

Abstract
Our understanding of the neural crest, a key vertebrate innovation, is built upon studies of multiple model organisms. Early research on neural crest cells (NCCs) was dominated by analyses of accessible amphibian and avian embryos, with mouse genetics providing complementary insights in more recent years. The zebrafish model is a relative newcomer to the field, yet it offers unparalleled advantages for the study of NCCs. Specifically, zebrafish provide powerful genetic and transgenic tools, coupled with rapidly developing transparent embryos that are ideal for high resolution real-time imaging of the dynamic process of neural crest development. While the broad principles of neural crest development are largely conserved across vertebrate species, there are critical differences in anatomy, morphogenesis, and genetics that must be considered before information from one model is extrapolated to another. Here, our goal is to provide the reader with a helpful primer specific to neural crest development in the zebrafish model. We focus largely on the earliest events—specification, delamination, and migration—discussing what is known about zebrafish NCC development and how it differs from NCC development in non-teleost species, as well as highlighting current gaps in knowledge.
Keywords: zebrafish, neural crest, epithelial-to-mesenchymal transition, cell migration
INTRODUCTION
The neural crest is a transient, multipotent cell population characterized by its extensive migratory capacity and remarkable differentiation potential. Neural crest cells (NCCs) give rise to a wide variety of derivatives, including neurons and glia of the peripheral nervous system, melanocytes, and craniofacial cartilage and bone (Le Douarin and Kalcheim, 1982). Together with ectodermal placodes, cranial NCCs were key to the evolution of vertebrate-specific cranial elements, including a hinged jaw, special sense organs, and novel neural structures. These features are lacking in the protochordates (non-vertebrate members of the phylum Chordata), leading Gans and Northcutt to hypothesize that the vertebrate-specific cranial elements could be viewed as a “new head” (Gans and Northcutt, 1983; Muñoz and Trainor, 2015). It was the evolution of the specialized vertebrate head, with its bony skull protecting a big brain, that facilitated the shift from passive to active feeding behaviors and enabled the extraordinary radiation of the vertebrate lineage (Hall, 2000).
Originally identified in chick embryos by William His in 1868 (Hall, 2000), the neural crest has fascinated developmental biologists for over a century. In 1893 Julia Platt suggested the neural crest origins of the skull based on research in the mudpuppy, a common salamander species (Platt, 1893). Platt’s work was highly contentious at the time because it challenged the dogma—based on germ-layer theory—that bone and cartilage must be of mesodermal origin. Her findings were nevertheless built upon by numerous subsequent researchers, in a series of reports that were summarized in a comprehensive and influential review from Sven Hörstadius (Hörstadius, 1950). In particular, Hörstadius provided a summary of work from his own group—primarily by his student Sellman—which used the classical embryology approaches of ablation and transplantation in amphibians to begin to reveal the remarkable migratory capacity of the neural crest and its broad contributions.
It was not until the 1970s, that Nicole LeDouarin’s pioneering quail-chick chimera experiments allowed the details of the migration pathways and derivatives of NCCs to be defined (Le Douarin, 1973; Le Douarin and Kalcheim, 1982; Le Lièvre and Le Douarin, 1975). These elegant experiments utilized transplantation of quail neural crest primordia from defined anteroposterior domains of the neural tube into equivalent positions within developing chick embryos. Quail-derived NCC contributions were then tracked, originally by histological visualization of the specialized quail cell nucleolus, and in more recent years using a quail-specific antibody. The quail-chick chimera approach proved highly informative and has played a major role in revealing the neural crest’s contributions to the vertebrate body. Influential transplantation experiments were also performed by Drew Noden in the early 1980s, providing key insights into the cranial neural crest. Noden’s experiments focused on heterotopic transplants, revealing a degree of intrinsic “pre-pattern” in the cranial neural crest primordium (Noden, 1983).
In recent years, NCC development has been investigated in a wide array of vertebrate organisms, from the agnathan lamprey, to the amphibians (especially Xenopus laevis), chick, and mouse. Here we highlight the contributions of the teleost zebrafish, Danio rerio, to the study of neural crest development. Zebrafish genetics has uncovered molecular mechanisms that underlie neural crest specification and differentiation, while the optical clarity of the embryos, coupled with transgenic tools, has enabled high-resolution imaging of cellular behaviors during the epithelial-to-mesenchymal transition (EMT) and subsequent migration. It is, in particular, the capacity to combine genetics with imaging that is allowing zebrafish to make novel contributions to the neural crest field.
Neural crest development takes place concurrently with late gastrulation and neurulation through a series of sequential steps: induction, specification, delamination, migration, and differentiation. Importantly, while Xenopus, chick, and mouse are all representatives of the lobe-finned fish lineage (sarcopterygia), the teleost zebrafish is the only well-studied developmental model within the large and diverse ray-finned fish lineage (actinopterygia). Although teleost fishes do not undergo open neurulation, as in sarcopterygian species, the general pathways and mechanisms underlying the processes of neural crest development are largely conserved across vertebrates. Nevertheless, there are interesting species-specific differences. By exploring both similarities and differences, we hope to gain a better understanding of the principles underlying the establishment, migration, and differentiation of NCCs, as well as of the diversity of NCC developmental mechanisms across vertebrates.
REGIONALIZATION OF THE ZEBRAFISH NEURAL CREST
Initial reports by Raible and colleagues (1992) and Schilling and Kimmel (1994) described the development of NCCs in the trunk and head of zebrafish embryos, respectively (Raible et al., 1992; Schilling and Kimmel, 1994). These early studies of the zebrafish neural crest relied on DIC light microscopy, electron microscopy, and microinjection of fluorescent lineage tracers, as the NCC-specific transgene reporter lines that have enabled more precise visualization (Table 1) were yet to be generated, and the anti-HNK1 antibody that conveniently recognizes NCCs in many other species—including the model teleost medaka (Sadaghiani and Vielkind, 1990)—fails to label zebrafish NCCs (Metcalfe et al., 1990). These foundational studies revealed that the basic organization of zebrafish NCC migratory routes, and the array of derivatives the NCCs produce, are conserved between the teleost zebrafish and other vertebrates (Figure 1). Briefly, specific NCC populations develop along the anteroposterior (AP) axis of the developing embryo, with the cranial NCC population—the most anterior midbrain and hindbrain derived NCCs—contributing to the cartilaginous and bony elements of the jaw and skull, as well as producing neurons and melanocytes (Kague et al., 2012; Schilling and Kimmel, 1994; Wada et al., 2005). The trunk NCC population, which contributes neurons and glia to the dorsal root and parasympathetic ganglia, as well as melanocytes and other cell types, arises along the entire extent of the developing spinal cord, immediately adjacent to the developing mesodermal somites (Figure 1) (Raible et al., 1992; Raible and Eisen, 1994). Although initial reports suggested that zebrafish NCCs contribute to the fin osteoblasts and connective tissues (Kague et al., 2012; Smith et al., 1994), more recent lineage tracing data has overturned this conclusion, revealing that in zebrafish, as in sarcopterygians, trunk crest does not generate ectomesenchymal derivatives (Lee et al., 2013a, 2013b).
Table 1:
Zebrafish neural crest genes, mutants and transgenes
Fig. 1: Zebrafish neural crest regionalization and derivatives.
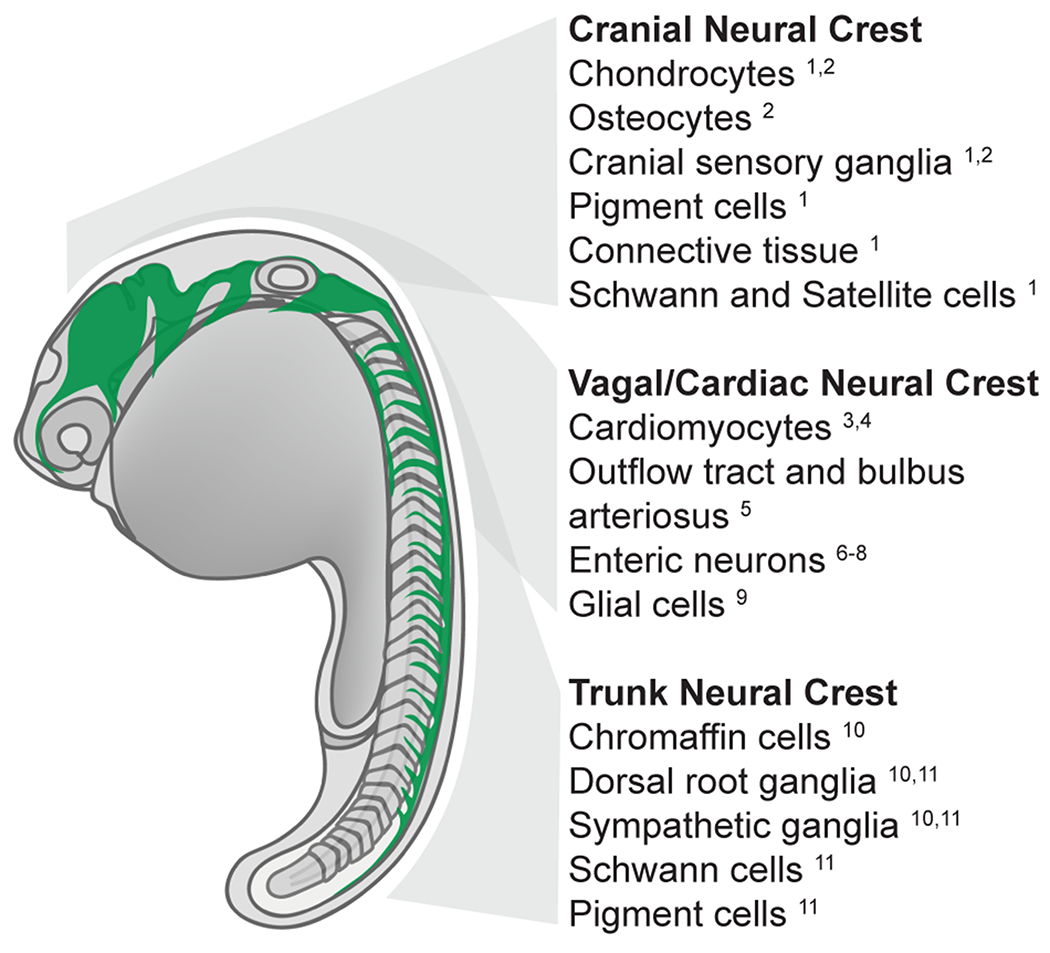
Schematic summarizing neural crest migration pathways (green) in the 22 hpf zebrafish embryo. Cell fates derived from specific anteroposterior levels are indicated. (1) Schilling et al., 1994, (2) Kague et al., 2012, (3) Li et al., 2003 (4) Sato and Yost 2003, (5) Cavanaugh et al., 2015, (6) Elworthy et al. 2005, (7) Olden et al., 2008, (8) Shepherd et al., 2004, (9) Shepherd and Eisen 2011 (10) An et al., 2002 (11) Raible et al., 1992.
The studies of the early 1990s also began to reveal zebrafish-specific aspects of NCC development. For example, zebrafish trunk NCCs migrate ventrally in streams along both medially and laterally restricted pathways, broadly similar to the NCCs of other species, but the details of the zebrafish trunk NCC migratory routes relative to the mesodermal somites differ significantly from the routes taken in sarcopterygian models (Raible et al., 1992), a topic we address in more detail below. In the cranial region, some NCCs remain laterally segregated from the developing neural tube as a coherent mass during the premigratory stages (Figure 2) and do not appear to reach the dorsal midline before migrating, likely as a consequence of teleost-specific neurulation mechanisms (Jimenez et al., 2016; Schilling and Kimmel, 1994).
Fig. 2: Zebrafish neurulation and neural crest migration pathways.
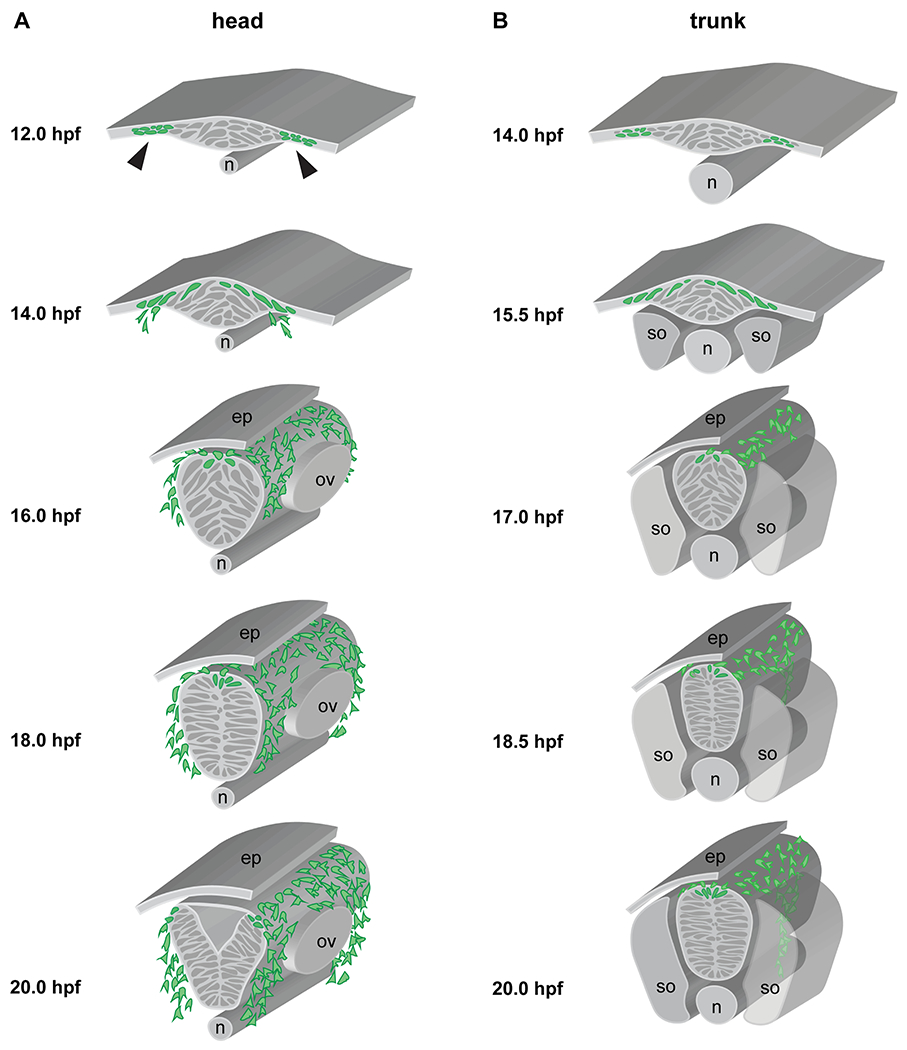
(A) Cranial neurulation and neural crest migration, schematized at the rhombomere (r)4-6 level, showing neural crest cells (green) migrating anterior and posterior of the otic vesicle towards pharyngeal arch (PA)2 and PA3, respectively. Stages are indicated, with laterally segregated cells apparent at 12 hpf (arrowheads), convergence movements forming the neural keel by 14 hpf, a neural rod at 18 hpf (with a clear midline now established), and finally a neural tube at 20 hpf. Note that the anterior terminus of the notochord lies under r4 in the zebrafish. (B) Trunk neurulation and neural crest migration, schematized at the somite 7/8 level, showing neural crest cells (green) migrating on the medial pathway adjacent to the center of each somite. Stages are indicated. n = notochord; ep = epidermis; ov = otic vesicle; s = somite.
Zebrafish cardiac and vagal crest populations also exhibit distinctive features (Hutchins et al., 2018) (Figure 1). In the chick, cardiac neural crest is thought to derive from a region between the posterior edge of the otic vesicle and the posterior limit of the third somite (Plein et al., 2015), but zebrafish reports differ on the precise origins of cardiac NCCs. Sato and Yost used both cell transplantation and lineage tracing, with a particular emphasis on labeling cells by uncaging fluorescein, to show that zebrafish cardiac NCCs contribute to the myocardium and derive from a region that extends all the way from the midbrain/hindbrain boundary to the level of the sixth somite (Sato and Yost, 2003). By contrast, results from Li and colleagues, who also labeled NCCs by uncaging fluorescein, show that NCCs posterior to the first somite do not contribute to cardiomyocytes (Li et al., 2003). Moreover, although Li and colleagues did not report the anterior limit of cardiac NCC progenitors in terms of hindbrain morphology, their data nevertheless additionally suggest that the cardiac NCC domain does not extend as far to the anterior as suggested by Sato and Yost. Of relevance, the caged fluorescein approach is known to be prone to artifact, so the results of NCC ablations should also be taken into consideration. Li and colleagues (2003) found that ablation of NCCs adjacent to somites 1-3 did not affect heart function, whereas ablation of more anterior NCC domains led to defects in heart looping and function, consistent with the zebrafish having a more limited cardiac NCC domain than the chick. Perhaps most importantly, the two reports concur that in zebrafish the NCCs contribute directly to the myocardial cell population, whereas any similar contribution in amniotes is much more contentious (Plein et al., 2015). More recently, Cavanaugh and colleagues have suggested that there are two distinct populations of NCCs that contribute to the zebrafish heart: one that migrates through pharyngeal arches 1 and 2, integrates into the heart tube and adopts a myocardial fate, and a second wave that migrates along the 6th pharyngeal arch artery, surrounds the ventral aorta and invades the bulbus arteriosus (Cavanaugh et al., 2015). While the precise origins of zebrafish vagal NCCs have not been determined, it is clear that post-otic NCCs born at the levels of somites 1-7 give rise to neurons and glia of the enteric nervous system (Hutchins et al., 2018; Shepherd and Eisen, 2011).
Efforts to generate, identify, and characterize zebrafish mutations affecting NCC derivatives have greatly advanced our understanding of the mechanisms underlying neural crest development. A screen conducted at the University of Oregon identified 29 mutations disrupting the development of NCC derivatives (Henion et al., 1996). Further, the large-scale screens conducted in Tübigen and Boston in the early-mid 1990s resulted in the identification of mutants in over 400 genes, many of which exhibited defects in neural crest development (Driever et al., 1996; Granato and Nüsslein-Volhard, 1996; Haffter et al., 1996). Since these initial genetic screens, significant work has gone into the analysis of these and other mutants (for some examples, see Table 1). Reciprocal ‘reverse genetic’ approaches have also been informative, with the development of morpholino knockdown tools in the early 2000s (Nasevicius and Ekker, 2000) providing a valuable approach to test zebrafish gene functions rapidly. More recently, the advent of CRISPR/Cas9 approaches has allowed targeted generation of novel zebrafish mutants (Irion et al., 2014), enabling genetic evaluation of candidate genes involved in neural crest development. Given the straightforward nature of this new mutagenesis technology, we can expect to see many more CRISPR/Cas mutants that impact NCC development reported and described in the coming years.
