

Abstract
Omiganan (OMN; a synthetic cationic peptide) and imiquimod (IMQ; a TLR7 agonist) have synergistic effects on interferon responses in vitro. The objective of this study was to translate this to a human model for proof‐of‐concept, and to explore the potential of OMN add‐on treatment for viral skin diseases. Sixteen healthy volunteers received topical IMQ, OMN, or a combination of both for up to 4 days on tape‐stripped skin. Skin inflammation was quantified by laser speckle contrast imaging and 2D photography, and molecular and cellular responses were analyzed in biopsies. IMQ treatment induced an inflammatory response of the skin. Co‐treatment with OMN enhanced this inflammatory response to IMQ, with increases in perfusion (+17.1%; 95% confidence interval (CI) 5.6%–30%; P < 0.01) and erythema (+1.5; 95% CI 0.25%–2.83; P = 0.02). Interferon regulatory factor‐driven and NFκB‐driven responses following TLR7 stimulation were enhanced by OMN (increases in IL‐6, IL‐10, MXA, and IFNɣ), and more immune cell infiltration was observed (in particular CD4+, CD8+, and CD14+ cells). These findings are in line with the earlier mechanistic in vitro data, and support evaluation of imiquimod/OMN combination therapy in human papillomavirus‐induced skin diseases.
Study Highlights.
WHAT IS THE CURRENT KNOWLEDGE ON THE TOPIC?
☑ Omiganan (OMN) enhances IFN responses induced by TLR7 (imiquimod (IMQ)) in vitro, which supports the future application of OMN as co‐treatment with IMQ for viral skin disease in humans.
WHAT QUESTION DID THIS STUDY ADDRESS?
☑ Does OMN treatment enhance IMQ‐induced IFN responses in the IMQ skin inflammation challenge model in healthy volunteers?
WHAT DOES THIS STUDY ADD TO OUR KNOWLEDGE?
☑OMN enhanced the inflammatory skin response to IMQ, supporting evaluation of IMQ/OMN combination therapy in HPV‐induced skin diseases, such as anogenital warts or high‐grade squamous intraepithelial lesion.
HOW MIGHT THIS CHANGE CLINICAL PHARMACOLOGY OR TRANSLATIONAL SCIENCE?
☑ This study shows that the IMQ skin inflammation challenge model is suitable to study proof‐of‐pharmacology of novel compounds targeting the innate immune system in healthy volunteers.
Cathelicidins are a family of antimicrobial (cationic) peptides that play an important role in the first line immune defense of the skin, related to their broad antimicrobial activity against bacteria, viruses, and fungi.1 LL‐37 is the only human member of the cathelicidin family.1 Besides its antimicrobial effects, this peptide also has direct immunomodulatory activity. LL‐37 affects the response of neutrophils to viruses, and modulates interferon (IFN) responses induced by viral triggers.2 LL‐37 converts self‐RNA into a ligand for toll‐like receptor (TLR)7 and TLR8 in human dendritic cells, thereby enhancing IFNα production in human skin.3
Omiganan (OMN) is a synthetic indolicidin (a cathelicidin isolated from bovine neutrophils), currently under development as topical gel for several clinical indications. OMN is known to have activity against a wide variety of microorganisms, such as gram‐positive and gram‐negative bacteria and fungi.4, 5 Moreover, OMN enhances IFN responses induced by TLR3 (Poly:IC), TLR7 (imiquimod (IMQ)), TLR8 (ssRNA), and TLR9 (CpG) in human immune cells, comparable but not similar to the effects observed for LL‐37 (unpublished data, Grievink et al.). These observations support the future application of OMN as co‐treatment with endosomal TLR ligands for viral skin disease in humans.
IMQ is the only registered endosomal TLR ligand, as Aldara topical cream. The mechanism of action of IMQ is based on TLR7‐dependent MyD88‐signaling.6, 7 This results in two responses: a tumoricidal effect by the release of several pro‐inflammatory cytokines (e.g., TNF‐α, IL‐6, and IL‐8, via NFκB) and an antiviral response by the induction of IFNα and IFN‐inducible genes (e.g., MX1 and MXA, via interferon regulatory factor (IRF)7).8 Based on these mechanisms IMQ is widely used in clinical practice for human papillomavirus (HPV)‐induced anogenital warts and high‐grade squamous intraepithelial lesions of the vulva (vulvar HSIL), actinic keratosis, and basal cell carcinoma.9 In most of these conditions, drug efficacy is suboptimal, and lesions may re‐occur after treatment discontinuation.10 Therefore, a treatment enhancing the efficacy of IMQ in these dermatological conditions would be of great benefit. Based on its observed preclinical activity, OMN may be a good candidate for combination treatment with IMQ.
