

Abstract
Survivin, a member of the inhibitor of apoptosis (IAP) family, is upregulated in multiple cancers including ovarian cancer, but is rarely detectable in normal tissues. We previously reported that survivin promoted epithelial to mesenchymal transition (EMT) in ovarian cancer cells, suggesting that survivin may contribute to ovarian tumor metastasis and chemoresistance. In this study, we tested whether knockout or pharmacological inhibition of survivin overcomes chemoresistance and suppresses tumor metastasis. The genetic loss of survivin suppressed tumor metastasis in an orthotopic ovarian cancer mouse model. To pharmacologically test the role of survivin on ovarian tumor metastasis, we treated chemoresistant ovarian cancer cells with a selective survivin inhibitor MX106 and found that MX106 effectively overcame chemoresistance in vitro. MX106 inhibited cell migration and invasion by attenuating the TGFβ pathway and inhibiting EMT in ovarian cancer cells. To evaluate the efficacy of MX106 in inhibiting ovarian tumor metastasis, we treated an orthotopic ovarian cancer mouse model with MX106, and found that MX106 efficiently inhibited primary tumor growth in ovaries and metastasis in multiple peritoneal organs as compared to vehicle-treated control mice. Our data demonstrate that inhibition of survivin using either genetic KO or a novel inhibitor MX106 suppresses primary ovarian tumor growth and metastasis, supporting that targeting survivin could be an effective therapeutic approach in ovarian cancer.
Keywords: BIRC5 (survivin), Lentiviral CRISPR/Cas9 nickasevector, ovarian tumor metastasis, epithelial to mesenchymal transition (EMT), orthotopic ovarian cancer mouse model, survivin inhibitor MX106
Introduction
Ovarian cancer has the highest mortality rate among women’s malignancies due to the lack of obvious symptoms at its early stages, and often tumors were detected at the late stages and tumors were disseminated in multiple peritoneal organs [1, 2]. The majority of ovarian cancer patients suffer relapse following chemotherapy [3, 4]. Ovarian cancer therapy is still a major challenge and little progress has been made in overall patient survival during last decades due to tumor metastasis and chemoresistance [5–7].
Epithelial to mesenchymal transition (EMT) is a biological process by which epithelial cells lose their cell polarity, and acquire migratory and invasive properties, and is associated with tumor metastasis and chemoresistance [8–10]. The EMT phenotypic switch is accompanied by downregulation of epithelial markers, such as E-cadherin or cytokeratin-7, and upregulation of mesenchymal markers, such as vimentin, snai2, and β-catenin [11]. Multiple signaling pathways, including TGFβ, WNT, Notch, ERK½ and NF-κB regulate EMT [11–16]. Several studies also showed that EMT contributes to ovarian tumor metastasis and chemotherapy drug resistance [10, 11, 17–19]. Inhibition of EMT is a strategy for cancer therapy. Several pharmacological inhibitors of TGFβ, EGFR or FAK induced EMT have been used in clinical trials, such as LY36497, ZD1839, GSK2256098 or PF-573228 [20].
Survivin (BIRC5) is the smallest member of the inhibitors of apoptosis protein (IAP) family, and found to be highly expressed in a variety of human cancers, including breast cancer and ovarian cancer, but low expression of survivin was observed in adult normal tissues [21–24]. Survivin expression is well-correlated with tumor metastasis and chemoresistance in several cancer types, such as melanoma, renal, prostate, breast [25–28], and ovarian [10, 29–32]. The significant difference in survivin expression between various types of tumors and normal tissues suggests that survivin may be important drug target for cancer treatment. We previously showed that survivin was highly expressed in ovarian cancer and correlated with overall patient survival. Survivin also promoted EMT in ovarian cancer cells by activating TGFβ pathway [24]. Knockout (KO) of survivin using CRSIPR/Cas9 nickase or inhibition of survivin expression using a small molecule inhibitor of survivin, YM155 leads to inhibition of EMT in ovarian cancer cells [24].
Several small molecule inhibitors of survivin have been developed and tested for efficacy in cancer cells. One of most studied survivin inhibitors, YM155, underwent clinical trials but failed to achieve adequate efficacy [33–35]. YM155 is a known substrate for the P-glycoprotein (Pgp) drug efflux pump, which may compromise its effect in Pgp-mediated multidrug resistance cancers [36]. YM155 also has a short half-life in vivo; hence, it requires continuous infusion [34]. Recent evidence suggests that YM155 is a DNA damage-inducing agent and that its inhibition of survivin could be secondary to this event [37]. We have recently discovered a highly selective survivin degrader, MX-106, which selectively degrades survivin via proteasome-mediated degradation [38–40]. MX-106 also effectively overcomes Pgp-mediated multidrug resistance [38–40]. In vivo studies indicate that MX-106 maintains its ability to induce survivin degradation and strongly suppresses melanoma tumor growth [38].
To further determine the role of survivin in ovarian cancer, in this study we tested the hypothesis that survivin contributes to primary ovarian tumor growth and metastasis in an orthotopic ovarian cancer mouse model by genetically knocking out survivin or pharmacologically inhibiting survivin with MX106. We demonstrate that both approaches significantly suppressed primary ovarian tumor growth and metastasis by inhibiting EMT through attenuating TGFβ pathway.
Materials and Methods
Cell culture.
Ovarian cancer cell lines, SKOV3 (HTB-77) and OVCAR3 (HTB-161) were obtained from ATCC and cultured as described previously [24]. Cell lines were authenticated using Short Tandem Repeat (STR) analysis by ATCC and tested negative for mycoplasma contamination using luciferase assay (Lonza, Allendale, NJ). Cells were frozen at early passages and used for less than 5 weeks in continuous culture.
Establishment of the paclitaxel resistant cell line OVCAR3/TxR: Paclitaxel-resistant OVCAR3/TxR cell line was established from the parental OVCAR3 cells by a stepwise increase of paclitaxel concentration. Paclitaxel was doubled each passage and concentration was increased from 20 nM to 320 nM. The resistant cell line was established once cells remained viable after 320 nM treatment during two and half month’s culture. These cells were cultured under the same conditions as the parental OVCAR3 cell line.
Lentiviral Vector Production
Survivin KO and control SKOV3 and OVCAR3 cells were generated using lentiviral CRISPR/Cas9 nickase as we described previously [24]. The lentiviral CRISPR/Cas9 nickase-mediated TGFβR2 gene editing vectors were constructed using the same method as we described previously [24] by annealing two gRNA oligonucleotide pairs, 5’TTCCAGAATAAAGTCATGGT and 5’TTCTCCAAAGTGCATTATGA to target exon4. Lentivirus was produced by packaging in 293FT cells as published previously[41].
