

Abstract
The disease ecology community has struggled to come to consensus on whether biodiversity reduces or increases infectious disease risk, a question that directly affects policy decisions for biodiversity conservation and public health. Here, we summarize the primary points of contention regarding biodiversity–disease relationships and suggest that vector-borne, generalist wildlife and zoonotic pathogens are the types of parasites most likely to be affected by changes to biodiversity. One synthesis on this topic revealed a positive correlation between biodiversity and human disease burden across countries, but as biodiversity changed over time within these countries, this correlation became weaker and more variable. Another synthesis—a meta-analysis of generally smaller-scale experimental and field studies—revealed a negative correlation between biodiversity and infectious diseases (a dilution effect) in various host taxa. These results raise the question of whether biodiversity–disease relationships are more negative at smaller spatial scales. If so, biodiversity conservation at the appropriate scales might prevent wildlife and zoonotic diseases from increasing in prevalence or becoming problematic (general proactive approaches). Further, protecting natural areas from human incursion should reduce zoonotic disease spillover. By contrast, for some infectious diseases, managing particular species or habitats and targeted biomedical approaches (targeted reactive approaches) might outperform biodiversity conservation as a tool for disease control. Importantly, biodiversity conservation and management need to be considered alongside other disease management options. These suggested guiding principles should provide common ground that can enhance scientific and policy clarity for those interested in simultaneously improving wildlife and human health.
Subject terms: Ecology, Medical research
There has been intense debate as to whether biodiversity increases or reduces the risk of infectious disease. This Review is the result of researchers from both sides of the debate attempting to reach a consensus.
Main
Humans alter biodiversity in complex ways; sometimes, human impacts add biodiversity to ecosystems through species introductions or restoration efforts, but in many cases they cause local biotic homogenization and loss of native biodiversity1–3. These changes to biodiversity have the potential to affect human and wildlife infectious diseases through a variety of mechanisms, and understanding when, where and how this might happen could be important for predicting and mitigating disease outbreaks. Biodiversity–infectious disease interactions have roots as far back as the classic work of Charles Elton4 and have influenced the fields of integrated pest management of crops5. More recently, this topic has generated intense debate in the literature and media, and the question of its generality has become a subject of lively discussion in the discipline of infectious disease ecology6–19. Much published research supports the dilution effect hypothesis, which proposes that biodiversity can reduce the abundance of a particular parasite species per host and thus reduce the risk of infectious disease caused by that parasite20–23. The dilution effect therefore predicts that biodiversity loss should increase infectious disease burden (with caveats noted below in ‘Points of agreement and contention’). Other studies support alternatives to the dilution effect, such as no relationship, a context-dependent relationship or an amplification effect6,11–14,24–26—defined by Keesing et al.27 as the opposite of the dilution effect, or a positive relationship between biodiversity and risk of a particular infectious disease. Debate has also centred on whether managing species composition or biodiversity in general is more effective at reducing risk and whether human diseases are exceptions to general rules about biodiversity–infectious disease associations6,9,13,14,16,17,28–30. Hypotheses regarding biodiversity–disease relationships have potentially important public health, management and policy implications, because they imply that changes to biodiversity could increase or decrease disease, thus suggesting that biodiversity conservation could have unaccounted costs or benefits, respectively1,6,11–14,20,24–26,31. Despite this, there are few examples where biodiversity–disease relationships have been used to set policy or reduce disease burdens. We define biodiversity conservation as preserving functioning ecosystems with predominantly native species and note that this is distinct from single-species conservation or restoration (see Supplementary Information for discussion of restoration and biodiversity augmentation versus biodiversity conservation), which might also affect parasite transmission. The policy and management implications of this debate are real, given that there is a precedent for wildlife management to be used to control disease. For example, based on guidance from ecological models and theory, estate owners in Scotland have culled mountain hares in an attempt to reduce nematode infections in grouse32, and the British government has culled European badgers in an attempt to limit the spread of Mycobacterium bovis, the causative agent of bovine tuberculosis33.
An improved understanding of biodiversity–disease relationships could lead to considerable progress towards disease control. As individuals with a diversity of perspectives on biodiversity–disease relationships, we joined together to summarize the primary points of contention underlying the debate and to identify potential common ground, building upon several recent reviews of biodiversity–disease relationships13,15,20,31,34,35. We first describe the host–parasite systems that are most likely to be affected by changes to biodiversity. Throughout this Review, ‘biodiversity’ is generally used to refer to species richness, whereas ‘species composition’ accounts for the identity or relative frequencies of species in a community. Next, we summarize the points of disagreement in the biodiversity–disease literature. We then describe the state of the science by comparing the results of two recent synthetic statistical analyses of biodiversity–disease relationships. If biodiversity management as a tool for disease control is considered in the broader context of other disease management options, we contend that it could have two primary benefits. First, it might prevent zoonotic and wildlife diseases from becoming problematic where they currently are not. Second, it might provide a means for managing existing diseases where no or few conventional interventions are available. If researchers can agree on this common ground, the resulting scientific and policy clarity could simultaneously improve ecosystem and human health.
How can biodiversity affect infectious disease?
Many pathogens might not interact with biodiversity (Fig. 1) or are well controlled in some settings by sanitation, drugs, pesticides or vaccines28. Other pathogens are likely to interact with biodiversity, including zoonotic diseases, which are caused by parasites that can be transmitted from animals to humans, and vector-borne diseases, which are transmitted by biting arthropods36–38. Hence, it is critical to identify the types of host–parasite systems most likely to be affected by biodiversity before asking whether positive, negative or neutral biodiversity–disease relationships predominate.
Fig. 1. The frequency of interactions with biodiversity and transmission potential are likely to influence whether a parasite will be weakly or strongly affected by biodiversity.
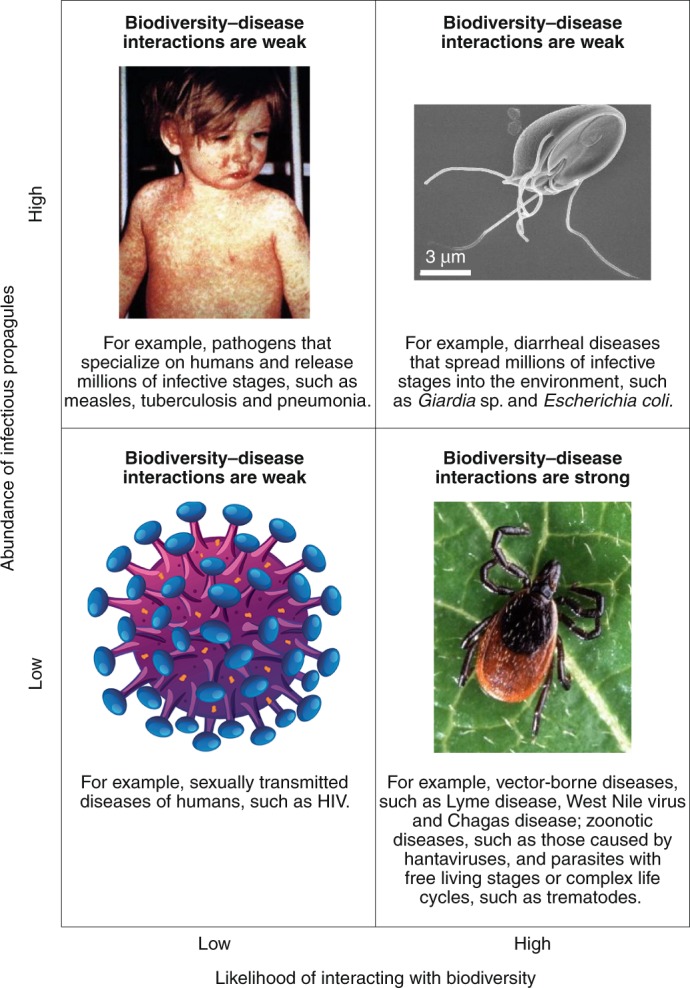
Transmission potential can be a product of releasing millions of infectious stages into the environment (high), or the number of blood meals a vector can take in its lifetime, or the number of sexual partners humans generally have in monogamous societies (low). When transmission potential is low, lost transmission events have a higher potential of reducing disease risk. However, to the extent that biodiversity regulates susceptible hosts27 and diverse microbiomes protect against infectious diseases (for example, refs. 20,34,47), some of these expectations will need re-evaluation.
Measles photo, CDC/NIP/Barbara Rice; Giardia lamblia, CDC/Janice Haney Carr; HIV image, Matthew Cole / Alamy Stock Vector; tick photo, Scott Bauer, USDA Agricultural Research Service.
