

Abstract
Aging-induced functional and phenotypic alterations of the vasculature (e.g., endothelial dysfunction, oxidative stress) have a central role in morbidity and mortality of older adults. It has become apparent in recent years that cell autonomous mechanisms alone are inadequate to explain all aspects of vascular aging. The present study was designed to test the hypothesis that age-related changes in circulating anti-geronic factors contribute to the regulation of vascular aging processes in a non-cell autonomous manner. To test this hypothesis, through heterochronic parabiosis we determined the extent, if any, to which endothelial function, vascular production of ROS, and shifts in the vascular transcriptome (RNA-seq) are modulated by the systemic environment. We found that in aortas isolated from isochronic parabiont aged (20-month-old) C57BL/6 mice [A-(A); parabiosis for 8 weeks] acetylcholine-induced endothelium-dependent relaxation was impaired and ROS production (dihydroethidium fluorescence) was increased as compared with those in aortas from young isochronic parabiont (6-month-old) mice [Y-(Y)]. The presence of young blood derived from young parabionts significantly improved endothelium-dependent vasorelaxation and attenuated ROS production in vessels of heterochronic parabiont aged [A-(Y)] mice. In aortas derived from heterochronic parabiont young [Y-(A)] mice, acetylcholine-induced relaxation and ROS production were comparable with those in aortas derived from Y-(Y) mice. Using RNA-seq we assessed transcriptomic changes in the aortic arch associated with aging and heterochronic parabiosis. We identified 347 differentially expressed genes in A-(A) animals compared with Y-(Y) controls. We have identified 212 discordant genes, whose expression levels differed in the aged phenotype, but have shifted back toward the young phenotype by the presence of young blood in aged A-(Y) animals. Pathway analysis shows that vascular protective effects mediated by young blood–regulated genes include mitochondrial rejuvenation. In conclusion, a relatively short-term exposure to young blood can rescue vascular aging phenotypes, including attenuation of oxidative stress, mitochondrial rejuvenation, and improved endothelial function. Our findings provide additional evidence supporting the significant plasticity of vascular aging and evidence for the existence of anti-geronic factors capable of exerting rejuvenating effects on the aging vasculature.
Keywords: Aging, Senescence, Geronic factors
Introduction
Cardiovascular and cerebrovascular diseases are the most common causes of serious long-term disability and mortality among older adults in the developed world (Ungvari et al. 2018a), accounting for approximately one-third of all deaths in the USA at the age of 65 and nearly two-thirds of all deaths at an age of 85 (Health, United States 2016). Importantly, epidemiological studies reveal that the effects of traditional risk factors (such as hypertension, dyslipidemia, cigarette smoking, obesity, diabetes mellitus, etc.) on the incidence of cardiovascular and cerebrovascular diseases are dwarfed by the single most important risk factor for these diseases: advanced aging (Ungvari et al. 2018a). In order to develop novel methods for prevention and treatment of vascular pathologies in older adults, it is important to understand the role of shared mechanisms of aging in the cellular and functional changes that occur in the vasculature during aging (Lakatta and Levy 2003).
In the past two decades, most studies on vascular aging have focused on cell autonomous mechanisms driving functional decline (Ungvari et al. 2018a). These studies have led to a comprehensive understanding of the critical roles of mitochondrial dysfunction (Tarantini et al. 2018; Gioscia-Ryan et al. 2014), increased production of reactive oxygen species(ROS) (Ungvari et al. 2018a; Csiszar et al. 2014; Csiszar et al. 2009; Toth et al. 2015a; Ungvari et al. 2007; Csiszar et al. 2002; Labinskyy et al. 2006; Csiszar et al. 2007a; Csiszar et al. 2007b; Csiszar et al. 2012; Fulop et al. 2018; Ungvari et al. 2011), impaired NO bioavailability (Csiszar et al. 2002; Tarantini et al. 2019a; Csiszar et al. 2015; Toth et al. 2014a), consequences of cellular NAD+ depletion (Tarantini et al. 2019b; Kiss et al. 2019a; Kiss et al. 2019b; Das et al. 2018), dysregulation of sirtuins(Csiszar et al. 2009; Mattison et al. 2014), and energetic dysfunction (Tarantini et al. 2018) in the etiology of vascular aging. However, it has also become apparent in recent years that these important cell autonomous mechanisms alone are inadequate to explain all aspects of vascular aging (Ungvari et al. 2018a).
In the field of geroscience, compelling evidence has been accumulated that implicates non-cell autonomous mechanisms as also playing important roles in driving aging processes and pathogenesis of age-related diseases (Ashpole et al. 2016; Ashpole et al. 2017; Lopez-Otin et al. 2013; Katsimpardi et al. 2014; Morrison et al. 2019; Rebo et al. 2016). Recently, the hypothesis was put forward that aging is associated with complex changes in inter-organ communication, which modulate cell autonomous aging processes in the vasculature contributing to pathogenesis of age-related vascular diseases(Ungvari et al. 2018a). An important prediction based on this hypothesis is that circulating pro-geronic factors (characterized by their increased production with age and deleterious effects on vascular homeostasis, e.g., inflammatory cytokines) and/or the presence of anti-geronic factors (which reverse or prevent development of cellular aging phenotypes and presumably decline with age), which are derived from other organs (e.g., adipose tissue, the brain, endocrine system, immune system) orchestrate aging processes in cells within the vascular wall. Yet, additional experimental evidence is needed to better understand the relative contribution of circulating factors versus cell autonomous mechanisms to the genesis of vascular aging phenotypes.
Parabiosis is a surgical approach for joining the circulatory systems of two animals that has been widely used to study the role of circulating (humoral and cellular) factors in the pathogenesis of various diseases, including endocrine and metabolic diseases (Harris 1997; Harris 1999; Coleman 1973; Coleman and Hummel 1969) as well as regulation of lifespan (Ludwig and Elashoff 1972; Horrington et al. 1960; Lunsford et al. 1963; McCay et al. 1957). Parabiosis re-emerged over the past 15 years in geroscience research to investigate the complex interaction between cell autonomous and non-cell autonomous mechanisms of aging comparing heterochronic (young–old) and isochronic (young–young and old–old) parabiont pairs of animals (Katsimpardi et al. 2014; Rebo et al. 2016; Lunsford et al. 1963; McCay et al. 1957; Bitto and Kaeberlein 2014; Cannata et al. 2017; Conboy et al. 2013; Fan et al. 2017; Flemming 2013; Ghosh et al. 2019; Gontier et al. 2018; Harrison and Astle 1982; Hirayama et al. 1993; Katsimpardi et al. 2020; Sousa-Victor et al. 2019; Villeda et al. 2011; Villeda et al. 2014; Zhang et al. 2019; DeCarolis et al. 2015; Middeldorp et al. 2016; Smith et al. 2015a). Using heterochronic parabiosis as an experimental tool, it was demonstrated that both pro-geronic and anti-geronic circulating factors mediate non-cell autonomous effects driving aging in multiple organs, including the skeletal muscle, central nervous system, and the heart (Katsimpardi et al. 2014; Fan et al. 2017; Villeda et al. 2011; Villeda et al. 2014; DeCarolis et al. 2015; Smith et al. 2015a; Castellano et al. 2016).
The present study was designed to experimentally test the hypothesis that age-related changes in circulating pro-geronic factors and/or anti-geronic factors contribute to the regulation of vascular aging processes in a non-cell autonomous manner. To test this hypothesis we leveraged heterochronic parabiosis in combination with vascular functional assays and transcriptomic profiling, to determine to what extent, if any, transposition of aging phenotypes could be observed in the young and old aorta by exposure to an old and young environment, respectively.
Methods
Animals and parabiosis surgery
Young (4-month-old) and aged (18-month-old) male C57BL/6 mice were obtained from the aging rodent colony maintained by the National Institute on Aging at Charles River Laboratories (Wilmington, MA). Mice were housed under specific pathogen-free conditions at the rodent barrier facility at Albert Einstein College of Medicine under a controlled photoperiod (12 h light; 12 h dark) with unlimited access to water and were fed a standard AIN-93G diet (ad libitum). Parabiosis surgery in young and aged animals was carried out by the Einstein Chronobiosis and Energetics/Metabolism of Aging Core, according to published protocols (Harris 1997; Harris 1999) as reported previously (Morrison et al. 2019). Surgical unions were performed between young animals (isochronic; young Y–(Y); n = 4 pairs), aged animals (isochronic old; A–(A); n = 4 pairs), and young and aged mice (heterochronic Y–(A) and A-(Y); n = 5 pairs) in the Einstein Health Span Core as described (Morrison et al. 2019). Following surgery, animals were kept on a partial heating pad overnight. Pairs were then intensively monitored and received subcutaneous (s.c) injections of Banamine (2 mg/kg each) immediately post-op and twice a day for t3 days and then once daily for 4 days. Animals also received 1 mL of Ringer’s lactate (s.c.) immediately after, daily for 3 days post-op to prevent dehydration. Animals remained joined for ~ 8 weeks prior to sacrifice. All experimental procedures were approved by the Institutional Animal Care and Use Committee (IACUC) at the Albert Einstein College of Medicine and the University of Oklahoma Health Sciences Center. All animal experiments were carried out in accordance with the National Institutes of Health guide for the care and use of Laboratory animals (NIH Publications No. 8023, revised 1978).
Assessment of endothelial function in the aorta
To assess the specific effect of circulating factors on endothelial function, endothelium-dependent vasorelaxation was assessed in isolated aorta ring preparations as described previously (Pearson et al. 2008). In brief, animals were killed as reported (Morrison et al. 2019) and the thoracic aortas were cleaned from the connective tissue and perivascular fat, cut into ring segments 1.5 mm in length and mounted in myographs chambers (Danish Myo Technology A/S, Inc., Denmark) for measurement of isometric tension. The vessels were superfused with Krebs buffer solution (118 mM NaCl, 4.7 mM KCl, 1.5 mM CaCl2, 25 mM NaHCO3, 1.1 mM MgSO4, 1.2 mM KH2PO4, and 5.6 mM glucose; at 37 °C; gassed with 95% air and 5% CO2). After an equilibration period of 1 h during which an optimal passive tension was applied to the rings (as determined from the vascular length-tension relationship), they were pre-contracted with 10−6 M phenylephrine and relaxation in response to acetylcholine was measured. The remaining aortic arch tissue was snap-frozen and stored at − 80 °C for transcriptomic analysis.
Assessment of vascular oxidative stress
To characterize vascular ROS production isolated segments of the aorta were loaded with the redox-sensitive dye dihydroethidium (DHE, Invitrogen, Carlsbad CA; 3 × 10−6 mol/L; for 30 min) in oxygenated Krebs solution (at 37 °C) as previously reported (Toth et al. 2015a; Csiszar et al. 2007a; Pearson et al. 2008; Ungvari et al. 2003; Labinskyy et al. 2009). After loading the dye was washed out five times with warm Krebs buffer, and the vessels were allowed to equilibrate for another 20 min. Then, the vessels were embedded in OCT medium and cryosectioned. Confocal images were captured using a Leica SP2 confocal laser scanning microscope (Leica Microsystems GmbH, Wetzlar, Germany). Average nuclear DHE fluorescence intensities were assessed using the Metamorph software (Molecular Devices LLC, Sunnyvale, CA) and values for each animal in each group were averaged.
RNA isolation, cDNA synthesis, library construction, and next generation sequencing
RNA was isolated from the aortic arch samples using AllPrep DNA/RNA Mini Kit (Qiagen) as previously described (Imperio et al. 2016; Valcarcel-Ares et al. 2019). Prior to 3′-tag RNA-seq analysis quality control measures were implemented. Concentration of RNA was ascertained via fluorometric analysis on a Thermo Fisher Qubit fluorometer. Overall quality of RNA was verified using an Agilent Tapestation instrument. Following initial QC steps sequencing libraries were generated using the Lexogen Quantseq FWD library prep kit according to the manufacturer’s protocol by the Quantitative Analysis Core of the Oklahoma Medical Research Foundation (Kiss et al. 2020). Briefly, the first strand of cDNA was generated using 5′-tagged poly-T oligomer primers. Following RNase digestion, the second strand of cDNA was generated using 5′-tagged random primers. A subsequent PCR step with additional primers added the complete adapter sequence to the initial 5′ tags, added unique indices for demultiplexing of samples, and amplified the library. Final libraries for each sample were assayed on the Agilent Tapestation for appropriate size and quantity. These libraries were then pooled in equimolar amounts as ascertained via fluorometric analyses. Final pools were absolutely quantified using qPCR on a Roche LightCycler 480 instrument with Kapa Biosystems Illumina Library Quantification reagents. Sequencing was performed by the Quantitative Analysis Core of the Oklahoma Medical Research Foundation (Kiss et al. 2020) using custom primers on an Illumina NextSeq 500 instrument with High Output chemistry and 75 bp single-ended reads.
RNA-Seq data analysis and visualization
Raw sequencing reads were trimmed of their Illumina TruSeq adapter sequences using Trimmomatic v0.35(Bolger et al. 2014), filtered for contaminants of ribosomal, mitochondrial, and hemoglobin transcripts, then aligned to the mouse genome version GRCm38 using Kallisto v0.43.03 (Bray et al. 2016). Samples were checked for outliers and separation by principle components analysis (PCA) with the R function prcomp. Raw expression counts were summarized at the gene level to transcript-length adjusted, library-size scaled counts per million (CPM) with the R/Bioconductor package tximport (Soneson et al. 2015). Differential expression analysis was performed using the empirical Bayes approach implemented in the R/Bioconductor package DESeq2 (Love et al. 2014). Significantly differentially expressed (DE) genes had an absolute log2 fold change ≥ 0.585 (corresponding to a change of 50% or more in expression) and the False Discovery Rate FDR-adjusted p value ≤ 0.05. Gene annotation was done using biomaRt (Durinck et al. 2009) in R/Bioconductor package. The R package pheatmap v1.0.12 was used to perform hierarchical clustering and to draw the heatmaps. The org.Mm.eg.db v3.8.2 R/Bioconductor package was used to collect Gene Ontology, KEGG, and Reactome terms associated with the DE genes. The same package was used to translate Ensemble IDs to Entrez IDs when it was required by the statistical packages.
