

The International Ocean Discovery Program Expedition 357—“Serpentinization and Life”—utilized seabed drills to collect rocks from the oceanic crust. The recovered rock cores represent the shallow serpentinite subsurface of the Atlantis Massif, where reactions between uplifted mantle rocks and water, collectively known as serpentinization, produce environmental conditions that can stimulate biological activity and are thought to be analogous to environments that were prevalent on the early Earth and perhaps other planets. The methodology and results of this project have implications for life detection experiments, including sample return missions, and provide a window into the diversity of microbial communities inhabiting subseafloor serpentinites.
KEYWORDS: Atlantis Massif, contamination, serpentinization
ABSTRACT
The Atlantis Massif rises 4,000 m above the seafloor near the Mid-Atlantic Ridge and consists of rocks uplifted from Earth’s lower crust and upper mantle. Exposure of the mantle rocks to seawater leads to their alteration into serpentinites. These aqueous geochemical reactions, collectively known as the process of serpentinization, are exothermic and are associated with the release of hydrogen gas (H2), methane (CH4), and small organic molecules. The biological consequences of this flux of energy and organic compounds from the Atlantis Massif were explored by International Ocean Discovery Program (IODP) Expedition 357, which used seabed drills to collect continuous sequences of shallow (<16 m below seafloor) marine serpentinites and mafic assemblages. Here, we report the census of microbial diversity in samples of the drill cores, as measured by environmental 16S rRNA gene amplicon sequencing. The problem of contamination of subsurface samples was a primary concern during all stages of this project, starting from the initial study design, continuing to the collection of samples from the seafloor, handling the samples shipboard and in the lab, preparing the samples for DNA extraction, and analyzing the DNA sequence data. To distinguish endemic microbial taxa of serpentinite subsurface rocks from seawater residents and other potential contaminants, the distributions of individual 16S rRNA gene sequences among all samples were evaluated, taking into consideration both presence/absence and relative abundances. Our results highlight a few candidate residents of the shallow serpentinite subsurface, including uncultured representatives of the Thermoplasmata, Acidobacteria, Acidimicrobia, and Chloroflexi.
IMPORTANCE The International Ocean Discovery Program Expedition 357—“Serpentinization and Life”—utilized seabed drills to collect rocks from the oceanic crust. The recovered rock cores represent the shallow serpentinite subsurface of the Atlantis Massif, where reactions between uplifted mantle rocks and water, collectively known as serpentinization, produce environmental conditions that can stimulate biological activity and are thought to be analogous to environments that were prevalent on the early Earth and perhaps other planets. The methodology and results of this project have implications for life detection experiments, including sample return missions, and provide a window into the diversity of microbial communities inhabiting subseafloor serpentinites.
INTRODUCTION
Subsurface environments may host the majority of microbial life on Earth (1, 2), but current estimates of subsurface biomass lack certainty due to a paucity of observations. Recent studies, including major ocean drilling expeditions, have greatly improved the quantification and characterization of microbial populations in deep marine sediments (3–6). Seafloor crustal rocks are also expected to be an important microbial habitat, in part because a significant fraction of the oceanic crust supports hydrothermal circulation of seawater that can bring nutrients and energy into subseafloor ecosystems (7–9). The mineral composition and geological history of the host rocks determine the local environmental conditions experienced by subseafloor microbes (10–13). Ultramafic habitats (high-iron, low-silica rocks derived from the mantle) for life may have unique attributes, since these rock types are unstable in the presence of seawater and undergo a series of geochemical reactions that can support chemolithotrophy (14–16). A few microbial diversity surveys of mafic basaltic and gabbroic seafloor rocks have been conducted (17–19), but no studies have characterized the microbial diversity of ultramafic rocks.
International Ocean Discovery Program (IODP) Expedition 357 addressed this knowledge gap by targeting the Atlantis Massif, a submarine mountain located in the north Atlantic Ocean on the western flank of the Mid-Atlantic Ridge (20, 21). The massif is approximately 16 km across and rises 4,267 m from the seafloor. It is composed of variable amounts of ultramafic rocks uplifted from the upper mantle along a major fault and altered into serpentinites through the geochemical process of serpentinization (22, 23). Oxidation of iron minerals in the serpentinizing rocks results in the release of hydrogen gas and hydroxyl ions, contributing to the formation of extremely reducing, high-pH fluids (24, 25). The high hydrogen concentrations that develop during serpentinization are conducive to the formation of methane and other organic compounds from the reduction of inorganic carbon (26–28). Circulation of organic-rich fluids through the serpentinites of the Atlantis Massif may support an active subseafloor ecosystem (29).
Much of our current knowledge of the biological implications of marine serpentinization (30) comes from the exploration of hydrothermal springs such as the “Lost City,” a collection of carbonate chimneys near the summit of the Atlantis Massif where high pH fluids exit the seafloor (22, 25, 31, 32). The chimneys are covered in mucilaginous biofilms formed by bacteria and archaea that are fueled by the high concentrations of hydrogen, formate, and methane in the venting fluids (33–36). The high density of the microbial biofilm communities is likely enabled by the mixing of warm, anoxic hydrothermal fluids with oxygenated ambient seawater, enabling a wide range of metabolic strategies (37). In contrast, the much more voluminous, rocky subsurface habitats underlying the Lost City chimneys and throughout the Atlantis Massif probably experience much more diffuse hydrothermal circulation and an extended alteration. Our knowledge of this subseafloor environment, however, is extremely limited (19).
The work reported here focused on two fundamental questions regarding serpentinization and life: How can we distinguish endemic microbial communities of low biomass subseafloor rocks from other environments? Which microbes inhabit marine serpentinites? To address these questions, we conducted a cultivation-independent census of microbial diversity in the shallow (<16 m below seafloor [mbsf]) rocks collected during IODP Expedition 357 (21) and compared these to the communities found in seawater and potential sources of contamination.
Laboratory contamination of DNA sequencing data sets is a well-known problem (38–41), especially for low-biomass subsurface samples (19, 42–44), that must be addressed by a combination of experimental and computational methods. No matter how cleanly DNA is prepared, contaminant sequences can appear for a myriad of reasons (39, 40). Sheik et al. (41) recently reviewed practices for identifying contaminants in DNA sequence data sets, highlighting the importance of collecting and sequencing control samples representing potential sources of environmental and laboratory contamination. These studies have highlighted the need to complement careful laboratory protocols with computational methods that can distinguish true residents from multiple potential environmental and laboratory contaminants (45–47).
Here, we report our methodological and computational strategies for minimizing and identifying contaminants in DNA sequencing data sets from extremely low-biomass seafloor serpentinites collected during IODP Expedition 357. Sequencing DNA from serpentinites required the development of a novel DNA extraction and purification protocol that recovers highly pure DNA without the use of any commercial kits or phenol. We then identified environmental and laboratory contaminants in the resulting DNA sequencing data with a computational workflow that considers the relative abundances of individual sequences among potential contamination sources. Finally, we describe the archaeal and bacterial taxa that are potential residents of seafloor serpentinites.
RESULTS
DNA yields from rocks.
DNA was extracted from at least one rock core sample collected from all sites (Fig. 1; see also Text S1 in the supplemental material) except M0073 (Table 1; see also Data Set S1 in the supplemental material). DNA yields from lysates (postlysis, prepurification) ranged from ∼100 to almost 500 ng of DNA per g of rock sample. DNA yields from serpentinite and nonserpentinite rocks (including metagabbro, carbonate sand, basalt breccia, and metadolerite) were similar (Table 1; see also Data Set S2). These values may be inflated by the many impurities in the lysates, which had not been purified and were visibly cloudy. No trends in DNA yield with respect to mineralogy or locations of the boreholes were apparent. DNA was quantified after lysis, washing, and the final purification (Table 1). DNA was below the detection limit for all final, purified preparations, indicating a substantial loss of DNA from the lysates.
FIG 1.