NEURAL CREST INDUCTION AND SPECIFICATION
Induction, specification, and the onset of migration of neural crest cells occur in concert with the morphogenetic processes of gastrulation and neurulation. In zebrafish and other teleosts, neurulation processes differ markedly from those observed in the sarcopterygian models. Thus, it is important to understand teleost-specific features of neurulation, and to consider how these impact neural crest development.
Zebrafish neurulation
In sarcopterygians—e.g. chick, mice, Xenopus—the majority of the epithelial neural plate undergoes “primary neurulation”, in which it folds or ‘rolls up,’ such that the lateral edges rise up and come together dorsally to form an epithelial tube with a central lumen. By contrast, the zebrafish neural plate is not strictly epithelial, nor does it fold (Papan and Campos-Ortega, 1994; Schmitz et al., 1993). Early descriptions of zebrafish neurulation referred to the process as a form of “secondary neurulation” (Papan and Campos-Ortega, 1994), a process limited exclusively to the most posterior neural tissue of sarcopterygians, which involves mesenchymal cells coalescing to produce a neural rod that subsequently cavitates to establish the lumen. However, secondary neurulation mechanisms also differ significantly from the processes of zebrafish neurulation, which involve pseudoepithelial cells and the organized establishment of a midline that allows the lumen to form. We therefore concur with those authors who suggest the term secondary neurulation be avoided in the context of zebrafish and other teleosts (Lowery and Sive, 2004).
Zebrafish neurulation begins at the tailbud stage (10 hours post fertilization; hpf) when gastrulation is reaching completion, at this point the zebrafish neural plate is a multilayered structure: in the anterior regions—fated to become fore, mid and hindbrain—the plate is 3-6 cells deep, thinning down to a single cell layer more posteriorly in the presumptive spinal cord (Clarke, 2009; Hong and Brewster, 2006). As development proceeds, neural plate cells converge, with lateral neural ectoderm cells moving towards the midline and intercalating while the most medial neural cells internalize and move ventrally. Together, these movements produce the neural keel, and subsequently the neural rod (Figure 2). It is at these same stages that molecular markers of neural crest progenitors are first expressed, implying that the first NCCs are specified as early as 11 hpf at the lateral edges of the anterior neural ectoderm (Odenthal and Nüsslein-Volhard, 1998; Thisse et al., 1995). As neurulation continues, neural ectoderm cells produce a pseudostratified epithelium, with the cells undergoing polarized cell divisions to establish a well-defined midline by 18 hpf (Buckley et al., 2013; Ciruna et al., 2006; Clarke, 2009; Tawk et al., 2007). Cells on either side of the midline then pull apart to produce the lumen of the neural tube, which despite its divergent mode of morphogenesis is now essentially equivalent to the neural tubes of other vertebrates.
Zebrafish NCCs begin to migrate away from the neural ectoderm in the cranial region by 14 hpf when the convergence processes of neurulation are still ongoing, although NCCs will also continue to emerge from the dorsal-most neural tissue after the neural tube is fully formed (Berndt et al., 2008; Jimenez et al., 2016; Schilling and Kimmel, 1994). Given the dynamic morphogenesis that occurs during neurulation, the earliest NCCs derive from cells that lie in lateral positions, with only the later NCCs deriving from the dorsal-most aspect of the neural tube. This may explain Schilling and Kimmel’s 1994 observation of coherent masses of NCCs lateral to the neural tube, a topic we return to in our discussion of EMT, below. As in other vertebrates, there is also an AP progression of NCC migration, such that the first cranial NCCs begin migration ~2 hours earlier than the most anterior trunk NCCs, with the remaining trunk NCCs commencing their migration in a distinct AP progression.
Placodes and Rohon-Beard cells
The neural plate border (NPB) in general gives rise both to NCCs and sensory cells. In the cranial region, the NPB is the source of NCCs and ectodermal placodes (Groves and LaBonne, 2014; Schlosser, 2006). The placodes are destined to produce specialized sensory structures of the vertebrate head (e.g. nose, eye, ear), and in aquatic organisms, such as zebrafish, the lateral line system (Schlosser, 2010, 2006). While placodes are not a focus of this review, it is important to be aware that NCCs collaborate with placodal cells in the formation of the cranial ganglia, which have a shared placodal/NCC origin, and during NCC migration (Theveneau et al., 2013).
The developmental and evolutionary relationship between the pre-placodal ectoderm (PPE) and neural crest has long been debated in the field. Gans and Northcutt initially proposed that these two cell types have a common evolutionary origin based on their similarities, including their migratory capacities and developmental potential (Northcutt and Gans, 1983). This hypothesis has been further supported by the suggestion that the two cell types are derived from a common neural plate border region. However, Gerhard Schlosser has proposed an alternative, “binary competence” model, which posits that competence to form neural crest and PPE is restricted to neural and non-neural ectoderm, respectively (Schlosser, 2008). This model highlights the difference between the two tissues, as revealed by developmental and comparative studies, and argues that they evolved independently. At this time, there is not sufficient evidence to conclusively decide which, if any, of these scenarios is correct. However, the ability to analyze the transcriptional state of cells at the neural border with single-cell resolution will likely prove critical to distinguishing between the models. Here, we describe the process of NCC development in accordance with the “neural plate border state” model, not as an endorsement of that particular scenario, but simply as a conceptual framework for NCC development.
In the trunk, the NPB is the source of both NCCs and dorsal sensory neurons. In zebrafish and amphibians the Rohon-Beard (RB) cells are a transient population of dorsal sensory neurons (Cornell and Eisen, 2000; Kelsh and Raible, 2002; Lamborghini, 1980; Moody, 1989; Selleck and Bronner-Fraser, 1995). The NCCs and RBs likely derive from a common progenitor cell type (Artinger et al., 1999), with their subsequent segregation mediated by Delta/Notch signaling (Cornell and Eisen, 2000). deltaA mutants exhibit supernumerary RBs and a loss of NCCs (Cornell and Eisen, 2000), and a similar phenotype is observed in mindbomb mutants, which have defects in the E3 ubiquitin ligase involved in Delta/Notch signaling (Jiang et al., 1996; Schier et al., 1996). Delta/Notch signaling in NCCs inhibits the expression of neurog1, which otherwise promotes the development of RBs at these stages (Cornell and Eisen, 2002). Reducing the levels of Neurog1 rescues NCCs in Delta/Notch signaling mutants (Cornell and Eisen, 2002), consistent with a model in which Neurog1 is incompatible with NCC fates.
Regulation of neural crest induction
During gastrulation, the neural crest, and in the cranial region the adjacent placodal cells, are induced at the interface between neural and non-neural ectoderm, a finding initially made in axolotls (Moury and Jacobson, 1990, 1989). In chick embryos, this process is largely dependent on Bmp4 and Bmp7 signaling, with both Bmps expressed at the edge of the neural plate (Liem et al., 1995). In the zebrafish, the bmp2b, bmp4, and bmp7 genes are expressed on the ventral side of the embryo during gastrulation and, in concert with the action of the dorsally-localized BMP antagonist Chordin, establish functional gradients that play key roles in patterning both the mesoderm and the ectoderm along the dorsoventral (DV) axis (Hammerschmidt et al., 1996; Nikaido et al., 1997). An intermediate level of BMP signaling induces neural crest fate (Schumacher et al., 2011), and altering the levels of BMP activity during gastrulation disrupts NCC formation (Neave et al., 1997; Nguyen et al., 1998). In addition, Wnt/β-catenin signaling plays a role in NCC induction, in part via regulation of expression of foxd3 and sox10 (Lewis et al., 2004). However, there is unlikely an absolute requirement for Wnt signaling in NCC induction or subsequent development, as some markers of NCCs and their chondrogenic and pigment cell derivatives are still expressed when a dominant inhibitor of Wnt signaling is induced at the onset of migration (Lewis et al., 2004).
Regulation of neural crest specification
In recent years, advances in molecular biology, and particularly next generation sequencing approaches, have fundamentally shaped our understanding of the gene regulatory interactions that underlie different events in neural crest development. This has led to the concept of a neural crest gene regulatory network (GRN), or rather a series of interlinked GRNs, that underlie the process of neural crest formation (Meulemans and Bronner-Fraser, 2004; Sauka-Spengler and Bronner-Fraser, 2008; Simões-Costa and Bronner, 2015). While the GRN has proven an invaluable tool, it is largely based on the integration of data from a variety of species. To appreciate the aspects of neural crest development that are either conserved or variable across species, it will be critical to generate species-specific GRNs. We have therefore begun to generate a zebrafish-specific neural crest GRN based upon published data (Figure 3). This is not an exhaustive summary, but is designed to initiate the process of organizing relevant interactions.
Fig. 3: A zebrafish-specific neural crest GRN.
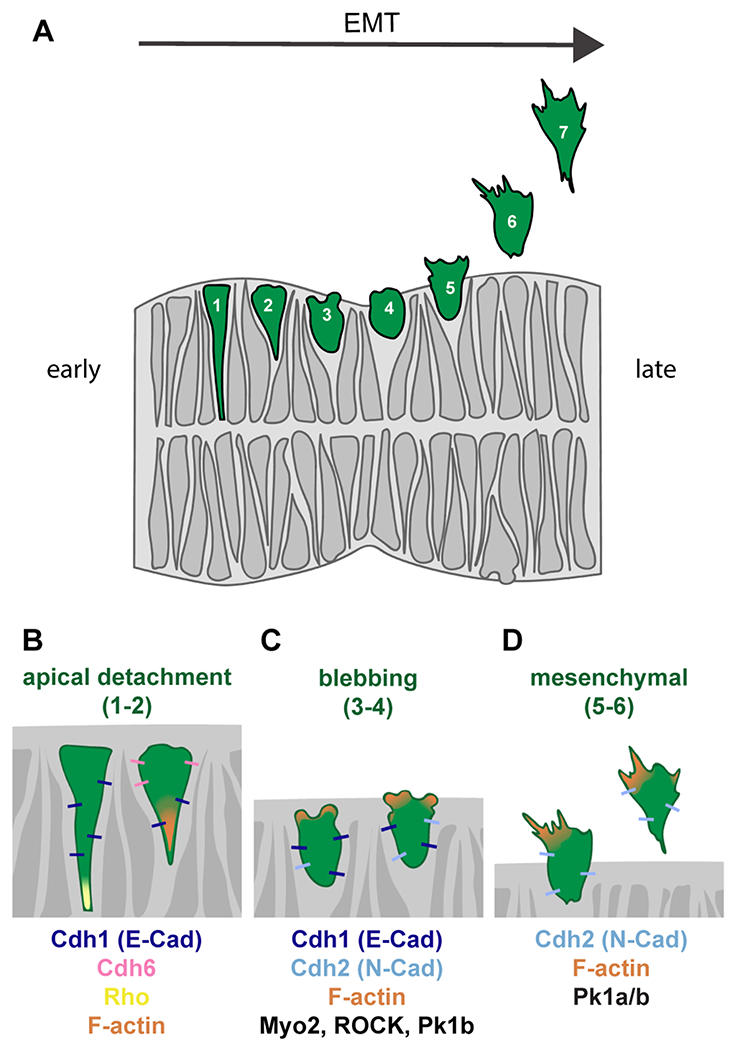
This simplified gene regulatory network is built exclusively from zebrafish data; see text for details. Direct interactions are depicted with solid lines, whereas dashed lines show interactions inferred from loss-of-function studies.
According to the hierarchy of the neural crest GRN (Sauka-Spengler and Bronner-Fraser, 2008; Simões-Costa and Bronner, 2015) inductive interactions between neural and non-neural ectoderm specify the NPB by driving the expression of a battery of transcription factor genes. In the zebrafish (Figure 3) these include pax3a (Garnett et al., 2012; Seo et al., 1998), tfap2a (Barrallo-Gimeno et al., 2004; Knight, 2003; Knight et al., 2004), msxBCE (Phillips et al., 2006), zic3 (Garnett et al., 2012), dlx5/6 (Narboux-Neme et al., 2019), and prdm1a (Artinger et al., 1999; Birkholz et al., 2009; Hernandez-Lagunas et al., 2005; Olesnicky et al., 2010; Powell et al., 2013; Roy and Ng, 2004). Three signals—BMP, Wnt, and FGF—are required for expression of pax3a at the NPB, whereas Wnt and intermediate levels of BMP are sufficient for zic3 expression (Garnett et al., 2012). Across vertebrate species, these signals are integrated through evolutionarily conserved enhancers that respond to particular combinations of signaling inputs (Garnett et al., 2012).
Cells at the NPB become specified to the neural crest fate as gastrulation draws to a close and segmentation begins, around 11 hpf. At this stage, several key transcription factor genes referred to as “neural crest specifiers” commence expression. These include foxd3 (Lister et al., 2006; Montero-Balaguer et al., 2006; Odenthal and Nüsslein-Volhard, 1998; Stewart et al., 2006), snai1b (Thisse et al., 1995), sox10 (Carney et al., 2006; Dutton et al., 2001; Kelsh and Eisen, 2000), and sox9a/b (Li et al., 2002; Yan et al., 2005, 2002). Additionally, twist (Das and Crump, 2012; Germanguz et al., 2007), mych (Hong et al., 2008), and id3 (Dickmeis et al., 2002), all of which have been shown to play key roles in NCC specification in other species (Rogers et al., 2012; Simões-Costa and Bronner, 2015), are expressed in zebrafish premigratory NCCs. In the zebrafish, these latter three genes play a role in restricting ectomesenchymal potential, survival, and proliferation, respectively. However, their role in NCC specification has not been formally demonstrated. As the hierarchical GRN model predicts, loss of upstream NPB gene function impedes NCC specification. For example, two different mutant alleles of prdm1a—narrowminded and ubo—display reduced NCC numbers and no RBs (Artinger et al., 1999; Roy and Ng, 2004). Prdm1a directly activates expression of the neural crest specifier genes foxd3 and tfap2 (Powell et al., 2013), and also regulates the expression of both the neural crest specifier sox10 and the RB marker islet1 (Olesnicky et al., 2010). In msxBCE morphants, expression of snai1b is lost and sox10 reduced, yet the expression of foxd3 remains unaffected (Phillips et al., 2006). Together these results indicate that the NPB genes activate expression of neural crest specifier genes through both specific and complex regulatory interactions.
Strikingly, phenotypic analysis of presumptive loss-of-function mutants in NPB and neural crest specifier genes has revealed that these transcription factors are required for the development of both distinct and overlapping NCC subpopulations. For example, colourless/sox10 mutants have defects in neural crest-derived melanoblasts, neurons, and glia, while skeletal derivatives remain unaffected (Dutton et al., 2001; Kelsh and Eisen, 2000). Further, the tfap2a/lockjaw mutant displays defects in NCC specification and migration, as well as in both chondrogenic and pigment derivatives (Knight, 2003; Knight et al., 2004). In a second mutant allele of tfap2a, termed mont blanc, defects in NCC induction, specification, and even migration were not noted, but these specimens again exhibit significant defects in chondrogenic differentiation (Barrallo-Gimeno et al., 2004). A more dramatic phenotype results from double morpholino knockdown of both tfap2a and tfap2c, which leads to the complete absence of NCCs (Hoffman et al., 2007; Li and Cornell, 2007). Similarly, both sym1/foxd3 mutants (Stewart et al., 2006) and foxd3 morphants (Lister et al., 2006) exhibit defects in several NCC derivatives, including peripheral neurons, glia, and cartilage, yet retain the proper number of melanocytes. On the other hand, mutants in the mother superior allele of foxd3, which disrupts a neural crest-specific regulatory element, exhibit a depletion of all NCC derivatives that is preceded by a reduction in expression of snail1b, sox9b, and sox10 (Montero-Balaguer et al., 2006). Finally, analysis of foxd3 and tfap2a double mutants has revealed that these embryos fail to express snai1b, sox9a/b and sox10 and lack all NCC derivatives (Arduini et al., 2009; Wang et al., 2011). In summary, mutations of individual zebrafish neural crest specifier genes are often insufficient to block NCC specification. Instead, experiments to date have revealed a robust combinatorial code that plays complex roles in both NCC specification and subsequent cell fate decisions.
The epithelial-to-mesenchymal transition
Following induction and specification of neural crest, the NCCs must pass through an epithelial-to-mesenchymal transition (EMT), during which their morphology, adhesive properties, polarity, and behavior change dramatically as they transition into actively migrating mesenchymal cells (Theveneau and Mayor, 2012a). EMT occurs in multiple contexts, and its roles in collective cell migrations, morphogenesis, and cancer have been broadly investigated (Friedl and Gilmour, 2009; Friedl and Wolf, 2003; Gallik et al., 2017; Lee et al., 2006; Thompson and Williams, 2008). However, the manner in which EMT has typically been discussed—as a ‘binary switch’ from tightly-packed, highly adhesive epithelial cells to dispersed, loosely-packed mesenchymal cells—has recently been called into question, both in the context of zebrafish neural crest (Ahsan et al., 2019; Berndt et al., 2008; Clay and Halloran, 2014, 2013), and beyond (Campbell and Casanova, 2016; Nieto et al., 2016). A growing body of evidence indicates that EMT is not in fact a binary switch, but is instead a more gradual process, during which cells pass through a spectrum of morphologies that are neither entirely epithelial nor entirely mesenchymal.
The process of EMT is complex, involving changes in cell adhesion, cell-cell, and cell-matrix interactions, as well as the extension of protrusions such as filopodia and pseudopodia (Berndt et al., 2008; Erickson and Perris, 1993; Halloran and Berndt, 2003; Savagner, 2010, 2001; Thiery and Sleeman, 2006). This morphological transition requires the regulation of many effector genes (Simões-Costa and Bronner, 2015). Critically, the GRN that underlies neural crest EMT in sarcopterygian models includes several transcription factors also involved in NCC fate specification, such as FoxD3, SoxE and Pax3/7, as well as Snail1/2 and Twist (Simões-Costa and Bronner, 2015). In zebrafish, the transcriptional regulation of EMT has not yet been investigated in detail. Zebrafish snai1b and twist1a/b are expressed in NCCs at the time of EMT, however their role in this process has not been formally evaluated (Das and Crump, 2012; Germanguz et al., 2007; Jimenez et al., 2016; Thisse et al., 1995).