Functional annotation
The org.Mm.eg.db v3.8.2 R/Bioconductor package was used to collect Gene Ontology terms associated with our differentially expressed genes. The hypergeometric test implemented in GOstats v2.51.0. R/Bioconductor package was used to calculate enrichment of the individual terms (Falcon and Gentleman 2007).
We used Upstream Regulator Analysis (URA) algorithm (Kramer et al. 2014) implemented in the Ingenuity Pathway Analysis (QIAGEN) software to identify upstream regulators that potentially explain the observed gene expression changes in our samples. The IPA uses a manually curated database (Ingenuity Knowledge Base) to calculate “enrichment” score [Fisher’s exact test (FET) p value] measures the overlap of observed and predicted regulated gene sets, and a z-score assessing the match of observed and predicted up/downregulation patterns.
Reactome pathway Knowledgebase (Fabregat et al. 2018)
Reactome is a free, open-source, curated, and peer-reviewed database of biological pathways. Enrichment of Reactome pathways was calculated and results were plotted by the ReacomePA v1.29.0 R/Bioconductor package (Yu and He 2016). org.Mm.eg.db v3.8.2 R/Bioconductor package was used to convert Ensemble IDs to Entrez IDs since that is the required input format of the ReactomePA package.
KEGG pathway
org.Mm.eg.db v3.8.2 R/Bioconductor package was used to convert Ensemble IDs to Entrez IDs. Pathview R/Bioconductor package was used to overlay the gene expression data with the curated KEGG pathways. Heat maps were drawn by the pheatmap v1.0.12 R package. [https://CRAN.R-project.org/package=pheatmap].
Results
Young blood rescues endothelial function and attenuates oxidative stress in aged aorta
There is strong experimental and clinical evidence that endothelial dysfunction significantly contributes to the pathogenesis of age-related diseases of the cardiovascular system (Ungvari et al. 2018b). To test the endothelial effects of circulating pro- and anti-geronic factors, endothelium-dependent vasodilator responses to acetylcholine were tested. In aorta rings derived from Y-(Y) mice, administration of acetylcholine resulted in significant relaxation, whereas these responses were significantly attenuated in aorta rings obtained from A-(A) mice (Fig. 1a). Acetylcholine-induced vasorelaxation was abolished by the pre-treatment of the aorta rings from each group with the NO synthase inhibitor L-NAME (data not shown). Exposure to young blood significantly improved NO-mediated, endothelium-dependent vasorelaxation in A-(Y) mice (Fig. 1a). In aorta rings derived from Y-(A) mice, acetylcholine-induced relaxation was comparable with that in aortas derived from Y-(Y) mice, suggesting that presence of old blood did not alter endothelial function considerably in young aorta.
Fig. 1.
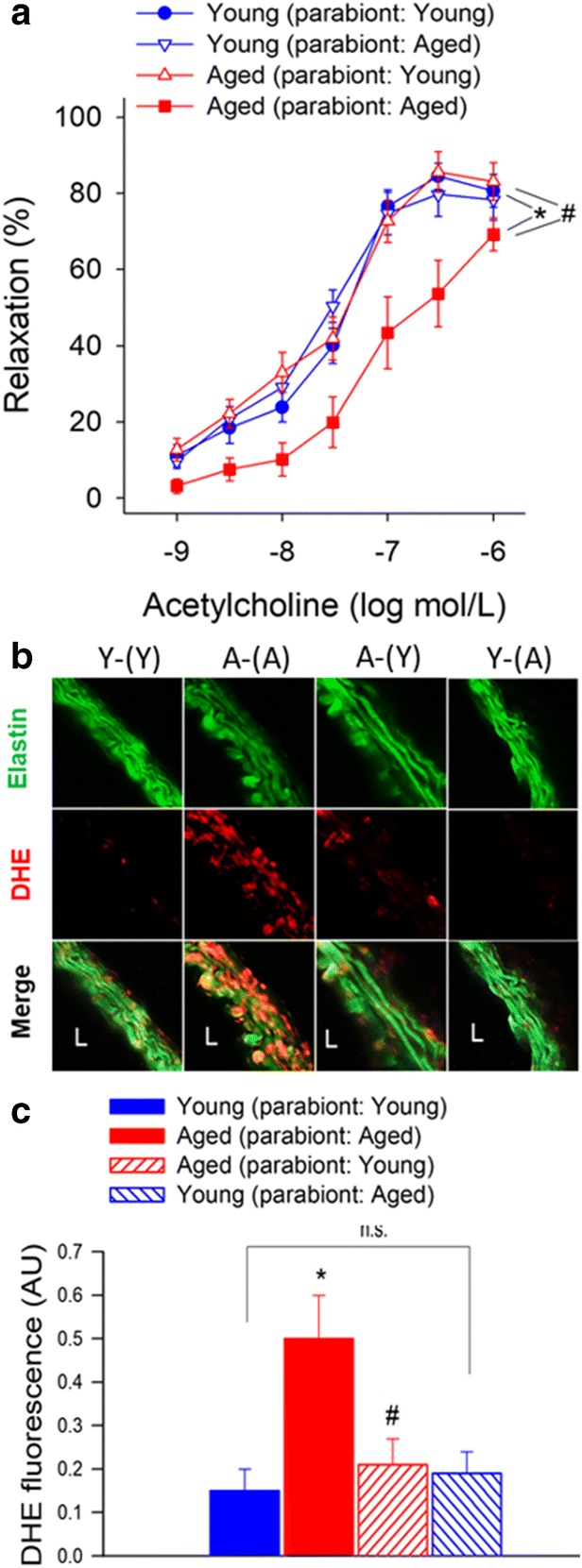
Young blood improves NO-mediated, endothelium-dependent vasorelaxation in aged mice. a Acetylcholine-induced relaxations in aortic ring preparations isolated from isochronic parabiont young mice [Y-(Y)], isochronic parabiont aged mice [A-(A)], heterochronic parabiont young mice [Y-(A)] and heterochronic parabiont aged mice [A-(Y)]. Age-related declines in endothelial function were reversed by the presence of circulating factors derived from young parabionts. Data are mean ± S.E.M. (n = 4–5 mice for each data point). b Representative confocal images showing stronger DHE staining (red fluorescence) indicating increased cellular ROS production in the wall of aortas isolated from A-(A) mice as compared with vessels isolated from Y-(Y) mice. Aortas isolated from A-(Y) mice exhibit weak DHE staining indicating decreased ROS production. Aortas isolated from Y-(A) mice exhibit DHE staining that is comparable to that in aortas isolated from Y-(Y) mice (original magnification: 10x). c Quantification of DHE fluorescence intensities. Data are means±S.E.M. (n = 4–5 in each group) *p < 0.05 aged (parabiont: aged) vs young (parabiont: young); #p < 0.05 aged (parabiont: young) vs. aged (parabiont: aged)
Previous studies demonstrate that aging significantly increases vascular superoxide production and that treatment with antioxidants can restore endothelium-mediated vasodilation in aged rodents (Tarantini et al. 2018; Csiszar et al. 2002; Tarantini et al. 2019b; Ungvari et al. 2018b; Wiedenhoeft et al. 2019; Csiszar et al. 2009; Csiszar et al. 2007c). We found that DHE staining in the wall of aortas isolated from A-(A) mice was significantly stronger as compared with that in vessels isolated from Y-(Y) mice, indicating increased cellular ROS production (Fig. 1b). Aortas isolated from A-(Y) mice exhibited weaker DHE staining than vessels of A-(A) mice, indicating that the presence of young blood in old mice decreased vascular ROS production. In agreement with the vasorelaxation responses, aortas isolated from Y-(A) mice exhibited DHE staining that was comparable with that in aortas isolated from Y-(Y) mice (Fig. 1b), suggesting that the old environment per se, is not sufficient to provoke ROS production.
Young blood reverses age-related changes in vascular mRNA expression profile
Using RNA-seq we assessed transcriptomic changes in the aorta arch associated with aging and heterochronic parabiosis. In Fig. 2a, a heat map is shown as a graphic representation of normalized expression values of differentially expressed genes in aorta samples from each group. Hierarchical clustering analysis revealed groups of genes whose expression changes with age per se, and which is reversed by the presence of young blood in aged mice (A-(Y)). Principal component analysis (PCA; Fig. 2c) of the transcriptomic data showed that biological replicates from the Y-(Y) and A-(A) groups, respectively, cluster together and that young samples segregate away from aged ones (Fig. 2b). The profiles from A-(A) mice also cluster separately from clusters representing A-(Y) mice. The Y-(Y) and A-(Y) expression profiles were more similar and clustered less discriminately in the PCA. These findings indicated a clear difference between the transcriptome profiles of young and old isochronic pairs, and a partial reversal of the aging effect by young blood. Meanwhile, the impact of old blood on expression in young aorta was less clear as the Y-(A) group did not exhibit good clustering in the space of the first three principal components due to the presence of an outlier (Y-(A) sample #3).
Fig. 2.

Young blood reverses age-related changes in vascular mRNA expression profile. a The heat map is a graphic representation of normalized expression values of differentially expressed genes in aorta samples derived from isochronic parabiont young mice [Y-(Y)], isochronic parabiont aged mice [A-(A)], heterochronic parabiont young mice [Y-(A)] and heterochronic parabiont aged mice [A-(Y)]. Hierarchical clustering analysis reveals groups of genes whose expression is similar in young mice and young blood exposed aged mice. It was noted that the sample Y-(A)#3 is an outlier, yet the data were included in further analyses. b Principal component analysis (PCA) plot of aortic mRNA expression profiles in the four experimental groups. The profiles from A-(A) mice (red) cluster separately from clusters representing Y-(Y) mice (blue) and A-(Y) mice (green). The profiles of Y-(A) mice (yellow) did not exhibit good clustering in the space of the first three principal components. PC1, PC2 and PC3: Principal components 1, 2, and 3, respectively. c Volcano plot depicting differentially expressed genes comparing aortic samples derived from Y-(Y) and A-(A) mice. Stratified p values are plotted against expression fold changes for results obtained in A-(A) samples normalized to Y-(Y) samples. Colored points refer to genes whose expression is significantly altered in A-(Y) mice. d Venn diagrams sowing the numbers of differentially expressed mRNAs in A-(A) mice (“aging effect”) and A-(Y) mice (“young blood effect”)
We then determined the number of genes that were significantly upregulated or downregulated (DE; fold change ≥ 1.5 or < 0.67; p < 0.05 adjusted for multiple comparisons) in the aorta by aging or by heterochronic parabiosis. We then filtered for genes that are significantly altered (adjusted p < 0.05), expressed at an appreciable level (fragments per kilobase of transcript per million mapped reads > 1). We identified 347 differentially expressed (DE) genes in A-(A) animals compared with Y-(Y) controls. We further identified 58 DE genes in A-(Y) mice compared with A-(A) controls. In Fig. 2c, a volcano plot shows statistical significance (p value) versus magnitude of age-related change in gene expression. Red symbols denote genes, whose expression levels differed in the aged phenotype, but have shifted back toward the young phenotype by the presence of young blood in aged animals (discordant DE genes). The Venn diagram in Fig. 2d shows that expression of 24 genes, which are differentially expressed in aged mice, was shifted back toward youthful levels by the presence of young blood in aged mice.
We realized that significance cut-offs to identify DE genes shared between the age-effect and young blood–effect datasets may be too stringent and the analysis illustrated in Fig. 2c may miss discordant patterns (youthful shifts) of gene expression with important biological relevance for young blood–induced vascular rejuvenation. Thus, we also used an approach to detect discordant transcriptional patterns (youthful shifts) by comparing the age-effect and young blood–effect gene expression datasets using combination criteria that took into account the effect direction. Genes were ranked by their effect size direction, and ranked lists were compared to identify overlapping genes across a continuous significance gradient. Our analysis required that discordant genes with youthful shifts (1) are “differentially expressed” based on both p value and fold-change criteria either in aging or the young blood effect group, (2) satisfy a fold-change criterion with a cutoff of ≥ 1.5 or < 0.67 in the group in which expression did not satisfy the statistical significance p < 0.05, and (3) satisfy the criterion that the effect directions of the age-effect and young blood effect are opposite. We found that these combination criteria found more biologically meaningful sets of genes than p values alone.
In Fig. 3a, the magnitude of age-related changes in gene expression is plotted against the magnitude of young blood–induced changes in gene expression in the aged aorta. Red symbols denote the 24 discordant DE genes, whose expression levels shifted back toward the young phenotype by the presence of young blood in aged animals with statistical significance. Genes which are DE only in one group but otherwise satisfy the other criteria are denoted by blue (DE in aging) and green (DE in A-(Y)) symbols. The Venn diagram in Fig. 3c shows that using this approach we have identified 188 discordant genes with youthful shifts, which changed in opposite directions between the two datasets. These data suggest that changes in circulating anti-geronic factor(s) have a critical role in age-related dysregulation of vascular gene expression.
Fig. 3.

