

Abstract
Purpose
Nonsteroidal anti-inflammatory drugs (NSAIDs) and selective COX-2 inhibitors (COXibs) inhibit the progression of endometrial cancer, ovarian cancer and cervical cancer. However, concerning the adverse effects of NSAIDs and COXibs, it is still urgent and necessary to explore novel and specific anti-inflammation targets for potential chemoprevention. The signaling of cyclooxygenase 2-prostaglandin E2-prostaglandin E2 receptors (COX-2-PGE2-EPs) is the central inflammatory pathway involved in the gynecological carcinogenesis.
Methods
Literature searches were performed to the function of COX-2-PGE2-EPs in gynecological malignancies.
Results
This review provides an overview of the current knowledge of COX-2-PGE2-EPs signaling in endometrial cancer, ovarian cancer and cervical cancer. Many studies demonstrated the upregulated expression of the whole signaling pathway in gynecological malignancies and some focused on the function of COX-2 and cAMP-linked EP2/EP4 and EP3 signaling pathway in gynecological cancer. By contrast, roles of EP1 and the exact pathological mechanisms have not been completely clarified. The studies concerning EP receptors in gynecological cancers highlight the potential advantage of combining COX enzyme inhibitors with EP receptor antagonists as therapeutic agents in gynecological cancers.
Conclusion
EPs represent promising anti-inflammation biomarkers for gynecological cancer and may be novel treatment targets in the near future.
Keywords: Cyclooxygenase-2 (COX-2), Prostaglandin E2 receptors (EPs), Endometrial cancer, Ovarian cancer, Cervical cancer
Introduction
Abundant literature has demonstrated a strong correlation between chronic inflammation and cancer development since chronic inflammation contributes to the development of over 15% of malignancies worldwide [1]. Plenty of pro-inflammatory factors mediate a role in carcinogenesis, such as tumour necrosis factor (TNF), interleukin (IL)-1α, IL-1β, IL-6, IL-8, IL-18, matrix metallopeptidase-9 (MMP-9), vascular endothelial growth factor (VEGF), cyclooxygenase 2 (COX-2), and arachidonate 5-lipoxygenase (5-LOX) [2]. Serum levels of C-reactive protein (CRP), IL-6, and IL-1 receptor antagonist (IL-1Ra) are significantly associated with endometrial cancer risk when analyzing 246,000 women in ten European countries [3]. The signaling of cyclooxygenase 2-prostaglandin E2-prostaglandin E2 receptors (COX-2-PGE2-EPs) is the central inflammatory pathway involved in the carcinogenesis (Fig. 1). Based on the current information, we aimed to supplement some additional knowledge of COX-2-PGE2-EPs in the carcinogenesis of gynecological cancer from the perspective of inflammation.
Fig. 1.
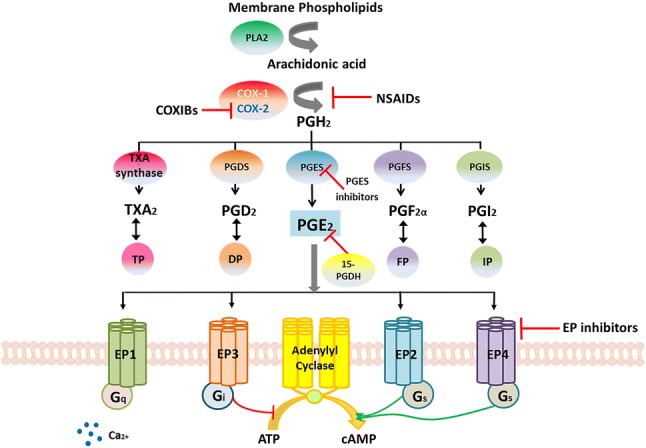
COX-2-PGE2-EPs signaling pathway. Arachidonic acid is released from the membrane phospholipids by PLA2 and then is metabolized by COX-1 and COX-2 into PGH2. PGH2 is converted by specific isomerases (PGDS, PGES, PGFS and PGIS) and TXA synthase to multiple prostaglandins (PGE2, PGD2, PGF2α, PGI2) and the thromboxane A2 [4]. Prostaglandins act through relative receptors (EP, DP, FP, IP and TP) to mediate their effects [5]. The inhibitors of COX-2-PGE2-EPs signaling pathway include nonsteroidal anti-inflammatory drugs (NSAIDs), COX-2 selective inhibitors (COXIBs), PGES inhibitor, 15-hydroxyprostaglandin dehydrogenase (15-PGDH) and EP inhibitors. NSAIDs inhibit the function of both COX-1 and COX-2 while COXIBs only inhibit the function of COX-2. PGE2 is degraded by 15-PGDH into an inactive 15-keto PGE2 after binding to EP receptors [6]. Both PGES inhibitors and EP inhibitors are novel inhibitors that have been investigating in these years. PLA2 phospholipases A2, COX-1 cyclooxygenase-1 COX-2 cyclooxygenase-2, PGDS prostaglandin D synthase, PGES prostaglandin G synthase, PGFS prostaglandin F synthase, PGIS prostaglandin I synthase, PG prostaglandin, EP prostaglandin E receptor, DP1.2 prostaglandin D receptor 1.2, FP prostaglandin F receptor, IP prostaglandin I receptor, ATP adenosine triphosphate, cAMP cyclic adenosine monophosphate
Biogenesis and signaling: COX-2-PGE2-EPs
Arachidonic acid is released from the membrane phospholipids by phospholipase A2 (PLA2) and then metabolized by the enzyme of COX-1 and COX-2 into prostaglandin H2 (PGH2). PGH2 is converted by specific isomerases (PGDS, PGES, PGFS and PGIS) and TXA synthase to various prostaglandins (PGE2, PGD2, PGF2α, PGI2) and the thromboxane A2 (TxA2) [4] (Fig. 1). All these prostaglandins (PGE2, PGD2, PGF2α, PGI2 and TXA2) act through relative specific G-protein coupled receptors (GPCR) to mediate their effects, referred to as the EP, DP, FP, IP and TP receptors [5] (Fig. 1).
COX enzymes are the primary enzymes in the synthesis of eicosanoids and exist in two isoforms: COX-1 is considered to be ubiquitously expressed [7], whereas COX-2 is expressed predominantly in inflammatory cells and upregulated in chronic and acute inflammations [8]. COX-1 and COX-2 are located on human chromosomes 9 and 1 respectively [9]. PGs produced by COX-1 are crucial for maintaining the integrity of gastric mucosa, normal platelet aggregation and renal function, while PGs derived by COX-2 contributes to cancer progression and metastasis [10]. The COX-2 expression is stimulated by different growth factors, cytokines and prostaglandins, which is associated with inflammatory response and is seen as a prognostic factor for malignancy [11, 12]. Furthermore, upregulation of COX-2 and PGE2 has been identified in many human cancers and precancerous lesions, and COX inhibitory drugs show protective effects in colorectal cancer and breast cancer [13].
The three distinct synthases contributing to PGE2 synthesis are consist of microsomal PGE synthase-1 (mPGES-1), mPGES-2 and cytosolic PGE synthase (cPGES) [14, 15]. There are two separate PGE2-biosynthetic routes: the cPLA2-COX-1-cPGES and cPLA2-COX-2-mPGES pathways [15]. COX-2 linked to mPGES is essential for delayed PGE2 biosynthesis, which may be linked to inflammation, fever, osteogenesis, and cancer [15]. mPGES-1 is primarily responsible for increasing PGE2 levels during inflammation and carcinogenesis, and elevated levels of mPGES-1 present in a number of human cancers, such as colon, lung, stomach, pancreas, cervix, prostate and head and neck squamous carcinoma [16].
PGE2 is the most abundant prostaglandin in humans and is known as a key mediator in inflammation. The functions of PGE2 are mainly facilitated by specific membrane-bound G-protein-coupled EP receptors (EP1-EP4) with various signaling pathways. EP1 is coupled to the G protein alpha q (Gq) to mobilize intracellular Ca2+, EP2 and EP4 are coupled to the G protein alpha stimulator (Gs) to activate adenylyl cyclase (AC), and EP3 is mainly coupled to the G protein alpha inhibitor (Gi) to suppress AC [17]. The EP3 receptor can also be coupled to G12/13 proteins, resulting in the activation of the small G protein Rho [18]. After binding its receptor, PGE2 can be catalyzed by 15-hydroxyprostaglandin dehydrogenase (15-PGDH) into an inactive 15-keto PGE2 [6].
