

Abstract
Background
Insect bite hypersensitivity (IBH) is the most common seasonal pruritic allergic dermatitis of horses occurring upon insect bites. In recent years, a major role for IL‐31 in allergic pruritus of humans, monkeys, dogs, and mice was acknowledged. Here, we investigate the role of IL‐31 in IBH of horses and developed a therapeutic vaccine against equine IL‐31 (eIL‐31).
Methods
IL‐31 levels were quantified in allergen‐stimulated peripheral blood mononuclear cells (PBMCs) and skin punch biopsies of IBH lesions and healthy skin from IBH‐affected and healthy horses. The vaccine consisted of eIL‐31 covalently coupled to a virus‐like particle (VLP) derived from cucumber mosaic virus containing a tetanus toxoid universal T‐cell epitope (CuMVTT). Eighteen IBH‐affected horses were recruited and immunized with 300 μg of eIL‐31‐CuMVTT vaccine or placebo and IBH severity score was recorded.
Results
IL‐31 was increased in PBMCs and exclusively detectable in skin lesions of IBH‐affected horses. Vaccination against eIL‐31 reduced delta clinical scores when compared to previous untreated IBH season of the same horses and to placebo‐treated horses in the same year. The vaccine was well tolerated without safety concerns throughout the study.
Conclusion
TH2‐derived IL‐31 is involved in IBH pathology and accordingly the immunotherapeutic vaccination approach targeting IL‐31 alleviated clinical scores in affected horses.
Keywords: allergic pruritus, interleukin 31, vaccination
Equine IL‐31 is detectable in insect bite hypersensitivity (IBH) skin lesions upon insect bites and mediates pruritus by targeting peripheral nerves. IL‐31 is absent in skin biopsies from nonlesional or healthy skin. eIL‐31‐CuMVTT vaccine successfully induces autoantibodies against IL‐31 and reduces lesion scores in horses.
1. INTRODUCTION
The inflammatory interleukin 31 (IL‐31) belongs to the gp130/IL‐6 cytokine family and is associated with cellular immunity against pathogens and numerous chronic inflammatory diseases.1, 2, 3, 4 The main source of IL‐31 is type 2 helper T (TH2) cells, however, to a lesser extent, other sources such as mature dendritic cells (DCs) have also been described.5, 6 IL‐31 signaling is initiated when binding to its receptor complex composed of the IL‐31 receptor A (IL‐31RA) that heterodimerizes with the oncostatin M receptor β (OSMRβ). Downstream intracellular signaling involves the activation of Janus kinase‐signal transducer and activator of transcription (JAK‐STAT), mitogen‐activated protein kinase (MAPK), and phosphatidylinositol 3‐kinase (PI3K) pathways. Epithelial cells constitutively express IL‐31RA and OSMRβ, whereas monocytes only do when activated.4 Other cell types including keratinocytes, macrophages, and eosinophils also do express the IL‐31 receptor complex.4, 7, 8 Moreover, a subset of nociceptive neurons in the dorsal root ganglia (DRG) express the receptor, thus allowing a direct targeting of peripheral nerves by the immune system.6, 9 Hence, IL‐31 directly links the immune system and the nervous system. Indeed, it is well appreciated that IL‐31 is involved in the induction of pruritus leading to itchy skin in an allergic TH2 context, which, however, was shown to be independent of mast cell or basophil degranulation or proteinase‐activated receptor‐2 (PAR‐2).5, 6 Transgenic mice overexpressing IL‐31 develop strong pruritus, as well as skin lesions often with hair loss, histologically characterized by increased inflammatory cell infiltration.2 Intradermal injection of IL‐31 was further shown to induce itch and consecutive scratching in murine wild‐type (WT) skin.5 In addition, in humans IL‐31 expression was shown in skin‐homing CD45RO+ memory cutaneous lymphocyte‐associated antigen (CLA)‐positive T cells derived from patients with atopic dermatitis. Also, in patients with atopic dermatitis, 60% of TH2 cells abundant in the dermis were positive for IL‐31.5
Insect bite hypersensitivity (IBH) in horses is the most common skin allergy of horses and is caused by insect bites. Clinical signs typically occur along the dorsal and ventral bodyline. The skin lesions manifest as hair loss, excoriations, exsudations sometimes even ulcers and look eczema‐like, with hyperkeratotic scales, bloody crust formation and lichenification in chronic phases.10, 11, 12, 13, 14, 15, 16, 17 The disease is characterized by type I/IVb allergic reactions with accompanied eosinophil infiltration into the skin.18, 19, 20, 21 Upon insect bites, allergic horses suffer from strong pruritus leading to intense scratching and self‐inflicted skin destruction thereby amplifying the allergic symptoms. Previously, thymic stromal lymphopoietin (TSLP) and monocyte chemoattractant protein 1 (MCP‐1) (or CCL2) were associated with IBH disease pathology.18, 22, 23 The role of IL‐31 in pruritic IBH lesions of the horse remains elusive. Similar to our earlier described therapeutic vaccine targeting equine IL‐5 (eIL‐5) in order to reduce eosinophil production and infiltration into the skin, we developed a therapeutic vaccine targeting equine IL‐31 (eIL‐31). The earlier described virus‐like particle (VLP)‐based eIL‐5‐CuMVTT vaccine was able to reduce IBH disease symptoms and even could show enhanced improvement of disease scores in a second follow‐up treatment year.19, 24 Accordingly, we performed a placebo‐controlled double‐blind randomized clinical study in IBH‐affected horses to evaluate the therapeutic potential of targeting IL‐31. Our results suggest a prominent role for IL‐31 in IBH disease pathology further strengthened by mitigated disease symptoms in IL‐31 vaccinated horses when comparing to placebo‐treated horses.
2. MATERIALS AND METHODS
2.1. Horses
All horses were privately held client‐owned horses, and all horse owners signed informed consent. Insect bite hypersensitivity‐affected horses showed recurrent seasonal clinical signs of IBH at least one season prior to untreated first evaluation season; healthy horses were IBH symptom‐free at sample collection and had no history of IBH. All interventions and clinical studies were approved by the cantonal veterinary authorities.
2.2. Sample collection, PBMCs, and biopsies
Peripheral blood mononuclear cells (PBMCs) were collected from 19 IBH‐affected horses and three healthy horses. Biopsies were collected from seven IBH‐affected horses and four healthy horses. From IBH‐affected horses, one biopsy was taken from lesional and one from nonlesional skin.
2.3. Clinical study horses
2.3.1. Case study
A single IBH‐affected Icelandic horse was recruited to the case study. The study consisted of 2 years with a first observational untreated season (“untreated”) and a second season including vaccination (“vaccinated”). In both seasons, the horse health was checked daily and the IBH severity was scored at least monthly from March until October.