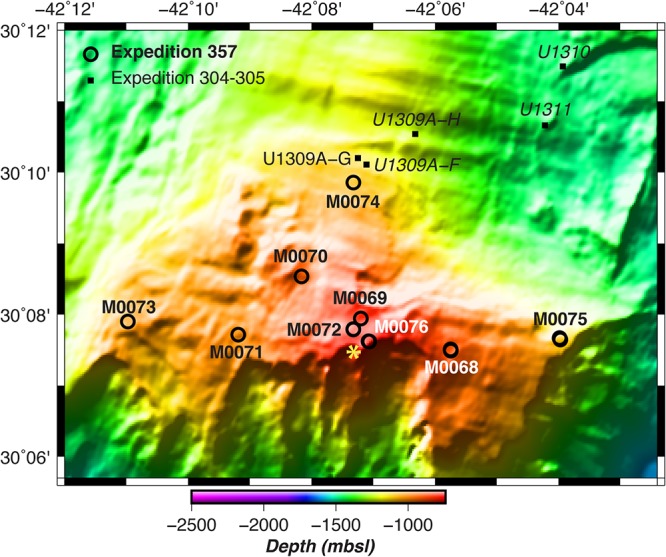
Multibeam bathymetry of the Atlantis Massif, with water depth in meters below sea level (mbsl) in color scale per legend. Drilled sites are shown as black circles. The yellow star is the location of the Lost City hydrothermal field. The distance between the most western site (M0073) and the most eastern site (M0075) is approximately 13 km. (Adapted from reference 20 with permission of the publisher.)
TABLE 1.
DNA yield from each IODP Expedition 357 serpentinite sample after lysis, washing, and purification stepsa
| Sample ID | IODP ID | Extracted rock (g) | Amt of DNA (ng/g of rock) |
No. of sequences |
|||
|---|---|---|---|---|---|---|---|
| Lysate | Postwashing | Purified | Total | After quality filtering | |||
| 0AMRd005A | 357-68B-8R-1,29-34cm | 30 | 325 | 4.07 | <0.46 | 2,985 | 1,318 |
| 0AMRd014A | 357-69A-7R-1,73-75cm | 40.5 | 104 | 2.51 | <0.55 | 2,928 | 1,926 |
| 0AMRd030A | 357-71B-2R-2,53-58cm | 37 | 224 | 2.34 | <0.29 | 3,893 | 2,957 |
| 0AMRd031A | 357-71B-2R-1,64-66cm | 37.5 | 215 | 2.32 | <0.49 | 2,780 | 2,210 |
| 0AMRd033A | 357-71C-2R-1,78-84cm | 39 | 371 | 4.99 | <0.2 | 3,920 | 3,291 |
| 0AMRd034B | 357-71C-2R-1,84-90cm | 39 | 213 | 2.97 | <0.19 | 2,901 | 2,294 |
| 0AMRd036A | 357-71C-5R-CC,5-10cm | 38 | 248 | 2.03 | <0.21 | 4,281 | 3,442 |
| 0AMRd045A | 357-72B-3R-1,34-36cm | 38 | 473 | 15.2 | <0.2 | 10,787 | 7,225 |
| 0AMRd057A | 357-75A-1R-CC,0-4cm | 26 | 136 | 2 | <0.16 | 153,054 | 113,994 |
| 0AMRd067A | 357-76B-5R-1,14-28cm | 37 | 299 | 4.39 | <0.22 | 4,699 | 2,831 |
| 0AMRd071A | 357-76B-7R-1,105-120cm | 34.5 | 100 | 7.14 | <0.41 | 10,095 | 7,742 |
| 0AMRd072A | 357-76B-9R-1,34-41cm | 37 | 110 | 2.64 | <0.49 | 122,361 | 82,246 |
| 0AMRd073A | 357-76B-9R-1,43-52cm | 47.4 | 149 | 4.71 | <0.82 | 5,189 | 2,079 |
| 0AMRd075A | 357-76B-10R-1,91-111cm | 38 | 300 | 1.49 | <0.52 | 12,082 | 9,758 |
| 0AMRd076A | 357-76B-10R-1,91-111cm | 33 | 223 | 4.3 | <1.24 | 9,399 | 7,478 |
The concentration of the final, purified DNA was below the stated detection limit for each sample. The numbers of 16S rRNA gene amplicon sequences are reported before and after quality filtering.
DNA sequencing results.
Despite the below-detection levels of purified DNA, a total of 1,125,191 sequencing read pairs of the 16S rRNA gene were obtained from all rock samples (50 samples, including sequencing duplicates). Here, we focus only on the DNA sequencing results from the 15 samples characterized as serpentinites, but the DNA sequencing data from all samples are included in our NCBI Sequence Read Archive (SRI) BioProject PRJNA575221. At least 2,780 read pairs were obtained from each sample, and >10,000 read pairs were obtained from five of the serpentinite rock samples (Table 1). We identified 2,063 amplicon sequence variants (ASVs) among all rock samples, 664 of which were found only in serpentinites. We obtained 27,524,256 read pairs from the water samples (76 samples of surface, shallow, and deep including sequencing duplicates) (see Data Set S3), from which we identified 15,142 ASVs. Six samples of lab air yielded 66,846 read pairs and 293 ASVs (see Data Set S4), despite below-detection levels of purified DNA.
The proportional abundances of all 17,081 unique ASVs among all rock, water, and air samples were used to calculate the community dissimilarity between each pair of samples. The results are visualized in the nonmetric multidimensional scaling (NMDS) plot in Fig. 2, which shows a clear split between water and rock samples. The bacterial communities of both serpentinite (orange points) and nonserpentinite (yellow points) rock samples are highly variable and are not consistently distinct from each other. The bacterial compositions of lab air samples (green points) overlap those of the rock samples in this visualization.
FIG 2.

A nonmetric multidimensional scaling plot shows general patterns in the 16S rRNA gene microbial community compositions of all types of samples. Each data point represents the microbial community composition of a sample of seawater (deep, shallow, or surface), rock core (serpentinite versus nonserpentinite), or laboratory air. The distances between the data points represent the Morisita-Horn dissimilarities among the microbial communities.
Evaluation of contamination sources.
To evaluate contamination in the serpentinites, we used SourceTracker2 (47) to estimate the proportion of each rock sample’s DNA sequences that could be attributed to contamination from water or air. These results show that very few of the DNA sequences in the serpentinites can be traced back to seawater (Fig. 3). The number of sequences attributed to air was highly variable among the serpentinites, from nearly zero to >50% of total sequences. Most serpentinite samples were dominated by sequences that could not be attributed to a single source by SourceTracker2. These ASVs are designated as having an “unknown” source and may represent true inhabitants of the serpentinites or may be derived from another source that was not sampled in this study. Based on these results, serpentinite samples with >15% of sequences attributed to lab air were excluded from further analyses (0AMRd014A, 031C, 033A, 033C, 034A, 036A, 045B, 067C, 071A, and 073A).
FIG 3.

Assessment of potential 16S rRNA gene library contamination in serpentinite samples with SourceTracker2. Each bar on this plot shows the percentage of ASVs from each source of contamination into each serpentinite sample. Green, lab air; blue, seawater samples; gray, undetermined.
SourceTracker2 computes probabilistic estimates of how the whole-community composition of samples relate to each other, but it is not designed to identify the source of each taxon. For example, multiple ASVs in sample 0AMRd057A occurred in both air and water samples, which some approaches would consider to be evidence of contamination, even though SourceTracker2 estimated that <1% of sequences in this sample could be assigned to water or air sources (Fig. 3). Therefore, we explored other methods for the identification of individual ASVs as contaminants.
Identification of contaminants from air and seawater.
Of the 293 ASVs detected in air samples, 187 also occurred in serpentinites and were therefore designated contaminants (Fig. 4A). Thus, all remaining ASVs were absent in air and were only detected in water and serpentinite samples. Next, two approaches, simple overlap (SO) and differential abundance (DA), were used in parallel to distinguish the serpentinite ASVs from the water ASVs. In the SO approach (Fig. 4B), the ASVs shared between water and serpentinite samples were removed from the data set regardless of their abundances in water or serpentinites (i.e., only considering presence or absence). Thus, the results from the SO approach include only those ASVs that are exclusively found in serpentinite samples (664 ASVs).
