

ABSTRACT
Thanks to many advances in genetic manipulation, mouse models have become very powerful in their ability to interrogate biological processes. In order to precisely target expression of a gene of interest to particular cell types, intersectional genetic approaches using two promoter/enhancers unique to a cell type are ideal. Within these methodologies, variants that add temporal control of gene expression are the most powerful. We describe the development, validation and application of an intersectional approach that involves three transgenes, requiring the intersection of two promoter/enhancers to target gene expression to precise cell types. Furthermore, the approach uses available lines expressing tTA/rTA to control the timing of gene expression based on whether doxycycline is absent or present, respectively. We also show that the approach can be extended to other animal models, using chicken embryos. We generated three mouse lines targeted at the Tigre (Igs7) locus with TRE-loxP-tdTomato-loxP upstream of three genes (p21, DTA and Ctgf), and combined them with Cre and tTA/rtTA lines that target expression to the cerebellum and limbs. Our tools will facilitate unraveling biological questions in multiple fields and organisms.
KEY WORDS: Cre, tTA, rtTA, Intersectional genetics, Inducible and reversible gene mis-expression, Tissue-specific, Tigre locus, DRAGON, p21, Ccn2/Ctgf, Diphtheria toxin, DTA, tox176
Summary: This study presents a collection of four mouse lines and genetic tools for mis-expression-mediated manipulation of cellular activity with high spatiotemporal control, in a reversible manner.
INTRODUCTION
The wealth of genetic tools that have been developed for using the mouse to study biological questions in vivo has enabled extremely sophisticated experiments to be conducted. Especially useful are tools that have a modular nature. One approach for inducing gene mis-expression is to combine a mouse line carrying a driver module that expresses a protein (transcription factor) that induces gene expression only when the right genetic elements (DNA binding sites) are present with a line carrying an effector module that contains the necessary genetic elements upstream of a gene of interest (GOI). In this way, only the offspring combining both modules will specifically express the GOI in particular cells defined by the driver module (Kaczmarczyk and Jackson, 2015; Rosen et al., 2015). This system can be modified to implement temporal and spatial control of gene expression by making it dependent on the activity of site-specific recombinases (SSRs) (e.g. Cre), and by expressing SSRs from gene promoters or loci with specific expression patterns (Smedley et al., 2011). One drawback of using SSR systems, including inducible versions that allow for temporal control (e.g. CreER) (Brocard et al., 1998; Feil et al., 1997, 2009; Metzger et al., 1995), is that recombination of the target sites (loxP in the case of Cre) is irreversible. Hence, if mis-expression of a GOI is dependent only on Cre activity, it cannot be reversed, and therefore studying any recovery process after a transient genetic manipulation is not possible.
The possibility of reversibility has been resolved by using other driver-effector combinations, the most prevalent in mouse being the Tet system, based on the tetracycline (Tet) resistance operon of E. coli (Hillen and Berens, 1994). In the first version of this system, a transactivator fusion protein composed of the tetracycline repressor (tetR) and the activating domain of the herpes simplex viral protein 16 (VP16) regulate the expression of a GOI by placing a Tet-responsive element (TRE) containing Tet-operator sequences (tetO) upstream of the gene sequence. This system is known as Tet-Off, because in the presence of Tet or its analog doxycycline (Dox), the affinity of tetR to tetO sequences diminishes by nine orders of magnitude, so that expression of the GOI is greatly reduced (Gossen and Bujard, 1992; St-Onge et al., 1996). Conversely, the Tet-On system is based on a reverse Tet-controlled transactivator (rtTA) derived from tTA through four amino acid changes in the tetR DNA binding moiety. This alters the binding properties of rtTA such that it can only recognize the tetO sequences in the presence of Dox (Gossen et al., 1995; Urlinger et al., 2000). A problem that for many years encumbered systems using tTA and rtTA collectively [referred to as (r)tTA] was that the TRE DNA sequences can be inactivated through epigenetic mechanisms, rendering them unreliable unless derepressed by additional genetic modules (Gödecke et al., 2017). This drawback was overcome when a systematic screen identified a locus in the mouse genome that does not show this inactivation (Zeng et al., 2008). Indeed, Tigre (also known as Igs7) is an intergenic region on mouse chromosome 9 that provides a safe harbor from which TRE-dependent expression can be tightly regulated (hence the name).
Despite these advances, all systems are limited by the fact that individual promoters/enhancers or gene loci often drive the expression of Cre/(r)tTA in a broader area (or more cell types) than is required for the experiment, leading to insufficient specificity. Several intersectional approaches have been developed that allow for greater control over which cells are genetically manipulated. In one approach separate fragments of the Cre recombinase (known as ‘split cre’) are expressed from distinct drivers (promoter/enhancer sequences), resulting in Cre activity (loxP excision) only in the intersectional cell population in which both halves of Cre are expressed (Hirrlinger et al., 2009). Another intersectional approach uses responder modules that depend on two types of recombinases (e.g. Cre and Flp), such as the systems developed by the Dymecki lab (Dymecki and Kim, 2007). However, neither approach allows for the GOI expression to be regulated in a reversible manner.
One strategy that allows for both precision of cell-type targeting and reversibility of gene mis-expression was recently developed by our labs (Madisen et al., 2015; Rosello-Diez et al., 2018). The approach involves combining the Cre/loxP and Tet systems such that the Tigre locus contains a double conditional allele in which expression of the GOI depends both on previous exposure to Cre (to prime the system through elimination of a floxed STOP sequence upstream of the GOI) and on the activity of (r)tTA (through a TRE sequence upstream of the loxP-STOP-loxP-GOI-polyA cassette). The version of this system that includes a tdTomato (tdT) coding sequence for the floxed-STOP cassette was called DRAGON (Fig. 1A), standing for doxycycline-dependent and recombinase-activated gene overexpression (Rosello-Diez et al., 2018). The GOI in the first DRAGON line was the cell-cycle suppressor gene p21 (also known as Cdkn1a) (TigreDragon-p21) and it was used to study cellular compensation mechanisms in the developing limb bones upon cell cycle blockade (Rosello-Diez et al., 2018). As the DRAGON approach relies on two promoter/enhancers, it is intersectional (Fig. 1B), enabling the study of specific tissue-type dependent effects of gene mis-expression. In addition, owing to its dependence on (r)tTA, the system is inducible and reversible (Fig. 1C,D), allowing the effect of different times and durations of mis-expression to be tested. Finally, by developing additional modules with particular GOIs involved in a specific cellular process (e.g. to reduce cell numbers by either blocking proliferation or inducing cell death), the system can effectively determine how different types of insults influence a biological process.
Fig. 1.

An intersectional genetic system for inducible and reversible gene mis-expression. (A) Basic components of the TigreDragon alleles. Ins: insulators; W: Woodchuck virus RNA stabilizing sequence (WPRE). Expression of the GOI depends on both Cre and (r)tTA. (B-D) In the presence of just (r)tTA (blue in C and D), only the tdTomato (tdT) embedded in the intact floxed cassette is expressed (top row in B, magenta in C and D). In the presence of just Cre (green in C and D), the floxed tdTomato-STOP cassette is eliminated, priming the system and deleting tdT, but the GOI is not expressed (middle row in B). It is only in the presence of both Cre and (r)tTA (hatched blue and green in C and D) that the GOI is expressed (bottom row in B, yellow in C and D). As the expression of both tdT and the GOI depends on (r)tTA activity, their expression is inducible and reversible, as it requires the presence of doxycycline (Dox) for rtTA to be active (C), or its absence in the case of tTA (D). (E) Table summarizing the available Dragon lines and their potential uses.
Here, we describe a series of new TigreDragon lines (Fig. 1E) and provide a proof-of-principle for the kind of uses that they enable in two developing organs, the cartilage that drives long-bone growth in the limb (Kronenberg, 2003) and the excitatory cells of the cerebellar nuclei required for cerebellum growth (Willett et al., 2019). We also describe non-intersectional applications for comparison purposes, and the possibility of using these tools for transient gene mis-expression in avian models.