2.3.2. Double‐blind placebo‐controlled randomized trial
Eighteen IBH‐affected horses were recruited to a double‐blind placebo‐controlled randomized clinical study: nine horses received vaccine and nine horses received placebo. The study consisted of 2 years, the recruitment and observational year and the treatment year including double‐blind vaccinations. The clinical study was performed as double‐blind placebo‐controlled randomized trial, neither the person that performed the clinical trial nor the horse owners knew in which group the horses were. The independent vaccine filler has randomized the study and was not involved in any clinical part of the study. Prior to first vaccination and in a monthly fashion after second vaccination, serum was withdrawn for antibody titer measurement. Insect bite hypersensitivity lesions were scored monthly during observation period from March until October. Design of the clinical study is illustrated in Figure S1.
2.4. Allergen stimulation in vitro
Peripheral blood mononuclear cells were isolated using Biocoll gradient in complete RPMI Glutamax 1640 medium (Sigma‐Aldrich) and for 24 hours stimulated with whole body extract (WBE) of Culicoides nubeculosus (5 μg/mL,25 Greer), concanavalin A (ConA, 5 μg/mL, Sigma‐Aldrich), or medium. Cells were harvested by centrifugation, resuspended in RNA lysis buffer, and stored at −80°C for RNA isolation.
2.5. Punch biopsies
Punch biopsies (2 mm) from lesions of IBH‐affected horses and from nonlesional skin of IBH‐affected horses and healthy skin of healthy non‐IBH horses were collected into RNAlater™ Stabilization Solution (Thermo Fisher) for RNA extraction.
2.6. RNA extraction and qPCR
Total RNA was extracted using RNAqueous‐Micro Kit (Thermo Fisher) for punch biopsies and NucleoSpin® RNA XS Kit (Macherrey‐Nagel) for PBMCs. Extractions were performed according to the manufacturer's protocol including DNase I treatment and inactivation. RNA was transcribed into cDNA using Reverse Transcription System (Promega). All qPCR experiments were performed using FastStart Universal SYBR Green Master (Roche) with duplicate samples on a Viia7 Real‐Time PCR System (Thermo Fisher). Gene expression levels were normalized by β‐actin expression. Primers are listed in Table S1. IL‐4 and IL‐31 primer were designed by us, β‐actin,26 MCP‐1,27 and TSLP28 were previously published.
2.7. Cloning, expression, and purification of recombinant eIL‐31
The DNA sequence encoding for mature equine IL‐31 (UniProt F7AHG9) was generated by gene synthesis. In addition, a three amino acid linker (GGC) was added C‐terminally and termed eIL‐31‐C‐His. This insert was flanked by 5′ NdeI and 3′ XhoI and was integrated into pET 42b (+), containing a hexa‐His‐tag and an in‐frame stop codon. The resulting eIL‐31 fusion protein was expressed in Escherichia coli BL21 (T7 Express C2566I) cells. Cell culturing, induction, harvest, inclusion body preparation, and affinity tag purification were performed as described in Fettelschoss‐Gabriel et al.19 Subsequently, eIL‐31 was refolded by sequential dialysis against the following buffers at pH 8.5 at 4°C: B1 (2 M Urea, 50 mM NaH2PO4, 5 mM glutathione reduced, 0.5 mM glutathione oxidized, 0.5 M arginine, 10% glycerol), B2 (50 mM NaH2PO4, 5 mM glutathione reduced, 0.5 mM glutathione oxidized, 0.5 M arginine, 10% glycerol), B3a (50 mM NaH2PO4, 0.5 M arginine, 10% glycerol), B3b (50 mM NaH2PO4, 10% glycerol), and B4 (PBS). Finally, refolded protein was concentrated and purified on a HiLoad 26/600 Superdex 75 prep grade (GE Healthcare) with PBS buffer to separate monomers and dimers. Protein concentration was determined by Bradford assay to BSA standard.
2.8. Circular dichroism (CD) spectroscopy of eIL‐31‐C‐His
The far‐UV CD spectrum of purified monomeric and dimeric eIL‐31‐C‐His (in PBS) was measured on a J‐710 spectropolarimeter (Jasco) at 25°C using a 1‐mm cuvette. After correction for the buffer spectrum, ellipticity was converted to mean residue ellipticity as described.29
2.9. Coupling of eIL‐31 to CuMVTT
CuMVTT‐VLP reacted with a 7.5‐fold molar excess of the heterobifunctional cross‐linker succinimidyl‐6‐(β‐maleimidopropionamido)hexanoate (SMPH) in 20 mM NaP/2 mM EDTA, pH 7.5 at 25°C (Pierce). Unreacted cross‐linker was removed by passage over a PD‐10 desalting column (GE Healthcare). The recombinant, purified, and refolded monomeric and dimeric eIL‐31‐C‐His (1:1 ratio) were reduced for 1h with an equimolar amount of tri(2‐carboxyethyl)phosphine hydrochloride (TCEP) in 20 mM NaP/2 mM EDTA, pH 7.5. The reduced eIL‐31‐C‐His was then mixed with the derivatized CuMVTT‐VLPs at a molar ratio of 2:1 and co‐incubated for 4 hours at 22°C in 20 mM NaP/2 mM EDTA, pH 7.5. Vaccine was purified on a HiLoad 26/600 Superdex 75 prep grade (GE Healthcare) with 20 mM NaP/2 mM EDTA, pH 7.5.
2.10. Vaccine analysis by SDS‐PAGE, Coomassie staining, and Western Blot
Described in Fettelschoss‐Gabriel et al.19
2.11. Electron Microscopy (EM) of CuMVTT and eIL‐31‐CuMVTT
Samples were applied onto Formvar‐coated 300‐mesh Cu‐grids (Plano, Germany) at a concentration of 0.1 mg/mL and incubated for 1 minute at room temperature. Grids were stained with 1% uranyl acetate solution for 1 minute before being dried for 30 minutes. Stained grids were observed under a 100 kV CM100 transmission electron microscope (FEI). Imaging was performed by support of the Center for Microscopy and Image Analysis, University of Zurich.
2.12. IBH lesion scoring
Described in Fettelschoss‐Gabriel et al.19 The delta (Δ) lesion score was calculated for placebo‐treated and vaccinated horses as follows: For each horse and each measurement, value of observational season was subtracted from corresponding vaccine season value. Many of the horses were recruited in the second half of the observational year, therefore, we only include IBH lesion score data from June onwards in both the observational and the vaccination year.
