

Graphical abstract
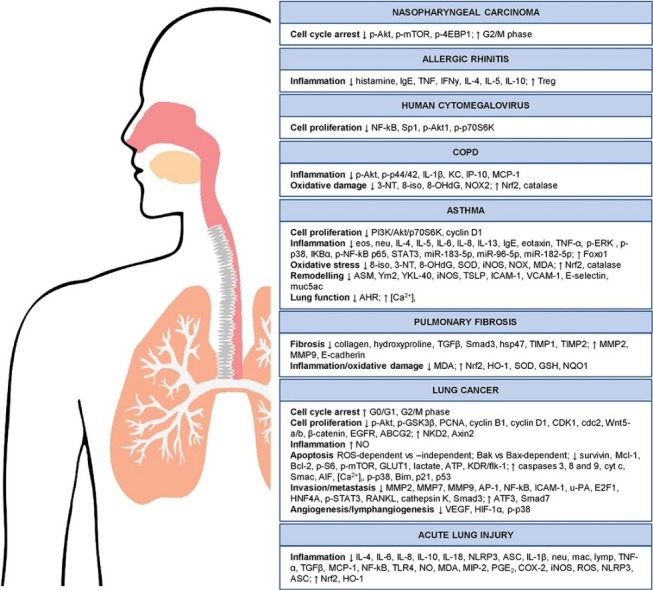
Molecular targets modulated by artemisinins in respiratory diseases.
Abbreviations: ABCG2, ATP-binding cassette subfamily member 2; ACE2, angiotensin-converting enzyme 2; AEC, alveolar epithelial cells; AHR, airways hyperresponsiveness; AIF, apoptosis-inducing factor; ALI, acute lung injury; AP-1, activator protein 1; ARTD, artemisinin-daumone hybrid 15; ASC, apoptosis-associated speck-like protein containing CARD; ATF3, activating transcription factor 3; ATP, adenosine triphosphate; Axin2, axis inhibition protein 2; BALF, bronchoalveolar lavage fluid; BDHA, biotinylated dihydroartesunate; CDDP, cisplatin; CDK, cyclin-dependent kinase; COPD, chronic obstructive pulmonary disease; COX-2, cyclooxygenase-2; CQ, chloroquine; HCQ, hydroxychloroquine; COVID-19, coronavirus disease 2019; CSE, cigarette smoke extract; CTX, cyclophosphamide; DHA, dihydroartemisinin; DHA-NLC, dihydroartemisinin-nanostructured lipid carriers; DLAedried, leaf artemisia extract; DNA, deoxyribonucleic acid; EGFR, epidermal growth factor receptor; EMT, epithelial-mesenchymal transition; eos, eosinophil; ER, endoplasmic reticulum; E2F1, E2F transcription factor 1; Foxo1, forkhead box O1; GLUT, glucose transporter; GPx, glutathione peroxidase; GSH, glutathione; GSK3β, glycogen synthase kinase 3 beta; H2AX, H2A histone family member X; HCMV, human cytomegalovirus; HDAC2, histone deacetylase 2; HELF, human embryonic lung fibroblasts; HIF-1α, hypoxia-inducible factor 1-alpha; HNF4A, hepatocyte nuclear factor 4 alpha; HO-1, heme oxygenase-1; hsp47, heat shock protein 47; ICAM-1, intercellular adhesion molecule 1; IκBα, inhibitor of NF-κB alpha; IL, interleukin; INF, interferon; iNOS, inducible nitric oxide synthase; IP-10, IFNγ-induced protein 10; JNK, c-Jun N-terminal kinase; KC, keratinocyte chemoattractant; KDR/flk-1, kinase insert domain receptor /fetal liver kinase-1; Keap1, kelch-like ECH-associated protein 1; LADPI, liposomal artesunate dry powder inhalers; LD50, lethal dose; LLC, lewis lung carcinoma; LMVD, lymphatic microvessel density; LPS, lipopolysaccharide; Lymp, lymphocyte; Mac, macrophage; MAPK, mitogen-activated protein kinases; Mcl-1, myeloid cell leukemia-1; MCP-1, monocyte chemoattractant protein-1; MDA, malondialdehyde; MIP-2, macrophage inflammatory protein 2; MMP, matrix metalloproteinase; MPO, myeloperoxidase; mTOR, mammalian target of rapamycin; neu, neutrophil; NF-κB, nuclear factor-kappa B; NKD2, naked cuticle homolog 2; NLRP3, NLR family pyrin domain containing 3; NO, nitric oxide; NOX, NADPH oxidase; NPC, nasopharyngeal carcinoma; NQO1, NAD(P)H quinone dehydrogenase 1; Nrf2, nuclear factor erythroid 2-related factor 2; NSCLC, non-small cell lung cancer; OVA, ovalbumin; PCNA, proliferating cell nuclear antigen; PEG, polyethylene glycol; PGE2, prostaglandin E2; PI3K, phosphoinositide 3-kinase; RA, FLS rheumatoid arthritis fibroblast-like synoviocytes; RANKL, receptor activator of nuclear factor kappa-B ligand; Rb, retinoblastoma; RIR, renal ischemia reperfusion; RNAi, RNA interference; ROS, reactive oxygen species; SCLC, small cell lung cancer; shRNA, short hairpin RNA; SLE, systemic lupus erythematosus; sm-α, actin smooth muscle-α actin; Smac, second mitochondrial activator of caspases; SOD, superoxide dismutase; STAT, signal transducers and activators of transcription; TGF, tumour growth factor; Th17, T helper 17; TIMP, tissue inhibitor of metalloproteinases; TLR4, toll-like receptor 4; TNF, tumour necrosis factor; Treg, regulatory T; TSLP, thymic stromal lymphopoietin; u-PA, urokinase-type plasminogen activator; XIAP, X-linked inhibitor of apoptosis protein; VCAM-1, vascular cell adhesion molecule 1; VDAC2, voltage-dependent anion channel 2; VEGF, vascular endothelial growth factor; YKL-40, chitinase-like glycoprotein; Ym2, chitinase 3-like protein 4; 2DG, 2-Deoxy-d-glucose; 3-NT, 3-nitrotyrosine; 4EBP1, eukaryotic translation initiation factor 4E-binding protein 1; 8-iso, 8-isoprostane; 8−OHdG, 8-hydroxy-2′-deoxyguanosine; [Ca2+]i, intracellular calcium ion
Keywords: Respiratory diseases, Artemisinin, Lung, Cell proliferation, Inflammation, COVID-19, Chemical compounds studied in this article Artemisinin (PubChem CID: 68827), Dihydroartemisinin (PubChem CID: 139073990), Artesunate (PubChem CID:156252), Artemisitene (PubChem CID: 11000442), Artemether (PubChem CID: 68911), Arteether (PubChem CID: 3000469)
Abstract
Artemisinins are sesquiterpene lactones with a peroxide moiety that are isolated from the herb Artemisia annua. It has been used for centuries for the treatment of fever and chills, and has been recently approved for the treatment of malaria due to its endoperoxidase properties. Progressively, research has found that artemisinins displayed multiple pharmacological actions against inflammation, viral infections, and cell and tumour proliferation, making it effective against diseases. Moreover, it has displayed a relatively safe toxicity profile. The use of artemisinins against different respiratory diseases has been investigated in lung cancer models and inflammatory-driven respiratory disorders. These studies revealed the ability of artemisinins in attenuating proliferation, inflammation, invasion, and metastasis, and in inducing apoptosis. Artemisinins can regulate the expression of pro-inflammatory cytokines, nuclear factor-kappa B (NF-κB), matrix metalloproteinases (MMPs), vascular endothelial growth factor (VEGF), promote cell cycle arrest, drive reactive oxygen species (ROS) production and induce Bak or Bax-dependent or independent apoptosis. In this review, we aim to provide a comprehensive update of the current knowledge of the effects of artemisinins in relation to respiratory diseases to identify gaps that need to be filled in the course of repurposing artemisinins for the treatment of respiratory diseases. In addition, we postulate whether artemisinins can also be repurposed for the treatment of COVID-19 given its anti-viral and anti-inflammatory properties.
1. Use of natural products for the treatment of respiratory diseases
Respiratory diseases refer to any disease or disorder of the airways and the lungs that interfere with respiration [1]. The respiratory system, comprising of the nose, nasal cavities and the lungs, is the sole internal system that is exposed to the external environment. Hence, it is easily susceptible to environmental agents, such as bacterial or viral infections, smoking, air pollution or cold weather [2], that can cause respiratory diseases. Importantly, chronic respiratory diseases are a great cause of concern as approximately one billion people suffer from them while four million succumb to these illnesses prematurely every year [2]. The Forum of International Respiratory societies identified chronic obstructive pulmonary disorder (COPD), asthma, acute respiratory infections, and lung cancer as the top few respiratory diseases that heavily burden society [2]. In brief, COPD is an obstructive lung disease characterized by long term breathing issues and poor airflow. It affects 200 million people and is the fourth leading cause of death worldwide [3]. It is currently being treated using inhaled bronchodilators and glucocorticoids [4]. Asthma is a condition in which the airways narrow and swell, and is accompanied by increased mucus production. It affects 300 million people worldwide [5]. The cause of asthma is still unknown, and it is furthermore uncurable. Treatment is limited to symptom relief using inhaled corticosteroids and bronchodilators [5]. Acute respiratory infections include pneumonia and viral respiratory infections. Annually, respiratory tract infections like influenza kill 250,000–500,000 people and cost billions of dollars. In addition, it occasionally causes epidemics that threaten the health of the global population [6]. Lung cancer is a malignant lung tumour that is characterized by uncontrolled cell growth in the lung tissues, and this growth can spread to other parts of the body, causing death. It has the highest fatality rate amongst the major cancers, killing more than 1.4 million people a year [7]. Patients are diagnosed and their disease is classified into different stages where earlier-stage patients are treated with surgery to remove the lung tumour, while late-stage patients are treated with chemotherapy or radiotherapy but often succumb to the disease [2]. Currently, the global approach to managing respiratory diseases is to provide better healthcare, reduce environmental pollution, and to create public awareness of the prevalence and risks of such diseases. Research in this field explores the causes of these respiratory diseases, prognostic markers to better diagnose patients and new therapies that can maintain and contain the disease [2]. Nonetheless, a lot more work will need to be done to find safer and more effective treatment methods.
Natural products have been used to treat respiratory diseases as far back as 2600BCE with the first records indicating that oils of Cedrus (cedar), Commiphora (myrrh), Cupressus sempervirens (cypress) and Glycyrrhiza glabra (licorice) were being used to treat inflammation, coughs, and colds [8]. Male newborns of the Indian tribes of southern California were bathed in hot Salvia ashes as it was believed to provide lifetime immunity from all respiratory diseases [9]. In 1952, erythromycin, derived from Saccharopolyspora erythraea, was launched commercially for bacterial infections affecting the upper respiratory tract [10]. Umckaloabo contains root extract of Pelargonium sidoides and was marketed in 1897 against tuberculosis but was later superseded by antibiotics. In the 2000s, it regained popularity for the treatment of acute bronchitis and is now one of the most commonly prescribed childhood medications [11]. Today, many natural-based products are still being investigated for its beneficial properties against respiratory diseases. In this review, we will provide a comprehensive update of the current knowledge of artemisinin, and its derivatives, for the treatment of various respiratory diseases.
2. Artemisinins chemical classification
Artemisinin is a sesquiterpene lactone with a peroxide constituent [12]. It is isolated from the leafy parts of Artemisia annua, a herb and medicinal plant that has been used for the treatment of chills and fever for centuries [13]. In the 1960s, the search for new anti-malarial drugs began in lieu of the increasing resistance of Plasmodium falciparum to chloroquine (CQ). Artemisinin, also known as Qinghaosu, was first isolated. Dihydroartemisinin (DHA) was subsequently the first generation of derivatives, made by modifying the carbonyl groups into hydroxyl groups [12]. Others like the more water-soluble artesunate and more oil-soluble artemether and arteether followed [14]. These derivatives were ten times more potent than artemisinin [12], with artesunate having a more favourable pharmacokinetic-pharmacodynamic profile [15]. They are also more easily produced [12] (Table 1 ). Artemisinins and its derivates are selectively taken up by parasites-infected erythrocytes and later localized in the parasite membranes, including that of the mitochondria, digestive vacuole and the parasite limiting membrane [12,14]. All forms of the drug contain an endoperoxide bridge (C-O-O-C) that is crucial for its anti-malarial activity, where the compound itself is catalyzed by heme or iron to form free radicals. These free radicals then alkylate malaria membrane-associated proteins, killing the parasite [14]. Artemisinin and its derivatives are found to be effective against different severities of malaria, especially those resistant to traditional gold standard drugs. They are highly efficacious, requiring only nanomolar concentrations in vitro [14]. They are also fast-acting, showing therapeutic potential as early as 20 h after administration. Moreover, artemisinins display a relatively safe toxicity profile, with the LD50 being 4223 mg/kg. In addition, whilst there was some evidence for neurotoxicity in neuronal cells and animals at high dosages, this was never reported in humans despite the wide usage of the drug [12,14].
Table 1.