Biodiversity is unlikely to increase or decrease directly transmitted, specialist parasites without free-living stages, intermediate hosts or vectors (Figs. 1 and 2). Examples include sexually transmitted diseases of wild vertebrate and human hosts, such as simian and human immunodeficiency viruses, and the causative agents of human tuberculosis, measles, non-pandemic influenza and pneumonia, all of which have few interactions with species other than their host13,14,28 (Fig. 1). By contrast, parasites such as West Nile virus, which can infect humans and numerous species of birds29, the causative agent of Chagas disease, which can infect humans and various wild and domestic animals39,40, the bacterium causing leptospirosis, which is typically transmitted from rat excreta to humans41, and hantavirus, which can infect humans and numerous mammals42–44, might be more likely to be influenced by biodiversity31,45. The sensitivity of these parasites to biodiversity also depends on the number and distribution of infectious stages and how they are transmitted (Fig. 1). For example, systems in which potential host contacts are limited—such as vectors that take a limited number of blood meals in their lifetime—are more likely to be affected by host and non-host biodiversity than are parasites that spread billions of infective stages into the environment (but see ref. 46) (Fig. 1). Hence, multi-host parasites, wildlife parasites, parasites with complex life cycles or free-living infectious stages, parasites with generalist vectors, and zoonotic parasites would be predicted to respond most readily to changes to biodiversity (that is, to be biodiversity-responsive parasites; Fig. 1). However, to the extent that biodiversity (1) regulates the density of susceptible wildlife hosts that might then pass directly transmitted parasites amongst themselves27 or (2) influences microbiota that protect against infectious diseases (for example, refs. 20,34,47), some of these expectations will need re-evaluation. Among these diversity-responsive parasites, negative effects of biodiversity on disease risk (that is, prevalence or per-capita parasite abundance) support the dilution effect, positive effects support the amplification effect, and unimodal relationships support both dilution and amplification under different circumstances. Finally, all of these patterns could be sensitive to the biodiversity and disease metrics used and the scale at which they are observed14,48,49.
Fig. 2. Venn diagram depicting two primary disease management strategies, general proactive and targeted reactive approaches, and examples of each.
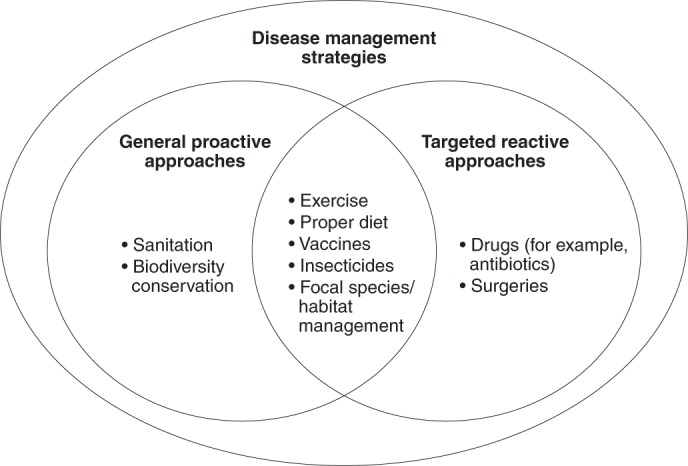
Most disease management strategies are either proactive or reactive but some can be both. If dilution occurs more frequently than amplification, we postulate that the value of general biodiversity conservation might be to prevent: (1) multi-host, zoonotic and wildlife diseases from becoming problematic; (2) diseases where specific key hosts are hard to manage; and (3) diseases where little is known about their ecology, because too little is known to hone any intervention to specific species. In contrast, when the key hosts are manageable, interventions might be targeted to specific species or habitats that are known to amplify or dilute disease, which might make the intervention more effective than general biodiversity conservation. To the extent that biodiversity regulates the density of susceptible hosts that might then pass directly transmitted pathogens amongst themselves27 or influences microbiota that protect against infectious diseases (for example, refs. 20,34,47), some of these hypotheses will need re-evaluation. Although this figure is presented as a dichotomy, it does not imply that each option is equally probable.
To facilitate policy and management decision making, it is important to (1) identify which diseases are likely to respond to biodiversity, (2) understand the relationships between biodiversity and disease risk, (3) link biodiversity change to change in both disease burden (that is, loss of fitness due to disease) and metrics of disease risk, such as infection prevalence, (4) consider the ecological and medical importance of a disease to a particular system or to humans, and (5) understand the trade-offs between biodiversity management and other land-use and public health options28,34 (see ‘Research frontiers’ section). Indeed, different response variables, such as disease burden, prevalence, intensity and force of infection, might respond differently to biodiversity.
Biologists generally agree that biodiversity affects some parasites and not others, so the measured effect of diversity on disease risk depends on whether all parasites or just diversity-responsive parasites are considered. This leaves two questions: (1) what fraction of infectious diseases interact with biodiversity (and through what mechanisms)? and (2) for those diseases that interact with biodiversity, how often is the outcome positive, negative, neutral or nonlinear? By analogy, because we would not expect an antibacterial compound to be effective against all pathogens, we would not ask how effective it is against pathogens other than bacteria. By this rationale, because we only expect biodiversity to interact with a subset of infectious diseases, when assessing whether biodiversity results in dilution or amplification, it is most relevant to consider only those pathogens that are likely to respond to biodiversity.
Points of agreement and contention
This section reviews the points of agreement and contention in biodiversity–infectious disease research.
What is the shape of the biodiversity–disease relationship?
For diversity-responsive parasites, the relationship between diversity and disease risk might be nonlinear. No parasites can exist where host richness equals zero11,12,35,50,51. For this reason, most dilution effect research has focused on how biodiversity reductions in intact communities affect some metric of disease given that a parasite can complete its entire life cycle in that community7,23,27,31,52.
Given the likelihood that biodiversity–disease relationships will be nonlinear, it is crucial to know the relevant ranges over which to evaluate diversity and over which diversity is related to conservation action. If unimodal biodiversity–disease relationships are often left-skewed or asymptotic, amplification effects should dominate, whereas if they are right-skewed, dilution should dominate14 (Fig. 3). Where the diversity of a community falls on these curves is also important, because if most communities fall in the right or left portions of unimodal diversity–disease curves, then dilution or amplification, respectively, will be most common, regardless of the direction of the skew (Fig. 3). We therefore must focus research on assessing the shape of diversity–disease risk relationships and identifying where systems generally fall along the x axis of diversity encompassed by these curves13,34,35,50 (see ‘Research frontiers’ section).
Fig. 3. Hypothetical relationships between biodiversity and disease risk.
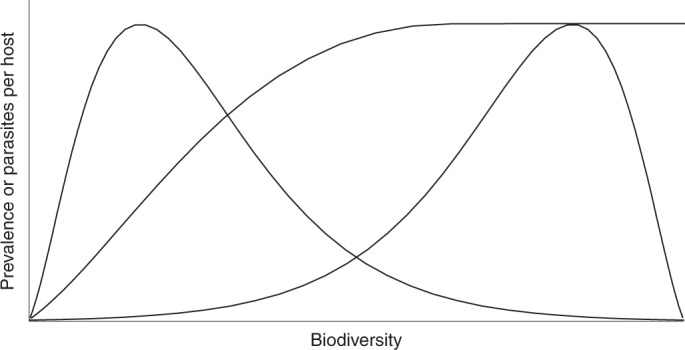
The right-skewed distribution suggests that dilution might occur more frequently, but less intensely than amplification because the relationship is moderately negative over a greater portion of the biodiversity gradient than it is strongly positive. The left-skewed distribution suggests that amplification might occur more frequently but less intensely than dilution, because the relationship is moderately positive over a greater portion of the biodiversity gradient than it is strongly negative. An asymptotic distribution suggests that amplification becomes increasingly moderate with biodiversity. In addition to the shape of biodiversity–disease relationships, the frequency with which each biodiversity level occurs in the environment will also affect the likelihood and intensity of dilution and amplification. These hypothetical curves underscore the importance of documenting the shape of biodiversity–disease relationships, which has rarely been accomplished empirically.
What is the influence of habitat transformation or species composition on pathogen transmission, relative to the direct influence of biodiversity?
Deforestation and urbanization can affect disease risk both directly and indirectly, via impacts on biodiversity, temperature, sanitation, access to health care, human population density, area of impervious surfaces, contact with biodiversity or other factors28,53. These multivariate relationships can make it difficult to disentangle the effects of biodiversity per se (see ref. 54 for a discussion of parasitism within the context of biodiversity–function relationships) on disease risk in changing landscapes9,28,35. Hence, it is crucial to control for these potentially confounding factors when testing for relationships between biodiversity and disease (for example, ref. 28). If these demographic, economic and environmental factors have stronger impacts on disease burden than does changing biodiversity per se, then management of biodiversity could have a relatively small effect on disease.