2.13. Blood samples from horses
Blood was collected from V. jugularis, sterile blood collection for PBMCs with NH Sodium Heparin VACUETTE® container (Greiner Bio‐one) and nonsterile blood collection for serum with serum tubes (IDEXX Diavet), centrifuged (1500 g, 10 minutes), and transferred into fresh tubes.
2.14. Vaccine administration, immunization regimen
Horses were injected subcutaneously with 300 μg of eIL31‐C‐His‐CuMVTT‐VLP in 1000 μL of 20 mM NaP/2 mM EDTA, pH 7.5 without additional adjuvants. Horses received a prime‐boost vaccination in weeks 0 and 4 and a booster in week 19 (June).
2.15. Anti‐CuMVTT and anti‐IL‐31 antibody titer
Maxisorp 96‐well ELISA plates (Nunc) were coated overnight with purified eIL‐31‐C‐His or purified CuMVTT (5 mg/L) and continued as described in Fettelschoss‐Gabriel et al.19 Absorbance was measured at 450 nm by Tecan Spark 10M spectrophotometer (Tecan). The antibody titers as OD50 were calculated (serum dilution on a logarithmic scale where OD450 was half maximal). All antibody titers were calculated with naïve serum subtracted on logarithmic scales and presented as delta OD50 (ΔOD50). Titers ≤ 10 were considered background.
2.16. Statistics
All graphs comparing vaccinated horses vs placebo horses show mean and standard error of mean (SEM). Peripheral blood mononuclear cells: Statistical analysis was performed by 2‐way ANOVA and if significant followed by Tukey's multiple comparisons test. Skin Biopsies: Statistical analysis was performed by Kruskal‐Wallis test and if significant followed by Dunn's multiple comparisons test. Placebo‐controlled clinical trial: Statistical analysis was performed by two‐tailed Wilcoxon test comparing lesion scores per month during untreated and treated IBH season. Considered to be statistically significant were P‐values lower than .05: *P < .05; **P < .01; ***P < .001.
3. RESULTS
3.1. IL‐31 expression in PBMCs and skin of healthy and IBH‐affected horses
Peripheral blood mononuclear cells were isolated from IBH‐affected and healthy horses and stimulated with the IBH allergen C nubeculosus (Cul n) extract or medium control. Due to the lack of monoclonal antibodies, we performed qPCR for quantification of equine cytokines. Cul n stimulated PBMCs from IBH‐affected horses showed increased levels of IL‐31 when compared to Cul n stimulated PBMCs from healthy horses. Moreover, there was no difference of IL‐31 levels in healthy horses when stimulating with the allergen or medium control. Insect bite hypersensitivity‐affected PBMCs showed increased IL‐31 levels upon Cul n stimulation compared to medium stimulated cells (Figure 1A). Concanavalin A (Con A) stimulation triggered similar levels of IL‐31, IL‐4, and Stat5a RNA in IBH and healthy horses showing comparable quality of PBMCs (Figure 1B). Donor‐matched skin punch biopsies from IBH‐affected horses were taken from IBH lesions and from nonlesional skin. In addition, we collected biopsies from healthy skin in healthy horses. Equine cytokines IL‐31, TSLP, and MCP‐1 were quantified by qPCR in relation to housekeeping gene β‐actin. IL‐31 was solely detectable in IBH‐affected skin, whereas it was detectable neither in nonlesional skin of IBH horses nor in healthy skin of healthy horses (Figure 1C). In addition, we also found a trend toward higher levels of TSLP (Figure 1D) and MCP‐1 (Figure 1E) in IBH lesions when comparing to healthy skin of IBH or healthy horses.
Figure 1.
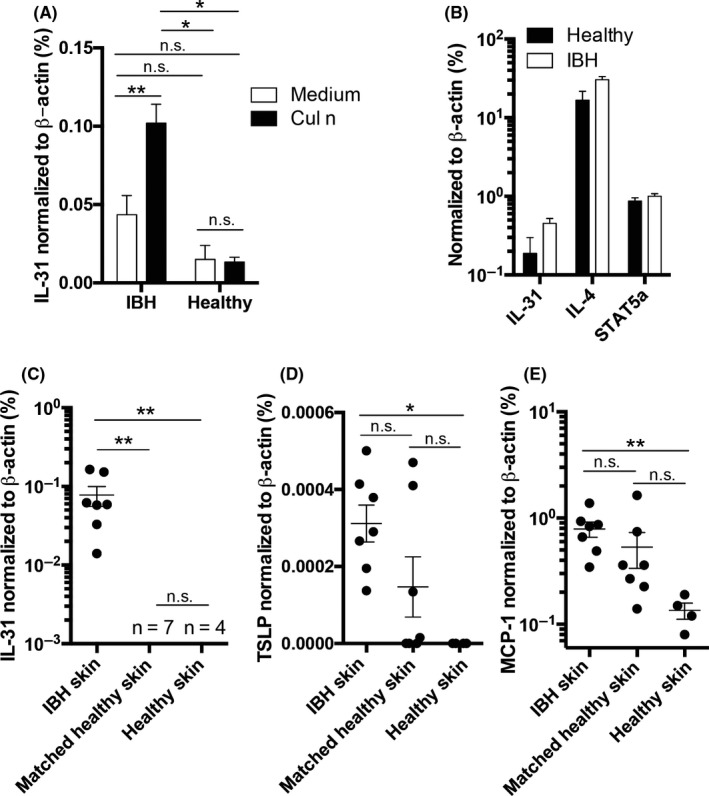
IL‐31 expression in IBH‐affected skin and allergen‐stimulated PBMCs. A and B, Culicoides nubeculosus (Cul n) (A) or Con A (B) allergen extract stimulated PBMCs from IBH‐affected horses (IBH, n = 19) and healthy horses (Healthy, n = 3). A, Percentage of eIL‐31 expression levels relative to eβ‐actin levels upon Cul n stimulation. B, Percentage of eIL‐31, eIL‐4, and Stat5a expression levels relative to eβ‐actin levels upon Con A stimulation. C, D, & E. Two mm skin punch biopsies from IBH‐affected skin (n = 7), healthy skin from IBH‐affected horses (n = 7), and healthy skin from healthy non‐IBH horses (n = 4). Quantification of eIL‐31 (C), eTSLP (D), and MCP‐1 (E) levels by qPCR shown as percent expression of eβ‐actin housekeeping gene, of skin biopsies taken from IBH lesion sites (n = 7), matched healthy skin from the same horses (n = 7), and healthy skin from healthy horses (n = 4)
3.2. IL‐31 and vaccine production
Recombinant eIL‐31 with a C‐terminal linker containing a free cysteine residue and a His‐tag (eIL‐31‐C‐His) was generated by gene synthesis for protein production in E coli. Purification and refolding steps are shown in Figure 2A. Far‐UV circular dichroism (CD) spectroscopy revealed mostly α‐helical secondary structures of the refolded eIL‐31‐C‐His indicated by the minimax points at 208 and 222 nm (Figure 2B).