We recently developed an in vivo challenge model with transient local skin inflammation, induced by 48 hours IMQ (Aldara cream, Aldara, Meda Pharma, Solna, Sweden) application under occlusion by a 12 mm Finn chamber to tape‐stripped skin.11 This model was used in the current study to explore the potential of combined IMQ and OMN treatment as novel therapeutic modality for HPV‐induced skin diseases (e.g., genital warts and vulvar HSIL). OMN was applied topically to IMQ‐primed skin, and the clinical, biophysical, cellular, and molecular responses to this combined treatment were investigated.
Methods
Study design and subjects
This was a randomized, open‐label, evaluator‐blinded, vehicle‐controlled, parallel‐cohort, dose‐ranging study. The study was conducted from February 2017 to March 2017 at the Centre for Human Drug Research, Leiden, The Netherlands, and was approved by the independent Medical Ethics Committee “Medisch Ethische Toetsingscommissie van de Stichting Beoordeling Ethiek Biomedisch Onderzoek” (Assen, The Netherlands). The study was conducted according to the Dutch Act on Medical Research involving Human Subjects (WMO). Before study procedures started, all subjects gave informed consent. Sixteen healthy male and female white (Fitzpatrick skin type III) volunteers, aged 18–45 years, were included. Subjects with a (family) history of psoriasis or any disease associated with immune system impairment were excluded.
Treatments and randomization
To explore the effect of OMN and the combination of OMN and IMQ on tape‐stripped skin, treatment combinations were applied and randomized over different treatment sites on the back (Table 1). All four treatment combinations were explored in each study participant. A standard daily dosage containing either 100 mg Aldara 5% (5 mg IMQ), 100 mg OMN 1% (1 mgOMN), 100 mg OMN 2.5%, 100 mg OMN vehicle (VehO), or cetomacrogol (which served as IMQ vehicle (VehI)) was applied under occlusion by a 12 mm Finn chamber (Smart Practice, Phoenix, AZ). The tape‐stripping procedure included 20 times stripping with tape (D‐Squame; CuDerm, Dallas, TX) to induce mild barrier disruption.
Table 1.
Treatment combinations
| Day 0 | Day 1 | Day 2 | Day 3 | |
|---|---|---|---|---|
| 1 | Imiquimod | Imiquimod | Vehicle (omiganan) | Vehicle (omiganan) |
| 2 | Imiquimod | Imiquimod | Omiganan 1% | Omiganan 1% |
| 3 | Imiquimod | Imiquimod | Omiganan 2.5% | Omiganan 2.5% |
| 4 | Vehicle (omiganan) | Vehicle (omiganan) | Vehicle (imiquimod) | Vehicle (imiquimod) |
It should be noted that within the same clinical study, alternative regimens and control conditions were explored within the same group of 16 volunteers. These additional conditions included the reverse treatment sequence (first IMQ, then OMN) and partial control groups VehI or VehO (1% or 2.5%). To increase the readability of this paper, it was decided to not present data related to these conditions.
Skin assessments
The skin was assessed daily for 5 days for signs of inflammation (erythema and hyperperfusion) by 2D photography, erythema index analysis, visual erythema grading (Clinician Erythema Assessment (CEA) scale; 0 represents absence of erythema, 4 very severe), colorimetry (a* value; DSM II ColorMeter; Cortex Technology, Hadslund, Denmark), and perfusion by laser speckle contrast imaging (LSCI; PeriCam PSI System, Perimed Jäfälla, Sweden). TAP (FibroTx, Estonia) were used to quantify skin surface biomarkers (IL‐8, IFNα, IL‐6, IL‐10, CCL20, and HBD‐2) by spot‐enzyme‐linked immunosorbent assay at predose and after end‐of‐treatment. Skin swabs were collected for microbiome analysis.
