

Abstract
Acute reactivity of the stress hormone cortisol is reflective of early adversity and stress exposure, with some studies finding that the impact of adversity on the stress response differs by race. The objectives of the current study were to characterize cortisol reactivity to two dyadically based stress paradigms across the first year of life, to examine cortisol reactivity within Black and White infants, and to assess the impact of correlates of racial inequity including socioeconomic status, experiences of discrimination, and urban life stressors, as well as the buffering by racial socialization on cortisol patterns. Salivary cortisol reactivity was assessed at 4 months of age during the Still Face paradigm (N = 207) and at 12 months of age across the Strange Situation procedure (N = 129). Infants demonstrated the steepest recovery after the Still Face paradigm and steepest reactivity to the Strange Situation procedure. Race differences in cortisol were not present at 4 months but emerged at 12 months of age, with Black infants having higher cortisol. Experiences of discrimination contributed to cortisol differences within Black infants, suggesting that racial discrimination is already “under the skin” by 1 year of age. These findings suggest that race-related differences in hypothalamic-pituitary-adrenal reactivity are present in infancy, and that the first year of life is a crucial time period during which interventions and prevention efforts for maternal-infant dyads are most likely able to shape hypothalamic-pituitary-adrenal reactivity thereby mitigating health disparities early across the life course.
Within the United States, Black individuals are more likely than non-Hispanic Whites to experience many leading causes of death including cardiovascular disease, diabetes, obesity, and stroke (Hasson, Apovian, & Istfan, 2015; Kurian & Cardarelli, 2007; Wong, Shapiro, Boscardin, & Ettner, 2002). According to the CDC, as recently as 2011, the average life span for Black individuals was 4.5 years less than that of non-Hispanic Whites despite increasing attention to health disparities (Barr, 2014; Case & Deaton, 2015; Kochanek, Murphy, & Xu, 2015). This increase in morbidity and mortality risk in Black individuals likely arises early in development with significant research implicating alterations in the stress-response systems as part of the underlying biological mechanism. To date, few studies have examined race differences in biological pathways, such as the hypothalamic-pituitary-adrenal (HPA) axis, during these early critical developmental windows (Ross & Desai, 2012).
Correlates of Racial Inequity May Help Explain Physiological Roots of Health Disparities
Exploring the earliest development of race differences in HPA function is expected to provide data to support that alterations to stress physiology are one biological pathway underlying racial health disparities. Linking this stress biomarker with measures that capture racism may further elucidate how health disparities emerge. Racism is a stressor disproportionately experienced by minority populations (Clark, Anderson, Clark, & Williams, 1999). While racism comes in many forms, for the purpose of this paper, we endorse the vantage point that racism constitutes, “beliefs, attitudes, institutional arrangements, and acts that tend to denigrate individuals or groups because of phenotypic characteristics or ethnic group affiliation” (Black, Johnson, & VanHoose, 2015; Clark et al., 1999; Jones, 2000). Central to this conceptualization is the allocation of resources, given that a core process of racism is the categorization of populations and the use of this categorization to disproportionally allocate societal goods and resources to groups regarded as superior (Williams & Mohammed, 2009). As such, racism constitutes an agglomerated web of processes that culminate in a group of individuals experiencing racial inequities. These racial inequities have physical and mental health consequences across the life course (Clark et al., 1999; Krieger, 1990; Merritt & Harrell, 1998; Pascoe & Smart Richman, 2009; Williams, Neighbors, & Jackson, 2003). We focus on the core inequities of racial discrimination, socioeconomic status (SES), and urban life stress exposure in this investigation as they capture objective and perceived racial inequities disproportionately experienced by minority populations. To avoid a deficit model approach, we also investigate maternal racial socialization as a potential buffer (Valencia, 2010).