RESULTS
A TigreDragon-DTA model can be used to generate acute tissue-specific cell ablation
A common theme in developmental and regenerative biology is that different responses are elicited by cell ablation compared with defective cell proliferation. Therefore, to complement our TigreDragon-p21 line, we generated an intersectional TigreDragon-DTA model (see Materials and Methods, Fig. 2A and Willett et al., 2019) to enable cell ablation with tissue specificity and temporal control, via expression of attenuated diphtheria toxin fragment A (DTA) (Breitman et al., 1990). To kill some of the excitatory cerebellar nuclei neurons (eCN) that are the main output neurons of the cerebellum, we combined it with two transgenes: Atoh1-tTA (Tet-Off) (Willett et al., 2019) and Atoh1-Cre (Matei et al., 2005). Both transgenes are expressed in cells after they leave the progenitor zone called the rhombic lip. As both the eCN and the precursors of the cerebellar granule cells, another excitatory cell type, express Atoh1 but at different time points [embryonic day (E) 9-E12 and after E13, respectively; Machold and Fishell, 2005], the mice were provided with Dox starting at E13.5 to shut down tTA activity after the eCN had extinguished Atoh1 expression, thus restricting DTA expression to the eCN and sparing the granule cell precursors (Fig. 2B). As shown in Fig. 2C-F, whereas control cerebella showed almost no cell death, in the cerebella of the Atoh1-tTA/+; Atoh1-Cre/+; TigreDragon-DTA/+ mice given Dox at E13.5 onwards (AC-eCN-DTA model: AC designates Atoh1-Cre and eCN designates eCN-expression using Atoh1-tTA) ectopic cell death (marked by TUNEL staining) was detected specifically in the nuclear transitory zone (NTZ), the region to which the precursors of the eCN first migrate after leaving the rhombic lip. tdTom+ cells were detected in the cells expected to express Atoh1-tTA, including the NTZ, granule cell precursors in the external granule cell layer (EGL) and in Atoh1-derived precerebellar hindbrain nuclei (Fig. 2E). Quantification of cell death revealed an 8.85±3.35-fold increase in TUNEL+ particles in the NTZ in AC-eCN-DTA animals compared with their control littermates (TigreDragon-DTA/+) at E13.5 (n=3 per condition, Fig. 2G). At postnatal day (P) 1, the number of eCN, revealed by MEIS2 nuclear immunostaining, was reduced by more than 50% in AC-eCN-DTA cerebella compared with controls (Fig. 2H-J).
Fig. 2.

A TigreDragon-DTA model to generate acute cell/tissue-specific cell ablation. (A,B) Schematic showing TigreDragon-DTA allele (A) and the experimental plan (B). (C-F) Sagittal sections through the cerebellum of three types of E13.5 control embryos (TigreDragon-DTA/+, Atoh1-Cre/+; TigreDragon-DTA/+ and Atoh1-tTA/+; TigreDragon-DTA/+) and an AC-eCN-DTA embryo in the absence of Dox. The boxed areas are shown magnified in the lower panels. Long arrows in the top panels show the direction of the tangential migration as the cells exit the RL. Arrowheads in the bottom panels point to apoptotic cells (TUNEL+). (G) Quantification of cell death in the NTZ identified by MEIS2 staining on adjacent sections (not shown) of E13.5 embryos as measured by the number of TUNEL+ particles. TigreDragon-DTA/+ animals were used as controls (n=3/genotype, P=0.016, Mann–Whitney unpaired test). (H,I) Sagittal sections through the cerebellum at P1, showing reduced numbers of MEIS2+ eCN in AC-eCN-DTA mutants compared with a TigreDragon-DTA/+ control. Right panels show magnification of the boxed areas in the left panels. (J) Quantification of MEIS2+ eCN number per half P1 cerebellum (every other section). TigreDragon-DTA/+ animals were used as controls (n=4/genotype, P=0.029, Mann–Whitney unpaired test). Data are mean± s.d. EGL, external granule layer; NTZ, nuclear transitory zone; RL, rhombic lip; VZ, ventricular zone. Scale bars: 100 μm.
One complication of using Atoh1-driven transgenes to target cells in the cerebellum is that they are also expressed in many precerebellar neurons in nuclei of the hindbrain at E10-E13, as well as ectopic expression elsewhere in the brain and expression in other organs. Therefore, we used an intersectional approach to limit the expression of DTA to cells in the cerebellum and only a few nuclei in the isthmus in the midbrain-hindbrain junction (Machold and Fishell, 2005) by incorporating our En1Cre knock-in line (Kimmel et al., 2000). This line expresses Cre in the midbrain and rhombomere 1 of the hindbrain starting at E8 and thus includes the rhombic lip (Fig. 3A). By using Atoh1-tTA/+; En1-Cre; TigreDragon-DTA/+ mice and providing Dox starting at E13.5 to shut down tTA activity in granule cell precursors (see Fig. 2B), we restricted DTA expression primarily to the eCN (Fig. 3B-E; EC-eCN-DTA model: EC designates En1Cre). Analysis of E13.5 embryos before Dox was administered showed ectopic cell death specifically in the region of the rhombic lip, where cells first turn on the Atoh1-tTA transgene in EC-eCN-DTA mice (marked by TUNEL staining, Fig. 3B-E). As En1Cre turns on earlier than Atoh1-Cre in the eCN precursors, cell death was observed in the rhombic lip rather than in the NTZ (where it was seen in AC-eCN-DTA embryos). Quantification of cell death revealed a 14.20±2.43-fold increase in TUNEL+ particles in the rhombic lip region of the EC-eCN-DTA embryos compared with controls (TigreDragon-DTA/+ or Atoh1-tTA/+; TigreDragon-DTA/+, n=3/condition, Fig. 3F). Furthermore, unlike in AC-eCN-DTA embryos, no tdTom+ cells were detected in the NTZ, consistent with the majority of eCN being ablated before tdTom is induced as Cre is present before the Atoh1-tTA transgene is expressed (Fig. 3E). In addition, at E13.5 we observed tdTom+ cells in the hindbrain posterior to rhombomere 1 that belong to the Atoh1-lineage and do not express En1 or Cre (Machold and Fishell, 2005) (Fig. 3D,E). At P30, the number of eCN, revealed by counting the large NeuN+ neurons in the cerebellar nuclei, was reduced by ∼60% in EC-eCN-DTA cerebella compared with the controls (Fig. 3G). As a consequence of the embryonic killing of the eCN, the size of the cerebellum was reduced by ∼40% (Fig. 3H-J). The decrease in cerebellum size at P30, and thus growth during development, was greater with the intersectional EC-eCN-DTA model than with the AC-eCN-DTA model (Willett et al., 2019), likely because in the latter some eCN do not express DTA because Atoh1-Cre is only transiently expressed in eCN precursors and once it is expressed it must first delete the floxed tdTom gene in the TigreDragon-DTA allele to allow DTA to be expressed and kill cells. Consistent with this, in AC-eCN-DTA E13.5 embryos tdTom+ cells were detected in the NTZ and granule cell precursors (Fig. 2). Comparison of the intersectional approach (Atoh1-tTA and En1Cre) with the double Atoh1-driven transgenes (Atoh1-tTA and Atoh1-Cre) demonstrates the unique utility of each approach, with the En1 and Atoh1 intersectional approach achieving more precise cell targeting of DTA, and also earlier and therefore more widespread killing of eCN. Furthermore, analysis of TigreDragon-DTA/+ and Atoh1-tTA/+; TigreDragon-DTA/+ mice demonstrates that the transgene does not have leaky expression of DTA when the tdTom STOP sequence is present, whereas analysis of either of the control transgenic embryos with Cre alone (Atoh1-Cre or En1Cre) demonstrates lack of leaky expression of the recombined allele in the absence of tTA (Figs 2C-E and 3B-D).
Fig. 3.

Intersectional capabilities of the TigreDragon-DTA model. (A) Schematic of En1-lineage (based on Sgaier et al., 2007) and Atoh1 expression (based on Machold and Fishell, 2005). Striped regions represent hindbrain nuclei that are outside of the cerebellum. (B-E) Representative sagittal sections showing the cerebellar anlagen at E13.5 in control embryos (either TigreDragon-DTA/+, En1Cre/+; TigreDragon-DTA/+ or Atoh1-tTA/+; TigreDragon-DTA/+) and EC-eCN-DTA embryos, in the absence of Dox. Asterisks indicate the loss of tdTom+ cells in the NTZ and the two hindbrain nuclei targeted by the intersectional approach; arrowheads point to the hindbrain nuclei that are not affected. The boxed areas are shown magnified in the right panels, rotated counterclockwise. These insets show cell death (TUNEL+) in the rhombic lip region (RL). Arrows show the direction of the tangential migration as the cells exit the RL. Dotted lines indicate the RL region. (F) Quantification of cell death in the RL of E13.5 embryos detected by TUNEL (n=3/genotype, P=0.001, Mann–Whitney unpaired test). (G) Quantification of large NeuN+ cells from every other sagittal section from half P30 cerebella shows the number of excitatory cerebellar nuclei in EC-eCN-DTA animals compared with controls (n=3 for both genotypes, P=0.029, Mann–Whitney unpaired test). (H,I) Representative images of the P30 cerebella of EC-eCN-DTA (I) and littermate control (H). (J) Quantification of cerebellar area of midline sections shows reduction in cerebellar area after eCN ablation. TigreDragon-DTA/+ or Atoh1-tTA/+; TigreDragon-DTA/+ animals were used as controls for the quantifications shown in F,G and J. Data are mean±s.d. EGL, external granule layer; NTZ, nuclear transitory zone; RL, rhombic lip; V, ventricle; VZ, ventricular zone. Scale bars: 100 μm.