Artemisinin derivatives and their differences.
| Derivatives | Chemical Structure | Structural modifications | Route of administration | Pharmacokinetics | Other Properties/ Comments | Ref. |
|---|---|---|---|---|---|---|
| Artemisinin | 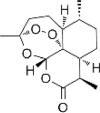 |
– | Oral | Tmax 1–3 h Vd 1420–1560 L Clearance 445–479 L/h T0.5 2.3 h |
– | [15,97] |
| i.m. | Tmax 3.4 h | [15] | ||||
| Rectal | Tmax 5.6 h | |||||
| DHA (1st generation) |
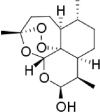 |
Carbonyl to hydroxyl group | Oral | Tmax 0.9–1.6 h | DHA pharmacokinetics are often measured following administration of artesunate as DHA is the active metabolite | [15] |
| Rectal | Tmax 4 h | |||||
| Artesunate (2nd generation) |
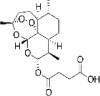 |
Reacted DHA with succinic acid anhydride | Oral | Bioavailability 61–88 % Tmax 15–39 mins Vd 14.8 L/kg Clearance 20.6 L/kg/h T0.5 0.36–1.2 h |
Water-soluble | [15,97] |
| i.m. | Bioavailability 86.4–88 % Tmax 7.2–12 mins Vd 1.09–3.98 L/kg Clearance 2.4–3.48 L/kg/h T0.5 25.2–48.2 mins |
[98] | ||||
| Rectal | Bioavailability 54.9 % Tmax 0.58–1.43 h T0.5 0.9-0.95 h |
[98] | ||||
| i.v. | Cmax 13000–16000 ng/ml Vd 0.1–0.3 L/kg Clearance 2–3 L/kg/h |
[98] | ||||
| Artemether (2nd generation) |
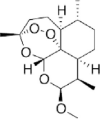 |
Methyl ether derivative of DHA | Oral | Tmax 1.7−6 h | Oil-soluble | [15] |
| i.m | Tmax 1.3−8.7 h | |||||
| Rectal | Tmax 3.1 h | |||||
| Arteether (2nd generation) |
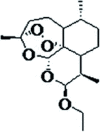 |
Ethyl ether derivative of DHA | i.m. | Tmax 4.8–7 h T0.5 12.4–30.2 h |
Oil-soluble | [15] |
| Artemisitene (2nd generation) |
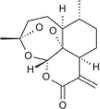 |
Oxidized form of artemisinin | Oral | Bioavailability (in rats) 3.7% Cmax 511 ng/ml Tmax 0.01 min Vd 40 L/kg Clearance 0.239 L/min/kg T0.5 91 min |
Novel Nrf2 activator, compared to artemisinin | [66,99] |
| i.v. | Bioavailability (in rats) 3.7 %Cmax511 ng/mL Tmax 0.01 min Vd 40 L/kg Clearance 0.239 L/min/kg T0.5 91 min |
|||||
| DLAe | – | Extracts from the dried leaves of artemisia plants | n.d. | n.d. | Less costly than artesunate | [34] |
| ARTD | 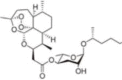 |
Glycolipid daumone group hybridized to artemisinin via covalent coupling | n.d. | n.d. | Enhanced bioavailability with less drug resistance | [58] |
| Compound 17 | 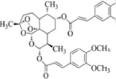 |
9α–OH DHA, prepared by biotransformation of DHA and cinnamic acid derivatives | n.d. | n.d. | IC50 = 0.2 μM, lower than that of DHA (80.42 μM) or 5-FU (6.76 μM) | [35] |
| BDHA | 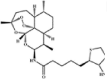 |
Biotinylated DHA | Likely similar to DHA | Likely similar to DHA | Used to identify cellular protein targets | [67] |
i.m.: intramuscular, i.v: intravenous, Cmax: maximum serum concentration of drug, Tmax: time Cmax is observed, Vd: Volume of distribution, T0.5: Elimination half-life, n.d.: no data.
Apart from its anti-malarial effects, artemisinin and its derivatives also exhibited additional properties in other diseases. For example, artesunate had anti-cancer effects as shown by its cytotoxic activity against 55 cancer cell lines through its regulation of various processes, including DNA damage and repair, apoptosis, and proliferation [16,17]. Artesunate displayed anti-inflammatory properties, as seen by its attenuation of the production of interleukin (IL)-1β, IL-6 and IL-8 in tumour necrosis factor (TNF)-α-stimulated rheumatoid arthritis fibroblast-like synoviocytes (RA FLS) via the regulation of NF-κB and phosphoinositide 3 kinase (PI3K) pathways [18]. It also displayed anti-viral properties where artemisinin inhibited the replication of human cytomegalovirus (HCMV) through a reduction in the DNA binding activity of NF-κB and Sp1, and subsequently downstream activities of Akt1 and p70S6K. [19]. Many of these pathophysiological processes are also present in respiratory diseases. Thus, artemisinin and its derivatives could potentially be repurposed for the treatment of respiratory diseases as well.
3. Artemisinin and its derivatives (effects in vitro)
The effects of artemisinin and its derivatives have been examined in various in vitro models (Tables 2 5 ) and these include: inhibition of cell proliferation; inductions of cell cycle arrest and apoptosis; inhibition of inflammation and oxidative stress; inhibition of angiogenesis, invasion and metastasis, and chemosensitization of cancer cells to chemotherapeutic agents.
Table 2.
Effects of artemisinins and its derivatives on cell proliferation, tumour growth, invasion and metastasis.
| Disease model | Cell line(s)/ stimulus/ allergen/ animal type | Derivative | Effective conc./ dose; route of administration | Outcomes | Ref. | |
|---|---|---|---|---|---|---|
| invitro | Asthma | ASM | Artesunate | 3−30 μM | ↓ cell number, p-Akt, p-p70S6K, cyclin D1 | [31] |
| Lung cancer | A549 | Artemisinin | 250−1000 μM | ↓ colony formation | [20] | |
| Artesunate | 100−150 μM | |||||
| A549 | DHA | 10−60 μM | ↓ colony formation | [21] | ||
| PC-9 | 8−64 μM | |||||
| A549, H1299 | DHA/ Arteminsinin/ Artesunate | 7.5−30 μM | ↓ cyclin D1, Wnt5-a/b, LRP6, Dvl2, β-catenin, invasion, migration, EMT, CSCs; ↑ G1 phase cell cycle arrest, NKD2, Axin2 | [22] | ||
| A549 | DHA | 10−30 μM | ↑ sub-G1 and G1 phase cell cycle arrest, p21; ↓ cyclin D1, PCNA, p-Akt, p-GSK3β | [23] | ||
| A549 | DHA | 3.2−1000 nmol/L | ↑ population doubling time, G0/G1 phase cell cycle arrest | [24] | ||
| PC-14 | DHA | 5−320 μM | ↓ cell proliferation | [25] | ||
| ASTC-a-1 | DHA | 1−30 μg/mL | ↓ cell proliferation | [28] | ||
| A549, NCI-H661, SK-MES-1, Spc-A-1 | DHA | 2.5 μM | ↓ IC50; Chemosensitize with onconase | [29] | ||
| A549 | Artesunate | 10−20 μg/L | ↓ cell proliferation, invasion | [30] | ||
| A549 | Artesunate | 75 μM | ↑ sub-G1 population; Chemosensitize with 50 μM CQ | [33] | ||
| DLAe | 50 μM | ↑ G2/M, p-H2AX; ↓ migration | [34] | |||
| H1299 | DLAe | 50−75 μM | ↑ G2/M, p-H2AX | [34] | ||
| A549 | DLAe | 100−150 μM | ↑ G1, p-H2AX; ↓ migration | [34] | ||
| A549 | Compound 17 | 0.2−30 μM | ↓ cell proliferation | [35] | ||
| LLC | DHA | 10−40 μg/mL | Chemosensitize with 25 μg/mL carboplatin | [36] | ||
| GLC-82 | DHA | 4−128 μg/mL | ↑ G0/G1 phase cell cycle arrest; ↓ S phase; Promote radiosensitization | [37] | ||
| H1975 | DHA | 10 μM | ↓ cyclin B1 and CDK1, migration and invasion; Chemosensitize with 10 μM gefitinib | [38] | ||
| A549 | DHA | 10−30 μg/mL | ↑ G2/M phase cell cycle arrest; Synergistic with low-dose ionising radiation (2 or 4 Gy) | [39] | ||
| A549 | Artesunate | 50−1600 μM | ↑ NO, G2/M phase cell cycle arrest; ↓ cyclin B1 and cdc2 mRNA; Radiosensitize with local radiotherapy | [40] | ||
| A549 | Artesunate | 1−100 μM | No ↓ in cell viability, ↓ cyclin D, CDK4, p-Rb; blockade at all cell cycle phases; Chemosensitize with 0.1−1 μM lentilomide | [42] | ||
| HCT116 | Artesunate | 1−100 μM | ↓ cell viability, cyclin D, CDK4, p-Rb; ↑ sub-G1, p21 | [42] | ||
| MCF7 | Artesunate | 1−100 μM | No ↓ in cell viability, ↓ cyclin D, CDK4, p-Rb; ↑ p21; blockade at all cell cycle phases (>30 μM), ↑ G1 and ↓ S phase (<30 μM); Chemosensitize with 0.1−1 μM lentilomide | [42] | ||
| H1975 | DHA | 15 μM | Chemosensitize with 2 μM ABT-263 | [48] | ||
| LLC | DHA | 20−80 μmol/L | ↓ cell proliferation | [49] | ||
| H460, H1299, Calu3, LXF289, A549, H1398 | Artesunate | 2.5 μM | ↓ cell proliferation, AP-1 activity, matrigel invasion | [54] | ||
| A549, H1975 | DHA | 7.5−30 μM | ↓ cyclin D1, migration and invasion | [55] | ||
| A549 | DHA | 0.71−11.36 mg/L (A549) 1.42−22.72 mg/L (A549/DDP) |
Chemosensitize with 0.46875−7.5 mg/L (A549) or 0.9375−15 mg/L (A549/DDP) cisplatin | [56] | ||
| A549 | ARTD | 2.5−7.5 μM | ↓ cell proliferation, E2F1, migration and invasion | [58] | ||
| A549 | DHA | 10 μg/mL | Chemosensitize with 10 μg/mL doxorubicin | [59] | ||
| A549, ASTC-a-1 | DHA | 20 μg/mL | Chemosensitize with 1−20 μM JNK inhibitor SP600125 | [60] | ||
| A549 | DHA | 6−12 μg/mL | Chemosensitize with 100−500 μM dictamine | [61] | ||
| A549 | DHA | 30−90 μM | ↓ S phase; ↑ G2/M phase cell cycle arrest in combination; Chemosensitize with 10−60 μM arsenic trioxide | [62] | ||
| A549 | DHA | 10−20 μg/mL | ↑ G2/M and sub-G1 phase cell cycle arrest; ↓ G0/G1 arrest; Chemosensitize with 1−10 μg/mL gemcitabine | [63] | ||
| ASTC-a-1, 95D, H446 | DHA | 5−40 μg/mL | ↓ cell proliferation; Chemosensitize with 1−10 μg/mL gemcitabine | [63] | ||
| A549 | Artesunate | 25–100 μM | ↓ cell proliferation | [83] | ||
| SCLC | H69, H69VP | Artemisinin | 2−20 nM | Pre-treatment with 880 nM transferrin ↓ IC50 | [64] | |
| EMT | TGFβ1-induced EMT RLE-6TN |
Artesunate | n.d. | ↓ EMT | [82] | |
| HCMV | HCMV infection in HELF | Artesunate | 12.5−400 μM | ↓ NF-κB, Sp1, p-Akt1, p-p70S6K | [32] | |
| ALI | LPS-induced ALI in A549 |
Artesunate | 1−4 μM | No change | [52] | |
| Nasopharyngeal carcinoma | C666−1, HONE-1, HK1, HNE1, CNE2 | Artesunate | 10−40 μM | ↑ G2/M phase cell cycle arrest, cyclin B1, p-Akt, p-mTOR, p-4EBP1; ↓ Rb, E2F-1; Chemosensitize with 10 μM cisplatin |
[41] | |
| in vivo | Lung cancer | A549; nude mice | DHA/ Artemisinin/ Artesunate | 60 mg/kg/day; gavage | ↓ tumour growth, β-catenin, oct3/4, sox2, nanog, vimentin, Wnt5-a/b, LRP6, Dvl2; ↑ NKD2, Axin2 | [22] |
| A549; nude mice | DHA | 100 mg/kg/day; oral gavage | ↓ tumour growth | [24] | ||
| A549; BALB/c nude mice | DHA | 10 mg/kg; i.p. | ↓, chemosensitize with 3 mg/kg onconase | [29] | ||
| A549; nu/nu mice | DLAe | 85 mg/kg; p.o. gavage | ↓ tumour growth | [34] | ||
| A549; nude mice | Artesunate | 7.5−30 mg/kg once; i.m. | Combination with local radiotherapy ↓ tumour growth | [40] | ||
| H1975; nude mice | DHA ABT-263 |
25 mg/kg 100 mg/kg/day; oral gavage |
Combination therapy ↓ tumour growth by >51 % | [48] | ||
| LLC; C57BL/6 mice | DHA | 50−100 mg/kg/day; i.g. | ↓ tumour growth; Chemosensitize with 50 mg/kg/day CTX to ↓ pulmonary metastasis | [49] | ||
| A549; BALB/c mice | DHA | 50−200 mg/kg/day; i.g. | ↓ tumour growth; Chemosensitize with 2 mg/kg/day CDDP | [49] | ||
| A549; nude mice | DHA | 50−100 mg/kg/day | ↓ metastasis | [55] | ||
| LLC; C57BL/6 mice | Artemisinin | 50 mg/kg/day; orally | No change in tumour growth; ↓ lung metastatic nodules, lymph node metastases | [57] | ||
| A549; BALB/c mice and ovariectomized mice | ARTD | 10−20 mg/kg; oral gavage | ↓ cancer-associated bone metastasis, E2F1; ↑ ATF3 | [58] | ||
| A549; BALB/c athymic nude mice | Artesunate | 60−120 mg/kg/day; oral gavage | ↓ p-EGFR, p-Akt, Akt, ABCG2 | [83] | ||
| Metastasis assay | H460; chicken embryo | Artesunate | i.v. | ↓ tumour growth and metastasis | [54] | |
| NSCLC | 120 advance stage patients | Artesunate | 120 mg/day; i.v. | Chemosensitize with vinorelbine and cisplatin therapy to ↑ time to progression, and disease controlled rate | [84] | |
| NPC | C666−1 or CNE2; SCID mice | Artesunate | 100 mg/kg/day; i.p. | ↓ tumour growth; Synergistic with 40 mg/kg/day cisplatin | [41] |
n.d.: no data.