Discrimination is an integral component of racism that is foundational to the denigration of minority populations and a contributor to variance in adult and child health outcomes (Williams et al., 2003). Racial discrimination can be viewed as a cultural process resulting in the enactment of a cohesive network of values, symbols, and practices that are learned and transmitted across generations and serve the purpose of systematically excluding minority populations (Causadias, 2013). It is difficult to quantify racial discrimination as discrimination can vary over time and include single and/or chronic exposures. In addition, discrimination can vary on a spectrum from traumatic exposures to systemic and structural racism to microaggressions (Williams et al., 2003). Kressin, Raymond, and Manze (2008) reviewed 34 measures of discrimination and concluded that few measures were theoretically based and most assessed only general dimensions of racism. The impact of racial discrimination on health can be conceptualized according to a stress and coping model. Stressors high in unpredictability and uncontrollability are potent predictors of health consequences, and these are also core components of racial discrimination (Williams & Mohammed, 2009). Effects of racial discrimination can be both cumulative and developmentally moderated such that exposure to discrimination during sensitive periods in development are the most likely to result in long-term physiological consequences (Doane, Sladek, & Adam, 2018). Thus, physical health outcomes associated with discrimination may be rooted in the role of discrimination as a potent stressor (Clark et al., 1999) impacting biological systems and leading to negative health and physiologic outcomes including alterations in cortisol and increased blood pressure (see Pascoe & Smart Richman, 2009, for a review).
Beyond the effect of discrimination, SES can impact the developing child. SES can interact with racism to impact health disparities across the life span (Schoenbaum & Waidmann, 1997). SES refers to “a multidimensional construct comprising diverse socioeconomic factors (typically, economic resources, power, and/or prestige)” (Braveman et al., 2005). Like discrimination, this construct is inherently interactive, tied as it is to power, social hierarchy, and access to resources. There are multiple pathways by which SES may impact health, such as the link between SES and geographic segregation and isolation (Iceland & Wilkes, 2006) or the link between SES and stress pathways (Needham et al., 2015). These SES pathways are contingent on other variables (e.g., discrimination) and are not limited to individual accumulated wealth but can operate at or across the level of the individual, the household, or the neighborhood. While there is variance in SES among all populations, Black individuals disproportionately experience low SES in the United States. According to 2016 data from the United States Federal Reserve Survey on Consumer Finances, Black families’ median and mean net worth is less than 15% that of White families, at $17,600 and $138,200, respectively (Federal Reserve, 2018). SES accounts for some of the observed racial disparities in health, though disparities by race often persist even among individuals with equivalent levels of SES (Williams, 1999). This makes sense given that race differences in SES capture relevant but limited aspects of the broader domain of racism (Williams, Mohammed, Leavell, & Collins, 2010). In sum, race/ethnicity and SES are intrinsically linked in the United States and the intersectionality of race and SES has a large impact on health (Williams, Priest, & Anderson, 2016).
Given the location of the parent study in a large urban environment, we also examined the impact of urban stressors. Urban stressors refer to subjective contextual community-level stressors as potential sources of psychological and emotional stress experienced by persons living in medium to large cities (Jaffee et al., 2005). Like SES, stressor exposure is not limited to minority populations. Nonetheless, these types of stressors are disproportionately experienced by urban-dwelling Black individuals (Goldmann et al., 2011; Norris, 1992; Williams et al., 2003) in part because this type of environmental context captures systematic neighborhood disadvantage. Thus, urban stressors likely impact health and behavior as a facet of stress exposure (Attar, Guerra, & Tolan, 1994).
Racial socialization, the process by which parents address group disadvantage and the meaning of race and ethnicity with their children, is proposed to buffer the impact of racial inequity on health outcomes (Fischer & Shaw, 1999), especially mental health (Hughes et al., 2006). In this context, racial socialization represents a resilience factor (Brown, 2008; Miller, 1999; Miller & MacIntosh, 1999) with the potential to mitigate the impact of racial inequalities and discrimination. It is important to note that racial socialization, in this investigation, is a three-generation process. Racial socialization is measured in parents of infant off-spring, and thus represents the amount of socialization grandparents engaged in with parents. Racial socialization may buffer infant stress response by increasing cultural pride and well-being in new parents, although it is also possible that greater exposure to racial socialization may indirectly index greater cumulative race-related stress, given that racial socialization and exposure to racism can be correlated (Stevenson & Arrington, 2009). That is, there may be a greater cultural need for racial socialization within contexts characterized by high levels of discrimination, so higher scores may reflect increased stress exposures.