To further demonstrate the utility of the TigreDragon-DTA line we bred it with mice bearing a left-lateral plate mesoderm-specific Pitx2-Cre (Shiratori et al., 2006) and either one of two possible cartilage-specific (r)tTAs: Col2a1-tTA (Rosello-Diez et al., 2018) (for E15.5 experiment, no Dox added) or Col2a1-rtTA (Posey et al., 2009) (for P1 experiment, Dox added at E12.5) to achieve unilateral ablation in the cartilage of the left limbs (PC-Cart-DTA model: PC designates Pitx2-Cre and Cart designates cartilage-specific expression using Col2a1-(r)tTA; Fig. 4A). At E15.5 (i.e. ∼3 days after activation), TUNEL staining revealed increased cell death in the left cartilage compared with the right one (Fig. 4B), which was also seen at P1 (Fig. 4D). Quite strikingly, maintenance of the injury until P0 led to the generation of acellular gaps in the left cartilage (dashed lines, Fig. 4C, n=3). This injury led to a significant left-right limb asymmetry at P0, with the left bones 10-15% shorter than the contralateral ones (Fig. 4E).
Fig. 4.
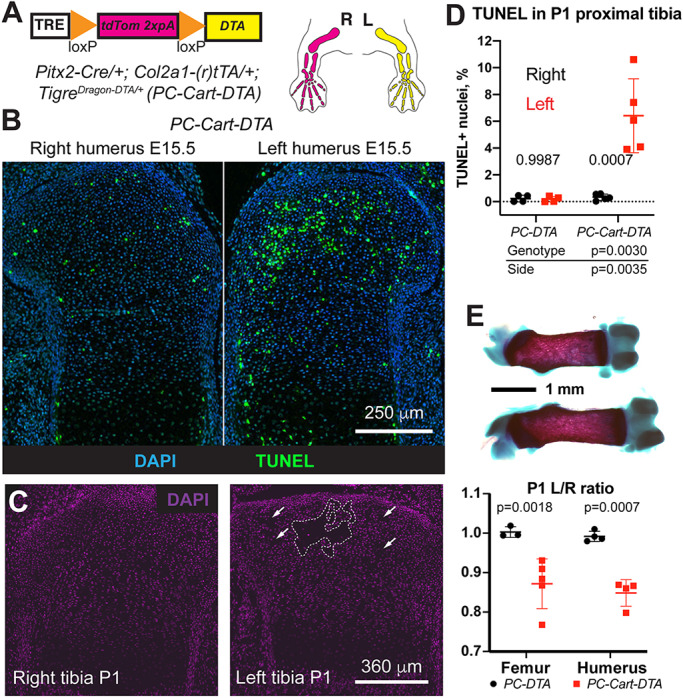
TigreDragon-DTA allows generation of structural damage and reduced local cell density due to cell death in tissues with low cell motility such as cartilage. (A) Schematic of the unilateral cartilage-specific cell ablation model. Col2a1-tTA (Tet-Off) starts to be expressed in the limbs at ∼E12.5, the same stage at which Dox was provided in the Tet-On model to activate Col2a1-driven rtTA (and maintained until collection). (B) TUNEL staining on E15.5 proximal humerus sections shows increased apoptosis in the left cartilage compared with the right. (C) One week of continuous DTA expression leads to the generation of acellular gaps in the left cartilage (dashed lines and arrows). (D) Quantification of TUNEL+ cells in cartilage at P1 (n=4 PC-DTA and 5 PC-Cart-DTA). Two-way ANOVA results are shown. The bottom table shows the P-values for the different variables. P-values within the graph are for Sidak's multiple comparisons test. (E) Cell death in the cartilage leads to impaired left-bone growth compared with the contralateral one at P1 (representative femora shown in top panel). Bone length ratios were analyzed by a mixed effects model (pGenotype <0.0001), as some samples lacked matching femur-humerus measurements. P-values for Sidak's multiple comparisons post-hoc test are shown in the chart. Data are mean±s.d.
With toxins as powerful as DTA, it is important to target only the population of interest, which is one of the main advantages of our intersectional system. Indeed, Pitx2-Cre is expressed not only in the left lateral plate mesenchyme, but also in some regions of the heart (Shiratori et al., 2006), which can lead to unwanted effects. For example, if expression of diphtheria toxin receptor is activated by Pitx2-Cre, injection of diphtheria toxin leads to cell death not only in the left limb mesenchyme, but also in the heart, affecting survival of the experimental animals (Fig. S1A and Rosello-Diez et al., 2017). Importantly, with our intersectional system, almost no cell death was detected in the hearts of PC-DTA controls or Dox-treated PC-Cart-DTA embryos (Fig. S1B-D), confirming the validity of the intersectional approach and lack of DTA expression in the absence of active (r)tTA. In summary, combining our TigreDragon-DTA line with different pairs of the available Cre and (r)tTA lines provides a valuable resource to selectively ablate cells with high spatiotemporal precision.
Unilateral mis-expression of connective tissue growth factor in the growing skeletal elements leads to bone asymmetry
We next searched for a GOI that could provide a versatile tool for transiently manipulating a variety of biological processes, such as development, homeostasis and repair. We reasoned that a molecule capable of interacting with several of the most common signaling pathways would be an ideal candidate. Connective tissue growth factor (CTGF) has been described to exert such a pleiotropic effect (Takigawa, 2013). Indeed, CTGF can interact with several signaling receptors, is considered as a hub in a network of molecular interactions, and plays a role in the harmonization of cartilage and bone development (Kubota and Takigawa, 2015), cardiovascular function (Ponticos, 2013), fibrosis, tendon healing (Tarafder et al., 2017), etc. We thus generated a TigreDragon-Ctgf mouse model (see Materials and Methods) and combined it with mice bearing left-lateral plate mesoderm-specific Pitx2-Cre (Shiratori et al., 2006) and a cartilage-specific Col2a1-tTA (Tet-Off) (Rosello-Diez et al., 2018) to achieve unilateral expression of CTGF in the cartilage of the left limbs (PC-Cart-Ctgf model; Fig. 5A). By P0-P1, whereas the expression of Ctgf (Ccn2) in the distal cartilage of the right PC-Cart-Ctgf radius was very similar to the expression in wild-type animals (Fig. 5B,C), the expression in the left PC-Cart-Ctgf element was more widespread (Fig. 5C). Interestingly, the ectopic expression of CTGF led to reduced growth of the left bones as early as E17.5, including deformation of the radius and ulna at or after P5 (Fig. 5D). Although mechanistic studies will be reported elsewhere, we are currently investigating whether this phenotype is related to increased vascular invasion and altered osteoclast activity, similar to that reported upon increased VEGF activity (Maes et al., 2010). Given its strong effects in a plethora of tissues and processes, we expect the TigreDragon-Ctgf mouse line to become a valuable tool for life scientists working on diverse topics.
Fig. 5.

Unilateral overexpression of CTGF leads to bone asymmetry. (A) Schematic of the PC-Cart-Ctgf model to achieve mis-expression of Ctgf in the cartilage of the left limb. Note that Col2a1-tTA (Tet-Off system) is used in this model, such that tTA is active without Dox (starting at ∼E12.5). (B,C) In situ hybridization for Ctgf mRNA in the distal radius cartilage from wild-type (WT) (B) and PC-Cart-Ctgf mice (C) at P0-P1 (n=3). (D) Representative examples of skeletal preparations of left (L) and right (R) PC-Cart-Ctgf bones at the indicated stages (n=7 at E15.5, 6 at E17.5, 4 at P1, 5 at P5-P7).
A model for reversible CTGF mis-expression in the growing skeletal elements reveals different outcomes of transient versus permanent expression
Besides its intersectional quality, a key aspect of the Dragon system is its reversibility. As a second proof-of-principle for testing this application, we repeated the PC-Cart-Ctgf experiment and supplied Dox to shut down tTA expression at E17.5 (Fig. 6A), soon after the phenotype is obvious (Fig. 5D). As expected, Ctgf ceased to be overexpressed in the cartilage by P0-P1 (Fig. 6B). Moreover, when the extent of asymmetry was compared between continuous and transient overexpression, we found that the latter led to a much milder asymmetry than the former (Fig. 6C). The transient expression experiment thus suggests that there is a catch-up mechanism that allows limb growth to recover, and that would not have been discovered by irreversible mis-expression, highlighting the utility of the new mouse lines.