Table 5.
Effects of artemisinins and its derivatives on other features.
| Feature | Disease model | Cell line(s)/ stimulus/ allergen/ animal type | Derivative | Effective conc./ dose; route of administration | Outcomes | in vitro/ in vivo | Ref. |
|---|---|---|---|---|---|---|---|
| Lung function | Asthma | Mouse/ human ASM cells | Artesunate | 0.75−2 mM | ↓ traction force; ↑ [Ca2+]i | in vitro | [78] |
| Asthma (airway inflammation) | CSE or OVA; BALB/c mice | Artesunate | 30 mg/kg; i.p. | ↓ AHR | in vivo | [50] | |
| Allergic asthma | OVA; BALB/c mice | Artesunate | 3−30 mg/kg; i.p. | ↓ AHR | [74] | ||
| Allergic asthma | OVA; BALB/c mice | DHA | 30 mg/kg/day; i.g. | ↓ AHR | [75] | ||
| Asthma | OVA; BALB/c mice | DHA | 50 mg/kg/day; i.p. | ↓ AHR | [76] | ||
| Asthma | OVA; BALB/c mice | Artesunate | 30−120 μg | ↓ AHR | [78] | ||
| Allergic rhinitis | OVA; BALB/c mice | Artemisinin | 10−100 mg/kg/day; nasally | ↓ sneezing, nasal rubbing | [77] | ||
| Angiogenesis | Tumour lymphangiogenesis | LLC | Artemisinin | 5−20 μM | ↓ VEGF-C | in vitro | [57] |
| Lung cancer | A549; BALB/c nude mice | DHA | 10 mg/kg; i.p. | ↓ VEGF, microvessel density | in vivo | [29] | |
| Lung cancer | A549 and A549/DDP; BALB/c athymic mice | DHA | 50, 100, 200 mg/kg/day; i.g. | ↓ HIP-1α, VEGF, tumour microvessel density | [56] | ||
| Cisplatin | 2 mg/kg/3 days, 12 (A549) or 28 (A54/DDP) days; i.p. | ||||||
| Lung cancer | LLC; C57BL/6 mice | Artemisinin | 50 mg/kg/day; orally | ↓ LMVD, VEGF-C, p-p38 | [57] | ||
| Fibrosis/ remodelling | Lung cancer | H460, H1299, Calu3, LXF289, A549, H1398 | Artesunate | 2.5 μM | ↓ u-PA, MMP2, MMP7 and NF-κB activities | in vitro | [54] |
| EMT | TGFβ1-induced EMT RLE-6TN |
Artesunate | n.d. | ↓ p-Smad3, Smad3; ↑ Smad7 | [82] | ||
| Pulmonary fibrosis | AEC | DHA | 5−7 mol/L | ↓ sm-α actin | [65] | ||
| Allergic asthma | OVA; C57BL/6 mice | Artesunate | 30 mg/kg; i.p. | ↓ sm-α actin, cyclin D1 | in vivo | [31] | |
| Allergic asthma | OVA; BALB/c mice | Artesunate | 3−30 mg/kg; i.p. | ↓ muc5ac, mucus hypersecretion | [74] | ||
| Allergic asthma | OVA; BALB/c mice | DHA | 30 mg/kg/day; i.g. | ↓ muc5ac | [75] | ||
| Asthma | OVA; BALB/c mice | DHA | 50 mg/kg/day; i.p. | ↓ mucus | [76] | ||
| Pulmonary fibrosis | Bleomycin; Sprague Dawley rats | DHA | 50 mg/kg/day; i.p. | ↓ alveolitis, fibrosis, sm-α actin, MDA; ↑ E-cadherin | [65] | ||
| Pulmonary fibrosis | Bleomycin; Sprague Dawley rats | DHA | 25−100 mg/kg/day; i.p. | ↓ hydroxyproline, TGFβ1, TNF-α, sm-α actin, NF-κB | [79] | ||
| Pulmonary fibrosis | Bleomycin; Sprague Dawley rats | Artesunate | 100 mg/kg/day; i.p. | ↓ hydroxyproline, TGFβ1, Smad3, hsp47, sm-α actin, collagen I | [80] | ||
| Pulmonary fibrosis | Bleomycin; Sprague Dawley rats | Artesunate | 100 mg/kg/day; i.p. | ↓ collagen-IV, TIMP1, TIMP2; ↑ MMP2, MMP9 | [81] | ||
| Lung cancer | A549; BALB/c nude mice | Artesunate | 10−40 mg/kg/day; i.p. | ↓ ICAM-1, MMP9 | [30] | ||
| Lung cancer | A549; BALB/c mice and ovariectomized mice | ARTD | 10−20 mg/kg; oral gavage | ↓ RANKL, MMP9, cathepsin K | [58] | ||
| Metastasis assay | H460; chicken embryo | Artesunate | i.v. | ↓ u-PA, MMP2, MMP7 | [54] | ||
| Oxidative stress | Lung cancer | A549, H1975 | DHA | 7.5−30 μM | ↓ Warburg effect, NF-κB, c-myc, GLUT1 | in vitro | [55] |
| Pulmonary fibrosis | Bleomycin; Sprague Dawley rats | DHA | 50 mg/kg/day; i.p. | ↑ Nrf2, HO-1, SOD, GSH | in vivo | [65] | |
| Others | Asthma | CSE/ OVA-induced airway inflammation in BEAS-2B | Artesunate | 10 μM | Reversed glucocorticoid (Dex 10−11-10-6 M) insensitivity | in vitro | [50] |
| Lung cancer | A549 | DHA | 10−60 μM | ↓ glucose uptake, glycolysis (ATP and lactate), glycolytic metabolism (p-mTOR, GLUT1) | [21] | ||
| PC-9 | 8−64 μM | ||||||
| Lung cancer | A549, ASTC-a-1 | DHA | 20 μg/mL | ↑ | [46] | ||
| Lung cancer | A549 | ARTD | 2.5−7.5 μM | ↓ HNF4A; ↑ ATF3 | [58] | ||
| ALI | LPS; BALB/c mice | Artesunate | 10−40 mg/kg; i.v. | ↓ lung wet-to-dry ratio | in vivo | [52] | |
| ALI | RIR; Sprague Dawley rats | Artesunate | 15 mg/kg; i.p. | ↓ lung wet-to-dry ratio | [53] | ||
| Pulmonary fibrosis | Bleomycin; Sprague Dawley rats | Artesunate | 100 mg/kg/day; i.p. | ↓ mortality | [80] |
n.d.: no data.
3.1. Inhibition of cell proliferation
The anti-proliferative effect of artemisinin and its derivatives are observed in a variety of lung cancer cell lines, including the non-small cell lung cancer (NSCLC) cell lines – lung adenocarcinoma A549 [[20], [21], [22], [23], [24]], PC-9 [21], PC-14 [25], H1299 [22], ASTC-a-1 [[26], [27], [28]] and Spc-A-1 [29] cells, squamous carcinoma SK-MES-1 cells and large cell lung cancer NCI-H661 cells [29]. Interestingly, one study noted that low concentrations (1.25−5 μg/L) of artesunate were unable to prevent the proliferation of A549 cells [30], suggesting that there may be a therapeutic window by which the drug would have anti-proliferative effects.
In addition, artesunate was found to have anti-proliferative effects in non-cancer cell types. Pre-treatment with artesunate reduced mitogen-stimulated increases in cyclin D1 protein expression and cell number in both asthmatic and non-asthmatic human cultured airway smooth muscle (ASM) cells. This effect was mediated by reductions in p-Akt and p-p70S6K protein expressions, which were not observed with dexamethasone treatment [31]. In HCMV, artesunate, but not ganciclovir, reduced the proliferation rates of infected human embryonic lung fibroblasts (HELF) [32]. It is worthy to note that they did not have cyotoxic effects on healthy, non-diseased cells, such as in normal human lung fibroblast WI-38 cells [21,33], non-cancerous human dermal fibroblasts CCD-1108Sk cells [34] and normal hepatic l-02 cells [35]. This finding lends support for the desired clinical property of arteminismins in regimes where inhibiton of cell growth is needed (such as in the cancer setting) without affecting the healthy, non-disease state condition.
3.2. Induction of cell cycle arrest
Various studies have shown that artemisinins induce cell cycle arrest at different phases in lung and nasopharyngeal cancer. Artemisinin, artesunate, and DHA inhibited cell proliferation in A549 and H1299 cells via cell cycle arrest in the G1 phase [22], with corresponding downregulation of p-Akt, p-glycogen synthase kinase 3 beta (GSK3β) as well as both mRNA and protein expressions of proliferating cell nuclear antigen (PCNA) and cyclin D1 [23]. DHA induced G0/G1 cell cycle arrest in Lewis lung carcinoma (LLC) cells and GLC-82 human lung cancer cells, with a reduction in the number of cells in the S and G2/M phases [36,37]. Chen and colleagues reported that DHA inhibited A549 cell proliferation in a concentration-dependent manner after 96 h of treatment, with an increase in the number of G0 and G1 cells. The population doubling time in DHA-treated cells also lengthened as compared to the control group (38.5 vs 21.3 h) [24]. On the other hand, DHA induced G2/M phase cell cycle arrest in NCI-H1975 human lung adenocarcinoma cells with a marked reduction in the protein expressions of cyclin B1 and cyclin-dependent kinase (CDK)1 [38]. Together with 24 h of ionizing radiation treatment, DHA induced a G2/M phase arrest that disappeared 36 h after treatment [39]. Similarly, artesunate enhanced the radiosensitivity of A549 cells with cell cycle arrest at the G2/M phase, with lower cyclin B1 mRNA levels and a heightened nitric oxide (NO) production [40]. G2/M phase arrest was also induced with artesunate in nasopharyngeal carcinoma (NPC) cells, and this effect was synergistic with cisplatin, with a reduction in phosphorylation of Akt, mTOR, and 4EBP1 [41]. Artesunate treatment reduced cell numbers in A549, HCT116, and MCF7 cells but did not reduce cell viability in A549 and MCF7 cells, suggesting a cytostatic effect. Interestingly, the sub-G1 population increased in HCT116 cells but there were no significant changes in the cell cycle phases in A549 and MCF7 cells, suggesting a simultaneous blockade at all cell cycle phases [42]. Artemisinin delivered as dried leaf artemisia extract (DLAe) induced G1 cell cycle arrest in A549 cells but induced G2/M arrest in PC-9 and H1299 cells [34]. Collectively, these studies show that whilst artemisinins bring about cell cycle arrest, caution should be taken into the study of artemisinins on cell cycle arrest as they affect different stages of the cell cycle and the effects may be cell-type dependent.
3.3. Pro-apoptotic effect
The apoptotic effects of artemisinins and its derivatives are largely observed in lung cancer cell lines and have been found to induce both the intrinsic and extrinsic pathways of apoptosis (Table 3 ).
Table 3.