Figure 2.
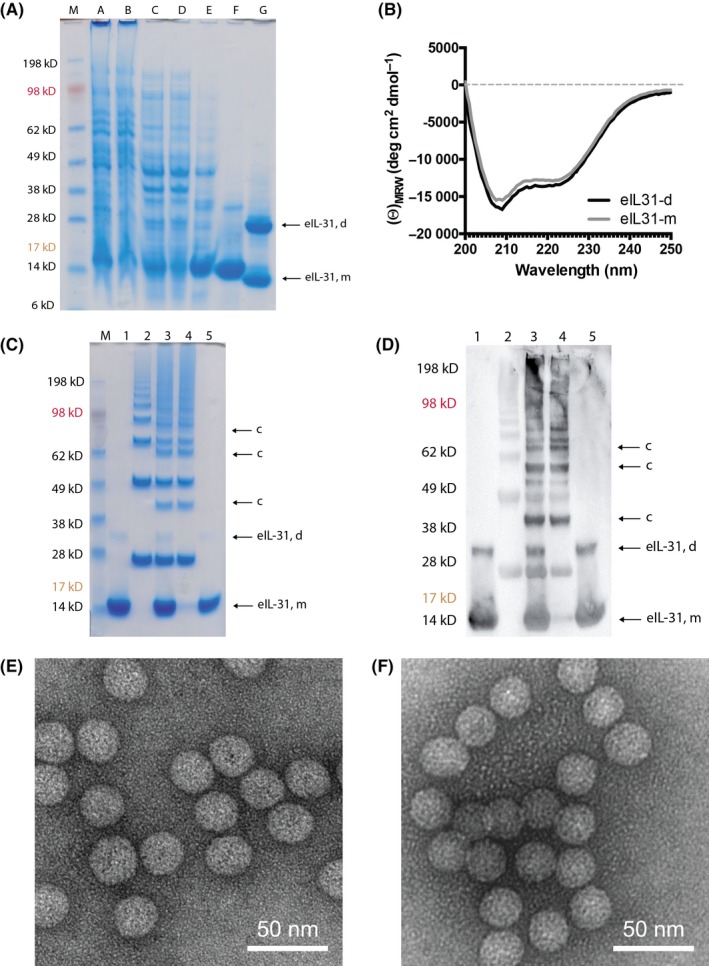
EIL‐31 and eIL‐31‐CuMVTT vaccine. A, SDS‐PAGE analysis of eIL‐31‐C‐His. Samples from various stages of the inclusion body preparation and purification were applied to a 4%‐12% B/T gel and run under reducing conditions. Proteins were stained with Coomassie blue. Lane M, size marker, lane 1, lysate (sample A), lane 2, soluble fraction (sample B), lane 3, solubilized inclusion bodies (sample C), lane 4, flow through (unbound material, sample D), lane 5, pooled eIL‐31 monomer (eIL‐31, m) eluate from Ni‐NTA column (sample E), lane 6, pooled eIL‐31 monomer and dimer eIL‐31 (eIL‐31, d) after refolding and size‐exclusion chromatography (sample F), lane 7, sample F run under native conditions (sample G). B, Far‐UV circular dichroism (CD) spectrum of purified eIL‐31‐C‐His monomer (eIL‐31‐m) and dimer (eIL‐31‐d). C & D, SDS‐PAGE of eIL‐31‐CuMVTT. Lane M, size marker, lane 1, TCEP activated eIL‐31‐C‐His, lane 2, CuMVTT‐VLP after derivatization with the chemical cross‐linker SMPH, lane 3, eIL‐31‐C‐His‐CuMVTT coupling reaction, lane 4, eIL‐31‐C‐His‐CuMVTT vaccine after size‐exclusion chromatography (SEC), lane 5, free eIL‐31 removed from eIL‐31‐C‐His‐CuMVTT vaccine during SEC. eIL‐31‐C‐His, CuMVTT, and eIL‐31‐CuMVTT were loaded equimolar in order to compare coupling efficacy. C, Proteins were stained with Coomassie blue. eIL‐31 monomer (eIL‐31, m), eIL‐31 dimer (eIL‐31, d), coupling (c). D, WB of eIL‐31‐CuMVTT. Stained with α‐His antibody. eIL‐31 monomer (eIL‐31, m), eIL‐5 dimer (eIL‐31, d), coupling (c). E, EM CuMVTT. F, EM eIL‐31‐CuMVTT
Chemical coupling of eIL‐31‐C‐His to CuMVTT‐VLPs was induced by a heterobifunctional cross‐linker. Derivatization of the VLP showed the typical “VLP‐ladder” caused by cross‐linking of monomeric and multimeric CuMVTT subunits (Figure 2C, lane 2). Coupling of CuMVTT subunits with eIL‐31‐C‐His molecules (Figure 2C, lane 1) led to additional coupling bands with a protein size corresponding to the molecular mass of monomeric or monomeric/dimeric eIL‐31‐C‐His plus monomeric or multimeric CuMVTT subunits (Figure 2C, lane 3). Successful covalent attachment of eIL‐31‐C‐His to CuMVTT was confirmed by Coomassie staining (Figure 2C) and Western blot using an anti‐His antibody (Figure 2D). Coupling efficiency for all batches was between 20% and 50%, that is, approximately 30‐75 eIL‐31 monomer and 15‐40 eIL‐31 dimer molecules per VLP (1:1 ratio of eIL‐31 monomer and dimer). A further vaccine polishing step by size‐exclusion chromatography was performed to remove free uncoupled eIL‐31‐C‐His from the vaccine (Figure 2C&D, lanes 4&5). Integrity of CuMVTT‐VLPs before and after coupling was analyzed and confirmed by EM (Figure 2E, CuMVTT, Figure 2F, eIL‐31‐CuMVTT).
3.3. Anti‐IL‐31 and anti‐CuMVTT antibody titers in horses
Antibody titers in serum of horses were evaluated monthly. Nine horses received a basic vaccination consisting of three immunizations, in weeks 0, 4, and 19 (June 1). Anti‐CuMVTT antibody titers of vaccinated horses were used as surrogate marker for successful vaccination as the immune system is expected to readily induce antibodies against the foreign CuMVTT particles. All nine vaccinated horses developed antibodies against both IL‐31 (Figure 3A&C) and CuMVTT (Figure 3B&D). Comparably to our previously published eIL‐5‐CuMVTT vaccine,19, 24 antibodies against eIL‐31 and CuMVTT can be observed already after the second vaccination, however, are not yet long‐lasting and a booster mid‐season was required (Figure 3A&C). Of note, upon booster a single horse did only re‐induce anti‐CuMVTT but not anti‐IL‐31 antibody titers.