Fig. 6.

Different outcomes of transient versus continuous expression of Ctgf in the cartilage of the growing bones. (A) Schematics of the transient injury experiment. (B) RNA in situ analysis of expression of Ctgf in the proximal cartilage of left and right tibia of PC-Cart-Ctgf P0 pups, 2 days after Dox administration at E17.5 (n=3). (C) When transgenic Ctgf expression was shut down by Dox treatment in the PC-Cart-Ctgf model (Dox at E17.5, analysis at P0-P1), the asymmetry was not as extreme as in the case of continuous Ctgf mis-expression (no-Dox condition). In the graphs, quantification of left/right ratio of bone length is shown for control (tTA–) and experimental animals (Exp; tTA+), treated or not with Dox. Analysis by two-way ANOVA for Genotype and Treatment. P-values for Sidak's multiple comparisons post-hoc test are shown. As we used an internal ratio (left/right length), data from femora and humeri were pooled together. They were treated as independent samples because Pitx2-Cre has been shown to recombine with different efficiencies in forelimb and hindlimb (Rosello-Diez et al., 2018). Data are mean±s.d.
The TigreDragon lines are effectively activated by (r)tTA and Cre, and are not leaky
We then performed in-depth characterization of the described lines, to ensure they were tightly and efficiently regulated in a variety of scenarios. In previous studies we showed that TRE-driven transgenes are highly expressed from the Tigre locus when insulator elements and a woodchuck virus RNA stabilizing sequence are included (Madisen et al., 2015; Rosello-Diez et al., 2018). However, it has not been tested whether the TRE in the TigreDragon lines drives expression of GOIs at a high level in the presence of other TREs that could compete for the transcriptional machinery and rtTA. This is important because one possible non-intersectional application of our system is to combine it with an (r)tTA and a TRE-driven Cre to achieve expression of both Cre and the GOI in a tissue during the time (r)tTA is activated. To address potential competition between TRE-driven transgenes, we generated mice bearing a TRE-Cre (Perl et al., 2002), our Col2a1-tTA allele – which initiates expression in the cartilage at ∼E12.5 (Rosello-Diez et al., 2018) – and the TigreDragon-p21 allele (TC-Cart-p21 model, Fig. 7A,B). We analyzed the mice at P0 in the absence of Dox, such that the tTA was active for at least a week before analysis. In TigreDragon-p21 mice that lacked both tTA and TRE-Cre, no tdTom or p21 expression was detected in the target tissue (or other tissues), confirming that there is no spontaneous expression of the Dragon allele (Fig. 7C,C′, left). In the presence of only tTA, ectopic expression of almost only tdTom was found in the cartilage (Fig. 7C, middle), and the intensity of the few p21+ cells was barely above background (Fig. 7C′, middle, D), revealing that there is negligible read-through activity past the STOP cassette. Finally, when both TRE-Cre and tTA were present along with the TigreDragon-p21 allele, we observed only p21 expression, demonstrating a full switch from TigreTRE-driven tdTom to p21 expression (Fig. 7C,C′,D). As no cells expressed only tdTom in this genetic combination, this result indicates that the two TRE-driven alleles are both effectively induced.
Fig. 7.
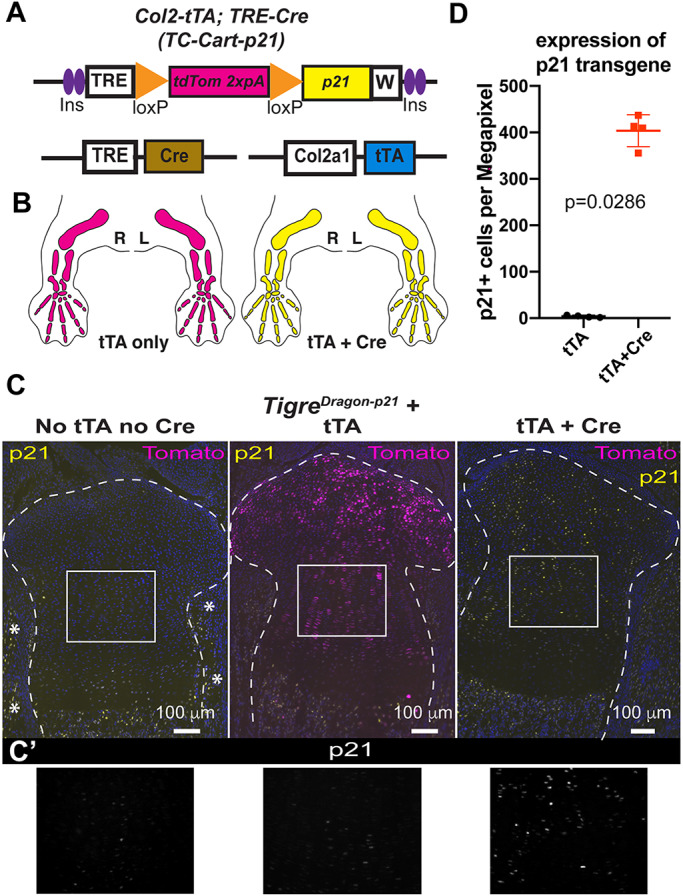
The Dragon lines are effectively activated by tTA and Cre, and not leaky. (A) Depiction of the alleles that a TC-Cart-p21 model is composed of (see text for details). (B) Prediction of expression domains (color code as in A) in the presence of Col2a1-tTA but absence of the Cre allele (left), and when both Cre and tTA alleles are present along with Dragon-p21 (right). Right (R) and left (L) limbs are shown. (C) Representative micrographs of immunohistochemistry for the indicated proteins on proximal tibia sections from newborn littermates of the indicated genotypes (TigreDragon-p21 is common to all three). Dashed lines delimit the cartilage. Asterisks indicate endogenous expression of p21 in muscle and perichondrium. No Dox was provided for these experiments. (C′) 1.8× magnification of boxed areas in C. Only p21 staining is shown. (D) Quantification of p21 expression in control and experimental mouse sections (n=4 each). P-value for unpaired Mann–Whitney test is shown. Data are mean±s.d.
A germline-recombined version of the TigreDragon allele is controlled by (r)tTA only
In cases where intersectionality is not a priority, to avoid the extra breeding steps required to introduce a third allele such as TRE-Cre, we have also generated a germline-recombined version of the TigreDragon-p21 allele (TigreTRE-p21 or p21rec, Fig. 8A) that only requires (r)tTA activity to express p21. To characterize this allele, we generated mice containing Col2a1-rtTA, p21rec and Pitx2-Cre (the latter for control purposes, Cart-p21 model). As expected, when we provided Dox to induce rtTA activity at E12.5, as p21 expression does not require intersection with Cre activity, there was no significant left-right difference in the number of chondrocytes expressing p21 (Fig. 8B,C). Moreover, a higher percentage of chondrocytes expressed p21 in the limbs (∼80%, Fig. 8B,C) than in the left limb of the intersectional approach [50-60% in Dox-treated Pitx2-Cre/+; Col2a1-rtTA; TigreDragon-p21 mice (Rosello-Diez et al., 2018)]. This new p21rec strain and similar derivatives that could be generated from our DTA and Ctgf Dragon alleles will be useful to interfere with cell functions in a variety of contexts, either by using existing tissue-specific (r)tTA mouse lines or Cre-dependent (r)tTAs activated by tissue-specific Cre.
Fig. 8.

A germline-recombined version of the TigreDragon-p21 allele becomes responsive to active (r)tTA only. (A) Depiction of the recombined allele (p21rec) and how it works in the presence of Col2-rtTA (Cart-p21 model). Note that the crosses include Pitx2-Cre for control purposes, even though it is not necessary for p21 expression. (B) Representative micrograph showing p21 expression in left and right proximal tibial cartilage at E16.5, 4 days after Dox administration. Dashed outline indicates the proliferative zone (PZ). (C) Quantification and statistical analysis of p21+ cells in the PZ. Data points are shown. P-value for Wilcoxon matched-pairs signed rank test is shown (n=3).