Young blood reverses age-related changes in vascular mRNA expression profile: identification of discordant genes. a Young blood–induced changes in gene expression (log2 fold changes; heterochronic parabiont aged [A-(Y)] mice vs. isochronic parabiont aged [A-(A)] mice) plotted against age-related changes (log2 fold changes; isochronic parabiont aged [A-(A)] mice vs. isochronic parabiont young [Y-(Y)] mice) in the aortic transcriptome. Red symbols indicate discordant differentially expressed genes, whose expression significantly changes with age and is restored by young blood toward youthful expression levels. Blue and green symbols denote discordant genes, whose expression changes in aging and is restored by young blood toward youthful expression levels, but only the aging (blue) or the young blood effect (green) reaches the cutoff for statistical significance. b Old blood-induced changes in gene expression (log2 fold changes; heterochronic parabiont young [Y-(A)] mice vs. isochronic parabiont young [Y-(Y)] mice) plotted against age-related changes (log2 fold changes; isochronic parabiont aged [A-(A)] mice vs. isochronic parabiont young [Y-(Y)] mice) in the aortic transcriptome. Red symbols indicate concordant differentially expressed genes, whose expression significantly changes with age and the direction of this effect is mimicked by exposure to old blood. Blue and green symbols denote concordant genes, whose expression similarly changes in aging and by old blood exposure, but only the aging (blue) or the old blood effect (green) reaches the cutoff for statistical significance. c Venn diagrams showing the numbers of differentially expressed mRNAs in each group. The blue circle represents genes, which are significantly up- or downregulated in aged mice as compared with young mice. The green circle represents genes, which are significantly up- or downregulated by the presence of young blood in aged mice. The red area represents discordant differentially expressed genes. Gray areas represent discordant genes, whose expression is changed by the presence of young blood in aged mice toward youthful levels, but the effect does not reach the cutoff for statistical significance. d Venn diagrams showing the effect of old blood on vascular gene expression. The blue circle represents genes, which are significantly up- or downregulated in aged mice as compared with young mice. Gray areas represent concordant genes, whose expression is changed by the presence of old blood in young mice toward the aging phenotype, but the effect does not reach the cutoff for statistical significance. The red area represents discordant differentially expressed genes. e The heat map is a graphic representation of normalized expression values of discordant genes in aorta samples derived from Y-(Y), A-(A) and A-(Y) mice
Using a similar approach, we also analyzed old blood-induced transcriptional changes in Y-(A) animals. In Fig. 3b, the magnitude of age-related changes in gene expression is plotted against the magnitude of old blood-induced changes in gene expression. Due to the presence of an outlier (Y-(A) sample #3) the analysis of the effects of the presence of old blood in young animals yielded fewer results. There was only one concordant DE gene (Prelid1; PRELI domain containing 1 pseudogene) whose expression was significantly upregulated by the presence of old blood in the young animals and whose expression was also significantly upregulated in aortas of A-(A) animals (Fig. 3b). Next, we used an approach to detect concordant transcriptional patterns (pro-geronic shifts) by comparing the age-effect and old blood-effect gene expression datasets using combination criteria that took into account the effect direction. Our analysis required that concordant genes with pro-geronic shifts (1) are “differentially expressed” based on both p value and fold-change criteria either in aging or the old blood effect group, (2) satisfy a fold-change criterion with a cutoff of ≥1.5 or < 0.67 in the group in which expression did not satisfy the statistical significance p < 0.05, and (3) satisfy the criterion that the effect directions of the age-effect and old blood effect are the same. Genes which are DE only in one group but otherwise satisfy the other criteria are denoted by blue in Fig. 3b. The Venn diagram in Fig. 3d shows that using this approach we have identified 114 concordant genes with pro-geronic shifts, which changed in the same directions between the two datasets. These data suggest that pro-geronic circulating factors may also have a role in age-related dysregulation of vascular gene expression, though these effects do not appear to compromise function in our model.
Young blood–induced vascular transcriptomic changes in aged mice predict mitochondrial rejuvenation
We performed GO enrichment analysis to explore potential biological functions of the young blood–induced discordant genes with youthful shifts. GO enrichment analysis of discordant genes with youthful shifts identified functions in mitochondrial regulation and production of ROS (Fig. 4a). Overrepresentation analysis of GO terms revealed that extracellular matrix related genes (including the GO terms “extracellular matrix” [GO:0031012] and “collagen-containing extracellular matrix” [GO:0062023]) were also overrepresented among the aging-induced DE genes, but these biological functions were unaffected by young blood. Figure 4 b depicts Reactome pathways, which are overrepresented (enriched) in the discordant genes induced by young blood exposure. This analysis also showed that exposure to young blood is associated with transcriptional changes in the aorta consistent with multifaceted mitochondrial protective effects as well as anti-inflammatory and anti-oxidative effects. Specifically, Reactome pathways identified included the Krebs cycle and oxidative phosphorylation as well as nonsense-mediated mRNA decay (NMD), a homeostatic mRNA quality maintenance system. Recently, the concept has emerged that activity of NMD decreases with aging, which may contribute to a decline in mRNA quality in older animals (Nguyen et al. 2018; Son and Lee 2017; Son et al. 2017; Tabrez et al. 2017).
Fig. 4.

Young blood reverses age-related changes in vascular expression of mitochondrion-related genes. a Most significantly enriched gene ontology (GO) terms for discordant genes induced by young blood exposure. b Reactome pathways, which are overrepresented (enriched) in the discordant genes induced by young blood exposure. Note that exposure to young blood is associated with transcriptional changes indicating multifaceted mitochondrial protective, anti-inflammatory and anti-oxidative effects. c Gene Set Enrichment Analysis (GSEA) to test the effect of aging on enrichment of the set of mitochondrion-targeted genes by comparing aorta samples derived from young mice (isochronic parabiont: young) and aged mice (isochronic parabiont: aged). Aging-induced gene expression changes were ranked from most upregulated (left, red) to most downregulated (right, green). Dots represent genes encoding mitochondrion-targeted proteins. Shown is a running-sum statistic (enrichment score) based on (e), increasing when a gene is a member of the mitochondrion-related gene set and decreasing when it is not. d GSEA showing the effect of exposure to young blood on enrichment of mitochondrion-targeted genes. Aorta samples derived from isochronic parabiont aged mice (parabiont: aged) and heterochronic parabiont aged mice (parabiont: young) were compared. Note that in aged mice enrichment scores increased predominantly on the right indicating downregulation of mitochondrion-related genes by aging. In contrast, in response to the presence of young blood in aged mice enrichment scores increased predominantly on the left indicating upregulation of mitochondrion-related genes. e The heat maps are graphic representations of normalized expression values of mitochondrion-related genes. Hierarchical clustering analysis revealed the similarities on aortic expression profiles of mitochondrion-related genes in young mice and young blood exposed aged mice. Note that expression of ~ 75% of mitochondrion-related genes are downregulated in aging, of which ~ 81% are restored toward youthful expression levels by the presence of young blood in aged animals. f, g KEGG pathway diagrams for oxidative phosphorylation (f) and TCA cycle (tricarboxylic acid cycle, g). Oxidative phosphorylation and the TCA (Krebs) cycle emerged as one of the most significantly enriched pathways for discordant genes induced by young blood exposure. The upper part of the figure in (f) displays the respiratory chain complexes and the lower portion represents their subunits as rectangles. Aging- (left side) and young blood–induced (right side) expressional changes are shown
Previous studies demonstrate that aging is associated with mitochondrial dysfunction and mitochondrial oxidative stress in endothelial and smooth muscle cells, which play a critical role in the genesis of endothelial dysfunction and vascular diseases (Tarantini et al. 2018; Csiszar et al. 2014; Ungvari et al. 2007; Csiszar et al. 2012; Tarantini et al. 2019b; Csiszar et al. 2019a; Dai et al. 2012; Springo et al. 2015). To elucidate how mitochondrion-related gene expression is altered by circulating factors present in young blood we analyzed expression of mitochondrion-related genes. We have used the existing database of the Broad Institute (MitoCarta2.0, accessed at: https://www.broadinstitute.org/files/shared/metabolism/mitocarta/mouse.mitocarta.2.0.html on 2019/11/01) to compile a list of genes with mitochondrial targeting sequences (; accessed on 2019/11/01) and known functions related to regulation of mitochondrial processes. We used Gene Set Enrichment Analysis (GSEA) for interpreting expression of mitochondrion-related genes (Subramanian et al. 2005). GSEA of mitochondrion-related genes was performed using a pre-ranked gene list based on the magnitude of the fold change (largest upregulation to most downregulated; Fig. 4c, d). Figure 4 c and d depict a running-sum statistic (enrichment score) based on Fig. 4, increasing when a gene is a member of the mitochondrion-related gene set and decreasing when it is not. Note that in aged mice enrichment scores increased predominantly on the right indicating downregulation of mitochondrion-related genes by aging. In contrast, in young blood exposed aged mice (A-(Y)) enrichment scores increased predominantly on the left indicating upregulation of mitochondrion-related genes by the presence of young blood in aortas of aged mice. The heatmap showing the expression pattern of mitochondrion-related genes is shown in Fig. 4e. Hierarchical clustering of the data showed that expression of ~ 40% of the mitochondrion-related genes tends to be downregulated in aging and these changes also tend to be returned toward youthful levels by the presence of young blood in the aged A-(Y) animals. Oxidative phosphorylation and the tricarboxylic acid (TCA; Krebs) cycle emerged as the most significantly enriched mitochondrial pathways for discordant genes induced by young blood exposure. KEGG pathway maps depicting age-related and young blood–induced changes in the expression of genes in the TCA cycle and oxidative phosphorylation pathways are shown in Fig. 4 f and g, respectively. These findings are consistent with the idea that young blood elicits mitochondrial rejuvenation in the aged vasculature.
Ingenuity upstream regulator analysis
We have performed IPA Upstream Regulator analysis (Kramer et al. 2014) to identify upstream transcriptional regulators that may contribute to the observed transcriptomic changes in our dataset, which can help to identify the mechanism of action of anti-geronic factors present in the young blood. The upstream regulator analysis is based on information in the Ingenuity Knowledge Base (a curated relational database of the available biomedical literature) on the expected effects between transcriptional regulators and their target genes. Using the IPA Upstream Regulator analysis it was examined how many known targets of each transcriptional regulator were differentially expressed in our samples and the direction of these gene expression changes were compared with what is expected from the literature. On the basis of the observed direction of change a prediction of the activation state of the predicted transcriptional regulators (“activated” or “inhibited”) was made (not shown). For each potential transcriptional regulator two statistical measures, an overlap p value and an activation z-score were computed. The overlap p value calls likely upstream regulators based on significant overlap between the DE genes and known targets regulated by that particular transcriptional regulator. The activation z-score is used to infer the activation state of the predicted transcriptional regulators (“activated” or “inhibited”) based on comparison with a model that assigns random regulation directions. The results of the IPA Upstream Regulator analysis are shown in Table 1. In particular, the IPA Upstream Regulator analysis predicts that young blood–induced vascular rejuvenation is associated with activation of PGC-1α (PPARGC1A) and IGF1R-mediated pathways and inhibition of mTOR signaling (Table 1).
Table 1.
Predicted upstream transcriptional regulators that may mediate the observed vasoprotective effects of circulating anti-geronic factors present in young blood. Results of the IPA Upstream Regulator analysis. Shown are predicted upstream transcriptional regulators that may contribute to the observed young blood–induced transcriptomic changes in our dataset. The overlap p value calls likely upstream regulators based on significant overlap between dataset genes and known targets regulated by a transcriptional regulator
| Upstream regulator | Name | Remark | Predicted activation state | Activation z-score | p value of overlap |
|---|---|---|---|---|---|
| RICTOR | Rapamycin-insensitive companion of mTOR | Subunit of mTORC2; regulates cell growth and survival in response to hormonal signals. Promotes inflammation, (Bhattacharya et al. 2013) atherosclerosis, (Babaev et al. 2018) and aneurysm formation (Parker et al. 2018); | Inhibited | − 3.873 | 5.8E−18 |
| KDM5A | lysine [K]-specific demethylase 5A | Histone demetylase, repressor of metabolic regulatory genes (Varaljai et al. 2015) | Inhibited | − 3 | 6.62E−12 |
| STK11 | Serine/threonine kinase 11 | Activates AMPK; regulates autophagy, energy metabolism; tumor suppressor; | Activated | 2.828 | 6.11E−09 |
| LMX1A | LIM homeobox transcription factor 1α | Regulates mitochondrial functions (Doucet-Beaupre et al. 2016) | Activated | 2 | 1.23E−08 |
| LMX1B | LIM homeobox transcription factor 1β | Regulates mitochondrial functions (Doucet-Beaupre et al. 2016) | Activated | 2 | 0.00000015 |
| PPARGC1A | PPARG coactivator 1 alpha; PGC-1α | Regulates the genes involved in energy metabolism; interacts with PPARγ; regulates the activities of CREB and nuclear respiratory factors; regulates mitochondrial biogenesis in endothelial cells (Patten and Arany 2012); | Activated | 2.789 | 3.36E−08 |
| RB1 | Retinoblastoma protein | Tumor suppressor; controls cell cycle progression and senescence | Activated | 3 | 0.000000256 |
| CAB39L | Calcium-binding protein 39-like | Activates AMPK, regulates mTOR pathway | Activated | 2 | 0.00000122 |
| MAP4K4 | Mitogen-activated protein kinase kinase kinase kinase 4 | Abundantly expressed in endothelial cells; regulates cell migration, proliferation and adhesion, and inflammatory processes; its ablation inhibits atherogenesis (Roth Flach et al. 2015) | Inhibited | − 2.236 | 0.00000491 |
| NRIP1 | Nuclear receptor-interacting protein 1 | Modulates hormone-dependent activation of nuclear receptors; coactivatot of NF-kB | Inhibited | − 2 | 0.00000831 |
| IGF1R | IGF-1 receptor | IGF-1 declines in aging; decline in circulating IGF-1 or knockdown of IGF1R in endothelial and smooth muscle cells mimics vascular aging phenotypes (Ashpole et al. 2017; Tarantini et al. 2017; Tarantini et al. 2016a; Toth et al. 2015b; Toth et al. 2014b; Sonntag et al. 2013; Bailey-Downs et al. 2012a; Fulop et al. 2019; Tarantini et al. 2016b) | Activated | 2.449 | 0.0000207 |
| ESRRA | Estrogen-related receptor alpha | Regulates mitochondrial function | Activated | 2 | 0.0000766 |
| PITX2 | Paired-like homeodomain 2 | Transcription factor; regulates Wnt-mediated beta catenin signaling; regulates differentiation of smooth muscle cells in aorta (Liu et al. 2002; Shang et al. 2008) | Activated | 2 | 0.000136 |
Effect of young blood on transcriptional footprint of shared mechanisms of vascular aging
Restoration of cellular NAD+ levels by treatment with the NAD+ precursor nicotinamide mononucleotide (NMN) (Tarantini et al. 2019b; Kiss et al. 2019a; Kiss et al. 2019b; Das et al. 2018; Csiszar et al. 2019b), activation of SIRT-1(Gano et al. 2014) and inhibition of mTOR pathways (Lesniewski et al. 2017; Lin et al. 2013) were shown to reverse vascular aging phenotypes. To investigate the contribution of these mechanisms to the vascular rejuvenation induced by young blood we analyzed expression of known NMN-regulated genes (Fig. 5a), SIRT-1-dependent and PGC1α-regulated genes (Fig. 5b, c) and mTOR-regulated genes (Fig. 5d). NMN-regulated genes were identified based on their differential expression in response to restoration of cellular NAD+ levels by treatment with the NAD+ precursor nicotinamide mononucleotide (NMN) in aged cerebromicrovascular endothelial cells (Kiss et al. 2020). SIRT1 dependent genes were identified based on their differential expression in the heart of mice with cardiac-specific overexpression of Sirt1 (Oka et al. 2011) and the brain of Sirt1−/− mice (Libert et al. 2011). The lists of PGC-1α- and mTOR-regulated genes were retrieved from Ingenuity Knowledge Base. Hierarchical clustering analysis revealed only weak similarities on aortic expression profiles of NMN-regulated, SIRT1- and PGC-1α-dependent, and mTOR-regulated genes in young mice and aged mice exposed to young blood (red box). The ratio of discordant genes within the gene list was 9.5% (NMN-regulated genes), ~ 3.6% (SIRT-1-regulated genes), 8.5% (PGC-1α-regulated genes), and 3.4% (mTOR-regulated genes).