Figure 3.
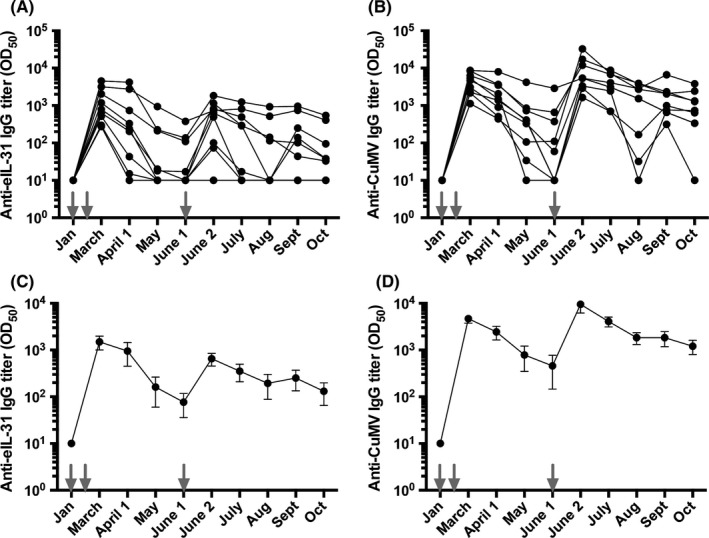
Antibody titer of vaccinated horses against eIL‐31 and CuMVTT‐VLP. Vaccinations are indicated by gray arrows. A, Antibody titer of anti‐eIL‐31 IgG of single horses. B, Antibody titer of anti‐CuMVTT IgG of single horses. C, Mean antibody titer of anti‐eIL‐31 IgG with SEM. D. Mean antibody titer of anti‐CuMVTT IgG with SEM. All antibody titers are calculated with naïve serum subtracted on logarithmic scales, and limit of detection is titers ≤10
3.4. Efficacy of eIL‐31‐CuMVTT vaccine by reduction of lesion scores
3.4.1. Case study
Lesion scores of a single IBH‐affected Icelandic horse were recorded in regular intervals in an untreated first IBH season (Figure 4A, untreated, gray) and a second IBH season with first year vaccination regimen using the eIL‐31‐CuMVTT vaccine (Figure 4A, vaccinated, black). The horse was vaccinated two times prior to IBH season start, which potentially delayed onset of IBH clinical signs. The booster vaccination in the middle of the IBH season further reduced IBH lesion scores during the vaccination year (Figure 4A). The anti‐IL‐31 antibody titer of that horse showed a strong, however, rapidly declining titer against IL‐31 and CuMVTT‐VLP after the second vaccination, and a booster effect upon mid‐season third vaccination, which also declined toward season end (Figure 4B).
Figure 4.
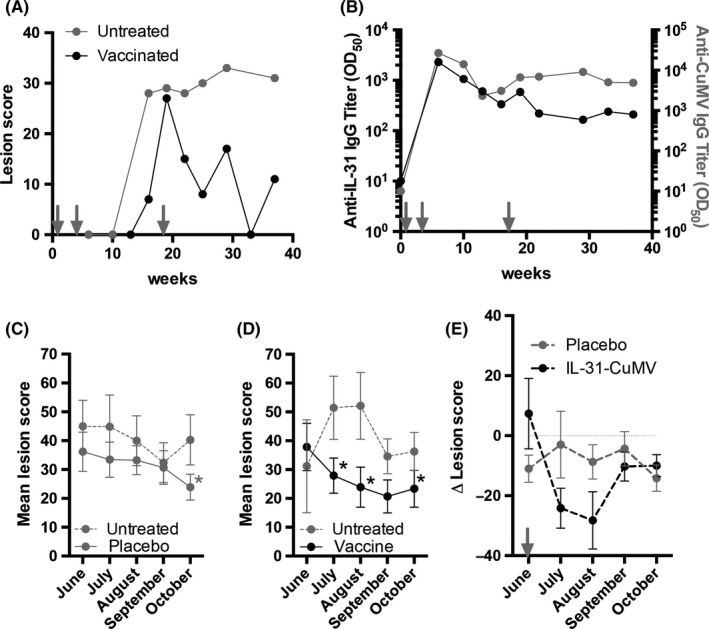
Efficacy of eIL‐31‐CuMVTT vaccine by reduction of lesion scores. A & B, Case report. A, IBH lesion scores of single horse during untreated previous season (gray) and vaccination season (black). B, Antibody titer against eIL‐31 (left y‐axis, black) and CuMVTT (right y‐axis, gray) of single horse. Vaccinations are indicated by arrows. C‐E, Double‐blind placebo‐controlled randomized clinical study. C, Monthly lesion score of placebo group during untreated (dotted gray line) and placebo‐treated (continuous gray line) season. Statistics is only indicated when significant. D, Monthly lesion score of vaccinated group during untreated (dotted gray line) and eIL‐31‐CuMVTT vaccinated (continuous black line) season. Statistics is only indicated when significant. E, Delta of monthly lesion scores after booster vaccination from previous IBH season subtracted by placebo‐ or vaccine‐treated season. Mid‐season booster indicated by arrow
3.4.2. Double‐blind placebo‐controlled randomized trial
Nine eIL‐31‐CuMVTT vaccine horses and nine placebo horses were scored monthly during a first observational year and a second year with first year vaccination regimen. In the first vaccination year, we established immunity against the vaccine by a total of three vaccine injections. Antibody titers are shown in Figure 3. We compared lesion scores from the third mid‐season injection onwards, that is, from June until October in untreated previous season and from June until October in vaccination season. No significant differences were found in the placebo group when comparing lesion scores from untreated and placebo‐treated season (Figure 4C). In contrast, significant differences were found in July and August in eIL‐31‐CuMVTT group when comparing scores from untreated and vaccine‐treated season (Figure 4D). In October, lesion scores of both groups improved independent of treatment when compared to the respective untreated seasons (Figure 4C&D). The delta lesion scores show the difference between both seasons. Mean delta lesion score of placebo horses was rather constant, indicating comparable clinical signs in both seasons (Figure 4E, gray). Mean delta lesion score of vaccinated horses was reduced upon booster vaccination in June, indicating improved clinical signs during vaccination season over untreated season (Figure 4E, black).