The Dragon targeting vectors can be used for in ovo electroporation
Lastly, we tested the possibility of using the TigreDragon tools in animal models other than the mouse. The chicken embryo has been a classical model for cell and developmental biology, immunology, virology and cancer, and is experiencing a revival since the introduction of modern genetic and imaging tools (Kain et al., 2014; Rashidi and Sottile, 2009; Stern, 2005). In ovo electroporation enables the introduction of exogenous genetic constructs in multiple locations of the embryo, in order to perform gain- and loss-of-function studies, lineage tracing, live imaging, etc. (Morin et al., 2017; Scaal et al., 2004). Therefore, we electroporated the right half of the neural tube of chicken embryos at Hamilton-Hamburger (HH) stage 14-17 (Hamburger and Hamilton, 1992) with a mix of pCAGGS-H2B-TagBFP (to visualize electroporated cells), pCAGGS-rtTA-IRES-Cre (to constitutively express rtTA and Cre) and a Tigre targeting vector (Ai62) bearing a TRE-loxSTOPlox-tdTomato cassette (Madisen et al., 2015). Dox was provided (1 μg per egg) at 16 h after electroporation, and at 24 h after electroporation, we observed robust and consistent electroporation (blue signal, Fig. 9A) in the right neural tube of most surviving embryos, as well as strong red signal in the electroporated area of Dox-treated embryos (Fig. 9A,C, n=11). Embryos not treated with Dox did not show any tdTom signal (Fig. 9B, n=3).
Fig. 9.

The Dragon vectors can be used for in ovo electroporation. (A) Schematic of the electroporation procedure in the neural tube of chicken embryos and representative examples of the outcome (micrographs taken on a fluorescence dissecting microscope). (B,C) Individual and merged channels of transverse neural tube sections (dashed lines) from electroporated chicken embryos, either untreated (B, n=3) or Dox-treated (C, n=11), 24 h after electroporation. In order to be able to visualize the native fluorescence of BFP, we used a green nuclear stain (SYTOX Green).
Lastly, we took advantage of chicken embryo accessibility to try an orthogonal approach. In this case, only pCAGGS-rtTA and the Tigre construct were electroporated, whereas the Cre was added as a fusion protein (TAT-Cre). The TAT peptide allows the Cre to cross membranes (Gitton et al., 2009), such that when beads loaded with TAT-Cre are implanted near the electroporated area, the Cre diffuses a few cell diameters away. The Cre then activates the GOI only in the electroporated cells that are close to the bead, and only in response to Dox (Fig. 10A,B, n=6). Control embryos electroporated with the full mix but without the bead (n=5, not shown), implanted with PBS-soaked beads (Fig. 10C, n=5) or electroporated with a mix devoid of rtTA and implanted with TAT-Cre beads (Fig. 10D, n=5) did not show any tdTom+ cells.
Fig. 10.

Orthogonal Dragon-driven gene mis-expression combining electroporation and implantation of Cre-soaked beads. (A) Schematic of the procedure of electroporation followed by bead implantation. (B) Implantation of TAT-Cre beads (3 mg/ml) activates tdT expression in some of the electroporated cells (TagBFP+) close by, but not in cells further than ∼100 μm from the bead. Native expression of TagBFP is shown. (C,D) Control experiments with PBS-only beads and a full electroporation mix (C, n=5) and Cre-beads in the presence of an electroporation mix lacking the rtTA plasmid (D, n=5). Scale bars in the right panels of B apply to C and D.
DISCUSSION
In this report, we have expanded and further tested the repertoire of TigreDragon intersectional tools and demonstrate their advantages for in vivo studies. Given that there are currently more than 3,000 Cre/CreER lines (http://www.informatics.jax.org/downloads/reports/MGI_Recombinase_Full.html), hundreds of which are available in repositories, and >200 (r)tTA lines (almost half of them in repositories, see https://www.tetsystems.com/fileadmin/downloads/transgenic_mouse_lines/18-06-07_table_1_ta.pdf), the potential combinations with the tools we describe here are numerous, allowing for very nuanced studies. As these numbers keep increasing, we anticipate that the TigreDragon system will be useful to many developmental biologists and other life scientists across different fields. The three main mouse lines described here (TigreDragon-p21, TigreDragon-DTA and TigreDragon-Ctgf) are being imported by the JAX repository (stocks # 034777-034779; The Jackson Laboratory) to make them available for the scientific community. It should be noted that these lines can be maintained in homozygosity with no apparent phenotype, although after several generations of inbreeding some of the homozygous animals show reduced fertility. Our labs mostly maintain these lines in an outbred Swiss Webster background. No parent-of-origin effects have been observed for these alleles. The DNA constructs are in the process of being submitted to Addgene (#140894-140896).
TigreDragon-DTA
We have shown, both in cerebella and limbs, that the TigreDragon-DTA line can be used for specific ablation of particular cell populations, restricted to a temporal window of interest. Coupled to the vast number of possible combinations of Cre and (r)tTA lines, the TigreDragon-DTA strain will be a valuable resource to ablate cells at any developmental stage and location, allowing for systemic and local responses to cell loss to be studied, including the activation of signaling pathways in response to altered cell density and regenerative mechanisms to replenish cells that are lost.
TigreDragon-Ctgf
CTGF is a pleiotropic molecule, which makes it difficult to elucidate its specific roles by traditional overexpression approaches in which several locations and cell types are affected. Conversely, the intersectionality of our approach enables targeting more precisely to a defined region, during a time window of interest. This approach will enable the elucidation of tissue-specific roles of CTGF, which should positively impact on regenerative medicine applications, tissue engineering and developmental biology research.
Avian experiments
Our results indicate that the plasmids we have generated for mouse embryonic stem (ES) cell targeting can be directly used for efficient in ovo electroporation, opening the door for precise gene manipulation in other animal models. The use of tissue-specific rather than ubiquitous promoters would make the system more versatile in the future. The orthogonal approach we describe using TAT-Cre enables a number of experimental approaches with high spatial and temporal precision. For example, after electroporation of the rtTA and the Dragon construct at an early stage, one can implant Cre-loaded beads in a very localized region of the embryo when it is still accessible, and defer Dox administration to a later time point, such that the GOI can be activated even if the embryo has sunk or turned in the egg.
Comparison with existing systems
Our system complements some of the existing sophisticated mouse genetic tools developed by the community over the last few years. For example, Belteki et al. (2005) combined tissue-specific Cre with Cre-inducible rtTA and a TRE-controlled transgene to achieve conditional, inducible and reversible transgene expression. However, that system is not intersectional, as the specificity depends only on the promoter driving Cre. Using the same number of transgenes (three) our strategy achieves the additional feature of refined spatial control. A similar system reduced the number of transgenes by making the rtTA cDNA and the TRE-controllable transgene part of a bicistronic cassette (Haenebalcke et al., 2013). However, the cassette is targeted to the Rosa26 locus, where the TRE is prone to epigenetic inactivation (Gödecke et al., 2017). Depending on the specifics of the study being undertaken, our tools should allow approaches that were technically hindered before.
With regards to more recent tools involving CRISPR/Cas9 approaches, overexpression of endogenous loci can be achieved via a nuclease-defective Cas9 fused to a transactivator domain in a structure-optimized manner (Konermann et al., 2015). This strategy could be used to achieve results similar to ours, if Cas9 were under the control of TRE with the sgRNAs in tandem, as recently described (Jo et al., 2019). However, such a strategy would not be intersectional, nor would it allow for mis-expression of genes not present endogenously, such as DTA or genes harboring coding mutations related to human disease. In summary, our approach fills a new niche in the ecosystem of genetic tools, thus enabling new types of studies.
Limitations and future directions
One potential limitation of our vectors is that, whereas the constructs work very well for transfection in vitro using HEK cells (not shown) and for chicken embryo electroporation, they are probably not suitable for efficient viral transduction, as they tend to be large (>10 kb) because of the inclusions of insulators and RNA-stabilizing sequences.
Moreover, with the current system it is not possible to trace the lineage of the cells where the intersectional activation took place, as the transgene stops being expressed when the (r)tTA is shut down or the cells transition to another fate where (r)tTA expression is lost. Future versions could be designed to express Flp recombinase in addition to the GOI, and combined with Flp-reporters to indelibly label the lineage of the targeted population.
Lastly, we envision that the use of CreER instead of Cre will introduce an additional level of control. For example, the CreER could be used to prime a stem cell population shortly after an injury, such that the descendants lose the loxSTOPlox cassette. In this scenario, only the descendants that differentiate into an (r)tTA-expressing cell population of interest would be targeted, as opposed to the whole set of descendants.
MATERIALS AND METHODS
Generation and maintenance of mouse lines
The generation of the TigreDragon-p21 mouse line was as previously described (Rosello-Diez et al., 2018). To generate the TigreTRE-p21 mouse line (aka p21rec), TigreDragon-p21 males were crossed with germline Cre-expressing females to delete the floxed cassette. The Cre allele was subsequently eliminated during the breeding to generate TigreTRE-p21 homozygous animals.