FIG 4.

Workflow for identifying DNA sequence contaminants. (A) All ASVs detected in air samples were removed from the data set. (B) The simple overlap (SO) approach was used to remove all ASVs detected in water samples. (C) The differential abundance (DA) approach was used to remove only ASVs that were significantly more abundant in water samples compared to serpentinite samples. Differentially abundant ASVs are shown as red data points in the plot. The significance threshold was a false detection rate of 0.05. Only one of the comparisons is shown here (deep water samples versus serpentinite samples). Shallow and surface water samples were compared in separate analyses not shown here.
In the DA approach (Fig. 4C), the sequence counts (i.e., the number of merged paired reads) of each ASV shared between water and serpentinite samples were compared. ASVs that were significantly more abundant in water samples compared to serpentinites were identified and then removed from the data set (Table S3). In this case, ASVs that are highly abundant in serpentinites and very rare (but still present) in water are not necessarily identified as contaminants, as they were in the SO approach. Three edgeR comparisons were performed between serpentinite samples and each depth category of water samples (serpentinites versus surface, serpentinites versus shallow, and serpentinites versus deep). The ASVs that were more abundant in each group of water samples (one example in Fig. 4C) were designated contaminants from water and excluded from further analyses. The ASVs identified as water contaminants and detected in serpentinites are listed in Data Set S5. The DA approach results in a final list of 684 ASVs putatively considered to be true inhabitants of serpentinites (Data Set S6), including 20 additional ASVs compared to the SO approach. Seven of these 20 ASVs were present at >100 counts in serpentinite samples and less than a total of 53 counts in all water samples (Data Set S7).
Taxonomic composition of serpentinites.
To conservatively report a shorter list of likely serpentinite inhabitants, the following analyses and visualizations only include ASVs with at least 100 total counts among all samples. Taxonomic classifications of the putative serpentinite ASVs from the SO approach are summarized in Fig. 5. For clarity, the taxonomic classifications of only the top 50 most abundant ASVs are shown, and more information about these 50 ASVs is provided in Data Set S7. The taxonomic compositions of serpentinite samples are variable, consistent with the spread of serpentinites shown in the NMDS plot (Fig. 2). No significant differences with respect to sample handling (flamed/unflamed, shaved/unshaved, and washed/unwashed) could be detected. Indeed, very few similarities between any two samples were apparent.
FIG 5.

Taxonomic summary of ASVs detected in serpentinite samples after contaminant removal. For clarity, only the top 50 ASVs among all samples (Data Set S7) are shown here. N is the total ASV counts for each sample after excluding rare ASVs. Samples 0AMRd076A, -075A, -072A, and -067A were collected from hole M0076B. Samples 0AMRd031A and -030A were collected from hole M0071B. No other samples in this figure were collected from the same hole.
Sample 0AMRd057A (Fig. 6) provided the deepest sequencing data set (98,462 merged paired reads among those ASVs with at least 100 counts). Most of the taxonomic diversity in this sample was represented by rare ASVs (i.e., those with lower abundance than the top 50 shown in Fig. 5). The most abundant ASV was classified as family Thermoplasmatales (phylum Euryarchaeota) and has 99% identity to a clone (NCBI accession number AB825685; Data Set S7) from a deep-sea sediment core in the Okinawa Trough (48). Two of the top ASVs in this sample were classified as two different classes of Chloroflexi (S085 and Anaerolineae), each of which was unique to sample 0AMRd057A and was most similar to clones from marine sediments (accession numbers AM998115 and HQ721355). Another top ASV unique to this sample was classified as Acidimicrobia clade OM1 (phylum Actinobacteria), which was nearly identical to a clone from a marine endolithic community (49). Two ASVs in 0AMRd057A were classified as Acidobacteria subgroup 21 and were most similar to sequences from marine sediments, including a clone (accession number KY977768) from altered rocks of the Mariana subduction zone, where serpentinization also occurs (50).
FIG 6.

(A) Taxonomic summary of serpentinite sample 0AMRd057 (357-75A-1R-CC,0-4cm). The relative abundance of each taxonomic group is its proportion of 103,229 total sequence counts among the final 331 ASVs identified by the differential abundance (DA) approach. (B) This sample was recovered from the core catcher section of the core from borehole M0075A. The scale is presented in millimeters (74).
The deepest sequencing data sets of the remaining samples were obtained from 0AMRd045A, 0AMRd072A, 0AMRd075A, and 0AMRd076A, which were collected from the sites closest to the Lost City chimneys (boreholes M0072B and M0076B; Fig. 1). Each of these samples were dominated by Clostridia and Alphaproteobacteria (Fig. 5), and most of these ASVs match sequences associated with animal digestive tracts or soil. An exception is a Rhodobacter ASV unique to 0AMRd045A that matches several sequences from lakes (e.g., accession number MF994002). An abundant Sphingomonadaceae ASV in 0AMRd075A matches a clone from groundwater near the Hanford Site, WA (accession number KT431076). A Dechloromonas ASV unique to 0AMRd076A (Site M0076) is identical to a clone from an ocean drilling expedition to the South Chamorro Seamount (accession number LC279322), which is also a site of subseafloor serpentinization (51). However, the genus Dechloromonas has also been detected on human skin (52).
Sample 0AMRd030A is dominated by two ASVs classified as genera Acidovorax (class Betaproteobacteria) and Cobetia (class Gammaproteobacteria), respectively. The Acidovorax ASV matches sequences from soil (e.g., accession number CP021359) and an anoxic lake (accession number KY515689) (53). Cobetia marina (previously Halomonas marina) is a ubiquitous marine bacterium (54) and also a model organism for the study of biofouling due to its ability to grow as a biofilm on a variety of surfaces (55).
Other abundant ASVs shown in Fig. 5 and listed in Data Set S7 include those classified as Acidobacteria, Actinobacteria, Betaproteobacteria, Gemmatimonadetes, and Ktedonobacteria, and these sequences were generally most similar to environmental sequences from soils or freshwater. One possible exception is an ASV that was abundant in sample 0AMRd031A and was classified as genus Modestobacter, a soil bacterium, but it was also identical to another species in the same family, Klenkia marina, which was isolated from marine sediment (accession number NR_156810).
DISCUSSION
Serpentinite samples have minimal environmental contamination.
Sequencing of environmental DNA from seafloor rocks is challenging due to their very low biomass and their abundance of minerals (e.g., phyllosilicates) that inhibit enzymatic reactions and bind DNA (56–58). The extremely low biomass of seafloor rocks also makes them susceptible to environmental and laboratory contamination. Many precautions were taken during handling of the rock core samples to minimize, eliminate, and detect contamination, and our results agree with previous reports that these efforts were generally successful (20, 59, 60).
To assess environmental contamination of the cores, perfluoromethylcyclohexane (PFC) was injected into the seabed drilling fluids during coring. Orcutt et al. (59) reported that the interior zones of rocks whose outer surfaces had been shaved off with a sterile rock saw had greatly reduced levels of PFC. Only one sample in our study (0AMRd073), whose exterior had been shaved, contained detectable levels of PFC. Two additional samples (0AMRd030 and 0AMRd036), which were too rubbly to be shaved with the rock saw and were instead washed with ultrapure water, contained detectable PFC. In general, however, washing rubbly samples with ultrapure water was largely effective in reducing the level of PFC and presumably of any other surface contamination (59). The PFC tracer, however, was not designed to track any seawater contamination that might have occurred after drilling or any contamination that might have resulted from handling the rock cores shipboard or in the laboratory.