SMAD dependent reporter gene luciferase assay.
The lentiviral vector pGF-SMAD2/¾-mCMV-Luciferase-EF1a-puro (System Biosciences, CA) containing SMAD2/¾ transcriptional response elements (TRE) was used to transduce survivin KO and control SKOV3 and OVCAR3 cells using a multiplicity of infection of 10. This same amount of virus was also used to transduce wild-type SKOV3 and OVCAR3 cells and then treated cells with 5 μM MX106 or vehicle for 4h. Survivin KO, control cells or MX106-treated cells, were treated with 6 ng/ml TGFβ for 12h to activate SMAD2/¾ pathway. Luciferase activity was measured and normalized by comparing to control or vehicle-treated groups.
MX106 compound production.
MX106 was synthesized and characterized as described previously[42].
Orthotopic ovarian cancer mouse model.
To track ovarian tumor growth and metastasis in vivo, wild-type, survivin KO and control SKOV3 cells were labeled with luciferase. To test the effect of survivin KO in ovarian cancer cells on ovarian tumor metastasis, 10 two-months-old immunocompromised NOD.Cg Prkdcscid Il2rgtm1Wjl/SzJ (NSG) female mice (#005557, the Jackson Laboratory) were randomized into two groups, 5×105 survivin KO and control SKOV3 cells were intrabursally injected into the mice. To test the efficacy of MX106, 10 NSG female mice were intrabursally injected with 5×105 wildtype SKOV3 cells and randomly divided into two groups after one week post-injection. One group of mice were treated with MX106 (20 mg/kg body weight) and another group of mice were treated with vehicle for 5 days a week through intraperitoneal injection for 4 more weeks. Tumor growth and metastasis were monitored by Xenogen bio-imaging system once a week. All mice from each group were sacrificed at 5 weeks after cell injection, and primary ovarian and metastatic tumors were collected and subjected to double-blind histopathological analysis. Animal work was performed in accordance with the protocol (#16–161) approved by the Institutional Animal Care and Use Committee of the University of Tennessee Health Science Center.
MTT assay.
OVCAR3 or OVCAR3/TxR cells (3,000/well) were plated into 96-well plates following treatment with different doses of MX106 or YM155 and cultured for 24h. Cell proliferation was determined by measuring the absorbance at 570 nm wavelength as described previously [24]
Cell migration assay.
The cell migration assay was performed using the transwell plates. Briefly, SKOV3 or OVCAR3 cells were treated with 5 μM MX106 or vehicle for 4 h and then 5 × 104 cells in 300 μl serum-free DMEM were added to the upper chamber. DMEM containing 10% FBS was added into the lower chamber and incubated for 8h. The migrated cells were counted as described previously [24].
Cell invasion assay.
SKOV3 and OVCAR3 cells were treated with 5 μM MX106 or vehicle for 4h and the invaded cells were stained and counted as described previously [24].
Immunofluorescent staining.
To detect survivin, vimentin, cytokeratin-7 and cleaved-caspase3 expression, the immunofluorescent staining was performed as described previously [24]. The primary antibodies include survivin (1:200 dilution, #2808S, Cell Signaling, Danvers, MA), vimentin (1:200 dilution, #5741S, Cell Signaling, Danvers, MA), cytokeratin-7 (1:200 dilution, ab181598, Abcam), cleaved-caspase3 (1:200 dilution, 9661S, Cell Signaling, Danvers, MA).
Cell apoptosis.
SKOV3 or OVCAR3 cancer cells were treated with MX106 at different doses (0, 1, 2, and 4 μM) for 24 h. Apoptosis was detected using a caspase3/7 assay kit (Promega, Madison, WI). Cell apoptosis was also examined in both SKOV3 and OVCAR3 cells treated with MX106 at different doses (0, 1, 2, and 4 μM) for 24 h using Western blot to detect cleaved PARP and active caspase 3.
Western blot.
Western blot was performed as described previously [24].The primary antibodies were purchased from Cell signaling including β-catenin(#8480S), snai2(#9585S), vimentin (#5741S), surviving (#2808S), XIAP(#14334S), cIAP1(#7065S), cIAP2(#3130S), livin (#5471S), cleaved-PARP(#5625S), cleaved-caspase3(#9661S), T-SMAD2/3 (#8685S), P-SMAD2 (#18338S). GAPDH was purchased from Sigma (#G9545) and cytokeratin-7 was purchased from Abcam (#ab181598).
Statistical analysis.
Statistical analysis was performed from at least two independent experiments in triplicate and presented as means ± SD using Student’s t-test. P < 0.05 was considered significant. All data from experiments were included in statistical analysis.
Results
Knockout of survivin using lentiviral CRISPR/Cas9 nickase vector suppressed primary ovarian tumor growth and metastasis by inhibiting EMT in an orthotopic ovarian mouse model.
We previously reported that the disruption of survivin inhibited EMT in ovarian cancer cells by attenuating the TGFβ pathway [24], suggesting that survivin may contribute to ovarian tumor metastasis. To test this hypothesis, we first established a stable KO of survivin in the SKOV3 cell line with lentiviral CRISPR/Cas9 nickase vector as described previously [24]. We then intrabursally injected 5×105 ovarian cancer SKOV3 KO and control cells into two-month-old immunodeficient NSG female mice. Tumor growth and metastasis were monitored weekly using live animal imaging. Mice xenografted with survivin KO cells showed inhibition of primary tumor growth in ovaries (Fig. 1A). After one month, all mice xenografted with survivin KO SKOV3 and control cells were sacrificed and tumors in ovaries were collected. Tumors were significantly smaller in mice injected with survivin KO cells compared with the control cells (Fig. 1B). We examined EMT markers and pSMAD2 expression in primary ovarian tumors using western blot, and KO of survivin downregulated the mesenchymal marker including β-catenin, snai2 (snail2), and vimentin and pSMAD2 expression, and upregulated the epithelial marker cytokeratin-7 and Ecadherin (Fig. 1C). Tumor sections of mouse ovary were immunostained with survivin, vimentin and cytokeratin-7 antibodies, and the results were consistent with the western blots (Fig. S1). We further examined multiple peritoneal organs and found that tumors mainly metastasized into the liver (Fig. 2A) and spleen (Fig. 2B) in mice injected with control cells. In contrast, mice injected with survivin KO SKOV3 cells did not display detectable metastatic tumors in those organs. Tumors were observed in livers of mice injected with control cells, but not with survivin KO cells as verified by H&E staining (Fig. 2C). Our results indicated that loss of survivin led to suppression of primary ovarian tumor growth and tumor metastasis by inhibiting EMT and attenuating TGFβ pathway in this orthotopic ovarian cancer mouse model.