Three‐millimeter punch biopsies were collected predose (after tape stripping) and at end‐of‐treatment. For all 16 subjects, a biopsy of the VehO + VehI, IMQ + OMN1%, and IMQ + OMN2.5% treated areas was collected. For only eight subjects the IMQ + VehO‐treated area was biopsied, to limit the number of biopsies per subject. Biopsies were snap frozen using liquid nitrogen and stored at ‐80°C until analysis at the Immunology Laboratory of Erasmus Medical Center, Rotterdam, The Netherlands, for determination of IFNα, IFNγ, IL‐1β, IL‐6, IL‐8, HBD‐2, MX1, MXA, CCL20, and IL‐10 mRNA expression relative to the housekeeping gene ABL by quantitative polymerase chain reaction. In addition, all biopsies were hematoxylin‐eosin stained to obtain histopathological scores of psoriasis and dermatitis; general infiltration, parakeratosis, acanthosis, papillomatosis, and spongiosis. The histopathological score for each characteristic was graded based on fold increase or decrease compared with a reference biopsy of a healthy subject not participating in the clinical trial (1; equal to the reference biopsy, 2; twofold increase compared with the reference biopsy, etc.). Furthermore, immunohistochemical staining was performed to obtain scoring of markers CD11c (Clone 5D11; Cell Marque), CD14 (Clone EPR3653; Cell Marque), CD1a (Clone EP3622; Cell Marque), CD4 (Clone SP35; Ventana), CD8 (Clone SP57; Ventana), and HLA‐DR (CR3/43; Dako).
Safety end points
Safety and tolerability were monitored by tracking adverse events, performing physical examination, measuring vital signs, 12‐lead electrocardiograms, and laboratory tests (i.e., hematology, chemistry, and urinalysis) at multiple time points throughout the study. IFNα, IFNβ, and IFNɣ were measured in blood samples to detect a possible systemic effect of the interventions.
Statistics
Treatment effects were analyzed with a mixed model analysis of variance with the baseline measurement as covariate. To determine the differences between the treatments, contrasts were calculated for all measurements. All calculations were performed using SAS for windows version 9.4 (SAS Institute, Cary, NC). Evaluation window for noninvasive measures was 0–96 hours (day 4), whereas biopsies were collected at 120 hours (day 5).
Results
Twelve female (75%) and 4 male (25%) white subjects participated in the study. All 16 included subjects completed the study according to the schedule in Table 1. The mean age was 24.6 (SD ± 5.8 years). Application site pruritus was the most frequent occurring adverse event (AE) in 14 of 16 subjects (87.5%). This can be related to the tape‐stripping procedure, occlusion procedure, or one of the treatments or vehicles. No serious AEs or discontinuations due to AEs occurred. No systemic effects of any of the treatments in terms of elevated circulating cytokines (serum IFNα, IFN‐β, or IFNɣ) were observed (data not shown).
IMQ treatment resulted in a modest inflammatory response, observed as enhanced erythema (quantified by 2D photograph; Figure 1, top panel) and perfusion (quantified by LSCI; Figure 1, bottom panel). The maximal IMQ response was reached after 1–2 days of treatment (Figure 2). After 48 hours of VehI exposure, the target areas were treated with OMN (or vehicle) for an additional 2 days. OMN treatment enhanced the IMQ‐driven increase in skin perfusion and erythema, without an indication of OMN dose dependency (Figure 2). OMN treatment significantly enhanced perfusion (profile 0–96 hours; Figure 2 a: IMQ + vehicle vs. IMQ + OMN; +17.1%; 95% confidence interval (CI) 5.6%–30%; P < 0.01 and + 15.1%; 95% CI 3.8%–27.7%; P < 0.01 for 1% and 2.5% OMN, respectively). For erythema, a statistically significant OMN effect was observed (profile 0–96 hours; for colorimetry, but only at the 1% OMN dose; Figure 2 b: IMQ + vehicle vs. IMQ + OMN +1.5; 95% CI 0.25%–2.83%; P = 0.02 and +0.92; 95% CI 0.37%–2.21%; P = 0.16 for 1% and 2.5% OMN, respectively). OMN treatment did not significantly alter IMQ‐related increases in erythema index (profile 0–96 hours; +0.8; 95% CI –1.62% to 3.25%; P = 0.51 and +2.21; 95% CI –0.23% to 4.64%; P = 0.08 for 1% and 2.5% OMN, respectively). The enhanced inflammatory responses were observed during the OMN treatment period (days 3 and 4; 48–96 hours). Hereafter, perfusion and erythema returned within 1 day to levels as observed for the IMQ + vehicle treatment within 1 day (Figure 2; 120 hours).
Figure 1.