Effects of artemisinins and its derivatives on apoptosis.
| Disease model | Cell line(s)/ stimulus/ allergen/ animal type | Derivative | Effective conc./ dose; route of administration | Outcomes | Ref. | |
|---|---|---|---|---|---|---|
| invitro | Lung cancer | A549 | Artemisinin | 250-1000 μM | ↑ cell death, LDH, ROS, DNA damage | [20] |
| Artesunate | 100-150 μM | |||||
| A549 | DHA | 10−60 μM | ↑ ROS, caspases 3, 8 and 9, cytochrome c and AIF when synergize with 2DG | [21] | ||
| PC-9 | 8−64 μM | |||||
| A549 | DHA | 10−30 μM | ↑ Bax, caspase 3, cytochrome c; ↓ Bcl-2 | [23] | ||
| PC-14 | DHA | 5−320 μM | ↑ [Ca2+]i, p-p38 | [25] | ||
| ASTC-a-1 | Artemisinin | 200−500 μM | ↑ ROS, caspases 3, 8 and 9 | [26,27] | ||
| ASTC-a-1 | DHA | 1−30 μg/mL | ↑ change in mitochondrial morphology, caspase 3; ↓ ΔΨm | [28] | ||
| A549, NCI-H661, SK-MES-1, Spc-A-1 | DHA | 2.5 μM | ↓ endothelial tube formation | [29] | ||
| A549 | Artesunate | 75 μM | ↑ ROS, cytochrome c, and cleaved caspase 3 | [33] | ||
| DLAe | 50 μM | ↑ caspases 3, 8 and 9 | [34] | |||
| H1299 | DLAe | 50−75 μM | ↑ caspases 3, 8 and 9 | [34] | ||
| A549 | DLAe | 100−150 μM | ↑ caspases 3, 8 and 9 | [34] | ||
| A549 | Compound 17 | 0.2−30 μM | ↑ ROS | [35] | ||
| LLC | DHA | 10−40 μg/mL | ↑ p-p38 | [36] | ||
| GLC-82 | DHA | 4−128 μg/mL | ↑ p53, p21; ↓ Bcl-2 | [37] | ||
| H1975 | DHA | 10 μM | ↓ p-Akt, p-mTOR, p-STAT3, and Bcl-2; ↑ Bax | [38] | ||
| A549 | DHA | 10−30 μg/mL | ↑ ROS, caspases 3, 8 and 9, tBid translocation, Bax; ↓ Bcl-xL | [39] | ||
| A549 | Artesunate | 50−1600 μM | No effect | [40] | ||
| ASTC-a-1 | DHA | 20 μg/mL | ↑ ROS, Bax translocation, change in mitochondrial morphology, cytochrome c, caspases 3, 8 and 9; ↓ ΔΨm | [43] | ||
| A549, ASTC-a-1 | Artesunate | 10−50 μg/mL | ↑ ROS, Smac, AIF, caspase 3, Bak; ↓ ΔΨm | [44] | ||
| A549 | Artemisinin | 400 μM | ↑ ROS, caspases 3, 8 and 9, Bak, Smac, AIF | [45] | ||
| A549, ASTC-a-1 | DHA | 20 μg/mL | ↑ Bim, Bim translocation | [46] | ||
| SPC-A-1 | DHA | 30 μM | ↑ [Ca2+]i; ↓ survivin | [47] | ||
| H1975 | DHA | 15 μM | ↓ p-STAT3, Mcl-1, and inflammation; ↑ Bim | [48] | ||
| LLC | DHA | 20−80 μmol/L | ↑ chromatin condensation, shrunken nucleus; ↓ KDR/flk-1 | [49] | ||
| H460, H1299, Calu3, LXF289, A549, H1398 | Artesunate | 2.5 μM | ↓ AP-1 activity | [54] | ||
| A549, H1975 | DHA | 7.5−30 μM | ↓ Bcl2, XIAP | [55] | ||
| A549 | DHA | 0.71−11.36 mg/L (A549) 1.42−22.72 mg/L (A549/DDP) |
↓ HIP-1α, VEGF | [56] | ||
| A549, ASTC-a-1 | DHA | 20 μg/mL | ↑ Bax translocation, cytochrome c release, and caspases 3 and 9; ↓ ΔΨm | [60] | ||
| A549 | DHA | 6−12 μg/mL | caspase 3-dependent | [61] | ||
| A549 | DHA | 30−90 μM | ↑ ROS, DNA damage | [62] | ||
| A549 | DHA | 10−20 μg/mL | ↑ ROS, Bak, caspases 3, 8 and 9, tBid; ↓ ΔΨm | [63] | ||
| ASTC-a-1, 95D, H446 | DHA | 5−40 μg/mL | ↑ apoptosis | [63] | ||
| A549 | Artesunate | 25–100 μM | ↓ p-EGFR, EGFR, p-Akt, ABCG2 | [83] | ||
| SCLC | H69, H69VP | Artemisinin | 2−20 nM | ↑ DNA fragmentation | [64] | |
| Nasopharyngeal carcinoma | C666−1, HONE-1, HK1, HNE1, CNE2 | Artesunate | 10−40 μM | ↑ caspase 3, cleaved PARP, mitochondrial superoxide, ROS; ↓ oxygen consumption rate, ATP | [41] | |
| in vivo | Lung cancer | H1975; nude mice | DHA | 25 mg/kg | ↓ p-STAT3, Mcl-1; ↑ Bim | [48] |
| ABT-263 | 100 mg/kg/day; oral gavage | |||||
| A549 and A549/DDP; BALB/c athymic mice | DHA | 50, 100, 200 mg/kg/day; i.g. | ↑ apoptosis | [56] | ||
| Cisplatin | 2 mg/kg/3 days, 12 (A549) or 28 (A54/DDP) days; i.p. |
DHA induces apoptosis in A549 and PC-9 cells. The glycolytic metabolism was attenuated, together with the inhibition of glucose uptake, and lactate and ATP production. DHA treatment also reduced the levels of p-S6 ribosomal protein, p-mammalian target of rapamycin (mTOR), and glucose transporter (GLUT)1. These effects were enhanced together with the glycolysis inhibitor 2-Deoxy-d-glucose (2DG), inducing apoptosis through the activation of caspases 3, 8, and 9, cytochrome c and apoptosis-inducing factor (AIF), without raising ROS levels [21]. Other studies found that artemisinins and its derivatives induced ROS-mediated apoptosis. DHA induced ROS-mediated apoptosis in ASTC-a-1 cells through Bax translocation, mitochondrial membrane depolarization, morphological changes, cytochrome c release and activation of caspases 3, 8 and 9 [43]. Similar observations were made by Lu and colleagues, who recorded cell shrinkage, membrane frilling, blebbing, ovalization and blurring of the nuclear and cytoplasm boundary via a caspase 3-dependent mechanism caused by DHA in the same cell line [28]. Artesunate was found to promote ROS-mediated apoptosis in A549 and ASTC-a-1 cells in a time- and concentration-dependent fashion with a loss of mitochondrial potential and release of Smac and AIF [44]. In both A549 and ASTC-a-1 cells, treatment triggered apoptosis through an increase in ROS levels, with the activation of caspases 3, 8 and 9 without the cleavage of Bid [26,27,45]. Interestingly, Xiao and colleagues observed that blockade of caspases 8 or 9, but not caspase 3, largely inhibited the pro-apoptotic effects of artemisinin [26,27]. In contrast, Gao and colleagues found that silencing either caspase led to almost no activation of all three caspases, suggesting the role of an amplification loop among these caspases [45]. In the latter, there was no loss of mitochondrial membrane potential and cytochrome c release, but an activation of Smac and AIF release [45]. Artesunate similarly induced ROS-mediated apoptosis through the release of Smac and AIF, but this was accompanied by the loss of mitochondrial membrane potential. Here, inhibiting caspases 8 or 9 did not have any effect whilst silencing AIF did prevent artesunate-induced apoptosis [44].
Certain differences regarding the apoptotic effects involving Bak and Bax were uncovered. Xiao and colleagues found that silencing Bax and Bak by RNAi did not have any effect on artemisinin-induced apoptosis, suggesting a Bax/Bak-independent apoptotic process [26,27]. However, others observed that silencing Bak, but not Bax inhibited artesunate-induced apoptosis and AIF release. In fact, artesunate was found to only activate Bak, not Bax [44,45]. On the other hand, silencing pro-apoptotic Bax, but not Bak hampered DHA-induced apoptosis [39].
Interestingly, whilst artesunate treatment in ASTC-a-1 and A549 cells did not induce a significant downregulation of voltage-dependent anion channel 2 (VDAC2) expression and upregulation of Bim, silencing VDAC2 strongly promoted artesunate-induced Bak activation and apoptosis, which were prevented when Bim was silenced [44]. On the other hand, silencing Bim using shRNA in the same cells did not prevent DHA-induced caspase 9 activation and cell apoptosis [46].
Non-canonical apoptotic pathways were also found to be involved in the action of artemisinins. DHA-induced PC-14 lung cancer cell apoptosis was accompanied by an increase in intracellular calcium ion levels ([Ca2+]i) and activation of p38 [25]. DHA also downregulated the mRNA and protein levels of survivin in SPC-A-1 lung cancer cells to induce apoptosis but did not affect caspase 4 expression [47]. DHA attenuated STAT3 phosphorylation and activation, resulting in the downregulation of myeloid cell leukemia-1 (Mcl-1) and survivin levels in ABT-263 NSCLC cells possessing epidermal growth factor receptor (EGFR) or RAS mutations [48]. DHA induced apoptosis in the mouse LLC cell line by lowering the mRNA and protein levels of KDR/flk-1 [49]. A DHA-cinnamic acid ester derivative time- and dose-dependently induced intracellular ROS generation and apoptosis by exploiting the elevated intracellular levels of ferrous ion and endogenous oxidation stress in A549 cells [35]. Together, these studies show that artemisinins induce apoptosis but utilize very different pathways to induce apoptosis even within the same cell lines itself.
3.4. Inhibition of inflammation and oxidative stress
Several studies have indicated the anti-inflammatory effects of artemisinins in vitro with most studies mainly in vivo (Table 4 ). BEAS-2B cells were found to be insensitive to dexamethasone after being exposed to cigarette smoke extract (CSE) and TNF-α stimulation. Treatment with artesunate was able to reverse this effect and restore HDAC2 deactivation that was induced by CSE [50]. BEAS-2B cells exposed to CSE saw reductions in p-Akt and p-p44/42 protein expressions with artesunate treatment, coupled with heightened Nrf2 nuclear expression after 24 h, suggesting that artesunate could mitigate PI3K/Akt and p44/42 mitogen-activated protein kinases (MAPK) signaling pathways that are known to be activated in COPD [51]. Similarly, pre-treatment of A549 cells with artesunate saw a reduction in lipopolysaccharide (LPS)-induced IL-6 and IL-8 generation, with no effect on cell viability [52]. Artesunate mitigated hypoxia/reoxygenation-mediated increase in ROS levels in alveolar macrophages, together with reductions in NLR family pyrin domain containing 3 (NLRP3) and apoptosis-associated speck-like protein containing CARD (ASC) protein abundance, caspase 1 activity and production, and IL-1β and IL-18 mRNA and protein levels [53].
Table 4.
Effects of artemisinins and its derivatives on inflammation.