Figure 1. Knockout of survivin inhibits primary ovarian tumor growth in an orthotopic ovarian cancer mouse model.

A. Live animal imaging of primary tumors in ovaries of mice at one month following intrabursally injecting SKOV3 survivin KO and control cells (***p<0.001, n=5). B. Tumors in ovaries of mice were dissected and imaged (*p<0.05, n=5). C. Western blot analysis and comparison of survivin, pSMAD2 and EMT markers in primary ovarian tumors from three different xenografted mice (*p<0.05, **p<0.01).
Figure 2. Knockout of survivin inhibits ovarian tumor metastasis in an orthotopic ovarian cancer mouse model.

A, B. Metastatic tumors in liver (A) and spleen (B) of xenografted mice were identified by live animal imaging. The maximal bioluminescence intensity was compared between control and survivin knockout mice (**p<0.01, ***p<0.001, n=5). C. H&E staining of metastatic tumors in livers of xenografted mice.
Selective survivin inhibitor MX106 overcomes chemoresistance in ovarian cancer cells.
Tumor metastasis and chemoresistance are major issues for cancer therapy. KO of survivin significantly inhibited tumor metastasis by suppressing EMT in our orthotopic mouse model. To test whether survivin inhibition also overcomes ovarian cancer chemoresistance, we established a paclitaxel-resistant ovarian cancer cell line OVCAR3/TxR by selection in the presence of increased concentrations of paclitaxel. Western blot analyses indicated that both survivin and Pgp (a well-known marker for paclitaxel resistance [43–45]) were significantly increased in OVCAR3/TxR as compared to parent OVCAR3 cells (Fig. 3A). To target survivin using a small molecule inhibitor, we have designed and developed a novel inhibitor MX106 to selectively degrade survivin [42]. MX106 treatment induced apoptosis in both OVCAR3 and OVCAR3/TxR cells, whereas YM155 only induced apoptosis in parent OVCAR3 cells as determined by detection of cleaved-PARP using Western blot (Fig. 3B). We further determined the effects of survivin inhibitors on cell proliferation by treating both parent and drug resistant OVCAR3/TxR cells for 24h with different doses of MX106 or YM155. We found that MX106 efficiently inhibits cell proliferation in both OVCAR3 and OVCAR3/TxR cells (Fig. 3C), whereas YM155 markedly inhibited the proliferation of OVCAR3 cells but not of OVCAR3/TxR cells (Fig. 3D).
Figure 3. MX106 overcomes ovarian tumor chemoresistance.

A. Western blot analysis and comparison of survivin and Pgp expression in parental OVCAR3 and OVCAR3/TxR cells (*p<0.05, **p<0.01). B. Apoptosis in OVCAR3 and OVCAR3/TxR cells was detected by examining for cleaved-PARP following different doses of YM155 and MX106 treatment for 24h, the expression of cleaved-PARP was compared between vehicle and different doses (*p<0.05, **p<0.01, ***p<0.001 ). One representative western blot was presented from three similar independent experiments. C, D. Cell proliferation was measured using MTT assay in parent OVCAR3 and OVCAR3/TxR cells following MX106 (C, p<0.05) and YM155 treatment (D, p<0.05).
MX106 inhibits EMT in ovarian cancer cells.
We previously reported that knockout of survivin using CRISPR/Cas9 nickase mediated editing resulted in the inhibition of EMT in ovarian cancer cells [24]. To test whether MX106 inhibits EMT in ovarian cancer cells, we examined EMT marker gene expression in both SKOV3 and OVCAR3 cells after treatment with different doses of MX106. As shown in Figure 4, MX106 inhibited survivin and mesenchymal markers and promoted epithelial marker in a dose-dependent manner. The epithelial cell marker cytokeratin-7 was upregulated and mesenchymal markers including vimentin, snai2 and β-catenin were downregulated in both SKOV3 and OVCAR3 cells following MX106 treatment for 24h (Fig. 4). To confirm the selective degradation of survivin as compared to other members of the IAP family, we also examined X-linked inhibitor of apoptosis protein (XIAP), cIAP1, cIAP2 and Livin expression using Western blot. MX106 did not alter expression of other members of IAP family in either SKOV3 or OVCAR3 cells (Fig. 4). Our data indicate that the MX106 inhibits EMT by selectively degrading survivin in ovarian cancer cells.
Figure 4. MX106 inhibits EMT in ovarian cancer cells.

A, B. Western blot analysis in both SKOV3 (A) and OVCAR3 (B) cells following treatment using different doses of MX106 for 24h, the expression of EMT markers and the other members of the inhibitor of apoptosis protein was compared between vehicle and different doses (*p<0.05, **p<0.01, ***p<0.001). One representative western blot was presented from three similar independent experiments.
MX106 inhibits ovarian cancer cell migration and invasion.
EMT promotes cell migration and invasion, and we found that MX106 inhibits EMT in ovarian cancer cells. We then examined whether MX106 inhibits ovarian cancer cell migration and invasion. SKOV3 and OVCAR3 cells were treated with 5μM MX106 for 4h and cell migration was assayed in transwell plates. MX106 significantly inhibited the migration in both SKOV3 and OVCAR3 cells following MX106 treatment (Fig. 5A) Cell invasion was also assessed using Matrigel-coated transwell plates. Following 5 μM MX106 treatment for 4h treatment in SKOV3 and OVCAR3 cells, MX106 was found to significantly inhibit invasion in both cell lines (Fig. 5B). To exclude the effect of apoptosis on cell migration and invasion, we treated both SKOV3 and OVCAR3 cells with 20 μM caspase inhibitor Z-VAD for 2h followed by 5 μM MX106 treatment for 4h and then performed cell migration and invasion [46]. Similarly, we found that MX106 inhibited cell migration and invasion independent of cell apoptosis (Fig. S2). To exclude cell toxicity induced by MX106, we also examined cell viability in the upper chambers of transwell following cell migration and invasion. Cells treated with 5 μM MX106 displayed similar viability to vehicle treated cells (Fig. S3, S4). Our results demonstrated that MX106 effectively inhibits ovarian cancer cell migration and invasion.