Fig. 5.

Effect of young blood on transcriptional footprint of shared mechanisms of vascular aging. Restoration of cellular NAD+ levels by treatment with the NAD+ precursor nicotinamide mononucleotide (NMN) (Tarantini et al. 2019b; Kiss et al. 2019a; Kiss et al. 2019b; Das et al. 2018; Csiszar et al. 2019b), activation of SIRT-1 (Gano et al. 2014) and inhibition of mTOR pathways(Lesniewski et al. 2017; Lin et al. 2013) were shown to reverse vascular aging phenotypes. The heat maps are graphic representation of normalized expression values of NMN-regulated genes (a), SIRT-1-dependent genes (b), PGC-1α-regulated genes (c) and mTOR-regulated genes (d) in aorta samples derived from isochronic parabiont young mice (parabiont: young), isochronic parabiont aged mice (parabiont: aged) and heterochronic parabiont aged mice (parabiont: young). NMN-regulated genes were identified based on their differential expression in response to restoration of cellular NAD+ levels by treatment with the NAD+ precursor nicotinamide mononucleotide (NMN) in aged cerebromicrovascular endothelial cells (Kiss et al. 2020). SIRT1 dependent genes were identified based on their differential expression in the heart of mice with cardiac-specific overexpression of Sirt1 (b) (Oka et al. 2011) and the brain of Sirt1−/− mice (Libert et al. 2011) (b, right). The lists of PGC-1α- and mTOR-regulated genes were retrieved from Ingenuity Knowledge Base (c, d). Hierarchical clustering analysis revealed the only weak similarities on aortic expression profiles of NMN-regulated, SIRT1- and PGC-1α-dependent and mTOR-regulated genes in young mice and aged mice exposed to young blood (red box). The ratio of discordant genes within the gene list was 9.5% (NMN-regulated genes), ~3.6% (SIRT-1 regulated genes), 8.5% (PGC-1α regulated genes) and 3.4% (mTOR-regulated genes)
Effect of young blood on the transcriptional footprint of signaling pathways activated by putative circulating anti-geronic factors in the vasculature
IGF-1 (insulin-like growth factor-1), GDF11 (growth differentiation factor 11; also known as bone morphogenetic protein 11 or BMP11), TGFβ (transforming growth factor-β), and oxytocin (Elabd et al. 2014) were suggested to act as circulating factors regulating aging phenotypes in a context-dependent manner (Ungvari et al. 2018a). IGF-1 has clearly established anti-aging effects in many tissues, including the vasculature, myocardium, and brain, which are unrelated to the effects of developmental IGF-1 deficiency on lifespan (Ashpole et al. 2016; Ashpole et al. 2017; Tarantini et al. 2017; Tarantini et al. 2016a; Toth et al. 2015b; Toth et al. 2014b; Sonntag et al. 2013; Bailey-Downs et al. 2012a; Mitschelen et al. 2011; Colon et al. 2019). To determine the extent, if any, to which changes in the aforementioned factors contribute to vascular rejuvenation through heterochronic parabiosis, we analyzed changes in the expression of known downstream targets of IGF-1, GDF11, TGFβ, and oxytocin. Figure 6 shows heatmaps representing normalized expression values of genes known to be regulated by IGF-1 via IGF1R activation (Fig. 6a), GDF11 (Fig. 6b), TGFβ (Fig. 6c), activation of SMAD2 and SMAD3 signaling (Fig. 6d, e,), and oxytocin (Fig. 6f). Hierarchical clustering analysis revealed weak similarities on aortic expression profiles of specific gene clusters known to be regulated by IGF-1, GDF11, TGFβ, and oxytocin, respectively, in Y-(Y) mice and young blood exposed A-(Y) mice.
Fig. 6.

Effect of young blood on transcriptional footprint of signaling pathways activated by putative circulating anti-geronic factors. IGF-1, GDF11 (Katsimpardi et al. 2014; Katsimpardi et al. 2020), TGFβ and oxytocin (Elabd et al. 2014) were suggested to act as circulating factors modulating cellular aging phenotypes. The heat maps are graphic representation of normalized expression values of genes regulated by IGF-1 via IGF1R activation (a), GDF11 (b), TGFβ (c), activation of SMAD2 and SMAD3 signaling (d, e) and oxytocin in aorta samples derived from isochronic parabiont young mice (parabiont: young), isochronic parabiont aged mice (parabiont: aged) and heterochronic parabiont aged mice (parabiont: young). Gene lists were retrieved from Ingenuity Knowledge Base. Hierarchical clustering analysis revealed the similarities on aortic expression profiles of genes regulated by circulating anti-geronic factors in young mice and aged mice exposed to young blood (red box)
Discussion
The key finding of this study is that a relatively short-term exposure to a young humoral environment can rescue vascular aging phenotypes, including opposing increased oxidative stress, mitochondrial impairment, and consequential endothelial dysfunction as well as dysregulation of vascular gene expression in a mouse model of aging that recapitulates key aspects of age-related vascular dysfunction manifested in elderly patients.
Elucidating the mechanisms by which aging impairs vascular responses and their potential reversal are critical for the development of new targets and effective therapies for prevention and treatment of cardiovascular and cerebrovascular diseases in older adults. Here, we show for the first time that circulating factors present in the blood of young mice rescue NO-mediated endothelium-dependent vasodilation and attenuate vascular oxidative stress in aged mice. Previous studies also demonstrated that presence of young blood in the circulation of aged heterochronic parabionts also reverses age-related cerebromicrovascular rarefaction (Katsimpardi et al. 2014). In that regard, it is also important that circulating non-cellular factors present in sera derived from caloric restricted non-human primates was also shown to improve endothelial angiogenic processes in cultured endothelial cells (Csiszar et al. 2013). The presence of young blood in the circulation of aged heterochronic parabionts (Fig. 7) also reversed select transcriptional changes of aging in the aorta supporting the concept that non-cell autonomous mechanisms play important roles in driving vascular aging processes and pathogenesis of age-related cardiovascular diseases (Ungvari et al. 2018a). The remarkable level of malleability of vascular aging phenotypes in response to anti-geronic circulating factors highlight the potential for therapeutic interventions to functionally reverse the deleterious effects of aging in the circulatory system via targeting the systemic milieu or the endothelium directly to prevent cardiovascular morbidity and mortality.
Fig. 7.
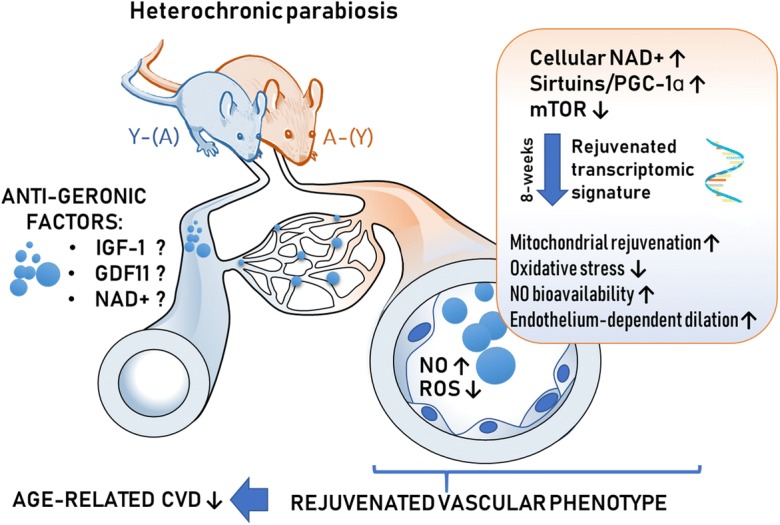
Proposed scheme for the mechanisms contributing to young blood–mediated vascular rejuvenation in aged heterochonic parabionts. The model, based on our present and previous findings and earlier data from the literature (Ungvari et al. 2018a; Tarantini et al. 2018; Tarantini et al. 2019b; Csiszar et al. 2019b; Van Skike et al. 2018), predicts that circulating anti-geronic factors present in young blood (derived from heterochronic parabiont young mice [Y-(A)]) restore cellular NAD+, activate sirtuins/PGC-1α and/or inhibit mTOR mediated pathways, promoting transcriptomic changes, which rescue a youthful vascular phenotype in and heterochronic parabiont aged mice [A-(Y)]. Young blood–mediated vascular rejuvenation is associated with restored mitochondrial function, attenuation of cellular and mitochondrial ROS production, increased bioavailability of NO and improved endothelium-mediated vasodilation. All of these effects are predicted to act to improve vascular health, preventing the pathogenesis of age-related vascular diseases. CVD: cardiovascular diseases; Y-(A)
Mitochondrial dysregulation plays a central role in the genesis of vascular aging phenotypes, including increases in oxidative stress and endothelial dysfunction (Tarantini et al. 2018; Csiszar et al. 2014; Ungvari et al. 2007; Springo et al. 2015; Ungvari et al. 2008). In the aged vasculature, the efficacy of the respiratory chain diminishes due to dysregulated expression of electron transport chain subunits, promoting electron leakage, and increased mitochondrial ROS (mtROS) production and reducing cellular ATP generation (Tarantini et al. 2019b). Importantly, treatments that restore mitochondrial gene expression and/or normalize mtROS production in the vasculature have been shown to improve endothelial function in rodent models of aging (Tarantini et al. 2018; Gioscia-Ryan et al. 2014; Tarantini et al. 2019b; Pearson et al. 2008). Here, we report that circulating factors present in the blood of young mice rescue aging-induced changes in mitochondrion-related gene expression in the aorta of aged parabionts. On the basis of previous findings, (Gomes et al. 2013) we posit that rescue of vascular mitochondrial function by restoring the expression of ETC subunits contributes to the endothelial protective effects of young blood. In addition, upregulation of enzymes involved in the TCA cycle and antioxidant defenses may also contribute to young blood–mediated vasoprotection.
Importantly, endothelial cells come in contact with each plasma constituent (including hormones, proteins, peptides, lipid mediators, micropeptides, metabolites, and circulating exosomes) as well as circulating cellular factors, all of which may exert important vasoprotective effects. Interestingly, circulating non-cellular factors present in the young blood were also shown to confer rejuvenating effects on the central nervous system of aged mice after intravenous delivery, including an increased neurogenesis, improved learning and memory, and attenuated neuro-inflammation (Wyss-Coray 2016), suggesting that key circulating anti-geronic factor(s) may penetrate the blood-brain barrier and/or exert their neuroprotective effects by rejuvenating the microcirculation.
The exact nature and the cellular origins of circulating anti-geronic factors responsible for the vascular rejuvenating effects observed in our studies remain obscure. Recent evidence supports the concept that aging is associated with cellular NAD+ depletion (Gomes et al. 2013; Massudi et al. 2012; Yoshino et al. 2018) and consequential SIRT1 dysfunction, which are critical driving forces of vascular aging processes contributing to increased oxidative stress and the genesis of endothelial dysfunction (Tarantini et al. 2019b; Kiss et al. 2019a; Kiss et al. 2019b). Another evolutionarily conserved fundamental mechanism involved in vascular aging and the endothelial dysfunction is mTOR (mechanistic/mammalian target of rapamycin) activation that governs cellular responses to nutrient availability and growth signals controlling biosynthesis of proteins, lipids, and nucleic acids (Lin et al. 2013; Parlar et al. 2010; Cheng et al. 2008; Corbin et al. 1994; Milliard et al. 1998). Accordingly, restoration of cellular NAD+ levels by treatment with NMN (Tarantini et al. 2019b; Kiss et al. 2019a; Kiss et al. 2019b; Das et al. 2018; Csiszar et al. 2019b), activation of SIRT1,(Gano et al. 2014) and inhibition of mTOR pathways (Lesniewski et al. 2017; Lin et al. 2013; Van Skike et al. 2018) were shown to reverse vascular aging phenotypes and improve endothelial function. Heterochoronic parabiosis is expected to increase levels of NAD+ precursors in the circulation, and consequently restore NAD+ levels in the vasculature, of aged parabionts. There are known circulating anti-geronic factors (e.g., induced by caloric restriction) that activate SIRT1 (Csiszar et al. 2009; Csiszar et al. 2013; de Cabo et al. 2003) and its downstream target PGC-1α and/or regulate mTOR. Transcriptomic analysis of the aortas suggests that these pathways may contribute to vascular rejuvenation induced by young blood. Further studies should investigate in detail the effects of heterochronic parabiosis or systemic administration of young plasma to aged mice on cellular NAD+ levels and SIRT1 and mTOR activities in the vasculature. In that regard, it is interesting that a recent study demonstrates that age-related decline in NAD+ levels in red blood cells is not corrected by heterochronic parabiosis (Morrison et al. 2019). The effects of young blood on vascular aging phenotypes should be also investigated in mice with genetic depletion of SIRT1.