We assessed environmental contamination of the serpentinites by sequencing DNA from samples of seawater collected above the boreholes. The contribution of seawater DNA sequences into the serpentinites was minimal (Fig. 3), further demonstrating that the sampling-handling precautions on the ship and at the Kochi Core Center were successful in eliminating environmental contamination that must have occurred at some level during core recovery on the seafloor. In addition, our results agree with those of Orcutt et al. (59) that there was no noticeable difference in contamination levels between rocks whose outer surfaces had been subjected to flame sterilization compared to those that were not flamed.
Serpentinite samples have minimal environmental DNA.
Our DNA yields (Table 1) were higher than expected from the previously reported numbers of cells for these serpentinites (20), all of which were only slightly greater than the quantification limit of ∼10 cells cm−3. Uncertainties and imprecision in the quantification of DNA and of cell density could partially explain this discrepancy, but it is also likely that much of the total environmental DNA was primarily extracellular. Nearly all of the DNA was lost during purification and fell below our detection limit, consistent with highly fragmented DNA. Neither the filtration units used for washing nor the electrophoretic purification method are expected to recover DNA below 600 bp. Additional analyses of DNA size distributions were not practical with such low concentrations of DNA from irreplaceable samples. Although the concentrations of purified DNA were below our detection limit, amplicon sequencing of 16S rRNA genes was moderately successful, thanks to the elimination of PCR inhibitors by the electrophoretic SCODA purification.
Identification of laboratory contaminants.
We identified laboratory contaminants as amplicon sequence variants (ASVs) that occurred in samples of both air and serpentinites. This simple overlap (SO) approach is commonly used in environmental microbiology studies to remove all sequences (or taxonomic categories) that are shared between any sources of contamination and the sample of interest (41). An important assumption of the SO approach is that the direction of contamination is from the suspected contamination source into the sample. However, this assumption is often untested. For example, if a low-DNA control sample is handled in close proximity (e.g., the same 96-well plate) to a high-DNA environmental sample, it may be more likely for “contamination” to occur from the environmental sample into the control sample. Subsequent use of the SO approach would erroneously identify an abundant member of the environmental sample as a laboratory contaminant.
We considered the possibility of “reverse contamination” from our core samples into our air samples to be unlikely for multiple reasons. First, most of the processing of the rock samples, including sawing, crushing, washing, and subsampling, occurred at the Kochi Core Center in Japan, and our samples of air were collected in our laboratory at the University of Utah. Second, our SourceTracker2 results implicated air as a major source of DNA into the serpentinites (Fig. 3). Third, nearly all of the taxa detected in the air samples are typically associated with humans (e.g., Pseudomonas, Staphylococcus, and Acinetobacter). Our list of laboratory air taxa (Data Set S4) may be useful for identifying contaminants in future studies as a complement to previously reported lists of contaminants from laboratory reagents (39–41).
Identification of environmental contaminants.
Identification of environmental contaminants from seawater into the serpentinites was more complex because of the greater possibility of mixing between seawater and rocks in both directions. The Lost City chimneys provide dramatic evidence of the flux of biological material from the rocky subsurface into ambient seawater (25), and flux from the boreholes in this study was also evident by the release of bubbles during drilling (20). Deep seawater near the Atlantis Massif cannot be assumed to be completely free of subsurface inhabitants, and samples of seawater at shallower depths or locations far removed from the Atlantis Massif would not be accurate representations of the environmental contamination that the serpentinite samples experienced during recovery on the seafloor. Therefore, the relative abundances of taxa in samples of deep seawater and serpentinites should be informative because true subsurface organisms that are mixed into ambient seawater should be immediately diluted and present in seawater only at very low abundances.
In such situations of environmental mixing, the SO approach can erroneously identify contaminants because it ignores abundance information. We identified at least one such example in our study by comparing the results of the SO approach to those of the differential abundance (DA) approach. An ASV classified as Acidobacteria subgroup 21 occurred 264 times in a serpentinite sample (0AMRd057A; 0.3% of total sequence counts in that sample) from borehole M0075 and also occurred a total of 12 times among five deep seawater samples collected from above boreholes M0068, M0072, and M0073 (Data Set S6). The ASV was identical to several sequences from marine sediments, as well as one from altered rocks of the Mariana subduction zone (accession number KY977768) and another from seafloor lavas near the Loi’hi seamount (accession number EU491098) (17).
This was the most striking example in our study, but there were 19 additional ASVs flagged as contaminants by the SO approach but not by the DA approach. This relatively small difference (664 compared to 684 total ASVs in serpentinites) is probably a consequence of the success in minimizing and eliminating environmental contamination during handling of the rock cores, as discussed above and demonstrated by the minimal level of contamination from seawater shown in Fig. 3. Differences between the SO and DA approaches are likely to be greater in studies where environmental contamination cannot be controlled so carefully.
Identification of contaminants by taxonomy.
After removal of DNA sequences identified as contaminants from seawater and air, the most abundant serpentinite ASVs (Data Set S6) still contained many taxa that seemed unlikely to be true inhabitants of the subseafloor. Some are typically associated with animal digestive tracts (e.g., Ruminiclostridium), some are found in soil (e.g., various Acidobacteria), and others have been previously identified as reagent contaminants (e.g., Micrococcus) (39, 40). The sources of these likely contaminants could not be determined in this study, but the extraction and sequencing reagents are strong candidates, even though we took great care in preparation of our own reagents at every step and did not use any commercial extraction or purification kits. Also, we only sampled and sequenced DNA from the air in our own laboratory at the University of Utah and did not sample the air inside the research vessel, the Kochi Core Center, or the DNA sequencing core facility at Michigan State University.
Identification of contaminants at the level of taxonomic category is risky, however, because organisms that are capable of persisting in the extreme environments of mostly sterile laboratory supplies may also be capable of inhabiting the extreme environments of mostly sterile subsurface rocks (61). For example, a relatively abundant serpentinite ASV that was absent in all seawater and air samples is nevertheless a suspected contaminant because it was classified as Acidovorax, a ubiquitous soil bacterium and plant pathogen that has also been identified as a contaminant in laboratory reagents (40). The ability of Acidovorax to form biofilms and perform anaerobic iron oxidation (62), however, would be advantageous in the serpentinite subsurface. Because we are suspicious of these taxa but lack direct experimental evidence of a source other than serpentinites, we include these ASVs in our final tables but flag them as suspected contaminants (Data Set S7).
Putative inhabitants of the serpentinite subsurface.
Excluding all air and seawater ASVs and ignoring all suspicious taxa leaves us with a few putative inhabitants of subseafloor serpentinites (Data Set S7). Many of these ASVs were unique to a single serpentinite sample shown in Fig. 6A. This serpentinite was recovered from the most eastern site (borehole M0075A) at a depth of ∼60 cm below the seafloor, and it had extensive talc-amphibole alteration, suggesting that it was altered during uplift and emplacement of the massif. The most abundant taxa in this sample included Euryarchaeota, Chloroflexi, Actinobacteria, and Acidobacteria, all of which matched DNA sequences previously found in marine sediments and rocks. The most abundant ASV was classified as Thermoplasmatales (an order within the Euryarchaeota) and was nearly identical to a clone from hydrothermal sediment collected during IODP Expedition 331 to the Mid-Okinawa Trough (48). Most, but not all, of the characterized members of the Thermoplasmatales are thermophilic, and they are typically involved in sulfur cycling (63, 64). The Thermoplasmatales are generally known as acidophiles (65), which is curious since serpentinization is associated with high pH.
None of these taxa are known to be capable of metabolizing hydrogen (H2) or methane, as is prevalent among the residents of Lost City chimney biofilm communities (25, 34, 66). Metagenomic studies have revealed that the taxonomic and environmental distribution of H2 metabolism is more widespread than has been previously appreciated (67), so future work should investigate the possibility that some of the taxa detected here encode genes for H2 metabolism. Nevertheless, one would not expect to find the same biofilm communities of the Lost City in the basement rocks of the Atlantis Massif, where hydrothermal circulation is much more diffuse compared to the high flux through Lost City chimneys. Furthermore, the sample in Fig. 6A was obtained from borehole M0075A, which exhibited low levels of H2 compared to other Expedition 357 boreholes (20).