Collectively, these experiences of racial discrimination, low SES, and urban life stressors contribute to the pervasive and recalcitrant racial disparities in health found in the United States. Increased racial socialization may buffer some of these effects. Each of these components of racial inequity can be thought of as operating, in part, through a stress framework. This impact may not be direct given that inequities often co-occur and the effects of SES, discrimination, and community-level events such as urban life stressors are intertwined. Nonetheless, because each of these components of racial inequity can be framed within a stress exposure model, it makes sense to examine them in conjunction with biological stress pathways.
Cortisol and Race
The HPA axis is a primary stress-response system that helps individuals mobilize resources in response to challenge and adapt to their environment and social context. The HPA axis begins with production of corticotropin-releasing hormone in the hypothalamus and ends with production of cortisol from the adrenal glands. Cortisol has been traditionally measured using diurnal rhythms (the change in cortisol concentration across the day), baseline cortisol (waking values), awakening response (the rapid increase in cortisol immediately after waking), as well as reactivity and recovery (the short-term rise and fall in cortisol through a proximal stressor). A recent meta-analysis by Adam et al. (2017) demonstrated a relationship between diurnal cortisol slope and a wide range of mental and physical health problems with the general pattern for flatter slopes to predict poorer mental and physical health. Cortisol reactivity and recovery to acute stress are also linked to physical and mental health, with flattened recovery linked to depression (Burke, Davis, Otte, & Mohr, 2005) and reactivity linked to depression and behavior problems (Susman, Dorn, Inoff-Germain, Nottel-mann, & Chrousos, 1997), health and weight (Epel, Lapidus, McEwen, & Brownell, 2001; Epel et al., 2000; Gluck, Geliebter, & Lorence, 2004), and heart disease (Hamer, O’Donnell, Lahiri, & Steptoe, 2010). These findings are not ubiquitous given that flattened cortisol reactivity has also been associated with negative physical and mental health outcomes (Phillips, Ginty, & Hughes, 2013).
Alterations in HPA regulation manifest across the life span in individuals exposed to early life stressors associated with racial inequity (Lupien, King, Meaney, & McEwen, 2001; Maniam, Antoniadis, & Morris, 2014; Miller, Chen, & Zhou, 2007). An extant literature demonstrates racial differences in cortisol levels in adolescents and adults. Korous, Causadias, and Casper (2017) suggest that the relation between racial discrimination and cortisol output is nonlinear. Their meta-analysis of participants aged 15–55 reported pronounced acute cortisol reactivity in response to racial discrimination, and four of six reported studies demonstrated flattened cortisol diurnal rhythm in response to discrimination. Lower basal cortisol, diminished cortisol reactivity (Hostinar, McQuillan, Mirous, Grant, & Adam, 2014), and flattened cortisol diurnal decline across the day (Cohen et al., 2006; DeSantis et al., 2007) have been observed in Black youth relative to White youth as general patterns. These race differences persist even when examining correlates of racial inequity such as SES, life stress, and adversity (Skinner, Shirtcliff, Haggerty, Coe, & Catalano, 2011), suggesting many pathways by which the impact of racial inequity can manifest in altered HPA axis regulation. Altered HPA axis regulation has implications for racial disparities in health across the life span. These alterations are associated with negative health outcomes including cardiovascular disease, diabetes, obesity, and stroke, which also are more prominent in Black compared to White individuals (Broderick et al., 1998; Hasson et al., 2015; Kurian & Cardarelli, 2007; Rosmond & Björntorp, 2000; Walker, 2007; Wong et al., 2002). This vantage point culminates in a traditional formulation of a stress model by which racism constitutes a source of aversive experiences that eventually lead to poor health outcomes (Harrell et al., 2011). Each component of this framework is potentially a source of targeted, developmentally informed intervention, yet the cumulative factors that contribute to race differences in cortisol levels, as well as the earliest age when measurable differences arise, remains ambiguous.
Given the existence of race differences in the HPA axis later in development (Cohen et al., 2006; DeSantis et al., 2007; Hostinar et al., 2014; Skinner et al., 2011) and the known plasticity of the HPA axis during the first years of life, it is critical to investigate whether race-related differences in HPA axis reactivity are emerging in infancy. To our knowledge, only one previous study has directly tested race differences in infant stress responses. In this study, Japanese infants displayed increased cortisol reactivity relative to North American White participants (Lewis, Ramsay, & Kawakami, 1993). The detection of race differences within the first year of life has significant implications for the timing and targeting of interventions.