In contrast to direct biodiversity–disease relationships, changing species composition, rather than diversity per se, can affect disease risk. This hypothesis states that the presence of certain host species increases or decreases disease, and because many experimental studies manipulate species composition in conjunction with biodiversity, the effects of these factors are difficult to disentangle51,52,55. Species vary in their diluting and amplifying capacity based on their abundance, susceptibility and transmission potential, and thus certain species can disproportionately affect disease risk56. However, there is also support for biodiversity in general affecting disease, particularly when changes in diversity are substitutive—that is, adding new species to a community reduces the abundance of existing species (for example, refs. 57–60). If species composition rather than diversity per se has a large effect on disease, then managing particular species might be more effective than managing overall biodiversity. Some species, including many rodents, thrive in communities with few predatory species and thus low overall diversity: communities in which many zoonotic diseases can be amplified10. However, adding or sustaining top predator species in a community without increasing or maintaining biodiversity, respectively, could be difficult for predators that require an ample abundance and diversity of prey species. This observation that low-diversity communities contain a nested subset of their higher-diversity counterparts57 is an example of how diversity and species composition can be correlated20 (see ‘Research frontiers’ section). This correlation makes it challenging to disentangle composition from diversity in nature and to manage composition independent of biodiversity. Importantly, however, this correlation can make it easier to manage diseases than if diversity and composition were related to one another idiosyncratically, because managing diversity will de facto result in management of composition20 (see ‘Research frontiers’ section).
What are the mechanisms underlying biodiversity–disease relationships?
Some have suggested that the biodiversity–disease literature lacks evidence for convincing causal mechanisms for a relationship between biodiversity and disease (for example, refs. 61,62). However, decades of literature on the dilution effect hypothesized explicit mechanisms23,50,52, and empirical support is growing for several of these. For example, communities with greater biodiversity might have greater densities of non-competent hosts, which can dilute disease by reducing encounters with vectors (for example, wasted bites)27, competing with competent hosts and reducing their densities44,49, or consuming free-living parasites or infected hosts (if the parasites are not trophically transmitted) (for example, refs. 58,59). When controlling for parasite exposure, traits of individual species may be predictive of whether they are likely to be a competent (amplifying) or non-competent (diluting) host for a focal parasite, including phylogenetic relatedness to focal hosts36,63–65, life history and immunity or other defence strategies66–70, numerical abundance63,64,71, predilection to consume parasites relative to hosts58,59, and contact rates or connectedness within communities72. Alternatively, increasing biodiversity might theoretically increase the density of competent hosts, which amplify parasite burden by increasing rates of parasite or vector reproduction, by serving as long-lived reservoirs of infection, or by having high contact rates with other hosts73.
The extent to which mechanisms that are affected by species composition relate to biodiversity depends on community (dis)assembly—how species are added to (or lost from) a community—which can be affected by the previously mentioned species traits55,74,75. Community assembly can range from substitutive, in which individuals of a new species replace individuals of existing species in a community, to additive, in which adding new species adds more individuals to a community. Often, both substitutive and additive assembly occur within a community, because systems typically start assembling additively and shift to substitutive assembly as niches become more fully occupied and competition increases76.
Some community assembly mechanisms could produce amplification. For example, when community (dis)assembly is additive, amplification is expected because host densities increase as new host species are added to a community. When community (dis)assembly is substitutive, amplification can occur when the addition of individuals of new, competent host species reduce the density of less competent host species27,50,52. Amplification or dilution can occur when competent hosts or non-competent hosts, respectively, are added to or subtracted from communities via the sampling effect (that is, more diverse communities are more likely to contain a host species that either strongly increases or decreases disease) (for example, ref. 77). Additionally, the overall burden of multiple diseases might increase through the cumulative effect of greater pathogen diversity78. If considering the cumulative effects of all pathogen species on a host population, host diversity could decrease the disease burden of individual pathogens (dilution) while simultaneously increasing the combined burden of all pathogens (see ‘Extrapolations beyond collected data’ section for further discussion and counter examples). Although this mechanism for an increase in disease is important for individual health, it would not meet the traditional definition of amplification27, which focuses on the prevalence or abundance of individual pathogen species in hosts, not the combined effects of all pathogens or an increase in pathogen richness.
Dilution should predominate when community disassembly is substitutive and competent hosts are abundant or are resilient to biodiversity loss. We focus on community disassembly because most of the literature on the dilution effect assumes biodiversity loss and thus disassembly. A commonly hypothesized mechanism for dilution assumes that: (1) parasites experience greater selection to infect abundant rather than rare hosts; (2) abundant hosts make considerable investments into reproduction, growth, and/or dispersal that might come at the expense of defences against parasites; (3) abundant hosts are more likely to colonize and less likely to be extirpated from ecosystems; and (4) adding rare hosts reduces the abundance of common host species in high-diversity communities50,70. When these assumptions are met, a right-skewed biodiversity–disease relationship should result (Fig. 3), and abundant and widespread hosts might be amplifying hosts, while hosts with greater diluting potential would be added to communities as biodiversity increases, or would be lost from communities when they become fragmented or disturbed7,34,76,79. A combination of mesocosm experiments and field surveys demonstrates that the most abundant and widespread amphibian hosts are also the most competent hosts for a parasitic trematode species, supporting the notion that community disassembly processes function in a manner consistent with the dilution effect for this parasite57. Similarly, community disassembly processes support dilution effects documented for Lyme disease20,45,55,74. In a recent study, when plant communities were (dis)assembled randomly, dilution was not observed, but when they (dis)assembled naturally, biodiversity significantly reduced disease, again highlighting the potential importance of natural assembly processes80. Nevertheless, the generality of this dilution mechanism remains unclear, in part because the frequency with which natural communities assemble and disassemble in a substitutive versus additive manner is not well characterized. Although we call for additional work, we emphasize that substantial progress has been made on mechanisms underlying biodiversity–disease relationships.
Extrapolations beyond collected data
Even when evidence supports a positive or negative biodiversity–disease relationship, there are often several links in the causal pathway between change in biodiversity and change in disease. Few studies have tracked all the links in this causal chain and links in the causal chain can act in opposite directions. For example, there are several links in the hypothesized causal chain connecting high vertebrate diversity to a high probability of zoonotic disease establishment and transmission in humans20, but not all of these pathways have strong scientific support10. There is considerable support for the argument that host diversity begets parasite diversity26,81–84. However, there is not consistent support for the hypothesis that increasing parasite diversity increases disease burden10. In fact, recent experimental work on wildlife and plants diseases57,78,85 showed that higher pathogen diversity can be associated with lower disease severity of individual pathogens in both plants85 and animals78. Ultimately, the impact of parasite species diversity on the health of hosts will depend on how the additional parasite species affect disease burden and severity (see ‘Research frontiers’ section).
Do biodiversity–disease relationships depend on scale?
The dependence of dilution and amplification on spatial scale has been a common thread throughout the biodiversity–disease debate11,12,35. There are two main ways in which scale might influence the relationship between biodiversity and disease. First, Johnson et al.34 proposed that any relationship between biodiversity and infectious disease should be strongest at local scales and weaken at larger scales. This prediction arises because species interactions affecting dilution and amplification occur at relatively small spatial scales, whereas abiotic factors like climate tend to dominate as drivers of biological pattern at larger spatial scales. Cohen et al.48 found support for this hypothesis in the amphibian chytrid fungus, West Nile virus and the bacterium that causes Lyme disease: at small spatial scales, host richness was a significant predictor of disease prevalence, whereas at larger spatial scales, the distribution of pathogens was more strongly influenced by climate and human population density48. Second, it is possible that the diluting capacity of a non-competent host might be most observable at small scales, where encounter reduction can occur, whereas the amplifying effect of a competent host might be most observable at larger temporal and spatial scales73. For example, for vector-transmitted diseases, removing a competent host can initially increase vector abundance on alternative hosts, suggesting a dilution effect, while over longer time and larger spatial scales, the removal of the competent host can ultimately cause decreases in the vector population73. Importantly, these results suggest that at least some of the variation in outcomes across biodiversity–disease studies could be a product of variation in the scales at which studies were conducted and at which mechanisms operate (see ‘Synthesizing the evidence’ and ‘Research frontiers’ sections). Indeed, biodiversity–disease studies have occurred from global to local scales6,81,86, while conservation actions and public health interventions generally occur at intermediate scales within countries6,28,86. In general, the most relevant studies are those conducted at temporal scales long enough to encompass pathogen life cycles and at spatial scales that are most relevant to both biodiversity–disease interactions and conservation, which are generally at the local to regional scales35.
