylinositol 3-kinase activation. PLoS One. 2009;4(9):e6922. doi: 10.1371/journal.pone.0006922 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wingrove CS, Garr E, Godsland IF, Stevenson JC. 17β-oestradiol enhances release of matrix metalloproteinase-2 from human vascular smooth muscle cells. Biochim Biophys Acta. 1998;1406(2):169-174. doi: 10.1016/S0925-4439(97)00097-5 [DOI] [PubMed] [Google Scholar]
- 31.Hahn-Dantona E, Ruiz JF, Bornstein P, Strickland DK. The low density lipoprotein receptor-related protein modulates levels of matrix metalloproteinase 9 (MMP-9) by mediating its cellular catabolism. J Biol Chem. 2001;276(18):15498-15503. doi: 10.1074/jbc.M100121200 [DOI] [PubMed] [Google Scholar]
- 32.LeMaire SA, McDonald ML, Guo DC, et al. Genome-wide association study identifies a susceptibility locus for thoracic aortic aneurysms and aortic dissections spanning FBN1 at 15q21.1. Nat Genet. 2011;43(10):996-1000. doi: 10.1038/ng.934 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sakai LY, Keene DR, Engvall E. Fibrillin, a new 350-kD glycoprotein, is a component of extracellular microfibrils. J Cell Biol. 1986;103(6 Pt 1):2499-2509. doi: 10.1083/jcb.103.6.2499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Dietz HC, Cutting GR, Pyeritz RE, et al. Marfan syndrome caused by a recurrent de novo missense mutation in the fibrillin gene. Nature. 1991;352(6333):337-339. doi: 10.1038/352337a0 [DOI] [PubMed] [Google Scholar]
- 35.von Hundelshausen P, Oexle K, Bidzhekov K, et al. Recurrent spontaneous coronary dissections in a patient with a de novo fibrillin-1 mutation without Marfan syndrome. Thromb Haemost. 2015;113(3):668-670. doi: 10.1160/TH14-11-0913 [DOI] [PubMed] [Google Scholar]
- 36.Ulitsky I, Bartel DP. lincRNAs: genomics, evolution, and mechanisms. Cell. 2013;154(1):26-46. doi: 10.1016/j.cell.2013.06.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Li J, Peng W, Du L, Yang Q, Wang C, Mo YY. The oncogenic potentials and diagnostic significance of long non-coding RNA LINC00310 in breast cancer. J Cell Mol Med. 2018;22(9):4486-4495. doi: 10.1111/jcmm.13750 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Tweet MS, Akhtar NJ, Hayes SN, Best PJ, Gulati R, Araoz PA. Spontaneous coronary artery dissection: acute findings on coronary computed tomography angiography. Eur Heart J Acute Cardiovasc Care. 2019;8(5):467-475. doi: 10.1177/2048872617753799 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.CARDIoGRAMplusC4D Consortium, Deloukas P, Kanoni S, Willenborg C, et al. Large-scale association analysis identifies new risk loci for coronary artery disease. Nat Genet 2013;45(1):25-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Nikpay M, Goel A, Won HH, et al. A comprehensive 1,000 Genomes-based genome-wide association meta-analysis of coronary artery disease. Nat Genet. 2015;47(10):1121-1130. doi: 10.1038/ng.3396 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Nelson CP, Goel A, Butterworth AS, et al. ; EPIC-CVD Consortium; CARDIoGRAMplusC4D; UK Biobank CardioMetabolic Consortium CHD working group . Association analyses based on false discovery rate implicate new loci for coronary artery disease. Nat Genet. 2017;49(9):1385-1391. doi: 10.1038/ng.3913 [DOI] [PubMed] [Google Scholar]
- 42.Webb TR, Erdmann J, Stirrups KE, et al. ; Wellcome Trust Case Control Consortium; MORGAM Investigators; Myocardial Infarction Genetics and CARDIoGRAM Exome Consortia Investigators . Systematic evaluation of pleiotropy identifies 6 further loci associated with coronary artery disease. J Am Coll Cardiol. 2017;69(7):823-836. doi: 10.1016/j.jacc.2016.11.056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Fils-Aimé N, Dai M, Guo J, et al. MicroRNA-584 and the protein phosphatase and actin regulator 1 (PHACTR1), a new signaling route through which transforming growth factor-β mediates the migration and actin dynamics of breast cancer cells. J Biol Chem. 2013;288(17):11807-11823. doi: 10.1074/jbc.M112.430934 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Reschen ME, Lin D, Chalisey A, Soilleux EJ, O’Callaghan CA. Genetic and environmental risk factors for atherosclerosis regulate transcription of phosphatase and actin regulating gene PHACTR1. Atherosclerosis. 2016;250:95-105. doi: 10.1016/j.atherosclerosis.2016.04.025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Xian X, Ding Y, Dieckmann M, et al. LRP1 integrates murine macrophage cholesterol homeostasis and inflammatory responses in atherosclerosis. Elife. 2017;6:e29292. doi: 10.7554/eLife.29292 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Au DT, Ying Z, Hernández-Ochoa EO, et al. LRP1 (low-density lipoprotein receptor-related protein 1) regulates smooth muscle contractility by modulating Ca2+ signaling and expression of cytoskeleton-related proteins. Arterioscler Thromb Vasc Biol. 2018;38(11):2651-2664. doi: 10.1161/ATVBAHA.118.311197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Gough RE, Goult BT. The tale of two talins: two isoforms to fine-tune integrin signalling. FEBS Lett. 2018;592(12):2108-2125. doi: 10.1002/1873-3468.13081 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Bax DV, Bernard SE, Lomas A, et al. Cell adhesion to fibrillin-1 molecules and microfibrils is mediated by alpha 5 beta 1 and alpha v beta 3 integrins. J Biol Chem. 2003;278(36):34605-34616. doi: 10.1074/jbc.M303159200 [DOI] [PubMed] [Google Scholar]
- 49.Wujak L, Böttcher RT, Pak O, et al. Low density lipoprotein receptor-related protein 1 couples β1 integrin activation to degradation. Cell Mol Life Sci. 2018;75(9):1671-1685. doi: 10.1007/s00018-017-2707-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sun Y, Chen Y, Li Y, et al. Association of TSR1 variants and spontaneous coronary artery dissection. J Am Coll Cardiol. 2019;74(2):167-176. doi: 10.1016/j.jacc.2019.04.062 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
eMethods. Supplementary Methods
eFigure 1. Quantile-Quantile Plot
eFigure 2. Allele-Specific Cis-eQTL Gene Expression in Arterial Tissues
eFigure 3. SNP Associations With SCAD Vs CAD
eTable 1. Prioritization of Genes Within Each Locus
eTable 2. Analysis of Regulatory Elements Overlapping Index SNPs
eTable 3. STRING Analysis of Candidate Gene Protein Interactions With TLN1
eTable 4. SNP-Trait Association and Genotyping Studies
eReferences