The Present Study
Cortisol reactivity may vary based on the type of stressor. Infants less than 24 months of age have primarily been studied using pain induction (e.g., vaccinations), discomfort paradigms (e.g., arm restraint), or novelty exposure (e.g., robot task). However, the HPA axis is socially responsive and the parent-infant relationship defines the infant social world (Gunnar & Donzella, 2002). Therefore, the targeted use of age-appropriate dyadic stressors may provide substantive information about the developmental patterns of HPA axis reactivity during the first year of life (Bowlby, 2008; Tarullo & Gunnar, 2006). The Face to Face interaction procedure, commonly referred to as the Still Face paradigm (Tronick, Als, Adamson, Wise, & Brazelton, 1978), induces a stress response by having a caregiver look at her infant for a short time without responding to the infant’s social cues. In older infants, the Strange Situation procedure (Ainsworth, Blehar, Waters, & Wall, 2015) uses separation-reunion episodes to characterize the organization of the infant’s attachment behaviors toward the caregiver. In both the Still Face paradigm and the Strange Situation procedure, the mother’s behavior deviates from the infant’s expectation. The mother engages visually with her child in the Still Face paradigm, but without expected responsiveness, leading to physiological stress induction (Haley & Stansbury, 2003). The induction of a physiologic stress response in the Strange Situation procedure occurs when the mother separates from her infant in an unfamiliar setting (Gunnar, Brodersen, Nachmias, Buss, & Rigatuso, 1996; Hertsgaard, Gunnar, Erickson, & Nachmias, 1995). In both paradigms, maternal behavior and sensitivity, as well as infant temperament, are thought to contribute to the behavioral and physiological responses of the infant.
Another challenge in the extant literature on infant development of the HPA axis is that the majority of HPA axis reactivity data are cross-sectional, precluding the ability to assess developmental changes (Gunnar, Talge, & Herrera, 2009; Jansen, Beijers, Riksen-Walraven, & de Weerth, 2010). Gunnar, Brodersen, Kreuger, et al. (1996) found an overall dampening of cortisol reactivity from birth into the second year of life, suggesting that as infants age, the HPA axis becomes less reactive to inoculation stress. In a more recent study examining dyadic stressors, infants did not show reactivity at 7 months to the Still Face procedure but did show reactivity to the Strange Situation procedure at 16 months of age (Martinez-Torteya et al., 2015). These differential longitudinal patterns between painful and dyadic stressors underscore the need for studies that carefully consider the type and the timing of the stressor. Measuring longitudinal changes in the HPA axis is vital for understanding how infants biologically integrate the caregiving relationship and other early environmental cues, particularly during the first years of life when both the attachment relationship and the HPA axis are simultaneously developing.
A third challenge is that when looking at longitudinal stress-related physiological studies in infants, race differences remain largely unexplored. As described above, race differences in cortisol and health outcomes related to stress are robust when comparing White and Black adults (Brondolo, 2015; Chong, Uhart, McCaul, Johnson, & Wand, 2008; Hajat et al., 2010; Wagner, Lampert, Tennen, & Feinn, 2015), young adults (Cohen et al., 2006; Skinner et al., 2011), and adolescents (DeSantis et al., 2007; Hostinar et al., 2014), but it is not yet known just how early race differences in HPA function can be detected and whether those early differences are explained by racial inequities. Because it is critical to move beyond contrasting race (Sen & Wasow, 2016), the impact of indices of racial inequity are considered within Black infants. This avoids the traps of a deficit approach in the consideration of race-related inequities and focuses the question on magnitude of impact within black participants.
To address these three challenges, the current study examined cortisol reactivity longitudinally at 4 and 12 months of age using two developmentally sensitive dyadic stress paradigms: the Still Face paradigm and the Strange Situation paradigm. We examined the following research questions:
Research Question 1: What is the effect of the Still Face paradigm and the Strange Situation procedure on cortisol output?