Figure 5. MX106 inhibits ovarian cell migration and invasion.

A. Cell migration in SKOV3 or OVCAR3 cells was examined using transwell plates following 5 μM MX106 treatment for 4h, and migrated cells were stained with crystal blue and counted (**P<0.01; ***P<0.001). B. Cell invasion in SKOV3 or OVCAR3 cells following 5 μM MX106 treatment was examined using Matrigel-coated plates, and invaded cells were stained with H.E. and counted (**P<0.01; ***P<0.001).
The association of survivin with the TGFβ pathway and MX106 attenuated TGFβ signaling pathway in ovarian cancer cells.
As showed in our previous study, KO of survivin attenuated the TGFβ signaling pathway in ovarian cancer cells [24]. To determine the correlation between survivin and this pathway, we knocked down (KD) of TGFβ receptor 2 (TGFβR2) using lentiviral CRISPR/Cas9 vector and then treated both control and TGFβR2 KD cells with 6 ng/ml TGFβ for 24h and examined survivin expression. Knockdown of TGFβR2 resulted in significant reduction of survivin in both SKOV3 and OVCAR3 cells, while TGFβ upregulated survivin expression (Fig. 6A). We examined the TGFβ pathway in both TGFβR2 KD and control SKOV3 and OVCAR3 cells by detecting p-SMAD2 and total SMAD2 expression following 6 ng/ml TGFβ treatment for different time points. We found that knockdown of TGFβR2 attenuated the TGFβ pathway (Fig. 6B). In addition to knocking down of TGFβR2 using lentiviral CRISPR/Cas9 nickase vector, we also inhibited TGFβR½ using its inhibitor SB431542 by pretreating wild-type SKOV3 and OVCAR3 cells with 20 μM SB431542 for 24h, followed by 6 ng/ml TGFβ treatment for 24h. The TGFβR½ inhibitor suppressed survivin expression, while TGFβ upregulated survivin expression in both SKOV3 and OVCAR3 cells (Fig. 6C). We reported previously that KO of survivin attenuated TGFβ pathway [24]. We further validated this finding by using luciferase reporter gene assay. KO of survivin significantly reduced SMAD2/¾ transcriptional activity in both ovarian cancer SKOV3 and OVCAR3 cells (Fig. S5A). We examined whether MX106 also attenuates TGFβ pathway by treating both SKOV3 and OVCAR3 cells with 5 μM MX106 and then treated with 6 ng/ml TGFβ for different time points. We also determined reporter gene luciferase activity following MX106 and TGFβ treatment, and found that MX106 significantly inhibited SMAD2/¾ transcriptional activity in both SKOV3 and OVCAR3 cells (Fig. S5B). In addition, we examined non-SMAD pathway by detecting phospho-ERK½ and AKT following 5 μM MX106 and 6 ng/ml TGFβ treatment in both SKOV3 and OVCAR3 cells (Fig. S6). We found that MX106 attenuated p-SMAD2 in both SKOV3 and OVCAR3 cells using western blot (Fig. 6D). Our data indicated that survivin associates with TGFβ pathway, and inhibition of survivin using MX106 attenuates SMAD dependent TGFβ pathway in ovarian cancer cells.
Figure 6. Association of survivin with the TGFβ pathway in ovarian cancer cells.

A. Western blot analysis and comparison of survivin in TGFβR2 KD and control SKOV3 and OVCAR3 cells following 6 ng/ml TGFβ treatment for 24h, respectively (*p<0.05, **p<0.01). B. Western blot analysis and comparison of pSMAD2 and total SMAD2 expression in TGFβR2 KD and control SKOV3 and OVCAR3 cells following 6 ng/ml TGFβ treatment at different time points, respectively (**P<0.01; ***P<0.001). C. Western blot analysis and comparison of survivin in SKOV3 and OVCAR3 cells following TGFβR inhibitor treatment for 24h and then treated with 6 ng/ml TGFβ for 24h, respectively (**P<0.01; ***P<0.001). D. Western blot analysis and comparison of phospho and total SMAD2 in SKOV3 and OVCAR3 cells following 5 μM MX106 treatment for 4h and then 6 ng/ml TGFβ at different time points, respectively (***P<0.001). One representative western blot was presented from three similar independent experiments.
MX106 suppresses tumor metastasis in an orthotopic ovarian cancer mouse model
Knockout of survivin inhibits primary tumor growth in ovaries and tumor metastasis in multiple peritoneal organs (Figs. 1, 2). To test whether MX106 suppresses ovarian tumor metastasis by inhibiting survivin, we examined the efficacy of MX106 using an orthotopic ovarian cancer mouse model by intrabursally injecting SKOV3-Luc2 cells into two-month-old immunodeficient NSG female mice and then treating the mice with MX106 daily through intraperitoneal injection for 4 weeks. Tumor growth and metastasis were monitored using live animal imaging, and ovarian tumor metastasis was observed approximately two weeks after cell injection. At 5 weeks following cell injection, tumors were dissected at necropsy. Primary tumors in ovaries were significantly reduced in MX106 treated mice compared to vehicle control (Figs. 7A and 7B). We also examined survivin, other members of IAP family, and EMT markers from primary ovarian tumors using Western blot. The epithelial markers cytokeratin-7 and Ecadherin were upregulated, while survivin, p-SMAD2 and mesenchymal markers including β-catenin, vimentin and snai2 were downregulated in mice treated with MX106 compared to vehicle treated mice. In contrast, expression of other members in IAP family were unaffected, indicating that MX106 inhibits EMT by selectively degrading survivin and attenuating the TGFβ pathway in vivo (Fig. 7C). To further assess the efficacy of MX106, we also examined its effect on apoptosis by detecting cleaved-caspase3 in ovarian tumors by immunostaining and western blotting. Strong staining for cleaved-caspase3 was observed in cell nuclei from ovarian tumor sections in MX106 treated mice but not in vehicle-treated mice (Fig. 8A). Similarly, cleaved-caspase3 expression was significantly induced in ovarian tumor following MX106 treatment compared to vehicle-treated mice (Fig. 8B). The ovarian tumor sections were also immunostained for the cell proliferation marker PCNA, survivin, vimentin and cytokeratin-7. MX106 inhibited EMT by upregulating cytokeratin7 and downregulating vimentin and survivin (Fig. S7). Tumors were found to metastasize into multiple peritoneal organs including the liver in vehicle-treated mice but not in MX106-treated mice as shown in H&E stained sections (Fig. 8C), which is consistent with what was observed in survivin KO mice (Fig. S1). Our data indicated that MX106 significantly suppresses primary ovarian tumor growth and metastasis by inhibiting EMT and attenuating TGFβ pathway in an orthotopic ovarian cancer mouse model.