Synthesizing the evidence
Two recent quantitative syntheses have examined the biodiversity–disease relationship; each has different strengths, limitations and foci. These two syntheses attempted to assess the relative frequencies of positive versus negative biodiversity–disease relationships, whether such relationships are detectable for human diseases despite the influence of other forces on disease burden (for example, wealth and disease control efforts), and whether these relationships depend on scale17,28.
In a systematic meta-analysis of the published literature on biodiversity–disease studies, which included 202 effect sizes for 61 parasite species, negative biodiversity–disease associations were common17 (see ref. 87 for similar findings among plant diseases). Negative diversity–disease associations were equally strong for zoonotic and wildlife diseases, and the meta-analysis did not reveal any significant context dependencies17, nor evidence of publication bias for zoonotic diseases18, though it is possible that null and amplification effects are underrepresented in the literature (that is, system selection rather than publication bias) (Table 1). Additionally, although Civitello et al.17 did not explicitly quantify the scale of the studies in their meta-analysis, there were no significant differences in effect sizes between smaller-scale experimental and larger-scale correlative field studies, indicating that, within the range of scales at which the studies were conducted, scale was unlikely to affect the nature or strength of dilution effects17,18. These results suggest that, among the diseases for which relationships with biodiversity have been tested, a negative association (1) is frequently reported for wildlife and human diseases, (2) is robust across ecological contexts, and (3) is consistent across the spatial scales studied and between experimental and observational studies. However, many important human parasites have not been studied under these conditions and therefore could not be included in this meta-analysis, and many of the studies in the meta-analysis quantified infection prevalence or parasite abundance rather than disease burden (Table 1). Finally, in a meta-analytic framework, it is difficult to separate effects of species composition (for example, sampling effects) from effects of biodiversity per se, although the meta-analysis included several studies that did separate these effects.
Table 1.
Characteristics that differ between the Civitello et al. and Wood et al. studies
| Studies within the Civitello et al.17 meta-analysis | Wood et al.28 within-country through time analysis | Wood et al.28 across-country analysis |
|---|---|---|
| Typically, include only relevant diversity. | Includes diversity that is of questionable relevance to hosts. | Includes diversity that is of questionable relevance to hosts. |
| Typically, do not homogenize patterns across communities. | Homogenizes patterns across communities. | Homogenizes patterns across communities. |
| Typically, compare same ecosystems and species. | Compares same ecosystems and species. | Compares different ecosystems and species. |
| Diversity varies mostly because of non-biogeographic differences, such as anthropogenic influences. | Diversity varies mostly because of non-biogeographic differences, such as anthropogenic influences. | Diversity varies mostly because of biogeographic differences. |
| Motivated by mechanism. | Motivated by mechanism. | Mechanism not necessarily clear. |
| Mix of manipulative and observational studies. | Only observational studies. | Only observational studies. |
| Generally small scale with some spatiotemporal scales smaller than what is relevant in nature. | Medium scale. | Largest scale. |
| Could have system selection bias. | No system selection bias. | No system selection bias. |
| Dilution | Ambiguous | Amplification |
A second synthesis tested for associations between environmental, demographic, economic and social factors and disease burden for the 24 most important human infectious diseases tracked by the World Health Organization, of which 11 were potentially affected by biodiversity28. To do so, Wood et al.28 conducted a global analysis investigating across-country associations between disease burden (disability adjusted life years; DALYs) and biodiversity (country-level bird and mammal species densities), while controlling for other potential drivers of disease burden, such as wealth, forestation and urbanization. All vector-borne or zoonotic diseases (that is, biodiversity-responsive) examined in this study were positively associated with biodiversity at the across-country scale, though only five of the associations were statistically significant. By contrast, when considering changes in biodiversity and disease burden over time within each country, coefficients for seven diseases were negative (one significantly), and four were positive (one significantly). Thus, biodiversity effects were less positive at the within-country scale, relative to comparisons among countries28. Because the study is global, it relies on country-scale estimates of biodiversity, which include species not directly relevant to each disease (for example, total bird diversity in disease systems for which only mammals can become infected) (Table 1). Additionally, the spatial scale of analysis (intermediate-size countries) is larger than the scale at which species interactions occur, but is a scale at which some policy decisions are made.
How can two synthetic analyses published two years apart come to such divergent conclusions? We hypothesize that the discrepancies arise in part from differences in the spatial scales and methods of the studies (Table 1). Civitello et al.17 used published studies that experimentally investigated diversity–disease associations at logistically feasible scales that were substantially smaller than the within- and across-country spatial extents and grains included in Wood et al.28 (see Source Data Fig. 4). In fact, the median replicate size for studies on zoonotic parasites in Civitello et al.17, as compiled by Halliday and Rohr88 (for studies with ≥3 biodiversity levels and rounding all replicate sizes to the nearest order of magnitude), was 1.5 km2, whereas the median replicate size for the analyses in Wood et al.28 was 321,489 km2. As discussed above, the association between biodiversity and chytrid, West Nile, and Lyme disease declined with spatial scale in multiple regression analyses48, consistent with the notion that mechanisms for dilution might operate at smaller scales than mechanisms for amplification. Clearly, the role of spatial scale in affecting the net effect of diversity on disease requires more attention.
We re-analysed data from Wood et al.28 to ask whether effect sizes in a single country through time differed from effect sizes across countries at a given time. Any disease designated as biodiversity-responsive by Wood et al.28 was considered biodiversity-responsive for this re-analysis. We conducted a general mixed-effects linear model in which we treated as the response variable their 11 pairs of regression coefficients (one for the across-space comparison and one for the across-time comparison) for the effect of biodiversity on human disease burden of biodiversity-responsive diseases, with time versus space as the predictors, and the disease identity as a random effect (coefficients taken from the full model; see Fig. 1 of ref. 28). The positive coefficient between biodiversity and disease decreased significantly when transitioning from the across-country global comparison to the within-country, across-time comparison (F1,10 = 13.59, P = 0.0042, Fig. 4), from significantly positive at the cross-country global scale to non-significant at the within-country scale over time. This re-analysis of the Wood et al.28 study suggests that when spatial scale is reduced from across to within countries (by considering a single country over time), biodiversity–disease relationships become less positive (Fig. 4). However, numerous other important factors differ between the Wood et al.28 and Civitello et al.17 studies (Table 1). For example, most studies in Civitello et al.17 assessed parasite prevalence or abundance, whereas Wood et al.28 analysed DALYs, which represent the burden of infection rather than parasite presence or abundance. Thus, for several reasons, we cannot definitively attribute differences in effect size and magnitude to differences in scale (Table 1).
Fig. 4. Hedges’ g effect sizes.
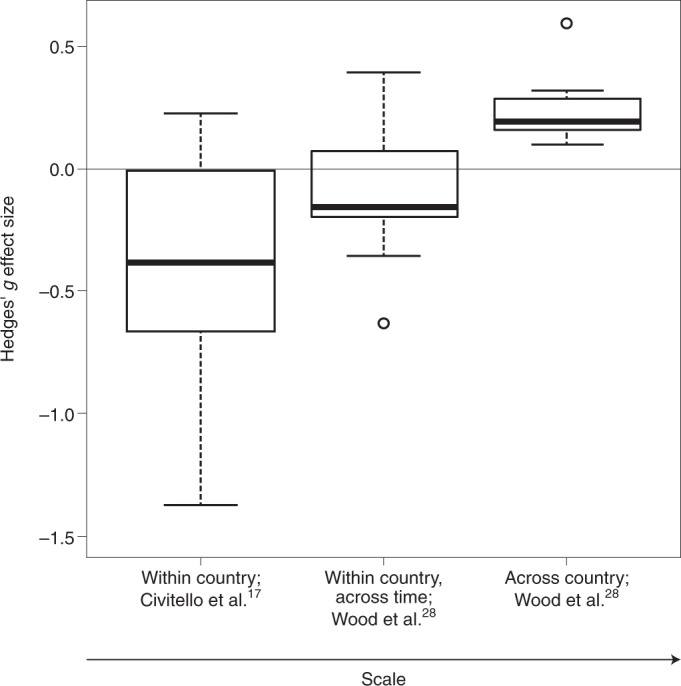
Effect sizes are shown for the association between biodiversity and zoonotic parasites (plus typhoid, because Wood et al.28 suggest it is biodiversity-responsive) at the cross-country and within-country (through time) scales (median replicate size: 321,489 km2; n = 11) from Wood et al.28 and various smaller-scale studies (median replicate size: 1.5 km2; n = 12) compiled by Civitello et al.17. Hedges’ g was provided by Civitello et al.17, whereas Wood et al.28 provided standardized regression coefficients. We converted the standardized regression coefficients to the Hedges’ g used in Civitello et al.17 by multiplying these coefficients by the sample size bias adjustment, . To properly account for the lack of independence among multiple effect sizes within studies and for the same diseases in Civitello et al.17, we calculated a mean effect size for each study weighting by the inverse of the variance, and then used inverse variance weighting on those study-wise means to obtain a weighted mean for each disease (see Source Data Fig. 4 for data used to generate this figure). In the Wood et al.28 study, the cross-country coefficient is significantly greater than zero (z = 5.82, P < 0.001), whereas the within-country (over time) coefficient is negative but not significantly different from zero (z = –1.20, P = 0.116). However, the relationship between these mean coefficients and scale is significantly positive (F1,10 = 13.59, P = 0.0042), indicating that positive diversity–disease associations are more likely for among-country comparisons than for comparisons within a country, over time. Relative to Wood et al.28, smaller-scale studies compiled in Civitello et al.17 were more likely to find negative diversity–disease associations. However, other factors also differ between the Wood et al.28 and Civitello et al.17 studies so we cannot confidently attribute all of this difference to the effect of scale. The midline of each boxplot is the median, the lower and upper limits of the box are first and third quartiles, respectively, the whiskers extend to 1.5 times the interquartile range, and the circles are extreme data points. Note that one extreme Hedges’ g value from Civitello et al.17 at –4.92 (Leptospira spp.) is not shown but was used to calculate the median, quartiles and whiskers of the boxplot.