The most abundant taxa in serpentinites recovered from the central drill sites (boreholes M0069, M0072, and M0076) were more difficult to interpret. At least two probable subsurface taxa (Sphingomonadaceae and Dechloromonas) were detected in these serpentinites, but many of the other sequences from these samples are identical to environmental sequences from studies of animal digestive tracts or soils. Therefore, serpentinites from the central sites, which are considered to be most representative of the massif’s basement, appear to be more susceptible to undetected contamination, perhaps due to extremely low DNA yields from these samples.
Recently, Quéméneur et al. (60) reported enrichment cultures obtained from IODP Expedition 357 rock cores, including two serpentinite samples. One of these was a section of a serpentinite core (357-71C-5R-CC) that was adjacent to a sample included in the present study (0AMRd036A). Our parallel sample was heavily contaminated with air DNA (Fig. 3) and did not contain any of the dominant taxa detected by Quéméneur et al. Many of the taxa detected in their cultivation experiments were also present in our results, but only one genus reported by Quéméneur et al. (Sphingomonas) was included in our final list of candidate serpentinite taxa (Data Set S7). Most of the successful enrichments reported by Quéméneur et al. were obtained from a carbonate-hosted basalt breccia, which was not included in our study.
In 2005, IODP expedition 304/305 recovered gabbroic rocks from the central dome of the Atlantis Massif, but no serpentinites were reported (Fig. 1). A few phylotypes of Proteobacteria were detected in the gabbroic rock cores by sequencing of environmental 16S rRNA genes (19). None of these matched the candidate serpentinite taxa reported here. However, most of the functional genes detected by Mason et al. (19) with the GeoChip microarray, including genes associated with metal toxicity, carbon degradation, denitrification, and carbon fixation, are consistent with the taxa detected in our serpentinite samples.
Conclusions.
This study provided a census of environmental DNA sequences from subseafloor serpentinites, enabled by the high recovery of rock cores by IODP Expedition 357 to the Atlantis Massif (20). The extremely low biomass of the serpentinites presented multiple challenges, necessitating the development of a novel DNA extraction and purification protocol. We developed strategies to increase the yield of DNA appropriate for PCR amplification, while eliminating many potential sources of laboratory contamination. We hope that our methodology, as well as our identification of laboratory contaminants derived from dust particles, will prove to be useful to other researchers studying extremely low-biomass environments. We were able to identify candidate residents of the serpentinite subseafloor by employing a series of contamination-detection procedures, including evaluations of whole-sample compositions and of individual sequences. Our results highlight the importance of a multifaceted approach to contamination control that features sampling of potential contamination sources as a complement to microbiologically clean sample-processing protocols. Multiple statistical and bioinformatic tools were used to identify contaminant DNA sequences in a careful process that did not unnecessarily ignore useful information, such as the relative abundances of individual DNA sequences in potential contamination sources. No computational tool can ever prove that an environmental DNA sequence is not a contaminant, and future studies will investigate candidate subseafloor organisms with experimental studies.
MATERIALS AND METHODS
Detailed descriptions of samples and sample-processing protocols are available in the supplemental methods (see Text S1). Frozen, homogenized subsamples of rock core samples prepared by the IODP Expedition 357 microbiology team at the Kochi Core Center were aliquoted (0.5 g) into 2-ml tubes containing a pH 10 DNA extraction buffer (0.03 M Tris-HCl, 0.01 M EDTA, 0.02 M EGTA, 0.1 M KH2PO4, 0.8 M guanidine-HCl, 0.5% Triton X-100) and mixed with 150 μl of 20% sodium pyrophosphate and 150 μl of 50 mM dATP (Jena Bioscience, Jena, Germany). Tubes were beaten with a MiniBeadBeater-16 Model 607 (Biospec, Bartlesville, OK). DNA purification was performed with SCODA (synchronous coefficient of drag alteration) technology implemented with the Aurora purification system (Boreal Genomics, Vancouver, BC, Canada). 16S rRNA gene amplicon sequencing was conducted by the Michigan State University genomics core facility on all of the samples (rock, water, and lab air). All samples were sequenced twice (i.e., sequencing replicates). The V4 region of the bacterial 16S rRNA gene was amplified with dual-indexed Illumina fusion primers (515F/806R) as described elsewhere (68). 16S rRNA gene amplicon sequences were processed with cutadapt v. 1.15 (69) and DADA2 v1.10.1 (70). Taxonomic classification of all ASVs was performed with DADA2 using the SILVA reference alignment (SSURefv132) and taxonomy outline (71). The major sources of contamination and the level of contamination in each sample were estimated with SourceTracker2, v2.0.1 (47). Differential abundance was tested with the R package edgeR v3.24.3 (72) as recommended elsewhere (73). We used edgeR to contrast the total read counts of ASVs in serpentinite rock samples compared to three groups of water samples (surface, shallow, and deep). ASVs that were absent in all serpentinites and ASVs with low variance (<1e–6) were excluded from the comparisons. The output of the three edgeR tests (serpentinite samples compared to each of the three groups of water samples) was three lists of ASVs with significant differential abundances (false discovery rate < 0.05) in serpentinite samples or water samples. The final list of ASVs was created by deleting ASVs with greater abundances in any of the categories of water samples (as determined by the edgeR tests) from the original list of ASVs from which all air ASVs had already been removed. Finally, rare ASVs (those that did not have ≥100 counts in a single sample) were excluded from the final results, merely as a conservative abundance filter for reporting a final list of ASVs expected to be present in serpentinite rocks. No comparisons of diversity were attempted after removing rare ASVs.
Data availability.
All unprocessed DNA sequence data are publicly available at the NCBI SRA under BioProject PRJNA575221. All supplementary data and protocols are available at https://github.com/Brazelton-Lab/Atlantis-Massif-2015. All custom software and scripts are available at https://github.com/Brazelton-Lab.
Supplementary Material
ACKNOWLEDGMENTS
This research used data and samples provided by the International Ocean Discovery Program (IODP) Expedition 357 supported by the European Consortium for Ocean Research Drilling (ECORD) and implemented by the ECORD Science Operator (ESO). The shipboard sample-processing protocols were executed by ESO Expedition Project Manager Carol Cotterill, ESO Operations Superintendent David Smith, the crews of the R.R.S. James Cook and the MeBo and RD2 seabed drills, and shipboard scientists Susan Lang, Marvin Lilley, Yuki Morono, Marianne Quéméneur, and Matthew Schrenk. We are indebted to Fumio Inagaki for welcoming our use of sampling equipment at the JAMSTEC Kochi Core Center (KCC); to Nan Xiao at KCC for coordinating our sample processing and operation of core sectioning equipment; to Christopher Thornton, Katherine Hickock, and Susan Lang for manually processing the rock samples at KCC; and to ESO staff Ursula Röhl and David McInroy for helping to coordinate sample transfer and tracking. Christopher Thornton, Alex Hyer, Emily Dart, and Julia McGonigle provided laboratory and computational assistance. We also thank Jackie Goordial for comments on an earlier draft. The Expedition 357 core description team helpfully provided descriptions of the core lithologies.
Funding for this work was provided by the NASA Astrobiology Institute Rock-Powered Life team, the U.S. National Science Foundation-funded US Science Support Program (subawards to W.J.B. and B.N.O.), the Deep Carbon Observatory funded by the Alfred P. Sloan Foundation (via subaward to B.N.O. from the Marine Biological Laboratory), and by the Swiss National Science Foundation (SNSF) project no. 200021_163187 (to G.L.F.-G.) and SNSF contributions to the Swiss IODP.
Footnotes
Supplemental material is available online only.