| Disease model | Cell line(s)/ stimulus/ allergen/ animal type | Derivative | Effective conc./ dose; route of administration | Outcomes | Ref. | |
|---|---|---|---|---|---|---|
| in vitro | Asthma | CSE/ OVA-induced airway inflammation in BEAS-2B | Artesunate | 10 μM | ↓ IL-8; ↑ HDAC2 activity | [50] |
| BEAS-2B | Artesunate | 10 μM | ↓ IL-6, IL-8, RANTES, p-Akt, p-tuberin, p-p70S6K, p-4EBP, MCP-1, and NF-κB transactivation | [74] | ||
| COPD | BEAS-2B | Artesunate | 30 μM | ↓ p-Akt, p-p44/42; ↑ Nrf2 | [51] | |
| ALI | LPS-induced ALI in A549 |
Artesunate | 1−4 μM | ↓ IL-6, IL-8 | [52] | |
| BEAS-2B | BDHA | 30 μM | ↑ Nrf2 | [67] | ||
| in vivo | Asthma | OVA; BALB/c mice | DHA | 50 mg/kg/day; i.p. | ↓ IL-17, IL-1b, TNFa, IL-6, STAT3, miR-183−5p, miR-96−5pm, miR-182−5p; ↑ IFN-γ, IL-10, Foxo1 | [76] |
| COPD (lung injury) | CSE exposure; BALB/c mice | Artesunate | 30 and 100 mg/kg; oral gavage | ↓ total and differential cell counts, IL-1β, MCP-1, IP-10, KC, TNF-α, MIP-2α, TGFβ, MMP9, TIMP1, iNOS, NOX2, 8-iso, 8-OHdG, 3-NT; ↑ catalase activity | [51] | |
| Allergic rhinitis | OVA; BALB/c mice | Artemisinin | 10−100 mg/kg/day; nasally | ↓ IgE, IL-4, IL-5, IL-10, TNF, IFNγ, IL-1β, ↑ Treg | [77] | |
| ALI | LPS; BALB/c mice | Artesunate | 10−40 mg/kg; i.v. | ↓ total cells, neu, mac, MPO, MDA, TNF-α, IL-1β, IL-6, TLR4, NF-κB; ↑ Nrf2, HO-1 | [52] | |
| RIR; Sprague Dawley rats | Artesunate | 15 mg/kg; i.p. | ↓ total cells, neu, mac, IL-1β, IL-18, MPO, MLRP3, ASC, caspase 1, ROS | [53] | ||
| Lung injury | Bleomycin; C57BL/6 mice | Artemisitene | 10 mg/kg; i.p. | ↑ Nrf2, NQO1, HO-1, IL-2, IFNy; ↓ hydroxyproline, total cells, neu, mac, lymph, IL-4, IL-6, TGFβ, MCP-1, sm-α actin | [66] | |
| Paraquat; Sprague Dawley rats | Artesunate | 100 mg/kg/day; i.p. | ↓ IL-10, TNF-α, TGFβ1 | [70] | ||
| Lung inflammation | RIR; Sprague Dawley rats | Artesunate | 15 mg/kg; i.p. | ↓ serum and pulmonary NO, MDA, IL-6, MIP-2, PGE2, arterial blood gas and biochemistry, lung wet/dry ratio, total cell number and [protein] in BALF, MPO, nuclear p65 NF-κB, p-IκB-α | [72] | |
| Sepsis lung injury | Cecal ligation and puncture; Kunming mice | Artesunate | 15 mg/kg; i.p. | ↓ TNF-α, IL-6, COX-2, iNOS, NF-κB; ↑ Nrf2, HO-1 | [73] |
3.5. Inhibition of angiogenesis, invasion, and metastasis
Angiogenesis, invasion, and metastasis are processes by which cancer cells spread. Artesunate impaired matrigel invasion of six NSCLC cell lines via the inhibition of urokinase-type plasminogen activator (u-PA), MMP2 and MMP7 promoter or enhancer activities, mRNA and protein expressions, with corresponding reductions in activator protein 1 (AP-1) and NF-κB [54]. Another group also observed that artesunate inhibited invasion of A549 cells using the transwell chamber invasion assay [30]. Similarly, a low concentration of DHA blocked in vitro migration and invasion of NSCLC cells through the downregulation of NF-κB levels that inhibited GLUT translocation and the Warburg effect [55]. DHA also enhanced the suppression of vascularisation-related proteins hypoxia-inducible factor 1-alpha (HIF-1α) and VEGF by cisplatin both in vitro and in vivo [56]. Artemisinin pre-treatment ablated IL-1β-induced p38 activation and VEGF-C mRNA and protein expression in LLC cells, factors associated with cancer and lymphangiogenesis. This effect was similar to that observed with a p38 MAPK inhibitor, suggesting the role of p38 as a pro-inflammatory cytokine-inducer of VEGF-C [57]. Artemisinin-daumone hybrid 15 (ARTD) was able to inhibit the invasion and metastasis of A549 cells, coupled with downregulation of E2F transcription factor 1 (E2F1) and hepatocyte nuclear factor 4 alpha (HNF4A), and upregulation of tumour-suppressive activating transcription factor 3 (ATF3) [58]. These show that artemisinins have the potential to impair angiogenesis and metastasis, but its effects were largely explored in the lung cancer setting. Thus, it would be interesting to see if similar effects are observed in other respiratory diseases that are implicated with angiogenic and metastatic events.
3.6. Chemosensitization of cancer cells to chemotherapeutic agents
Multiple studies have shown that artemisinin and its derivatives could chemosensitize other drugs. 10 μg/mL of DHA and 10 μg/mL of doxorubicin was found to be the most optimal concentrations that could reduce A549 cell viability [59]. DHA promoted the cytotoxic and apoptotic levels of carboplatin in LLC cells via the phosphorylation of p38 [36]. DHA together with ABT-263 could activate Bax-dependent apoptosis in NSCLC cells. This was because DHA induced downregulation of survivin and an upregulation of Bim, contributing to cotreatment-induced cytotoxicity. Also, DHA downregulated Mcl-1 expression which is responsible for drug resistance to ABT-263. This anti-tumour effect was also observed in vivo on H1975 xenograft growth in nude mice [48]. Similarly, DHA also upregulated Bax expression in the presence of gefitinib in H1975 cells, alongside an attenuation of p-Akt, p-mTOR, p-signal transducers and activators of transcription (STAT)3 and Bcl-2 to prevent migration and invasion [38]. Surprisingly, the JNK inhibitor SP600125 synergistically promoted DHA-induced cell apoptosis in A549 and ASTC-a-1 cells by activating Bax translocation, mitochondrial membrane depolarisation, cytochrome c release and caspase 3 and 9, unlike its usual anti-apoptotic function that suppresses c-Jun N-terminal kinase (JNK) and Bax [60]. DHA also interestingly promoted dictamine-induced apoptosis via a caspase 3-mediated pathway in A549 cells, even though dictamine alone induces S phase cell cycle arrest at low concentrations and cell apoptosis at higher concentrations without the involvement of caspases or mitochondria [61]. DHA and cisplatin ablated cell proliferation and induced apoptosis in both A549 and cisplatin-insensitive A549/DDP cells [56]. DHA could also reverse the high resistance of A549 cells to arsenic trioxide to reduce cell viability and promote cell death via higher levels of ROS and DNA damage, with no adverse effects on normal human bronchial epithelial cells [62]. On the contrary, apoptosis triggered by a combination of DHA and gemcitabine in A549 cells was not associated with additional generation of ROS as compared to either treatments alone. Instead, the combination strongly activated both the Bak-mediated intrinsic apoptosis pathway as well as the Fas-caspase 8-mediated extrinsic apoptosis pathway [63]. Moreover, DHA can enhance radiosensitization in GLC-82 lung cancer cells, inducing apoptosis with heightened expressions of p53 and p21, and lowered expression of Bcl-2 [37]. DHA coupled with a low dose of ionizing radiation led to irreparable G2/M phase cell cycle arrest as well as apoptosis due to ROS generation and the activation of caspases 3 and 8 [39]. In SCLC, pre-treatment with transferrin sensitized the multi-resistant H69VP phenotype to artemisinin as they had double the number of transferrin receptors. This combination induced DNA fragmentation and apoptosis [64]. Treating A549 cells with CQ prior to artesunate treatment synergistically promoted cell death, where an increase in the sub-G1 population of cells was observed, and the build-up of acidic vacuoles and ROS resulted in cytochrome c release followed by caspase 3-mediated apoptosis [33]. Conversely, artesunate did not induce A549 cell apoptosis when administered alone or in the presence of local radiotherapy. Instead, it induced G2/M phase cell cycle arrest, with heightened NO protein, and lessened cyclin B1 and cdc2 mRNA expression [40]. All these studies show that various therapies can be used in conjunction with artemisinins to promote its therapeutic effectiveness.
4. Pre-clinical/ in vivo studies of artemisinin and its derivatives
The effects of artemisinins in in vivo models are summarized in Table 2, Table 3, Table 4, Table 5 and these include disease models of pulmonary fibrosis, acute lung injury (ALI), asthma, COPD, lung cancer, and NPC. In general, the underlying mechanisms implicated in these models include inhibition of oxidative stress, inflammation, airway remodelling features, and tumour formation.
4.1. Inhibition of oxidative stress
The effect on oxidative stress has been studied in mouse and rat models of bleomycin-induced pulmonary fibrosis (IPF), ALI, asthma, and COPD.
DHA treatment increased lung tissue mRNA and protein levels of nuclear factor erythroid 2-related factor 2 (Nrf2) and heme oxygenase-1 (HO-1) as compared to rats stimulated with intratracheal administration of bleomycin over 14 days. This was with heightened levels of superoxide dismutase (SOD) and glutathione (GSH), and suppressed levels of malondialdehyde (MDA) [65]. Bleomycin-induced lung damage in mice was significantly inhibited by i.p. administration of 10 mg/kg artemisitene through its activation of Nrf2 and subsequently an increase in the mRNA expression of its downstream genes NAD(P)H quinone dehydrogenase 1 (NQO1) and HO-1 [66]. Artesunate treatment led to a further increment in LPS-induced increase in Nrf2 and HO-1 expressions in LPS-induced ALI [52]. Artesunate and biotinylated dihydroartesunate (BDHA) increased nuclear Nrf2 levels in BEAS-2B human bronchial epithelial cells via direct molecular interaction with Kelch-like ECH-associated protein 1 (Keap1) to free Nrf2 for transcriptional activity [67]. Similarly, artesunate promoted nuclear levels of Nrf2 in asthmatic mouse lung tissues sensitized and challenged with ovalbumin (OVA), and TNF-α stimulated BEAS-2B cells. Surprisingly, HO-1 expression was reduced by artesunate in this mouse model of allergic asthma. Artesunate also reduced the expressions of oxidative damage markers 8-isoprostane, 8-hydroxy-2-deoxyguanosine (8−OHdG) and 3-nitrotyrosine (3-NT), and the gene expression of regulatory subunits of NADPH oxidase (NOX)2, NOX4, p22phox, and p67phox. Expressions of SOD, inducible nitric oxide synthase (iNOS), and NOX1−4 were also reduced while that of catalase increased with artesunate treatment [68]. In cigarette smoke-induced lung oxidative damage in mice, artesunate also stimulated Nrf2 expression while reducing SOD activity, 3-NT and MDA [69]. In both allergic asthma and cigarette smoke-induced lung oxidative damage, glutathione peroxidase (GPx) levels were consistently unaffected by artesunate treatment [68,69]. In COPD animal models, artesunate dose-dependently ablated the expression of 8-isoprostane, 8OHdG and 3-NT in the bronchoalveolar lavage fluid (BALF), promoted catalase activity and reduced NOX2 protein expression in the mice lungs [51].
4.2. Inhibition of inflammation
Artemisitene reduced bleomycin-induced acute inflammatory responses through the activation of the Nrf2 pathway, as seen by a reduction in the total number of inflammatory cells, neutrophils, macrophages and lymphocytes, together with lower IL-4, IL-6, tumomur growth factor (TGF)β and monocyte chemoattractant protein-1 (MCP-1) mRNA expressions [66]. Artesunate treatment attenuated lung injury in paraquat-intoxicated rats via reductions in TGFβ1, IL-10 and TNF-α [70]. 30 mg/kg of artesunate suppressed total, eosinophil and neutrophil inflammatory cell counts as well in an OVA-induced model of allergic asthma [68]. It also reduced IL-8 levels and total inflammatory and neutrophil cell counts that were increased in a 40 days cigarette smoke-induced lung oxidative damage mouse model. IL-8 levels were similarly lowered by artesunate in 16HBE cells exposed to cigarette smoke extract [69]. Artesunate pre-treatment inhibited ALI that was induced by either LPS or NLRP3 activation by renal ischemia-reperfusion (RIR) [52,53], with both observing a reduction in total inflammatory, macrophage and neutrophil cell counts and IL-1β levels. In addition, artesunate reduced IL-18 levels in RIR-mediated ALI [53], and IL-6, TNF-α [71], NF-κB and TLR4 levels in LPS-induced ALI [52]. Interestingly, the activation of NLRP3 inflammasome was dependent on pulmonary ROS generation accompanied by higher ASC and caspase 1 levels [53]. In another study, Liu and colleagues also found that artesunate suppressed many RIR-stimulated factors involved in lung inflammation, including the production of serum and pulmonary NO, MDA, macrophage inflammatory protein 2 (MIP-2) and prostaglandin E2 (PGE2), and attenuated NF-κB translocation [72]. Artesunate also protected against sepsis-induced lung injury by reducing IL-6 and TNF-α levels in both the serum and BALF. In the lung tissues, artesunate suppressed cyclooxygenase-2 (COX-2), iNOS and NF-κB levels and activated Nrf2 through and increase in HO-1 expression and enzymatic activity [73]. The effect of artesunate on COPD was similar to that of ALI where artesunate dose-dependently suppressed total and differential inflammatory cell counts and IL-1β levels, together with a drop in keratinocyte chemoattractant (KC), IFNγ-induced protein 10 (IP-10) and MCP-1 [51]. In asthma, artesunate dose-dependently suppressed airway inflammation as observed in acute mouse models of allergic asthma. 30 mg/kg of artesunate attenuated total inflammatory and eosinophil counts when house dust mites were used as the allergen, while total inflammatory and eosinophil cell counts, IL-4, IL-5, IL-13 and eotaxin in BALF were found to be attenuated when OVA was used [74]. Luo and colleagues also noted similar reductions in total and differential cell counts, IL-4, IL-8, IL-13 and TNF-α, accompanied by reductions in p110δPI3K and p-Akt1, suggesting the involvement of PI3K/Akt pathway [50]. Likewise, the same dose of DHA, although administered intragastrically, also attenuated airway inflammation by reducing the number of infiltrating inflammatory cells, IL-4, IL-5, IL-13 and IgE levels. This was accompanied by reductions in p-ERK, p-p38, (inhibitor of NF-κB alpha) IκBα and p-NF-κB p65 protein abundance [75]. DHA and artesunate were also found to ablate the mRNA levels of molecules involved in promoting airway inflammation and remodelling, including chitinase 3-like protein 4 (Ym2) [75], chitinase-like glycoprotein (YKL-40), iNOS, thymic stromal lymphopoietin (TSLP), intercellular adhesion molecule 1 (ICAM-1), vascular cell adhesion molecule 1 (VCAM-1) and E-selectin [74]. Zhu and colleagues found that DHA significantly relieved airways hyperresponsiveness (AHR) and mucus secretion in OVA-stimulated mice by reducing the percentage of T helper 17 (Th17) cells. This was through the abolishment of IL-6 and STAT3 expression, which was due to a reduction in the transcriptional levels of miR-183−5p, miR-96−5p and miR-182−5p, and an increase in transcriptional and translational expression of forkhead box O1 (Foxo1) [76]. Lastly, artemisinin with or without neurectomy of pterygoid canal reduced the levels of histamine, IgE, TNF, IFNγ, IL-1β, IL-4, IL-5 and IL-10, and raised the proportion of regulatory T (Treg) cells in OVA-induced allergic rhinitis in mice [77].