Previous studies identified age-related changes in circulating anti-geronic factors (e.g., IGF-1 (Ashpole et al. 2016; Ashpole et al. 2017; Tarantini et al. 2017; Toth et al. 2015b; Toth et al. 2014b; Bailey-Downs et al. 2012a; Mitschelen et al. 2011; Bailey-Downs et al. 2012b; Fulop et al. 2019; Tarantini et al. 2016b; Ungvari and Csiszar 2012), which can be causally linked to the development of aging phenotypes in multiple organ systems. Importantly, decreases in circulating levels of IGF-1 (Tarantini et al. 2016b) were shown to contribute to key aspects of vascular aging, including endothelial dysfunction (Toth et al. 2015b), impaired autoregulation of cerebral blood flow (Toth et al. 2014b), pathological vascular remodeling, atherogenesis (Higashi et al. 2012), impaired vascular stress resilience (Bailey-Downs et al. 2012a; Bailey-Downs et al. 2012b), impaired angiogenic processes, and microvascular rarefaction (Lahteenvuo and Rosenzweig 2012). In the present study, IGF1R signaling was identified as a likely upstream regulator involved in young blood–mediated vascular rejuvenation. Future studies should further interrogate experimentally the role of circulating IGF-1 in mediation of the vasoprotective effects of young blood transfer (e.g., using animal models with endothelium-specific knockdown of IGF1R as parabionts).
Other circulating factors, whose levels are rescued in aged mice by heterochronic parabiosis or systemic administration of young plasma and which may confer rejuvenating effects in multiple organs (e.g., brain, heart, and skeletal muscle (Katsimpardi et al. 2014; Katsimpardi et al. 2020; Loffredo et al. 2013; Poggioli et al. 2016; Sinha et al. 2014)) include GDF-11, a member of the TGF-β superfamily of growth factors (Katoh and Katoh 2006). GDF11 is known to activate SMAD2/SMAD3 signaling in endothelial cells (Poggioli et al. 2016; Finkenzeller et al. 2015). However, transcriptomic analysis of the aortas did not confirm that GDF11/SMAD2/3 signaling plays a major role in young blood–mediated vascular rejuvenation. Importantly, the possible rejuvenating effects of GDF11 were also questioned by several recent studies (Egerman et al. 2015; Rodgers and Eldridge 2015; Smith et al. 2015b). It is of note that the pleiotropic growth factor TGFβ also induces SMAD2/SMAD3 (Nakao et al. 1997) and that increased TGFβ signaling has been linked to the genesis of vascular aging phenotypes (Gaertner et al. 2005; Masliah et al. 2001; Wyss-Coray et al. 1997a; Wyss-Coray et al. 1995; Wyss-Coray et al. 2000; Wyss-Coray et al. 1997b). Oxytocin was also proposed to mediate rejuvenating effects in the aged skeletal muscle (Elabd et al. 2014), yet it is unlikely that it plays an important role in young blood–induced changes in the vascular transcriptome.
Perspectives
Our findings provide additional evidence supporting the significant plasticity of vascular aging and its amenability to modulation by circulating anti-geronic factors. Future studies will be necessary to mechanistically expand on the present observations, including investigations aimed at defining the exact nature of the non-autonomous factors that drive vascular aging. The role of age-related changes in circulating hormones, cytokines, and growth factors in development of vascular aging phenotypes should be methodologically elucidated. Studies using mass spectrometry–based approaches led to the identification of complex age-related changes in circulating metabolites, proteins (Lehallier et al. 2019), micropeptides, and lipid mediators. Subsequent studies should identify changes that are not strain-, species-, or sex-dependent and elucidate their role in modulating vascular health. Further studies are also needed to identify the critical organ(s), tissues, and cell types that contribute to the regulation of vascular aging processes via circulating factors. It should be noted that in heterochronic parabionts all the organs are shared via the conjoined circulation. In addition to the brain (e.g., by releasing hypothalamic hormones and peptides and neuroendocrine factors) and endocrine organs, aged mice can benefit from access to the young bone marrow, liver, lungs, immune system, adipose tissue, skeletal muscle, heart, kidneys, and the gastrointestinal system and microbiome. In addition to the decline in anti-geronic factors, pro-geronic circulating factors, whose levels increase with age, may also contribute to the genesis of vascular aging phenotypes (Rebo et al. 2016). For example, there is strong evidence that TNFα(Csiszar et al. 2007c) and other cytokines (e.g., CCL11, TGFβ (Masliah et al. 2001) may serve as pro-geronic factors. Importantly, many of these factors are secreted by senescent cells in distant organs (e.g., the adipose tissue (Baker et al. 2011; Tchkonia et al. 2013). Further analysis of our dataset may lead to the identification of important circulating pro-geronic factors that contribute to the induction of aging phenotypes in young parabionts. It will also be important to determine which of the non-autonomous changes observed in the vascular tissues are actually causative of age-related vascular pathologies. Targeting these mechanisms might offer potential new treatments for prevention of age-related vascular diseases.
Funding information
This work was supported by grants from the American Heart Association (ST), the Oklahoma Center for the Advancement of Science and Technology (to AC, AY, ZU), the National Institute on Aging (R01-AG047879; R01-AG038747; R01-AG055395), the National Institute of Neurological Disorders and Stroke (NINDS; R01-NS056218 to AC, R01-NS100782 to ZU), the Oklahoma Shared Clinical and Translational Resources (OSCTR) program funded by the National Institute of General Medical Sciences (GM104938, to AY and JW), the Presbyterian Health Foundation (to ZU, AC, AY), the NIA-supported Geroscience Training Program in Oklahoma (T32AG052363), the Oklahoma Nathan Shock Center (P30AG050911), and the Cellular and Molecular GeroScience CoBRE (1P20GM125528, sub#5337). DMH is supported by R21AG055026, the American Federation for Aging Research (AFAR) and the Einstein Nathan Shock Center (P30AG038072). The funding sources had no role in the study design; in the collection, analysis, and interpretation of data; in the writing of the report; and in the decision to submit the article for publication.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Tamas Kiss, Stefano Tarantini, Tamas Csipo, Priya Balasubramanian and Ádám Nyúl-Tóth contributed equally to this work.
Contributor Information
Derek M. Huffman, Email: derek.huffman@einstein.yu.edu
Zoltan Ungvari, Email: zoltan-ungvari@ouhsc.edu.
References
- Ashpole NM, Herron JC, Mitschelen MC, Farley JA, Logan S, Yan H, et al. IGF-1 regulates vertebral bone aging through sex-specific and time-dependent mechanisms. J Bone Miner Res. 2016;31(2):443–54. doi: 10.1002/jbmr.2689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashpole NM, Logan S, Yabluchanskiy A, Mitschelen MC, Yan H, Farley JA, Hodges EL, Ungvari Z, Csiszar A, Chen S, Georgescu C, Hubbard GB, Ikeno Y, Sonntag WE. IGF-1 has sexually dimorphic, pleiotropic, and time-dependent effects on healthspan, pathology, and lifespan. Geroscience. 2017;39:129–145. doi: 10.1007/s11357-017-9971-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Babaev VR, Huang J, Ding L, Zhang Y, May JM, Linton MF. Loss of Rictor in monocyte/macrophages suppresses their proliferation and viability reducing atherosclerosis in LDLR null mice. Front Immunol. 2018;9:215. doi: 10.3389/fimmu.2018.00215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailey-Downs LC, Mitschelen M, Sosnowska D, Toth P, Pinto JT, Ballabh P, Valcarcel-Ares MN, Farley J, Koller A, Henthorn JC, Bass C, Sonntag WE, Ungvari Z, Csiszar A. Liver-specific knockdown of IGF-1 decreases vascular oxidative stress resistance by impairing the Nrf2-dependent antioxidant response: a novel model of vascular aging. J Gerontol A Biol Sci Med Sci. 2012;67:313–329. doi: 10.1093/gerona/glr164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailey-Downs LC, Sosnowska D, Toth P, Mitschelen M, Gautam T, Henthorn JC, Ballabh P, Koller A, Farley JA, Sonntag WE, Csiszar A, Ungvari Z. Growth hormone and IGF-1 deficiency exacerbate high-fat diet-induced endothelial impairment in obese Lewis dwarf rats: implications for vascular aging. J Gerontol A Biol Sci Med Sci. 2012;67:553–564. doi: 10.1093/gerona/glr197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker DJ, Wijshake T, Tchkonia T, LeBrasseur NK, Childs BG, van de Sluis B, Kirkland JL, van Deursen JM. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature. 2011;479:232–236. doi: 10.1038/nature10600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhattacharya I, Dragert K, Albert V, Contassot E, Damjanovic M, Hagiwara A, Zimmerli L, Humar R, Hall MN, Battegay EJ, Haas E. Rictor in perivascular adipose tissue controls vascular function by regulating inflammatory molecule expression. Arterioscler Thromb Vasc Biol. 2013;33:2105–2111. doi: 10.1161/ATVBAHA.112.301001. [DOI] [PubMed] [Google Scholar]
- Bitto A, Kaeberlein M. Rejuvenation: it's in our blood. Cell Metab. 2014;20:2–4. doi: 10.1016/j.cmet.2014.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolger AM, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics. 2014;30:2114–2120. doi: 10.1093/bioinformatics/btu170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bray NL, Pimentel H, Melsted P, Pachter L. Near-optimal probabilistic RNA-seq quantification. Nat Biotechnol. 2016;34:525–527. doi: 10.1038/nbt.3519. [DOI] [PubMed] [Google Scholar]
- Cannata A, Marcon G, Cimmino G, Camparini L, Ciucci G, Sinagra G, Loffredo FS. Role of circulating factors in cardiac aging. J Thorac Dis. 2017;9:S17–S29. doi: 10.21037/jtd.2017.03.95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castellano JM, Palner M, Li SB, Freeman GM, Jr, Nguyen A, Shen B, Stan T, Mosher KI, Chin FT, de Lecea L, Luo J, Wyss-Coray T. In vivo assessment of behavioral recovery and circulatory exchange in the peritoneal parabiosis model. Sci Rep. 2016;6:29015. doi: 10.1038/srep29015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng C, Tempel D, Oostlander A, Helderman F, Gijsen F, Wentzel J, van Haperen R, Haitsma DB, Serruys PW, van der Steen AF, de Crom R, Krams R. Rapamycin modulates the eNOS vs. shear stress relationship. Cardiovasc Res. 2008;78:123–129. doi: 10.1093/cvr/cvm103. [DOI] [PubMed] [Google Scholar]
- Coleman DL. Effects of parabiosis of obese with diabetes and normal mice. Diabetologia. 1973;9:294–298. doi: 10.1007/BF01221857. [DOI] [PubMed] [Google Scholar]
- Coleman DL, Hummel KP. Effects of parabiosis of normal with genetically diabetic mice. Am J Phys. 1969;217:1298–1304. doi: 10.1152/ajplegacy.1969.217.5.1298. [DOI] [PubMed] [Google Scholar]
- Colon G, Saccon T, Schneider A, Cavalcante MB, Huffman DM, Berryman D, List E, Ikeno Y, Musi N, Bartke A, Kopchick J, Kirkland JL, Tchkonia T, Masternak MM. The enigmatic role of growth hormone in age-related diseases, cognition, and longevity. Geroscience. 2019;41:759–774. doi: 10.1007/s11357-019-00096-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conboy MJ, Conboy IM, Rando TA. Heterochronic parabiosis: historical perspective and methodological considerations for studies of aging and longevity. Aging Cell. 2013;12:525–530. doi: 10.1111/acel.12065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corbin F, Blaise GA, Parent M, Chen H, Daloze PM. Effect of rapamycin on rat aortic ring vasomotion. J Cardiovasc Pharmacol. 1994;24:813–817. doi: 10.1097/00005344-199424050-00017. [DOI] [PubMed] [Google Scholar]
- Csiszar A, Ungvari Z, Edwards JG, Kaminski PM, Wolin MS, Koller A, Kaley G. Aging-induced phenotypic changes and oxidative stress impair coronary arteriolar function. Circ Res. 2002;90:1159–1166. doi: 10.1161/01.res.0000020401.61826.ea. [DOI] [PubMed] [Google Scholar]
- Csiszar A, Labinskyy N, Orosz Z, Xiangmin Z, Buffenstein R, Ungvari Z. Vascular aging in the longest-living rodent, the naked mole rat. Am J Phys. 2007;293:H919–H927. doi: 10.1152/ajpheart.01287.2006. [DOI] [PubMed] [Google Scholar]
- Csiszar A, Labinskyy N, Zhao X, Hu F, Serpillon S, Huang Z, Ballabh P, Levy RJ, Hintze TH, Wolin MS, Austad SN, Podlutsky A, Ungvari Z. Vascular superoxide and hydrogen peroxide production and oxidative stress resistance in two closely related rodent species with disparate longevity. Aging Cell. 2007;6:783–797. doi: 10.1111/j.1474-9726.2007.00339.x. [DOI] [PubMed] [Google Scholar]