A recent report of single cell RNAseq of murine NCCs uncovered a sequence of transcriptional events that take place during delamination and early migration. In particular, this study revealed that the murine premigratory neural crest can be separated into two distinct subpopulations: a pre-EMT population composed of cells that have not yet started delaminating from the neural tube, and a delaminating subpopulation that is marked by expression of Snai1 (Soldatov et al., 2019). Importantly, the upregulation of neural crest specifier genes and downregulation of NPB genes exhibits variable timing, with some NPB genes—e.g. Msx1, Msx2, Lmx1a—continuing to be expressed even as the specifier genes upregulate. Furthermore, this analysis also uncovered a subset of migratory progenitors that do not yet express known markers of downstream fates (Soldatov et al., 2019). These recent results from a mammalian system provide further evidence that EMT is a gradual process, not just in regards to cell morphologies but also transcriptional states.
Although the molecular underpinnings of zebrafish EMT remain incompletely understood, our understanding of the cellular basis of EMT has benefitted from live imaging approaches available in the zebrafish model. In particular, a series of studies from the Halloran lab has provided detailed insights into EMT of the zebrafish cranial NCCs. An initial study used single cell labeling to reveal that the earliest cranial NCCs originate from lateral to, or overlaying, the developing neuroepithelium at the 13-15 hpf stage, with later NCCs arising at 15-19 hpf from the pseudostratified neuroepithelium (Berndt et al., 2008). This recognition of two separate populations of cranial NCCs is consistent with the initial morphological description of the region (Schilling and Kimmel, 1994), and with follow up studies using transgenic markers (Jimenez et al., 2016). Subsequent detailed analyses showed that the later migrating NCC population initiates EMT at the point of apical detachment from the midline of the neuroepithelium (Figure 4) (Berndt et al., 2008; Clay and Halloran, 2014), when activation of the small GTPase Rho becomes restricted to the apical region by the RhoGAP Arhgap1, leading to F-actin enrichment (Clay and Halloran, 2013). Subsequently, the NCCs round up and form membrane-based blebs—rounded bulges on one side of the cell—and then begin to extend filopodial and lamellipodial protrusions as they exit the neuroepithelium and migrate away (Ahsan et al., 2019; Berndt et al., 2008; Clay and Halloran, 2014). Interestingly, blebs and filopodia exhibit different underlying actin localization and dynamics, consistent with an ongoing series of morphological transitions (Berndt et al., 2008). Bleb formation is crucial for EMT to progress, as inhibition of myosin-II or Rho-kinase (ROCK), which disrupt blebbing, inhibits EMT (Berndt et al., 2008). Our own recent work has also implicated Prickle1a and Prickle1b—core Planar Cell Polarity (PCP) molecules—in zebrafish cranial NCC EMT. While in wild type embryos the blebbing phase of EMT typically completes within 20 minutes, in Prickle1-deficient embryos the majority of NCCs are delayed in the transition from blebbing to mesenchymal morphologies (Ahsan et al., 2019), indicating a block in the EMT process. Future work will be required to address whether the entire core PCP suite of molecules is necessary for the EMT that precedes overt NCC migration, both in zebrafish and other models.
Fig. 4: Zebrafish neural crest cells undergoing the epithelial-to-mesenchymal transition.
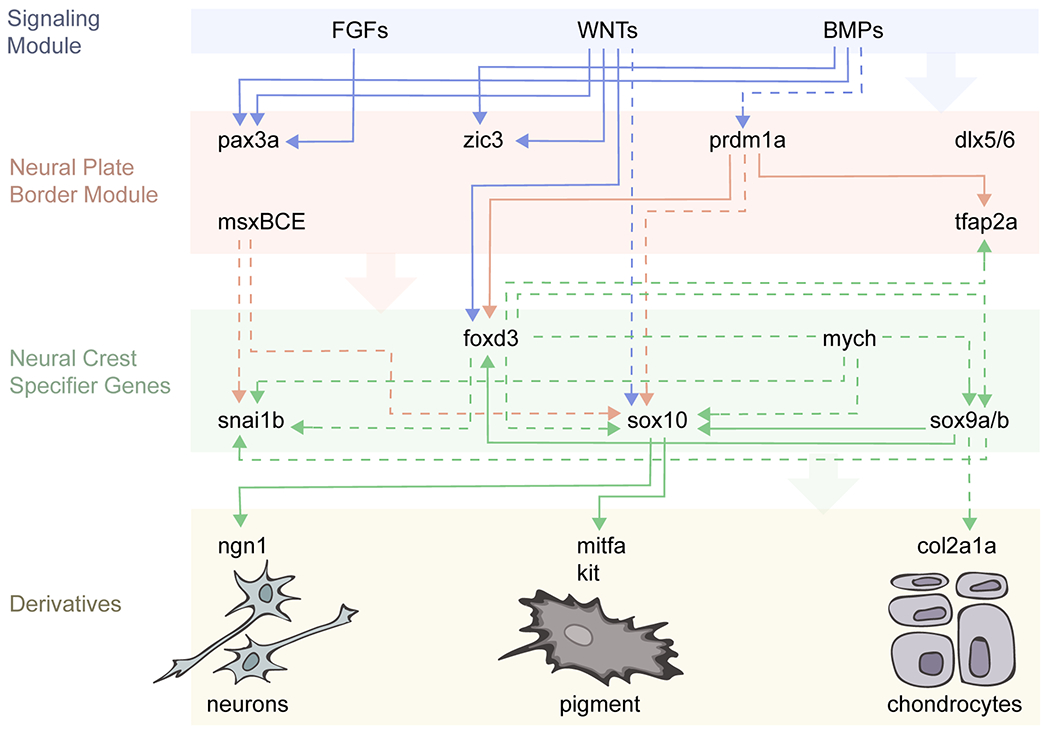
(A) A single premigratory neural crest cell (1) is shown over successive time points (from left to right) undergoing apical detachment from the neuroepithelium (2), blebbing behavior (3-4), and ultimately becoming a fully mesenchymal and migratory neural crest cell (5-7). (Based on Berndt et al., 2008, Clay and Halloran, 2010, and Ahsan, 2019). (B-D) Molecular regulators of apical detachment (B), blebbing (C), and mesenchymal (D) states; see text for details.
EMT also involves cell surface changes and the dissolution of cadherin-mediated adherens junctions. Initially, evidence from several model species indicated that NCCs undergo a “cadherin switch” from E-cadherin to N-cadherin during EMT (Taneyhill and Padmanabhan, 2014; Taneyhill and Schiffmacher, 2017). Consistent with these findings, in zebrafish the premigratory NCCs predominantly express E-cadherin (Cdh1), whereas migratory NCCs predominantly express N-cadherin (Cdh2) (Ahsan et al., 2019; Scarpa et al., 2015). E-cadherin represses contact inhibition of locomotion (CIL) by controlling protrusions via Rac1 and p120 (Scarpa et al., 2015). N-cadherin is expressed in migratory NCCs, where it plays a critical role in promoting migration (Piloto and Schilling, 2010; Powell et al., 2015). The proper cellular localization of N-cadherin is regulated by several inputs associated with Shh and Wnt signaling, and disruption of these inputs causes NCC migration to stall. For example, mutant analysis has revealed that the putative Shh receptor Cdon localizes N-cadherin to allow directed migration of trunk NCCs (Powell et al., 2015). Similarly, deficiencies in Rabconnectin-3a and its associated v-ATPase subunit lead to misregulation of cadherins by disrupting vesicle endocytosis and Wnt signaling, resulting in a failure of NCCs to migrate—with phenotypic analysis in this study focusing on the cranial region (Tuttle et al., 2014). Further, disruption of the Wnt target gene Ovo1 similarly disrupts intracellular trafficking, thus blocking localization of N-cadherin at the membrane and again disrupting cranial NCC migration (Piloto and Schilling, 2010). Interestingly, Prickle1b-deficient embryos have elevated levels of E-Cad in both premigratory and migratory cranial NCCs, and decreased levels of N-Cad in the migratory cells, indicating a potential link between PCP and cadherin regulation (Ahsan et al., 2019).
Recent studies in other models have revealed a more complex and dynamic regulation of cadherins (Taneyhill and Schiffmacher, 2017). Briefly, a second switch from type I cadherins (E- and N-cadherin), which mediate strong cell-cell interactions, to more mesenchymal type II cadherins (including Cadherin-7 and Cadherin-11) also takes place (Martik and Bronner, 2017; Mayor and Theveneau, 2013; Simões-Costa and Bronner, 2015; Taneyhill and Schiffmacher, 2017). In zebrafish, Cadherin-6 (Cdh6) is upregulated prior to EMT and promotes apical detachment by regulating the spatiotemporal dynamics of F-actin and active Rho GTPase (Clay and Halloran, 2014). On the other hand, the roles of Cadherin-7 and Cadherin-11 have not been investigated in zebrafish; however, given their roles in NCC EMT in other models it is possible that the zebrafish homologs of these molecules will similarly prove to play important roles in the zebrafish neural crest.
An interesting facet of neural crest development in zebrafish, and presumably of other teleosts too, is the derivation of early NCCs from laterally located cells in the developing neural keel. This may in turn confer different properties on these cells, and lineage studies have indeed shown that the lateral versus medial zebrafish cranial NCCs have differing fates (Schilling and Kimmel, 1994). An intriguing commentary piece from Weston and Thierry (2015) lays out a related argument that the cranial neural crest is made up of two very different populations, with a more medial “authentic” neural crest deriving from the neural folds and an early lateral “metablast” arising from the adjacent non-neural epithelium (Weston and Thiery, 2015). While Weston and Thierry base their arguments on results from multiple species, their hypothesis is driven primarily by chick data, with a central aspect of their model being that the ‘metablast’ cells alone have skeletogenic potential. Importantly, Schilling and Kimmel’s zebrafish fate mapping data (1994) are at odds with this model, as it is the most medial cranial NCCs that give rise to the skeletogenic fates, with the lateral cells giving rise not to skeletal elements but instead to neurons. Ultimately, to determine whether there are significant species-specific differences in the origins of the skeletogenic neural crest, we will require a more nuanced understanding of both the precise sources of the differing populations of cranial neural crest cells, and of the cellular mechanisms that allow the cells to become migratory, across a variety of phylogenetically informative species.
In closing this section, we note that the term EMT is often used interchangeably with ‘delamination’ in the neural crest literature, which is appropriate when delamination is used to refer to individual neural crest cells leaving the neuroepithelium. However, delamination may equally be used to describe the separation of two tissue layers, such as the separation of the neural epithelium from the overlying epidermal epithelium. These alternative uses of the word can lead to confusion, especially when a neural crest ‘layer’ is implied to delaminate from the neuroepithelial layer, as it is unclear which of the two definitions of delaminate—or perhaps some combination of both— is being referred to. In general, we recommend care be taken in appropriate use of terminology.
NEURAL CREST MIGRATION
After completing EMT, NCC cells migrate collectively along stereotypic pathways, often traveling great distances before reaching their destination and differentiating into a variety of derivatives (Bronner and Simões-Costa, 2016; Mayor and Theveneau, 2013). Cranial and trunk NCCs exhibit important differences in their routes and modes of migration, at least in part as a result of differences in their local environments. However, in both contexts migration is generally regulated by a combination of attractive and repulsive cues, as well as by cell-cell interactions (Theveneau and Mayor, 2012b). In this section, we describe the regulation of cranial and trunk neural crest migration, and highlight proposed models that explain how collective migration is regulated.
Cranial neural crest migration
Zebrafish cranial NCCs begin their migration around 13 hpf. A subset of NCCs from the midbrain migrate anteriorly between and around the eyes to contribute to the neurocranium (Knight and Schilling, 2006) (see cranial morphogenesis section), whereas posterior midbrain and hindbrain-derived NCCs segregate into distinct streams that migrate ventrally into the pharyngeal arches (PAs) and produce the viscerocranium (Figure 1) (Knight and Schilling, 2006; Schilling and Kimmel, 1994). NCCs from the midbrain and rhombomeres (r) 1-3 contribute to the mandibular arch (PA1), while those from r3-5 populate the hyoid arch (PA2), and cells from r5-r7 fill the branchial arches (PA4-7) (Schilling and Kimmel, 1994). Studies from multiple model organisms have revealed that cranial NCC migration is regulated by a variety of repulsive cues and chemoattractants (Figure 5A). Of note, semaphorins and their neuropilin/plexin receptors have been shown to play an important role in zebrafish cranial NCC stream formation, while Sdf1 (Cxcl12b) is an important chemoattractant (Gallik et al., 2017; Halloran and Berndt, 2003; Theveneau and Mayor, 2012b, 2012a). However, many of the signals that have been demonstrated to control directional migration in NCCs of other species (Theveneau and Mayor, 2012b), including Eph/Ephrin signaling and VEGF, have not been evaluated in zebrafish.
Fig. 5: Models of neural crest migration.
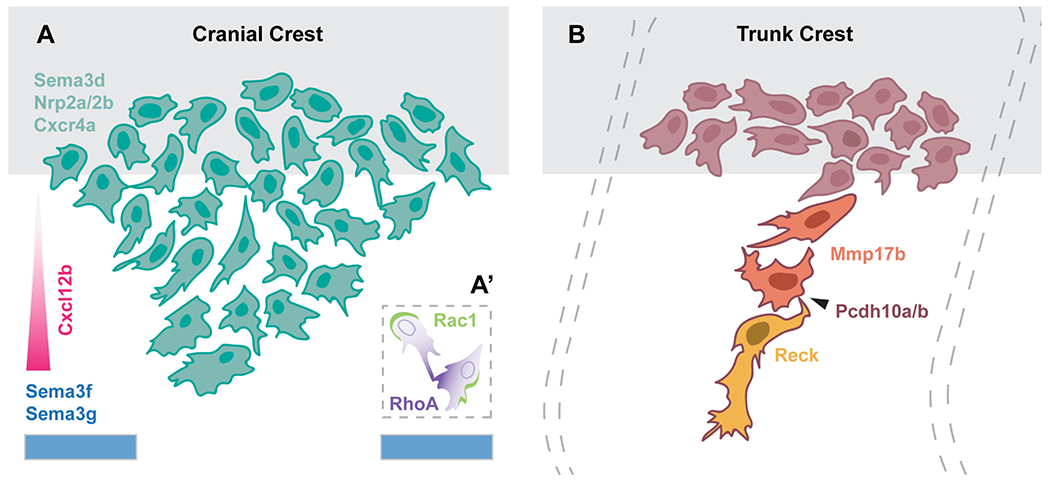
(A) Schematic of molecular regulators of cranial neural crest migration. A stream of cranial NCCs migrates away from the neural tube (grey) avoiding repulsive Sema3f and Sema3g cues in the environment, shown in blue, and towards the chemokine Cxcl12b, shown in pink. These cells express Sema3d, as well as the receptors Nrp2a/2b and Cxcr4a. The inset (A’) shows the molecular basis of CIL, during which Rac1 and RhoA activity become recruited to the trailing and leading end, respectively. (B) Schematic of trunk neural crest cell migration, away from the neural tube (grey), depicting the leader (yellow), follower (orange), and premigratory (red) populations. Leader cells express Reck, whereas follower cells express Mmp17b. All trunk neural crest cells express Pcdh10a/b, which mediates cell-cell contacts (arrowhead). Segmental migration of trunk neural crest cells is restricted by adaxial cells; see text for details.
Zebrafish sema3d is expressed as early as 11 hpf in premigratory NCCs and is eventually expressed in all three streams of NCCs that migrate towards the pharynx (Berndt and Halloran, 2006; Halloran et al., 1999). Knockdown of Sema3d reduces migratory NCC marker expression and disrupts NCC-derived tissues (Berndt and Halloran, 2006). Further, sema3f and sema3g are expressed in the neural crest-free regions of the head and coordinate the migration of cranial NCCs, which express the corresponding receptors nrp2a and nrp2b (Yu and Moens, 2005). In addition to environmental repulsive cues, small, secreted chemoattractants are important for directing neural crest migration (Theveneau and Mayor, 2012b). In zebrafish, cxcr4a is expressed in migrating cranial, but not trunk, NCCs, while sdf1b is expressed in the pharyngeal endoderm, the target tissue of directionally migrating cranial NCCs (Olesnicky Killian et al., 2009). Knockdown and overexpression approaches have indicated that Sdf1b/Cxcr4a signaling promote proper migration of NCCs into the pharyngeal arches (Olesnicky Killian et al., 2009), possibly mediated via Fascin1-dependent filpodia (Boer et al., 2015). Our schematic (Figure 5A) hypothesizes that Cxcl12b functions in a gradient; future experiments—perhaps using beads to provide localized protein sources—will be needed to test this hypothesis. Finally proper dispersal and migration of the anterior population of NCCs that contribute to the palate relies on an interplay of the attractant signal Pdgf, and its negative regulator, microRNA Mirn140 (Eberhart et al., 2008).
Trunk neural crest migration
Along the AP extent of the trunk, NCCs migrate on two distinct pathways: an initial, medial pathway, between the neural tube and the somites (Figures 2, 5B), and a later, lateral pathway, between the somites and the overlying ectoderm (Raible et al., 1992). NCCs located at the most lateral aspect of the dorsal neural tube are the first to begin migration and enter the medial pathway starting after 15 hpf. The onset of their migration shows a distinct anterior to posterior sequence, although occasionally more posterior NCCs do migrate out of sequence, ahead of more anterior ones (Raible et al., 1992). NCCs that migrate along the medial pathway give rise to all types of derivative, including neurons of the sensory and sympathetic ganglia, Schwann cells, and pigment cells (Raible and Eisen, 1994). Approximately 4 hours later, NCCs initially located in the dorsal-most region of the neural tube begin to enter the lateral pathway (Raible et al., 1992). Notably, these cells only differentiate into pigment cells (Raible and Eisen, 1994). However, factors other than time of migration likely regulate pathway decision, as some cells continue to enter the medial pathway even after migration along the lateral pathway has begun (Raible et al., 1992). Here, we will focus on the mechanisms that regulate migration along the medial pathway.