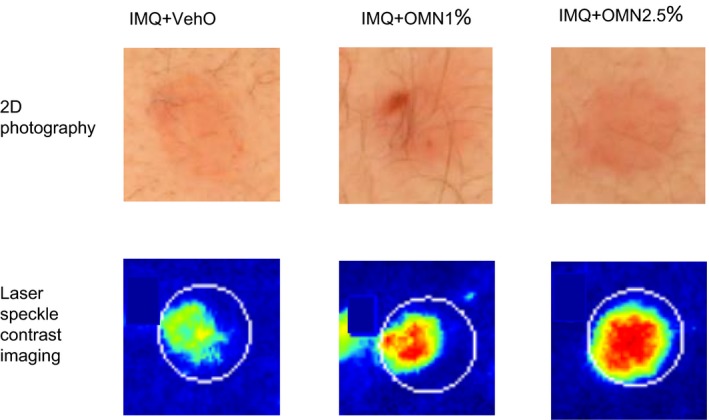
Clinical impression of imiquimod (IMQ) response (left panel) and IMQ + omiganan (OMN; middle and right panel) of one subject at day 4, 24 hours after the last application of omiganan or vehicle. VehO, omiganan vehicle.
Figure 2.
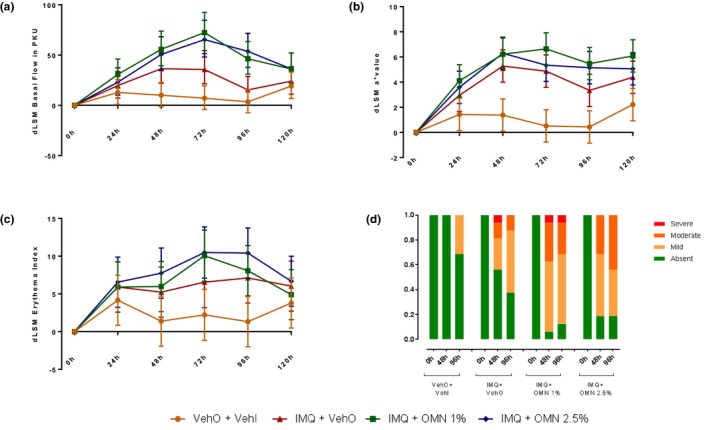
Skin inflammation induced by imiquimod (IMQ) and omiganan (OMN), as quantified by laser speckle contrast imaging (perfusion/basal flow, a), and erythema assessments (b: colorimetry, c: erythema, d: visual grading). dLSM, delta least square mean; VehI, imiquimod vehicle; VehO, omiganan vehicle.
In addition to the above noninvasive assessments, skin punch biopsies were taken from the target areas. Biopsies were stained for dermal immune cell infiltration, and independently analyzed by two investigators blinded to treatment compared with a reference biopsy (healthy unaffected skin). IMQ treatment resulted in an influx of immune cells in the skin, reflected by an increase in macrophages, HLA‐DR cells, myeloid dendritic cells, Langerhans cells, and CD4+ and CD8+ T cells (Figure 3 a–f, second bars vs. first bars). Consistent with the observations for perfusion and erythema, OMN treatment enhanced the IMQ‐driven inflammatory response as quantified in skin punch biopsies. When IMQ‐exposed skin was treated with OMN, this resulted in a strong increase of infiltrating immune cells (Figure 3 a–f, third and fourth bars vs. second bars). There was no indication of a clear OMN dose‐dependency, although the response to the 1% OMN formulation seemed slightly higher.
Figure 3.