To generate the TigreDragon-Ctgf mouse line, the NruI-STOP-loxP-tdTomato-SnaBI fragment in the Ai62(TITL-tdT) Flp-in replacement vector (Madisen et al., 2015) was replaced by a custom NruI-tdTomato-STOP-loxP-MluI-HpaI-SnaBI cassette, to generate an empty DRAGON vector. A PCR-amplified Kozak-Ctgf cassette was subsequently cloned into the MluI and SnaBI sites to generate the DRAGON-Ctgf vector. This vector was then used for recombinase-mediated cassette exchange into Igs7-targeted G4 ES cells (Madisen et al., 2015). Two successfully targeted clones (#1 and #2) were injected into C2J blastocysts to generate chimeras, obtaining 11 chimeric males that survived (out of 30 animals born) with 25-85% chimaerism. One male from clone #1 and three males from clone #2 were crossed to Swiss Webster mice (Charles River) to assess germline transmission. Only clone #2 was successfully transmitted and used to establish the new mouse line.
A similar process was used to generate the TigreDragon-DTA mouse line, with the only difference being that an attenuated version of DTA (tox176) was used to PCR-amplify the Kozak-DTA cassette. Two successfully targeted clones (#8 and #13) were injected into C2J blastocysts to generate chimeras, obtaining 34 chimeric males that survived (out of 52 animals born) with 35-100% chimaerism. Four males from clone #8 and two from clone #13 were used to assess germline transmission and establish the new mouse line.
All mouse lines were maintained on an outbred Swiss Webster background. Atoh1-tTA, En1Cre and TetO-Cre were genotyped as described (Kimmel et al., 2000; Perl et al., 2002; Willett et al., 2019). All reagents and resources are shown in Table S1. The DNA primers and programs used for genotyping the rest of alleles and transgenes are shown in Table S2.
Animal experiments
All mouse work was carried out according to institutional guidelines after obtaining their approval (Sloan Kettering Institute Institutional Animal Care and Use Committee and Monash University Animal Ethics Committee). For time-mated crosses, vaginal plugs were checked in the mornings, and noon of the day the plug was detected was considered E0.5. When Dox was needed, it was added to the drinking water. To activate Col2a1-rtTA, the final concentration was 1 mg/ml, including 1% sucrose to increase palatability. To inactivate Col2a1-tTA and Atoh1-tTA, the final concentration of Dox was 0.02 mg/ml.
Upon reception, chicken eggs were kept at 18°C for up to 2 weeks. Two and half days before the experiment, they were transferred to a watered incubator at 37°C with forced ventilation.
In ovo electroporation
In ovo electroporation was performed as previously described (Blank et al., 2007). Briefly, on the day of the experiment, eggs were laid out horizontally; 3 ml of albumin were removed from the blunt end of each egg, and a window was opened on top with blunt forceps. A few drops of Ringer's solution containing Penicillin-Streptomycin (Gibco, 15140122, diluted 1/50) were pipetted on top, and the membranes removed with a hooked needle. Indian ink (drawing ink A, Pelikan, 201665, diluted 1/100 in Ringer's solution) was injected under the vitelline membranes to provide contrast. A pulled glass capillary (Harvard Apparatus, GC120T-10) and a mouth pipette were used to inject into the caudal neural tube the electroporation mix [1 μg/μl each plasmid in injection solution: high viscosity carboxymethylcellulose 0.33% (Sigma-Aldrich); Fast Green 1% (Sigma-Aldrich); MgCl2 1 mM in PBS]. Custom-made electrodes (using 0.5-mm wide platinum wire for the positive electrode and a tungsten wire for the negative one) were placed flanking the embryo, and a pulse generator (Intracel TSS20 with current amplifier) used to deliver three square pulses (27 V, 30 ms ON, 100 ms OFF). After electroporation, 200 μl of doxycycline solution (5 ng/μl in Ringer's solution) was micropipetted underneath the embryo. Eggs were then sealed with tape and incubated for 24 h. See Table S1 for full reagent and resource details.
Bead implantation
Affigel blue beads were soaked for at least 30 min in a solution of TAT-Cre (0.5 mg/ml in PBS), and rinsed in PBS before grafting. Implantation was performed as previously described (https://bio-protocol.org/e963) using a tungsten needle to open up a small section of the neural tube, and Dumont #5 forceps to insert the bead inside the socket.
Cloning of pCAGGS-rtTA-IRES-Cre
The vector for constitutive expression of rtTA and Cre was built via Gibson cloning using Addgene #20910 (digested SmaI-PacI) as backbone, and the following fragments: an IRES sequence amplified from Addgene #102423 using primers CTTGACATGCTCCCCGGGTAACCCCTCTCCCTCCCCCCC and GCTCCATTCATTTATCATCGTGTTTTTCAAAGGAAAACCACG, and a Cre sequence amplified from genomic DNA of a Cre+ mouse with primers AAACACGATGATAAATGTCCAATTTACTGACCG and ATGCTCTCGTTAATCTAATCGCCATCTTCCAG. The molecular assembly was carried out using the NEBuilder HiFi DNA Assembly cloning kit (New England BioLabs) following the manufacturer's instructions.
Sample collection and processing
Mouse limbs: Upon embryo or pup collection, the limbs were dissected out in cold PBS, skinned and fixed in 4% paraformaldehyde (PFA) for 2 days. No decalcification was carried out for samples P1 or younger. After PBS washes, the limbs were cryoprotected in PBS with 15% sucrose and then equilibrated at 37°C in PBS with 15% sucrose and 7.5% porcine skin gelatin (Sigma-Aldrich) until they sank. The limbs were oriented inside the wells of multi-welled plates and cooled down until solid, and then flash-frozen using a dry-ice-cold bath of isopentane (Merck). The cryoblocks were sectioned at 7 μm on a cryostat (Leica, CM3050S) onto Superfrost slides.
Mouse brains: The brains of all embryonic stages and P1 pups were dissected in cold PBS and immersion fixed in 4% PFA for 24 h at 4°C. P30 animals were transcardially perfused with PBS followed by 4% PFA, and brains were post-fixed in 4% PFA overnight. Specimens for cryosectioning were placed in 30% sucrose/PBS until they sank, embedded in optimal cutting temperature compound (OCT) using cryomoulds (Tissue-Tek), frozen in dry-ice-cooled isopentane, and sectioned at 14 μm on a cryostat (Leica, CM3050S) onto Superfrost slides.
Chicken embryos: The embryos were scooped out of the egg and dissected in cold PBS. The electroporated section of the trunk was isolated under the fluorescence scope and then fixed in 4% PFA overnight. After two PBS washes, specimens were cryoprotected in 15% sucrose and then 30% sucrose in PBS, until they sank. They were then embedded in OCT using cryomoulds (Tissue-Tek), frozen in dry-ice-cooled isopentane and sectioned at 7 μm on a cryostat (Leica, CM3050S) onto Superfrost slides. See Table S1 for full reagent and resource details.
Immunohistochemistry and TUNEL staining
Mouse brains: Sections were incubated in PBS for 10 min and then blocked with blocking buffer [5% bovine serum albumin (BSA, Sigma-Aldrich) in PBS with 0.1% Triton X-100]. Primary antibodies (MEIS2, 1:3000; NeuN, 1:1000) in blocking buffer were placed on slides overnight at 4°C, washed in PBS with 0.1% Triton X-100 (PBST) and applied with secondary antibodies (1:500 Alexa Fluor-conjugated secondary antibodies in blocking buffer) for 1 h at room temperature. Counterstaining was performed using Hoechst 33258 (Invitrogen). The slides were then washed in PBST, mounted with a coverslip and Fluorogel mounting medium (Electron Microscopy Sciences). TUNEL staining was performed before secondary antibody incubation using the Biotin-based in situ cell death detection kit (Roche) and following the manufacturer's instructions, including an Avidin-Biotin blocking step (Vector Laboratories). See Table S1 for full reagent and resource details.
Chicken embryos: Sections were incubated in PBS for 10 min and then permeabilized for 10 min with 0.5% Triton X-100 in PBS. Primary antibodies (mCherry, 1:1000; p21 1:300) in PBS with 0.1% Triton X-100 (PBST) were placed on slides overnight at 4°C, washed in PBST and applied with secondary antibodies (1:500, Alexa Fluor-conjugated secondary antibodies in PBS) for 1 h at room temperature. Counterstaining was performed using DAPI (Invitrogen). The slides were washed in PBST then mounted with a coverslip and Fluorogel mounting medium (Electron Microscopy Sciences).
In situ hybridization
The DIG-labeled RNA probe for Ctgf was prepared by in vitro transcription of a PCR-amplified template from E12.5 cDNA, using the following primers: Fwd: 5′-CAGCTGGGAGAACTGTGTAC-3′, Rev(T7): 5′-CGATGTTAATACGACTCACTATAGGGGCTCGCATCATAGTTGGGTC-3′. The hybridization procedure was previously described in Rosello-Diez et al. (2018).