As in other species, zebrafish NCC migration on the medial pathway is spatially restricted along the AP axis, such that a series of reiterated streams, one per somite, is established along each side of the trunk (Figure 1) (Raible et al., 1992). However, unlike in chick and mouse embryos, where NCCs migrate through the sclerotome of the anterior half of each somite (Bronner and Simões-Costa, 2016; Newgreen et al., 1990), zebrafish trunk NCCs show no obvious AP restrictions until they reach the ventral edge of the neural tube, at which point they become restricted to migrating on the medial surface of the center of each somite (Figure 5B) (Raible et al., 1992). In zebrafish, as in other teleosts, the majority of somite-derived cells are myotomal, with only a small ventrally localized sclerotome compartment forming (Honjo and Eisen, 2005; Morin-Kensicki and Eisen, 1997). Ablation of the zebrafish sclerotome does not disrupt the segmental pattern or subsequent development of dorsal root ganglia (DRG) (Morin-Kensicki and Eisen, 1997). Instead, the slow muscle precursors, termed adaxial cells, regulate the pattern of neural crest migration on the medial pathway (Honjo and Eisen, 2005). This is perhaps not surprising as myotome cells are the first encountered by migrating NCCs and, in fact, the onset of neural crest migration coincides with contact between the two cell types (Raible et al., 1992).
Although the molecular basis of trunk NCC stream restriction and migration has not been fully established, interactions between migrating NCCs and myotome cells, the extracellular matrix, and other NCCs are likely critical. The muscle-specific receptor kinase (MuSK) and its ligand Wnt11r play a key role in restricting NCC migration to the center of each somite (Banerjee et al., 2011). In the absence of MuSK, NCCs do not retract non-productive leading edges, leading to impaired stream formation (Banerjee et al., 2011). Similarly, the glycosyltransferase lh3 (now plod3) is expressed in a subset of ventral somitic cells and, with its substrate Collagen18A1, is necessary for segmental stream formation (Banerjee et al., 2013). Additionally, the GPI-linked matrix metalloproteinase mmp17b and the metalloproteinase inhibitor reck are both required in a cell-autonomous fashion for NCC migration (Leigh et al., 2013; Prendergast et al., 2012) (Figure 5B). Interestingly, Mmp17b and Reck are expressed in NCCs found in close apposition, and interact biochemically, raising the possibility that they function in concert (Leigh et al., 2013). Finally, loss of the proto-cadherin encoding gene pcdh10a, either singly or together with its paralog pcdh10b, disrupts migration along the medial pathway, in part due to a loss of cell-cell contact with other migrating NCCs (Williams et al., 2018).
Models of neural crest collective migration
Several studies support a model whereby contact inhibition of locomotion (Abercrombie and Heaysman, 1953) or “CIL”–based dispersion and mutual attraction promote the collective directional migration of NCCs (Theveneau and Mayor, 2012a, 2012b). Research from the Mayor group showed that in Xenopus neural crest explants cells at the edge were polarized, projected large lamellopodia, and had higher persistence of migration than interior cells (Carmona-Fontaine et al., 2008). When two explanted migrating NCCs came into contact, they collapsed their protrusions, and migrated away from each other, suggesting that CIL plays a critical role in directional migration of NCCs (Carmona-Fontaine et al., 2008). In follow up studies, the molecular basis of CIL has also been investigated in some detail in Xenopus. These studies showed that the Wnt/planar cell polarity (PCP) pathway mediates CIL during neural crest migration, in part by promoting RhoA activation at the site of contact and Rac1 activity at the free edge in an N-cadherin-dependent manner (Carmona-Fontaine et al., 2008; De Calisto et al., 2005; Matthews et al., 2008; Theveneau et al., 2010). Further, the chemoattractant Sdf1 amplifies and stabilizes cell polarity, which results in directional migration (Theveneau et al., 2010). Critically, Xenopus NCCs also exhibit mutual attraction mediated by the complement factor C3a and its receptor C3aR (Carmona-Fontaine et al., 2011). In supplement to comprehensive Xenopus explant experiments, live imaging of zebrafish embryos transgenic for the NCC marker sox10:gfp demonstrated that migrating cranial NCCs also exhibit CIL in vivo (Figure 5A’) (Carmona-Fontaine et al., 2008). While syndecan4 (Matthews et al., 2008) and par3 (Moore et al., 2013) have been shown to play key roles in zebrafish neural crest migration and CIL, the degree to which the molecular mechanisms elucidated in Xenopus are conserved in zebrafish remains to be explored further.
Interestingly, mathematical models from the Kulesa lab show that simple chemotaxis is insufficient to explain the experimental observations of long-distance migration by NCCs (McLennan et al., 2012). On the other hand, when the model is refined to include two NCC subpopulations—cells at the migratory front that respond to a VEGF gradient and late–emerging trailing cells that respond to directional cues from leading NCCs—it predicts that the majority of cells invade in a multicellular stream (McLennan et al., 2012). Moreover, transcriptional profiling of migrating chick cranial NCCs revealed that leading and trailing cells possess distinct gene expression profiles (McLennan et al., 2012): the leading cells show upregulation of genes involved in cell guidance and navigation, whereas the trailing cells show upregulation of distinct cadherins (McLennan et al., 2012). Additional profiling of single chick cranial NCCs uncovered a unique gene expression signature in a small fraction of lead cells termed “trailblazers” (McLennan et al., 2015a). This signature includes 16 genes that are up- or down-regulated, with the TGF-beta signaling antagonist BAMBI, and the CXCR1 and NOTCH1 receptors, consistently expressed at high levels by trailblazers across two different phases of NCC migration. Notably, exposure of NCCs to the chemoattractant VEGF in vitro upregulates a subset of genes associated with the lead cell signature, suggesting that it induces trailblazer cell identity (McLennan et al., 2015b). Future experiments will be needed to address whether trailblazer cells direct the collective migration of zebrafish cranial NCCs, and whether the molecular signature of lead cell identity is conserved.
A recent report from the Linker lab reveals that cranial and trunk NCCs utilize distinct migratory strategies (Figure 5). Cell tracking experiments in both chick and zebrafish show that in the cranial region NCCs dynamically rearrange within the multicellular streams without regard to their initial time of emergence. Further, when cells at the front of a cranial NCC stream are ablated in zebrafish, migration remains unperturbed (Richardson et al., 2016). Future experiments will be needed to determine whether the cells at the front of the chick cranial neural crest streams are required for successful migration. However, even if chick leading cranial NCCs prove not to be required for migration, the molecularly-defined trailblazer cells—potentially a population that changes over time—may still prove to influence migration in important ways.
In the trunk, zebrafish NCCs consist of three distinct subpopulations: motile premigratory cells that remain in the dorsal region, leader cells that initiate migration along the medial pathway, and follower cells that trail the leader cell and connect it to the premigratory population (Figure 5B). Leader cells retain their position at the front of each stream throughout migration, whereas follower cells actively rearrange within a stream. Using laser ablation, Richardson and colleagues demonstrated that the leader cells are required for, and direct, trunk NCC migration. Further, leader and follower identities are non-interchangeable and seemingly acquired before migration. After leader cell ablation, migration pauses until a new leader emerges from within the premigratory cell population, and migrates down the stalled stream to resume leading it onward (Richardson et al., 2016). To fully understand this process, the mechanisms that confer leader versus follower trunk NCC identity will need to be uncovered.
NEURAL CREST DIFFERENTIATION
The zebrafish has proven helpful to unpacking how intrinsic gene regulatory states interact with extrinsic environmental signals to mediate the differentiation of NCC derivatives. Zebrafish NCCs are fate-restricted from an early stage, as individually labeled premigratory NCCs typically produce differentiated cells of only a single class (Kelsh and Raible, 2002; Raible et al., 1992; Schilling and Kimmel, 1994). Dorsky et al. (1998) confirmed the initial report from Schilling and Kimmel (1994) of clonal restrictions in cranial NCCs, and further showed that cell fates are biased by Wnt signaling, with medial Wnt signals promoting pigment cell fates. Further, clonal analysis revealed that those NCCs that do give rise to multiple kinds of derivatives likely do so by first forming type-restricted precursors (Raible and Eisen, 1994). These results suggest that NCCs are specified to their final fates well before they reach their ultimate locations.
Recently, single cell RNAseq of murine NCCs revealed that migrating NCCs undergo a series of sequential binary fate restrictions. Further, in situ sequencing showed that these distinct states show spatial segregation. Specifically, the first bifurcation separates the sensory lineage from the common progenitors of autonomic and mesenchymal fates, and the second split separates the autonomic neuronal fate from mesenchymal progenitors (Soldatov et al., 2019). This analysis is especially valuable because it elucidates the transcriptional regulation of cell fate decisions. Namely, each decision consists of an initial coactivation phase, where genes of competing cell fate programs are coexpressed, followed by a gradual biasing towards a particular fate, and finally a commitment phase where mutually exclusive, fate-specific gene expression programs becomes activated.
In addition to intrinsic regulatory progams, proper migration and environmental signals are also necessary for NCC differentiation. For example, erbB3 mutants lack DRG not because of an inability to specify neurons, but rather because NCCs fail to pause at the location where DRGs should form and thus do not receive the necessary signals to differentiate (Honjo et al., 2008). A great deal of research has been published on the differentiation of the various neural crest cell types—below, we present a brief discussion of zebrafish NCC differentiation into three specific cell types: pigment cells, chondrocytes, and neurons.
Pigment Cells
Zebrafish are an invaluable model for the study of pigment development and pattern formation due to the wealth of pigment mutants identified during the large-scale mutagenesis screens of the 1990s (Kelsh et al., 1996; Odenthal et al., 1996). However, as several recent reviews have already covered this topic in significant depth (Cooper, 2017; Irion et al., 2016; Rawls et al., 2001), we touch on only a few highlights in the section below.
Zebrafish NCCs give rise to three major pigment cell types—yellow xanthophores, iridescent iridophores, and black melanocytes (Rawls et al., 2001). Melanocyte precursors begin expressing melanin pigment around 24 hpf and the embryonic melanocyte pattern is largely established by 48 hpf (Rawls et al., 2001). During larval-to-adult metamorphosis, this pattern is gradually replaced by the adult pigment pattern (Johnson et al., 1995). Importantly, the adult pigment cells are largely derived from post-embryonic melanocyte stem cells (MSCs) that originate from the neural crest (Budi et al., 2011; Hultman and Johnson, 2010; Tryon et al., 2011). These MSCs are established within the first 2 days and remain closely associated with peripheral nerves and ganglia, including the DRG, which likely provide a niche (Budi et al., 2011; Dooley et al., 2013). Further, the developmental mechanisms of embryonic and metamorphic pigment cells may be decoupled, as in puma (tuba8l3) mutants that lack adult melanophores while embryonic melanophores remain unaffected (Parichy et al., 2003; Parichy and Turner, 2003).
As in other species, the basic helix–loop–helix/leucine zipper transcription factor Mitfa plays a central role in melanocyte differentiation (Lister et al., 1999). Melanophores are absent throughout embryonic and larval development in nacre/mitfa mutants, a phenotype which persists through adulthood. On the other hand, the mutants show an increase in the number of iridophores. The mitfa gene functions cell-autonomously in the melanophores, where it is required for the expression of trp2 and kit (Lister et al., 1999). Analysis of the mitfa promoter has revealed Tcf/Lef binding sites (Dorsky et al., 2000), suggesting that Wnt signaling, which promotes pigment cell fate at the expense of neurons and glia (Dorsky et al., 1998), may do so by directly regulating mitfa expression (Dorsky et al., 2000, 1998). Moreover, the mitfa promoter contains Sox10 binding sites, which are necessary for its expression both in vitro and in vivo and expression of mitfa is sufficient to rescue melanophore development in sox10 mutant embryos (Elworthy et al., 2003).
The establishment of the adult pigment pattern requires cell-cell interactions between different pigment cell types (Eom et al., 2015; Frohnhofer et al., 2013; Hamada et al., 2014; Maderspacher and Nüsslein-Volhard, 2003; Nakamasu et al., 2009; Patterson et al., 2014; Patterson and Parichy, 2013; Volkening and Sandstede, 2018). The genetic and molecular mechanisms by which the self-organizing stripe pattern is generated were recently reviewed in detail by Irion and colleagues (2016). An exciting recent finding from the Parichy lab is that macrophages are necessary to relay long-distance signals between xanthophores and melanophores via specialized cellular projections termed airinemes (Eom and Parichy, 2017). Whether airineme/macrophage-mediated signaling functions in other contexts remains to be explored.
Chondrocytes and cranial morphogenesis
The elaborated cranium, a key feature of all vertebrates (Gans and Northcutt, 1983), is largely derived from the cranial neural crest. In humans, defects in crest-derived, or crest-influenced, craniofacial structures are relatively common (Mayor and Theveneau, 2013; Trainor, 2016). Unsurprisingly, there is a great deal of interest in understanding the patterning and differentiation of the neural crest-derived cartilaginous and bony elements of the skull. Two recent reviews have highlighted the benefits of the zebrafish in this context (Mork and Crump, 2015; Van Otterloo et al., 2016).
Foundational fate mapping of the zebrafish cranial neural crest was established by Schilling and Kimmel (1994). By dextran-labeling of individual cells, they not only revealed the AP regionalization of the cranial NCCs, but additionally showed that particular cell fates are influenced by the initial mediolateral (ML) position of NCCs. The most laterally-derived cells tend to differentiate into neurons of the cranial ganglia, whereas more medially located cells migrate longer distances into the pharyngeal arches to differentiate into cartilage. Other fates—glia and melanocyte derivatives—are less spatially restricted.
More recently, transgenic reporter lines have allowed detailed fate mapping of the cranial neural crest contribution to the anterodorsally-located neurocranium (Dougherty et al., 2013; Wada et al., 2005). These studies revealed that NCCs derived from the anterior midbrain migrate over the eye to contribute to the medially-located ethmoid plate of the neurocranium, whereas NCCs from the posterior midbrain migrate behind the eye to contribute to the bilateral trabeculae elements. The more posterior components of the neurocranium do not derive from the neural crest, but rather from the mesoderm. The precise interface between the NCC–derived and mesoderm–derived elements was revealed via tracing experiments that used a tamoxifen-inducible Cre-based lineage marker driven by the sox10 regulatory sequences (Mongera et al., 2013). The indelible nature of the marker allowed lineage tracing though to adult stages, revealing late NCC contributions to additional cranial structures, including the gill pillar cells and the barbels, which are tentacle-like chemosensory structures. A variety of genetic manipulations have revealed significant conservation in the fundamental molecular mechanisms that pattern the zebrafish anterior neurocranium and the human hard palate (Swartz et al., 2011), suggesting that zebrafish can provide a useful model of palate development (Mork and Crump, 2015).
Differentiation of NCCs into chondrocytes depends on gene regulatory interactions (Simões-Costa and Bronner, 2015). Studies in mice suggest that the NCC specifier gene Sox9 is a key regulator of chondrogenic fate, functioning in part by regulating the expression of extracellular matrix (ECM) components and genes encoding ECM modification enzymes (Bi et al., 1999; Oh et al., 2014). In zebrafish, mutations in jellyfish/sox9a (Yan et al., 2005, 2002) or sox9b (Yan et al., 2005) result in craniofacial defects and reduced expression of the collagen–encoding gene col2a1a (Yan et al., 2005). Notably, the sox9 paralogs play distinct roles during cartilage morphogenesis: sox9a is necessary for chondrocyte stacking, while sox9b mutants fail to attain proper number of chondrocytes (Yan et al., 2005). Double mutants in both sox9a and sox9b completely lack both pharyngeal cartilage and the neurocranium, suggesting that the sox9 paralogs have both overlapping and distinct functions during pharyngeal cartilage development (Yan et al., 2005).
The cranial NCCs carry some intrinsic information with them into the pharyngeal region (Noden, 1988, 1983), however signals from the surrounding tissues, including Endothelin 1 (Clouthier and Schilling, 2004; Kimmel et al., 2003; Miller et al., 2003, 2000; Nair et al., 2007), Shh (Eberhart, 2006; Wada et al., 2005), and Bmp (Alexander et al., 2011; Zuniga et al., 2011) are necessary for proper formation and patterning of the skeletal elements in the pharyngeal arches (Knight and Schilling, 2006; Medeiros and Crump, 2012; Minoux and Rijli, 2010). Fgf signals are also critical for chondrocyte development, with early expression of Fgfs in the neural tube and lateral plate mesoderm, as well as later expression in the pharyngeal endoderm, influencing distinct aspects of pharyngeal cartilage development. If early Fgf signals are lost, the endodermal pouches of the pharyngeal arches fail to form, and subsequently, the pharyngeal cartilages are reduced or absent (Crump et al., 2004a; Reifers et al., 2000, 1998). Later, Fgf signals from the pharyngeal endoderm are required for induction and survival of chondrogenic precursors (David et al., 2002; Walshe and Mason, 2003). Another example of the importance of pharyngeal endoderm in patterning adjacent NCC derivatives, is the requirement for endodermal Integrinα5 function in development of hyoid (second) arch derivatives (Crump et al., 2004b). Integrinα5 is necessary for development of the first endodermal “pouch”, which in turn controls cartilage development from the adjacent NCCs, apparently by regulating not gene expression, but rather the compaction and survival of NCC-derived cells.
Overall, the zebrafish has shown particular utility in allowing researchers to visualize chondrogenic precursors in vivo using transgenic tools, which when coupled with genetic manipulation can provide new insights into how signals are integrated to drive cartilage development and morphogenesis. A powerful example of this approach uncovered the integration of Notch, Endothelin and Bmp signaling in the control of skeletal patterning in the upper face (Barske et al., 2016). Further, a recent study has coupled quantitative measurements of gene expression, and live imaging, with a computational approach, to model the complexities of the signaling networks that pattern the mandibular (first) arch structures that give rise to the jaw (Meinecke et al., 2018). Such detailed models not only provide new insights into zebrafish pattern formation, but have the potential to be extrapolated to other species and ultimately shed light on how widely differing jaw structures have evolved.
Neurons/Glia
DRG and Sympathetic Neurons.
NCCs at all axial levels are capable of giving rise to neurogenic derivatives. In the cranial region, NCCs contribute to the cranial ganglia as well as to Schwann and satellite cells (Kague et al., 2012; Schilling and Kimmel, 1994). In the trunk, NCCs give rise to sensory neurons of the dorsal root ganglia (DRG), autonomic sympathetic neurons, and Schwann cells (Raible et al., 1992; Raible and Eisen, 1994). In zebrafish embryos, DRG neurons begin their differentiation at 36 hpf, about 20 hours after the onset of trunk neural crest migration (An et al., 2002). In contrast, sympathetic neurons are not detected until at least 2 days after the differentiation of DRG neurons (An et al., 2002). Individual DRGs and sympathetic ganglia initially contain only a few differentiated neurons (An et al., 2002); however, the number of neurons progressively increases due to continued neuronal cell division until at least 4 weeks of age (An et al., 2002). In this regard, the zebrafish is very different to chick and mouse embryos, in which neuronal differentiation is concomitant with terminal mitosis, and an early over-population of neurons is reduced via apoptosis (Carr and Simpson, 1978; Marusich et al., 1994; Rohrer and Thoenen, 2018).