REFERENCES
- 1.Whitman WB, Coleman DC, Wiebe WJ. 1998. Prokaryotes: the unseen majority. Proc Natl Acad Sci U S A 95:6578–6583. doi: 10.1073/pnas.95.12.6578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bar-On YM, Phillips R, Milo R. 2018. The biomass distribution on Earth. Proc Natl Acad Sci U S A 115:6506–6511. doi: 10.1073/pnas.1711842115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lipp JS, Morono Y, Inagaki F, Hinrichs K-U. 2008. Significant contribution of Archaea to extant biomass in marine subsurface sediments. Nature 454:991–994. doi: 10.1038/nature07174. [DOI] [PubMed] [Google Scholar]
- 4.Kallmeyer J, Pockalny R, Adhikari RR, Smith DC, D’Hondt S. 2012. Global distribution of microbial abundance and biomass in subseafloor sediment. Proc Natl Acad Sci U S A 109:16213–16216. doi: 10.1073/pnas.1203849109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Orcutt BN, Sylvan JB, Knab NJ, Edwards KJ. 2011. Microbial ecology of the dark ocean above, at, and below the seafloor. Microbiol Mol Biol Rev 75:361–422. doi: 10.1128/MMBR.00039-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Takai K, IODP Expedition 331 Scientists, Mottl MJ, Nielsen SH. 2012. IODP Expedition 331: strong and expansive subseafloor hydrothermal activities in the Okinawa Trough. Sci Drill 13:19–27. doi: 10.5194/sd-13-19-2012. [DOI] [Google Scholar]
- 7.Schrenk MO, Huber JA, Edwards KJ. 2010. Microbial provinces in the subseafloor. Annu Rev Mar Sci 2:279–304. doi: 10.1146/annurev-marine-120308-081000. [DOI] [PubMed] [Google Scholar]
- 8.Mottl MJ, Wheat CG. 1994. Hydrothermal circulation through mid-ocean ridge flanks: fluxes of heat and magnesium. Geochim Cosmochim Acta 58:2225–2237. doi: 10.1016/0016-7037(94)90007-8. [DOI] [Google Scholar]
- 9.Fisher AT, Becker K. 2000. Channelized fluid flow in oceanic crust reconciles heat-flow and permeability data. Nature 403:71–74. doi: 10.1038/47463. [DOI] [PubMed] [Google Scholar]
- 10.Santelli CM, Edgcomb VP, Bach W, Edwards KJ. 2009. The diversity and abundance of bacteria inhabiting seafloor lavas positively correlate with rock alteration. Environ Microbiol 11:86–98. doi: 10.1111/j.1462-2920.2008.01743.x. [DOI] [PubMed] [Google Scholar]
- 11.Singer E, Chong LS, Heidelberg JF, Edwards KJ. 2015. Similar microbial communities found on two distant seafloor basalts. Front Microbiol 6:1409. doi: 10.3389/fmicb.2015.01409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Edwards KJ, Wheat CG, Sylvan JB. 2011. Under the sea: microbial life in volcanic oceanic crust. Nat Rev Microbiol 9:703–712. doi: 10.1038/nrmicro2647. [DOI] [PubMed] [Google Scholar]
- 13.Smith AR, Fisk MR, Thurber AR, Flores GE, Mason OU, Popa R, Colwell FS. 2017. Deep crustal communities of the Juan de Fuca Ridge are governed by mineralogy. Geomicrobiol J 34:147–156. doi: 10.1080/01490451.2016.1155001. [DOI] [Google Scholar]
- 14.Amend JP, Rogers KL, Shock EL, Gurrieri S, Inguaggiato S. 2003. Energetics of chemolithoautotrophy in the hydrothermal system of Vulcano Island. South Italy Geobiol 1:37–58. doi: 10.1046/j.1472-4669.2003.00006.x. [DOI] [Google Scholar]
- 15.Lollar BS. 2004. Life’s chemical kitchen. Science 304:972–973. doi: 10.1126/science.1098112. [DOI] [PubMed] [Google Scholar]
- 16.Okland I, Huang S, Dahle H, Thorseth IH, Pedersen RB. 2012. Low temperature alteration of serpentinized ultramafic rock and implications for microbial life. Chem Geol 318–319:75–87. doi: 10.1016/j.chemgeo.2012.05.015. [DOI] [Google Scholar]
- 17.Santelli CM, Orcutt BN, Banning E, Bach W, Moyer CL, Sogin ML, Staudigel H, Edwards KJ. 2008. Abundance and diversity of microbial life in ocean crust. Nature 453:653–656. doi: 10.1038/nature06899. [DOI] [PubMed] [Google Scholar]
- 18.Mason OU, Meo-Savoie CAD, Van Nostrand JD, Zhou J, Fisk MR, Giovannoni SJ. 2009. Prokaryotic diversity, distribution, and insights into their role in biogeochemical cycling in marine basalts. ISME J 3:231–242. doi: 10.1038/ismej.2008.92. [DOI] [PubMed] [Google Scholar]
- 19.Mason OU, Nakagawa T, Rosner M, Van Nostrand JD, Zhou J, Maruyama A, Fisk MR, Giovannoni SJ. 2010. First investigation of the microbiology of the deepest layer of ocean crust. PLoS One 5:e15399. doi: 10.1371/journal.pone.0015399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Früh-Green GL, Orcutt BN, Rouméjon S, Lilley MD, Morono Y, Cotterill C, Green S, Escartin J, John BE, McCaig AM, Cannat M, Ménez B, Schwarzenbach EM, Williams MJ, Morgan S, Lang SQ, Schrenk MO, Brazelton WJ, Akizawa N, Boschi C, Dunkel KG, Quéméneur M, Whattam SA, Mayhew L, Harris M, Bayrakci G, Behrmann J-H, Herrero-Bervera E, Hesse K, Liu H-Q, Ratnayake AS, Twing K, Weis D, Zhao R, Bilenker L. 2018. Magmatism, serpentinization, and life: insights through drilling the Atlantis Massif (IODP Expedition 357). Lithos 323:137–155. doi: 10.1016/j.lithos.2018.09.012. [DOI] [Google Scholar]
- 21.Früh-Green GL, Orcutt BN, Green SL, Cotterill C, Morgan S, Akizawa N, Bayrakci G, Behrmann JH, Boschi C, Brazleton WJ. 2017. Expedition 357 summary. Proc Int Ocean Discov Program, vol 357. doi: 10.14379/iodp.proc.357.101.2017. [DOI] [Google Scholar]
- 22.Kelley DS, AT3-60 Shipboard Party, Karson JA, Blackman DK, Früh-Green GL, Butterfield DA, Lilley MD, Olson EJ, Schrenk MO, Roe KK, Lebon GT, Rivizzigno P. 2001. An off-axis hydrothermal vent field near the Mid-Atlantic Ridge at 30°N. Nature 412:145–149. doi: 10.1038/35084000. [DOI] [PubMed] [Google Scholar]
- 23.Blackman DK, Karson JA, Kelley DS, Cann JR, Früh-Green GL, Gee JS, Hurst SD, John BE, Morgan J, Nooner SL, Ross DK, Schroeder TJ, Williams EA. 2002. Geology of the Atlantis Massif (Mid-Atlantic Ridge, 30°N: implications for the evolution of an ultramafic oceanic core complex. Mar Geophys Res 23:443–469. doi: 10.1023/B:MARI.0000018232.14085.75. [DOI] [Google Scholar]
- 24.Allen DE, Seyfried WE. 2004. Serpentinization and heat generation: constraints from Lost City and Rainbow hydrothermal systems. Geochim Cosmochim Acta 68:1347–1354. doi: 10.1016/j.gca.2003.09.003. [DOI] [Google Scholar]
- 25.Kelley DS, Karson JA, Früh-Green GL, Yoerger DR, Shank TM, Butterfield DA, Hayes JM, Schrenk MO, Olson EJ, Proskurowski G, Jakuba M, Bradley A, Larson B, Ludwig K, Glickson D, Buckman K, Bradley AS, Brazelton WJ, Roe K, Elend MJ, Delacour A, Bernasconi SM, Lilley MD, Baross JA, Summons RE, Sylva SP. 2005. A serpentinite-hosted ecosystem: the Lost City hydrothermal field. Science 307:1428–1434. doi: 10.1126/science.1102556. [DOI] [PubMed] [Google Scholar]
- 26.McCollom TM, Seewald JS. 2007. Abiotic synthesis of organic compounds in deep-sea hydrothermal environments. Chem Rev 107:382–401. doi: 10.1021/cr0503660. [DOI] [PubMed] [Google Scholar]
- 27.Holm NG, Charlou JL. 2001. Initial indications of abiotic formation of hydrocarbons in the Rainbow ultramafic hydrothermal system, Mid-Atlantic Ridge. Earth Planet Sci Lett 191:1–8. doi: 10.1016/S0012-821X(01)00397-1. [DOI] [Google Scholar]