It is important that we determine whether the relationship between biodiversity and disease does vary with spatial scale and, if so, that we identify the mechanisms that drive this relationship. Another key research frontier is evaluating the scale at which diversity can be feasibly managed to reduce the burden of disease. When diversity at the scale of nations or continents predicts disease patterns, information on the diversity–disease association seems most useful in assessing general risk at large scales. Biogeographic patterns of diversity, however, are rarely if ever amenable to direct management for disease reduction. When diversity at local and regional scales predicts disease patterns, management of diversity will sometimes be feasible as a means of managing disease.
Towards common ground
Given that scientists struggle to predict which, when and where infectious diseases will become problematic, general preventative approaches that produce net reductions in disease could have considerable value (Fig. 2). Ecosystems regularly pose a threat of disease to humans and wildlife, but ecosystems vary in these threats. Thus, targeting conservation toward protecting ecosystems that are not currently posing a major threat of problematic disease to humans or wildlife might prevent increases in disease (Fig. 2). In contrast, when the goal is to manage a specific disease whose ecology is reasonably well understood, it may be simpler and more effective to manage the particular species (vectors or amplifying or diluting hosts) or habitats that are known to decrease or increase disease (for example, through vaccination, culling, predator supplementation and habitat manipulation) than to conserve biodiversity in general (Fig. 2). However, management of particular species or habitats might only be effective against a focal pathogen. By contrast, preservation of intact, functioning ecosystems and finding sustainable, equitable interventions that discourage human incursions into those ecosystems (for example, for logging and bush-meat hunting), could reduce the risk of transmission of multiple pathogens, even if these interventions are not the single most efficient control method for individual diseases. Thus, they could represent win–win scenarios for conservation and disease control.
When considering disease management strategies, the costs and benefits of each tactic and their alternatives must be evaluated thoroughly before implementation. For example, it is possible that the land needed for biodiversity conservation might have greater value to humans if it is used differently, such as for agriculture or development, though it is also worth noting that sometimes the value arising from agriculture or development disproportionately accrues to outside commercial interests, while the health tolls are disproportionately borne by local communities89. Just as importantly, there are many reasons to conserve, restore and manage biodiversity that are unrelated to infectious disease, including other ecosystem services as well as ethical, aesthetic and cultural motivations. Additional health benefits from conservation can help make such actions more palatable when weighed against other land-use options.
Research frontiers
There are several outstanding questions in the biodiversity–disease literature that we have organized into five research frontiers: (1) pattern and process in biodiversity–disease relationships; (2) the shape of biodiversity–disease relationships; (3) metrics of disease and diversity; (4) context dependencies; and (5) public health and conservation.
Pattern and process in biodiversity–disease relationships
The foundational principles of disease ecology rest on a few well-studied disease agents, with many studies describing associations between biodiversity and disease rather than revealing the mechanisms that drive those patterns. We submit that the field needs to diversify, by both developing a broader understanding of biodiversity–disease patterns (that is, testing associations between biodiversity and disease beyond the disease agents that are already well studied) and narrowing in on the processes (that is, the mechanisms) that generate these patterns. A few macro-ecological investigations of biodiversity–disease pattern have already been conducted at the global scale (for example, refs. 26,28,86), but there is substantial promise in adopting this approach at regional or local spatial scales, which could reveal the conditions under which biodiversity–disease relationships are ecologically influential48. To discover what drives these ecologically influential relationships, experimental studies are needed to isolate mechanisms. For instance, most of the mechanisms posited to explain the dilution effect involve substitutive community (dis)assembly, but it remains unclear how often community (dis)assembly is additive versus substitutive, how strongly this affects biodiversity–disease relationships, and whether the most competent hosts for infections are also the most robust to biodiversity loss under historical and current conditions20,31,34,75,90. The degree of correlation between species composition and biodiversity and the relative importance of stochastic and deterministic community assembly mechanisms are also not well established. The less deterministic community assembly is, the less effective biodiversity management might be as a tool for disease control. Finally, ecological communities can also change stochastically through time (that is, ecological drift), which could cause temporal shifts in the roles of different dilution mechanisms at a given location.
The shape of biodiversity–disease relationships
Although there is consensus that biodiversity–disease relationships must be unimodal for dilution to occur, their propensity to be skewed to the left or right, and where they sit relative to the peak is unclear. For right-skewed biodiversity–disease relationships, a larger proportion of the diversity axis would produce dilution, whereas for left-skewed biodiversity–disease relationships, a larger proportion of the diversity axis would produce amplification. An even more challenging but equally important issue is determining how species richness values in nature are distributed along the diversity axis, and whether these tend to cluster on the portion of that axis where increasing biodiversity increases disease, or vice versa (Fig. 3).
Metrics of disease and diversity
Metrics of disease and diversity can vary widely across biodiversity–disease studies, but whether the choice of metric influences the study outcome is poorly understood. For instance, because infections do not necessarily manifest in disease burden (that is, loss of host fitness), it is possible that parasite prevalence or intensity might be more sensitive to biodiversity than is disease burden. Like disease, biodiversity can also be measured or represented in many ways and this too has the potential to affect outcomes91. For instance, whether alpha, beta or gamma diversity affect disease differently is uncertain. Moreover, different subsets of biodiversity might have different effects on disease. For example, it remains unclear how parasite diversity affects the number of infections per host or disease78,85. Additionally, natural enemies of parasites (for example, predators58), symbionts of hosts (for example, host microbiomes47), and other non-host species can influence biodiversity–disease relationships, but the common mechanisms by which non-hosts affect infectious disease remain equivocal27,34,58.
Context dependencies
Some attribute disagreements over the biodiversity–disease relationship to an overemphasis on generality30; while there might be context dependencies for the biodiversity–disease relationship, we think it is worthwhile to seek the rules that govern when and where each form of biodiversity–disease relationship might emerge. For example, there might be common traits of parasite species that are affected by biodiversity and common traits of host species that amplify or dilute disease13,14,58,59. If all host species are unequal, then it raises the question: how strongly are biodiversity–disease relationships a product of the identity of particular host species (‘sampling effects’)1,34,51,58,92, an emergent property of biodiversity in general (‘complementarity effects’), or spurious correlations driven by other factors?
Other ecological disciplines have identified relationships similar to the dilution effect. For instance, in the plant–herbivore literature, host plant diversity decreases herbivory, a phenomenon coined associational resistance93. The biodiversity–ecosystem functioning literature has documented consistent positive relationships between biodiversity and ecosystem functions1, and the Red Queen hypothesis is based on the notion that genetic diversity within a host species decreases infections per host65. What are the commonalities and differences among the dilution effect, associational resistance, biodiversity–ecosystem functioning and Red Queen hypotheses1,17,54,58,65,71,93, and are any differences a product of context dependencies?