4. DISCUSSION
The present study demonstrates involvement of IL‐31 in the pathophysiology of IBH. In vitro allergen‐stimulated PBMCs from IBH‐affected horses and skin biopsies from IBH lesions showed a prominent role for IL‐31. In healthy horses, only low levels of IL‐31 were detectable in allergen‐stimulated PBMCs and moreover, IL‐31 was completely absent in skin biopsies from nonlesional or healthy skin. Moreover, we found a trend toward higher levels of TSLP and MCP‐1 in IBH lesions compared to healthy skin, confirming earlier suggested data.18, 22, 23 TSLP in the skin mainly derives from epithelial cells upon skin barrier disruption, MCP‐1 recruits monocytes, dendritic cells, and memory T cells to inflammatory tissue sites, thus both contributing to the exacerbation of allergy. In a case report and a placebo‐controlled double‐blind clinical trial, eIL‐31‐CuMVTT vaccinated IBH‐affected horses developed anti‐IL‐31‐specific antibodies and showed improved clinical signs compared to placebo‐treated IBH‐affected horses and previous untreated season. No safety concerns occurred throughout the clinical trials. Similar to the eIL‐5‐CuMVTT vaccine, two initial vaccinations of eIL‐31‐CuMVTT were sufficient to induce autoantibodies against IL‐31 and the third injection prolonged antibody titers throughout the IBH season.24 Anti‐CuMVTT antibody titers against the foreign VLP as surrogate marker demonstrate successful vaccination. In contrast, detection of anti‐IL‐31 antibodies could be masked by anti‐IL‐31 antibody/IL‐31 antigen complexes in the serum leading to different rates of anti‐IL‐31 antibody depletion. Along these lines, it is likely that anti‐IL‐31 antibody levels require a certain threshold to be protective rather than showing a linear dependency. Also, allergen exposure levels during the IBH season are not constant and vary with weather conditions and activity of the midges. For example, seasonal dynamics of the main trigger associated with IBH Culicoides spp. show highest activity in spring and in autumn30 leading to varying intensities of allergic symptoms including varying levels of associated effector molecules.
Insect bite hypersensitivity is defined as type I allergy with type IVb characteristics.18, 19, 20, 21 Type I pathology includes IgE‐mediated mast cell and basophil activation when allergen‐specific IgE binds to its Fc ε receptor type I (FcεRI), leading to histamine release from cellular granules.31 Nevertheless, the use of antihistamines did not show any efficacy against IBH and thus is not recommended for IBH therapy.32 This led to the assumption that there might be an additional causative mediator for the allergic itch. IL‐31 is a TH2 cell–derived cytokine, mediating pruritus by targeting peripheral nerves and acting independent of histamine.5
In humans, IL‐31 appears to be involved in the pruritus development of patients with several skin diseases such as atopic dermatitis, psoriasis or lupus erythematodes.33, 34, 35, 36 A monoclonal antibody (mAb) against human IL‐31 receptor (nemolizumab, Chugai Pharma) for the treatment of pruritus in atopic dermatitis entered clinical testing in 2012 and significantly reduced itching, however, did not improve eczema.37 A human double‐blind safety study of nemolizumab administered every 4 or 8 weeks for up to 64 weeks was well tolerated in patients with moderate‐to‐severe atopic dermatitis and was without any safety concerns. Furthermore, an anti‐IL‐31 mAb by Bristol‐Myers Squibb completed clinical phase 1 study in healthy subjects and adults with atopic dermatitis in 2015 and so far without safety concerns. In animals, Zoetis Inc developed an anti‐canine IL‐31 mAb (Lokivetmab, Cytopoint®), which recently entered the market as a treatment of atopic dermatitis in dogs. Such antibody treatment is expected to have fewer side effects than the currently used JAK inhibitor (oclacitinib, APOQUEL™). Regarding therapy costs and given the size and weight of a horse, systemically daily chemical inhibition of JAK or intravenous monthly monoclonal antibody injections, both applied based on body weight, limit the choice of therapeutics to the class of vaccines, which are applied at low dose and will be injected subcutaneously for three times in the first year and a single booster in the following years.24 Moreover, vaccines induce self‐made polyclonal active immunity with additive neutralization capacities over blocking a single epitope, that is, by passive immunity transfer of mAbs. In addition, mAbs commonly have a modified protein sequence adjusting the use to the target species such as humanization,thus, equine mAbs would need an “equinization.” However, 786 existing equine breeds (2006, FAO Global Data Bank) will turn this into a very complex task. Even in humans, humanization can be challenging and a common side effect is the development of anti‐therapeutic antibodies limiting the choice of therapeutics in affected individuals.38, 39
To our knowledge, we first describe a role for IL‐31 in IBH and the eIL‐31‐CuMVTT vaccine is the first therapy targeting IL‐31 in horses. It is the second therapeutic vaccine proposed for the use in IBH‐affected horses, besides our recently described eIL‐5‐CuMVTT vaccine.19, 24 While the eIL‐5‐CuMVTT vaccine targets the allergy‐associated hyper‐eosinophilia and thereby reduces the number of toxic cells potentially causing tissue damage of the skin, the eIL‐31‐CuMVTT vaccine rather targets the allergic itching caused by the underlying allergic immune response in the skin. Comparably to the mAb against IL‐31, we suggest that the vaccine‐induced anti‐IL‐31 antibodies mitigate the IL‐31‐mediated pruritus40, 41 and thus lowering the self‐inflicted trauma caused by the horse scratching its skin. When comparing the clinical benefit of blocking IL‐31 in horses with other species, we noticed a reduction of skin lesion severity in IBH‐affected horses, which is in contrast to atopic dermatitis trials in dogs and humans that only showed reduction of pruritus whereas no improvement of eczema.37, 42 This suggests a meaningful impact of the self‐inflicted trauma in horses with IBH. Although an anti‐IL‐31 therapy will not prevent the allergic reaction itself, it might represent an option stopping the vicious circle of self‐reinforcing pruritus to alleviate clinical symptoms.
Taken together, eIL‐31‐CuMVTT successfully induced autoantibodies against IL‐31 and simultaneously reduced lesion scores in vaccinated horses when compared to placebo‐treated horses. This is the second anticytokine vaccine that has shown clinical efficacy for the treatment of a disease in the target species horse and might facilitate the development of a similar vaccine targeting IL‐31 in human or other companion animal subjects. The next step will be the combination of the eIL‐5‐CuMVTT and eIL‐31‐CuMVTT vaccinations.