4.3. Effect on respiratory/ lung function
Whether or not changes in inflammation brought about by artesunate was associated with changes in lung function parameters have been reported by one group. Here, artesunate treatment in mice exposed to cigarette smoke and OVA saw a reduction in methacholine-induced AHR, with efficacies similar to the extent produced by dexamethasone [50]. Both artesunate and DHA treatment in an OVA-induced model of allergic asthma brought about a reduction in AHR [74,75]. Another study found that 120 μg of artesunate relieved OVA-induced airway resistance with comparable efficacy to 3 μg of salbutamol through an increase in [Ca2+]i and reduced traction force in airway smooth muscle cells, mediated by bitter taste receptor signaling [78]. However, the concentration and dose used in this study are high; suggesting the need to explore whether the same effects could be observed at the lower therapeutic range. Artemisinin was also able to improve the behavior scores (sneezing, nasal rubbing) in a mouse model of allergic rhinitis, where mice were given nasal drip of 500 μg of OVA [77].
4.4. Inhibition of airway remodeling features
4.4.1. Fibrosis
Artemisitene inhibited bleomycin-induced collagen and hydroxyproline expression in mice. The expression of key players of fibrosis, smooth muscle (sm)-α actin and TGFβ were also reduced in bleomycin-treated mice [66]. Similar observations were made with DHA, which reduced the Szapiel fibrotic score and hydroxyproline content with comparable efficacy to dexamethasone in bleomycin-induced pulmonary fibrosis in rats [79]. Another study also showed that DHA treatment reduced interstitial fibrosis, leukocyte infiltration, collagen deposition and sm-α actin expression in lung tissues with heightened E-cadherin expression. The reduction in sm-α actin, normally heightened in the event of oxidative stress, was also seen in DHA-treated rat alveolar epithelial cells (AECs) cultured in hypoxia, which shows that DHA could inhibit the hypoxia-induced increase in myofibroblastic-like process [65]. Artesunate attenuated bleomycin-induced pulmonary fibrosis in Sprague Dawley rats through a reduction in pro-fibrotic proteins such as TGFβ1, Smad3, heat shock protein 47 (hsp47), sm-α actin, and collagen I [80]. The same group also observed that artesunate upregulated MMP2 and MMP9 expressions while reducing tissue inhibitor of metalloproteinases (TIMP) and TIMP2 levels, which then contributed to a decrease in collagen IV protein expression, which is otherwise heightened in bleomycin-induced pulmonary fibrosis [81].
4.4.2. Angiogenesis and metastasis
Artemisinin, artesunate, and DHA inhibited processes that contribute to tumour malignancy, including migration, invasion, cancer stem cells and epithelial-mesenchymal transition (EMT) transition. This was through attenuation of the Wnt/β-catenin pathway that contributes to tumour cell proliferation and malignancy, as seen by a reduction in Wnt5-a/b protein level and a simultaneous increase in naked cuticle homolog 2 (NKD2) and axis inhibition protein 2 (Axin2) that eventually led to a drop in β-catenin levels [22]. Artesunate post-treatment also reportedly prevented TGFβ1-induced EMT in RLE-6TN alveolar epithelial cells by reducing p-Smad3 and Smad3 and upregulating Smad7 protein expressions [82]. Artesunate impaired tumour growth and metastasis in a chicken embryo metastasis model, together with corresponding reductions in MMP2, MMP7, and u-PA mRNA expression [54]. Artesunate also inhibited RIR-mediated lung damage, vascular permeability and edema in rats [53]. ARTD blocked cancer-associated bone metastasis more potently than artemisinin when mice were inoculated with lung cancer A549 cells. It induced the expression of tumour-suppressive ATF3 and reduced the mRNA and protein levels of oncogenic E2F1, receptor activator of nuclear factor kappa-B ligand (RANKL), and secreted levels of MMP9 and cathepsin K that contribute to the bone-resorbing activity [58]. In addition, oral administration of artemisinin inhibited lymph node and lung metastasis, with no effect on tumour growth in a LLC mouse model, promoting longer survival. Tumour lymphangiogenesis was also inhibited, with corresponding reduction in VEGF-C levels [57]. Interestingly, studies done by two different groups found that a combination of DHA with either cisplatin or onconase could more effectively ablate the density of the microvasculature and microvessels in an A549 mouse xenograft model [29,56].
4.4.3. Mucus production
Studies by Wong and colleagues found that artesunate mitigated mucus hypersecretion via a reduction in muc5ac mRNA expression in the lung tissues of OVA-challenged asthmatic mice [74,75]. Whether or not similar effects on mucus production and muc5ac expression can be observed using a more clinically relevant allergen such as house dust mite remains to be observed. Unfortunately, not much research has looked at the effect of artemisinins on mucus production and alleviating it would be beneficial since excessive mucus production occurs in many lung diseases and impede on patients’ comfort levels.
4.5. Tumour proliferation
Tong and colleagues observed that artemisinin, DHA and artesunate were all able to reduce tumour growth in an A549-induced mouse xenograft model via inhibition of the Wnt-5a/b/β-catenin signaling pathway [22]. Artesunate dose-dependently attenuated A549 xenograft growth in mice with a reduction in EGFR, Akt and ATP-binding cassette subfamily member 2 (ABCG2) mRNA and protein expressions [83]. In addition, artesunate radiosensitized tumour cells to the effects of local radiotherapy [40]. Conversely, another group found that 10 mg/kg of artesunate was not sufficient to inhibit A549-induced xenograft growth in mice, although it could potentially block invasion as observed by a reduction in ICAM-1 and MMP9 protein abundance [30]. Unexpectedly, oral administration of either DLAe or artesunate was able to inhibit A549 xenograft growth but only DLAe was able to inhibit PC-9 induced tumour growth [34]. The tumour-inhibiting rate of DHA as studied in nude mice bearing A549 cells was 54.3 % [24]. A combination of DHA and ABT-263 reduced xenograft growth in nude mice [48]. Using an acute model of allergic asthma in mice, artesunate pre-treatment was found to reduce the area of sm-α actin positive cells in the airways and cyclin D1 protein expression [31].
5. Artesunate (Clinical studies)
Currently, only one study has investigated the use of artesunate for lung cancer in humans. Adding on 120 mg/day of artesunate treatment to vinorelbine and cisplatin chemotherapy was found to promote better disease control and slow the time to disease progression as compared to advanced stage NSCLC patients treated with vinorelbine and cisplatin chemotherapy alone. However, no significant differences to short term survival rate, mean survival time and one-year survival rates were observed. Importantly, this treatment combination did not produce significant toxic effects [84].
6. Artemisinins for the treatment of COVID-19
The recent Coronavirus Disease 2019 (COVID-19) pandemic has affected and taken many lives [85]. Since vaccines against the novel SARS-CoV-2 virus, which causes COVID-19, may take a long time to be developed, many are repurposing drugs for its treatment. CQ and hydroxychloroquine (HCQ) are anti-malarial drugs being tested for COVID-19 [86,87] that have also been used against autoimmune diseases such as rheumatoid arthritis and systemic lupus erythematosus (SLE). Whilst HCQ has displayed a safer toxicity profile than CQ [86], there are still side effects that are of concern. One example is cardiac toxicity, which would be especially dangerous for patients with pre-existing health conditions, like that of cardiovascular diseases, as they would have poorer prognosis for COVID-19 [88]. Whilst the anti-malarial mode of action of artemisinins are different from CQ or HCQ, their immunomodulant effects against inflammatory disorders and viral replications are overlapping. Traditionally, artemisinins have been used for the treatment of fevers, and could be useful given that 83.3 % of patients with COVID-19 have fever [89]. Given its ability to reduce TNF-α and IL-6, key mediators of acute respiratory distress syndrome (ARDS) that leads to the worsening of COVID-19 patient conditions [90], artemisinins may be a promising therapy. Other molecular targets of artemisinin and its derivatives, as shown in Fig. 1 , may also be involved in the pathogenesis of COVID-19 and thus may have other benefits that may not yet be known. Moreover, artemisinins are known to display a safe toxicity profile so higher doses can be prescribed with less worry about potential side effects. With the understanding that CQ and HCQ are affective against viruses due to the pH altering activities that affect viral replication, artemisinins could alternatively be used as adjunct therapy to lower the dose required of CQ or HCQ, and reduce side effects, while also suppressing the cytokine storm. Unfortunately, no study to date has investigated the effects or interactions of artemisinins on the angiotensin-converting enzyme 2 (ACE2) receptor, that is known to be the critical binding cellular receptor of SARS-CoV-2 [91]. This can greatly influence the favourability of trying out the effectiveness of artemisinins for COVID-19.
Fig. 1.

Molecular targets modulated by artemisinins in respiratory diseases.
7. Modifications to artemisinins for drug delivery
Artemisinins have been used for a long time with high efficacies and relatively safe toxicity profiles. Some groups have looked into modifications to artemisinins in order to improve its efficacy and lower the risk of toxic side effects. DHA was observed to display poor water solubility and short plasma half-life. Dai and colleagues connected DHA with a multiarm polyethylene glycol (PEG) to produce PEG-DHA and found that it was 82–163 times more water-soluble and its blood circulation half-time was 5.75–16.75 times that of DHA, all while retaining or improving its anti-cancer efficacy [92]. Sun and colleagues encapsulated DHA with gelatin or hyaluronan nanoparticles using an electrostatic field system and observed that it inhibited proliferation and promoted apoptosis of A549 cells better than DHA [93]. DHA loaded with nanostructured lipid carriers (DHA-NLC) resided more greatly in organs such as the lung, liver, spleen, brain, and muscle, and less in the heart and kidneys, promoting sustained-release and better drug-targeted effects, therefore allowing for lower dosages and systemic toxic side effects [94]. A C-10 acetal artemisinin synthesized using the Sonogashira cross-coupling reaction displayed higher growth inhibition of A549 cells compared to artemisinin. However, it only had moderate effects on other cancer cell lines such as breast, prostate, and neuroblastoma [95]. Lastly, Yang and colleagues noted that transferrin receptors were overexpressed in cancer cells. They, therefore, developed adducts of transferrin with artemisinin, DHA or artesunate and found that their anti-cancer effects were stronger in A549 cells with improved cellular uptake, whilst having minimal effects on normal human liver HL-7702 cells [96].
8. Conclusion and future perspectives
We present here an up-to-date overview of the current knowledge of artemisinins and its derivatives as potential therapeutic targets for the treatment of respiratory diseases. Fig. 1 summaries the plethora of signaling pathways that are regulated by artemisinin and its derivatives in the treatment of different respiratory diseases to date. These include inhibition of the cell cycle, cell proliferation, inflammation, apoptosis, angiogenesis, invasion and metastasis, and promotion of oxidative stress. In vivo studies additionally examined the effects of artemisinins in structural changes and functional assays. To date, a large proportion of studies have been conducted in lung cancer, with not much done in other respiratory diseases. Yet, many of the physiological processes that occur in lung cancer are observed in other respiratory diseases. This includes, but is not limited to, cell proliferation and inflammatory events. Therefore, there is potential for artemisinins to be used to mitigate other respiratory diseases and that needs to be explored. Fig. 2 summarizes the different ranges of concentrations of artemisinins used in the in vitro studies of different processes involved in the respiratory diseases explored. It is interesting to observe that larger and higher ranges of concentrations were used in cell proliferation and apoptosis studies, which could possibly be attributed to the higher concentrations needed to produce a cytotoxic effect in cancer cells. A moderate range of artemisinin concentrations was applied in studies looking at inflammation, angiogenesis, invasion and metastasis, whilst lowest concentrations were used in airway wall remodelling (AWR) and lung function studies. This could indicate that different concentrations, and even drug formulation, could be pegged according to the disease type and process to reduce the potential side effects. It is also interesting to note that a large proportion of studies have been conducted in vitro, especially for studying the mechanisms affecting cell/tumour proliferation and apoptosis. More of these could be studied in vivo as the dosages of artemisinins are known and well-tolerated in animals. Nonetheless, given that artemisinins are already approved for use in clinics and many clinical studies conducted in malaria have shown that the drug is well-tolerated, artemisinins would make a strong drug candidate that can be repurposed as therapy for a wide spectrum of respiratory diseases, and could potentially be an option for the urgent treatment for the COVID-19 pandemic.
Fig. 2.

Concentration of artemisinins used in the in vitro study of respiratory diseases.