Skeletal preparations
After embryo collection, the skin, internal organs and adipose tissue were removed. The samples were then fixed in 95% EtOH overnight at room temperature. To remove excess fat, the samples were then incubated in acetone overnight at room temperature. To stain the cartilage, the samples were submerged in a glass scintillation vial containing Alcian Blue solution [0.04% (w/v), 70% EtOH, 20% acetic acid] and incubated at least overnight at room temperature. The samples were destained by incubating them in 95% EtOH overnight, and then equilibrated in 70% EtOH, before being pre-cleared in 1% KOH solution for 1-10 h at room temperature (until blue skeletal elements were seen through). The KOH solution was replaced with Alizarin Red solution [0.005% (w/v) in water] for 3-4 h at room temperature. The Alizarin Red solution was then replaced with 1-2% KOH until most soft tissues were cleared. For final clearing, the samples were progressively equilibrated through 20% glycerol:80% (1%) KOH, 50% glycerol:50% (1%) KOH and finally transferred to 100% glycerol for long-term storage. See Table S1 for full reagent and resource details.
Image analysis and quantification
The regions of interest were isolated from imaged sections of the multiple models. All the parameters including the brightness, contrast, filters and layers were kept the same for all the images in the same study. Cells positive for the signal of interest were counted manually using ImageJ, after acquisition on an Imager Z1 microscope (Zeiss).
Statistical analysis
The comparisons were carried out by an unpaired Mann–Whitney test (1 variable and 2 conditions), or by two-way ANOVA (2 variables and 2 or more conditions) following a matched (paired) design when possible (indicated when not). When left and right measurements were compared within experimental animals only, Wilcoxon matched-pairs signed rank test was used. Non-parametric tests were chosen when normality could not be confirmed. All the statistical analyses were carried out using Prism7 software (Graphpad). No blinding was used except for Fig. 6C. Electroporated chicken embryos were randomly assigned to control or experimental groups. No previous power analysis was carried out to estimate the sample size.
Supplementary Material
Acknowledgements
We thank members of the Joyner and Rosello-Diez laboratories for discussions and technical help. We are grateful to Dr Miguel Torres for providing a polyclonal antibody to all isoforms of mouse MEIS2, and to Megan Davey for sharing her expertise with TAT-Cre beads. We acknowledge the assistance from Monash Animal Research Platform, Monash Micro Imaging Platform and the Sloan Kettering Institute Research Animal Resource Center and Mouse Genetics Core Facility. We also thank the Center for Comparative Medicine and Pathology and the Mouse Genetics Core Facilities and Dr Yas Furuta of Memorial Sloan Kettering Cancer Center for outstanding technical support. Plasmid pT2-CAGGS-H2B-BFP was a gift of the Marcelle lab. Addgene plasmids #20910 (PB-CA-rtTA Adv) and #102423 (PB-CAGGS-rtTA-IRES-Hygro) were donated by the Nagy and Koh labs, respectively. The Australian Regenerative Medicine Institute is supported by grants from the State Government of Victoria and the Australian Government.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Author contributions
Conceptualization: A.R.-D.; Methodology: A.R.-D., A.L.J.; Formal analysis: N.S.B., A.S., A.R.-D.; Investigation: E.A., N.S.B., X.Q., A.S., L.M., D.S., A.R.-D.; Writing - original draft: E.A., A.R.-D.; Writing - review & editing: N.S.B., A.L.J., A.R.-D.; Supervision: A.L.J., A.R.-D.; Project administration: A.L.J., A.R.-D.; Funding acquisition: A.L.J., A.R.-D.
Funding
This work was supported by grants from the Human Frontier Science Program (LT000521/2012-L and CDA00021/2019) and the Charles H. Revson Foundation (grant number 15-34) to A.R-D.; Monash University Graduate Scholarship, Monash University International Tuition Scholarship and Islamic Development Bank (600037455) funds to E.A.; Postdoctoral fellowships from New York State Stem Cell Science (C32599GG) and the National Institute of Neurological Disorders and Stroke (K99 NS112605-01) to N.S.B.; the National Institutes of Health (R21 HD083860, R37 MH085726 and R01 NS092096) to A.L.J. and a National Cancer Institute Cancer Center Support Grant (P30 CA008748-48) to Memorial Sloan Kettering Cancer Center Deposited in PMC for release after 12 months.
Supplementary information
Supplementary information available online at http://dev.biologists.org/lookup/doi/10.1242/dev.186650.supplemental
Peer review history
The peer review history is available online at https://dev.biologists.org/lookup/doi/10.1242/dev.186650.reviewer-comments.pdf
References
- Belteki G., Haigh J., Kabacs N., Haigh K., Sison K., Costantini F., Whitsett J., Quaggin S. E. and Nagy A. (2005). Conditional and inducible transgene expression in mice through the combinatorial use of Cre-mediated recombination and tetracycline induction. Nucleic Acids Res. 33, e51 10.1093/nar/gni051 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blank M. C., Chizhikov V. Millen K. J. (2007). In ovo electroporations of HH stage 10 chicken embryos. J. Vis. Exp. 9, e408 10.3791/408 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breitman M. L., Rombola H., Maxwell I. H., Klintworth G. K. and Bernstein A. (1990). Genetic ablation in transgenic mice with an attenuated diphtheria toxin A gene. Mol. Cell. Biol. 10, 474-479. 10.1128/MCB.10.2.474 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brocard J., Feil R., Chambon P. and Metzger D. (1998). A chimeric Cre recombinase inducible by synthetic,but not by natural ligands of the glucocorticoid receptor. Nucleic Acids Res. 26, 4086-4090. 10.1093/nar/26.17.4086 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dymecki S. M. and Kim J. C. (2007). Molecular neuroanatomy's “Three Gs”: a primer. Neuron 54, 17-34. 10.1016/j.neuron.2007.03.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feil R., Wagner J., Metzger D. and Chambon P. (1997). Regulation of Cre recombinase activity by mutated estrogen receptor ligand-binding domains. Biochem. Biophys. Res. Commun. 237, 752-757. 10.1006/bbrc.1997.7124 [DOI] [PubMed] [Google Scholar]
- Feil S., Valtcheva N. and Feil R. (2009). Inducible Cre mice. Methods Mol. Biol. 530, 343-363. 10.1007/978-1-59745-471-1_18 [DOI] [PubMed] [Google Scholar]
- Gitton Y., Tibaldi L., Dupont E., Levi G. and Joliot A. (2009). Efficient CPP-mediated Cre protein delivery to developing and adult CNS tissues. BMC Biotechnol. 9, 40 10.1186/1472-6750-9-40 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gödecke N., Zha L., Spencer S., Behme S., Riemer P., Rehli M., Hauser H. and Wirth D. (2017). Controlled re-activation of epigenetically silenced Tet promoter-driven transgene expression by targeted demethylation. Nucleic Acids Res. 45, e147 10.1093/nar/gkx601 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gossen M. and Bujard H. (1992). Tight control of gene expression in mammalian cells by tetracycline-responsive promoters. Proc. Natl. Acad. Sci. USA 89, 5547-5551. 10.1073/pnas.89.12.5547 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gossen M., Freundlieb S., Bender G., Muller G., Hillen W. and Bujard H. (1995). Transcriptional activation by tetracyclines in mammalian cells. Science 268, 1766-1769. 10.1126/science.7792603 [DOI] [PubMed] [Google Scholar]
- Haenebalcke L., Goossens S., Dierickx P., Bartunkova S., D'hont J., Haigh K., Hochepied T., Wirth D., Nagy A. and Haigh J. J. (2013). The ROSA26-iPSC mouse: a conditional, inducible, and exchangeable resource for studying cellular (De)differentiation. Cell Rep 3, 335-341. 10.1016/j.celrep.2013.01.016 [DOI] [PubMed] [Google Scholar]
- Hamburger V. and Hamilton H. L. (1992). A series of normal stages in the development of the chick embryo. 1951. Dev. Dyn. 195, 231-272. 10.1002/aja.1001950404 [DOI] [PubMed] [Google Scholar]
- Hillen W. and Berens C. (1994). Mechanisms underlying expression of Tn10 encoded tetracycline resistance. Annu. Rev. Microbiol. 48, 345-369. 10.1146/annurev.mi.48.100194.002021 [DOI] [PubMed] [Google Scholar]