In cancer development, EP1 mediates tumor cell migration, invasion and adjustment to hypoxia environment; EP2 induces angiogenesis and suppresses the anti-tumor immune response; EP4 can mediate tumor cell migration, metastasis, as well as promote aberrant DNA methylation [18]. The role of EP3 in carcinogenesis is still unclear with conflicting effects in distinct cancer cells. EP3 is a unique PGE2 receptor, since the human EP3 gene consists of ten exons and nine introns, encoding at least eight distinct EP3 splice variants [19]. EP3 isoforms differ in the amino acid sequences in their specific C-terminal tails and signal transduction pathways by activating different second messengers [20, 21]. This might increase the complexity of investigating the effects of EP3 on the pathological mechanism of cancer development. Studies concerning the COX-2-PGE2-EPs expression have been investigated recently and are summarized in this review.
Crosstalks with other signaling pathways in cancer
Wang et al. elucidated crosstalks interacting with COX-2-PGE2-EPs signaling pathways in carcinogenesis, mainly consisting of the epidermal growth factor receptor (EGFR) pathway, nuclear receptor pathway, and Ras-mitogen-activated protein kinase cascade (Ras-MAPK) pathway [7]. The classic and most studied signaling pathway is EGFR pathway since both COX-2 and PGE2 are involved in the proliferation, migration and invasion of human colon carcinoma cells through EGFR [22, 23]. Combining EGFR tyrosine kinase inhibitor (erlotinib) with COX-2 inhibitor (celecoxib) can inhibit the tumor cells proliferation of head and neck cancer cell lines and the tumor growth of nude mouse xenograft models compared with either single agent [24]. Moveover, the biomarker expression (antigen Ki67, phosphorylated S6 and CD34) of head and neck cancer is decreased in 11 cancer patients received the combined treatment with erlotinib and celecoxib [24]. PGE2 can also trans-activate peroxisome proliferator-activated receptor β/δ (PPAR β/δ) via phosphatidylinositol 3-kinase/protein kinase B (PI3K/Akt) signaling to promote cell survival of intestinal adenoma [25]. In the mice model, PGE2 can stimulate tumor growth of intestinal adenoma in Apcmin mice, but not in Apcmin mice lacking PPAR β/δ [25]. As a nuclear transcription factor, PPAR β/δ binds as heterodimers with a retinoid X receptor (RXR) for transcription initiation [26], and the natural ligands for PPAR β/δ include fatty acid and PGE2 [27]. Additionally, PGE2 activates Ras-MAPK cascade and high expression of PGE2 can induce COX-2 expression in intestinal adenomas [28]. Studies concerning the COX-2-PGE2-EPs signaling pathway are limited in gynecological cancers compared with that in gastrointestinal cancer and breast cancer [29].
Endometrial cancer
Endometrial cancer (EC) is the most common gynecological malignancy in developed countries, including the United States, Canada and Western Europe [30]. There estimated to be more than 61,000 new cases of EC and over 12,000 deaths in the United States according to the 2019 cancer statistics [31]. The main risk factor for EC is exposure to endogenous and exogenous estrogens, which is linked to obesity, diabetes, early age at menarche, null parity, late menopause and use of tamoxifen [32]. EC is classified into two subtypes: type I and type II. Type I is the most common subtype, and it is low-grade, endometrioid, diploid, hormone-receptor-positive endometrial cancer with a good prognosis [32]. By contrast, type II EC is high-grade, non-endometrioid, aneuploidy, hormone-receptor-negative, TP53-mutated with a poor prognosis and a higher risk of metastasis [32].
COX-2 and endometrial cancer
COX-2 is expressed in the cytoplasm of normal proliferative glandular epithelium and endometrial cancer cells [33]. The mRNA level of COX-2 is elevated in 51 cancerous endometria compared with 16 normal endometria [34]. COX-2 is proved to be a negative predictor of disease relapse for EC patients in the univariate analysis. COX-2 plays a key role not only in the maintenance of the endometrium during the menstrual cycle but also in EC carcinogenesis [35]. COX-2 overexpression increases angiogenesis, migration, invasiveness and tumor-induced immmuno-suppression, as well as prevents apoptosis [35]. A combined treatment with celecoxib (a COX-2 inhibitor) and rapamycin (a mammalian target of rapamycin complex 1 inhibitor, a mTORC1 inhibitor) reduces EC progression in mouse models of EC and human EC cell lines [36]. Brasky et al. demonstrated that treatment of aspirin could reduce the risk of EC, especially in estrogen-mediated cases by analyzing 22,268 female Americans after up to 10 years [37]. In clinical studies, the correlation of COX-2 expression and EC patients’ prognosis still remains conflicting [38, 39]. PGE2 is associated with both endometrial functions and disorders. Ke et al. found that prostaglandin E synthase 2 (PTGES2) is upregulated in the 119 endometrial cancer tissues compared with 50 normal endometria, and PTGES2 is associated with the endometrial carcinoma stage, grade and the depth of myometrial invasion [40].
EPs expression in endometrial cancer
Zhu et al. suggested that six patients with higher EP1 staining survived after seven years follow-up, although EP1 expression was not correlated to progression-free survival or overall survival of endometrial cancer patients [41]. More recruited EC patients’ samples might further prove the connection of EP1 expression and EC (Fig. 2). The biosynthesis of EP2, EP4 and cAMP are significantly elevated in response to PGE2 in endometrial adenocarcinoma tissues compared with normal endometria by quantitative PCR [42]. PGE2 stimulates vascular endothelial growth factor (VEGF) expression in Ishikawa cells (a human endometrial adenocarcinoma cell line) via EP2-cAMP-mediated transactivation of the epidermal growth factor receptor (EGFR) and extracellular signal-regulated kinases 1/2 (ERK1/2) pathways [43]. Battersby and his colleagues proved that PGE2 upregulates the expression of fibroblast growth factor 2 (FGF2) via the EP2 receptor in a cAMP-, c-Src-, epidermal growth factor receptor (EGFR)- and extracellular signal-regulated kinase (ERK)-dependent manner in Ishikawa cells [44]. FGF2 is a potent mitogenic and angiogenic factor, causing adenocarcinoma cell proliferation in nude mice transplanted subcutaneously with endometrial adenocarcinoma [45]. PGE2 can enhance proliferation and invasion of two human endometrial cancer cells (Ishikawa and HEC-1B) by stimulating EP4 receptor and small ubiquitin-like modifier-1 (SUMO-1) via the Wnt/β-catenin signaling pathway [40].
Fig. 2.
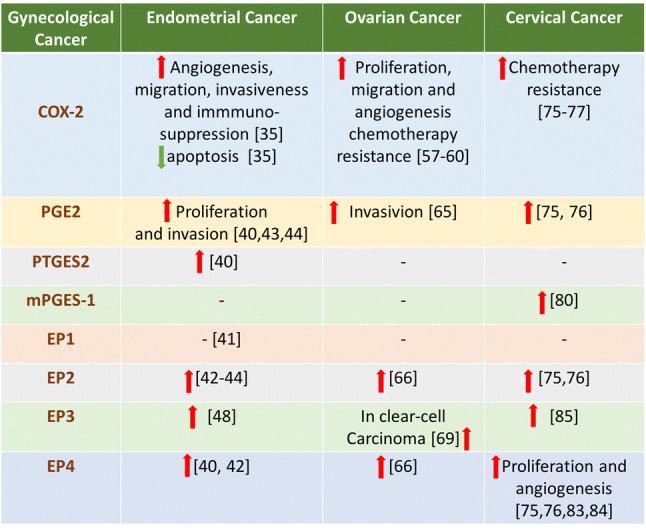
Overview the expression of COX-2-PGE2-EPs signaling in gynecological malignancies (endometrial cancer, ovarian cancer and cervical cancer). Red arrow: upregulation; green arrow:downregulation; -: uncertain
The proliferation and angiogenesis of implanted tumor can be directly inhibited in EP3−/− mice, as well as suppressed by an EP3 antagonist (ONO-AE3–240) in wild-type mice [46]. EP3 mRNA is expressed abundantly in the uterus [47]. The latest study by our research group demonstrated that high expression of EP3 correlates with poor progression-free survival and overall survival in endometrial carcinoma [48]. Our group also proved that L-798,106 (a specific EP3 antagonist) induces the expression of estrogen receptor β and inhibits the activity of Ras, leading to decreased proliferation and migration of RL95-2 cells [48]. Overexpression of estrogen receptor β (ERβ) inhibits proliferation and invasion of tumor cells in breast cancer and endometrium [49, 50].