- Csiszar A, Labinskyy N, Smith K, Rivera A, Orosz Z, Ungvari Z. Vasculoprotective effects of anti-TNFalfa treatment in aging. Am J Pathol. 2007;170:388–698. doi: 10.2353/ajpath.2007.060708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Csiszar A, Labinskyy N, Jimenez R, Pinto JT, Ballabh P, Losonczy G, Pearson KJ, de Cabo R, Ungvari Z. Anti-oxidative and anti-inflammatory vasoprotective effects of caloric restriction in aging: role of circulating factors and SIRT1. Mech Ageing Dev. 2009;130:518–527. doi: 10.1016/j.mad.2009.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Csiszar A, Sosnowska D, Wang M, Lakatta EG, Sonntag WE, Ungvari Z. Age-associated proinflammatory secretory phenotype in vascular smooth muscle cells from the non-human primate Macaca mulatta: reversal by resveratrol treatment. J Gerontol A Biol Sci Med Sci. 2012;67:811–820. doi: 10.1093/gerona/glr228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Csiszar A, Sosnowska D, Tucsek Z, Gautam T, Toth P, Losonczy G, Colman RJ, Weindruch R, Anderson RM, Sonntag WE, Ungvari Z. Circulating factors induced by caloric restriction in the nonhuman primate Macaca mulatta activate angiogenic processes in endothelial cells. J Gerontol A Biol Sci Med Sci. 2013;68:235–249. doi: 10.1093/gerona/gls158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Csiszar A, Gautam T, Sosnowska D, Tarantini S, Banki E, Tucsek Z, Toth P, Losonczy G, Koller A, Reglodi D, Giles CB, Wren JD, Sonntag WE, Ungvari Z. Caloric restriction confers persistent anti-oxidative, pro-angiogenic, and anti-inflammatory effects and promotes anti-aging miRNA expression profile in cerebromicrovascular endothelial cells of aged rats. Am J Physiol Heart Circ Physiol. 2014;307:H292–H306. doi: 10.1152/ajpheart.00307.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Csiszar A, Pinto JT, Gautam T, Kleusch C, Hoffmann B, Tucsek Z, et al. Resveratrol encapsulated in novel fusogenic liposomes activates Nrf2 and attenuates oxidative stress in cerebromicrovascular endothelial cells from aged rats. J Gerontol A Biol Sci Med Sci. 2015;70(3):303–13. doi: 10.1093/gerona/glu029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Csiszar A, Yabluchanskiy A, Ungvari A, Ungvari Z, Tarantini S. Overexpression of catalase targeted to mitochondria improves neurovascular coupling responses in aged mice. Geroscience. 2019;41:609–617. doi: 10.1007/s11357-019-00111-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Csiszar A, Tarantini S, Yabluchanskiy A, Balasubramanian P, Kiss T, Farkas E, et al. Role of endothelial NAD+ deficiency in age-related vascular dysfunction. Am J Physiol Heart Circ Physiol. 2019b; in press. [DOI] [PMC free article] [PubMed]
- Dai DF, Rabinovitch PS, Ungvari Z. Mitochondria and cardiovascular aging. Circ Res. 2012;110:1109–1124. doi: 10.1161/CIRCRESAHA.111.246140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das A, Huang GX, Bonkowski MS, Longchamp A, Li C, Schultz MB, Kim LJ, Osborne B, Joshi S, Lu Y, Trevino-Villarreal JH, Kang MJ, Hung TT, Lee B, Williams EO, Igarashi M, Mitchell JR, Wu LE, Turner N, Arany Z, Guarente L, Sinclair DA. Impairment of an endothelial NAD(+)-H2S signaling network is a reversible cause of vascular aging. Cell. 2018;173:74–89. doi: 10.1016/j.cell.2018.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Cabo R, Furer-Galban S, Anson RM, Gilman C, Gorospe M, Lane MA. An in vitro model of caloric restriction. Exp Gerontol. 2003;38:631–639. doi: 10.1016/s0531-5565(03)00055-x. [DOI] [PubMed] [Google Scholar]
- DeCarolis NA, Kirby ED, Wyss-Coray T, Palmer TD. The role of the microenvironmental niche in declining stem-cell functions associated with biological aging. Cold Spring Harb Perspect Med. 2015;5(12). 10.1101/cshperspect.a025874. [DOI] [PMC free article] [PubMed]
- Doucet-Beaupre H, Gilbert C, Profes MS, Chabrat A, Pacelli C, Giguere N, Rioux V, Charest J, Deng Q, Laguna A, Ericson J, Perlmann T, Ang SL, Cicchetti F, Parent M, Trudeau LE, Levesque M. Lmx1a and Lmx1b regulate mitochondrial functions and survival of adult midbrain dopaminergic neurons. Proc Natl Acad Sci U S A. 2016;113:E4387–E4396. doi: 10.1073/pnas.1520387113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durinck S, Spellman PT, Birney E, Huber W. Mapping identifiers for the integration of genomic datasets with the R/Bioconductor package biomaRt. Nat Protoc. 2009;4:1184–1191. doi: 10.1038/nprot.2009.97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egerman MA, Cadena SM, Gilbert JA, Meyer A, Nelson HN, Swalley SE, Mallozzi C, Jacobi C, Jennings LL, Clay I, Laurent G, Ma S, Brachat S, Lach-Trifilieff E, Shavlakadze T, Trendelenburg AU, Brack AS, Glass DJ. GDF11 increases with age and inhibits skeletal muscle regeneration. Cell Metab. 2015;22:164–174. doi: 10.1016/j.cmet.2015.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elabd C, Cousin W, Upadhyayula P, Chen RY, Chooljian MS, Li J, Kung S, Jiang KP, Conboy IM. Oxytocin is an age-specific circulating hormone that is necessary for muscle maintenance and regeneration. Nat Commun. 2014;5:4082. doi: 10.1038/ncomms5082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fabregat A, Jupe S, Matthews L, Sidiropoulos K, Gillespie M, Garapati P, Haw R, Jassal B, Korninger F, May B, Milacic M, Roca CD, Rothfels K, Sevilla C, Shamovsky V, Shorser S, Varusai T, Viteri G, Weiser J, Wu G, Stein L, Hermjakob H, D'Eustachio P. The reactome pathway knowledgebase. Nucleic Acids Res. 2018;46:D649–D655. doi: 10.1093/nar/gkx1132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falcon S, Gentleman R. Using GOstats to test gene lists for GO term association. Bioinformatics. 2007;23:257–258. doi: 10.1093/bioinformatics/btl567. [DOI] [PubMed] [Google Scholar]
- Fan X, Wheatley EG, Villeda SA. Mechanisms of hippocampal aging and the potential for rejuvenation. Annu Rev Neurosci. 2017;40:251–272. doi: 10.1146/annurev-neuro-072116-031357. [DOI] [PubMed] [Google Scholar]
- Finkenzeller G, Stark GB, Strassburg S. Growth differentiation factor 11 supports migration and sprouting of endothelial progenitor cells. J Surg Res. 2015;198:50–56. doi: 10.1016/j.jss.2015.05.001. [DOI] [PubMed] [Google Scholar]
- Flemming A. Cardiovascular disease: rejuvenating the ageing heart. Nat Rev Drug Discov. 2013;12:503. doi: 10.1038/nrd4064. [DOI] [PubMed] [Google Scholar]
- Fulop GA, Kiss T, Tarantini S, Balasubramanian P, Yabluchanskiy A, Farkas E, Bari F, Ungvari Z, Csiszar A. Nrf2 deficiency in aged mice exacerbates cellular senescence promoting cerebrovascular inflammation. Geroscience. 2018;40:513–521. doi: 10.1007/s11357-018-0047-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fulop GA, Ramirez-Perez FI, Kiss T, Tarantini S, Valcarcel Ares MN, Toth P, et al. IGF-1 deficiency promotes pathological remodeling of cerebral arteries: a potential mechanism contributing to the pathogenesis of Intracerebral hemorrhages in aging. J Gerontol A Biol Sci Med Sci. 2019;74(4):446–54. doi: 10.1093/gerona/gly144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaertner RF, Wyss-Coray T, Von Euw D, Lesne S, Vivien D, Lacombe P. Reduced brain tissue perfusion in TGF-beta 1 transgenic mice showing Alzheimer’s disease-like cerebrovascular abnormalities. Neurobiol Dis. 2005;19:38–46. doi: 10.1016/j.nbd.2004.11.008. [DOI] [PubMed] [Google Scholar]
- Gano LB, Donato AJ, Pasha HM, Hearon CM, Jr, Sindler AL, Seals DR. The SIRT1 activator SRT1720 reverses vascular endothelial dysfunction, excessive superoxide production, and inflammation with aging in mice. Am J Physiol Heart Circ Physiol. 2014;307:H1754–H1763. doi: 10.1152/ajpheart.00377.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghosh AK, O'Brien M, Mau T, Qi N, Yung R. Adipose tissue senescence and inflammation in aging is reversed by the young milieu. J Gerontol A Biol Sci Med Sci. 2019;74:1709–1715. doi: 10.1093/gerona/gly290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gioscia-Ryan RA, LaRocca TJ, Sindler AL, Zigler MC, Murphy MP, Seals DR. Mitochondria-targeted antioxidant (MitoQ) ameliorates age-related arterial endothelial dysfunction in mice. J Physiol. 2014;592:2549–2561. doi: 10.1113/jphysiol.2013.268680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomes AP, Price NL, Ling AJ, Moslehi JJ, Montgomery MK, Rajman L, White JP, Teodoro JS, Wrann CD, Hubbard BP, Mercken EM, Palmeira CM, de Cabo R, Rolo AP, Turner N, Bell EL, Sinclair DA. Declining NAD(+) induces a pseudohypoxic state disrupting nuclear-mitochondrial communication during aging. Cell. 2013;155:1624–1638. doi: 10.1016/j.cell.2013.11.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gontier G, Iyer M, Shea JM, Bieri G, Wheatley EG, Ramalho-Santos M, Villeda SA. Tet2 rescues age-related regenerative decline and enhances cognitive function in the adult mouse brain. Cell Rep. 2018;22:1974–1981. doi: 10.1016/j.celrep.2018.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris RB. Loss of body fat in lean parabiotic partners of Ob/Ob mice. Am J Phys. 1997;272:R1809–R1815. doi: 10.1152/ajpregu.1997.272.6.R1809. [DOI] [PubMed] [Google Scholar]
- Harris RB. Parabiosis between db/db and ob/ob or db/+ mice. Endocrinology. 1999;140:138–145. doi: 10.1210/endo.140.1.6449. [DOI] [PubMed] [Google Scholar]
- Harrison DE, Astle CM. Loss of stem cell repopulating ability upon transplantation. Effects of donor age, cell number, and transplantation procedure. J Exp Med. 1982;156:1767–1779. doi: 10.1084/jem.156.6.1767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Health, United States. 2016: With chartbook on long-term trends in health Hyattsville (MD); 2017. [PubMed]
- Higashi Y, Sukhanov S, Anwar A, Shai SY, Delafontaine P. Aging, atherosclerosis, and IGF-1. J Gerontol A Biol Sci Med Sci. 2012;67:626–639. doi: 10.1093/gerona/gls102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirayama R, Takemura K, Nihei Z, Ichikawa W, Takagi Y, Mishima Y, Utsuyama M, Hirokawa K. Differential effect of host microenvironment and systemic humoral factors on the implantation and the growth rate of metastatic tumor in parabiotic mice constructed between young and old mice. Mech Ageing Dev. 1993;71:213–221. doi: 10.1016/0047-6374(93)90085-6. [DOI] [PubMed] [Google Scholar]
- Horrington EM, Pope F, Lunsford W, Mc CC. Age changes in the bones, blood pressure, and diseases of rats in parabiosis. Gerontologia. 1960;4:21–31. doi: 10.1159/000210970. [DOI] [PubMed] [Google Scholar]
- Imperio CG, McFalls AJ, Colechio EM, Masser DR, Vrana KE, Grigson PS, Freeman WM. Assessment of individual differences in the rat nucleus accumbens transcriptome following taste-heroin extended access. Brain Res Bull. 2016;123:71–80. doi: 10.1016/j.brainresbull.2015.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katoh Y, Katoh M. Comparative integromics on BMP/GDF family. Int J Mol Med. 2006;17:951–955. [PubMed] [Google Scholar]
- Katsimpardi L, Litterman NK, Schein PA, Miller CM, Loffredo FS, Wojtkiewicz GR, Chen JW, Lee RT, Wagers AJ, Rubin LL. Vascular and neurogenic rejuvenation of the aging mouse brain by young systemic factors. Science. 2014;344:630–634. doi: 10.1126/science.1251141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katsimpardi L, Kuperwasser N, Camus C, Moigneu C, Chiche A, Tolle V, et al. Systemic GDF11 stimulates the secretion of adiponectin and induces a calorie restriction-like phenotype in aged mice. Aging Cell. 2020;19(1):e13038. doi: 10.1111/acel.13038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiss T, Balasubramanian P, Valcarcel-Ares MN, Tarantini S, Yabluchanskiy A, Csipo T, et al. Nicotinamide mononucleotide (NMN) treatment attenuates oxidative stress and rescues angiogenic capacity in aged cerebromicrovascular endothelial cells: a potential mechanism for prevention of vascular cognitive impairment. GeroScience. 2019a; in press. [DOI] [PMC free article] [PubMed]