Research Question 2: Are there race differences in cortisol levels, reactivity, and recovery at the Still Face paradigm and the Strange Situation procedure?
Research Question 3: Do markers of racial inequity, including SES, experiences of discrimination, urban life stressors, and the buffer racial socialization, moderate racial differences in cortisol?
Research Question 4: What is the impact of racial inequity measures and cortisol trajectory within black infants?
Method
Participants
Participants included dyads recruited as part of an ongoing longitudinal cohort study in New Orleans, Louisiana, designed to examine the effect of cumulative maternal life course and prenatal stress on infant and child development. The longitudinal study sought to examine the impact of maternal life course adversity on infant stress responsivity and how the mother-infant relationship influences the development of infant stress-response systems. To achieve this goal, the study conducted age-appropriate dyadic stress paradigms at 4 months (i.e., Still Face paradigm) and at 12 months of age (i.e., Strange Situation procedure; Gray, Jones, Theall, Glackin, & Drury, 2017; Martinez-Torteya et al., 2015). A convenience sample of pregnant mothers, age 18 to 41, was recruited from prenatal and Women, Infant and Children clinics, as well as from other ongoing studies of maternal health and pregnancy outcomes at Tulane University. Mothers were excluded if they were less than 18 years of age. Data on 233 dyads were available for analysis. Of these, 144 (62%) infants were Black and 111 (48%) were female. Two hundred and seven individuals had data at 4 months and 129 individuals had data at 12 months, and 109 had full cortisol at both visits. The Tulane University Institutional Review Board approved this study.
Cortisol
Cortisol was obtained contemporaneously with the Still Face paradigm (age 4 months) and the Strange Situation procedure (12 months) at the lab. Sample collection occurred 15 min after arrival at site, 15 min after peak stress, and 45 min after peak stress for each task to track reactivity. Saliva was collected via Salimetrics SalivaBio Children’s Swab (Rodriguez et al., 2016). For both stressors, cortisol peak collection was timed based on the expected greatest dyadic stress point: the beginning of the Still Face paradigm episode in which the mother looks at her infant with a neutral expression for the Still Face paradigm and the second separation period for the Strange Situation procedure. Saliva was centrifuged out of the collection swab and stored immediately in Wheaton Cryovials at −80 °C until assayed. Assays were performed on samples in duplicate using commercially available, validated enzyme-immunoassays (www.salimetrics.com) kits. Samples with intra-assay coefficients of variation greater than 7% were reassayed. Plates with unreliable performance (e.g., standard curves with R2 less than .996 or control samples that vary by 15% from standardized controls) were reassayed. Cortisol levels across the Still Face paradigm and Strange Situation procedure are available in Table 1.
Table 1.
Average cortisol by time point
| Condition | Collection | Count | Cortisol mean (SD) |
|---|---|---|---|
| SF | 1 | 207 | 3.82 (0.95) |
| 2 | 207 | 3.93 (0.90) | |
| 3 | 204 | 3.72 (0.81) | |
| SSP | 1 | 129 | 3.52 (0.76) |
| 2 | 128 | 3.84 (0.85) | |
| 3 | 122 | 3.86 (0.83) |
Note: SF, Still Face procedure. SSP, Strange Situation procedure. Values are log transformed and windzorised and reported as mean (SD).
The Still Face paradigm
The Still Face paradigm is a validated interactive social stressor that has been used to evaluate an infant’s stress response (Haley & Stansbury, 2003; Lewis & Ramsay, 2005) and regulatory capacities in the context of the maternal relationship (Tronick et al., 1978). A trained research assistant conducted the Still Face paradigm. The following standard procedure was used with three distinct epochs: talking and interacting (2 min), maternal neutral face and no interaction (2 min), and resumption of talking and interaction (2 min). Twenty seconds of sustained distress resulted in the termination of the epoch. Infants were seated in an age-appropriate chair or car seat for the procedure, facing the mother at eye level. A research assistant was present but hidden from the infant throughout the procedure. To systematically examine infant stress response to the Still Face paradigm across all infants, the peak cortisol sample was collected 15 min after the start of the second epoch where the mother maintains a neutral expression. Peak cortisol was not estimated from infant affective distress real time due to the potential incongruency of affective distress and HPA axis activation. The mother was given careful instructions not to have physical contact with the infant during the entire procedure but to “play” with her child as she would normally during the first and third epochs.