- Hirrlinger J., Scheller A., Hirrlinger P. G., Kellert B., Tang W., Wehr M. C., Goebbels S., Reichenbach A., Sprengel R., Rossner M. J. et al. (2009). Split-cre complementation indicates coincident activity of different genes in vivo. PLoS ONE 4, e4286 10.1371/journal.pone.0004286 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jo N., Sogabe Y., Yamada Y., Ukai T., Kagawa H., Mitsunaga K., Woltjen K. and Yamada Y. (2019). Platforms of in vivo genome editing with inducible Cas9 for advanced cancer modeling. Cancer Sci. 110, 926-938. 10.1111/cas.13924 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaczmarczyk L. and Jackson W. S. (2015). Astonishing advances in mouse genetic tools for biomedical research. Swiss Med. Wkly. 145, w14186. [DOI] [PubMed] [Google Scholar]
- Kain K. H., Miller J. W., Jones-Paris C. R., Thomason R. T., Lewis J. D., Bader D. M., Barnett J. V. and Zijlstra A. (2014). The chick embryo as an expanding experimental model for cancer and cardiovascular research. Dev. Dyn. 243, 216-228. 10.1002/dvdy.24093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimmel R. A., Turnbull D. H., Blanquet V., Wurst W., Loomis C. A. and Joyner A. L. (2000). Two lineage boundaries coordinate vertebrate apical ectodermal ridge formation. Genes Dev. 14, 1377-1389. [PMC free article] [PubMed] [Google Scholar]
- Konermann S., Brigham M. D., Trevino A. E., Joung J., Abudayyeh O. O., Barcena C., Hsu P. D., Habib N., Gootenberg J. S., Nishimasu H. et al. (2015). Genome-scale transcriptional activation by an engineered CRISPR-Cas9 complex. Nature 517, 583-588. 10.1038/nature14136 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kronenberg H. M. (2003). Developmental regulation of the growth plate. Nature 423, 332-336. 10.1038/nature01657 [DOI] [PubMed] [Google Scholar]
- Kubota S. and Takigawa M. (2015). Cellular and molecular actions of CCN2/CTGF and its role under physiological and pathological conditions. Clin. Sci. (Lond.) 128, 181-196. 10.1042/CS20140264 [DOI] [PubMed] [Google Scholar]
- Machold R. and Fishell G. (2005). Math1 is expressed in temporally discrete pools of cerebellar rhombic-lip neural progenitors. Neuron 48, 17-24. 10.1016/j.neuron.2005.08.028 [DOI] [PubMed] [Google Scholar]
- Madisen L., Garner A. R., Shimaoka D., Chuong A. S., Klapoetke N. C., Li L., Van Der Bourg A., Niino Y., Egolf L., Monetti C. et al. (2015). Transgenic mice for intersectional targeting of neural sensors and effectors with high specificity and performance. Neuron 85, 942-958. 10.1016/j.neuron.2015.02.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maes C., Goossens S., Bartunkova S., Drogat B., Coenegrachts L., Stockmans I., Moermans K., Nyabi O., Haigh K., Naessens M. et al. (2010). Increased skeletal VEGF enhances beta-catenin activity and results in excessively ossified bones. EMBO J. 29, 424-441. 10.1038/emboj.2009.361 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matei V., Pauley S., Kaing S., Rowitch D., Beisel K. W., Morris K., Feng F., Jones K., Lee J. and Fritzsch B. (2005). Smaller inner ear sensory epithelia in Neurog 1 null mice are related to earlier hair cell cycle exit. Dev. Dyn. 234, 633-650. 10.1002/dvdy.20551 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Metzger D., Clifford J., Chiba H. and Chambon P. (1995). Conditional site-specific recombination in mammalian cells using a ligand-dependent chimeric Cre recombinase. Proc. Natl. Acad. Sci. USA 92, 6991-6995. 10.1073/pnas.92.15.6991 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morin V., Veron N. and Marcelle C. (2017). CRISPR/Cas9 in the Chicken Embryo. Methods Mol. Biol. 1650, 113-123. 10.1007/978-1-4939-7216-6_7 [DOI] [PubMed] [Google Scholar]
- Perl A. K., Wert S. E., Nagy A., Lobe C. G. and Whitsett J. A. (2002). Early restriction of peripheral and proximal cell lineages during formation of the lung. Proc. Natl. Acad. Sci. USA 99, 10482-10487. 10.1073/pnas.152238499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ponticos M. (2013). Connective tissue growth factor (CCN2) in blood vessels. Vascul. Pharmacol. 58, 189-193. 10.1016/j.vph.2013.01.004 [DOI] [PubMed] [Google Scholar]
- Posey K. L., Veerisetty A. C., Liu P., Wang H. R., Poindexter B. J., Bick R., Alcorn J. L. and Hecht J. T. (2009). An inducible cartilage oligomeric matrix protein mouse model recapitulates human pseudoachondroplasia phenotype. Am. J. Pathol. 175, 1555-1563. 10.2353/ajpath.2009.090184 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rashidi H. and Sottile V. (2009). The chick embryo: hatching a model for contemporary biomedical research. BioEssays 31, 459-465. 10.1002/bies.200800168 [DOI] [PubMed] [Google Scholar]
- Rosello-Diez A., Stephen D. and Joyner A. L. (2017). Altered paracrine signaling from the injured knee joint impairs postnatal long bone growth. Elife 6, e27210 10.7554/eLife.27210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosello-Diez A., Madisen L., Bastide S., Zeng H. and Joyner A. L. (2018). Cell-nonautonomous local and systemic responses to cell arrest enable long-bone catch-up growth in developing mice. PLoS Biol. 16, e2005086 10.1371/journal.pbio.2005086 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosen B., Schick J. and Wurst W. (2015). Beyond knockouts: the International Knockout Mouse Consortium delivers modular and evolving tools for investigating mammalian genes. Mamm. Genome 26, 456-466. 10.1007/s00335-015-9598-3 [DOI] [PubMed] [Google Scholar]
- Scaal M., Gros J., Lesbros C. and Marcelle C. (2004). In ovo electroporation of avian somites. Dev. Dyn. 229, 643-650. 10.1002/dvdy.10433 [DOI] [PubMed] [Google Scholar]
- Sgaier S. K., Lao Z., Villanueva M. P., Berenshteyn F., Stephen D., Turnbull R. K. and Joyner A. L. (2007). Genetic subdivision of the tectum and cerebellum into functionally related regions based on differential sensitivity to engrailed proteins. Development 134, 2325-2335. 10.1242/dev.000620 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shiratori H., Yashiro K., Shen M. M. and Hamada H. (2006). Conserved regulation and role of Pitx2 in situs-specific morphogenesis of visceral organs. Development 133, 3015-3025. 10.1242/dev.02470 [DOI] [PubMed] [Google Scholar]
- Smedley D., Salimova E. and Rosenthal N. (2011). Cre recombinase resources for conditional mouse mutagenesis. Methods 53, 411-416. 10.1016/j.ymeth.2010.12.027 [DOI] [PubMed] [Google Scholar]
- St-Onge L., Furth P. A. and Gruss P. (1996). Temporal control of the Cre recombinase in transgenic mice by a tetracycline responsive promoter. Nucleic Acids Res. 24, 3875-3877. 10.1093/nar/24.19.3875 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stern C. D. (2005). The chick; a great model system becomes even greater. Dev. Cell 8, 9-17. 10.1016/S1534-5807(04)00425-3 [DOI] [PubMed] [Google Scholar]
- Takigawa M. (2013). CCN2: a master regulator of the genesis of bone and cartilage. J Cell Commun. Signal 7, 191-201. 10.1007/s12079-013-0204-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarafder S., Chen E., Jun Y., Kao K., Sim K. H., Back J., Lee F. Y. and Lee C. H. (2017). Tendon stem/progenitor cells regulate inflammation in tendon healing via JNK and STAT3 signaling. FASEB J. 31, 3991-3998. 10.1096/fj.201700071R [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urlinger S., Baron U., Thellmann M., Hasan M. T., Bujard H. and Hillen W. (2000). Exploring the sequence space for tetracycline-dependent transcriptional activators: novel mutations yield expanded range and sensitivity. Proc. Natl. Acad. Sci. USA 97, 7963-7968. 10.1073/pnas.130192197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willett R. T., Bayin N. S., Lee A. S., Krishnamurthy A., Wojcinski A., Lao Z., Stephen D., Rosello-Diez A., Dauber-Decker K. L., Orvis G. D. et al. (2019). Cerebellar nuclei excitatory neurons regulate developmental scaling of presynaptic Purkinje cell number and organ growth. Elife 8, e50617 10.7554/eLife.50617 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeng H., Horie K., Madisen L., Pavlova M. N., Gragerova G., Rohde A. D., Schimpf B. A., Liang Y., Ojala E., Kramer F. et al. (2008). An inducible and reversible mouse genetic rescue system. PLoS Genet. 4, e1000069 10.1371/journal.pgen.1000069 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.