Ovarian cancer
Ovarian cancer ranks the fifth as a cause of neoplastic death among women worldwide [30] and the first lethal gynecological malignancy [31]. There would be around 22,530 new cases and 13,980 deaths in the United States in 2019 [31]. Incidence rates are highest in more developed regions, with rates of more than 7.5 per 100,000 [30]. The overall 5-year survival rate of ovarian cancer is just approximately 30–40% [51]. The fundamental problem in treating ovarian cancer is that it is not easy to discover it at an early stage and accomplish complete curative resection. Ovarian cancer histological groups include type I epithelial, type II epithelial, germ cell, sex cord-stromal, other specific non-epithelial and non-specific tumors [52]. Among all the subtypes, type II epithelial tumors are the most common in Oceania, North America and Europe, while type I epithelial tumors are more common in Asia during 2005–2009 [52]. Type II epithelial tumors are associated with poorer survival than type I epithelial, germ cell and sex cord-stromal tumors [52].
COX and ovarian cancer
COX-2−/− female mice show defective ovulation, fertilization and implantation [53]. The mRNA expressions of COX-1, COX-2, EP2 and EP4 are detected in both granulosa and cumulus cells in mice periovulatory follicles during superovulation [54]. COX-1, COX-2, mPGES-1, EP1 and EP2 are expressed predominantly in epithelial cells of human epithelial ovarian cancer [55]. Kino et al. (2005) believed that COX-1 is the primary enzyme for producing PGE2 instead of COX-2 in ovarian cancer cells. Because the elevated expression of COX-1 instead of COX-2 was detected in 22 ovarian cancer tissues compared with that in normal cases [56], and SC-560 (a COX-1 inhibitor) can suppress the production of PGE2 in three ovarian cancer cell lines while NS-398 and rofecoxib (COX-2 inhibitors) can not [56]. However, the latest meta-analysis (2017) suggested that COX-2 expression is correlated with FIGO stage, histological type and patients’ age and the high expression of COX-2 is associated with reduced OS and DFS of ovarian cancer patients [57]. In addition, high expression of COX-2 is associated with a shorter progression time and overall survival time in the patients who firstly underwent explorative laparotomy and then received chemotherapy [58]. It implies that COX-2 is also correlated with chemotherapy resistance.
For in vitro studies, COX-2 can stimulate the proliferation, migration and angiogenesis of ovarian cancer cells. COX-2 enhances proliferation and migration of human ovarian cancer CAOV-3 cells mainly through the activation of phosphatidylinositol 3-kinase/protein kinase B (PI3-k/Akt) pathway [59]. By analyzing the epithelial ovarian cancer (EOC) tissues and EOC cell lines (MDAH2274 and SKOV3), Uddin and his colleagues demonstrated that COX-2 modulates cell growth and apoptosis also through PI3K/AKT signaling pathway in EOC [60]. Furthermore, Uddin et al. proved that COX-2 could be a potential therapeutic target in EOC because treatment of xenografts together with aspirin can inhibit tumor growth in nude mice through decreasing the expression of COX-2 and Akt [60]. COX-2 protein levels correlate with VEGF protein levels and microvessel counts in ovarian carcinoma [61].
The expression of COX-2 is regulated by various cytokines in ovarian cancer cells, such EGF, vitamin D, IL-1β. EGF induces the production of both COX-2 and PGE2 via the activation of the PI3K/Akt signaling pathway, resulting in an invasion of SKOV3 and OVCAR5 cells (two human ovarian cancer cell lines) [62]. A reduced level of vitamin D receptor (VDR) and an induced level of COX-2, 15-PGDH and PGE2 are found in the serum of ovarian cancer patients older than 45 years [63], suggesting an interaction between PG and vitamin D-metabolism in ovarian cancer. The mRNA and protein expression of COX-2 can be stimulated by IL-1β and phorbol ester (TPA) in OVCAR-3 cells and by TPA in CAOV-3 cells [64].
EPs and ovarian cancer
PGE2 can induce cell invasiveness via increasing the expression of MMP-2 and MMP-9 in two human ovarian cancer cell lines (CaOV-3 and SKOV-3) [65]. PGE2 stimulates the VEGF production in HEY ovarian cancer cells mainly via activating EP2 and EP4 receptors, which can be reversed by AH23848 (an antagonist of both EP2 and EP4) [66]. In addition, PGE2-induced EP4 receptor signaling induces MMP production and ovarian cancer cell invasion through Src-mediated EGFR transactivation [66]. EP2 plays a vital role in the process of ovulation and fertilization because cumulus expansion becomes decreased in EP2−/− mice [67]. By feeding hens 10% flaxseed-enriched or standard diet for four years, Eilati et al. proved a decreased expression of COX-2 and PGE2, as well as a reduction in ovarian cancer severity and incidence [68]. Our research group (2019) has recently demonstrated that EP3 expression was higher in clear-cell carcinoma compared to three other histological subtypes of ovarian cancer [69]. Moreover, EP3 negative patients showed longer overall survival in a subgroup with negative expression of a tumor-specific epitope of tumor-associated epithelial mucin 1 (MUC1) [69].
Cervical cancer
Cervical cancer is the fourth most common cancer and the fourth most common cause of cancer-related death among women worldwide [70]. The incidence and prevalence of cervical cancer are higher in the developing countries than in the developed countries owing to a lack of screening, availability of vaccine, and awareness of HPV infections [71]. In the United States in 2019, there were an estimated 13,170 cases and 4250 deaths from cervical cancer [31]. In many developing countries, cervical cancer causes more than one-quarter of a million deaths per year [72]. The 5-year survival rate of cervical cancer is between 63–79% in China, Singapore, South Korea and Turkey, less than 25% in Gambia and Uganda [73]. The two main malignant epithelial cervical cancer types are the squamous cell carcinoma and the adenocarcinoma [74].
COX-2 and cervical cancer
High expressions of COX-2 and PGE2 have been found in the cervical carcinoma [75, 76]. COX-2 is overexpressed in various types of cervical neoplasm such as cervical intraepithelial neoplasia (CIN), adenocarcinoma and squamous cell carcinoma, implying that COX-2 expression is highly associated with cervical carcinoma development and progression [10]. Many studies show that COX-2 contributes to carcinogenesis and progression of cervical cancer. High expression of COX-2 is related to poor overall survival (OS) and poor disease-free survival (DFS) in cervical cancer patients [77]. COX-2 is also associated with poor DFS in chemo-radiation subgroup, implying COX-2 is a chemo-radiation resistance predictive factor for cervical cancer [77]. The COX-2 expression is more frequently expressed in adenocarcinoma than in squamous cell carcinoma by immunohistochemistry [78]. The co-expression of COX-2 and thymidine phosphorylase (TP) is related to poor 5-year disease-free and overall survival rates, suggesting that the combination of COX-2 and TP is a prognosticator for squamous cell carcinoma of the cervical cancer [79].
The expression of mPGES-1 is higher in squamous intraepithelial lesions and carcinoma of the uterine cervix compared with the normal cervical epithelium [80]. Radilova et al. found that COX-1 is also coupled with mPGES-1 for co-regulating PGE2 synthesis in human cervix cancer cells [81]. Dimethylcelecoxib (a non-COX-2 inhibitor) inhibits the early growth response protein 1 (EGR1) and transcription of mGPES-1 via an enhanced complex of NF-κB and histone deacetylase 1 (HDAC1) that binds to the EGR1 promoter in Hela cells [82].
EPs and cervical cancer
Sales et al. reported that the syntheses of COX-2, PGE2, EP2, EP4 and cyclic adenosine monophosphate (cAMP) are up-regulated in cervical cancer tissue compared to that in the healthy cervix, suggesting that PGE2 may regulate neoplastic cell function via the EP2/EP4 receptors [75]. Sales and his colleagues further in 2002 proved that PGE2 could induce the expression of COX-2, EP4 and cAMP in Hela cells which were transiently transfected with EP2 or EP4 cDNA [76]. However, this research did not detect whether the expression of cAMP would be decreased after knocking out or knocking down the expression of EP2 or EP4. Kurt et al. observed that rapid accumulation of cAMP is produced in Hela cells after being stimulated with PGE2, which is mediated via the cAMP-linked EP2/EP4 receptors [76]. Both studies imply that PGE2 regulates the function of cervical cancer cells mainly via cAMP-linked EP2/EP4 signaling pathway.