- 28.Konn C, Charlou J-L, Donval J-P, Holm NG, Dehairs F, Bouillon S. 2009. Hydrocarbons and oxidized organic compounds in hydrothermal fluids from Rainbow and Lost City ultramafic-hosted vents. Chem Geol 258:299–314. doi: 10.1016/j.chemgeo.2008.10.034. [DOI] [Google Scholar]
- 29.Lang SQ, Früh-Green GL, Bernasconi SM, Lilley MD, Proskurowski G, Méhay S, Butterfield DA. 2012. Microbial utilization of abiogenic carbon and hydrogen in a serpentinite-hosted system. Geochim Cosmochim Acta 92:82–99. doi: 10.1016/j.gca.2012.06.006. [DOI] [Google Scholar]
- 30.Schrenk MO, Brazelton WJ, Lang SQ. 2013. Serpentinization, carbon, and deep life. Rev Mineral Geochem 75:575–606. doi: 10.2138/rmg.2013.75.18. [DOI] [Google Scholar]
- 31.Früh-Green GL, Kelley DS, Bernasconi SM, Karson JA, Ludwig KA, Butterfield DA, Boschi C, Proskurowski G. 2003. 30,000 years of hydrothermal activity at the Lost City vent field. Science 301:495–498. doi: 10.1126/science.1085582. [DOI] [PubMed] [Google Scholar]
- 32.Ludwig KA, Kelley DS, Butterfield DA, Nelson BK, Früh-Green G. 2006. Formation and evolution of carbonate chimneys at the Lost City hydrothermal field. Geochim Cosmochim Acta 70:3625–3645. doi: 10.1016/j.gca.2006.04.016. [DOI] [Google Scholar]
- 33.Schrenk MO, Kelley DS, Bolton SA, Baross JA. 2004. Low archaeal diversity linked to subseafloor geochemical processes at the Lost City hydrothermal field, Mid-Atlantic Ridge. Environ Microbiol 6:1086–1095. doi: 10.1111/j.1462-2920.2004.00650.x. [DOI] [PubMed] [Google Scholar]
- 34.Brazelton WJ, Schrenk MO, Kelley DS, Baross JA. 2006. Methane-and sulfur-metabolizing microbial communities dominate the Lost City hydrothermal field ecosystem. Appl Environ Microbiol 72:6257–6270. doi: 10.1128/AEM.00574-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Brazelton WJ, Mehta MP, Kelley DS, Baross JA. 2011. Physiological differentiation within a single-species biofilm fueled by serpentinization. mBio 2:e00127-11. doi: 10.1128/mBio.00127-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lang SQ, Früh-Green GL, Bernasconi SM, Brazelton WJ, Schrenk MO, McGonigle JM. 2018. Deeply-sourced formate fuels sulfate reducers but not methanogens at Lost City hydrothermal field. Sci Rep 8:755. doi: 10.1038/s41598-017-19002-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lang SQ, Brazelton WJ. 2020. Habitability of the marine serpentinite subsurface: a case study of the Lost City hydrothermal field. Philos Trans R Soc A 378:20180429. doi: 10.1098/rsta.2018.0429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Tanner MA, Goebel BM, Dojka MA, Pace NR. 1998. Specific ribosomal DNA sequences from diverse environmental settings correlate with experimental contaminants. Appl Environ Microbiol 64:3110–3113. doi: 10.1128/AEM.64.8.3110-3113.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Barton HA, Taylor NM, Lubbers BR, Pemberton AC. 2006. DNA extraction from low-biomass carbonate rock: an improved method with reduced contamination and the low-biomass contaminant database. J Microbiol Methods 66:21–31. doi: 10.1016/j.mimet.2005.10.005. [DOI] [PubMed] [Google Scholar]
- 40.Salter SJ, Cox MJ, Turek EM, Calus ST, Cookson WO, Moffatt MF, Turner P, Parkhill J, Loman NJ, Walker AW. 2014. Reagent and laboratory contamination can critically impact sequence-based microbiome analyses. BMC Biol 12:87. doi: 10.1186/s12915-014-0087-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sheik CS, Reese BK, Twing KI, Sylvan JB, Grim SL, Schrenk MO, Sogin ML, Colwell FS. 2018. Identification and removal of contaminant sequences from ribosomal gene databases: lessons from the census of deep life. Front Microbiol 9:840. doi: 10.3389/fmicb.2018.00840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Labonté JM, Lever MA, Edwards KJ, Orcutt BN. 2017. Influence of igneous basement on deep sediment microbial diversity on the eastern Juan de Fuca Ridge flank. Front Microbiol 8:1434. doi: 10.3389/fmicb.2017.01434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Santelli CM, Banerjee N, Bach W, Edwards KJ. 2010. Tapping the subsurface ocean crust biosphere: low biomass and drilling-related contamination calls for improved quality controls. Geomicrobiol J 27:158–169. doi: 10.1080/01490450903456780. [DOI] [Google Scholar]
- 44.Lever MA, Alperin M, Engelen B, Inagaki F, Nakagawa S, Steinsbu BO, Teske A. 2006. Trends in basalt and sediment core contamination during IODP Expedition 301. Geomicrobiol J 23:517–530. doi: 10.1080/01490450600897245. [DOI] [Google Scholar]
- 45.Davis NM, Proctor DM, Holmes SP, Relman DA, Callahan BJ. 2018. Simple statistical identification and removal of contaminant sequences in marker-gene and metagenomics data. Microbiome 6:226. doi: 10.1186/s40168-018-0605-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Inagaki F, Hinrichs K-U, Kubo Y, Bowles MW, Heuer VB, Hong W-L, Hoshino T, Ijiri A, Imachi H, Ito M, Kaneko M, Lever MA, Lin Y-S, Methe BA, Morita S, Morono Y, Tanikawa W, Bihan M, Bowden SA, Elvert M, Glombitza C, Gross D, Harrington GJ, Hori T, Li K, Limmer D, Liu C-H, Murayama M, Ohkouchi N, Ono S, Park Y-S, Phillips SC, Prieto-Mollar X, Purkey M, Riedinger N, Sanada Y, Sauvage J, Snyder G, Susilawati R, Takano Y, Tasumi E, Terada T, Tomaru H, Trembath-Reichert E, Wang DT, Yamada Y. 2015. Exploring deep microbial life in coal-bearing sediment down to ∼2.5 km below the ocean floor. Science 349:420–424. doi: 10.1126/science.aaa6882. [DOI] [PubMed] [Google Scholar]
- 47.Knights D, Kuczynski J, Charlson ES, Zaneveld J, Mozer MC, Collman RG, Bushman FD, Knight R, Kelley ST. 2011. Bayesian community-wide culture-independent microbial source tracking. Nat Methods 8:761–763. doi: 10.1038/nmeth.1650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Yanagawa K, Nunoura T, McAllister S, Hirai M, Breuker A, Brandt L, House C, Moyer CL, Birrien J-L, Aoike K, Sunamura M, Urabe T, Mottl M, Takai K. 2013. The first microbiological contamination assessment by deep-sea drilling and coring by the D/V Chikyu at the Iheya North hydrothermal field in the Mid-Okinawa Trough (IODP Expedition 331). Front Microbiol 4:327. doi: 10.3389/fmicb.2013.00327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Couradeau E, Roush D, Guida BS, Garcia-Pichel F. 2017. Diversity and mineral substrate preference in endolithic microbial communities from marine intertidal outcrops (Isla de Mona, Puerto Rico). Biogeosciences 14:311–324. doi: 10.5194/bg-14-311-2017. [DOI] [Google Scholar]
- 50.Hyndman RD, Peacock SM. 2003. Serpentinization of the forearc mantle. Earth Planet Sci Lett 212:417–432. doi: 10.1016/S0012-821X(03)00263-2. [DOI] [Google Scholar]