Figure 7. MX106 inhibits primary ovarian tumor growth and EMT in an orthotopic ovarian cancer mouse model.

A. Live animal imaging of primary tumor in ovaries at one month following intrabursal injection of wildtype SKOV3 cells and MX106 (20 mg/kg body weight) or vehicle treatment daily. B. Ovarian tumors were dissected and imaged. C. Western blot analysis and comparison of survivin, pSMAD2 and EMT markers in primary ovarian tumors from three different xenografted mice (*p<0.05, **p<0.01).
Fig.8. inhibits ovarian tumor metastasis and induced cell apoptosis in an orthotopic ovarian cancer mouse model.
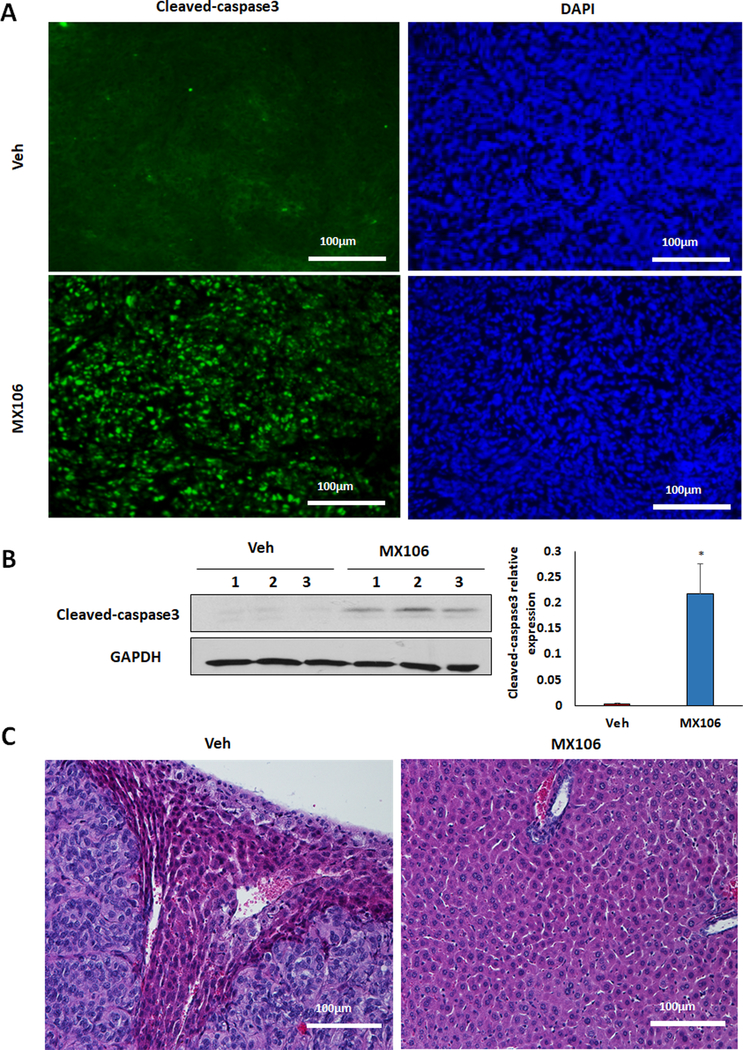
A. Ovarian tumor sections were immunostained using cleaved-caspase3 antibody following MX106 or vehicle treatment. B. Cell apoptosis in ovaries was detected and compared by examining cleaved-caspase3 using Western blot in primary ovarian tumors from three different xenografted mice (*p<0.05). C. H&E stained metastatic ovarian tumor in liver following MX106 or vehicle treatment.
Discussion
Based on the expression of survivin in ovarian cancer, survivin is a potential biomarker for diagnosis and prognosis in ovarian cancer, and also a potential drug target for therapy. In particular, we reported previously that survivin promoted EMT in ovarian cancer cells [24], suggesting a role in ovarian tumor metastasis and chemoresistance. In this study we have demonstrated that knockout of survivin using lentiviral CRISPR/Cas9 nickase vector suppresses primary tumor growth in ovaries as well as peritoneal metastasis in an orthotopic ovarian cancer mouse model xenografted with survivin KO and control SKOV3 cell line (Figs. 1 and 2). Although SKOV3 cell line is not a high grade serous carcinoma, [47] it is still the most frequently used ovarian carcinoma cell line due to its aggressive properties in inducing ovarian tumor metastasis in mouse models[48, 49] In addition, survivin expression is higher than that in OVCAR3 cells. Therefore, we selected SKOV3 cell line to investigate ovarian tumor metastasis in this study. Our studies using this in vivo orthotopic ovarian cancer mouse model indicate that survivin indeed contributes to ovarian tumor metastasis by promoting EMT in vivo. Furthermore, mice injected with survivin KO cells displayed reduced p-SMAD2 in ovarian tumors as compared to control mice. For the first time we showed that loss of survivin inhibited primary ovarian tumor growth and metastasis by attenuating the TGFβ pathway. Our study provided experimental evidence that survivin is an EMT-associated drug target for ovarian cancer therapy.
Although survivin as a therapeutic target for cancer has been studied over a decade, only a few small molecule inhibitors of survivin have been developed. For example, YM155 was in clinical trials for several different cancers, but was found to be ineffective in lung cancer [50], lymphoma [51] and rectal cancer [52], and had only modest activity in castration-resistant taxane-pretreated prostate cancer [53]. YM155 was developed in screening for inhibitors of the promoter activity of survivin, but was found not to directly bind survivin. In contrast, MX106 was developed based on its direct interaction with survivin, and selectively targets and degrades survivin. We found that MX106 inhibited ovarian cancer cell migration and invasion. Moreover, MX106 overcame chemoresistance in OVCAR3/TxR paclitaxel-resistant ovarian cancer cells, and induced ovarian cancer cell apoptosis and inhibited cell proliferation. In contrast, YM155 had no effect in OVCAR3/TxR drug resistant ovarian cancer cells (Fig. 3). However, YM155 increased the toxicity of chemotherapy drugs if Pgp transport was inhibited in glioma [54]. We further tested the efficacy of MX106 using orthotopic ovarian cancer mouse models and MX106 effectively inhibited primary tumor growth in ovaries and metastasis in peritoneal organs (Fig. 8). We found that MX106 mimics the phenotype of survivin KO in ovarian cancer cells and orthotopic ovarian cancer mouse models by degrading survivin and inhibiting EMT (Fig.4). Taken together, our data indicate that MX106 functionally inhibits ovarian tumor metastasis and overcomes chemoresistance by reversing EMT.