Jung-Min et al. showed an increased expression of EP4 in 52 cervical cancer tissues compared with four healthy controls by immunohistochemistry [83]. This study also demonstrated that HPV16 E5 upregulated the activity of PGE2-EP4-cAMP signaling pathways by inducing the binding of cyclic adenosine monophosphate response element-binding protein (CREB) to a variant CRE site in the promoter of the human EP4 gene [83]. EP4 plays a role in the proliferation and angiogenesis of cervical cancer cells since GW627368X (a highly selective EP4 antagonist) inhibits the proliferation and angiogenesis of cervical carcinoma by blocking EP4/EGFR signaling pathway in cervical cancer cell lines (HeLa, SiHa and ME 180) and suppresses the tumor size in xenograft mice model [84].
Our group recently proved that overexpression of EP3 in cervical cancer patients is associated with impaired prognosis in overall survival rates when evaluating 250 cervical cancer patients with immunohistochemistry [85]. As an independent prognosticator for cervical carcinoma, the EP3 receptor is also significantly correlated with lymph node stage and FIGO stage [85]. However, the pathological mechanism of how EP3 signaling regulates in cervical cancer is still unclear.
Drug targeting of COX-2-PGE2-EPs signaling
Chemoprevention has long been recognized as an important prophylactic strategy to reduce the burden of cancer on the health care system. In addition, nonsteroidal anti-inflammatory drugs (NSAIDs) as chemoprevention chemicals have been proved to reduce the risk of several cancers in human, such as gastrointestinal cancer, breast cancer, prostate cancer, lung cancer and skin cancer [86]. Nan et al. (2015) found that regular use of aspirin or NSAIDs is linked to lower risk of colorectal cancer compared with no regular use after analyzing 8634 colorectal cancer cases and 8553 matched controls between 1976 and 2011 [87]. Long-term use of COX inhibitors in humans leads to a 50% decrease in risk for colorectal cancer [25]. A meta-analysis by Banndrup et al. suggested the risk of invasive ovarian cancer is significantly reduced with the use of aspirin [88].
NSAIDs include aspirin, ibuprofen and naproxen, and act by inhibiting both COX-1 and COX-2. These unspecific inhibitors cause many adverse effects, such as gastrointestinal ulcers and bleeds, heart attack and kidney disease [89]. Selective COX-2 inhibitors (COXibs) has been successfully documented and showed less toxicity to gastrointestinal tract as compared to traditional NSAIDs [90]. However, the long-term use of COX-2 selective inhibitors still has other side effects. The adverse effects of COX-2 selective and non-selective inhibitors are summarized in the latest review by Rayar et al. including myocardial infarction, hypertension, stroke, reduced glomerular filtration rate and renal plasma flow, acute renal failure, acute interstitial nephritis, inhibition of ulcer healing, hepatic complications, allergy, fatal skin reaction, depression, delayed follicular rupture and so on [91]. Therefore, further exploration of novel anti-inflammation targets is needed.
Clinical studies show elevated levels of mPGES-1 are identified in colon, lung, stomach, pancreas, cervix, prostate, papillary thyroid carcinoma, head and neck squmaous carcinoma and brain tumors, suggesting mPGES-1 inhibitors might be a potential chemopreventive agent [16]. However, a limited number of compounds that inhibiting mPGES-1 has not been successfully developed as anti-cancer agents, such as celecoxib, MF-63, NS-398, MK-866 and triclosan [16].
In recent years, extensive efforts have been made into elucidating the function of PGE2 and the EP receptors in health and carcinogenesis with the aim of exploring promising targets and selective inhibitors as a novel therapy. Many researchers have found the strong correlation of EP2/EP4 receptors with colon cancer, skin cancer, mucosa cancer of the pharynx and the esophagus, prostate cancer, urothelial cancer and non-small cell lung cancer [18]. The EP4 promotes migration, invasion, angiogenesis and lymphangiogenesis of mammary tumor cells [92]. EP4 receptor is responsible for the PGE2-induced colorectal tumor cell proliferation and morphogenic changes via PI3k/Akt signaling pathway [93]. Mice are not able to have inflammatory responses to PGE2, IL-1β or lipopolysaccharide when lacking the EP3 receptor, but not EP1, EP2 or EP4 receptor [94]. Deletion or inhibition of EP3 receptors could ameliorate the neuronal apoptosis in the ischemic cortex in EP3 knock-out mice or EP3 antagonist-treated mice compared with wild-type mice or vehicle-treated mice, respectively. It suggests that EP3 is involved in the inflammatory and apoptotic reactions during stroke injury [95]. The investigations concerning EP receptors in gynecological cancers highlight the potential advantage of combining COX enzyme inhibitors with EP receptor antagonists as therapeutic agents in gynecological cancers.
Wang et al. proposed the possible PGE2 downstream targets that might also serve as promising specific chemopreventive agents for cancer prevention and treatment, which include angiogenic factors (VEGF, bFGF), anti-apoptotic factors (Bcal-2), chemokines (MIP-1α, MIP-1β, RANTES, CXCR4) and their receptors, and immunosuppressive mediators [7].
Conclusions
Studies show that COX-2 is a negative predicator of EC, ovarian cancer and cervical cancer, contributing to carcinogenesis and progression of gynecological cancer. The expression of mPGES-1 is highly expressed in human epithelial ovarian cancer and cervical cancer, while PTGES2 is associated with the endometrial carcinoma carcinogenesis. Many studies focus the function of cAMP-linked EP2/EP4 signaling pathway in EC, ovarian cancer and cervical cancer, while roles of EP1 and EP3 have not been completely clarified. Further investigations concerning EP receptors in gynecological cancers are necessary. EPs may represent novel and specific anti-inflammation targets for gynecological cancer chemoprevention. It might highlight the potential advantage of combining COX enzyme inhibitors with EP receptor antagonists as therapeutic agents in gynecological cancer.
Acknowledgements
Open Access funding provided by Projekt DEAL. China Scholarship Council supported Yao Ye scholarships. The sponsors did not participate in the manuscript writing.
Author contributions
YY and XPW: literature searching, data analysis and manuscript writing. UJ and VvS: MANUSCRIPT editing and revising.
Compliance with ethical standards
Conflict of interest
All authors declare no conflict of interest.
Informed consent
Consent statement is not applicable for this review manuscript.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Xipeng Wang, Email: wangxipeng@xinhuamed.com.cn.
Udo Jeschke, Email: Udo.Jeschke@med.uni-muenchen.de.
References
- 1.Kuper H, Adami HO, Trichopoulos D. Infections as a major preventable cause of human cancer. J Intern Med. 2000;248(3):171–183. doi: 10.1046/j.1365-2796.2000.00742.x. [DOI] [PubMed] [Google Scholar]
- 2.Aggarwal BB, Shishodia S, Sandur SK, Pandey MK, Sethi G. Inflammation and cancer: how hot is the link? Biochem Pharmacol. 2006;72(11):1605–1621. doi: 10.1016/j.bcp.2006.06.029. [DOI] [PubMed] [Google Scholar]
- 3.Dossus L, Rinaldi S, Becker S, Lukanova A, Tjonneland A, Olsen A, Stegger J, Overvad K, Chabbert-Buffet N, Jimenez-Corona A, Clavel-Chapelon F, Rohrmann S, Teucher B, Boeing H, Schutze M, Trichopoulou A, Benetou V, Lagiou P, Palli D, Berrino F, Panico S, Tumino R, Sacerdote C, Redondo ML, Travier N, Sanchez MJ, Altzibar JM, Chirlaque MD, Ardanaz E, Bueno-de-Mesquita HB, van Duijnhoven FJ, Onland-Moret NC, Peeters PH, Hallmans G, Lundin E, Khaw KT, Wareham N, Allen N, Key TJ, Slimani N, Hainaut P, Romaguera D, Norat T, Riboli E, Kaaks R. Obesity, inflammatory markers, and endometrial cancer risk: a prospective case-control study. Endocr Relat Cancer. 2010;17(4):1007–1019. doi: 10.1677/ERC-10-0053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Simmons DL, Botting RM, Hla T. Cyclooxygenase isozymes: the biology of prostaglandin synthesis and inhibition. Pharmacol Rev. 2004;56(3):387–437. doi: 10.1124/pr.56.3.3. [DOI] [PubMed] [Google Scholar]
- 5.Coleman RA, Smith WL, Narumiya S. International Union of Pharmacology classification of prostanoid receptors: properties, distribution, and structure of the receptors and their subtypes. Pharmacol Rev. 1994;46(2):205–229. [PubMed] [Google Scholar]
- 6.Legler DF, Bruckner M, Uetz-von Allmen E, Krause P. Prostaglandin E2 at new glance: novel insights in functional diversity offer therapeutic chances. Int J Biochem Cell Biol. 2010;42(2):198–201. doi: 10.1016/j.biocel.2009.09.015. [DOI] [PubMed] [Google Scholar]
- 7.Wang D, Dubois RN. Prostaglandins and cancer. Gut. 2006;55(1):115–122. doi: 10.1136/gut.2004.047100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ferrer MD, Busquests-Cortes C, Capo X, Tejada S, Tur JA, Pons A, Sureda A. Cyclooxygenase-2 inhibitors as a therapeutic target in inflammatory diseases. Curr Med Chem. 2018 doi: 10.2174/0929867325666180514112124. [DOI] [PubMed] [Google Scholar]
- 9.Tay A, Squire JA, Goldberg H, Skorecki K. Assignment of the human prostaglandin-endoperoxide synthase 2 (PTGS2) gene to 1q25 by fluorescence in situ hybridization. Genomics. 1994;23(3):718–719. doi: 10.1006/geno.1994.1569. [DOI] [PubMed] [Google Scholar]
- 10.Kim HS, Kim T, Kim MK, Suh DH, Chung HH, Song YS. Cyclooxygenase-1 and -2: molecular targets for cervical neoplasia. J Cancer Prev. 2013;18(2):123–134. doi: 10.15430/JCP.2013.18.2.123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.O'Banion MK, Winn VD, Young DA. cDNA cloning and functional activity of a glucocorticoid-regulated inflammatory cyclooxygenase. Proc Natl Acad Sci USA. 1992;89(11):4888–4892. doi: 10.1073/pnas.89.11.4888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Williams CS, DuBois RN. Prostaglandin endoperoxide synthase: why two isoforms? Am J Physiol. 1996;270(3 Pt 1):G393–400. doi: 10.1152/ajpgi.1996.270.3.G393. [DOI] [PubMed] [Google Scholar]
- 13.Howe LR. Inflammation and breast cancer. Cyclooxygenase/prostaglandin signaling and breast cancer. Breast Cancer Res. 2007;9(4):210. doi: 10.1186/bcr1678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jakobsson PJ, Thoren S, Morgenstern R, Samuelsson B. Identification of human prostaglandin E synthase: a microsomal, glutathione-dependent, inducible enzyme, constituting a potential novel drug target. Proc Natl Acad Sci USA. 1999;96(13):7220–7225. doi: 10.1073/pnas.96.13.7220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Murakami M, Naraba H, Tanioka T, Semmyo N, Nakatani Y, Kojima F, Ikeda T, Fueki M, Ueno A, Oh S, Kudo I. Regulation of prostaglandin E2 biosynthesis by inducible membrane-associated prostaglandin E2 synthase that acts in concert with cyclooxygenase-2. J Biol Chem. 2000;275(42):32783–32792. doi: 10.1074/jbc.M003505200. [DOI] [PubMed] [Google Scholar]
- 16.Nakanishi M, Gokhale V, Meuillet EJ, Rosenberg DW. mPGES-1 as a target for cancer suppression: a comprehensive invited review "Phospholipase A2 and lipid mediators". Biochimie. 2010;92(6):660–664. doi: 10.1016/j.biochi.2010.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sokolowska M, Chen LY, Liu Y, Martinez-Anton A, Qi HY, Logun C, Alsaaty S, Park YH, Kastner DL, Chae JJ, Shelhamer JH. Prostaglandin E2 inhibits NLRP3 inflammasome activation through EP4 receptor and intracellular cyclic AMP in human macrophages. J Immunol. 2015;194(11):5472–5487. doi: 10.4049/jimmunol.1401343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.O'Callaghan G, Houston A. Prostaglandin E2 and the EP receptors in malignancy: possible therapeutic targets? Br J Pharmacol. 2015;172(22):5239–5250. doi: 10.1111/bph.13331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kotani M, Tanaka I, Ogawa Y, Usui T, Tamura N, Mori K, Narumiya S, Yoshimi T, Nakao K. Structural organization of the human prostaglandin EP3 receptor subtype gene (PTGER3) Genomics. 1997;40(3):425–434. doi: 10.1006/geno.1996.4585. [DOI] [PubMed] [Google Scholar]
- 20.Namba T, Sugimoto Y, Negishi M, Irie A, Ushikubi F, Kakizuka A, Ito S, Ichikawa A, Narumiya S. Alternative splicing of C-terminal tail of prostaglandin E receptor subtype EP3 determines G-protein specificity. Nature. 1993;365(6442):166–170. doi: 10.1038/365166a0. [DOI] [PubMed] [Google Scholar]
- 21.Kotani M, Tanaka I, Ogawa Y, Suganami T, Matsumoto T, Muro S, Yamamoto Y, Sugawara A, Yoshimasa Y, Sagawa N, Narumiya S, Nakao K. Multiple signal transduction pathways through two prostaglandin E receptor EP3 subtype isoforms expressed in human uterus. J Clin Endocrinol Metab. 2000;85(11):4315–4322. doi: 10.1210/jcem.85.11.6989. [DOI] [PubMed] [Google Scholar]
- 22.Yoshimoto T, Takahashi Y, Kinoshita T, Sakashita T, Inoue H, Tanabe T. Growth stimulation and epidermal growth factor receptor induction in cyclooxygenase-overexpressing human colon carcinoma cells. Adv Exp Med Biol. 2002;507:403–407. doi: 10.1007/978-1-4615-0193-0_62. [DOI] [PubMed] [Google Scholar]
- 23.Buchanan FG, Wang D, Bargiacchi F, DuBois RN. Prostaglandin E2 regulates cell migration via the intracellular activation of the epidermal growth factor receptor. J Biol Chem. 2003;278(37):35451–35457. doi: 10.1074/jbc.M302474200. [DOI] [PubMed] [Google Scholar]
- 24.Shin DM, Zhang H, Saba NF, Chen AY, Nannapaneni S, Amin AR, Muller S, Lewis M, Sica G, Kono S, Brandes JC, Grist WJ, Moreno-Williams R, Beitler JJ, Thomas SM, Chen Z, Shin HJ, Grandis JR, Khuri FR, Chen ZG. Chemoprevention of head and neck cancer by simultaneous blocking of epidermal growth factor receptor and cyclooxygenase-2 signaling pathways: preclinical and clinical studies. Clin Cancer Res. 2013;19(5):1244–1256. doi: 10.1158/1078-0432.CCR-12-3149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wang D, Wang H, Shi Q, Katkuri S, Walhi W, Desvergne B, Das SK, Dey SK, DuBois RN. Prostaglandin E(2) promotes colorectal adenoma growth via transactivation of the nuclear peroxisome proliferator-activated receptor delta. Cancer Cell. 2004;6(3):285–295. doi: 10.1016/j.ccr.2004.08.011. [DOI] [PubMed] [Google Scholar]
- 26.Berger J, Moller DE. The mechanisms of action of PPARs. Annu Rev Med. 2002;53:409–435. doi: 10.1146/annurev.med.53.082901.104018. [DOI] [PubMed] [Google Scholar]
- 27.Pestka A, Fitzgerald JS, Toth B, Markert UR, Jeschke U. Nuclear hormone receptors and female reproduction. Curr Mol Med. 2013;13(7):1066–1078. doi: 10.2174/1566524011313070002. [DOI] [PubMed] [Google Scholar]
- 28.Wang D, Buchanan FG, Wang H, Dey SK, DuBois RN. Prostaglandin E2 enhances intestinal adenoma growth via activation of the Ras-mitogen-activated protein kinase cascade. Cancer Res. 2005;65(5):1822–1829. doi: 10.1158/0008-5472.CAN-04-3671. [DOI] [PubMed] [Google Scholar]
- 29.Nakanishi M, Rosenberg DW. Multifaceted roles of PGE2 in inflammation and cancer. Semin Immunopathol. 2013;35(2):123–137. doi: 10.1007/s00281-012-0342-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, Parkin DM, Forman D, Bray F. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer. 2015;136(5):E359–386. doi: 10.1002/ijc.29210. [DOI] [PubMed] [Google Scholar]
- 31.Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin. 2019;69(1):7–34. doi: 10.3322/caac.21551. [DOI] [PubMed] [Google Scholar]
- 32.Morice P, Leary A, Creutzberg C, Abu-Rustum N, Darai E. Endometrial cancer. Lancet. 2016;387(10023):1094–1108. doi: 10.1016/S0140-6736(15)00130-0. [DOI] [PubMed] [Google Scholar]
- 33.Uotila PJ, Erkkola RU, Klemi PJ. The expression of cyclooxygenase-1 and -2 in proliferative endometrium and endometrial adenocarcinoma. Ann Med. 2002;34(6):428–433. doi: 10.1080/078538902321012379. [DOI] [PubMed] [Google Scholar]
- 34.Jarzabek K, Koda M, Walentowicz-Sadlecka M, Grabiec M, Laudanski P, Wolczynski S. Altered expression of ERs, aromatase, and COX2 connected to estrogen action in type 1 endometrial cancer biology. Tumour Biol. 2013;34(6):4007–4016. doi: 10.1007/s13277-013-0991-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ohno S, Ohno Y, Suzuki N, Inagawa H, Kohchi C, Soma G, Inoue M. Multiple roles of cyclooxygenase-2 in endometrial cancer. Anticancer Res. 2005;25(6A):3679–3687. [PubMed] [Google Scholar]
- 36.Daikoku T, Terakawa J, Hossain MM, Yoshie M, Cappelletti M, Yang P, Ellenson LH, Dey SK. Mammalian target of rapamycin complex 1 and cyclooxygenase 2 pathways cooperatively exacerbate endometrial cancer. Am J Pathol. 2014;184(9):2390–2402. doi: 10.1016/j.ajpath.2014.05.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Brasky TM, Moysich KB, Cohn DE, White E. Non-steroidal anti-inflammatory drugs and endometrial cancer risk in the VITamins And Lifestyle (VITAL) cohort. Gynecol Oncol. 2013;128(1):113–119. doi: 10.1016/j.ygyno.2012.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Menczer J, Schreiber L, Berger E, Levy T. The effect of cyclooxygenase-2 expression in uterine carcinosarcoma on survival: a reassessment based on mature data. Int J Gynecol Cancer. 2015;25(8):1415–1417. doi: 10.1097/IGC.0000000000000527. [DOI] [PubMed] [Google Scholar]
- 39.Cai S, Zhang YX, Han K, Ding YQ. Expressions and clinical significance of COX-2, VEGF-C, and EFGR in endometrial carcinoma. Arch Gynecol Obstet. 2017;296(1):93–98. doi: 10.1007/s00404-017-4386-9. [DOI] [PubMed] [Google Scholar]
- 40.Ke J, Yang Y, Che Q, Jiang F, Wang H, Chen Z, Zhu M, Tong H, Zhang H, Yan X, Wang X, Wang F, Liu Y, Dai C, Wan X. Prostaglandin E2 (PGE2) promotes proliferation and invasion by enhancing SUMO-1 activity via EP4 receptor in endometrial cancer. Tumour Biol. 2016;37(9):12203–12211. doi: 10.1007/s13277-016-5087-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zhu J, Mayr D, Kuhn C, Mahner S, Jeschke U, von Schonfeldt V. Prostaglandin E2 receptor EP1 in healthy and diseased human endometrium. Histochem Cell Biol. 2018;149(2):153–160. doi: 10.1007/s00418-017-1616-y. [DOI] [PubMed] [Google Scholar]
- 42.Jabbour HN, Milne SA, Williams AR, Anderson RA, Boddy SC. Expression of COX-2 and PGE synthase and synthesis of PGE(2)in endometrial adenocarcinoma: a possible autocrine/paracrine regulation of neoplastic cell function via EP2/EP4 receptors. Br J Cancer. 2001;85(7):1023–1031. doi: 10.1054/bjoc.2001.2033. [DOI] [PubMed] [Google Scholar]
- 43.Sales KJ, Maudsley S, Jabbour HN. Elevated prostaglandin EP2 receptor in endometrial adenocarcinoma cells promotes vascular endothelial growth factor expression via cyclic 3',5'-adenosine monophosphate-mediated transactivation of the epidermal growth factor receptor and extracellular signal-regulated kinase 1/2 signaling pathways. Mol Endocrinol. 2004;18(6):1533–1545. doi: 10.1210/me.2004-0022. [DOI] [PubMed] [Google Scholar]
- 44.Battersby S, Sales KJ, Williams AR, Anderson RA, Gardner S, Jabbour HN. Seminal plasma and prostaglandin E2 up-regulate fibroblast growth factor 2 expression in endometrial adenocarcinoma cells via E-series prostanoid-2 receptor-mediated transactivation of the epidermal growth factor receptor and extracellular signal-regulated kinase pathway. Hum Reprod. 2007;22(1):36–44. doi: 10.1093/humrep/del328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Giavazzi R, Sennino B, Coltrini D, Garofalo A, Dossi R, Ronca R, Tosatti MP, Presta M. Distinct role of fibroblast growth factor-2 and vascular endothelial growth factor on tumor growth and angiogenesis. Am J Pathol. 2003;162(6):1913–1926. doi: 10.1016/S0002-9440(10)64325-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Amano H, Hayashi I, Endo H, Kitasato H, Yamashina S, Maruyama T, Kobayashi M, Satoh K, Narita M, Sugimoto Y, Murata T, Yoshimura H, Narumiya S, Majima M. Host prostaglandin E(2)-EP3 signaling regulates tumor-associated angiogenesis and tumor growth. J Exp Med. 2003;197(2):221–232. doi: 10.1084/jem.20021408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kotani M, Tanaka I, Ogawa Y, Usui T, Mori K, Ichikawa A, Narumiya S, Yoshimi T, Nakao K. Molecular cloning and expression of multiple isoforms of human prostaglandin E receptor EP3 subtype generated by alternative messenger RNA splicing: multiple second messenger systems and tissue-specific distributions. Mol Pharmacol. 1995;48(5):869–879. [PubMed] [Google Scholar]
- 48.Zhu J, Trillsch F, Mayr D, Kuhn C, Rahmeh M, Hofmann S, Vogel M, Mahner S, Jeschke U, von Schonfeldt V. Prostaglandin receptor EP3 regulates cell proliferation and migration with impact on survival of endometrial cancer patients. Oncotarget. 2018;9(1):982–994. doi: 10.18632/oncotarget.23140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Ma L, Liu Y, Geng C, Qi X, Jiang J. Estrogen receptor beta inhibits estradiol-induced proliferation and migration of MCF-7 cells through regulation of mitofusin 2. Int J Oncol. 2013;42(6):1993–2000. doi: 10.3892/ijo.2013.1903. [DOI] [PubMed] [Google Scholar]
- 50.Fatima I, Saxena R, Kharkwal G, Hussain MK, Yadav N, Hajela K, Sankhwar PL, Dwivedi A. The anti-proliferative effect of 2-[piperidinoethoxyphenyl]-3-[4-hydroxyphenyl]-2H-benzo(b) pyran is potentiated via induction of estrogen receptor beta and p21 in human endometrial adenocarcinoma cells. J Steroid Biochem Mol Biol. 2013;138:123–131. doi: 10.1016/j.jsbmb.2013.04.005. [DOI] [PubMed] [Google Scholar]
- 51.Allemani C, Weir HK, Carreira H, Harewood R, Spika D, Wang XS, Bannon F, Ahn JV, Johnson CJ, Bonaventure A, Marcos-Gragera R, Stiller C, Azevedo e Silva G, Chen WQ, Ogunbiyi OJ, Rachet B, Soeberg MJ, You H, Matsuda T, Bielska-Lasota M, Storm H, Tucker TC, Coleman MP, Group CW. Global surveillance of cancer survival 1995–2009: analysis of individual data for 25,676,887 patients from 279 population-based registries in 67 countries (CONCORD-2) Lancet. 2015;385(9972):977–1010. doi: 10.1016/S0140-6736(14)62038-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Matz M, Coleman MP, Sant M, Chirlaque MD, Visser O, Gore M, Allemani C, The CWG The histology of ovarian cancer: worldwide distribution and implications for international survival comparisons (CONCORD-2) Gynecol Oncol. 2017;144(2):405–413. doi: 10.1016/j.ygyno.2016.10.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Lim H, Paria BC, Das SK, Dinchuk JE, Langenbach R, Trzaskos JM, Dey SK. Multiple female reproductive failures in cyclooxygenase 2-deficient mice. Cell. 1997;91(2):197–208. doi: 10.1016/S0092-8674(00)80402-X. [DOI] [PubMed] [Google Scholar]
- 54.Segi E, Haraguchi K, Sugimoto Y, Tsuji M, Tsunekawa H, Tamba S, Tsuboi K, Tanaka S, Ichikawa A. Expression of messenger RNA for prostaglandin E receptor subtypes EP4/EP2 and cyclooxygenase isozymes in mouse periovulatory follicles and oviducts during superovulation. Biol Reprod. 2003;68(3):804–811. doi: 10.1095/biolreprod.102.003590. [DOI] [PubMed] [Google Scholar]
- 55.Rask K, Zhu Y, Wang W, Hedin L, Sundfeldt K. Ovarian epithelial cancer: a role for PGE2-synthesis and signalling in malignant transformation and progression. Mol Cancer. 2006;5:62. doi: 10.1186/1476-4598-5-62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kino Y, Kojima F, Kiguchi K, Igarashi R, Ishizuka B, Kawai S. Prostaglandin E2 production in ovarian cancer cell lines is regulated by cyclooxygenase-1, not cyclooxygenase-2. Prostaglandins Leukot Essent Fatty Acids. 2005;73(2):103–111. doi: 10.1016/j.plefa.2005.04.014. [DOI] [PubMed] [Google Scholar]
- 57.Sun H, Zhang X, Sun D, Jia X, Xu L, Qiao Y, Jin Y. COX-2 expression in ovarian cancer: an updated meta-analysis. Oncotarget. 2017;8(50):88152–88162. doi: 10.18632/oncotarget.21538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ferrandina G, Lauriola L, Zannoni GF, Fagotti A, Fanfani F, Legge F, Maggiano N, Gessi M, Mancuso S, Ranelletti FO, Scambia G. Increased cyclooxygenase-2 (COX-2) expression is associated with chemotherapy resistance and outcome in ovarian cancer patients. Ann Oncol. 2002;13(8):1205–1211. doi: 10.1093/annonc/mdf207. [DOI] [PubMed] [Google Scholar]
- 59.Gu P, Su Y, Guo S, Teng L, Xu Y, Qi J, Gong H, Cai Y. Over-expression of COX-2 induces human ovarian cancer cells (CAOV-3) viability, migration and proliferation in association with PI3-k/Akt activation. Cancer Invest. 2008;26(8):822–829. doi: 10.1080/07357900801941860. [DOI] [PubMed] [Google Scholar]
- 60.Uddin S, Ahmed M, Hussain A, Assad L, Al-Dayel F, Bavi P, Al-Kuraya KS, Munkarah A. Cyclooxygenase-2 inhibition inhibits PI3K/AKT kinase activity in epithelial ovarian cancer. Int J Cancer. 2010;126(2):382–394. doi: 10.1002/ijc.24757. [DOI] [PubMed] [Google Scholar]
- 61.Fujimoto J, Toyoki H, Sakaguchi H, Jahan I, Alam SM, Tamaya T. Clinical implications of expression of cyclooxygenase-2 related to angiogenesis in ovarian cancer. Oncol Rep. 2006;15(1):21–25. [PubMed] [Google Scholar]
- 62.Qiu X, Cheng JC, Chang HM, Leung PC. COX2 and PGE2 mediate EGF-induced E-cadherin-independent human ovarian cancer cell invasion. Endocr Relat Cancer. 2014;21(4):533–543. doi: 10.1530/ERC-13-0450. [DOI] [PubMed] [Google Scholar]
- 63.Thill M, Fischer D, Kelling K, Hoellen F, Dittmer C, Hornemann A, Salehin D, Diedrich K, Friedrich M, Becker S. Expression of vitamin D receptor (VDR), cyclooxygenase-2 (COX-2) and 15-hydroxyprostaglandin dehydrogenase (15-PGDH) in benign and malignant ovarian tissue and 25-hydroxycholecalciferol (25(OH2)D3) and prostaglandin E2 (PGE2) serum level in ovarian cancer patients. J Steroid Biochem Mol Biol. 2010;121(1–2):387–390. doi: 10.1016/j.jsbmb.2010.03.049. [DOI] [PubMed] [Google Scholar]
- 64.Denkert C, Kobel M, Pest S, Koch I, Berger S, Schwabe M, Siegert A, Reles A, Klosterhalfen B, Hauptmann S. Expression of cyclooxygenase 2 is an independent prognostic factor in human ovarian carcinoma. Am J Pathol. 2002;160(3):893–903. doi: 10.1016/S0002-9440(10)64912-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lau MT, Wong AS, Leung PC. Gonadotropins induce tumor cell migration and invasion by increasing cyclooxygenases expression and prostaglandin E(2) production in human ovarian cancer cells. Endocrinology. 2010;151(7):2985–2993. doi: 10.1210/en.2009-1318. [DOI] [PubMed] [Google Scholar]
- 66.Spinella F, Rosano L, Di Castro V, Natali PG, Bagnato A. Endothelin-1-induced prostaglandin E2-EP2, EP4 signaling regulates vascular endothelial growth factor production and ovarian carcinoma cell invasion. J Biol Chem. 2004;279(45):46700–46705. doi: 10.1074/jbc.M408584200. [DOI] [PubMed] [Google Scholar]
- 67.Hizaki H, Segi E, Sugimoto Y, Hirose M, Saji T, Ushikubi F, Matsuoka T, Noda Y, Tanaka T, Yoshida N, Narumiya S, Ichikawa A. Abortive expansion of the cumulus and impaired fertility in mice lacking the prostaglandin E receptor subtype EP(2) Proc Natl Acad Sci USA. 1999;96(18):10501–10506. doi: 10.1073/pnas.96.18.10501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Eilati E, Bahr JM, Hales DB. Long term consumption of flaxseed enriched diet decreased ovarian cancer incidence and prostaglandin E(2)in hens. Gynecol Oncol. 2013;130(3):620–628. doi: 10.1016/j.ygyno.2013.05.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Czogalla B, Kuhn C, Heublein S, Schmockel E, Mayr D, Kolben T, Trillsch F, Burges A, Mahner S, Jeschke U, Hester A. EP3 receptor is a prognostic factor in TA-MUC1-negative ovarian cancer. J Cancer Res Clin Oncol. 2019;145(10):2519–2527. doi: 10.1007/s00432-019-03017-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.den Boon JA, Pyeon D, Wang SS, Horswill M, Schiffman M, Sherman M, Zuna RE, Wang Z, Hewitt SM, Pearson R, Schott M, Chung L, He Q, Lambert P, Walker J, Newton MA, Wentzensen N, Ahlquist P. Molecular transitions from papillomavirus infection to cervical precancer and cancer: role of stromal estrogen receptor signaling. Proc Natl Acad Sci USA. 2015;112(25):E3255–3264. doi: 10.1073/pnas.1509322112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Daniyal M, Akhtar N, Ahmad S, Fatima U, Akram M, Asif HM. Update knowledge on cervical cancer incidence and prevalence in Asia. Asian Pac J Cancer Prev. 2015;16(9):3617–3620. doi: 10.7314/APJCP.2015.16.9.3617. [DOI] [PubMed] [Google Scholar]
- 72.Small W, Jr, Bacon MA, Bajaj A, Chuang LT, Fisher BJ, Harkenrider MM, Jhingran A, Kitchener HC, Mileshkin LR, Viswanathan AN, Gaffney DK. Cervical cancer: a global health crisis. Cancer. 2017;123(13):2404–2412. doi: 10.1002/cncr.30667. [DOI] [PubMed] [Google Scholar]