As in other model systems, zebrafish sox10 plays a crucial role in the specification of neuronal lineages, as well as glia and pigment cells (Carney et al., 2006; Dutton et al., 2001; Kelsh and Eisen, 2000). sox10 mutants exhibit marked reduction of both DRG neurons and the associated Schwann cells and satellite glia (Carney et al., 2006). However, the observed reduction in DRG neurons is not due to cell death or changes in proliferation rates, and DRG survival is not dependent on proximity to glial cells. Instead, these mutants show reduced expression of the proneural gene neurog1 and exhibit frequent gaps in its stereotyped segmental expression pattern (Carney et al., 2006). Furthermore, Sox10 is transiently expressed in DRG sensory neuron progenitors, where it induces the expression of neurog1 (Carney et al., 2006). Neurog1, in turn, plays a key role in the formation of sensory neurons (Carney et al., 2006; Delfino-Machín et al., 2017; McGraw et al., 2008). Interestingly, if neurog1 function is lost, cells fated to become DRG neurons instead adopt a glial fate (McGraw et al., 2008). It is noteworthy that at earlier stages Neurog1 promotes RB sensory neuron fates, and at that point must be excluded from the NCCs; yet later, this same transcription factor acts within the NCCs to promote DRG fates.
The baz1/sox10 mutant, in which a single nucleotide substitution impairs the DNA-binding HMG domain, exhibits features typical of other sox10 mutant alleles but also displays supernumerary sensory neurons (Carney et al., 2006; Delfino-Machín et al., 2017). The baz1 allele retains the ability to drive expression of neurog1 (Delfino-Machín et al., 2017). Further, overexpression of neurog1 rescues the loss of sensory neurons in sox10 morphant embryos (Delfino-Machín et al., 2017), suggesting that the activation of neurog1 by Sox10 is critical for the establishment of neuronal fate. Additionally, lateral inhibition likely plays an important role in the balanced production of neurons and glia during DRG formation. Consistent with this model, notch1a, deltaA, and deltaD are expressed in non-neuronal cells associated with the DRGs (McGraw et al., 2012) and inhibition of Delta/Notch signaling results in an increased number of DRG neurons (Delfino-Machín et al., 2017; McGraw et al., 2012).
Enteric Neurons.
The enteric nervous system (ENS) of the zebrafish is derived entirely from the vagal neural crest (Figure 1), unlike in amniotes where the ENS arises from a combination of vagal and sacral NCCs (Shepherd and Eisen, 2011). The role of the ENS is to innervate the intestine to regulate various aspects of its function, including motility, secretion, and local blood flow (Ganz et al., 2016). In humans, improper development of the ENS can result in devastating syndromes, such as Hirschsprung’s disease where the distal portion of the intestinal tract lacks innervation and is thus unable to generate proper bowel movements (Ganz et al., 2016). The zebrafish has emerged as a useful model in which to interrogate ENS development.
Zebrafish ENS precursors enter the anterior portion of the digestive system at 32 hpf and migrate posteriorly along the length of the developing gut as two parallel chains, reaching its posterior end by 66 hpf (Elworthy et al., 2005; Olden et al., 2008; Shepherd et al., 2004). The expression of phox2b in ENS precursors is conserved in zebrafish, where it is necessary for proper development (Elworthy et al., 2005). Similarly, components of the GDNF/Ret signaling pathway are expressed in the subset of vagal NCCs that give rise to the ENS after the onset of migration towards the gut and play a crucial role in precursor migration (Shepherd et al., 2004, 2001). In zebrafish, ret is expressed as two isoforms, ret9 and ret51, and the Ret9 isoform is sufficient for colonization of the gut by enteric neurons (Heanue and Pachnis, 2008). Moreover, when gfra1a/gfra1b or ret are knocked down, enteric NCCs are still capable of expressing phox2b and entering the anterior gut, indicating that their initial specification is not affected. However, they are not able to migrate posteriorly or increase in numbers, resulting in markedly fewer enteric neurons along the length of the gut (Shepherd et al., 2004). These results, together with the fact that sox10 mutants exhibit reduced numbers of ENS neurons (Dutton et al., 2001; Kelsh and Eisen, 2000), suggest that aspects of ENS development are largely conserved across the vertebrates.
Olden and colleagues (2008) proposed that ENS development can be divided into four distinct phases. At 32-50 hpf, all precursors express crestin prior to neural differentiation. At 55-58 hpf, anterior precursors begin neuronal differentiation as a group and downregulate crestin, while posterior precursors retain crestin expression, do not yet undergo differentiation, and proliferate at a higher rate (Olden et al., 2008). At 62-69 hpf, anterior precursors no longer express crestin, while posterior precursors have begun downregulating its expression. Posterior precursors also begin to differentiate into neurons in an anterior to posterior wave. During this stage, proliferation rates are equal between anterior and posterior regions. Finally, at 69-99 hpf, posterior precursors no longer express crestin and continue to differentiate throughout the intestine while the rate of proliferation decreases (Olden et al., 2008). More recently, analysis of the spatial and temporal colocalization patterns of phox2b, ret, and sox10 revealed three major ENS progenitor subpopulations that are indicative of distinct developmental states (Taylor et al., 2016). Namely, less mature progenitors at the wave front express all three genes, while more mature progenitors lose expression of sox10 and/or ret (Taylor et al., 2016).
Although several factors involved in ENS specification and development have been uncovered, the mechanisms that regulate enteric NCC migration along the gut are less well understood. Because of their amenability for live imaging and experimental manipulations, zebrafish embryos are proving useful to probe this question in vivo. As an example, Uribe and colleagues demonstrated that Retinoic Acid (RA) is necessary for the migration and survival of enteric NCCs along the developing gut (Uribe et al., 2018). Specifically, RA functions during the time window when NCCs migrate into and along the foregut to maintain collective chain migration and ensure complete colonization of the gut (Uribe et al., 2018). This function is mediated at least in part by regulation of ret and meis3, genes previously shown to be required for colonization of the gut by NCCs (Uribe and Bronner, 2015).
Olfactory neurons.
The sense organs of the head, as well as neuronal portions of the cranial ganglia, have long been thought to derive from ectodermal placodes (Gans and Northcutt, 1983). However, the contribution of NCCs to these structures, particularly to the olfactory epithelium (OE), has been controversial. The OE arises from the olfactory ectodermal placode and gives rise primarily to ciliated and microvillous sensory neurons (Hansen and Zeiske, 1993), which are ensheathed by supporting glial cells that in chick arise from cranial NCCs (Barraud et al., 2010). Recently, photoconversion-based fate mapping coupled with live cell-tracking and laser ablation experiments demonstrated that Sox10-expressing cranial NCCs migrate into the OE to give rise to the microvillous sensory neurons, whereas the ciliated neurons are placode-derived (Saxena et al., 2013). Further, these authors used a Sox10 morpholino to show that Sox10 knockdown causes a depletion of microvillous neurons (Saxena et al., 2013).
A third subtype of neurons, expressing gonadotrophin releasing hormone 3 (gnrh3), is also associated with the OE. Previous analysis suggested that zebrafish Gnrh3-expressing cells, similar to microvillous neurons, rely on Sox10 function and are derived from NCCs (Whitlock et al., 2005, 2003). However, a more recent analysis used cell-lineage reconstructions—achieved by ‘backtracking’ cell movements through time-lapse datasets, as well as by following photoconverted cells forward through time—to establish that most if not all Gnrh3-expressing neurons are derived from the PPE (Aguillon et al., 2018). Moreover, these authors showed that the numbers of islet1/2-expressing Gnrh3 neurons in the OE were unaltered in sox10 mutants (Aguillon et al., 2018). Surprisingly, using similar approaches, these authors reported that the microvillous sensory neurons also derive from PPE, and develop independent of Sox10 function.
In summary, the studies of Saxena et al. (2013) and Aguillon et al. (2018) come to very different conclusions regarding the origins of the microvillous sensory neurons. Caution must be exercised when using transgene markers to follow cells—given the potential for ectopic expression sites to arise—yet this caveat seems inadequate to explain the discrepancy in results, as Saxena and colleagues not only traced NCCs to the microvillous neurons when using a transgene marker to “follow” NCCs, but obtained similar results when they labeled small numbers of NCCs via photoconversion and followed them from their neural place of origin all the way to the nasal cavity. Moving forward, ensuring a full and common understanding of the precise extent of the PPE will be critical to resolving the discrepancies between these independent studies. In the long term, it will be important to determine whether and how interactions between NCCs and placodal cells shape the development of the OE.
Conclusions and Perspectives
The neural crest is a remarkable cell population that continues to fascinate researchers. Its central role in the evolution of vertebrates, implications for human birth defects and cancer, and astonishing biology all continue to attract intensive investigation across a variety of model and non-model systems. In this review, we have highlighted the invaluable contributions of the zebrafish towards our understanding of NCC development.
Recently, systems biology approaches, including transcriptional and epigenomic profiling, have helped to define the gene regulatory circuits that govern neural crest development while providing new insights into underlying molecular mechanisms. For example, a recent zebrafish study led to the discovery that FoxD3 acts initially as a ‘pioneer factor’ that activates the expression of genes regulating NCC specification and migration, but later functions as a repressor by directly remodeling nucleosome and chromatin organization (Lukoseviciute et al., 2018). Nevertheless, an important limitation of modern systems biology approaches is that they often lack critical spatial information. As discussed above, a recent tour de force study in the mouse model has addressed this challenge by coupling single cell and spatial transcriptomics with lineage tracing, to reveal a series of branching cell fate bifurcation points in the NCC lineage, while also pinpointing key differences in trunk versus cranial potential (Soldatov et al. 2019). Because the sequence of fate restrictions described in murine NCCs does not correspond with the observed lineage tracing results in zebrafish, it will be critical to determine whether the process of transcriptional regulation that underlies cell fate decisions at these bifurcation points is conserved.
A major challenge that remains to be addressed is to deepen our understanding of how such intrinsic regulatory networks are influenced by the complex 3D-environment though which the NCCs migrate, a challenge that requires an appropriate balance of large-scale ‘omics approaches and in depth functional analyses rooted in the complexity of the underlying biology. To this end, recent approaches to enable the quantitative visualization of gene expression with cellular resolution in a multiplexed manner, such as hybridization chain reaction, will be a valuable tool for providing spatial context. One great advantage of the zebrafish is that over the years multiple transgenic reporters of key neural crest genes have been generated (Table 1); these tools allow real-time in vivo spatiotemporal insights into neural crest development that are unique to the zebrafish model. Going forward, we hope to see researchers finding new ways to productively merge genomic analyses with live imaging in the zebrafish, to ultimately allow a deeper integration of the spatial, temporal, and cellular context of neural crest development with our increasing understanding of the underlying molecular changes that direct its development.
Key Findings:
The zebrafish provides a powerful model for studies of neural crest
Despite teleost-specific neurulation mechanisms, the general principles of neural crest development are well conserved across vertebrates
The combination of genetic and transgenic tools with high-resolution live imaging is allowing the zebrafish model to provide unique insights into neural crest developmental mechanisms
ACKNOWLEDGEMENTS
We are grateful to our colleagues in the neural crest field for helpful conversations that have influenced this article, and we apologize to the many individuals whose excellent work we were unable to cite due to space limitations. In particular, we thank Jon Clarke for his input to the section on neurulation, and Mary Halloran, Ankur Saxena, and Robert Ho and members of his group, for many helpful discussions. We also thank the expert reviewers of the initial version of the manuscript, whose insightful comments helped us to make significant improvements. Related work in the Prince lab was funded by NSF award 1528911 and by the Chicago Biomedical Consortium with support from the Searle Funds at The Chicago Community Trust (award C-070 to VEP and Ankur Saxena). KA was supported by the George A. Hines Scholarship fund. MR and AB were supported by the Eunice Kennedy Shriver National Institute Of Child Health & Human Development of the National Institutes of Health under award number T32HD055164. This material is additionally based upon work supported by award number F31HD097957 to MR, and by the National Science Foundation Graduate Research Fellowship Program under Grant No #DGE-1144082 to AB. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the authors and do not necessarily reflect the views of the National Institutes of Health or the National Science Foundation. Finally, this work benefitted from the resources of the ZFIN database (zfin.org).
Grant Support: NSF 1528911; NSF DGE-1144082; NIH T32HD055164; NIH F31HD097957; CBC C-070
REFERENCES
- Abercrombie M, Heaysman JEM, 1953. Observations on the social behaviour of cells in tissue culture. I. Speed of movement of chick heart fibroblasts in relation to their mutual contacts. Exp. Cell Res 5, 111–131. 10.1016/0014-4827(53)90098-6 [DOI] [PubMed] [Google Scholar]
- Aguillon R, Batut J, Subramanian A, Madelaine R, Dufourcq P, Schilling TF, Blader P, 2018. Cell-type heterogeneity in the early zebrafish olfactory epithelium is generated from progenitors within preplacodal ectoderm. Elife 7, 1–17. 10.7554/eLife.32041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahsan K, Singh N, Rocha M, Huang C, Prince VE, 2019. Prickle1 is required for EMT and migration of zebrafish cranial neural crest. Dev. Biol 448, 16–35. 10.1016/j.ydbio.2019.01.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander C, Zuniga E, Blitz IL, Wada N, Le Pabic P, Javidan Y, Zhang T, Cho KW, Crump JG, Schilling TF, 2011. Combinatorial roles for BMPs and Endothelin 1 in patterning the dorsal-ventral axis of the craniofacial skeleton. Development 138, 5135–5146. 10.1242/dev.067801 [DOI] [PMC free article] [PubMed] [Google Scholar]
- An M, Luo R, Henion PD, 2002. Differentiation and maturation of zebrafish dorsal root and sympathetic ganglion neurons. J. Comp. Neurol 446, 267–275. 10.1002/cne.10214 [DOI] [PubMed] [Google Scholar]
- Arduini BL, Bosse KM, Henion PD, 2009. Genetic ablation of neural crest cell diversification. Development 136, 1987–1994. 10.1242/dev.033209 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Artinger KB, Chitnis a B., Mercola M, Driever W, 1999. Zebrafish narrowminded suggests a genetic link between formation of neural crest and primary sensory neurons. Development 126, 3969–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banerjee S, Gordon L, Donn TM, Berti C, Moens CB, Burden SJ, Granato M, 2011. A novel role for MuSK and non-canonical Wnt signaling during segmental neural crest cell migration. Development 138, 3287–3296. 10.1242/dev.067306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banerjee S, Isaacman-Beck J, Schneider VA, Granato M, 2013. A Novel Role for Lh3 Dependent ECM Modifications during Neural Crest Cell Migration in Zebrafish. PLoS One 8, 1–8. 10.1371/journal.pone.0054609 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrallo-Gimeno A, Holzschuh J, Driever W, Knapik EW, 2004. Neural crest survival and differentiation in zebrafish depends on mont blanc/tfap2a gene function. Development 131, 1463–1477. 10.1242/dev.01033 [DOI] [PubMed] [Google Scholar]