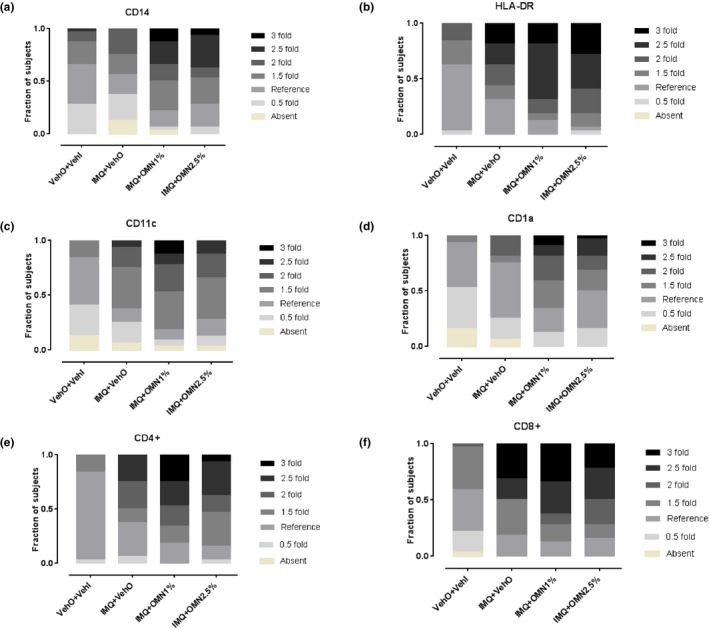
Skin inflammation induced by imiquimod (IMQ) and omiganan (OMN) on day 5 (scored compared with a reference biopsy), as quantified by immune cell influx. (a) CD14+ macrophages. (b) HLA‐DR cells. (c) CD11c+ myeloid dendritic cells. (d) CD1a+ Langerhans cells. (e) CD4+ T cells. (f) CD8+ T cells. VehI, imiquimod vehicle; VehO, omiganan vehicle.
Subsequently, the effects of IMQ and OMN add‐on treatment on local cytokine responses were investigated. As expected, IMQ treatment resulted in an NFκB‐driven increase in IL‐6 and IL‐10 (Figure 4 a; IL‐6 VehI vs. vehicle/vehicle + 120.9%; 95% CI 2.6%–375.6%; P = 0.04; IL‐10 VehI vs. vehicle/vehicle + 132.1%; 95% CI 40.8%–282.8%; P = 0.001). In line with this, IMQ increased the expression of type I IFN‐driven MXA (Figure 4 b, left panel, VehI vs. vehicle/vehicle + 213.3%; 95% CI 50.7%–551%; P = 0.002) and IFNɣ (Figure 4 b right panel, VehI vs. vehicle/vehicle + 542.4%; 95% CI 132.1%–1678.3%; P < 0.001). No treatment effect was observed for MX1 expression. Subsequently, OMN was applied for 2 days to the target areas. Although OMN did not significantly alter any of the IMQ‐driven responses, a higher level of cytokines was consistently found in the IMQ/OMN treatment group when compared with the VehI treatment group (Figure 4 a; IL‐6 and IL‐10: for 1% OMN + 26.3%; 95% CI –41.6% to 173.1%; P = 0.55 and + 36.1%; 95% CI –17.7% to 125.1%; P = 0.23 for IL‐6 and IL‐10, respectively; Figure 4 b: +88.4%; 95% CI –9.4% to 291.5%; P = 0.09 and +44.4%; 95% CI –48.1% to 302.4%; P = 0.48, for MXA and IFNɣ, respectively). Overall, the response induced by 1% OMN was more outspoken than the response to 2.5% OMN. IL‐8 was induced by IMQ but no enhancement was seen with OMN addition (data not shown). No effects of IMQ and OMN add‐on treatment were observed for the skin surface biomarkers by transdermal analysis patch, or on skin microbiome (data not shown).
Figure 4.
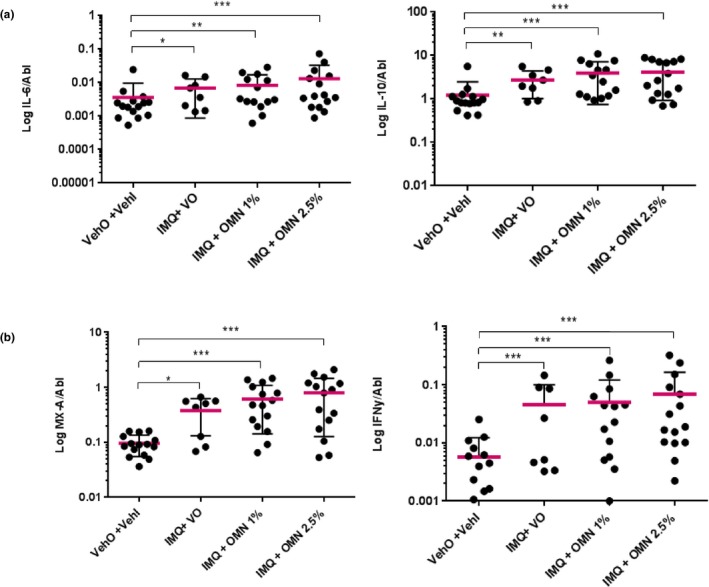
Skin inflammation induced by imiquimod (IMQ), omiganan (OMN), vehicle imiquimod (VehI) and vehicle omiganan (VehO) on day 5, as quantified by cytokine production (quantitative polymerase chain reaction) relative to ABL. (a) IL‐6 (left panel) and IL‐10 (right panel). (b) MXA (left panel) and IFNɣ (right panel). N = 8 for the IMQ + VehO contrast and N = 16 for the other contrasts.