- 51.Kawagucci S, Miyazaki J, Morono Y, Seewald JS, Wheat CG, Takai K. 2018. Cool, alkaline serpentinite formation fluid regime with scarce microbial habitability and possible abiotic synthesis beneath the South Chamorro Seamount. Prog Earth Planet Sci 5:74. doi: 10.1186/s40645-018-0232-3. [DOI] [Google Scholar]
- 52.Grice EA, NISC Comparative Sequencing Program, Kong HH, Renaud G, Young AC, Bouffard GG, Blakesley RW, Wolfsberg TG, Turner ML, Segre JA. 2008. A diversity profile of the human skin microbiota. Genome Res 18:1043–1050. doi: 10.1101/gr.075549.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Schiff SL, Tsuji JM, Wu L, Venkiteswaran JJ, Molot LA, Elgood RJ, Paterson MJ, Neufeld JD. 2017. Millions of boreal shield lakes can be used to probe archaean ocean biogeochemistry. Sci Rep 7:46708. doi: 10.1038/srep46708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Arahal DR, Castillo AM, Ludwig W, Schleifer KH, Ventosa A. 2002. Proposal of Cobetia marina gen. nov., comb. nov., within the family Halomonadaceae, to include the species Halomonas marina. Syst Appl Microbiol 25:207–211. doi: 10.1078/0723-2020-00113. [DOI] [PubMed] [Google Scholar]
- 55.Mieszkin S, Martin-Tanchereau P, Callow ME, Callow JA. 2012. Effect of bacterial biofilms formed on fouling-release coatings from natural seawater and Cobetia marina, on the adhesion of two marine algae. Biofouling 28:953–968. doi: 10.1080/08927014.2012.723696. [DOI] [PubMed] [Google Scholar]
- 56.Direito SO, Marees A, Röling WF. 2012. Sensitive life detection strategies for low-biomass environments: optimizing extraction of nucleic acids adsorbing to terrestrial and Mars analogue minerals. FEMS Microbiol Ecol 81:111–123. doi: 10.1111/j.1574-6941.2012.01325.x. [DOI] [PubMed] [Google Scholar]
- 57.Saeki K, Sakai M. 2009. The influence of soil organic matter on DNA adsorptions on andosols. Microbes Environ 24:175–179. doi: 10.1264/jsme2.ME09117. [DOI] [PubMed] [Google Scholar]
- 58.Lever MA, Torti A, Eickenbusch P, Michaud AB, Šantl-Temkiv T, Jørgensen BB. 2015. A modular method for the extraction of DNA and RNA, and the separation of DNA pools from diverse environmental sample types. Front Microbiol 6:476. doi: 10.3389/fmicb.2015.00476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Orcutt BN, Bergenthal M, Freudenthal T, Smith D, Lilley MD, Schnieders L, Green S, Früh-Green GL. 2017. Contamination tracer testing with seabed drills: IODP Expedition 357. Sci Drill 23:39–46. doi: 10.5194/sd-23-39-2017. [DOI] [Google Scholar]
- 60.Quéméneur M, Erauso G, Frouin E, Zeghal E, Vandecasteele C, Ollivier B, Tamburini C, Garel M, Ménez B, Postec A. 2019. Hydrostatic pressure helps to cultivate an original anaerobic bacterium from the Atlantis Massif subseafloor (IODP Expedition 357): Petrocella atlantisensis gen. nov. sp. nov. Front Microbiol 10:1497. doi: 10.3389/fmicb.2019.01497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Swanner ED, Nell RM, Templeton AS. 2011. Ralstonia species mediate Fe-oxidation in circumneutral, metal-rich subsurface fluids of Henderson mine. CO Chem Geol 284:339–350. doi: 10.1016/j.chemgeo.2011.03.015. [DOI] [Google Scholar]
- 62.Byrne-Bailey KG, Weber KA, Chair AH, Bose S, Knox T, Spanbauer TL, Chertkov O, Coates JD. 2010. Completed genome sequence of the anaerobic iron-oxidizing bacterium Acidovorax ebreus strain TPSY. J Bacteriol 192:1475–1476. doi: 10.1128/JB.01449-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Arce-Rodríguez A, Puente-Sánchez F, Avendaño R, Martínez-Cruz M, de Moor JM, Pieper DH, Chavarría M. 2019. Thermoplasmatales and sulfur-oxidizing bacteria dominate the microbial community at the surface water of a CO2-rich hydrothermal spring located in Tenorio Volcano National Park, Costa Rica. Extremophiles 23:177–187. doi: 10.1007/s00792-018-01072-6. [DOI] [PubMed] [Google Scholar]
- 64.Barton LL, Fardeau M-L, Fauque GD. 2014. Hydrogen sulfide: a toxic gas produced by dissimilatory sulfate and sulfur reduction and consumed by microbial oxidation, p 237–277. In The metal-driven biogeochemistry of gaseous compounds in the environment. Springer, New York, NY. [DOI] [PubMed] [Google Scholar]
- 65.Yelton AP, Comolli LR, Justice NB, Castelle C, Denef VJ, Thomas BC, Banfield JF. 2013. Comparative genomics in acid mine drainage biofilm communities reveals metabolic and structural differentiation of co-occurring archaea. BMC Genomics 14:485. doi: 10.1186/1471-2164-14-485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Brazelton WJ, Nelson B, Schrenk MO. 2012. Metagenomic evidence for H2 oxidation and H2 production by serpentinite-hosted subsurface microbial communities. Front Microbiol 2:268. doi: 10.3389/fmicb.2011.00268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Greening C, Biswas A, Carere CR, Jackson CJ, Taylor MC, Stott MB, Cook GM, Morales SE. 2016. Genomic and metagenomic surveys of hydrogenase distribution indicate H2 is a widely utilized energy source for microbial growth and survival. ISME J 10:761–777. doi: 10.1038/ismej.2015.153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Kozich JJ, Westcott SL, Baxter NT, Highlander SK, Schloss PD. 2013. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl Environ Microbiol 79:5112–5120. doi: 10.1128/AEM.01043-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Martin M. 2011. Cutadapt removes adapter sequences from high-throughput sequencing reads. Embnet J 17:10–12. doi: 10.14806/ej.17.1.200. [DOI] [Google Scholar]
- 70.Callahan BJ, McMurdie PJ, Rosen MJ, Han AW, Johnson AJA, Holmes SP. 2016. DADA2: high-resolution sample inference from Illumina amplicon data. Nat Methods 13:581–583. doi: 10.1038/nmeth.3869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Pruesse E, Peplies J, Glöckner FO. 2012. SINA: accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 28:1823–1829. doi: 10.1093/bioinformatics/bts252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Robinson MD, McCarthy DJ, Smyth GK. 2010. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26:139–140. doi: 10.1093/bioinformatics/btp616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.McMurdie PJ, Holmes S. 2014. Waste not, want not: why rarefying microbiome data is inadmissible. PLoS Comput Biol 10:e1003531. doi: 10.1371/journal.pcbi.1003531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Früh-Green GL, Orcutt BN, Green SL, Cotterill C, Morgan S, Akizawa N, Bayrakci G, Behrmann J-H, Boschi C, Brazleton WJ. 2017. Supplementary material for volume 357 expedition reports. Proc Int Ocean Discov Program, vol 357. doi: 10.14379/iodp.proc.357supp.2017. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All unprocessed DNA sequence data are publicly available at the NCBI SRA under BioProject PRJNA575221. All supplementary data and protocols are available at https://github.com/Brazelton-Lab/Atlantis-Massif-2015. All custom software and scripts are available at https://github.com/Brazelton-Lab.