Our previous and current studies indicate that knockout or inhibition of survivin suppresses EMT in ovarian cancer cells in vitro or orthotopic ovarian cancer mouse model in vivo. Although multiple signaling pathways are involved in EMT regulation, we identified that survivin regulates EMT through its association with the TGFβ pathway in ovarian cancer cells. Genetic KO of survivin or pharmacological inhibition of survivin attenuates TGFβ pathway as evidenced by reduced p-SMAD2 in ovarian cancer cells in vitro and orthotopic ovarian cancer mouse model in vivo. Our previous work showed that TGFβ promotes EMT in ovarian cancer cells [55]. Therefore, we hypothesized that survivin promotes EMT by activating the TGFβ pathway. However, it is unclear how survivin affects the TGFβ pathway in regulating EMT in ovarian cancer cells. XIAP, a member of the IAP family, directly interacts with TGFβ receptor1 (TGFβR1) through its baculovirus IAP repeat (BIR) domain [56]. It is possible that survivin as another IAP family member interacts with TGFβR½ through its BIR domain, thus regulating TGFβ pathway in ovarian cancer cells.
To define the interaction between survivin and TGFβ pathway, we knocked down TGFβR2 in ovarian cancer SKOV3 and OVCAR3 cells. Survivin levels were significantly reduced in TGFβR2 KD cells as compared to control cells, indicating the interplay between survivin and TGFβ pathway in ovarian cancer cells. It was reported previously that SMAD¾ promoted survivin expression through transcriptional regulation [57]. Therefore, it is possible that survivin may interact with SMAD¾ complex to form a positive feedback loop, thus regulating TGFβ pathway in ovarian cancer cells. However, further investigation is required to determine whether the BIR domain of survivin or interaction with SMAD¾ complex is responsible for this interaction in ovarian cancer cells.
In conclusion, we demonstrated that survivin contributes to primary ovarian tumor growth and metastasis and chemoresistance by promoting EMT through activation of the TGFβ pathway in ovarian cancer. Survivin is a therapeutic drug target for ovarian cancer. MX106 as a selective degrader of survivin inhibited survivin expression but had no effect on other members of IAP family. Compared with YM155, MX106 overcomes chemoresistance and suppresses primary ovarian tumor growth and metastasis by inhibiting EMT through attenuating the TGFβ pathway, which provides a new approach for ovarian cancer therapy by targeting survivin.
Supplementary Material
Acknowledgments
Financial Support:
This study was partially supported by grants from NIH R01CA193609–01A1 (W. Li) and NIH R21 CA216585–01A1 (J. Yue), National Natural Science Foundation of China 81572574(Y. Guo),Natural Science Foundation of Henan Province (182300410311(Y. Guo),Foundation of Henan Educational Committee 162102410011(YG). The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Footnotes
Disclosure of Potential Conflicts of Interest: The authors declare no competing interests
References:
- 1.Siegel RL, Miller KD, Jemal A. Cancer Statistics, 2017. CA Cancer J Clin. 2017; 67: 7–30. [DOI] [PubMed] [Google Scholar]
- 2.Gloss BS, Samimi G. Epigenetic biomarkers in epithelial ovarian cancer. Cancer Lett. 2014; 342: 257–63. [DOI] [PubMed] [Google Scholar]
- 3.Jayson GC, Kohn EC, Kitchener HC, Ledermann JA. Ovarian cancer. Lancet. 2014; 384: 1376–88. [DOI] [PubMed] [Google Scholar]
- 4.Roy L, Cowden Dahl KD. Can Stemness and Chemoresistance Be Therapeutically Targeted via Signaling Pathways in Ovarian Cancer? Cancers (Basel). 2018; 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Scalici JM, Arapovic S, Saks EJ, Atkins KA, Petroni G, Duska LR, et al. Mesothelium expression of vascular cell adhesion molecule-1 (VCAM-1) is associated with an unfavorable prognosis in epithelial ovarian cancer (EOC). Cancer. 2017; 123: 977–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ahmed N, Stenvers KL. Getting to know ovarian cancer ascites: opportunities for targeted therapy-based translational research. Front Oncol. 2013; 3: 256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Nagai T, Oshiro H, Sagawa Y, Sakamaki K, Terauchi F, Nagao T. Pathological Characterization of Ovarian Cancer Patients Who Underwent Debulking Surgery in Combination With Diaphragmatic Surgery A Cross-Sectional Study. Medicine (Baltimore). 2015; 94: e2296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zhou XM, Zhang H, Han X. Role of epithelial to mesenchymal transition proteins in gynecological cancers: pathological and therapeutic perspectives. Tumour Biol. 2014; 35: 9523–30. [DOI] [PubMed] [Google Scholar]
- 9.Deng J, Wang L, Chen H, Hao J, Ni J, Chang L, et al. Targeting epithelial-mesenchymal transition and cancer stem cells for chemoresistant ovarian cancer. Oncotarget. 2016; 7: 55771–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ahmed N, Abubaker K, Findlay J, Quinn M. Epithelial mesenchymal transition and cancer stem cell-like phenotypes facilitate chemoresistance in recurrent ovarian cancer. Curr Cancer Drug Targets. 2010; 10: 268–78. [DOI] [PubMed] [Google Scholar]
- 11.Yoshida S, Furukawa N, Haruta S, Tanase Y, Kanayama S, Noguchi T, et al. Expression profiles of genes involved in poor prognosis of epithelial ovarian carcinoma: a review. Int J Gynecol Cancer. 2009; 19: 992–7. [DOI] [PubMed] [Google Scholar]
- 12.Arend RC, Londono-Joshi AI, Straughn JM Jr., Buchsbaum DJ. The Wnt/beta-catenin pathway in ovarian cancer: a review. Gynecol Oncol. 2013; 131: 772–9. [DOI] [PubMed] [Google Scholar]