- Barraud P, Seferiadis AA, Tyson LD, Zwart MF, Szabo-Rogers HL, Ruhrberg C, Liu KJ, Baker CVH, 2010. Neural crest origin of olfactory ensheathing glia. Proc. Natl. Acad. Sci 107, 21040–21045. 10.1073/pnas.1012248107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barske L, Askary A, Zuniga E, Balczerski B, Bump P, Nichols JT, Crump JG, 2016. Competition between Jagged-Notch and Endothelin1 Signaling Selectively Restricts Cartilage Formation in the Zebrafish Upper Face. PLoS Genet. 12, 1–33. 10.1371/journal.pgen.1005967 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berndt JD, Clay MR, Langenberg T, Halloran MC, 2008. Rho-kinase and myosin II affect dynamic neural crest cell behaviors during epithelial to mesenchymal transition in vivo. Dev. Biol 324, 236–244. 10.1016/j.ydbio.2008.09.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berndt JD, Halloran MC, 2006. Semaphorin 3d promotes cell proliferation and neural crest cell development downstream of TCF in the zebrafish hindbrain. Development 133, 3983–92. 10.1242/dev.02583 [DOI] [PubMed] [Google Scholar]
- Bi W, Deng JM, Zhang Z, Behringer RR, de Crombrugghe B, 1999. Sox9 is required for cartilage formation. Nat. Genet 22, 85–9. 10.1038/8792 [DOI] [PubMed] [Google Scholar]
- Birkholz DA, Olesnicky Killian EC, George KM, Artinger KB, 2009. Prdm1a is necessary for posterior pharyngeal arch development in zebrafish. Dev. Dyn 238, 2575–2587. 10.1002/dvdy.22090 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boer EF, Howell ED, Schilling TF, Jette CA, Stewart RA, 2015. Fascin1-Dependent Filopodia are Required for Directional Migration of a Subset of Neural Crest Cells. PLoS Genet 11, 1–25. 10.1371/journal.pgen.1004946 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bronner ME, Simões-Costa M, 2016. The Neural Crest Migrating into the Twenty-First Century. Curr. Top. Dev. Biol 116, 115–34. 10.1016/bs.ctdb.2015.12.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buckley CE, Ren X, Ward LC, Girdler GC, Araya C, Green MJ, Clark BS, Link BA, Clarke JDW, 2013. Mirror-symmetric microtubule assembly and cell interactions drive lumen formation in the zebrafish neural rod. EMBO J. 32, 30–44. 10.1038/emboj.2012.305 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Budi EH, Patterson LB, Parichy DM, 2011. Post-embryonic nerve-associated precursors to adult pigment cells: Genetic requirements and dynamics of morphogenesis and differentiation. PLoS Genet. 7 10.1371/journal.pgen.1002044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campbell K, Casanova J, 2016. A common framework for EMT and collective cell migration. Development 143, 4291–4300. 10.1242/dev.139071 [DOI] [PubMed] [Google Scholar]
- Carmona-Fontaine C, Matthews HK, Kuriyama S, Moreno M, Dunn GA, Parsons M, Stern CD, Mayor R, 2008. Contact inhibition of locomotion in vivo controls neural crest directional migration. Nature 456, 957–61. 10.1038/nature07441 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carmona-Fontaine C, Theveneau E, Tzekou A, Tada M, Woods M, Page KM, Parsons M, Lambris JD, Mayor R, 2011. Complement Fragment C3a Controls Mutual Cell Attraction during Collective Cell Migration. Dev. Cell 21, 1026–1037. 10.1016/j.devcel.2011.10.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carney TJ, Dutton KA, Greenhill E, Delfino-Machin M, Dufourcq P, Blader P, Kelsh RN, 2006. A direct role for Sox10 in specification of neural crest-derived sensory neurons. Development 133, 4619–4630. 10.1242/dev.02668 [DOI] [PubMed] [Google Scholar]
- Carr VM, Simpson SB, 1978. Proliferative and degenerative events in the early development of chick dorsal root ganglia. I. Normal development. J. Comp. Neurol 182, 727–39. 10.1002/cne.901820410 [DOI] [PubMed] [Google Scholar]
- Cavanaugh AM, Huang J, Chen JN, 2015. Two developmentally distinct populations of neural crest cells contribute to the zebrafish heart. Dev. Biol 404, 103–112. 10.1016/j.ydbio.2015.06.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ciruna B, Jenny A, Lee D, Mlodzik M, Schier AF, 2006. Planar cell polarity signalling couples cell division and morphogenesis during neurulation. Nature 439, 220–4. 10.1038/nature04375 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke J, 2009. Role of polarized cell divisions in zebrafish neural tube formation. Curr. Opin. Neurobiol 19, 134–138. 10.1016/j.conb.2009.04.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clay MR, Halloran MC, 2014. Cadherin 6 promotes neural crest cell detachment via F-actin regulation and influences active Rho distribution during epithelial-to-mesenchymal transition. Development 141, 2506–2515. 10.1242/dev.105551 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clay MR, Halloran MC, 2013. Rho activation is apically restricted by Arhgap1 in neural crest cells and drives epithelial-to-mesenchymal transition. Development 140, 3198–3209. 10.1242/dev.095448 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clouthier DE, Schilling TF, 2004. Understanding endothelin-1 function during craniofacial development in the mouse and zebrafish. Birth Defects Res. Part C - Embryo Today Rev 72, 190–199. 10.1002/bdrc.20007 [DOI] [PubMed] [Google Scholar]
- Cooper CD, 2017. Insights from zebrafish on human pigment cell disease and treatment. Dev. Dyn 246, 889–896. 10.1002/dvdy.24550 [DOI] [PubMed] [Google Scholar]
- Cornell RA, Eisen JS, 2002. Delta/Notch signaling promotes formation of zebrafish neural crest by repressing Neurogenin 1 function. Development 129, 2639–48. 10.1126/science.282.5394.1711 [DOI] [PubMed] [Google Scholar]
- Cornell RA, Eisen JS, 2000. Delta signaling mediates segregation of neural crest and spinal sensory neurons from zebrafish lateral neural plate. Development 127, 2873–82. [DOI] [PubMed] [Google Scholar]
- Crump JG, Maves L, Lawson ND, Weinstein BM, Kimmel CB, 2004a. An essential role for Fgfs in endodermal pouch formation influences later craniofacial skeletal patterning. Development 131, 5703–16. 10.1242/dev.01444 [DOI] [PubMed] [Google Scholar]
- Crump JG, Swartz ME, Kimmel CB, 2004b. An integrin-dependent role of pouch endoderm in hyoid cartilage development. PLoS Biol. 2 10.1371/journal.pbio.0020244 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das A, Crump JG, 2012. Bmps and Id2a act upstream of twist1 to restrict ectomesenchyme potential of the cranial neural crest. PLoS Genet. 8 10.1371/journal.pgen.1002710 [DOI] [PMC free article] [PubMed] [Google Scholar]
- David NB, Saint-Etienne L, Tsang M, Schilling TF, Rosa FM, 2002. Requirement for endoderm and FGF3 in ventral head skeleton formation. Development 129, 4457–4468. [DOI] [PubMed] [Google Scholar]
- De Calisto J, Araya C, Marchant L, Riaz CF, Mayor R, 2005. Essential role of non-canonical Wnt signalling in neural crest migration. Development 132, 2587–97. 10.1242/dev.01857 [DOI] [PubMed] [Google Scholar]
- Delfino-Machín M, Madelaine R, Busolin G, Nikaido M, Colanesi S, Camargo-Sosa K, Law EWP, Toppo S, Blader P, Tiso N, Kelsh RN, 2017. Sox10 contributes to the balance of fate choice in dorsal root ganglion progenitors. PLoS One 12, 1–29. 10.1371/journal.pone.0172947 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickmeis T, Rastegar S, Lam CS, Aanstad P, Clark M, Fischer N, Rosa F, Korzh V, Strähle U, 2002. Expression of the helix-loop-helix gene id3 in the zebrafish embryo. Mech. Dev 113, 99–102. 10.1016/S0925-4773(02)00006-0 [DOI] [PubMed] [Google Scholar]
- Dooley CM, Mongera A, Walderich B, Nusslein-Volhard C, 2013. On the embryonic origin of adult melanophores: the role of ErbB and Kit signalling in establishing melanophore stem cells in zebrafish. Development 140, 1003–1013. 10.1242/dev.087007 [DOI] [PubMed] [Google Scholar]
- Dorsky RI, Moon RT, Raible DW, 1998. Control of neural crest cell fate by the Wnt signalling pathway. Nature 396, 370–373. 10.1038/24620 [DOI] [PubMed] [Google Scholar]
- Dorsky RI, Raible DW, Moon RT, 2000. Direct regulation of nacre, a zebrafish MITF homolog required for pigment cell formation, by the Wnt pathway. Genes Dev. 14, 158–62. 10.1101/gad.14.2.158 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dougherty M, Kamel G, Grimaldi M, Gfrerer L, Shubinets V, Ethier R, Hickey G, Cornell RA, Liao EC, 2013. Distinct requirements for wnt9a and irf6 in extension and integration mechanisms during zebrafish palate morphogenesis. Development 140, 76–81. 10.1242/dev.080473 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Driever W, Schier AF, Neuhauss SCF, Malicki J, Stemple DL, Stainier DYR, Zwartkruis F, Abdelilah S, Rangini Z, Belak J, Boggs C, 1996. A genetic screen for mutations affecting embryogenesis in zebrafish. Development 123, 37–46. [DOI] [PubMed] [Google Scholar]
- Dutton KA, Pauliny A, Lopes SS, Elworthy S, Carney TJ, Rauch J, Geisler R, Haffter P, Kelsh RN, 2001. Zebrafish colourless encodes sox10 and specifies non-ectomesenchymal neural crest fates. Development 128, 4113–25. 10.1093/hmg/4.12.2407 [DOI] [PubMed] [Google Scholar]
- Eberhart JK, 2006. Early Hedgehog signaling from neural to oral epithelium organizes anterior craniofacial development. Development 133, 1069–1077. 10.1242/dev.02281 [DOI] [PubMed] [Google Scholar]
- Eberhart JK, He X, Swartz ME, Yan YL, Song H, Boling TC, Kunerth AK, Walker MB, Kimmel CB, Postlethwait JH, 2008. MicroRNA Mirn140 modulates Pdgf signaling during palatogenesis. Nat. Genet 40, 290–298. 10.1038/ng.82 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elworthy S, Lister JA, Carney TJ, Raible DW, Kelsh RN, 2003. Transcriptional regulation of mitfa accounts for the sox10 requirement in zebrafish melanophore development. Development 130, 2809–18. 10.1242/dev.00461 [DOI] [PubMed] [Google Scholar]
- Elworthy S, Pinto JP, Pettifer A, Cancela ML, Kelsh RN, 2005. Phox2b function in the enteric nervous system is conserved in zebrafish and is sox10-dependent. Mech. Dev 122, 659–669. 10.1016/j.mod.2004.12.008 [DOI] [PubMed] [Google Scholar]
- Eom DS, Bain EJ, Patterson LB, Grout ME, Parichy DM, 2015. Long-distance communication by specialized cellular projections during pigment pattern development and evolution. Elife 4, 1–25. 10.7554/eLife.12401 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eom DS, Parichy DM, 2017. A macrophage relay for long-distance signaling during postembryonic tissue remodeling. Science (80-.). 1320, 1317–1320. 10.1126/science.aam6622 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erickson CA, Perris R, 1993. The role of cell-cell and cell-matrix interactions in the morphogenesis of the neural crest. Dev. Biol 159, 60–74. 10.1006/dbio.1993.1221 [DOI] [PubMed] [Google Scholar]
- Friedl P, Gilmour D, 2009. Collective cell migration in morphogenesis, regeneration and cancer. Nat. Rev. Mol. Cell Biol 10, 445–57. 10.1038/nrm2720 [DOI] [PubMed] [Google Scholar]
- Friedl P, Wolf K, 2003. Tumour-cell invasion and migration: diversity and escape mechanisms. Nat. Rev. Cancer 3, 362–74. 10.1038/nrc1075 [DOI] [PubMed] [Google Scholar]
- Frohnhofer HG, Krauss J, Maischein H-M, Nusslein-Volhard C, 2013. Iridophores and their interactions with other chromatophores are required for stripe formation in zebrafish. Development 140, 2997–3007. 10.1242/dev.096719 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gallik KL, Treffy RW, Nacke LM, Ahsan K, Rocha M, Green-Saxena A, Saxena A, 2017. Neural crest and cancer: Divergent travelers on similar paths. Mech. Dev 148, 89–99. 10.1016/j.mod.2017.08.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gans C, Northcutt RG, 1983. Neural Crest and the Origin of Vertebrates: A New Head. Science (80-.). 220, 268–273. 10.1126/science.220.4594.268 [DOI] [PubMed] [Google Scholar]
- Ganz J, Melancon E, Eisen JS, 2016. Zebrafish as a model for understanding enteric nervous system interactions in the developing intestinal tract. Methods Cell Biol. 134, 139–64. 10.1016/bs.mcb.2016.02.003 [DOI] [PubMed] [Google Scholar]
- Garnett a. T., Square T. a., Medeiros DM, 2012. BMP, Wnt and FGF signals are integrated through evolutionarily conserved enhancers to achieve robust expression of Pax3 and Zic genes at the zebrafish neural plate border. Development 139, 4220–4231. 10.1242/dev.081497 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Germanguz I, Lev D, Waisman T, Kim CH, Gitelman I, 2007. Four twist genes in zebrafish, four expression patterns. Dev. Dyn 236, 2615–2626. 10.1002/dvdy.21267 [DOI] [PubMed] [Google Scholar]
- Granato M, Nüsslein-Volhard C, 1996. Fishing for genes controlling development. Curr. Opin. Genet. Dev 6, 461–468. 10.1016/S0959-437X(96)80068-2 [DOI] [PubMed] [Google Scholar]
- Groves AK, LaBonne C, 2014. Setting appropriate boundaries: Fate, patterning and competence at the neural plate border. Dev. Biol 389, 2–12. 10.1016/j.ydbio.2013.11.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haffter P, Granato M, Brand M, Mullins MC, Hammerschmidt M, Kane DA, Odenthal J, van Eeden FJ, Jiang YJ, Heisenberg CP, Kelsh RN, Furutani-Seiki M, Vogelsang E, Beuchle D, Schach U, Fabian C, Nüsslein-Volhard C, 1996. The identification of genes with unique and essential functions in the development of the zebrafish, Danio rerio. Development 123, 1–36. [DOI] [PubMed] [Google Scholar]
- Hall BK, 2000. The neural crest as a fourth germ layer and vertebrates as quadroblastic not triploblastic. Evol. Dev 2, 3–5. 10.1046/j.1525-142X.2000.00032.x [DOI] [PubMed] [Google Scholar]
- Halloran MC, Berndt JD, 2003. Current Progress in Neural Crest Cell Motility and Migration and Future Prospects for the Zebrafish Model System. Dev. Dyn 228, 497–513. 10.1002/dvdy.10374 [DOI] [PubMed] [Google Scholar]
- Halloran MC, Severance SM, Yee CS, Gemza DL, Raper JA, Kuwada JY, 1999. Analysis of a zebrafish semaphorin reveals potential functions in vivo. Dev. Dyn 214, 13–25. 10.1002/(SICI)1097-0177(199901)214:1<13::AID-DVDY2>3.0.CO;2-3 [DOI] [PubMed] [Google Scholar]
- Hamada H, Watanabe M, Lau HE, Nishida T, Hasegawa T, Parichy DM, Kondo S, 2014. Involvement of Delta/Notch signaling in zebrafish adult pigment stripe patterning. Development 141, 318–24. 10.1242/dev.099804 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammerschmidt M, Serbedzija GN, McMahon AP, 1996. Genetic analysis of dorsoventral pattern formation in the zebrafish: Requirement of a BMP-like ventralizing activity and its dorsal repressor. Genes Dev. 10, 2452–2461. 10.1101/gad.10.19.2452 [DOI] [PubMed] [Google Scholar]
- Hansen A, Zeiske E, 1993. Development of the olfactory organ in the zebrafish, Brachydanio rerio. J. Comp. Neurol 333, 289–300. 10.1002/cne.903330213 [DOI] [PubMed] [Google Scholar]
- Heanue TA, Pachnis V, 2008. Ret isoform function and marker gene expression in the enteric nervous system is conserved across diverse vertebrate species. Mech. Dev 125, 687–699. 10.1016/j.mod.2008.04.006 [DOI] [PubMed] [Google Scholar]
- Henion PD, Raible DW, Beattie CE, Stoesser KL, Weston J. a, Eisen JS, 1996. Screen for mutations affecting development of Zebrafish neural crest. Dev. Genet 18, 11–17. 10.1002/(SICI)1520-6408(1996)18:1<11::AID-DVG2>3.0.CO;2-4 [DOI] [PubMed] [Google Scholar]
- Hernandez-Lagunas L, Choi IF, Kaji T, Simpson P, Hershey C, Zhou Y, Zon L, Mercola M, Artinger KB, 2005. Zebrafish narrowminded disrupts the transcription factor prdm1 and is required for neural crest and sensory neuron specification. Dev. Biol 278, 347–357. 10.1016/j.ydbio.2004.11.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffman TL, Javier AL, Campeau SA, Knight RD, Schilling TF, 2007. Tfap2 transcription factors in zebrafish neural crest development and ectodermal evolution. J. Exp. Zool. B. Mol. Dev. Evol 308, 679–91. 10.1002/jez.b.21189 [DOI] [PubMed] [Google Scholar]
- Hong E, Brewster R, 2006. N-cadherin is required for the polarized cell behaviors that drive neurulation in the zebrafish. Development 133, 3895–905. 10.1242/dev.02560 [DOI] [PubMed] [Google Scholar]
- Hong SK, Tsang M, Dawid IB, 2008. The Mych gene is required for neural crest survival during zebrafish development. PLoS One 3 10.1371/journal.pone.0002029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honjo Y, Eisen JS, 2005. Slow muscle regulates the pattern of trunk neural crest migration in zebrafish. Development 132, 4461–4470. 10.1242/dev.02026 [DOI] [PubMed] [Google Scholar]
- Honjo Y, Kniss J, Eisen JS, 2008. Neuregulin-mediated ErbB3 signaling is required for formation of zebrafish dorsal root ganglion neurons. Development 135, 2993–2993. 10.1242/dev.027763 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hörstadius S, 1950. The neural crest: its properties and derivatives in the light of experimental research. Oxford University Press. [Google Scholar]
- Hultman KA, Johnson SL, 2010. Differential contribution of direct-developing and stem cell-derived melanocytes to the zebrafish larval pigment pattern. Dev. Biol 337, 425–431. 10.1016/j.ydbio.2009.11.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutchins EJ, Kunttas E, Piacentino ML, Howard AGA, Bronner ME, Uribe RA, 2018. Migration and diversification of the vagal neural crest. Dev. Biol 444 10.1016/j.ydbio.2018.07.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Irion U, Krauss J, Nusslein-Volhard C, 2014. Precise and efficient genome editing in zebrafish using the CRISPR/Cas9 system. Development 141, 4827–4830. 10.1242/dev.115584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Irion U, Singh AP, Nüsslein-Volhard C, 2016. The Developmental Genetics of Vertebrate Color Pattern Formation: Lessons from Zebrafish. Curr. Top. Dev. Biol 117, 141–169. 10.1016/bs.ctdb.2015.12.012 [DOI] [PubMed] [Google Scholar]