CONFLICT OF INTEREST
Dr Fettelschoss‐Gabriel reports grants from CTI during the conduct of the study, personal fees and nonfinancial support from Evax AG, during the conduct of the study; personal fees and non‐financial support from Evax AG, outside the submitted work; In addition, Dr Fettelschoss‐Gabriel has a patent WO 2017/042212 A1 licensed to Benchmark Animal Health Limited, and a patent WO 2018/162577 A1 pending. V Fettelschoss reports personal fees from Evax AG, during the conduct of the study and outside the submitted work; In addition, V Fettelschoss has a patent WO 2018/162577 A1 pending. Dr Thoms is an employee of HypoPet AG formerly Saiba Biotech GmbH. Dr Birkmann reports personal fees from Evax AG, outside the submitted work. Dr Bachmann reports personal fees from Evax AG, during the conduct of the study; personal fees and non‐financial support from Saiba GmbH, outside the submitted work; In addition, Dr Bachmann has a patent WO 2017/042212 A1 licensed to Benchmark Animal Health Limited, and a patent WO 2018/162577 A1 pending and MFB is an employee of the HypoPet AG formerly Saiba Biotech GmbH. Dr Kündig reports grants from CTI during the conduct of the study; personal fees from Evax AG, outside the submitted work. Dr Marti reports grants from Swiss National Science Foundation, grants from Morris Animal Foundation, grants from Stiftung Forschung für das Pferd, outside the submitted work. F Olomski has nothing to disclose. Dr Jonsdottir has nothing to disclose.
AUTHOR CONTRIBUTIONS
FO and VF planned, performed, and analyzed experiments. SJ planned and performed experiments. KB, FT, EM, MFB and TMK contributed to discussions. AFG planned and analyzed experiments, performed the clinical studies, interpreted the data, and wrote the manuscript.
Supporting information
ACKNOWLEDGMENTS
We thank all horse owners who participated with their horse(s) in our clinical studies.
Olomski F, Fettelschoss V, Jonsdottir S, et al. Interleukin 31 in insect bite hypersensitivity—Alleviating clinical symptoms by active vaccination against itch. Allergy. 2020;75:862–871. 10.1111/all.14145
Olomski and Fettelschoss equally contributed to this study.
Funding information
This project was supported by the Commission for Technology and Innovation (CTI Grant 25758.1 PFLS‐LS) and EVAX AG, Switzerland.
REFERENCES
- 1. Bagci IS, Ruzicka T. IL‐31: a new key player in dermatology and beyond. J Allergy Clin Immunol. 2018;141:858‐866. [DOI] [PubMed] [Google Scholar]
- 2. Dillon SR, Sprecher C, Hammond A, et al. Interleukin 31, a cytokine produced by activated T cells, induces dermatitis in mice. Nat Immunol. 2004;5:752‐760. [DOI] [PubMed] [Google Scholar]
- 3. Pflanz S, Hibbert L, Mattson J, et al. WSX‐1 and glycoprotein 130 constitute a signal‐transducing receptor for IL‐27. J Immunol. 2004;172:2225‐2231. [DOI] [PubMed] [Google Scholar]
- 4. Zhang Q, Putheti P, Zhou Q, Liu Q, Gao W. Structures and biological functions of IL‐31 and IL‐31 receptors. Cytokine Growth Factor Rev. 2008;19:347‐356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Cevikbas F, Wang X, Akiyama T, et al. A sensory neuron‐expressed IL‐31 receptor mediates T helper cell‐dependent itch: involvement of TRPV1 and TRPA1. J Allergy Clin Immunol. 2014;133:448‐460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Sonkoly E, Muller A, Lauerma AI, et al. IL‐31: a new link between T cells and pruritus in atopic skin inflammation. J Allergy Clin Immunol. 2006;117:411‐417. [DOI] [PubMed] [Google Scholar]
- 7. Kasraie S, Niebuhr M, Baumert K, Werfel T. Functional effects of interleukin 31 in human primary keratinocytes. Allergy. 2011;66:845‐852. [DOI] [PubMed] [Google Scholar]
- 8. Kasraie S, Niebuhr M, Werfel T. Interleukin (IL)‐31 induces pro‐inflammatory cytokines in human monocytes and macrophages following stimulation with staphylococcal exotoxins. Allergy. 2010;65:712‐721. [DOI] [PubMed] [Google Scholar]
- 9. Mizuno T, Kanbayashi S, Okawa T, Maeda S, Okuda M. Molecular cloning of canine interleukin‐31 and its expression in various tissues. Vet Immunol Immunopathol. 2009;131:140‐143. [DOI] [PubMed] [Google Scholar]
- 10. Anderson GS, Belton P, Kleider N. Culicoides obsoletus (Diptera: Ceratopogonidae) as a causal agent of Culicoides hypersensitivity (sweet itch) in British Columbia. J Med Entomol. 1991;28:685‐693. [DOI] [PubMed] [Google Scholar]
- 11. Baker KP, Quinn PJ. A report on clinical aspects and histopathology of sweet itch. Equine Vet J. 1978;10:243‐248. [DOI] [PubMed] [Google Scholar]
- 12. Braverman Y, Ungar‐Waron H, Frith K, et al. Epidemiological and immunological studies of sweet itch in horses in Israel. Vet Rec. 1983;112:521‐524. [DOI] [PubMed] [Google Scholar]
- 13. Fadok VA, Greiner EC. Equine insect hypersensitivity: skin test and biopsy results correlated with clinical data. Equine Vet J. 1990;22:236‐240. [DOI] [PubMed] [Google Scholar]
- 14. Greiner EC, Fadok VA, Rabin EB. Equine Culicoides hypersensitivity in Florida: biting midges aspirated from horses. Med Vet Entomol. 1990;4:375‐381. [DOI] [PubMed] [Google Scholar]
- 15. Kurotaki T, Narayama K, Oyamada T, Yoshikawa H, Yoshikawa T. Immunopathological study on equine insect hypersensitivity (“kasen”) in Japan. J Comp Pathol. 1994;110:145‐152. [DOI] [PubMed] [Google Scholar]
- 16. Quinn PJ, Baker KP, Morrow AN. Sweet itch: responses of clinically normal and affected horses to intradermal challenge with extracts of biting insects. Equine Vet J. 1983;15:266‐272. [DOI] [PubMed] [Google Scholar]
- 17. Schaffartzik A, Hamza E, Janda J, Crameri R, Marti E, Rhyner C. Equine insect bite hypersensitivity: what do we know? Vet Immunol Immunopathol. 2012;147:113‐126. [DOI] [PubMed] [Google Scholar]
- 18. Benarafa C, Collins ME, Hamblin AS, Cunningham FM. Role of the chemokine eotaxin in the pathogenesis of equine sweet itch. Vet Rec. 2002;151:691‐693. [PubMed] [Google Scholar]