- 13.Chou JL, Chen LY, Lai HC, Chan MW. TGF-beta: friend or foe? The role of TGF-beta/SMAD signaling in epigenetic silencing of ovarian cancer and its implication in epigenetic therapy. Expert Opin Ther Targets. 2010; 14: 1213–23. [DOI] [PubMed] [Google Scholar]
- 14.Wu J, Liu Z, Shao C, Gong Y, Hernando E, Lee P, et al. HMGA2 overexpression-induced ovarian surface epithelial transformation is mediated through regulation of EMT genes. Cancer Res. 2011; 71: 349–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Yang J, Guo W, Wang L, Yu L, Mei H, Fang S, et al. Notch signaling is important for epithelial-mesenchymal transition induced by low concentrations of doxorubicin in osteosarcoma cell lines. Oncol Lett. 2017; 13: 2260–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Natsuizaka M, Whelan KA, Kagawa S, Tanaka K, Giroux V, Chandramouleeswaran PM, et al. Interplay between Notch1 and Notch3 promotes EMT and tumor initiation in squamous cell carcinoma. Nat Commun. 2017; 8: 1758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Amankwah EK, Lin HY, Tyrer JP, Lawrenson K, Dennis J, Chornokur G, et al. Epithelial-Mesenchymal Transition (EMT) Gene Variants and Epithelial Ovarian Cancer (EOC) Risk. Genet Epidemiol. 2015; 39: 689–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Helleman J, Smid M, Jansen MP, van der Burg ME, Berns EM. Pathway analysis of gene lists associated with platinum-based chemotherapy resistance in ovarian cancer: the big picture. Gynecol Oncol. 2010; 117: 170–6. [DOI] [PubMed] [Google Scholar]
- 19.Takai M, Terai Y, Kawaguchi H, Ashihara K, Fujiwara S, Tanaka T, et al. The EMT (epithelial-mesenchymal-transition)-related protein expression indicates the metastatic status and prognosis in patients with ovarian cancer. J Ovarian Res. 2014; 7: 76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Davis FM, Stewart TA, Thompson EW, Monteith GR. Targeting EMT in cancer: opportunities for pharmacological intervention. Trends Pharmacol Sci. 2014; 35: 479–88. [DOI] [PubMed] [Google Scholar]
- 21.Liguang Z, Peishu L, Hongluan M, Hong J, Rong W, Wachtel MS, et al. Survivin expression in ovarian cancer. Exp Oncol. 2007; 29: 121–5. [PubMed] [Google Scholar]
- 22.Li XJ, Pang JS, Li YM, Ahmed FA, He RQ, Ma J, et al. Clinical value of survivin and its underlying mechanism in ovarian cancer: A bioinformatics study based on GEO and TCGA data mining. Pathol Res Pract. 2018; 214: 385–401. [DOI] [PubMed] [Google Scholar]
- 23.Gunaldi M, Isiksacan N, Kocoglu H, Okuturlar Y, Gunaldi O, Topcu TO, et al. The value of serum survivin level in early diagnosis of cancer. J Cancer Res Ther. 2018; 14: 570–3. [DOI] [PubMed] [Google Scholar]
- 24.Zhao G, Wang Q, Gu Q, Qiang W, Wei JJ, Dong P, et al. Lentiviral CRISPR/Cas9 nickase vector mediated BIRC5 editing inhibits epithelial to mesenchymal transition in ovarian cancer cells. Oncotarget. 2017; 8: 94666–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Chen L, Liang L, Yan X, Liu N, Gong L, Pan S, et al. Survivin status affects prognosis and chemosensitivity in epithelial ovarian cancer. Int J Gynecol Cancer. 2013; 23: 256–63. [DOI] [PubMed] [Google Scholar]
- 26.Parvani JG, Davuluri G, Wendt MK, Espinosa C, Tian M, Danielpour D, et al. Deptor enhances triple-negative breast cancer metastasis and chemoresistance through coupling to survivin expression. Neoplasia. 2015; 17: 317–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Nestal de Moraes G, Delbue D, Silva KL, Robaina MC, Khongkow P, Gomes AR, et al. FOXM1 targets XIAP and Survivin to modulate breast cancer survival and chemoresistance. Cell Signal. 2015; 27: 2496–505. [DOI] [PubMed] [Google Scholar]
- 28.Czyz M, Lesiak-Mieczkowska K, Koprowska K, Szulawska-Mroczek A, Wozniak M. Cell context-dependent activities of parthenolide in primary and metastatic melanoma cells. Br J Pharmacol. 2010; 160: 1144–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zhou L, Liu P, Chen B, Wang Y, Wang X, Chiriva Internati M, et al. Silibinin restores paclitaxel sensitivity to paclitaxel-resistant human ovarian carcinoma cells. Anticancer Res. 2008; 28: 1119–27. [PubMed] [Google Scholar]
- 30.Jiang L, Luo RY, Yang J, Cheng YX. Knockdown of survivin contributes to antitumor activity in cisplatin-resistant ovarian cancer cells. Mol Med Rep. 2013; 7: 425–30. [DOI] [PubMed] [Google Scholar]
- 31.Lili LN, Matyunina LV, Walker LD, Wells SL, Benigno BB, McDonald JF. Molecular profiling supports the role of epithelial-to-mesenchymal transition (EMT) in ovarian cancer metastasis. J Ovarian Res. 2013; 6: 49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yan H, Sun Y. Evaluation of the mechanism of epithelial-mesenchymal transition in human ovarian cancer stem cells transfected with a WW domain-containing oxidoreductase gene. Oncol Lett. 2014; 8: 426–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Satoh T, Okamoto I, Miyazaki M, Morinaga R, Tsuya A, Hasegawa Y, et al. Phase I study of YM155, a novel survivin suppressant, in patients with advanced solid tumors. Clin Cancer Res. 2009; 15: 3872–80. [DOI] [PubMed] [Google Scholar]
- 34.Nakahara T, Kita A, Yamanaka K, Mori M, Amino N, Takeuchi M, et al. YM155, a novel small-molecule survivin suppressant, induces regression of established human hormone-refractory prostate tumor xenografts. Cancer Res. 2007; 67: 8014–21. [DOI] [PubMed] [Google Scholar]
- 35.Giaccone G, Zatloukal P, Roubec J, Floor K, Musil J, Kuta M, et al. Multicenter phase II trial of YM155, a small-molecule suppressor of survivin, in patients with advanced, refractory, non-small-cell lung cancer. J Clin Oncol. 2009; 27: 4481–6. [DOI] [PubMed] [Google Scholar]