Discussion
In human peripheral blood mononuclear cells, OMN enhances inflammatory responses driven by endosomal TLRs (unpublished data, Grievink et al.). OMN strongly increased type 1 IFN responses when cells were incubated with ligands for TLR3 (Poly:IC), TLR7 (IMQ), TLR8 (ssRNA), or TLR9 (CpG). IRF and NFκB pathways induced by these endosomal TLRs, drive tumoricidal and antiviral responses. Therefore, enhancement of endosomal TLR signaling in the skin may be of therapeutic interest for a variety of pathophysiological conditions. To investigate the clinical translation of OMN’s enhancement of endosomal TLR signaling, a healthy volunteer study was designed exploring the effects of IMQ combined with OMN add‐on treatment. This combination was well‐tolerated by the study participants, the main AE being mild application site pruritus, which was equal to the IMQ alone and OMN alone treatment groups. The clinical skin response was evaluated with LSCI (perfusion) and erythema assessments (colorimetry, erythema, and visual grading by the physician). Two days of IMQ treatment induced an inflammatory response similar as previously described,11 with erythema, increased perfusion, and increased inflammatory cell infiltration on histopathology lasting for at least 5 days. This effect was enhanced when IMQ was combined with OMN treatment. The influx of immune cells coincided with an increased cytokine response. IMQ induces an inflammatory response via TLR7‐driven IRF and NFκB signaling,8 which plays a role in a variety of dermal cells (T cells, keratinocytes, macrophages, Langerhans cells, and dendritic cells). In this study, OMN treatment increased the IMQ‐driven production of IL‐6 and IL‐10, reflecting NFκB activity. In addition, IRF‐driven pathways were enhanced: after application of OMN, elevated expression levels MXA were observed. MXA is a downstream mediator of interferons; its expression indicates an IFNα response.12 Moreover, OMN treatment increased type II interferon (IFNɣ) levels, which is mainly produced by T cells. Importantly, cellular and molecular responses were quantified in skin biopsies collected at day 5 (120 hours), where OMN (or vehicle) was applied at 0, 24, 48, and 72 hours. It could be contemplated that at earlier (uninvestigated) time points, the additive effect of OMN on immune responses was more outspoken, as observed for laser speckle and 2D photography data at time points 72–96 hours.
Our results relate to experimental conditions where skin of healthy human volunteers was first primed with IMQ, and subsequently treated with OMN. The reverse sequence was also studied, with OMN pretreatment for 2 days followed by 2 day’s application of IMQ. With this treatment sequence, the enhanced effects of OMN on IMQ responses were not observed (data not shown). This is in line with mechanistic in vitro experiments on human peripheral blood mononuclear cells, which suggest that coinciding exposure to OMN and endosomal TLR ligands result in the strongest immune response (unpublished data, Grievink et al.). Furthermore, OMN treatment alone did not induce any clinical, molecular, or cellular immune response (data not shown), which also corroborates with earlier peripheral blood mononuclear cell‐based experiments. It is hypothesized that the immune enhancing effects of OMN on endosomal TLR signaling requires a complex formation between the cationic peptide and the TLR ligand. Such complex formation has been demonstrated earlier, for example, between TLR9 ligand CpG and the bovine host defense peptide indolicidin, thereby enhancing innate and adaptive immune responses.13
The potentiating effect of OMN on IMQ induced responses, and potentially on the effect of other endosomal TLR ligands that are currently under development as immunostimulatory compounds, may be interesting from a drug development perspective. The effectiveness of IMQ treatment for HPV‐induced skin disease is suboptimal. In anogenital warts, for example, the estimated complete clearance is ~ 50%, with a recurrence rate of 13–19%. For HSIL, effectiveness of IMQ is estimated to be 58% with a 16% recurrence rate.10, 14, 15, 16 These data underline the need for enhanced treatment modalities. The combination treatment of IMQ with OMN may be considered as such. Although OMN’s effect size on top of IMQ‐induced responses was relatively small in our study, and no clear dose‐dependency for OMN was observed, our findings support the mechanistic concept of OMN‐dependent enhancement of endosomal TLR signaling. Thus, optimization of combined OMN/IMQ treatment seems to be a rational way forward.