- 19. Fettelschoss‐Gabriel A, Fettelschoss V, Thoms F, et al. Treating insect‐bite hypersensitivity in horses with active vaccination against IL‐5. J Allergy Clin Immunol. 2018;142(4):1194‐1205.e3. [DOI] [PubMed] [Google Scholar]
- 20. McKelvie J, Foster AP, Cunningham FM, Hamblin AS. Characterisation of lymphocyte subpopulations in the skin and circulation of horses with sweet itch (Culicoides hypersensitivity). Equine Vet J. 1999;31:466‐472. [DOI] [PubMed] [Google Scholar]
- 21. Rf R. Studies on allergic dermatitis (Queensland itch) of the horse I. Description, distribution, symptoms and pathology. Aust Vet J. 1953;28:177‐184. [Google Scholar]
- 22. Klumplerova M, Vychodilova L, Bobrova O, et al. Major histocompatibility complex and other allergy‐related candidate genes associated with insect bite hypersensitivity in Icelandic horses. Mol Biol Rep. 2013;40:3333‐3340. [DOI] [PubMed] [Google Scholar]
- 23. Vychodilova L, Matiasovic J, Bobrova O, et al. Immunogenomic analysis of insect bite hypersensitivity in a model horse population. Vet Immunol Immunopathol. 2013;152:260‐268. [DOI] [PubMed] [Google Scholar]
- 24. Fettelschoss‐Gabriel A, Fettelschoss V, Olomski F, et al. Active vaccination against interleukin‐5 as long‐term treatment for insect‐bite hypersensitivity in horses. Allergy. 2019;74:572‐582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Peeters LM, Janssens S, Goddeeris BM, et al. Evaluation of an IgE ELISA with Culicoides spp. extracts and recombinant salivary antigens for diagnosis of insect bite hypersensitivity in Warmblood horses. Vet J. 2013;198:141‐147. [DOI] [PubMed] [Google Scholar]
- 26. Bogaert L, Van Poucke M, De Baere C, Peelman L, Gasthuys F, Martens A. Selection of a set of reliable reference genes for quantitative real‐time PCR in normal equine skin and in equine sarcoids. BMC Biotechnol. 2006;6:24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Benarafa C, Cunningham FM, Hamblin AS, Horohov DW, Collins ME. Cloning of equine chemokines eotaxin, monocyte chemoattractant protein (MCP)‐1, MCP‐2 and MCP‐4, mRNA expression in tissues and induction by IL‐4 in dermal fibroblasts. Vet Immunol Immunopathol. 2000;76:283‐298. [DOI] [PubMed] [Google Scholar]
- 28. Klukowska‐Rotzler J, Marti E, Lavoie JP, et al. Expression of thymic stromal lymphopoietin in equine recurrent airway obstruction. Vet Immunol Immunopathol. 2012;146:46‐52. [DOI] [PubMed] [Google Scholar]
- 29. Milburn MV, Hassell AM, Lambert MH, et al. A novel dimer configuration revealed by the crystal structure at 2.4 Å resolution of human interleukin‐5. Nature. 1993;363:172‐176. [DOI] [PubMed] [Google Scholar]
- 30. Sanders CJ, Shortall CR, Gubbins S, et al. Influence of season and meteorological parameters on flight activity of Culicoides biting midges. J Appl Ecol. 2011;48:1355‐1364. [Google Scholar]
- 31. Prussin C, Metcalfe DD. 4. IgE, mast cells, basophils, and eosinophils. J Allergy Clin Immunol. 2003;111:S486‐S494. [DOI] [PubMed] [Google Scholar]
- 32. Olsen L, Bondesson U, Brostrom H, et al. Pharmacokinetics and effects of cetirizine in horses with insect bite hypersensitivity. Vet J. 2011;187:347‐351. [DOI] [PubMed] [Google Scholar]
- 33. Huang HT, Chen JM, Guo J, Lan Y, Wei YS. The association of interleukin‐31 polymorphisms with interleukin‐31 serum levels and risk of systemic lupus erythematosus. Rheumatol Int. 2016;36:799‐805. [DOI] [PubMed] [Google Scholar]
- 34. Lu J, Wu K, Zeng Q, Xiang Y, Gao L, Huang J. Serum interleukin‐31 level and pruritus in atopic dermatitis: a meta‐analysis. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2018;43:124‐130. [DOI] [PubMed] [Google Scholar]
- 35. Nattkemper LA, Tey HL, Valdes‐Rodriguez R, et al. The genetics of chronic itch: gene expression in the skin of patients with atopic dermatitis and psoriasis with severe itch. J Invest Dermatol. 2018;138:1311‐1317. [DOI] [PubMed] [Google Scholar]
- 36. Siniewicz‐Luzenczyk K, Stanczyk‐Przyluska A, Zeman K. Correlation between serum interleukin‐31 level and the severity of disease in children with atopic dermatitis. Postepy Dermatol Alergol. 2013;30:282‐285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Ruzicka T, Hanifin JM, Furue M, et al. Anti‐interleukin‐31 receptor A antibody for atopic dermatitis. N Engl J Med. 2017;376:826‐835. [DOI] [PubMed] [Google Scholar]
- 38. Steenholdt C, Bendtzen K, Brynskov J, Thomsen OØ, Ainsworth MA. Measurement of infliximab and anti‐infliximab antibody levels can help distinguish maintenance versus loss of response. Gastroenterol Hepatol. 2012;8:131‐134. [PMC free article] [PubMed] [Google Scholar]
- 39. van Schie KA, Kruithof S, van Schouwenburg PA, et al. Neutralizing capacity of monoclonal and polyclonal anti‐natalizumab antibodies: the immune response to antibody therapeutics preferentially targets the antigen‐binding site. J Allergy Clin Immunol. 2017;139:1035‐1037.e6. [DOI] [PubMed] [Google Scholar]
- 40. Gonzales AJ, Fleck TJ, Humphrey WR, et al. IL‐31‐induced pruritus in dogs: a novel experimental model to evaluate anti‐pruritic effects of canine therapeutics. Vet Dermatol. 2016;27:34‐e10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Lewis KE, Holdren MS, Maurer MF, et al. Interleukin (IL) 31 induces in cynomolgus monkeys a rapid and intense itch response that can be inhibited by an IL‐31 neutralizing antibody. J Eur Acad Dermatol Venereol. 2017;31:142‐150. [DOI] [PubMed] [Google Scholar]
- 42. Tamamoto‐Mochizuki C, Paps JS, Olivry T. Proactive maintenance therapy of canine atopic dermatitis with the anti‐IL‐31 lokivetmab. Can a monoclonal antibody blocking a single cytokine prevent allergy flares? Vet Dermatol. 2019;30:98‐e26. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials