

Abstract
Banana peels (BP), an under-utilized waste material, was studied for the production of xylanase and pectinase by Aspergillus fumigates MS16. The factors affecting the co-production of both the enzymes were separately studied for their influence under submerged (Smf) and solid-state fermentation (SSF) of BP. The strain was cultivated in the presence of mineral salt (MS) solution containing BP powder as a sole source of carbon and physical and nutritional factors varied to observe the change in the enzyme titers. The data revealed that the MS-based medium was appropriate for the production of both the enzymes; therefore, in subsequent experiments, the same medium was used. A temperature of 30–35°C was found better for the production of the two enzymes under Smf; however, the titers of pectinase dropped significantly at 40°C. Contrarily, xylanase production was inhibited at 40°C under SSF but not under Smf. Whereas, supplementation of xylan or pectin to BP induced the production of xylanase and pectinase, respectively. Lowering the pH value favored the production of both the enzymes under Smf; however, the production of pectinase improved significantly when a higher concentration of BP (1%) was used compared to the concentration (0.25%) required for the production of xylanase. Interestingly, the enzyme preparation obtained under SSF exhibited optimal activities of both the enzymes at higher temperatures when compared to those obtained under Smf. The data indicated that the physiology of the fungus differed greatly when the cultivation pattern varied from Smf to SSF and, hence, the enzymes produced were characteristically distinct.
Key words: Aspergillus fumigatus, banana peels, pectinase, Solid-State Fermentation, xylanase
Introduction
Lignocellulose (LC) present in the plant cell wall provides strength and protection against pathogens. Lignin, cellulose, and hemicelluloses are the main constituents of the LC (Yang et al. 2004). Cellulose is a homopolysaccharide while hemicelluloses are derived from different sugars, the composition of which depends on the type of plants and weather conditions. Xylose is the main constituent of most of the hemicellulose and it is usually arranged in shorter chains of ~ 200 units (Brown et al. 2009) to comprise xylan.
Considering heterogeneity in the structure of hemicelluloses, microbial degradation of hemicellulose requires various enzymes; xylanases play a major role in this context. Pectinases, on the other hand, attack on a pectic portion of the LC masses. Since abundantly found LC substrates are rich in fermentable components, therefore, these are perceived as the promising raw materials for biorefineries. Hence, the bioconversion of LC is an important subject in biological research (Otero and Nielsen 2010). Previously, enzymatic digestion of several LC materials has been reported such as corncob (Kahar et al. 2010), Kraft paper-mill sludges (Kang et al. 2010), and sugarcane bagasse (Buaban et al. 2010). It suggests that LC materials can be saccharified using an enzyme(s) and can provide a cheaper source of raw materials for many industries. However, the selection of an LC substrate for industrial exploitation mainly depends on local agricultural practices, as availability of the substrates remains a decisive factor. Banana peels (BP) is a waste LC substrate that is generated by the fruit that remains available throughout the year in Pakistan and, hence, can be investigated for its possible application as a promising LC substrate.
Banana is one of the most popular fruit in the world owing to its nutritional value, hygiene, cost-effectiveness, and its digestibility. In South Asia, it is the leading fruit, grown over a hundred thousand hectares of cultivated land. Consequently, thousands of tons of BP are accumulated as the domestic and food industry’s waste, which is not effectively utilized. With considerable quantities of lignin (6–12%), pectin (10–21%), cellulose (7–10%), and hemicelluloses (6–9.4%) (Emaga et al. 2008), BP has been reported as an effective fermentation substrate (Kiranmayi et al. 2011; Rehman et al. 2014). Various studies have been conducted to produce industrially important enzymes using BP including alpha-amylase by Bacillus subtilis and Penicillium species (Akkarachaneeyakorn et al. 2018), and laccase by Trametes pubescens (Osma et al. 2007). Moreover, the substrate has also been utilized to obtain citric acid from Aspergillus niger (Karthikeyan and Sivakumar 2010), biohydrogen by adopting two-phase anaerobic fermentation (Nathoa et al. 2014), and for alcohol production (Janveja et al. 2013). Considering the availability of BP in Pakistan and the suitability of the substrate for fermentation processes, this study was conducted to compare coproduction of xylanase and pectinase by A. fumigatus MS16 under two types of fermentation i.e. submerged and solid-state fermentation. The enzymes obtained under the two sets of processes were investigated to reveal the differences in their activity.
Experimental
Material and Methods
Organism and culture maintenance. The strain of A. fumigatus MS16, previously reported to produce cellulase, xylanase, and pectinase under the fermentation of apple peels (Jalis et al. 2014) and some other crude substrates (Naseeb et al. 2015), was retrieved on Sabouraud’s dextrose agar (SDA) slants, stored at 4°C and sub-cultured periodically.
Substrate. The substrate, BP, was collected from a fruit juice seller and was solar-dried for 5 days, cut into pieces, ground, autoclaved, passed through 100 mesh particle-size sieve, and stored under refrigeration until used.
Inoculum. A. fumigates MS16 was grown on SDA plates for 6 days at 35°C. Spores were collected and a spore count of 1 × 108 spores ml–1 in mineral salt medium (MSM; Shariq et al. 2018) was maintained by using a hemocytometer. The spore suspension was used as inoculum for the submerged and solid-state fermentation.
Solid-state fermentation (SSF). BP (2 g) was autoclaved in a 100 ml Erlenmeyer flask at 121°C for 30 min and moistened with 3.6 ml of MSM to achieve the desired moisture level. An inoculum of 2 ml was sprinkled over the substrate and the flasks were incubated at 30°C for one week. In another set of experiments, BP was separately supplemented with 0.5% xylan (Sigma-Aldrich, Germany) and 0.5% pectin (Sigma-Aldrich, Germany), and SSF was carried out as described above. In yet another set of experiments, the effect of temperature on SSF of BP was studied by carrying out the cultivation at a temperature from 25–40°C. Subsequently, the variable moisture contents and incubation period were investigated.
The enzyme preparation was harvested by shaking the fermented BP in 40 ml of 0.05 M sodium citrate buffer (pH – 4.8) and Tween 80 (0.5%) on an orbital shaker for 1 h at 150 rpm at 30°C and, then, filtering the slurry through several layers of muslin cloth and a Whatman number 1 filter paper. The extract was centrifuged at 6000 × g for 20 min, and the supernatant was used as a crude enzyme preparation that was stored at –20°C until used for the determination of the enzyme activity.
Submerged fermentation (Smf). Smf was carried out in 250 ml Erlenmeyer flask containing 100 ml of MSM supplemented with BP (1%, w/v). The inoculum was transferred to the flasks to obtain a final concentration of 1 × 106 spores g–1 substrate and incubated for one week under static condition. To harvest enzyme after submerged fermentation, the contents were passed through the glass wool and filtrate was centrifuged at 8000 × g for 30 min. The cell-free culture supernatant (CFCS) was stored at –20°C until used as an enzyme preparation.
The submerged fermentation was further investigated by adopting one variable at a time strategy (Shariq et al. 2018). To optimize the temperature for the production of xylanase and pectinase the flasks were incubated at a temperature from 25 to 40°C. Likewise, optimum pH was determined by cultivating the strain MS 16 at an optimum temperature in the medium with pH adjusted to 4–6.5. In a similar manner, the substrate concentration (0–1.6%), the effect of supplementation (0.5% xylan or pectin), and the incubation period (5–7 days) were studied.
Enzyme activity assays. The xylanase and pectinase activities were determined by using 0.5% birchwood xylan or 0.5% citrus pectin as a substrate (Rehman et al. 2014). The liberation of reducing sugars was estimated by the dinitrosalicylic acid (DNS) method (Miller 1959) against standard curves of xylose or galacturonic acid. One international unit of enzymes (xylanase and pectinase) was defined as the amount of enzyme that produced the reducing sugar equivalent to 1 μmol of the product (xylose or galacturonic acid) per minute.
Statistical Analysis. All the experiments were conducted in triplicate and mean values along with standard deviation have been presented. The analysis was carried out using SigmaPlot (version 12.5, USA).
Results and Discussion
Fungal strains have frequently been reported for the production of industrially important enzymes from waste materials. The hyphal mode of growth enables them to penetrate through the substrate and to utilize fermentable components by elaborating hydrolases. The availability of banana makes its waste, BP, a promising substrate for the fungal fermentation to obtain value-added products. Previously, A. fumigatus was found to have better growth on the banana peels medium when compared to the other fungal strains (Essien et al. 2005). Whereas, the strain MS16 of A. fumigatus reportedly produced cellulase, xylanase, and pectinase on apple peels (Jalis et al. 2014) and other waste materials (Naseeb et al. 2015). Subsequently, the ability of this fungus to produce xylanase and pectinase under submerged and solid-state fermentation of BP is being reported here. Considering the fact that fermentation processes are greatly influenced by environmental conditions and nutritional factors, a comparative study was conducted to investigate the effect of these factors on the production of the enzymes under two different sets of conditions i.e. Smf and SSF.
Submerged fermentation. The studies on the effect of the medium on the enzyme production showed that the maximum activity of xylanase was found in mineral salt medium (MSM), whereas, titers of pectinase were slightly higher in Czapeck dox broth than in MSM (Fig. 1). Therefore, MSM was used in the remaining experiments.
Fig. 1.
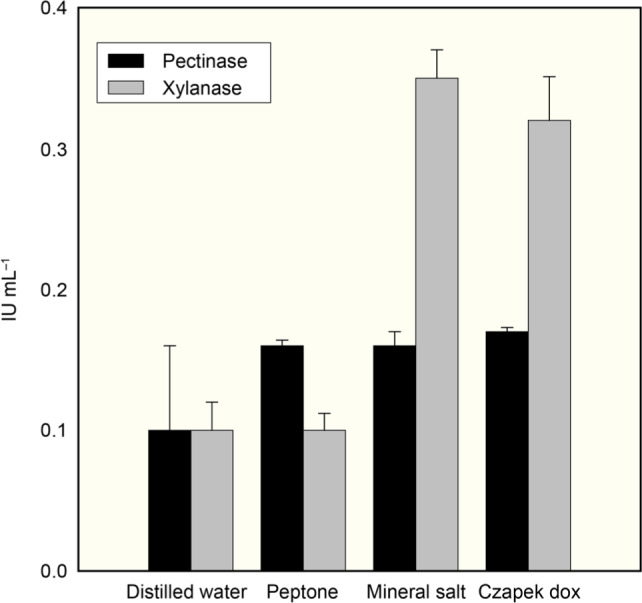
Effect of medium along with banana peels (BP) on the pectinase and xylanase production by A. fumigatus MS16 under submerged fermentation.
When the effect of temperature on pectinase production was studied, it was observed that there wasn’t much variation in the titers of pectinase when the cultivation temperature varied from 25 to 35°C (Fig. 2). However, a drastic decrease in the levels of pectinase was observed when the temperature was adjusted to 40°C. Temperature affected the production of xylanase differently as it was observed that Smf could be carried out at a temperature from 20–40°C without affecting the enzyme yield, though titers of the enzymes were slightly higher at 30 and 35°C. This finding was in agreement with the results obtained by Naseeb et al. (2015).
Fig. 2.

Effect of temperature on pectinase and xylanase production under submerged (SmF) and Solid state fermentation (SSF) of banana peels by A. fumigatus MS16.
To investigate the induction of pectinase and xylanase by the corresponding substrate (pectin or xylan), the two substrates were added separately or in combination, in BP containing MSM and the production of both the enzymes was studied. It was noted that pectinase production was enhanced when only pectin or pectin with xylan was added to the MSM + BP (Fig. 3); however, the production was negatively affected if only xylan was added to the MSM + BP. Likewise, xylanase production by MS16 was induced in the presence of xylan supplemented MSM + BP or xylan and pectin containing MSM + BP. Previously, supplementation of pectin and xylan to apple peel powder was found to induce pectinase and xylanase, respectively, under SSF by A. fumigtus MS16 (Jalis et al. 2014).
Fig. 3.
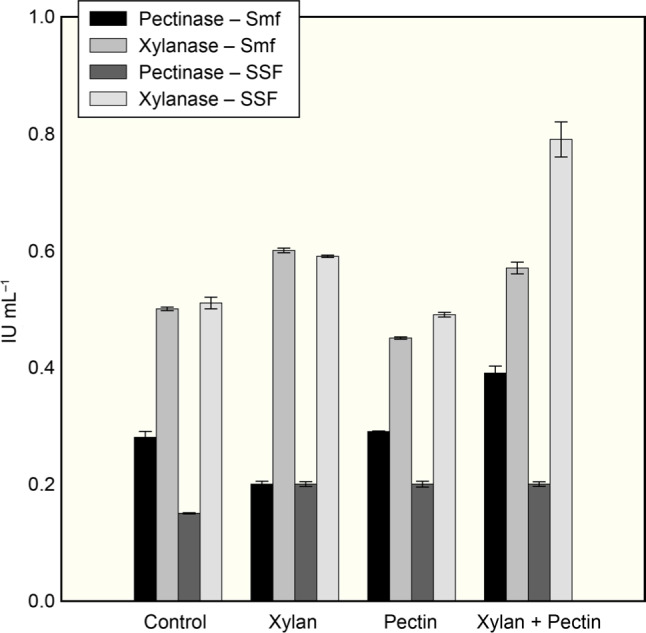
Effect of supplementation of pectin and xylan to banana peels containing medium on the production of pectinase and xylanase from A. fumigatus MS16 under solid-state (SSF) and submerged fermentation (Smf).
The pH value of the medium influences the production of an enzyme by regulating the solubility of the nutrients, the permeability of the cell membrane, and ionization of amino acids and/or proteins. Usually, fungal strains grow better under acidic conditions; A. fumigates MS16 did not show any exception, indeed a value of 5 as initial pH of the medium favored the production of pectinase (Fig. 4) and pH 6 spurred xylanase production. The production of extracellular enzyme by A. fumigates under acidic conditions using municipal waste as a substrate (Gautam et al. 2011) and xylanase by a mutated strain of A. niger (Haq et al. 2004) was reported previously.
Fig. 4.

Effect of pH on pectinase and xylanase production from A. fumigatus MS16 under submerged fermentation.
In the submerged fermentation the varying concentrations (0.25–1.6%) of BP were tested to determine the optimal concentration for the production of the enzymes. MS16 produced the highest titers of pectinase in MSM with 1% BP and xylanase in 0.25% BP containing medium (Fig. 5).
Fig. 5.

Effect of banana peels concentration on pectinase and xylanase production from A. fumigatus MS16 under submerged fermentation.
After studying optimum levels of the above-mentioned factors, the enzyme production was studied by taking aliquots intermittently and the optimum incubation period for both the enzymes was determined. The data showed that pectinase production was maximum on day six, thereafter, it decreased slightly, while xylanase production dropped significantly when the incubation period extended from five days (data not shown).
Solid-State Fermentation. The production of pectinase and xylanase was also studied in the absence of water, i.e. under SSF. In this case, the optimum levels of the factors were different than was observed for Smf. For instance, a cultivation temperature of 25°C appeared as the most suitable temperature when the production of pectinase was studied under SSF (Fig. 2). Earlier, pectinase production by Fusarium sp. was reportedly found better at a lower temperature (Sohail et al. 2009) while A. niger also produced pectinase under similar conditions (Haq et al. 2004). The production of the enzyme under SSF appeared as sensitive to temperature as the levels of xylanase declined by 50% when the temperature was increased from 35 to 40°C. It can be attributed to poor control of heat transfer under SSF that usually results in an increase in the temperature inside the substrate and hence the enzyme can be denatured or organism’s growth can be restricted. Palaniswamy et al. (2012) also observed the decrease in xylanase production by Aspergillus sp. in rice bran containing medium at higher temperatures.
In SSF, the xylanase production increased with the supplementation of 0.5% xylan alone or with pectin, while it decreased when the pectin was supplemented without xylan (Fig. 3). The supplementation of pectin and/or xylan did not exert the significant effect on the pectinase production by MS16. In their studies, Rehman et al. (2014) reported about the indifferent pattern of pectinase production upon supplementation of pectin to BP under SSF by a fungal co-culture.
The moisture content is one of the important factors for solid-state fermentation but it did not affect the activity of xylanase; it was almost the same but the pectinase activity was maximal at 65% moisture content (Fig. 6) that corroborated with the findings of Padma et al. (2012) where 65% moisture was optimal to obtain the highest levels of polygalactouronase by SSF of BP.
Fig. 6.

Effect of moisture content on pectinase and xylanase production under solid-state fermentation of banana peels.
Finally, the production of the enzyme was studied under optimum conditions for a period of 5–7 days and enzyme activity was assayed. The maximum pectinase was produced in five days of incubation while xylanase production was the highest after seven days of incubation (data not shown).
Factors affecting enzyme activities. The optimum temperatures and pH for the activities of xylanase and pectinase produced under Smf and SSF were also determined. The pectinase activity produced under Smf was found to be the highest at 45°C (Fig. 7) while the enzyme produced under SSF exhibited optimal activity at 60°C. Likewise, xylanase activity produced under SSF had higher temperature optima of 55–60°C compared to the enzyme produced under smf that was optimally active at 50°C. However, the extraction methods to obtain crude enzyme from Smf and SSF differed greatly and might have influenced the stability of the enzymes. Nonetheless, both the enzymes were more heat-stable as compared to the cellulase (Topt 40°C) produced on BP (Kiranmayi et al. 2011).
Fig. 7.

Effect of temperature on pectinase and xylanase activities produced under solid-state (SSF) and submerged fermentation (Smf).
The studies on the effect of pH on the enzyme activity showed that pectinase and xylanase were catalytically most active at pH 5.0 and 5.5 (Fig. 8), respectively, and this property remained unaffected when the production method was changed from Smf to SSF. Generally, fungal enzymes work well under acidic conditions. For instance, Kiranmayi et al. (2011) reported that a value of 6.0 as the most suitable pH for cellulase activity from A. niger on BP as a substrate.
Fig. 8.

Effect of pH on pectinase and xylanase activity produced from A. fumigatus MS16 under solid-state (SSF) and submerged fermentation (Smf).
Conclusions
Banana peels appeared as a suitable substrate for the simultaneous production of pectinase and xylanase from A. fumigatus MS16 under solid-state and submerged fermentation. The conditions affecting the process varied greatly and hence can be adjusted to obtain the higher titers of any one of the enzymes. Moreover, the type of fermentation influenced the characteristics of the enzyme as the pectinase produced under solid-state fermentation worked at a higher optimum temperature than produced under submerged fermentation.
Footnotes
Conflict of interest
The authors do not report any financial or personal connections with other persons or organizations, which might negatively affect the contents of this publication and/or claim authorship rights to this publication.
ORCID
Muhammad Sohail https://orcid.org/0000-0002-7208-9441
Literature
- Akkarachaneeyakorn S, Suwakrai A, Pewngam D.. Optimization of reducing sugar production from enzymatic hydrolysis of banana peels using response surface methodology. Songklanakarin J Sci Technol. 2018. 10.14456/sjst-psu.2018.1 [DOI] [Google Scholar]
- Brown WH, Foote CS, Iverson BL, Anslyn E.. Organic Chemistry. Belmont (USA): Brooks Cole Cengage Learning; 2009. [Google Scholar]
- Buaban B, Inoue H, Yano S, Tanapongpipat S, Ruanglek V, Champreda V, Pichyangkura R, Rengpipat S, Eurwilaichitr L.. Bioethanol production from ball milled bagasse using an on-site produced fungal enzyme cocktail and xylose-fermenting Pichia stipitis. J Biosci Bioeng. 2010. Jul;110(1):18–25. 10.1016/j.jbiosc.2009.12.003 [DOI] [PubMed] [Google Scholar]
- Essien JP, Akpan EJ, Essien EP.. Studies on mould growth and biomass production using waste banana peel. Bioresour Technol. 2005. Sep;96(13):1451–1456. 10.1016/j.biortech.2004.12.004 [DOI] [PubMed] [Google Scholar]
- Gautam SP, Bundela PS, Pandey AK, Awasthi MK, Sarsaiya S.. Isolation, identification and cultural optimization of indigenous fungal isolates as a potential bioconversion agent of municipal solid waste. Ann Environ Sci (Boston Mass). 2011;5:23–34. [Google Scholar]
- Happi Emaga T, Robert C, Ronkart SN, Wathelet B, Paquot M.. Dietary fibre components and pectin chemical features of peels during ripening in banana and plantain varieties. Bioresour Technol. 2008. Jul;99(10):4346–4354. 10.1016/j.biortech.2007.08.030 [DOI] [PubMed] [Google Scholar]
- Haq I-U, Tasneem M, Raana K, Khan A, Mukhtar H, Javed M.. Optimization of cultural conditions for the production of xylanase by chemically mutated strain of Aspergillus niger GCBCX-20. Int J Agric Biol. 2004;6(6):1115–1118. [Google Scholar]
- Jalis H, Ahmad A, Khan SA, Sohail M.. Utilization of apple peels for the production of plant cell-wall degrading enzymes by Aspergillus fumigatus MS16. J Anim Plant Sci. 2014;24(2). [Google Scholar]
- Janveja C, Rana SS, Soni KS.. Kitchen waste residues as potential renewable biomass resources for the production of multiple fungal carbohydrases and second generation bioethanol. J Technol Innov Renew Energy. 2013;2(2): 10.6000/1929-6002.2013.02.02.11 [DOI] [Google Scholar]
- Kahar P, Taku K, Tanaka S.. Enzymatic digestion of corncobs pretreated with low strength of sulfuric acid for bioethanol production. J Biosci Bioeng. 2010. Oct;110(4):453–458. 10.1016/j.jbiosc.2010.05.002 [DOI] [PubMed] [Google Scholar]
- Kang L, Wang W, Lee YY.. Bioconversion of kraft paper mill sludges to ethanol by SSF and SSCF. Appl Biochem Biotechnol. 2010. May; 161(1–8):53–66. 10.1007/s12010-009-8893-4 [DOI] [PubMed] [Google Scholar]
- Karthikeyan A, Sivakumar N.. Citric acid production by Koji fermentation using banana peel as a novel substrate. Bioresour Technol. 2010. Jul;101(14):5552–5556. 10.1016/j.biortech.2010.02.063 [DOI] [PubMed] [Google Scholar]
- Kiranmayi MU, Poda S, Vijayalakshmi M, Krishna PV, Krishna PV.. Studies on influence of natural biowastes on cellulase production by Aspergillus niger. J Environ Biol. 2011. Nov;32(6):695–699. [PubMed] [Google Scholar]
- Miller GL. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal Chem. 1959. Mar;31(3):426–428. 10.1021/ac60147a030 [DOI] [Google Scholar]
- Naseeb S, Sohail M, Ahmad A, Khan SA.. Production of xylanases and cellulases by Aspergillus fumigatus MS16 using crude lignocellulosic substrates. Pak J Bot. 2015;47(2). [Google Scholar]
- Nathoa C, Sirisukpoca U, Pisutpaisal N.. Production of hydrogen and methane from banana peel by two phase anaerobic fermentation. Energy Procedia. 2014;50:702–710. 10.1016/j.egypro.2014.06.086 [DOI] [Google Scholar]
- Osma JF, Toca Herrera JL, Rodríguez Couto S.. Banana skin: A novel waste for laccase production by Trametes pubescens under solid-state conditions. Application to synthetic dye decolouration. Dye Pigment. 2007;75(1):32–37. 10.1016/j.dyepig.2006.05.021 [DOI] [Google Scholar]
- Otero JM, Nielsen J.. Industrial systems biology. Biotechnol Bioeng. 2010. Feb 15;105(3):439–460. 10.1002/bit.22592 [DOI] [PubMed] [Google Scholar]
- Padma PN, Anuradha K, Nagaraju B.. Use of pectin rich fruit wastes for polygalacturonase production by Aspergillus awamori MTCC 9166 in Solid State Fermentation. J Bioprocess Biotech. 2012;02(02). 10.4172/2155-9821.1000116 [DOI] [Google Scholar]
- Palaniswamy M, Arulanandham TV, Angayarkanni J.. Production of xylanase by litter degrading fungal species using agro-industrial wastes as substrates by solid state fermentation. Res J Pharm Biol Chem Sci. 2012;3(1):143–149. [Google Scholar]
- Rehman S, Aslam H, Ahmad A, Khan SA, Sohail M.. Production of plant cell wall degrading enzymes by monoculture and co-culture of Aspergillus niger and Aspergillus terreus under SSF of banana peels. Braz J Microbiol. 2014. Dec;45(4):1485–1492. 10.1590/S1517-83822014000400045 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shariq M, Muhammad F, Ahmad A, Khan SA, Moin SF, Sohail M.. Production and characterization of endoglucanase from an indigenous yeast strain. Pak J Bot. 2018;50(6):2413–2421. [Google Scholar]
- Sohail M, Naseeb S, Sherwani SK, Sultana S, Aftab S, Shahzad S, Ahmad A, Khan SA.. Distribution of hydrolytic enzymes among native fungi: aspergillus the pre-dominant genus of hydrolase producer. Pak J Bot. 2009;41(5):2567–2582. [Google Scholar]
- Yang YH, Wang BC, Wang QH, Xiang LJ, Duan CR.. Research on Solid-State Fermentation on rice chaff with a microbial consortium. Colloids Surf B Biointerfaces. 2004. Mar;34(1):1–6. 10.1016/j.colsurfb.2003.10.009 [DOI] [PubMed] [Google Scholar]