For practical reasons, IMQ and OMN could only be administered as alternating treatments. Because a plausible mechanistic basis for OMN‐enhanced IMQ effects is the complex formation between TLR ligand and cationic peptide, it is not likely that pharmaceutical adjustments can be made to increase the desired effects. This may consist of optimization of the formulation containing a mixture of both compounds, or application of treatment regimens with rapid alternation of OMN and IMQ. Importantly, the observed enhanced IMQ responses by OMN co‐treatment also support further exploration of treatments combining OMN with other endosomal TLR ligands. The limitation is that currently no other endosomal TLR ligands besides IMQ are available for clinical application in the European Union. Rintatolimod, a TLR3 ligand, is only accessible via an early access program for chronic fatigue syndrome. Other interesting candidates for combined treatment with OMN include resiquimod, a TLR7/8 agonist, or one of the TLR9 agonists that are currently being evaluated in phase III clinical programs.
In summary, OMN enhanced the inflammatory skin response to IMQ, as studied in healthy volunteers with LSCI (perfusion), 2D photography (colorimetry, erythema, and visual grading), and analysis of molecular and cellular responses in skin biopsies. Figure 5 provides a graphical summary of key biomarkers, and underlines the OMN‐induced increase of IMQ‐driven responses. These findings are in line with the observations of enhanced endosomal TLR responses by OMN in in vitro experiments on primary human immune cells, and are supporting evaluation of IMQ/OMN combination therapy in HPV‐induced skin diseases, such as anogenital warts or HSIL.
Figure 5.
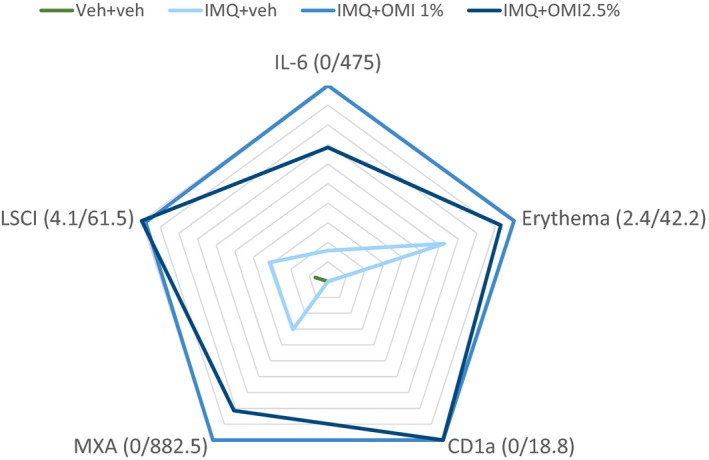
Graphical summary of key biomarkers: NF‐κB‐driven immune response (IL‐6), IRF‐driven immune response (MXA), perfusion (laser speckle contrast imaging (LSCI)), colorimetry (erythema), and immune cell infiltration (CD1a Langerhans cells). Responses were normalized to the maximal effects. Category labels indicate the actual minimum and maximum response. IMQ, imiquimod; OMI, omiganan; VehI, imiquimod vehicle; VehO, omiganan vehicle.
Funding
This study was co‐funded by Cutanea Life Sciences.
Conflict of Interest
One of the co‐authors (Gary Feiss) works at the co‐funder (Cutanea Life Sciences) of the trial. The final draft was approved by this co‐author on behalf of the co‐funder. However, no major comments were made. All other authors declared no competing interests for this work.
Author Contributions
T.N.v.d.K., S.A., T.P.B., M.R., E.S.K., G.F., E.P.P., J.B., M.B.A.v.D., R.R., and M.M. wrote the manuscript. T.N.v.d.K., S.A., E.S.K., G.F., E.P.P., J.B., M.B.A.v.D., R.R., and M.M. designed the research. T.N.v.d.K., S.A., T.P.B., M.R., E.F., E.P.P., J.B., M.B.A.v.D., R.R., and M.M. performed the research. T.N.v.d.K., S.A., E.S.K., G.F., E.F., E.P.P., J.B., M.B.A.v.D., R.R., and M.M. analyzed the data.
Acknowledgments
The authors thank Dr. Karen Broekhuizen for her careful review of the manuscript.