- 36.Iwai M, Minematsu T, Li Q, Iwatsubo T, Usui T. Utility of P-glycoprotein and organic cation transporter 1 double-transfected LLC-PK1 cells for studying the interaction of YM155 monobromide, novel small-molecule survivin suppressant, with P-glycoprotein. Drug Metab Dispos. 2011; 39: 2314–20. [DOI] [PubMed] [Google Scholar]
- 37.Glaros TG, Stockwin LH, Mullendore ME, Smith B, Morrison BL, Newton DL. The “survivin suppressants” NSC 80467 and YM155 induce a DNA damage response. Cancer Chemother Pharmacol. 2012; 70: 207–12. [DOI] [PubMed] [Google Scholar]
- 38.Xiao M, Wang J, Lin Z, Lu Y, Li Z, White SW, et al. Design, Synthesis and Structure-Activity Relationship Studies of Novel Survivin Inhibitors with Potent Anti-Proliferative Properties. PLoS One. 2015; 10: e0129807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ning Z, Wang A, Liang J, Xie Y, Liu J, Yan Q, et al. USP22 promotes epithelial-mesenchymal transition via the FAK pathway in pancreatic cancer cells. Oncol Rep. 2014; 32: 1451–8. [DOI] [PubMed] [Google Scholar]
- 40.Qi J, Dong Z, Liu J, Peery RC, Zhang S, Liu JY, et al. Effective Targeting of the Survivin Dimerization Interface with Small-Molecule Inhibitors. Cancer Res. 2016; 76: 453–62. [DOI] [PubMed] [Google Scholar]
- 41.Yue J, Sheng Y, Ren A, Penmatsa S. A miR-21 hairpin structure-based gene knockdown vector. Biochem Biophys Res Commun. 2010; 394: 667–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Xiao M, Xue Y, Wu Z, Lei ZN, Wang J, Chen ZS, et al. Design, synthesis and biological evaluation of selective survivin inhibitors. J Biomed Res. 2019; 33: 82–100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Bradley G, Ling V. P-glycoprotein, multidrug resistance and tumor progression. Cancer Metastasis Rev. 1994; 13: 223–33. [DOI] [PubMed] [Google Scholar]
- 44.Li L, Jiang AC, Dong P, Wan Y, Yu ZW. The characteristics of Hep-2 cell with multiple drug resistance induced by Taxol. Otolaryngol Head Neck Surg. 2007; 137: 659–64. [DOI] [PubMed] [Google Scholar]
- 45.Mechetner E, Kyshtoobayeva A, Zonis S, Kim H, Stroup R, Garcia R, et al. Levels of multidrug resistance (MDR1) P-glycoprotein expression by human breast cancer correlate with in vitro resistance to taxol and doxorubicin. Clin Cancer Res. 1998; 4: 389–98. [PubMed] [Google Scholar]
- 46.Kashyap VK, Wang Q, Setua S, Nagesh PKB, Chauhan N, Kumari S, et al. Therapeutic efficacy of a novel betaIII/betaIV-tubulin inhibitor (VERU-111) in pancreatic cancer. J Exp Clin Cancer Res. 2019; 38: 29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Domcke S, Sinha R, Levine DA, Sander C, Schultz N. Evaluating cell lines as tumour models by comparison of genomic profiles. Nat Commun. 2013; 4: 2126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Xu X, Ayub B, Liu Z, Serna VA, Qiang W, Liu Y, et al. Anti-miR182 reduces ovarian cancer burden, invasion, and metastasis: an in vivo study in orthotopic xenografts of nude mice. Mol Cancer Ther. 2014; 13: 1729–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Peng JM, Chen YH, Hung SW, Chiu CF, Ho MY, Lee YJ, et al. Recombinant viral protein promotes apoptosis and suppresses invasion of ovarian adenocarcinoma cells by targeting alpha5beta1 integrin to down-regulate Akt and MMP-2. Br J Pharmacol. 2012; 165: 479–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kelly RJ, Thomas A, Rajan A, Chun G, Lopez-Chavez A, Szabo E, et al. A phase I/II study of sepantronium bromide (YM155, survivin suppressor) with paclitaxel and carboplatin in patients with advanced non-small-cell lung cancer. Ann Oncol. 2013; 24: 2601–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Cheson BD, Bartlett NL, Vose JM, Lopez-Hernandez A, Seiz AL, Keating AT, et al. A phase II study of the survivin suppressant YM155 in patients with refractory diffuse large B-cell lymphoma. Cancer. 2012; 118: 3128–34. [DOI] [PubMed] [Google Scholar]
- 52.Sprenger T, Rodel F, Beissbarth T, Conradi LC, Rothe H, Homayounfar K, et al. Failure of downregulation of survivin following neoadjuvant radiochemotherapy in rectal cancer is associated with distant metastases and shortened survival. Clin Cancer Res. 2011; 17: 1623–31. [DOI] [PubMed] [Google Scholar]
- 53.Tolcher AW, Quinn DI, Ferrari A, Ahmann F, Giaccone G, Drake T, et al. A phase II study of YM155, a novel small-molecule suppressor of survivin, in castration-resistant taxane-pretreated prostate cancer. Ann Oncol. 2012; 23: 968–73. [DOI] [PubMed] [Google Scholar]
- 54.Radic-Sarikas B, Halasz M, Huber KVM, Winter GE, Tsafou KP, Papamarkou T, et al. Lapatinib potentiates cytotoxicity of YM155 in neuroblastoma via inhibition of the ABCB1 efflux transporter. Sci Rep. 2017; 7: 3091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Chen Z, Wang Y, Liu W, Zhao G, Lee S, Balogh A, et al. Doxycycline inducible Kruppel-like factor 4 lentiviral vector mediates mesenchymal to epithelial transition in ovarian cancer cells. PLoS One. 2014; 9: e105331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Neil JR, Tian M, Schiemann WP. X-linked inhibitor of apoptosis protein and its E3 ligase activity promote transforming growth factor-{beta}-mediated nuclear factor-{kappa}B activation during breast cancer progression. J Biol Chem. 2009; 284: 21209–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Yang J, Song K, Krebs TL, Jackson MW, Danielpour D. Rb/E2F4 and Smad2/3 link survivin to TGF-beta-induced apoptosis and tumor progression. Oncogene. 2008; 27: 5326–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.