An important potential context dependency for biodiversity–disease relationships is spatial scale. How much of the variation in outcomes of biodiversity–disease studies arises from differences in the scale at which studies are conducted (spatial and temporal scales and the scope of the biodiversity gradient)12,34,48? There is a rich literature on the scale-dependent effects of ecological drivers on biodiversity patterns91. Can it inform the biodiversity–disease discipline? More specifically, recent work shows that species composition often changes when it is measured at different spatial scales, and that measures of biodiversity are therefore often uncorrelated across scales94. Can this phenomenon, coupled with neutral sampling effects, cause different biodiversity–disease patterns to emerge at different scales? How important is the temporal scale of community assembly in revealing novel mechanisms linking host diversity to disease95?
Public health and conservation
Much of the controversy surrounding the biodiversity–disease discipline has arisen from the suggestion that human health would benefit from biodiversity conservation. Not surprisingly, many questions and considerable work remain on the application of biodiversity management to public health. For instance, some studies suggest mutual interference among co-infecting pathogens, such that an increase in parasite diversity generally decreases overall infections75. What is the general shape of the response surface for the relationships among disease and per-capita parasite diversity and abundance, and what are the public health trade-offs between more cases of one disease versus more different types of parasitic infections? How large is the unique effect of biodiversity compared to the unique effect of factors that commonly co-vary with biodiversity loss due to habitat conversion to more urban settings, such as increases in sanitation, access to health care and area of impervious surfaces? What are the trade-offs of investing in conservation as a disease management approach versus other public health interventions?
Conclusions
We believe that most disease ecologists can agree on several elements of the biodiversity–disease debate. First, biodiversity should have a greater effect on multi-host, wildlife, vector-borne and zoonotic diseases, especially those parasites with complex life cycles and free-living stages, than on directly transmitted, host-specialist diseases. Second, the relationship between biodiversity and disease can be nonlinear, and identifying the shape of these relationships and where the diversity of a community falls on this continuum is critical for understanding when decreasing biodiversity will increase, decrease or have no effect on disease risk. Third, not all diseases are equal. Ebola is not the same as head lice, and from a conservation perspective, a disease causing widespread extirpations or extinctions is different from one that is not consistently causing wildlife declines. Fourth, biodiversity–disease relationships may be scale-dependent. Understanding the mechanisms that underlie this possible scale dependency could lead to insights into which management targets (biodiversity, particular species and human behaviour) and which scales of action are most effective for disease control. Outcomes with co-benefits for biodiversity and human health should be actively sought. Establishing consensus on these guiding principles will help to set a research and policy agenda for simultaneously improving ecosystem and human health.
Supplementary information
Supplementary Discussion
Acknowledgements
We are thankful to J. Cohen, A. Dobson, R. Holt, P. T. J. Johnson, A. M. Kilpatrick, T. Levi, C. Lively and M. Venesky for insightful discussions on biodiversity–disease relationships over the years, and J. Mihaljevic for constructive comments on this manuscript. We also cannot thank R. Ostfeld and F. Keesing enough for >1.5 years of invaluable input on this manuscript as invited co-authors and their contributions to the biodiversity–disease discipline in general. All co-authors were encouraged to document any points of disagreement or compromise in an Appendix to this paper and passed on that opportunity. This research was supported by grants from the National Science Foundation (EF-1241889), National Institutes of Health (R01GM109499, R01TW010286), US Department of Agriculture (NRI 2006-01370, 2009-35102-0543), and US Environmental Protection Agency (CAREER 83518801) to J.R.R., by a grant from the National Science Foundation (OCE-1829509), an Alfred P. Sloan Foundation Sloan Research Fellowship, a University of Washington Innovation Award, and a University of Washington Royalty Research Fund award to C.L.W., and by grants from the National Science Foundation (DEB-1518681), the Stanford University Woods Institute for the Environment, and the Hellman Faculty Scholars fund to E.A.M. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US Government.
Source data
Data used to generate Fig. 4.
Author contributions
J.R.R. initiated and wrote the Review and J.R.R., D.J.C., F.W.H., P.J.H., K.D.L., C.L.W. and E.A.M. contributed ideas and edited the manuscript.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary information is available for this paper at 10.1038/s41559-019-1060-6.
References
- 1.Cardinale BJ, et al. Biodiversity loss and its impact on humanity. Nature. 2012;486:59–67. doi: 10.1038/nature11148. [DOI] [PubMed] [Google Scholar]
- 2.Rohr JR, Bernhardt ES, Cadotte MW, Clements WH. The ecology and economics of restoration: when, what, where, and how to restore ecosystems. Ecol. Soc. 2018;23:15. [Google Scholar]
- 3.Dornelas M, et al. Assemblage time series reveal biodiversity change but not systematic loss. Science. 2014;344:296–299. doi: 10.1126/science.1248484. [DOI] [PubMed] [Google Scholar]
- 4.Elton, C. S. The Ecology of Invasions by Animals and Plants (Methuen Publishing, 1958).
- 5.Van der Plank, J. E. Plant Diseases: Epidemics and Control (Academic Press, 1963).
- 6.Randolph SE, Dobson ADM. Pangloss revisited: a critique of the dilution effect and the biodiversity-buffers-disease paradigm. Parasitology. 2012;139:847–863. doi: 10.1017/S0031182012000200. [DOI] [PubMed] [Google Scholar]
- 7.Levi T, et al. Does biodiversity protect humans against infectious disease? Comment. Ecology. 2016;97:536–542. doi: 10.1890/15-354.1. [DOI] [PubMed] [Google Scholar]
- 8.Ostfeld RS. A Candide response to Panglossian accusations by Randolph and Dobson: biodiversity buffers disease. Parasitology. 2013;140:1196–1198. doi: 10.1017/S0031182013000541. [DOI] [PubMed] [Google Scholar]
- 9.Ostfeld RS, Keesing F. Straw men don’t get Lyme disease: response to Wood and Lafferty. Trends Ecol. Evol. 2013;28:502–503. doi: 10.1016/j.tree.2013.05.009. [DOI] [PubMed] [Google Scholar]
- 10.Ostfeld RS, Keesing F. Is biodiversity bad for your health? Ecosphere. 2017;8:e01676. [Google Scholar]
- 11.Lafferty KD, Wood CL. It’s a myth that protection against disease is a strong and general service of biodiversity conservation: response to Ostfeld and Keesing. Trends Ecol. Evol. 2013;28:503–504. doi: 10.1016/j.tree.2013.06.012. [DOI] [PubMed] [Google Scholar]
- 12.Wood CL, Lafferty KD. Biodiversity and disease: a synthesis of ecological perspectives on Lyme disease transmission. Trends Ecol. Evol. 2013;28:239–247. doi: 10.1016/j.tree.2012.10.011. [DOI] [PubMed] [Google Scholar]
- 13.Wood CL, et al. Does biodiversity protect humans against infectious disease? Ecology. 2014;95:817–832. doi: 10.1890/13-1041.1. [DOI] [PubMed] [Google Scholar]
- 14.Wood CL, et al. Does biodiversity protect humans against infectious disease? Reply. Ecology. 2016;97:542–545. doi: 10.1890/15-1503.1. [DOI] [PubMed] [Google Scholar]
- 15.Salkeld DJ, Padgett KA, Jones JH. A meta‐analysis suggesting that the relationship between biodiversity and risk of zoonotic pathogen transmission is idiosyncratic. Ecol. Lett. 2013;16:679–686. doi: 10.1111/ele.12101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Salkeld DJ, Padgett KA, Jones JH, Antolin MF. Public health perspective on patterns of biodiversity and zoonotic disease. Proc. Natl Acad. Sci. USA. 2015;112:E6261. doi: 10.1073/pnas.1517640112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Civitello DJ, et al. Biodiversity inhibits parasites: broad evidence for the dilution effect. Proc. Natl Acad. Sci. USA. 2015;112:8667–8671. doi: 10.1073/pnas.1506279112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Civitello DJ, et al. Reply to Salkeld et al.: Diversity-disease patterns are robust to study design, selection criteria, and publication bias. Proc. Natl Acad. Sci. USA. 2015;112:E6262. doi: 10.1073/pnas.1518473112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wilcox, C. The Hidden Dispute Over Biodiversity’s Health Benefits. The Atlantic (31 October 2017).