- 73.Sankaranarayanan R, Swaminathan R, Brenner H, Chen K, Chia KS, Chen JG, Law SC, Ahn YO, Xiang YB, Yeole BB, Shin HR, Shanta V, Woo ZH, Martin N, Sumitsawan Y, Sriplung H, Barboza AO, Eser S, Nene BM, Suwanrungruang K, Jayalekshmi P, Dikshit R, Wabinga H, Esteban DB, Laudico A, Bhurgri Y, Bah E, Al-Hamdan N. Cancer survival in Africa, Asia, and Central America: a population-based study. Lancet Oncol. 2010;11(2):165–173. doi: 10.1016/S1470-2045(09)70335-3. [DOI] [PubMed] [Google Scholar]
- 74.Young RH, Clement PB. Endocervical adenocarcinoma and its variants: their morphology and differential diagnosis. Histopathology. 2002;41(3):185–207. doi: 10.1046/j.1365-2559.2002.01462.x. [DOI] [PubMed] [Google Scholar]
- 75.Sales KJ, Katz AA, Davis M, Hinz S, Soeters RP, Hofmeyr MD, Millar RP, Jabbour HN. Cyclooxygenase-2 expression and prostaglandin E(2) synthesis are up-regulated in carcinomas of the cervix: a possible autocrine/paracrine regulation of neoplastic cell function via EP2/EP4 receptors. J Clin Endocrinol Metab. 2001;86(5):2243–2249. doi: 10.1210/jcem.86.5.7442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Sales KJ, Katz AA, Millar RP, Jabbour HN. Seminal plasma activates cyclooxygenase-2 and prostaglandin E2 receptor expression and signalling in cervical adenocarcinoma cells. Mol Hum Reprod. 2002;8(12):1065–1070. doi: 10.1093/molehr/8.12.1065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Huang M, Chen Q, Xiao J, Liu C, Zhao X. Prognostic significance of cyclooxygenase-2 in cervical cancer: a meta-analysis. Int J Cancer. 2013;132(2):363–373. doi: 10.1002/ijc.27686. [DOI] [PubMed] [Google Scholar]
- 78.Kim YB, Kim GE, Pyo HR, Cho NH, Keum KC, Lee CG, Seong J, Suh CO, Park TK. Differential cyclooxygenase-2 expression in squamous cell carcinoma and adenocarcinoma of the uterine cervix. Int J Radiat Oncol Biol Phys. 2004;60(3):822–829. doi: 10.1016/j.ijrobp.2004.04.030. [DOI] [PubMed] [Google Scholar]
- 79.Pyo H, Kim YB, Cho NH, Suh CO, Park TK, Yun YS, Kim GE. Coexpression of cyclooxygenase-2 and thymidine phosphorylase as a prognostic indicator in patients with FIGO stage IIB squamous cell carcinoma of uterine cervix treated with radiotherapy and concurrent chemotherapy. Int J Radiat Oncol Biol Phys. 2005;62(3):725–732. doi: 10.1016/j.ijrobp.2004.10.044. [DOI] [PubMed] [Google Scholar]
- 80.Herfs M, Herman L, Hubert P, Minner F, Arafa M, Roncarati P, Henrotin Y, Boniver J, Delvenne P. High expression of PGE2 enzymatic pathways in cervical (pre)neoplastic lesions and functional consequences for antigen-presenting cells. Cancer Immunol Immunother. 2009;58(4):603–614. doi: 10.1007/s00262-008-0584-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Radilova H, Libra A, Holasova S, Safarova M, Viskova A, Kunc F, Buncek M. COX-1 is coupled with mPGES-1 and ABCC4 in human cervix cancer cells. Mol Cell Biochem. 2009;330(1–2):131–140. doi: 10.1007/s11010-009-0126-1. [DOI] [PubMed] [Google Scholar]
- 82.Deckmann K, Rorsch F, Geisslinger G, Grosch S. Dimethylcelecoxib induces an inhibitory complex consisting of HDAC1/NF-kappaB(p65)RelA leading to transcriptional downregulation of mPGES-1 and EGR1. Cell Signal. 2012;24(2):460–467. doi: 10.1016/j.cellsig.2011.09.025. [DOI] [PubMed] [Google Scholar]
- 83.Oh JM, Kim SH, Lee YI, Seo M, Kim SY, Song YS, Kim WH, Juhnn YS. Human papillomavirus E5 protein induces expression of the EP4 subtype of prostaglandin E2 receptor in cyclic AMP response element-dependent pathways in cervical cancer cells. Carcinogenesis. 2009;30(1):141–149. doi: 10.1093/carcin/bgn236. [DOI] [PubMed] [Google Scholar]
- 84.Parida S, Pal I, Parekh A, Thakur B, Bharti R, Das S, Mandal M. GW627368X inhibits proliferation and induces apoptosis in cervical cancer by interfering with EP4/EGFR interactive signaling. Cell Death Dis. 2016;7:e2154. doi: 10.1038/cddis.2016.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Heidegger H, Dietlmeier S, Ye Y, Kuhn C, Vattai A, Aberl C, Jeschke U, Mahner S, Kost B. The prostaglandin EP3 receptor is an independent negative prognostic factor for cervical cancer patients. Int J Mol Sci. 2017 doi: 10.3390/ijms18071571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Rao CV, Reddy BS. NSAIDs and chemoprevention. Curr Cancer Drug Targets. 2004;4(1):29–42. doi: 10.2174/1568009043481632. [DOI] [PubMed] [Google Scholar]
- 87.Nan H, Hutter CM, Lin Y, Jacobs EJ, Ulrich CM, White E, Baron JA, Berndt SI, Brenner H, Butterbach K, Caan BJ, Campbell PT, Carlson CS, Casey G, Chang-Claude J, Chanock SJ, Cotterchio M, Duggan D, Figueiredo JC, Fuchs CS, Giovannucci EL, Gong J, Haile RW, Harrison TA, Hayes RB, Hoffmeister M, Hopper JL, Hudson TJ, Jenkins MA, Jiao S, Lindor NM, Lemire M, Le Marchand L, Newcomb PA, Ogino S, Pflugeisen BM, Potter JD, Qu C, Rosse SA, Rudolph A, Schoen RE, Schumacher FR, Seminara D, Slattery ML, Thibodeau SN, Thomas F, Thornquist M, Warnick GS, Zanke BW, Gauderman WJ, Peters U, Hsu L, Chan AT, Ccfr G. Association of aspirin and NSAID use with risk of colorectal cancer according to genetic variants. JAMA. 2015;313(11):1133–1142. doi: 10.1001/jama.2015.1815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Baandrup L, Faber MT, Christensen J, Jensen A, Andersen KK, Friis S, Kjaer SK. Nonsteroidal anti-inflammatory drugs and risk of ovarian cancer: systematic review and meta-analysis of observational studies. Acta Obstet Gynecol Scand. 2013;92(3):245–255. doi: 10.1111/aogs.12069. [DOI] [PubMed] [Google Scholar]
- 89.Bally M, Dendukuri N, Rich B, Nadeau L, Helin-Salmivaara A, Garbe E, Brophy JM. Risk of acute myocardial infarction with NSAIDs in real world use: bayesian meta-analysis of individual patient data. BMJ. 2017;357:j1909. doi: 10.1136/bmj.j1909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Sarkar FH, Adsule S, Li Y, Padhye S. Back to the future: COX-2 inhibitors for chemoprevention and cancer therapy. Mini Rev Med Chem. 2007;7(6):599–608. doi: 10.2174/138955707780859431. [DOI] [PubMed] [Google Scholar]
- 91.Rayar AM, Lagarde N, Ferroud C, Zagury JF, Montes M, Sylla-Iyarreta Veitia M. Update on COX-2 selective inhibitors: chemical classification, side effects and their use in cancers and neuronal diseases. Curr Top Med Chem. 2017;17(26):2935–2956. doi: 10.2174/1568026617666170821124947. [DOI] [PubMed] [Google Scholar]
- 92.Majumder M, Nandi P, Omar A, Ugwuagbo KC, Lala PK. EP4 as a therapeutic target for aggressive human breast cancer. Int J Mol Sci. 2018 doi: 10.3390/ijms19041019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Sheng H, Shao J, Washington MK, DuBois RN. Prostaglandin E2 increases growth and motility of colorectal carcinoma cells. J Biol Chem. 2001;276(21):18075–18081. doi: 10.1074/jbc.M009689200. [DOI] [PubMed] [Google Scholar]
- 94.Ushikubi F, Segi E, Sugimoto Y, Murata T, Matsuoka T, Kobayashi T, Hizaki H, Tuboi K, Katsuyama M, Ichikawa A, Tanaka T, Yoshida N, Narumiya S. Impaired febrile response in mice lacking the prostaglandin E receptor subtype EP3. Nature. 1998;395(6699):281–284. doi: 10.1038/26233. [DOI] [PubMed] [Google Scholar]
- 95.Ikeda-Matsuo Y, Tanji H, Narumiya S, Sasaki Y. Inhibition of prostaglandin E2 EP3 receptors improves stroke injury via anti-inflammatory and anti-apoptotic mechanisms. J Neuroimmunol. 2011;238(1–2):34–43. doi: 10.1016/j.jneuroim.2011.06.014. [DOI] [PubMed] [Google Scholar]