- Jiang YJ, Brand M, Heisenberg CP, Beuchle D, Furutani-Seiki M, Kelsh RN, Warga RM, Granato M, Haffter P, Hammerschmidt M, Kane DA, Mullins MC, Odenthal J, van Eeden FJ, Nusslein-Volhard C, 1996. Mutations affecting neurogenesis and brain morphology in the zebrafish, Danio rerio. Development 123, 205–216. 10.1242/dev.025148 [DOI] [PubMed] [Google Scholar]
- Jimenez L, Wang J, Morrison MA, Whatcott C, Soh KK, Warner S, Bearss D, Jette CA, Stewart RA, 2016. Phenotypic chemical screening using a zebrafish neural crest EMT reporter identifies retinoic acid as an inhibitor of epithelial morphogenesis. Dis. Model. Mech 9, 389–400. 10.1242/dmm.021790 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson SL, Africa D, Walker C, Weston JA, 1995. Genetic control of adult pigment stripe development in zebrafish. Dev. Biol 167, 27–33. 10.1006/dbio.1995.1004 [DOI] [PubMed] [Google Scholar]
- Kague E, Gallagher M, Burke S, Parsons M, Franz-Odendaal T, Fisher S, 2012. Skeletogenic Fate of Zebrafish Cranial and Trunk Neural Crest. PLoS One 7, 1–13. 10.1371/journal.pone.0047394 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelsh RN, Brand M, Jiang YJ, Heisenberg CP, Lin S, Haffter P, Odenthal J, Mullins MC, van Eeden FJ, Furutani-Seiki M, Granato M, Hammerschmidt M, Kane DA, Warga RM, Beuchle D, Vogelsang L, Nusslein-Volhard C, 1996. Zebrafish pigmentation mutations and the processes of neural crest development. Development 123, 369–389. [DOI] [PubMed] [Google Scholar]
- Kelsh RN, Eisen JS, 2000. The zebrafish colourless gene regulates development of non-ectomesenchymal neural crest derivatives. Development 127, 515–525. 10.1242/dev.065193 [DOI] [PubMed] [Google Scholar]
- Kelsh RN, Raible DW, 2002. Specification of zebrafish neural crest. Results Probl Cell Differ 40, 216–236. [DOI] [PubMed] [Google Scholar]
- Kimmel CB, Ullmann B, Walker M, Miller CT, Crump JG, 2003. Endothelin 1-mediated regulation of pharyngeal bone development in zebrafish. Development 130, 1339–51. 10.1242/dev.00338 [DOI] [PubMed] [Google Scholar]
- Knight RD, 2003. Lockjaw Encodes a Zebrafish Tfap2a Required for Early Neural Crest Development. Development 130, 5755–5768. 10.1242/dev.00575 [DOI] [PubMed] [Google Scholar]
- Knight RD, Javidan Y, Nelson S, Zhang T, Schilling TF, 2004. Skeletal and Pigment Cell Defects in the lockjaw Mutant Reveal Multiple Roles for Zebrafish tfap2a in Neural Crest Development. Dev. Dyn 229, 87–98. 10.1002/dvdy.10494 [DOI] [PubMed] [Google Scholar]
- Knight RD, Schilling TF, 2006. Cranial neural crest and development of the head skeleton. Adv. Exp. Med. Biol 589, 120–33. 10.1007/978-0-387-46954-6_7 [DOI] [PubMed] [Google Scholar]
- Lamborghini JE, 1980. Rohon‐beard cells and other large neurons in Xenopus embryos originate during gastrulation. J. Comp. Neurol 189, 323–333. 10.1002/cne.901890208 [DOI] [PubMed] [Google Scholar]
- Le Douarin N, 1973. A biological cell labeling technique and its use in experimental embryology. Dev. Biol 30, 217–222. 10.1016/0012-1606(73)90061-4 [DOI] [PubMed] [Google Scholar]
- Le Douarin N, Kalcheim C, 1982. The Neural Crest, Cambridge New York: Cambridge University Press. [Google Scholar]
- Le Lièvre CS, Le Douarin NM, 1975. Mesenchymal derivatives of the neural crest: analysis of chimaeric quail and chick embryos. J. Embryol. Exp. Morphol 34, 125–154. [PubMed] [Google Scholar]
- Lee JM, Dedhar S, Kalluri R, Thompson EW, 2006. The epithelial-mesenchymal transition: new insights in signaling, development, and disease. J. Cell Biol 172, 973–81. 10.1083/jcb.200601018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee RTH, Knapik EW, Thiery JP, Carney TJ, 2013a. An exclusively mesodermal origin of fin mesenchyme demonstrates that zebrafish trunk neural crest does not generate ectomesenchyme. Development 140, 2923–2932. 10.1242/dev.093534 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee RTH, Thiery JP, Carney TJ, 2013b. Dermal fin rays and scales derive from mesoderm, not neural crest. Curr. Biol 23, R336–7. 10.1016/j.cub.2013.02.055 [DOI] [PubMed] [Google Scholar]
- Leigh NR, Schupp MO, Li K, Padmanabhan V, Gastonguay A, Wang L, Chun CZ, Wilkinson GA, Ramchandran R, 2013. Mmp17b Is Essential for Proper Neural Crest Cell Migration In Vivo. PLoS One 8, 1–12. 10.1371/journal.pone.0076484 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis JL, Bonner J, Modrell M, Ragland JW, Moon RT, Dorsky RI, Raible DW, 2004. Reiterated Wnt signaling during zebrafish neural crest development. Development 131, 1299–1308. 10.1242/dev.01007 [DOI] [PubMed] [Google Scholar]
- Li M, Zhao C, Wang Y, Zhao Z, Meng A, 2002. Zebrafish sox9b is an early neural crest marker. Dev. Genes Evol 212, 203–206. 10.1007/s00427-002-0235-2 [DOI] [PubMed] [Google Scholar]
- Li W, Cornell RA, 2007. Redundant activities of Tfap2a and Tfap2c are required for neural crest induction and development of other non-neural ectoderm derivatives in zebrafish embryos. Dev. Biol 304, 338–354. 10.1016/j.ydbio.2006.12.042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li YX, Zdanowicz M, Young L, Kumiski D, Leatherbury L, Kirby ML, 2003. Cardiac neural crest in zebrafish embryos contributes to myocardial cell lineage and early heart function. Dev. Dyn 226, 540–550. 10.1002/dvdy.10264 [DOI] [PubMed] [Google Scholar]
- Liem KF, Tremml G, Roelink H, Jessell TM, 1995. Dorsal differentiation of neural plate cells induced by BMP-mediated signals from epidermal ectoderm. Cell 82, 969–979. 10.1016/0092-8674(95)90276-7 [DOI] [PubMed] [Google Scholar]
- Lister J, Robertson C, Lepage T, Johnson S, Raible D, 1999. Nacre Encodes a Zebrafish Microphthalmia-Related Protein That Regulates Neural-Crest-Derived Pigment Cell Fate. Development 126, 3757–3767. [DOI] [PubMed] [Google Scholar]
- Lister JA, Cooper C, Nguyen K, Modrell M, Grant K, Raible DW, 2006. Zebrafish Foxd3 is required for development of a subset of neural crest derivatives. Dev. Biol 290, 92–104. 10.1016/j.ydbio.2005.11.014 [DOI] [PubMed] [Google Scholar]
- Lowery LA, Sive H, 2004. Strategies of vertebrate neurulation and a re-evaluation of teleost neural tube formation. Mech. Dev 121, 1189–1197. 10.1016/j.mod.2004.04.022 [DOI] [PubMed] [Google Scholar]
- Lukoseviciute M, Gavriouchkina D, Williams RM, Hochgreb-Hagele T, Senanayake U, Chong-Morrison V, Thongjuea S, Repapi E, Mead A, Sauka-Spengler T, 2018. From Pioneer to Repressor: Bimodal foxd3 Activity Dynamically Remodels Neural Crest Regulatory Landscape In Vivo. Dev. Cell 47, 608–628.e6. 10.1016/j.devcel.2018.11.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maderspacher F, Nüsslein-Volhard C, 2003. Formation of the adult pigment pattern in zebrafish requires leopard and obelix dependent cell interactions. Development 130, 3447–57. 10.1242/dev.00519 [DOI] [PubMed] [Google Scholar]
- Martik ML, Bronner ME, 2017. Regulatory Logic Underlying Diversification of the Neural Crest. Trends Genet. 33, 715–727. 10.1016/j.tig.2017.07.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marusich MF, Furneaux HM, Henion PD, Weston JA, 1994. Hu neuronal proteins are expressed in proliferating neurogenic cells. J. Neurobiol 25, 143–155. 10.1002/neu.480250206 [DOI] [PubMed] [Google Scholar]
- Matthews HK, Marchant L, Carmona-Fontaine C, Kuriyama S, Larrain J, Holt MR, Parsons M, Mayor R, 2008. Directional migration of neural crest cells in vivo is regulated by Syndecan-4/Rac1 and non-canonical Wnt signaling/RhoA. Development 135, 1771–1780. 10.1242/dev.017350 [DOI] [PubMed] [Google Scholar]
- Mayor R, Theveneau E, 2013. The neural crest. Development 140, 2247–51. 10.1242/dev.091751 [DOI] [PubMed] [Google Scholar]
- McGraw HF, Nechiporuk A, Raible DW, 2008. Zebrafish Dorsal Root Ganglia Neural Precursor Cells Adopt a Glial Fate in the Absence of Neurogenin1. J. Neurosci 28, 12558–12569. 10.1523/JNEUROSCI.2079-08.2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGraw HF, Snelson CD, Prendergast A, Suli A, Raible DW, 2012. Postembryonic neuronal addition in Zebrafish dorsal root ganglia is regulated by Notch signaling. Neural Dev. 7, 1–13. 10.1186/1749-8104-7-23 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLennan R, Dyson L, Prather KW, Morrison JA, Baker RE, Maini PK, Kulesa PM, 2012. Multiscale mechanisms of cell migration during development: theory and experiment. Development 139, 2935–44. 10.1242/dev.081471 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLennan R, Schumacher LJ, Morrison JA, Teddy JM, Ridenour DA, Box AC, Semerad CL, Li H, McDowell W, Kay D, Maini PK, Baker RE, Kulesa PM, 2015a. Neural crest migration is driven by a few trailblazer cells with a unique molecular signature narrowly confined to the invasive front. Development 142, 2014–2025. 10.1242/dev.117507 [DOI] [PubMed] [Google Scholar]
- McLennan R, Schumacher LJ, Morrison JA, Teddy JM, Ridenour DA, Box AC, Semerad CL, Li H, McDowell W, Kay D, Maini PK, Baker RE, Kulesa PM, 2015b. VEGF signals induce trailblazer cell identity that drives neural crest migration. Dev. Biol 407, 12–25. 10.1016/j.ydbio.2015.08.011 [DOI] [PubMed] [Google Scholar]
- Medeiros DM, Crump JG, 2012. New perspectives on pharyngeal dorsoventral patterning in development and evolution of the vertebrate jaw. Dev. Biol 371, 121–135. 10.1016/j.ydbio.2012.08.026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meinecke L, Sharma PP, Du H, Zhang L, Nie Q, Schilling TF, 2018. Modeling craniofacial development reveals spatiotemporal constraints on robust patterning of the mandibular arch, PLoS Computational Biology. 10.1371/journal.pcbi.1006569 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Metcalfe WK, Myers PZ, Trevarrow B, Bass MB, Kimmel CB, 1990. Primary neurons that express the L2/HNK-1 carbohydrate during early development in the zebrafish. Development 110, 491–504. [DOI] [PubMed] [Google Scholar]
- Meulemans D, Bronner-Fraser M, 2004. Gene-regulatory interactions in neural crest evolution and development. Dev. Cell 7, 291–299. 10.1016/j.devcel.2004.08.007 [DOI] [PubMed] [Google Scholar]
- Miller CT, Schilling TF, Lee K, Parker J, Kimmel CB, 2000. sucker encodes a zebrafish Endothelin-1 required for ventral pharyngeal arch development. Development 127, 3815–28. [DOI] [PubMed] [Google Scholar]
- Miller CT, Yelon D, Stainier DYR, Kimmel CB, 2003. Two endothelin 1 effectors, hand2 and bapx1, pattern ventral pharyngeal cartilage and the jaw joint. Development 130, 1353–65. 10.1242/dev.00339 [DOI] [PubMed] [Google Scholar]
- Minoux M, Rijli FM, 2010. Molecular mechanisms of cranial neural crest cell migration and patterning in craniofacial development. Development 137, 2605–2621. [DOI] [PubMed] [Google Scholar]
- Mongera A, Singh AP, Levesque MP, Chen Y-Y, Konstantinidis P, Nusslein-Volhard C, 2013. Genetic lineage labeling in zebrafish uncovers novel neural crest contributions to the head, including gill pillar cells. Development 140, 916–925. 10.1242/dev.091066 [DOI] [PubMed] [Google Scholar]
- Montero-Balaguer M, Lang MR, Sachdev SW, Knappmeyer C, Stewart RA, De La Guardia A, Hatzopoulos AK, Knapik EW, 2006. The mother superior mutation ablates foxd3 activity in neural crest progenitor cells and depletes neural crest derivatives in zebrafish. Dev. Dyn 235, 3199–3212. 10.1002/dvdy.20959 [DOI] [PubMed] [Google Scholar]
- Moody SA, 1989. Quantitative lineage analysis of the origin of frog primary motor and sensory neurons from cleavage stage blastomeres. J. Neurosci 9, 2919–30. 10.1523/jneurosci.09-08-02919 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore R, Theveneau E, Pozzi S, Alexandre P, Richardson J, Merks A, Parsons M, Kashef J, Linker C, Mayor R, 2013. Par3 controls neural crest migration by promoting microtubule catastrophe during contact inhibition of locomotion. Development 140, 4763–4775. 10.1242/dev.098509 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morin-Kensicki EM, Eisen JS, 1997. Sclerotome development and peripheral nervous system segmentation in embryonic zebrafish. Development 124, 159–167. [DOI] [PubMed] [Google Scholar]
- Mork L, Crump G, 2015. Zebrafish Craniofacial Development: A Window into Early Patterning. Curr. Top. Dev. Biol 115, 235–69. 10.1016/bs.ctdb.2015.07.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moury JD, Jacobson AG, 1990. The origins of neural crest cells in the axolotl. Dev. Biol 141, 243–253. 10.1016/0012-1606(90)90380-2 [DOI] [PubMed] [Google Scholar]
- Moury JD, Jacobson AG, 1989. Neural fold formation at newly created boundaries between neural plate and epidermis in the axolotl. Dev. Biol 133, 44–57. [DOI] [PubMed] [Google Scholar]
- Muñoz WA, Trainor PA, 2015. Neural crest cell evolution: how and when did a neural crest cell become a neural crest cell. Curr. Top. Dev. Biol 111, 3–26. 10.1016/bs.ctdb.2014.11.001 [DOI] [PubMed] [Google Scholar]
- Nair S, Li W, Cornell R, Schilling TF, 2007. Requirements for Endothelin type-A receptors and Endothelin-1 signaling in the facial ectoderm for the patterning of skeletogenic neural crest cells in zebrafish. Development 134, 335–345. 10.1242/dev.02704 [DOI] [PubMed] [Google Scholar]
- Nakamasu A, Takahashi G, Kanbe A, Kondo S, 2009. Interactions between zebrafish pigment cells responsible for the generation of Turing patterns. Proc. Natl. Acad. Sci. U. S. A 106, 8429–34. 10.1073/pnas.0808622106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Narboux-Neme N, Ekker M, Levi G, Heude E, 2019. Posterior axis formation requires Dlx5/Dlx6 expression at the neural plate border. PLoS One 14, e0214063 10.1371/journal.pone.0214063 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nasevicius A, Ekker SC, 2000. Effective targeted gene “knockdown” in zebrafish. Nat. Genet 26, 216–20. 10.1038/79951 [DOI] [PubMed] [Google Scholar]
- Neave B, Holder N, Patient R, 1997. A graded response to BMP-4 spatially coordinates patterning of the mesoderm and ectoderm in the zebrafish. Mech. Dev 62, 183–195. 10.1016/S0925-4773(97)00659-X [DOI] [PubMed] [Google Scholar]
- Newgreen DF, Powell ME, Moser B, 1990. Spatiotemporal changes in HNK-1/L2 glycoconjugates on avian embryo somite and neural crest cells. Dev. Biol 139, 100–120. 10.1016/0012-1606(90)90282-N [DOI] [PubMed] [Google Scholar]
- Nguyen VH, Schmid B, Trout J, Connors S. a, Ekker M, Mullins MC, 1998. Ventral and lateral regions of the zebrafish gastrula, including the neural crest progenitors, are established by a bmp2b/swirl pathway of genes. Dev. Biol 199, 93–110. 10.1006/dbio.1998.8927 [DOI] [PubMed] [Google Scholar]
- Nieto MA, Huang RYYJ, Jackson RAA, Thiery JPP, 2016. Emt: 2016. Cell 166, 21–45. 10.1016/j.cell.2016.06.028 [DOI] [PubMed] [Google Scholar]
- Nikaido M, Tada M, Saji T, Ueno N, 1997. Conservation of BMP signaling in zebrafish mesoderm patterning. Mech. Dev 61, 75–88. 10.1016/S0925-4773(96)00625-9 [DOI] [PubMed] [Google Scholar]
- Noden DM, 1988. Interactions and fates of avian craniofacial mesenchyme. Development 103 Suppl, 121–40. [DOI] [PubMed] [Google Scholar]
- Noden DM, 1983. The role of the neural crest in patterning of avian cranial skeletal, connective, and muscle tissues. Dev. Biol 96, 144–165. 10.1016/0012-1606(83)90318-4 [DOI] [PubMed] [Google Scholar]
- Northcutt RG, Gans C, 1983. The genesis of neural crest and epidermal placodes: a reinterpretation of vertebrate origins. Q. Rev. Biol 58, 1–28. https://doi.org/6346380 [DOI] [PubMed] [Google Scholar]
- Odenthal J, Nüsslein-Volhard C, 1998. Fork Head Domain Genes in Zebrafish. Dev. Genes Evol 208, 245–258. 10.1007/s004270050179 [DOI] [PubMed] [Google Scholar]
- Odenthal J, Rossnagel K, Haffter P, Kelsh RN, Vogelsang E, Brand M, van Eeden FJ, Furutani-Seiki M, Granato M, Hammerschmidt M, Heisenberg CP, Jiang YJ, Kane DA, Mullins MC, Nüsslein-Volhard C, 1996. Mutations affecting xanthophore pigmentation in the zebrafish, Danio rerio. Development 123, 391–8. [DOI] [PubMed] [Google Scholar]
- Oh C. Do, Lu Y, Liang S, Mori-Akiyama Y, Chen D, De Crombrugghe B, Yasuda H, 2014. SOX9 regulates multiple genes in chondrocytes, including genes encoding ECM proteins, ECM modification enzymes, receptors, and transporters. PLoS One 9 10.1371/journal.pone.0107577 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olden T, Akhatar T, Beckman SA, Wallace KN, 2008. Differentiation of the zebrafish Enteric nervous system and intestinal smooth muscle. Genesis 46, 484–498. 10.1002/dvg.20429 [DOI] [PubMed] [Google Scholar]
- Olesnicky E, Hernandez-Lagunas L, Artinger KB, 2010. prdm1a Regulates sox10 and islet1 in the development of neural crest and Rohon-Beard sensory neurons. Genesis 48, 656–666. 10.1002/dvg.20673 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olesnicky Killian EC, Birkholz DA, Artinger KB, 2009. A role for chemokine signaling in neural crest cell migration and craniofacial development. Dev. Biol 333, 161–172. 10.1016/j.ydbio.2009.06.031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papan C, Campos-Ortega JA, 1994. On the formation of the neural keel and neural tube in the zebrafishDanio (Brachydanio) rerio. Roux’s Arch. Dev. Biol. Off. organ EDBO 203, 178–186. 10.1007/BF00636333 [DOI] [PubMed] [Google Scholar]