- 20.Keesing F, et al. Impacts of biodiversity on the emergence and transmission of infectious diseases. Nature. 2010;468:647–652. doi: 10.1038/nature09575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Laurenson MK, Norman R, Gilbert L, Reid HW, Hudson PJ. Identifying disease reservoirs in complex systems: mountain hares as reservoirs of ticks and louping‐ill virus, pathogens of red grouse. J. Anim. Ecol. 2003;72:177–185. [Google Scholar]
- 22.Norman R, Bowers R, Begon M, Hudson PJ. Persistence of tick-borne virus in the presence of multiple host species: tick reservoirs and parasite mediated competition. J. Theor. Biol. 1999;200:111–118. doi: 10.1006/jtbi.1999.0982. [DOI] [PubMed] [Google Scholar]
- 23.Van Buskirk J, Ostfeld RS. Controlling Lyme disease by modifying the density and species composition of tick hosts. Ecol. Appl. 1995;5:1133–1140. [Google Scholar]
- 24.Young H, Griffin RH, Wood CL, Nunn CL. Does habitat disturbance increase infectious disease risk for primates? Ecol. Lett. 2013;16:656–663. doi: 10.1111/ele.12094. [DOI] [PubMed] [Google Scholar]
- 25.Dunn RR. Global mapping of ecosystem disservices: the unspoken reality that nature sometimes kills us. Biotropica. 2010;42:555–557. [Google Scholar]
- 26.Dunn RR, Davies TJ, Harris NC, Gavin MC. Global drivers of human pathogen richness and prevalence. Proc. Royal Soc. Lond. B. 2010;277:2587–2595. doi: 10.1098/rspb.2010.0340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Keesing F, Holt RD, Ostfeld RS. Effects of species diversity on disease risk. Ecol. Lett. 2006;9:485–498. doi: 10.1111/j.1461-0248.2006.00885.x. [DOI] [PubMed] [Google Scholar]
- 28.Wood CL, McInturff A, Young HS, Kim D, Lafferty KD. Human infectious disease burdens decrease with urbanization but not with biodiversity. Philos. Trans. Royal Soc. B. 2017;372:20160122. doi: 10.1098/rstb.2016.0122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kilpatrick AM. Globalization, land use, and the invasion of West Nile Virus. Science. 2011;334:323–327. doi: 10.1126/science.1201010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Halsey S. Defuse the dilution effect debate. Nature Ecol. Evol. 2019;3:145–146. doi: 10.1038/s41559-018-0764-3. [DOI] [PubMed] [Google Scholar]
- 31.Ostfeld RS, Keesing F. Effects of host diversity on infectious disease. Annu. Rev. Ecol. Evol. Syst. 2012;43:157–182. [Google Scholar]
- 32.Laurenson MK, Norman R, Gilbert L, Reid HW, Hudson PJ. Mountain hares, louping-ill, red grouse and harvesting: complex interactions but few data. J. Anim. Ecol. 2004;73:811–813. [Google Scholar]
- 33.Donnelly CA, et al. Positive and negative effects of widespread badger culling on tuberculosis in cattle. Nature. 2006;439:843–846. doi: 10.1038/nature04454. [DOI] [PubMed] [Google Scholar]
- 34.Johnson PTJ, Ostfeld RS, Keesing F. Frontiers in research on biodiversity and disease. Ecol. Lett. 2015;18:1119–1133. doi: 10.1111/ele.12479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kilpatrick AM, Salkeld DJ, Titcomb G, Hahn MB. Conservation of biodiversity as a strategy for improving human health and well-being. Philos. Trans. Royal Soc. B. 2017;372:20160131. doi: 10.1098/rstb.2016.0131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lloyd-Smith JO, et al. Epidemic dynamics at the human-animal interface. Science. 2009;326:1362–1367. doi: 10.1126/science.1177345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Plowright RK, et al. Pathways to zoonotic spillover. Nat. Rev. Microbiol. 2017;15:502–510. doi: 10.1038/nrmicro.2017.45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Taylor LH, Latham SM, Woolhouse MEJ. Risk factors for human disease emergence. Philos. Trans. Royal Soc. Lond. B. 2001;356:983–989. doi: 10.1098/rstb.2001.0888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.das Chagas Xavier SC, et al. Lower richness of small wild mammal species and Chagas disease risk. PLOS Neglect. Trop. Dis. 2012;6:e1647. doi: 10.1371/journal.pntd.0001647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gottdenker NL, Chaves LF, Calzada JE, Saldaña A, Carroll CR. Host life history strategy, species diversity, and habitat influence Trypanosoma cruzi vector infection in changing landscapes. PLoS Neglect. Trop. Dis. 2012;6:e1884. doi: 10.1371/journal.pntd.0001884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Derne BT, Fearnley EJ, Lau CL, Paynter S, Weinstein P. Biodiversity and leptospirosis risk: a case of pathogen regulation? Med. Hypotheses. 2011;77:339–344. doi: 10.1016/j.mehy.2011.05.009. [DOI] [PubMed] [Google Scholar]
- 42.Dizney LJ, Ruedas LA. Increased host species diversity and decreased prevalence of Sin Nombre virus. Emerg. Infect. Dis. 2009;15:1012–1018. doi: 10.3201/eid1507.081083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Suzán G, et al. Experimental evidence for reduced rodent diversity causing increased hantavirus prevalence. PLoS ONE. 2009;4:e5461. doi: 10.1371/journal.pone.0005461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Luis AD, Kuenzi AJ, Mills JN. Species diversity concurrently dilutes and amplifies transmission in a zoonotic host–pathogen system through competing mechanisms. Proc. Natl Acad. Sci. USA. 2018;115:7979–7984. doi: 10.1073/pnas.1807106115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Ostfeld RS, Keesing F. Biodiversity and disease risk: the case of lyme disease. Conserv. Biol. 2000;14:722–728. [Google Scholar]
- 46.Herrera D, et al. Upstream watershed condition predicts rural children’s health across 35 developing countries. Nat. Commun. 2017;8:811. doi: 10.1038/s41467-017-00775-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Knutie SA, Wilkinson CL, Kohl KD, Rohr JR. Early-life disruption of host microbiota decreases later-life resistance to infections. Nat. Commun. 2017;8:86. doi: 10.1038/s41467-017-00119-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Cohen JM, et al. Spatial scale modulates the strength of ecological processes driving disease distributions. Proc. Natl Acad. Sci. USA. 2016;113:E3359–E3364. doi: 10.1073/pnas.1521657113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Strauss AT, Civitello DJ, Cáceres CE, Hall SR. Success, failure and ambiguity of the dilution effect among competitors. Ecol. Lett. 2015;18:916–926. doi: 10.1111/ele.12468. [DOI] [PubMed] [Google Scholar]
- 50.Ostfeld RS, Keesing F. Biodiversity series: the function of biodiversity in the ecology of vector-borne zoonotic diseases. Can. J. Zool. 2000;78:2061–2078. [Google Scholar]
- 51.Ostfeld, R. S., Thomas, M. B. & Keesing, F. in Biodiversity, Ecosystem Functioning, and Human Well-Being: An Ecological and Economic Perspective (eds Naeem, S. et al.) 209–216 (Oxford Univ. Press, 2009).
- 52.Schmidt KA, Ostfeld RS. Biodiversity and the dilution effect in disease ecology. Ecology. 2001;82:609–619. [Google Scholar]
- 53.Linske MA, Williams SC, Stafford KC, Ortega IM. Ixodes scapularis (Acari: Ixodidae) reservoir host diversity and abundance impacts on dilution of Borrelia burgdorferi (Spirochaetales: Spirochaetaceae) in residential and woodland habitats in Connecticut, United States. J. Med. Entomol. 2018;55:681–690. doi: 10.1093/jme/tjx237. [DOI] [PubMed] [Google Scholar]
- 54.Frainer A, McKie BG, Amundsen P-A, Knudsen R, Lafferty KD. Parasitism and the biodiversity-functioning relationship. Trends Ecol. Evol. 2018;33:260–268. doi: 10.1016/j.tree.2018.01.011. [DOI] [PubMed] [Google Scholar]
- 55.LoGiudice K, Ostfeld RS, Schmidt KA, Keesing F. The ecology of infectious disease: effects of host diversity and community composition on Lyme disease risk. Proc. Natl Acad. Sci. USA. 2003;100:567–571. doi: 10.1073/pnas.0233733100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Levi T, Keesing F, Holt RD, Barfield M, Ostfeld RS. Quantifying dilution and amplification in a community of hosts for tick-borne pathogens. Ecol. Appl. 2016;26:484–498. doi: 10.1890/15-0122. [DOI] [PubMed] [Google Scholar]
- 57.Johnson PTJ, Preston DL, Hoverman JT, Richgels KLD. Biodiversity decreases disease through predictable changes in host community competence. Nature. 2013;494:230–233. doi: 10.1038/nature11883. [DOI] [PubMed] [Google Scholar]
- 58.Rohr JR, et al. Predator diversity, intraguild predation, and indirect effects drive parasite transmission. Proc. Natl Acad. Sci. USA. 2015;112:3008–3013. doi: 10.1073/pnas.1415971112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Venesky MD, Liu X, Sauer EL, Rohr JR. Linking manipulative experiments to field data to test the dilution effect. J. Anim. Ecol. 2014;83:557–565. doi: 10.1111/1365-2656.12159. [DOI] [PubMed] [Google Scholar]
- 60.Mitchell CE, Tilman D, Groth JV. Effects of grassland plant species diversity, abundance, and composition on foliar fungal disease. Ecology. 2002;83:1713–1726. [Google Scholar]
- 61.Young HS, et al. Conservation, biodiversity and infectious disease: scientific evidence and policy implications. Philos. Trans. Royal Soc. B. 2017;372:20160124. doi: 10.1098/rstb.2016.0124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hosseini PR, et al. Does the impact of biodiversity differ between emerging and endemic pathogens? The need to separate the concepts of hazard and risk. Philos. Trans. R. Soc. B-Biol. Sci. 2017;372:20160129. doi: 10.1098/rstb.2016.0129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Clay, K. et al. in Infectious Disease Ecology: Effects of Ecosystems on Disease and of Disease on Ecosystems (eds Ostfeld, R. S., Keesing, F. & Eviner, V. T.) 145–178 (Princeton Univ. Press, 2008).