Funding
Thai Tran is supported by the Singapore Ministry of Education’s Academic Research Fund Tier 1 (T1−2014 Apr-03). WS Fred Wong is partly supported by the Singapore National Research Foundation through the grant NRF/CREATE/R-184-000-269-592.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- 1.Hansen-Flaschen J., Bates D.V. Respiratory disease. 2019;16 https://www.britannica.com/science/respiratory-disease January 2019 February 03, 2019]; Available from: [Google Scholar]
- 2.2013. Forum of International Respiratory Societies, Respiratory Diseases in the World. [Google Scholar]
- 3.World Health Organization . 2007. Global Surveillance, Prevention and Control of Chronic Respiratory Diseases. [Google Scholar]
- 4.World Health Organization . 2019. COPD Management.https://www.who.int/respiratory/copd/management/en/ Available from: [Google Scholar]
- 5.GINA . 2018. Pocket Guide for Asthma Management and Prevention. [Google Scholar]
- 6.World Health Organization . 2003. Influenza.https://www.who.int/mediacentre/factsheets/2003/fs211/en/ Available from: [Google Scholar]
- 7.Ferlay J. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer. 2010;127(12):2893–2917. doi: 10.1002/ijc.25516. [DOI] [PubMed] [Google Scholar]
- 8.Cragg G.M., Newman D.J. Biodiversity: a continuing source of novel drug leads. Pure Appl. Chem. 2005;77(1):7–24. [Google Scholar]
- 9.Hicks S. 2nd ed. Nayler.; San Antonio, Texas: 1966. Desert Plants and People. [Google Scholar]
- 10.Dias D.A., Urban S., Roessner U. A historical overview of natural products in drug discovery. Metabolites. 2012;2(2):303–336. doi: 10.3390/metabo2020303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Timmer A. Pelargonium sidoides extract for treating acute respiratory tract infections. Cochrane Database Syst. Rev. 2013;(10):Cd006323. doi: 10.1002/14651858.CD006323.pub3. [DOI] [PubMed] [Google Scholar]
- 12.Tu Y. The development of the antimalarial drugs with new type of chemical structure--qinghaosu and dihydroqinghaosu. Southeast Asian J. Trop. Med. Public Health. 2004;35(2):250–251. [PubMed] [Google Scholar]
- 13.Cheng C. Anti-malarial drug artesunate attenuates experimental allergic asthma via inhibition of the phosphoinositide 3-Kinase/Akt pathway. PLoS One. 2011;6(6) doi: 10.1371/journal.pone.0020932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Meshnick S.R., Taylor T.E., Kamchonwongpaisan S. Artemisinin and the antimalarial endoperoxides: from herbal remedy to targeted chemotherapy. Microbiol. Rev. 1996;60(2):301–315. doi: 10.1128/mr.60.2.301-315.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Karunajeewa H. 2012. Artemisinins: Artemisinin, Dihydroartemisinin, Artemether and Artesunate, in Milestones in Drug Therapy; pp. 157–190. [Google Scholar]
- 16.Efferth T. Molecular modes of action of artesunate in tumor cell lines. Mol. Pharmacol. 2003;64(2):382–394. doi: 10.1124/mol.64.2.382. [DOI] [PubMed] [Google Scholar]
- 17.Efferth T. The anti-malarial artesunate is also active against cancer. Int. J. Oncol. 2001;18(4):767–773. doi: 10.3892/ijo.18.4.767. [DOI] [PubMed] [Google Scholar]
- 18.Xu H. Anti-malarial agent artesunate inhibits TNF-alpha-induced production of proinflammatory cytokines via inhibition of NF-kappaB and PI3 kinase/Akt signal pathway in human rheumatoid arthritis fibroblast-like synoviocytes. Rheumatology (Oxford) 2007;46(6):920–926. doi: 10.1093/rheumatology/kem014. [DOI] [PubMed] [Google Scholar]
- 19.Efferth T. The antiviral activities of artemisinin and artesunate. Clin. Infect. Dis. 2008;47(6):804–811. doi: 10.1086/591195. [DOI] [PubMed] [Google Scholar]
- 20.Li X. The selectivity of artemisinin-based drugs on human lung normal and cancer cells. Environ. Toxicol. Pharmacol. 2018;57:86–94. doi: 10.1016/j.etap.2017.12.004. [DOI] [PubMed] [Google Scholar]
- 21.Mi Y.J. Dihydroartemisinin inhibits glucose uptake and cooperates with glycolysis inhibitor to induce apoptosis in non-small cell lung carcinoma cells. PLoS One. 2015;10(3):e0120426. doi: 10.1371/journal.pone.0120426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Tong Y. Artemisinin and its derivatives can significantly inhibit lung tumorigenesis and tumor metastasis through Wnt/beta-catenin signaling. Oncotarget. 2016;7(21):31413–31428. doi: 10.18632/oncotarget.8920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Liao K., Li J., Wang Z. Dihydroartemisinin inhibits cell proliferation via AKT/GSK3beta/cyclinD1 pathway and induces apoptosis in A549 lung cancer cells. Int. J. Clin. Exp. Pathol. 2014;7(12):8684–8691. [PMC free article] [PubMed] [Google Scholar]
- 24.Chen W. [Effect of dihydroartemisinin on proliferation of human lung adenocarcinoma cell line A549] Zhongguo Fei Ai Za Zhi. 2005;8(2):85–88. doi: 10.3779/j.issn.1009-3419.2005.02.02. [DOI] [PubMed] [Google Scholar]
- 25.Mu D. The role of calcium, P38 MAPK in dihydroartemisinin-induced apoptosis of lung cancer PC-14 cells. Cancer Chemother. Pharmacol. 2008;61(4):639–645. doi: 10.1007/s00280-007-0517-5. [DOI] [PubMed] [Google Scholar]
- 26.Xiao F. Amplification activation loop between caspase-8 and -9 dominates artemisinin-induced apoptosis of ASTC-a-1 cells. Apoptosis. 2012;17(6):600–611. doi: 10.1007/s10495-012-0706-5. [DOI] [PubMed] [Google Scholar]
- 27.Xiao F.L. Artemisinin induces caspase-8/9-mediated and Bax/Bak-independent apoptosis in human lung adenocarcinoma (ASTC-a-1) cells. J. Xray Sci. Technol. 2011;19(4):545–555. doi: 10.3233/XST-2011-0313. [DOI] [PubMed] [Google Scholar]
- 28.Lu Y.Y. Dihydroartemisinin (DHA) induces caspase-3-dependent apoptosis in human lung adenocarcinoma ASTC-a-1 cells. J. Biomed. Sci. 2009;16:16. doi: 10.1186/1423-0127-16-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Shen R. Combination of onconase and dihydroartemisinin synergistically suppresses growth and angiogenesis of non-small-cell lung carcinoma and malignant mesothelioma. Acta Biochim Biophys Sin (Shanghai) 2016;48(10):894–901. doi: 10.1093/abbs/gmw082. [DOI] [PubMed] [Google Scholar]
- 30.Chen X. [Effects of artesunate on the invasion of lung adenocarcinoma A549 cells and expression of ICAM-1 and MMP-9] Zhongguo Fei Ai Za Zhi. 2013;16(11):567–571. doi: 10.3779/j.issn.1009-3419.2013.11.01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Tan S.S. The antimalarial drug artesunate inhibits primary human cultured airway smooth muscle cell proliferation. Am. J. Respir. Cell Mol. Biol. 2014;50(2):451–458. doi: 10.1165/rcmb.2013-0273OC. [DOI] [PubMed] [Google Scholar]
- 32.Zeng A.H. Human embryonic lung fibroblasts treated with artesunate exhibit reduced rates of proliferation and human cytomegalovirus infection in vitro. J. Thorac. Dis. 2015;7(7):1151–1157. doi: 10.3978/j.issn.2072-1439.2015.07.05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ganguli A. Inhibition of autophagy by chloroquine potentiates synergistically anti-cancer property of artemisinin by promoting ROS dependent apoptosis. Biochimie. 2014;107:338–349. doi: 10.1016/j.biochi.2014.10.001. Pt B. [DOI] [PubMed] [Google Scholar]
- 34.Rassias D.J., Weathers P.J. Dried leaf Artemisia annua efficacy against non-small cell lung cancer. Phytomedicine. 2019;52:247–253. doi: 10.1016/j.phymed.2018.09.167. [DOI] [PubMed] [Google Scholar]
- 35.Xu C.C. Synthesis and in vitro antitumor evaluation of dihydroartemisinin-cinnamic acid ester derivatives. Eur. J. Med. Chem. 2016;107:192–203. doi: 10.1016/j.ejmech.2015.11.003. [DOI] [PubMed] [Google Scholar]
- 36.Zhang B. Dihydroartemisinin sensitizes Lewis lung carcinoma cells to carboplatin therapy via p38 mitogen-activated protein kinase activation. Oncol. Lett. 2018;15(5):7531–7536. doi: 10.3892/ol.2018.8276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zuo Z.J. [Effect of dihydroartemisinin combined irradiation on the apoptosis of human lung cancer GLC-82 cells and its mechanism study] Zhongguo Zhong Xi Yi Jie He Za Zhi. 2014;34(10):1220–1224. [PubMed] [Google Scholar]
- 38.Jin H. Dihydroartemisinin and gefitinib synergistically inhibit NSCLC cell growth and promote apoptosis via the Akt/mTOR/STAT3 pathway. Mol. Med. Rep. 2017;16(3):3475–3481. doi: 10.3892/mmr.2017.6989. [DOI] [PubMed] [Google Scholar]
- 39.Chen T., Chen M., Chen J. Ionizing radiation potentiates dihydroartemisinin-induced apoptosis of A549 cells via a caspase-8-dependent pathway. PLoS One. 2013;8(3):e59827. doi: 10.1371/journal.pone.0059827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhao Y. Artesunate enhances radiosensitivity of human non-small cell lung cancer A549 cells via increasing NO production to induce cell cycle arrest at G2/M phase. Int. Immunopharmacol. 2011;11(12):2039–2046. doi: 10.1016/j.intimp.2011.08.017. [DOI] [PubMed] [Google Scholar]
- 41.Li Q. Targeting nasopharyngeal carcinoma by artesunate through inhibiting Akt/mTOR and inducing oxidative stress. Fundam. Clin. Pharmacol. 2017;31(3):301–310. doi: 10.1111/fcp.12266. [DOI] [PubMed] [Google Scholar]
- 42.Liu W.M., Gravett A.M., Dalgleish A.G. The antimalarial agent artesunate possesses anticancer properties that can be enhanced by combination strategies. Int. J. Cancer. 2011;128(6):1471–1480. doi: 10.1002/ijc.25707. [DOI] [PubMed] [Google Scholar]
- 43.Lu Y.Y. Single-cell analysis of dihydroartemisinin-induced apoptosis through reactive oxygen species-mediated caspase-8 activation and mitochondrial pathway in ASTC-a-1 cells using fluorescence imaging techniques. J. Biomed. Opt. 2010;15(4):46028. doi: 10.1117/1.3481141. [DOI] [PubMed] [Google Scholar]
- 44.Zhou C. Artesunate induces apoptosis via a Bak-mediated caspase-independent intrinsic pathway in human lung adenocarcinoma cells. J. Cell. Physiol. 2012;227(12):3778–3786. doi: 10.1002/jcp.24086. [DOI] [PubMed] [Google Scholar]
- 45.Gao W. Artemisinin induces A549 cell apoptosis dominantly via a reactive oxygen species-mediated amplification activation loop among caspase-9, -8 and -3. Apoptosis. 2013;18(10):1201–1213. doi: 10.1007/s10495-013-0857-z. [DOI] [PubMed] [Google Scholar]
- 46.Chen M. Dihydroarteminsin-induced apoptosis is not dependent on the translocation of bim to the endoplasmic reticulum in human lung adenocarcinoma cells. Pathol. Oncol. Res. 2012;18(4):809–816. doi: 10.1007/s12253-012-9508-x. [DOI] [PubMed] [Google Scholar]
- 47.Mu D. Calcium and survivin are involved in the induction of apoptosis by dihydroartemisinin in human lung cancer SPC-A-1 cells. Methods Find. Exp. Clin. Pharmacol. 2007;29(1):33–38. doi: 10.1358/mf.2007.29.1.1063493. [DOI] [PubMed] [Google Scholar]
- 48.Yan X. Dihydroartemisinin suppresses STAT3 signaling and Mcl-1 and Survivin expression to potentiate ABT-263-induced apoptosis in Non-small Cell Lung Cancer cells harboring EGFR or RAS mutation. Biochem. Pharmacol. 2018;150:72–85. doi: 10.1016/j.bcp.2018.01.031. [DOI] [PubMed] [Google Scholar]
- 49.Zhou H.J. Dihydroartemisinin improves the efficiency of chemotherapeutics in lung carcinomas in vivo and inhibits murine Lewis lung carcinoma cell line growth in vitro. Cancer Chemother. Pharmacol. 2010;66(1):21–29. doi: 10.1007/s00280-009-1129-z. [DOI] [PubMed] [Google Scholar]
- 50.Luo Q. The anti-malaria drug artesunate inhibits cigarette smoke and ovalbumin concurrent exposure-induced airway inflammation and might reverse glucocorticoid insensitivity. Int. Immunopharmacol. 2015;29(2):235–245. doi: 10.1016/j.intimp.2015.11.016. [DOI] [PubMed] [Google Scholar]