Study site: Centre for Human Drug Research, Leiden, The Netherlands.
References
- 1. Kosciuczuk, E.M. et al. Cathelicidins: family of antimicrobial peptides. A review. Mol. Biol. Rep. 39, 10957–10970 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Takiguchi, T. , Morizane, S. , Yamamoto, T. , Kajita, A. , Ikeda, K. & Iwatsuki, K. Cathelicidin antimicrobial peptide LL‐37 augments interferon‐beta expression and antiviral activity induced by double‐stranded RNA in keratinocytes. Br. J. Dermatol. 171, 492–498 (2014). [DOI] [PubMed] [Google Scholar]
- 3. Ganguly, D. et al. Self‐RNA‐antimicrobial peptide complexes activate human dendritic cells through TLR7 and TLR8. J. Exp. Med. 206, 1983–1994 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Fritsche, T.R. , Rhomberg, P.R. , Sader, H.S. & Jones, R.N. Antimicrobial activity of omiganan pentahydrochloride against contemporary fungal pathogens responsible for catheter‐associated infections. Antimicrob. Agents Chemother. 52, 1187–1189 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Fritsche, T.R. , Rhomberg, P.R. , Sader, H.S. & Jones, R.N. Antimicrobial activity of omiganan pentahydrochloride tested against contemporary bacterial pathogens commonly responsible for catheter‐associated infections. J. Antimicrob. Chemother. 61, 1092–1098 (2008). [DOI] [PubMed] [Google Scholar]
- 6. Hemmi, H. et al. Small anti‐viral compounds activate immune cells via the TLR7 MyD88‐dependent signaling pathway. Nat. Immunol. 3, 196–200 (2002). [DOI] [PubMed] [Google Scholar]
- 7. Lee, J. et al. Molecular basis for the immunostimulatory activity of guanine nucleoside analogs: activation of Toll‐like receptor 7. Proc. Natl. Acad. Sci. USA 100, 6646–6651 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Schon, M.P. & Schon, M. Imiquimod: mode of action. Br. J. Dermatol. 157 (suppl. 2), 8–13 (2007). [DOI] [PubMed] [Google Scholar]
- 9. Wagstaff, A.J. & Perry, C.M. Topical imiquimod: a review of its use in the management of anogenital warts, actinic keratoses, basal cell carcinoma and other skin lesions. Drugs 67, 2187–2210 (2007). [DOI] [PubMed] [Google Scholar]
- 10. Grillo‐Ardila, C.F. , Angel‐Muller, E. , Salazar‐Diaz, L.C. , Gaitan, H.G. , Ruiz‐Parra, A.I. & Lethaby, A. Imiquimod for anogenital warts in non‐immunocompromised adults. Cochrane Database Syst. Rev. (11), CD010389 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. van der Kolk, T. et al. Comprehensive, multi‐modal characterization of an imiquimod‐induced human skin inflammation model for drug development. Clin. Transl. Sci. 11, 607–615 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Haller, O. & Kochs, G. Human MxA protein: an interferon‐induced dynamin‐like GTPase with broad antiviral activity. J. Interferon Cytokine Res. 31, 79–87 (2011). [DOI] [PubMed] [Google Scholar]
- 13. Kovacs‐Nolan, J. , Mapletoft, J.W , Lawman, Z. , Babiuk, L.A & van Drunen Littel‐van den Hurk, S . Formulation of bovine respiratory syncytial virus fusion protein with CpG oligodeoxynucleotide, cationic host defence peptide and polyphosphazene enhances humoral and cellular responses and induces a protective type 1 immune response in mice. J. Gen. Virol. 90 (Pt 8), 1892–1905 (2009). [DOI] [PubMed] [Google Scholar]
- 14. Wiley, D.J. et al. External genital warts: diagnosis, treatment, and prevention. Clin. Infect. Dis. 35 (suppl. 2), S210–S224 (2002). [DOI] [PubMed] [Google Scholar]
- 15. Moore, R.A. , Edwards, J.E. , Hopwood, J. & Hicks, D. Imiquimod for the treatment of genital warts: a quantitative systematic review. BMC Infect. Dis. 1, 3 (2001). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Lawrie, T.A. , Nordin, A. , Chakrabarti, M. , Bryant, A. , Kaushik, S. & Pepas, L. Medical and surgical interventions for the treatment of usual‐type vulval intraepithelial neoplasia. Cochrane Database Syst. Rev. (11), CD011837 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]