- Kiss T, Giles CB, Tarantini S, Yabluchanskiy A, Balasubramanian P, Gautam T, et al. Nicotinamide mononucleotide (NMN) supplementation promotes anti-aging miRNA expression profile in the aorta of aged mice, predicting epigenetic rejuvenation and anti-atherogenic effects. Geroscience. 2019;41(4):419–39. doi: 10.1007/s11357-019-00095-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiss T, Nyul-Toth A, Balasubramanian P, Tarantini S, Ahire C, Yabluchanskiy A, et al. Nicotinamide mononucleotide (NMN) supplementation promotes neurovascular rejuvenation in aged mice: transcriptional footprint of SIRT1 activation, mitochondrial protection, anti-inflammatory, and anti-apoptotic effects. Geroscience. 2020 doi: 10.1007/s11357-020-00165-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kramer A, Green J, Pollard J, Jr, Tugendreich S. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics. 2014;30:523–530. doi: 10.1093/bioinformatics/btt703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Labinskyy N, Csiszar A, Orosz Z, Smith K, Rivera A, Buffenstein R, Ungvari Z. Comparison of endothelial function, O2.- and H2O2 production, and vascular oxidative stress resistance between the longest-living rodent, the naked mole rat, and mice. Am J Physiol. 2006;291:H2698–H2704. doi: 10.1152/ajpheart.00534.2006. [DOI] [PubMed] [Google Scholar]
- Labinskyy N, Mukhopadhyay P, Toth J, Szalai G, Veres M, Losonczy G, Pinto JT, Pacher P, Ballabh P, Podlutsky A, Austad SN, Csiszar A, Ungvari Z. Longevity is associated with increased vascular resistance to high glucose-induced oxidative stress and inflammatory gene expression in Peromyscus leucopus. Am J Physiol Heart Circ Physiol. 2009;296:H946–H956. doi: 10.1152/ajpheart.00693.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lahteenvuo J, Rosenzweig A. Effects of aging on angiogenesis. Circ Res. 2012;110:1252–1264. doi: 10.1161/CIRCRESAHA.111.246116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lakatta EG, Levy D. Arterial and cardiac aging: major shareholders in cardiovascular disease enterprises: part I: aging arteries: a “set up” for vascular disease. Circulation. 2003;107:139–146. doi: 10.1161/01.cir.0000048892.83521.58. [DOI] [PubMed] [Google Scholar]
- Lehallier B, Gate D, Schaum N, Nanasi T, Lee SE, Yousef H, Moran Losada P, Berdnik D, Keller A, Verghese J, Sathyan S, Franceschi C, Milman S, Barzilai N, Wyss-Coray T. Undulating changes in human plasma proteome profiles across the lifespan. Nat Med. 2019;25:1843–1850. doi: 10.1038/s41591-019-0673-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lesniewski LA, Seals DR, Walker AE, Henson GD, Blimline MW, Trott DW, Bosshardt GC, LaRocca TJ, Lawson BR, Zigler MC, Donato AJ. Dietary rapamycin supplementation reverses age-related vascular dysfunction and oxidative stress, while modulating nutrient-sensing, cell cycle, and senescence pathways. Aging Cell. 2017;16:17–26. doi: 10.1111/acel.12524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Libert S, Pointer K, Bell EL, Das A, Cohen DE, Asara JM, Kapur K, Bergmann S, Preisig M, Otowa T, Kendler KS, Chen X, Hettema JM, van den Oord EJ, Rubio JP, Guarente L. SIRT1 activates MAO-A in the brain to mediate anxiety and exploratory drive. Cell. 2011;147:1459–1472. doi: 10.1016/j.cell.2011.10.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin AL, Zheng W, Halloran JJ, Burbank RR, Hussong SA, Hart MJ, Javors M, Shih YY, Muir E, Solano Fonseca R, Strong R, Richardson AG, Lechleiter JD, Fox PT, Galvan V. Chronic rapamycin restores brain vascular integrity and function through NO synthase activation and improves memory in symptomatic mice modeling Alzheimer’s disease. J Cereb Blood Flow Metab. 2013;33:1412–1421. doi: 10.1038/jcbfm.2013.82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu C, Liu W, Palie J, Lu MF, Brown NA, Martin JF. Pitx2c patterns anterior myocardium and aortic arch vessels and is required for local cell movement into atrioventricular cushions. Development. 2002;129:5081–5091. doi: 10.1242/dev.129.21.5081. [DOI] [PubMed] [Google Scholar]
- Loffredo FS, Steinhauser ML, Jay SM, Gannon J, Pancoast JR, Yalamanchi P, Sinha M, Dall'Osso C, Khong D, Shadrach JL, Miller CM, Singer BS, Stewart A, Psychogios N, Gerszten RE, Hartigan AJ, Kim MJ, Serwold T, Wagers AJ, Lee RT. Growth differentiation factor 11 is a circulating factor that reverses age-related cardiac hypertrophy. Cell. 2013;153:828–839. doi: 10.1016/j.cell.2013.04.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez-Otin C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013;153:1194–1217. doi: 10.1016/j.cell.2013.05.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15:550. doi: 10.1186/s13059-014-0550-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ludwig FC, Elashoff RM. Mortality in syngeneic rat parabionts of different chronological age. Trans N Y Acad Sci. 1972;34:582–587. doi: 10.1111/j.2164-0947.1972.tb02712.x. [DOI] [PubMed] [Google Scholar]
- Lunsford WR, Mc CC, Lupien PJ, Pope FE, Sperling G. Parabiosis as a method for studying factors which affect aging in rats. Gerontologia. 1963;7:1–8. doi: 10.1159/000211170. [DOI] [PubMed] [Google Scholar]
- Masliah E, Ho G, Wyss-Coray T. Functional role of TGF beta in Alzheimer’s disease microvascular injury: lessons from transgenic mice. Neurochem Int. 2001;39:393–400. doi: 10.1016/s0197-0186(01)00046-8. [DOI] [PubMed] [Google Scholar]
- Massudi H, Grant R, Braidy N, Guest J, Farnsworth B, Guillemin GJ. Age-associated changes in oxidative stress and NAD+ metabolism in human tissue. PLoS One. 2012;7:e42357. doi: 10.1371/journal.pone.0042357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mattison JA, Wang M, Bernier M, Zhang J, Park SS, Maudsley S, An SS, Santhanam L, Martin B, Faulkner S, Morrell C, Baur JA, Peshkin L, Sosnowska D, Csiszar A, Herbert RL, Tilmont EM, Ungvari Z, Pearson KJ, Lakatta EG, de Cabo R. Resveratrol prevents high fat/sucrose diet-induced central arterial wall inflammation and stiffening in nonhuman primates. Cell Metab. 2014;20:183–190. doi: 10.1016/j.cmet.2014.04.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCay CM, Pope F, Lunsford W, Sperling G, Sambhavaphol P. Parabiosis between old and young rats. Gerontologia. 1957;1:7–17. doi: 10.1159/000210677. [DOI] [PubMed] [Google Scholar]
- Middeldorp J, Lehallier B, Villeda SA, Miedema SS, Evans E, Czirr E, Zhang H, Luo J, Stan T, Mosher KI, Masliah E, Wyss-Coray T. Preclinical assessment of young blood plasma for Alzheimer disease. JAMA Neurol. 2016;73:1325–1333. doi: 10.1001/jamaneurol.2016.3185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milliard S, Silva A, Blaise G, Chen H, Xu D, Qi S, Daloze P. Rapamycin’s effect on vasomotion in the rat. Transplant Proc. 1998;30:1036–1038. doi: 10.1016/s0041-1345(98)00140-7. [DOI] [PubMed] [Google Scholar]
- Mitschelen M, Yan H, Farley JA, Warrington JP, Han S, Herenu CB, Csiszar A, Ungvari Z, Bailey-Downs LC, Bass CE, Sonntag WE. Long-term deficiency of circulating and hippocampal insulin-like growth factor I induces depressive behavior in adult mice: a potential model of geriatric depression. Neuroscience. 2011;185:50–60. doi: 10.1016/j.neuroscience.2011.04.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrison EJ, Champagne DP, Dzieciatkowska M, Nemkov T, Zimring JC, Hansen KC, et al. Parabiosis incompletely reverses aging-induced metabolic changes and oxidant stress in mouse red blood cells. Nutrients. 2019;11(6). 10.3390/nu11061337. [DOI] [PMC free article] [PubMed]
- Nakao A, Imamura T, Souchelnytskyi S, Kawabata M, Ishisaki A, Oeda E, Tamaki K, Hanai J, Heldin CH, Miyazono K, ten Dijke P. TGF-beta receptor-mediated signalling through Smad2, Smad3 and Smad4. EMBO J. 1997;16:5353–5362. doi: 10.1093/emboj/16.17.5353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nguyen AD, Nguyen TA, Zhang J, Devireddy S, Zhou P, Karydas AM, Xu X, Miller BL, Rigo F, Ferguson SM, Huang EJ, Walther TC, Farese RV., Jr Murine knockin model for progranulin-deficient frontotemporal dementia with nonsense-mediated mRNA decay. Proc Natl Acad Sci U S A. 2018;115:E2849–E2858. doi: 10.1073/pnas.1722344115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oka S, Alcendor R, Zhai P, Park JY, Shao D, Cho J, Yamamoto T, Tian B, Sadoshima J. PPARalpha-Sirt1 complex mediates cardiac hypertrophy and failure through suppression of the ERR transcriptional pathway. Cell Metab. 2011;14:598–611. doi: 10.1016/j.cmet.2011.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parker SJ, Stotland A, MacFarlane E, Wilson N, Orosco A, Venkatraman V, Madrid K, Gottlieb R, Dietz HC, Van Eyk JE. Proteomics reveals Rictor as a noncanonical TGF-beta signaling target during aneurysm progression in Marfan mice. Am J Physiol Heart Circ Physiol. 2018;315:H1112–H1126. doi: 10.1152/ajpheart.00089.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parlar A, Can C, Erol A, Ulker S. Posttransplantation therapeutic rapamycin concentration protects nitric oxide-related vascular endothelial function: comparative effects in rat thoracic aorta and coronary endothelial cell culture. Transplant Proc. 2010;42:1923–1930. doi: 10.1016/j.transproceed.2010.03.134. [DOI] [PubMed] [Google Scholar]
- Patten IS, Arany Z. PGC-1 coactivators in the cardiovascular system. Trends Endocrinol Metab. 2012;23:90–97. doi: 10.1016/j.tem.2011.09.007. [DOI] [PubMed] [Google Scholar]
- Pearson KJ, Baur JA, Lewis KN, Peshkin L, Price NL, Labinskyy N, Swindell WR, Kamara D, Minor RK, Perez E, Jamieson HA, Zhang Y, Dunn SR, Sharma K, Pleshko N, Woollett LA, Csiszar A, Ikeno Y, Le Couteur D, Elliott PJ, Becker KG, Navas P, Ingram DK, Wolf NS, Ungvari Z, Sinclair DA, de Cabo R. Resveratrol delays age-related deterioration and mimics transcriptional aspects of dietary restriction without extending life span. Cell Metab. 2008;8:157–168. doi: 10.1016/j.cmet.2008.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poggioli T, Vujic A, Yang P, Macias-Trevino C, Uygur A, Loffredo FS, Pancoast JR, Cho M, Goldstein J, Tandias RM, Gonzalez E, Walker RG, Thompson TB, Wagers AJ, Fong YW, Lee RT. Circulating growth differentiation factor 11/8 levels decline with age. Circ Res. 2016;118:29–37. doi: 10.1161/CIRCRESAHA.115.307521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rebo J, Mehdipour M, Gathwala R, Causey K, Liu Y, Conboy MJ, Conboy IM. A single heterochronic blood exchange reveals rapid inhibition of multiple tissues by old blood. Nat Commun. 2016;7:13363. doi: 10.1038/ncomms13363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodgers BD, Eldridge JA. Reduced circulating GDF11 is unlikely responsible for age-dependent changes in mouse heart, muscle, and brain. Endocrinology. 2015;156:3885–3888. doi: 10.1210/en.2015-1628. [DOI] [PubMed] [Google Scholar]
- Roth Flach RJ, Skoura A, Matevossian A, Danai LV, Zheng W, Cortes C, Bhattacharya SK, Aouadi M, Hagan N, Yawe JC, Vangala P, Menendez LG, Cooper MP, Fitzgibbons TP, Buckbinder L, Czech MP. Endothelial protein kinase MAP4K4 promotes vascular inflammation and atherosclerosis. Nat Commun. 2015;6:8995. doi: 10.1038/ncomms9995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shang Y, Yoshida T, Amendt BA, Martin JF, Owens GK. Pitx2 is functionally important in the early stages of vascular smooth muscle cell differentiation. J Cell Biol. 2008;181:461–473. doi: 10.1083/jcb.200711145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinha M, Jang YC, Oh J, Khong D, Wu EY, Manohar R, Miller C, Regalado SG, Loffredo FS, Pancoast JR, Hirshman MF, Lebowitz J, Shadrach JL, Cerletti M, Kim MJ, Serwold T, Goodyear LJ, Rosner B, Lee RT, Wagers AJ. Restoring systemic GDF11 levels reverses age-related dysfunction in mouse skeletal muscle. Science. 2014;344:649–652. doi: 10.1126/science.1251152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith LK, He Y, Park JS, Bieri G, Snethlage CE, Lin K, Gontier G, Wabl R, Plambeck KE, Udeochu J, Wheatley EG, Bouchard J, Eggel A, Narasimha R, Grant JL, Luo J, Wyss-Coray T, Villeda SA. beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis. Nat Med. 2015;21:932–937. doi: 10.1038/nm.3898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith SC, Zhang X, Zhang X, Gross P, Starosta T, Mohsin S, Franti M, Gupta P, Hayes D, Myzithras M, Kahn J, Tanner J, Weldon SM, Khalil A, Guo X, Sabri A, Chen X, MacDonnell S, Houser SR. GDF11 does not rescue aging-related pathological hypertrophy. Circ Res. 2015;117:926–932. doi: 10.1161/CIRCRESAHA.115.307527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Son HG, Lee SV. Longevity regulation by NMD-mediated mRNA quality control. BMB Rep. 2017;50:160–161. doi: 10.5483/BMBRep.2017.50.4.