- 51.Ng D.S. Anti-malarial drug artesunate protects against cigarette smoke-induced lung injury in mice. Phytomedicine. 2014;21(12):1638–1644. doi: 10.1016/j.phymed.2014.07.018. [DOI] [PubMed] [Google Scholar]
- 52.Zhao D. Artesunate protects LPS-Induced acute lung injury by inhibiting TLR4 expression and inducing Nrf2 activation. Inflammation. 2017;40(3):798–805. doi: 10.1007/s10753-017-0524-6. [DOI] [PubMed] [Google Scholar]
- 53.Liu Z. Artesunate Inhibits Renal Ischemia-Reperfusion-Mediated Remote Lung Inflammation Through Attenuating ROS-Induced Activation of NLRP3 Inflammasome. Inflammation. 2018;41(4):1546–1556. doi: 10.1007/s10753-018-0801-z. [DOI] [PubMed] [Google Scholar]
- 54.Rasheed S.A.K. First evidence that the antimalarial drug artesunate inhibits invasion and in vivo metastasis in lung cancer by targeting essential extracellular proteases. Int. J. Cancer. 2010;127(6):1475–1485. doi: 10.1002/ijc.25315. [DOI] [PubMed] [Google Scholar]
- 55.Jiang J. Repurposing the anti-malarial drug dihydroartemisinin suppresses metastasis of non-small-cell lung cancer via inhibiting NF-kappaB/GLUT1 axis. Oncotarget. 2016;7(52):87271–87283. doi: 10.18632/oncotarget.13536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Zhang J.L. DHA regulates angiogenesis and improves the efficiency of CDDP for the treatment of lung carcinoma. Microvasc. Res. 2013;87:14–24. doi: 10.1016/j.mvr.2013.02.006. [DOI] [PubMed] [Google Scholar]
- 57.Wang J. Artemisinin inhibits tumor lymphangiogenesis by suppression of vascular endothelial growth factor C. Pharmacology. 2008;82(2):148–155. doi: 10.1159/000148261. [DOI] [PubMed] [Google Scholar]
- 58.Ma G.T. Artemisinin-daumone hybrid inhibits Cancer cell-Mediated osteolysis by targeting Cancer cells and osteoclasts. Cell. Physiol. Biochem. 2018;49(4):1460–1475. doi: 10.1159/000493449. [DOI] [PubMed] [Google Scholar]
- 59.Tai X. In vitro and in vivo inhibition of tumor cell viability by combined dihydroartemisinin and doxorubicin treatment, and the underlying mechanism. Oncol. Lett. 2016;12(5):3701–3706. doi: 10.3892/ol.2016.5187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Lu Y.-Y. The JNK inhibitor SP600125 enhances dihydroartemisinin-induced apoptosis by accelerating Bax translocation into mitochondria in human lung adenocarcinoma cells. FEBS Lett. 2010;584(18):4019–4026. doi: 10.1016/j.febslet.2010.08.014. [DOI] [PubMed] [Google Scholar]
- 61.An F.F. Dihydroartemisinine enhances dictamnine-induced apoptosis via a caspase dependent pathway in human lung adenocarcinoma A549 cells. Asian Pac. J. Cancer Prev. 2013;14(10):5895–5900. doi: 10.7314/apjcp.2013.14.10.5895. [DOI] [PubMed] [Google Scholar]
- 62.Chen H. Dihydroartemisinin sensitizes human lung adenocarcinoma A549 cells to arsenic trioxide via apoptosis. Biol. Trace Elem. Res. 2017;179(2):203–212. doi: 10.1007/s12011-017-0975-5. [DOI] [PubMed] [Google Scholar]
- 63.Zhao C., Gao W., Chen T. Synergistic induction of apoptosis in A549 cells by dihydroartemisinin and gemcitabine. Apoptosis. 2014;19(4):668–681. doi: 10.1007/s10495-013-0953-0. [DOI] [PubMed] [Google Scholar]
- 64.Sadava D. Transferrin overcomes drug resistance to artemisinin in human small-cell lung carcinoma cells. Cancer Lett. 2002;179(2):151–156. doi: 10.1016/s0304-3835(02)00005-8. [DOI] [PubMed] [Google Scholar]
- 65.Yang D.X. Dihydroartemisinin alleviates oxidative stress in bleomycin-induced pulmonary fibrosis. Life Sci. 2018;205:176–183. doi: 10.1016/j.lfs.2018.05.022. [DOI] [PubMed] [Google Scholar]
- 66.Chen W. Artemisitene activates the Nrf2-dependent antioxidant response and protects against bleomycin-induced lung injury. FASEB J. 2016;30(7):2500–2510. doi: 10.1096/fj.201500109R. [DOI] [PubMed] [Google Scholar]
- 67.Ravindra K.C. Untargeted proteomics and systems-based mechanistic investigation of artesunate in human bronchial epithelial cells. Chem. Res. Toxicol. 2015;28(10):1903–1913. doi: 10.1021/acs.chemrestox.5b00105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ho W.E. Anti-malarial drug artesunate ameliorates oxidative lung damage in experimental allergic asthma. Free Radic. Biol. Med. 2012;53(3):498–507. doi: 10.1016/j.freeradbiomed.2012.05.021. [DOI] [PubMed] [Google Scholar]
- 69.Luo Q.Z. [Effects of artesunate on cigarette smoke-induced lung oxidative damage in mice and the expression of Nrf2 and the possible mechanism] Zhonghua Yi Xue Za Zhi. 2016;96(12):960–965. doi: 10.3760/cma.j.issn.0376-2491.2016.12.011. [DOI] [PubMed] [Google Scholar]
- 70.Jiang L. Artesunate attenuates lung injury in paraquat-intoxicated rats via downregulation of inflammatory cytokines. Clin. Lab. 2015;61(11):1601–1607. doi: 10.7754/clin.lab.2015.141244. [DOI] [PubMed] [Google Scholar]
- 71.Hu Y.Z. [Preparation of liposomal artesunate dry powder inhalers and the effect on the acute lung injury of rats] Yao Xue Xue Bao. 2016;51(12):1906–1912. [PubMed] [Google Scholar]
- 72.Liu Z. Artesunate inhibits renal ischemia reperfusion-stimulated lung inflammation in rats by activating HO-1 pathway. Inflammation. 2018;41(1):114–121. doi: 10.1007/s10753-017-0669-3. [DOI] [PubMed] [Google Scholar]
- 73.Cao T.H. Artesunate protects against sepsis-induced lung injury via heme Oxygenase-1 modulation. Inflammation. 2016;39(2):651–662. doi: 10.1007/s10753-015-0290-2. [DOI] [PubMed] [Google Scholar]
- 74.Cheng C. Anti-malarial drug artesunate attenuates experimental allergic asthma via inhibition of the phosphoinositide 3-kinase/Akt pathway. PLoS One. 2011;6(6):e20932. doi: 10.1371/journal.pone.0020932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Wei M. Dihydroartemisinin suppresses ovalbumin-induced airway inflammation in a mouse allergic asthma model. Immunopharmacol. Immunotoxicol. 2013;35(3):382–389. doi: 10.3109/08923973.2013.785559. [DOI] [PubMed] [Google Scholar]
- 76.Zhu H., Ji W. Dihydroartemisinin ameliorated ovalbumin-induced asthma in mice via regulation of MiR-183C. Med. Sci. Monit. 2019;25:3804–3814. doi: 10.12659/MSM.915399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Li J. Effect of artemisinin and neurectomy of pterygoid canal in ovalbumin-induced allergic rhinitis mouse model. Allergy Asthma Clin. Immunol. 2018;14:22. doi: 10.1186/s13223-018-0249-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Wang Y. Artesunate attenuates airway resistance in vivo and relaxes airway smooth muscle cells in vitro via bitter taste receptor-dependent calcium signalling. Exp. Physiol. 2019;104(2):231–243. doi: 10.1113/EP086824. [DOI] [PubMed] [Google Scholar]
- 79.Yang D. Dihydroartemisinin supresses inflammation and fibrosis in bleomycine-induced pulmonary fibrosis in rats. Int. J. Clin. Exp. Pathol. 2015;8(2):1270–1281. [PMC free article] [PubMed] [Google Scholar]
- 80.Wang C. Anti-profibrotic effects of artesunate on bleomycin-induced pulmonary fibrosis in Sprague Dawley rats. Mol. Med. Rep. 2015;12(1):1291–1297. doi: 10.3892/mmr.2015.3500. [DOI] [PubMed] [Google Scholar]
- 81.Wang Y. Artesunate modulates expression of matrix metalloproteinases and their inhibitors as well as collagen-IV to attenuate pulmonary fibrosis in rats. Genet. Mol. Res. 2016;15(2) doi: 10.4238/gmr.15027530. [DOI] [PubMed] [Google Scholar]
- 82.Wang C.M. [Relationship between artesunate influence on the process of TGF-beta1 induced alveolar epithelial cells transform into mesenchymal cells and on idiopathic pulmonary fibrosis] Yao Xue Xue Bao. 2014;49(1):142–147. [PubMed] [Google Scholar]
- 83.Ma H. The effects of artesunate on the expression of EGFR and ABCG2 in A549 human lung cancer cells and a xenograft model. Molecules. 2011;16(12):10556–10569. doi: 10.3390/molecules161210556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Zhang Z.Y. [Artesunate combined with vinorelbine plus cisplatin in treatment of advanced non-small cell lung cancer: a randomized controlled trial] Zhong Xi Yi Jie He Xue Bao. 2008;6(2):134–138. doi: 10.3736/jcim20080206. [DOI] [PubMed] [Google Scholar]
- 85.World Health Organisation . 2020. Coronavirus Disease (COVID-19) Pandemic.https://www.who.int/emergencies/diseases/novel-coronavirus-2019 Available from: [Google Scholar]
- 86.Gautret P. Hydroxychloroquine and azithromycin as a treatment of COVID-19: results of an open-label non-randomized clinical trial. Int. J. Antimicrob. Agents. 2020:105949. doi: 10.1016/j.ijantimicag.2020.105949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.McKee D.L. Candidate drugs against SARS-CoV-2 and COVID-19. Pharmacol. Res. 2020:104859. doi: 10.1016/j.phrs.2020.104859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Megarbane B. Chloroquine and hydroxychloroquine to treat COVID-19: between hope and caution. Clin Toxicol (Phila) 2020:1–2. doi: 10.1080/15563650.2020.1748194. [DOI] [PubMed] [Google Scholar]
- 89.Fu L. Clinical characteristics of coronavirus disease 2019 (COVID-19) in China: a systematic review and meta-analysis. J. Infect. 2020 doi: 10.1016/j.jinf.2020.03.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Guastalegname M., Vallone A. Could chloroquine /hydroxychloroquine be harmful in Coronavirus Disease 2019 (COVID-19) treatment? Clin. Infect. Dis. 2020 doi: 10.1093/cid/ciaa321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Zhou P. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020;579(7798):270–273. doi: 10.1038/s41586-020-2012-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Dai L. Novel multiarm polyethylene glycol-dihydroartemisinin conjugates enhancing therapeutic efficacy in non-small-cell lung cancer. Sci. Rep. 2014;4:5871. doi: 10.1038/srep05871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Sun Q. Enhanced apoptotic effects of dihydroartemisinin-aggregated gelatin and hyaluronan nanoparticles on human lung cancer cells. J Biomed Mater Res B Appl Biomater. 2014;102(3):455–462. doi: 10.1002/jbm.b.33023. [DOI] [PubMed] [Google Scholar]
- 94.Zhang X. Dihydroartemisinin loaded nanostructured lipid carriers (DHA-NLC): evaluation of pharmacokinetics and tissue distribution after intravenous administration to rats. Pharmazie. 2010;65(9):670–678. [PubMed] [Google Scholar]
- 95.Buragohain P. Synthesis of a novel series of artemisinin dimers with potent anticancer activity involving Sonogashira cross-coupling reaction. Bioorg. Med. Chem. Lett. 2014;24(1):237–239. doi: 10.1016/j.bmcl.2013.11.032. [DOI] [PubMed] [Google Scholar]
- 96.Yang Y. Enhanced delivery of artemisinin and its analogues to cancer cells by their adducts with human serum transferrin. Int. J. Pharm. 2014;467(1):113–122. doi: 10.1016/j.ijpharm.2014.03.044. [DOI] [PubMed] [Google Scholar]
- 97.Hien T.T. Orally formulated artemisinin in healthy fasting Vietnamese male subjects: a randomized, four-sequence, open-label, pharmacokinetic crossover study. Clin. Ther. 2011;33(5):644–654. doi: 10.1016/j.clinthera.2011.04.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Morris C.A. Review of the clinical pharmacokinetics of artesunate and its active metabolite dihydroartemisinin following intravenous, intramuscular, oral or rectal administration. Malar. J. 2011;10:263. doi: 10.1186/1475-2875-10-263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Wu L.-l. Vol. 31. 2017. pp. 1121–1128. (Determination of Artemisitene in Rat Plasma by Ultra-Performance Liquid Chromatography/Tandem Mass Spectrometry and Its Application in Pharmacokinetics). (13) [DOI] [PubMed] [Google Scholar]