- 64.Parker IM, et al. Phylogenetic structure and host abundance drive disease pressure in communities. Nature. 2015;520:542–544. doi: 10.1038/nature14372. [DOI] [PubMed] [Google Scholar]
- 65.Lively CM. The effect of host genetic diversity on disease spread. Am. Nat. 2010;175:E149–E152. doi: 10.1086/652430. [DOI] [PubMed] [Google Scholar]
- 66.Han BA, Schmidt JP, Bowden SE, Drake JM. Rodent reservoirs of future zoonotic diseases. Proc. Natl Acad. Sci. USA. 2015;112:7039–7044. doi: 10.1073/pnas.1501598112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Luis AD, et al. A comparison of bats and rodents as reservoirs of zoonotic viruses: are bats special? Proc. Royal Soc. B. 2013;280:20122753. doi: 10.1098/rspb.2012.2753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Sears BF, Snyder PW, Rohr JR. Host life history and host-parasite syntopy predict behavioural resistance and tolerance of parasites. J. Anim. Ecol. 2015;84:625–636. doi: 10.1111/1365-2656.12333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Johnson PTJ, et al. Living fast and dying of infection: host life history drives interspecific variation in infection and disease risk. Ecol. Lett. 2012;15:235–242. doi: 10.1111/j.1461-0248.2011.01730.x. [DOI] [PubMed] [Google Scholar]
- 70.Previtali MA, et al. Relationship between pace of life and immune responses in wild rodents. Oikos. 2012;121:1483–1492. [Google Scholar]
- 71.Lively CM, Dybdahl MF. Parasite adaptation to locally common host genotypes. Nature. 2000;405:679–681. doi: 10.1038/35015069. [DOI] [PubMed] [Google Scholar]
- 72.Lloyd-Smith JO, Schreiber SJ, Kopp PE, Getz WM. Superspreading and the effect of individual variation on disease emergence. Nature. 2005;438:355–359. doi: 10.1038/nature04153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Buck JC, Perkins SE. Study scale determines whether wildlife loss protects against or promotes tick-borne disease. Proc. Royal Soc. Lond. B. 2018;285:20180218. doi: 10.1098/rspb.2018.0218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ostfeld RS, LoGiudice K. Community disassembly, biodiversity loss, and the erosion of an ecosystem service. Ecology. 2003;84:1421–1427. [Google Scholar]
- 75.Keesing F, Ostfeld RS. Is biodiversity good for your health? Science. 2015;349:235–236. doi: 10.1126/science.aac7892. [DOI] [PubMed] [Google Scholar]
- 76.Mihaljevic JR, Joseph MB, Orlofske SA, Paull SH. The scaling of host density with richness affects the direction, shape, and detectability of diversity-disease relationships. PLoS ONE. 2014;9:e97812. doi: 10.1371/journal.pone.0097812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Halliday, F. W., Heckman, R. W., Wilfahrt, P. A. & Mitchell, C. E. A multivariate test of disease risk reveals conditions leading to disease amplification Proc. Royal Soc. B. 284, 20171340 (The Royal Society). [DOI] [PMC free article] [PubMed]
- 78.Johnson PTJ, Hoverman JT. Parasite diversity and coinfection determine pathogen infection success and host fitness. Proc. Natl Acad. Sci. USA. 2012;109:9006–9011. doi: 10.1073/pnas.1201790109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Joseph MB, Mihaljevic JR, Orlofske SA, Paull SH. Does life history mediate changing disease risk when communities disassemble? Ecol. Lett. 2013;16:1405–1412. doi: 10.1111/ele.12180. [DOI] [PubMed] [Google Scholar]
- 80.Liu X, Chen F, Lyu S, Sun D, Zhou S. Random species loss underestimates dilution effects of host diversity on foliar fungal diseases under fertilization. Ecol. Evol. 2018;8:1705–1713. doi: 10.1002/ece3.3749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Hechinger RF, Lafferty KD. Host diversity begets parasite diversity: bird final hosts and trematodes in snail intermediate hosts. Proc. Royal Soc. Lond. B. 2005;272:1059–1066. doi: 10.1098/rspb.2005.3070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Johnson PTJ, et al. Habitat heterogeneity drives the host-diversity-begets-parasite-diversity relationship: evidence from experimental and field studies. Ecol. Lett. 2016;19:752–761. doi: 10.1111/ele.12609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Kamiya T, O’Dwyer K, Nakagawa S, Poulin R. Host diversity drives parasite diversity: meta‐analytical insights into patterns and causal mechanisms. Ecography. 2014;37:689–697. [Google Scholar]
- 84.Wood CL, Johnson PT. How does space influence the relationship between host and parasite diversity? J. Parasitol. 2016;102:485–494. doi: 10.1645/15-920. [DOI] [PubMed] [Google Scholar]
- 85.Rottstock T, Joshi J, Kummer V, Fischer M. Higher plant diversity promotes higher diversity of fungal pathogens, while it decreases pathogen infection per plant. Ecology. 2014;95:1907–1917. doi: 10.1890/13-2317.1. [DOI] [PubMed] [Google Scholar]
- 86.Guernier V, Hochberg ME, Guegan JFO. Ecology drives the worldwide distribution of human diseases. PLoS Biol. 2004;2:740–746. doi: 10.1371/journal.pbio.0020141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Huang Z, Van Langevelde F, Estrada-Peña A, Suzán G, De Boer W. The diversity–disease relationship: evidence for and criticisms of the dilution effect. Parasitology. 2016;143:1075–1086. doi: 10.1017/S0031182016000536. [DOI] [PubMed] [Google Scholar]
- 88.Halliday FW, Rohr JR. Measuring the shape of the biodiversity-disease relationship across systems reveals new findings and key gaps. Nat. Commun. 2019;10:5032. doi: 10.1038/s41467-019-13049-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Myers SS, et al. Human health impacts of ecosystem alteration. Proc. Natl Acad. Sci. USA. 2013;110:18753–18760. doi: 10.1073/pnas.1218656110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Strona G, Lafferty KD. Environmental change makes robust ecological networks fragile. Nat. Commun. 2016;7:12462. doi: 10.1038/ncomms12462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Chase JM, Knight TM. Scale‐dependent effect sizes of ecological drivers on biodiversity: why standardised sampling is not enough. Ecol. Lett. 2013;16:17–26. doi: 10.1111/ele.12112. [DOI] [PubMed] [Google Scholar]
- 92.Becker CG, et al. Partitioning the net effect of host diversity on an emerging amphibian pathogen. Proc. Royal Soc. Lond. B. 2014;281:20141796. doi: 10.1098/rspb.2014.1796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Barbosa P, et al. Associational resistance and associational susceptibility: having right or wrong neighbors. Annu. Rev. Ecol. Evol. Syst. 2009;40:1–20. [Google Scholar]
- 94.Chase JM, et al. Embracing scale‐dependence to achieve a deeper understanding of biodiversity and its change across communities. Ecol. Lett. 2018;21:1737–1751. doi: 10.1111/ele.13151. [DOI] [PubMed] [Google Scholar]
- 95.Halliday FW, Heckman RW, Wilfart PA, Mitchell CE. Past is prologue: host community assembly and the risk of infectious disease over time. Ecol. Lett. 2019;22:138–148. doi: 10.1111/ele.13176. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Discussion