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Son HG, Seo M, Ham S, Hwang W, Lee D, An SW, Artan M, Seo K, Kaletsky R, Arey RN, Ryu Y, Ha CM, Kim YK, Murphy CT, Roh TY, Nam HG, Lee SV. RNA surveillance via nonsense-mediated mRNA decay is crucial for longevity in daf-2/insulin/IGF-1 mutant C. elegans. Nat Commun. 2017;8:14749. doi: 10.1038/ncomms14749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soneson C, Love MI, Robinson MD. Differential analyses for RNA-seq: transcript-level estimates improve gene-level inferences. F1000Res. 2015;4:1521. doi: 10.12688/f1000research.7563.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sonntag WE, Deak F, Ashpole N, Toth P, Csiszar A, Freeman W, Ungvari Z. Insulin-like growth factor-1 in CNS and cerebrovascular aging. Front Aging Neurosci. 2013;5:27. doi: 10.3389/fnagi.2013.00027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sousa-Victor P, Neves J, Cedron-Craft W, Ventura PB, Liao CY, Riley RR, Soifer I, van Bruggen N, Kolumam GA, Villeda SA, Lamba DA, Jasper H. MANF regulates metabolic and immune homeostasis in ageing and protects against liver damage. Nat Metab. 2019;1:276–290. doi: 10.1038/s42255-018-0023-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Springo Z, Tarantini S, Toth P, Tucsek Z, Koller A, Sonntag WE, Csiszar A, Ungvari Z. Aging exacerbates pressure-induced mitochondrial oxidative stress in mouse cerebral arteries. J Gerontol A Biol Sci Med Sci. 2015;70:1355–1359. doi: 10.1093/gerona/glu244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub TR, Lander ES, Mesirov JP. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A. 2005;102:15545–15550. doi: 10.1073/pnas.0506580102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tabrez SS, Sharma RD, Jain V, Siddiqui AA, Mukhopadhyay A. Differential alternative splicing coupled to nonsense-mediated decay of mRNA ensures dietary restriction-induced longevity. Nat Commun. 2017;8:306. doi: 10.1038/s41467-017-00370-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarantini S, Giles CB, Wren JD, Ashpole NM, Valcarcel-Ares MN, Wei JY, Sonntag WE, Ungvari Z, Csiszar A. IGF-1 deficiency in a critical period early in life influences the vascular aging phenotype in mice by altering miRNA-mediated post-transcriptional gene regulation: implications for the developmental origins of health and disease hypothesis. Age (Dordr) 2016;38:239–258. doi: 10.1007/s11357-016-9943-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarantini S, Tucsek Z, Valcarcel-Ares MN, Toth P, Gautam T, Giles CB, Ballabh P, Wei JY, Wren JD, Ashpole NM, Sonntag WE, Ungvari Z, Csiszar A. Circulating IGF-1 deficiency exacerbates hypertension-induced microvascular rarefaction in the mouse hippocampus and retrosplenial cortex: implications for cerebromicrovascular and brain aging. Age (Dordr) 2016;38:273–289. doi: 10.1007/s11357-016-9931-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarantini S, Valcarcel-Ares NM, Yabluchanskiy A, Springo Z, Fulop GA, Ashpole N, Gautam T, Giles CB, Wren JD, Sonntag WE, Csiszar A, Ungvari Z. Insulin-like growth factor 1 deficiency exacerbates hypertension-induced cerebral microhemorrhages in mice, mimicking the aging phenotype. Aging Cell. 2017;16:469–479. doi: 10.1111/acel.12583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarantini S, Valcarcel-Ares NM, Yabluchanskiy A, Fulop GA, Hertelendy P, Gautam T, et al. Treatment with the mitochondrial-targeted antioxidant peptide SS-31 rescues neurovascular coupling responses and cerebrovascular endothelial function and improves cognition in aged mice. Aging Cell. 2018;17(2). 10.1111/acel.12731. [DOI] [PMC free article] [PubMed]
- Tarantini S, Yabluchanskiy A, Csipo T, Fulop G, Kiss T, Balasubramanian P, et al. Treatment with the poly(ADP-ribose) polymerase inhibitor PJ-34 improves cerebromicrovascular endothelial function, neurovascular coupling responses and cognitive performance in aged mice, supporting the NAD+ depletion hypothesis of neurovascular aging. Geroscience. 2019a;41(5):533–42. 10.1007/s11357-019-00101-2. [DOI] [PMC free article] [PubMed]
- Tarantini S, Valcarcel-Ares MN, Toth P, Yabluchanskiy A, Tucsek Z, Kiss T, Hertelendy P, Kinter M, Ballabh P, Sule Z, Farkas E, Baur JA, Sinclair DA, Csiszar A, Ungvari Z. Nicotinamide mononucleotide (NMN) supplementation rescues cerebromicrovascular endothelial function and neurovascular coupling responses and improves cognitive function in aged mice. Redox Biol. 2019;24:101192. doi: 10.1016/j.redox.2019.101192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tchkonia T, Zhu Y, van Deursen J, Campisi J, Kirkland JL. Cellular senescence and the senescent secretory phenotype: therapeutic opportunities. J Clin Invest. 2013;123:966–972. doi: 10.1172/JCI64098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toth P, Tarantini S, Tucsek Z, Ashpole NM, Sosnowska D, Gautam T, Ballabh P, Koller A, Sonntag WE, Csiszar A, Ungvari ZI. Resveratrol treatment rescues neurovascular coupling in aged mice:role of improved cerebromicrovascular endothelial function and down-regulation of NADPH oxidas. Am J Physiol Heart Circ Physiol. 2014;306:H299–H308. doi: 10.1152/ajpheart.00744.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toth P, Tucsek Z, Tarantini S, Sosnowska D, Gautam T, Mitschelen M, Koller A, Sonntag WE, Csiszar A, Ungvari Z. IGF-1 deficiency impairs cerebral myogenic autoregulation in hypertensive mice. J Cereb Blood Flow Metab. 2014;34:1887–1897. doi: 10.1038/jcbfm.2014.156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toth P, Tarantini S, Springo Z, Tucsek Z, Gautam T, Giles CB, Wren JD, Koller A, Sonntag WE, Csiszar A, Ungvari Z. Aging exacerbates hypertension-induced cerebral microhemorrhages in mice: role of resveratrol treatment in vasoprotection. Aging Cell. 2015;14:400–408. doi: 10.1111/acel.12315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toth P, Tarantini S, Ashpole NM, Tucsek Z, Milne GL, Valcarcel-Ares NM, Menyhart A, Farkas E, Sonntag WE, Csiszar A, Ungvari Z. IGF-1 deficiency impairs neurovascular coupling in mice: implications for cerebromicrovascular aging. Aging Cell. 2015;14:1034–1044. doi: 10.1111/acel.12372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ungvari Z, Csiszar A. The emerging role of IGF-1 deficiency in cardiovascular aging: recent advances. J Gerontol A Biol Sci Med Sci. 2012;67:599–610. doi: 10.1093/gerona/gls072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ungvari Z, Csiszar A, Huang A, Kaminski PM, Wolin MS, Koller A. High pressure induces superoxide production in isolated arteries via protein kinase C-dependent activation of NAD(P)H oxidase. Circulation. 2003;108:1253–1258. doi: 10.1161/01.CIR.0000079165.84309.4D. [DOI] [PubMed] [Google Scholar]
- Ungvari Z, Orosz Z, Labinskyy N, Rivera A, Xiangmin Z, Smith K, Csiszar A. Increased mitochondrial H2O2 production promotes endothelial NF-kappaB activation in aged rat arteries. Am J Physiol Heart Circ Physiol. 2007;293:H37–H47. doi: 10.1152/ajpheart.01346.2006. [DOI] [PubMed] [Google Scholar]
- Ungvari ZI, Labinskyy N, Gupte SA, Chander PN, Edwards JG, Csiszar A. Dysregulation of mitochondrial biogenesis in vascular endothelial and smooth muscle cells of aged rats. Am J Physiol Heart Circ Physiol. 2008;294:H2121–H2128. doi: 10.1152/ajpheart.00012.2008. [DOI] [PubMed] [Google Scholar]
- Ungvari Z, Bailey-Downs L, Sosnowska D, Gautam T, Koncz P, Losonczy G, Ballabh P, de Cabo R, Sonntag WE, Csiszar A. Vascular oxidative stress in aging: a homeostatic failure due to dysregulation of Nrf2-mediated antioxidant response. Am J Physiol Heart Circ Physiol. 2011;301:H363–H372. doi: 10.1152/ajpheart.01134.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ungvari Z, Tarantini S, Donato AJ, Galvan V, Csiszar A. Mechanisms of vascular aging. Circ Res. 2018;123:849–867. doi: 10.1161/CIRCRESAHA.118.311378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ungvari Z, Tarantini S, Kiss T, Wren JD, Giles CB, Griffin CT, Murfee WL, Pacher P, Csiszar A. Endothelial dysfunction and angiogenesis impairment in the ageing vasculature. Nat Rev Cardiol. 2018;15:555–565. doi: 10.1038/s41569-018-0030-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valcarcel-Ares MN, Tucsek Z, Kiss T, Giles CB, Tarantini S, Yabluchanskiy A, et al. Obesity in aging exacerbates neuroinflammation, dysregulating synaptic function-related genes and altering eicosanoid synthesis in the mouse hippocampus: potential role in impaired synaptic plasticity and cognitive decline. J Gerontol A Biol Sci Med Sci. 2019;74(3):290–8. doi: 10.1093/gerona/gly127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Skike CE, Jahrling JB, Olson AB, Sayre NL, Hussong SA, Ungvari Z, Lechleiter JD, Galvan V. Inhibition of mTOR protects the blood-brain barrier in models of Alzheimer’s disease and vascular cognitive impairment. Am J Physiol Heart Circ Physiol. 2018;314:H693–H703. doi: 10.1152/ajpheart.00570.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varaljai R, Islam AB, Beshiri ML, Rehman J, Lopez-Bigas N, Benevolenskaya EV. Increased mitochondrial function downstream from KDM5A histone demethylase rescues differentiation in pRB-deficient cells. Genes Dev. 2015;29:1817–1834. doi: 10.1101/gad.264036.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villeda SA, Luo J, Mosher KI, Zou B, Britschgi M, Bieri G, Stan TM, Fainberg N, Ding Z, Eggel A, Lucin KM, Czirr E, Park JS, Couillard-Despres S, Aigner L, Li G, Peskind ER, Kaye JA, Quinn JF, Galasko DR, Xie XS, Rando TA, Wyss-Coray T. The ageing systemic milieu negatively regulates neurogenesis and cognitive function. Nature. 2011;477:90–94. doi: 10.1038/nature10357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villeda SA, Plambeck KE, Middeldorp J, Castellano JM, Mosher KI, Luo J, Smith LK, Bieri G, Lin K, Berdnik D, Wabl R, Udeochu J, Wheatley EG, Zou B, Simmons DA, Xie XS, Longo FM, Wyss-Coray T. Young blood reverses age-related impairments in cognitive function and synaptic plasticity in mice. Nat Med. 2014;20:659–663. doi: 10.1038/nm.3569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiedenhoeft T, Tarantini S, Nyul-Toth A, Yabluchanskiy A, Csipo T, Balasubramanian P, Lipecz A, Kiss T, Csiszar A, Csiszar A, Ungvari Z. Fusogenic liposomes effectively deliver resveratrol to the cerebral microcirculation and improve endothelium-dependent neurovascular coupling responses in aged mice. Geroscience. 2019;41:711–725. doi: 10.1007/s11357-019-00102-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wyss-Coray T. Ageing, neurodegeneration and brain rejuvenation. Nature. 2016;539:180–186. doi: 10.1038/nature20411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wyss-Coray T, Feng L, Masliah E, Ruppe MD, Lee HS, Toggas SM, Rockenstein EM, Mucke L. Increased central nervous system production of extracellular matrix components and development of hydrocephalus in transgenic mice overexpressing transforming growth factor-beta 1. Am J Pathol. 1995;147:53–67. [PMC free article] [PubMed] [Google Scholar]
- Wyss-Coray T, Borrow P, Brooker MJ, Mucke L. Astroglial overproduction of TGF-beta 1 enhances inflammatory central nervous system disease in transgenic mice. J Neuroimmunol. 1997;77:45–50. doi: 10.1016/s0165-5728(97)00049-0. [DOI] [PubMed] [Google Scholar]
- Wyss-Coray T, Masliah E, Mallory M, McConlogue L, Johnson-Wood K, Lin C, Mucke L. Amyloidogenic role of cytokine TGF-beta1 in transgenic mice and in Alzheimer’s disease. Nature. 1997;389:603–606. doi: 10.1038/39321. [DOI] [PubMed] [Google Scholar]
- Wyss-Coray T, Lin C, von Euw D, Masliah E, Mucke L, Lacombe P. Alzheimer's disease-like cerebrovascular pathology in transforming growth factor-beta 1 transgenic mice and functional metabolic correlates. Ann N Y Acad Sci. 2000;903:317–323. doi: 10.1111/j.1749-6632.2000.tb06382.x. [DOI] [PubMed] [Google Scholar]
- Yoshino J, Baur JA, Imai SI. NAD(+) intermediates: the biology and therapeutic potential of NMN and NR. Cell Metab. 2018;27:513–528. doi: 10.1016/j.cmet.2017.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu G, He QY. ReactomePA: an R/Bioconductor package for reactome pathway analysis and visualization. Mol BioSyst. 2016;12:477–479. doi: 10.1039/c5mb00663e. [DOI] [PubMed] [Google Scholar]
- Zhang H, Cherian R, Jin K. Systemic milieu and age-related deterioration. Geroscience. 2019;41:275–284. doi: 10.1007/s11357-019-00075-1. [DOI] [PMC free article] [PubMed] [Google Scholar]