- Parichy DM, Turner JM, 2003. Zebrafish puma mutant decouples pigment pattern and somatic metamorphosis. Dev. Biol 256, 242–257. 10.1016/S0012-1606(03)00015-0 [DOI] [PubMed] [Google Scholar]
- Parichy DM, Turner JM, Parker NB, 2003. Essential role for puma in development of postembryonic neural crest-derived cell lineages in zebrafish. Dev. Biol 256, 221–241. 10.1016/S0012-1606(03)00016-2 [DOI] [PubMed] [Google Scholar]
- Patterson LB, Bain EJ, Parichy DM, 2014. Pigment cell interactions and differential xanthophore recruitment underlying zebrafish stripe reiteration and Danio pattern evolution. Nat. Commun 5, 1–9. 10.1038/ncomms6299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patterson LB, Parichy DM, 2013. Interactions with Iridophores and the Tissue Environment Required for Patterning Melanophores and Xanthophores during Zebrafish Adult Pigment Stripe Formation. PLoS Genet. 9, 1–14. 10.1371/journal.pgen.1003561 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phillips BT, Kwon HJ, Melton C, Houghtaling P, Fritz A, Riley BB, 2006. Zebrafish msxB, msxC and msxE function together to refine the neural-nonneural border and regulate cranial placodes and neural crest development. Dev. Biol 294, 376–390. 10.1016/j.ydbio.2006.03.001 [DOI] [PubMed] [Google Scholar]
- Piloto S, Schilling TF, 2010. Ovo1 links Wnt signaling with N-cadherin localization during neural crest migration. Development 137, 2603–2603. 10.1242/dev.055913 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Platt JB, 1893. Ectodermic origin of the cartilages of the head. Anat Anz 8, 506–509. [Google Scholar]
- Plein A, Fantin A, Ruhrberg C, 2015. Neural crest cells in cardiovascular development. Curr. Top. Dev. Biol 111, 183–200. 10.1016/bs.ctdb.2014.11.006 [DOI] [PubMed] [Google Scholar]
- Powell DR, Hernandez-Lagunas L, LaMonica K, Artinger KB, 2013. Prdm1a directly activates foxd3 and tfap2a during zebrafish neural crest specification. Development 140, 3445–3455. 10.1242/dev.096164 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Powell DR, Williams JS, Hernandez-Lagunas L, Salcedo E, O’Brien JH, Artinger KB, 2015. Cdon promotes neural crest migration by regulating N-cadherin localization. Dev. Biol 407, 289–299. 10.1016/j.ydbio.2015.07.025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prendergast A, Linbo TH, Swarts T, Ungos JM, McGraw HF, Krispin S, Weinstein BM, Raible DW, 2012. The metalloproteinase inhibitor Reck is essential for zebrafish DRG development. Development 139, 1141–1152. 10.1242/dev.072439 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raible DW, Eisen JS, 1994. Restriction of neural crest cell fate in the trunk of the embryonic zebrafish. Development 120, 495–503. [DOI] [PubMed] [Google Scholar]
- Raible DW, Wood A, Hodsdon W, Henion PD, Weston J. a, Eisen JS, 1992. Segregation and early dispersal of neural crest cells in the embryonic zebrafish. Dev. Dyn 195, 29–42. 10.1002/aja.1001950104 [DOI] [PubMed] [Google Scholar]
- Rawls JF, Mellgren EM, Johnson SL, 2001. How the zebrafish gets its stripes. Dev. Biol 240, 301–314. 10.1006/dbio.2001.0418 [DOI] [PubMed] [Google Scholar]
- Reifers F, Böhli H, Walsh EC, Crossley PH, Stainier DY, Brand M, 1998. Fgf8 is mutated in zebrafish acerebellar (ace) mutants and is required for maintenance of midbrain-hindbrain boundary development and somitogenesis. Development 125, 2381–95. [DOI] [PubMed] [Google Scholar]
- Reifers F, Walsh EC, Léger S, Stainier DY, Brand M, 2000. Induction and differentiation of the zebrafish heart requires fibroblast growth factor 8 (fgf8/acerebellar). Development 127, 225–35. [DOI] [PubMed] [Google Scholar]
- Richardson J, Gauert A, Briones Montecinos L, Fanlo L, Alhashem ZM, Assar R, Marti E, Kabla A, Härtel S, Linker C, 2016. Leader Cells Define Directionality of Trunk, but Not Cranial, Neural Crest Cell Migration. Cell Rep. 2076–2088. 10.1016/j.celrep.2016.04.067 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers CD, Jayasena CS, Nie S, Bronner ME, 2012. Neural crest specification: Tissues, signals, and transcription factors. Wiley Interdiscip. Rev. Dev. Biol 1, 52–68. 10.1002/wdev.8 [DOI] [PubMed] [Google Scholar]
- Rohrer H, Thoenen H, 2018. Relationship between differentiation and terminal mitosis: chick sensory and ciliary neurons differentiate after terminal mitosis of precursor cells, whereas sympathetic neurons continue to divide after differentiation. J. Neurosci 7, 3739–3748. 10.1523/jneurosci.07-11-03739.1987 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy S, Ng T, 2004. Blimp-1 specifies neural crest and sensory neuron progenitors in the zebrafish embryo. Curr. Biol 14, 1772–7. 10.1016/j.cub.2004.09.046 [DOI] [PubMed] [Google Scholar]
- Sadaghiani B, Vielkind JR, 1990. Distribution and migration pathways of HNK-1-immunoreactive neural crest cells in teleost fish embryos. Development 110, 197–209. [DOI] [PubMed] [Google Scholar]
- Sato M, Yost HJ, 2003. Cardiac neural crest contributes to cardiomyogenesis in zebrafish. Dev. Biol 257, 127–139. 10.1016/S0012-1606(03)00037-X [DOI] [PubMed] [Google Scholar]
- Sauka-Spengler T, Bronner-Fraser M, 2008. A gene regulatory network orchestrates neural crest formation. Nat. Rev. Mol. Cell Biol 9, 557–68. 10.1038/nrm2428 [DOI] [PubMed] [Google Scholar]
- Savagner P, 2010. The epithelial-mesenchymal transition (EMT) phenomenon. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol 21 Suppl 7, vii89–92. 10.1093/annonc/mdq292 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savagner P, 2001. Leaving the neighborhood: molecular mechanisms involved during epithelial-mesenchymal transition. Bioessays 23, 912–23. 10.1002/bies.1132 [DOI] [PubMed] [Google Scholar]
- Saxena A, Peng BN, Bronner ME, 2013. Sox10-dependent neural crest origin of olfactory microvillous neurons in zebrafish. Elife 2013, 1–17. 10.7554/eLife.00336 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scarpa E, Szabó A, Bibonne A, Theveneau E, Parsons M, Mayor R, 2015. Cadherin Switch during EMT in Neural Crest Cells Leads to Contact Inhibition of Locomotion via Repolarization of Forces. Dev. Cell 34, 421–34. 10.1016/j.devcel.2015.06.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schier AF, Neuhauss SC, Harvey M, Malicki J, Solnica-Krezel L, Stainier DY, Zwartkruis F, Abdelilah S, Stemple DL, Rangini Z, Yang H, Driever W, 1996. Mutations affecting the development of the embryonic zebrafish brain. Development 123, 165–78. [DOI] [PubMed] [Google Scholar]
- Schilling TF, Kimmel CB, 1994. Segment and cell type lineage restrictions during pharyngeal arch development in the zebrafish embryo. Development 120, 483–94. [DOI] [PubMed] [Google Scholar]
- Schlosser G, 2010. Making senses development of vertebrate cranial placodes. Int. Rev. Cell Mol. Biol 283, 129–234. 10.1016/S1937-6448(10)83004-7 [DOI] [PubMed] [Google Scholar]
- Schlosser G, 2008. Do vertebrate neural crest and cranial placodes have a common evolutionary origin? BioEssays 30, 659–672. 10.1002/bies.20775 [DOI] [PubMed] [Google Scholar]
- Schlosser G, 2006. Induction and specification of cranial placodes. Dev. Biol 294, 303–51. 10.1016/j.ydbio.2006.03.009 [DOI] [PubMed] [Google Scholar]
- Schmitz B, Papan C, Campos-Ortega JA, 1993. Neurulation in the anterior trunk region of the zebrafish Brachydanio rerio. Roux’s Arch. Dev. Biol 202, 250–259. 10.1007/BF00363214 [DOI] [PubMed] [Google Scholar]
- Schumacher JA, Hashiguchi M, Nguyen VH, Mullins MC, 2011. An intermediate level of bmp signaling directly specifies cranial neural crest progenitor cells in zebrafish. PLoS One 6 10.1371/journal.pone.0027403 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selleck M. a, Bronner-Fraser M, 1995. Origins of the avian neural crest: the role of neural plate-epidermal interactions. Development 121, 525–538. [DOI] [PubMed] [Google Scholar]
- Seo HC, Sætre BO, Håvik B, Ellingsen S, Fjose A, 1998. The zebrafish Pax3 and Pax7 homologues are highly conserved, encode multiple isoforms and show dynamic segment-like expression in the developing brain. Mech. Dev 70, 49–63. 10.1016/S0925-4773(97)00175-5 [DOI] [PubMed] [Google Scholar]
- Shepherd I, Eisen J, 2011. Development of the Zebrafish Enteric Nervous System, Third Edit. ed, Methods in Cell Biology. Elsevier Ltd; 10.1016/B978-0-12-387036-0.00006-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shepherd IT, Beattie CE, Raible DW, 2001. Functional analysis of zebrafish GDNF. Dev. Biol 231, 420–435. 10.1006/dbio.2000.0145 [DOI] [PubMed] [Google Scholar]
- Shepherd IT, Pietsch J, Elworthy S, Kelsh RN, Raible DW, 2004. Roles for GFRalpha1 receptors in zebrafish enteric nervous system development. Development 131, 241–9. 10.1242/dev.00912 [DOI] [PubMed] [Google Scholar]
- Simões-Costa M, Bronner ME, 2015. Establishing neural crest identity: a gene regulatory recipe. Development 142, 242–57. 10.1242/dev.105445 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith M, Hickman A, Amanze D, Lumsden A, Thorogood P, 1994. Trunk neural crest origin of caudal fin mesenchyme in the zebrafish Brachydanio rerio. Proc. R. Soc. B Biol. Sci 10.1098/rspb.1994.0061 [DOI] [Google Scholar]
- Soldatov R, Kaucka M, Kastriti ME, Petersen J, Chontorotzea T, Englmaier L, Akkuratova N, Yang Y, Häring M, Dyachuk V, Bock C, Farlik M, Piacentino ML, Boismoreau F, Hilscher MM, Yokota C, Qian X, Nilsson M, Bronner ME, Croci L, Hsiao W-Y, Guertin DA, Brunet J-F, Consalez GG, Ernfors P, Fried K, Kharchenko PV, Adameyko I, 2019. Spatiotemporal structure of cell fate decisions in murine neural crest. Science (80-.). 364, eaas9536 10.1126/science.aas9536 [DOI] [PubMed] [Google Scholar]
- Stewart RA, Arduini BL, Berghmans S, George RE, Kanki JP, Henion PD, Look AT, 2006. Zebrafish foxd3 is selectively required for neural crest specification, migration and survival. Dev. Biol 292, 174–88. 10.1016/j.ydbio.2005.12.035 [DOI] [PubMed] [Google Scholar]
- Swartz ME, Sheehan-Rooney K, Dixon MJ, Eberhart JK, 2011. Examination of a palatogenic gene program in zebrafish. Dev. Dyn 240, 2204–2220. 10.1002/dvdy.22713 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taneyhill LA, Padmanabhan R, 2014. The Cell Biology of Neural Crest Cell Delamination and EMT. Neural Crest Cells Evol. Dev. Dis 51–72. 10.1016/B978-0-12-401730-6.00003-X [DOI] [Google Scholar]
- Taneyhill LA, Schiffmacher AT, 2017. Should I stay or should I go? Cadherin function and regulation in the neural crest. Genesis 55, 1–21. 10.1002/dvg.23028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tawk M, Araya C, Lyons DA, Reugels AM, Girdler GC, Bayley PR, Hyde DR, Tada M, Clarke JDW, 2007. A mirror-symmetric cell division that orchestrates neuroepithelial morphogenesis. Nature 446, 797–800. 10.1038/nature05722 [DOI] [PubMed] [Google Scholar]
- Taylor CR, Montagne WA, Eisen JS, Ganz J, 2016. Molecular fingerprinting delineates progenitor populations in the developing zebrafish enteric nervous system. Dev. Dyn 245, 1081–1096. 10.1002/dvdy.24438 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theveneau E, Marchant L, Kuriyama S, Gull M, Moepps B, Parsons M, Mayor R, 2010. Collective Chemotaxis Requires Contact-Dependent Cell Polarity. Dev. Cell 19, 39–53. 10.1016/j.devcel.2010.06.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theveneau E, Mayor R, 2012a. Neural crest delamination and migration: From epithelium-to-mesenchyme transition to collective cell migration. Dev. Biol 10.1016/j.ydbio.2011.12.041 [DOI] [PubMed] [Google Scholar]
- Theveneau E, Mayor R, 2012b. Neural crest migration: Interplay between chemorepellents, chemoattractants, contact inhibition, epithelial-mesenchymal transition, and collective cell migration. Wiley Interdiscip. Rev. Dev. Biol 1, 435–445. 10.1002/wdev.28 [DOI] [PubMed] [Google Scholar]
- Theveneau E, Steventon B, Scarpa E, Garcia S, Trepat X, Streit A, Mayor R, 2013. Chase-and-run between adjacent cell populations promotes directional collective migration. Nat. Cell Biol 15, 763–72. 10.1038/ncb2772 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thiery JP, Sleeman JP, 2006. Complex networks orchestrate epithelial-mesenchymal transitions. Nat. Rev. Mol. Cell Biol 7, 131–42. 10.1038/nrm1835 [DOI] [PubMed] [Google Scholar]
- Thisse C, Thisse B, Postlethwait JH, 1995. Expression of snail2, a second member of the zebrafish snail family, in cephalic mesendoderm and presumptive neural crest of wild-type and spadetail mutant embryos. Dev. Biol 172, 86–99. 10.1006/dbio.1995.0007 [DOI] [PubMed] [Google Scholar]
- Thompson EW, Williams ED, 2008. EMT and MET in carcinoma--clinical observations, regulatory pathways and new models. Clin. Exp. Metastasis 25, 591–2. 10.1007/s10585-008-9189-8 [DOI] [PubMed] [Google Scholar]
- Trainor PA, 2016. Developmental Biology: We Are All Walking Mutants, 1st ed, Current Topics in Developmental Biology. Elsevier Inc. 10.1016/bs.ctdb.2015.11.029 [DOI] [PubMed] [Google Scholar]
- Tryon RC, Higdon CW, Johnson SL, 2011. Lineage relationship of direct-developing melanocytes and melanocyte stem cells in the zebrafish. PLoS One 6 10.1371/journal.pone.0021010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tuttle AM, Hoffman TL, Schilling TF, 2014. Rabconnectin-3a Regulates Vesicle Endocytosis and Canonical Wnt Signaling in Zebrafish Neural Crest Migration. PLoS Biol. 12 10.1371/journal.pbio.1001852 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uribe RA, Bronner ME, 2015. Meis3 is required for neural crest invasion of the gut during zebrafish enteric nervous system development. Mol. Biol. Cell 26, 3728–3740. 10.1091/mbc.E15-02-0112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uribe RA, Hong SS, Bronner ME, 2018. Retinoic acid temporally orchestrates colonization of the gut by vagal neural crest cells. Dev. Biol 433, 17–32. 10.1016/j.ydbio.2017.10.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Otterloo E, Williams T, Artinger KB, 2016. The old and new face of craniofacial research: How animal models inform human craniofacial genetic and clinical data. Dev. Biol 415, 171–187. 10.1016/j.ydbio.2016.01.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volkening A, Sandstede B, 2018. Iridophores as a source of robustness in zebrafish stripes and variability in Danio patterns. Nat. Commun 9, 1–14. 10.1038/s41467-018-05629-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wada N, Javidan Y, Nelson S, Carney TJ, Kelsh RN, Schilling TF, 2005. Hedgehog signaling is required for cranial neural crest morphogenesis and chondrogenesis at the midline in the zebrafish skull. Development 132, 3977–88. 10.1242/dev.01943 [DOI] [PubMed] [Google Scholar]
- Walshe J, Mason I, 2003. Fgf signalling is required for formation of cartilage in the head. Dev. Biol 264, 522–536. 10.1016/j.ydbio.2003.08.010 [DOI] [PubMed] [Google Scholar]
- Wang W Der, Melville DB, Montero-Balaguer M, Hatzopoulos AK, Knapik EW, 2011. Tfap2a and Foxd3 regulate early steps in the development of the neural crest progenitor population. Dev. Biol 360, 173–185. 10.1016/j.ydbio.2011.09.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weston JA, Thiery JP, 2015. Pentimento: Neural Crest and the origin of mesectoderm. Dev. Biol 401, 37–61. 10.1016/j.ydbio.2014.12.035 [DOI] [PubMed] [Google Scholar]
- Whitlock KE, Smith KM, Kim H, Harden M V, 2005. A role for foxd3 and sox10 in the differentiation of gonadotropin-releasing hormone (GnRH) cells in the zebrafish Danio rerio. Development 132, 5491–502. 10.1242/dev.02158 [DOI] [PubMed] [Google Scholar]
- Whitlock KE, Wolf CD, Boyce ML, 2003. Gonadotropin-releasing hormone (GnRH) cells arise from cranial neural crest and adenohypophyseal regions of the neural plate in the zebrafish, Danio rerio. Dev. Biol 257, 140–152. 10.1016/S0012-1606(03)00039-3 [DOI] [PubMed] [Google Scholar]
- Williams JS, Hsu JY, Rossi CC, Artinger KB, 2018. Requirement of zebrafish pcdh10a and pcdh10b in melanocyte precursor migration. Dev. Biol 10.1016/j.ydbio.2018.03.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan Y-L, Willoughby J, Liu D, Crump JG, Wilson C, Miller CT, Singer A, Kimmel C, Westerfield M, Postlethwait JH, 2005. A pair of Sox: distinct and overlapping functions of zebrafish sox9 co-orthologs in craniofacial and pectoral fin development. Development 132, 1069–1083. 10.1242/dev.01674 [DOI] [PubMed] [Google Scholar]
- Yan YL, Miller CT, Nissen RM, Singer A, Liu D, Kirn A, Draper B, Willoughby J, Morcos PA, Amsterdam A, Chung BC, Westerfield M, Haffter P, Hopkins N, Kimmel C, Postlethwait JH, 2002. A zebrafish sox9 gene required for cartilage morphogenesis. Development 129, 5065–5079. [DOI] [PubMed] [Google Scholar]
- Yu HH, Moens CB, 2005. Semaphorin signaling guides cranial neural crest cell migration in zebrafish. Dev. Biol 280, 373–385. 10.1016/j.ydbio.2005.01.029 [DOI] [PubMed] [Google Scholar]
- Zuniga E, Rippen M, Alexander C, Schilling TF, Crump JG, 2011. Gremlin 2 regulates distinct roles of BMP and Endothelin 1 signaling in dorsoventral patterning of the facial skeleton. Development 138, 5147–5156. 10.1242/dev.067785 [DOI] [PMC free article] [PubMed] [Google Scholar]