The Strange Situation procedure
Ainsworth’s Strange Situation procedure is an extensively studied laboratory assessment using a separation-reunion paradigm to classify the extent to which the child uses the parent as a secure base for exploration (Ainsworth et al., 2015). Briefly, the Strange Situation procedure is a structured series of events that include interactions with the mother alone, involvement of a stranger, separation from the mother, and a reunion with the mother. The Strange Situation procedure is approximately 21 min in duration and consists of eight episodes. There are two separation episodes of the Strange Situation procedure. In the second separation, the child is left alone in a room, and this is considered the peak stressor event of the paradigm. The Strange Situation procedure has been found to elicit HPA axis activation in infants though developmental differences and differences by attachment classification occur (Gunnar et al., 2009; Gunnar, Brodersen, Krueger, et al., 1996; Van Bakel & Riksen-Walraven, 2004). The Strange Situation procedure was conducted by trained research assistants.
Measures
Mothers provided information about multiple levels of their health, behavior, and environment using an approximately 90-min interview-assisted computer survey completed by all participants, administered face-to-face at the research site or at prenatal clinics (Questionnaire Development System, QDS, Nova Research, Bethesda, MD) prior to delivery, and additional information was obtained at each subsequent study visit. Race-related inequity variables were utilized from the prenatal survey. Oral responses were recorded onto the computer by trained interviewers. Mean differences in covariates and control variables by race are available in Table 2.
Table 2.
Covariate means and standard deviations within Black participants
| EOD | RS | SES | ULSS | Age |
|---|---|---|---|---|
| 1.79 (2.18) | 5.45 (4.29) | 2.10 (1.71) | 38.18 (13.42) | 38.63 (1.99) |
Note: EOD, experiences of discrimination. RS, racial socialization. SES, socioeconomic status. ULSS, urban life stressors as measured by the Urban Life Stressors Scale. Age represents gestational age of the child.
SES.
Maternal report was used to measure SES, which reflected the family/household ecological level. A summary score of SES, generated in our laboratory, was indexed from education, employment, home ownership, income, savings, and government assistance status (range: 0–6). Education was scored “0” for less than high school and “1” for high school or greater. Employment was scored “0” for less than full time and “1” for full time. Home ownership was scored “0” for renting and “1” for ownership. Income was scored “0” for less than $24,999 annual income and “1” for greater than $25,000. Savings was scored “0” for less than $500 and “1” for $500 or greater. Government assistance was scored “0” for assistance and “1” for no assistance. A score of 6 represents the highest tier of SES.
Discrimination.
The Experiences of Discrimination Scale was used to assess discrimination. This reliable and valid instrument (Krieger, Smith, Naishadham, Hartman, & Bar-beau, 2005) captures the degree to which mothers, in their lifetime, have been made to feel inferior, hassled, discriminated against, or prevented from doing something they wanted to do, or called derogatory terms based on race or gender. The Cronbach’s a coefficient in this sample was 0.86. This survey is especially relevant as it was designed to predict health outcomes. At the prenatal assessment, mothers reported whether or not they worried about racial discrimination (1) as a child and (2) in the past year, (3) personally and (3) for their racial/ethnic group. Only race-related discrimination was utilized in this study. The total worry index ranged from 0 (did not endorsed worrying about discrimination) to 4 (endorsed worrying about discrimination on all four items).
Racial socialization.
The measure developed by Hughes and Chen (1997), specifically three items from the preparation for bias subscale, was used to assess racial socialization. These items asked whether the mother’s own parent(s) talked to her about racism, the fight for equality, or told her that people may try to limit her because of her race (never = 0 to very often = 4). Items were summed to create an overall socialization scale that ranged from 0 to 12.
Urban life stressors.
The Urban Life Stressor Scale (ULSS; Jaffee et al., 2005) was used to capture urban life stressors. The ULSS is a 21-item instrument that is used to measure subjective contextual community-level stressors as potential sources of psychological and emotional stress experienced by persons living in medium to large cities. Each item on the ULSS is answered with reference to a 5-point scale ranging from 1 (no stress at all) to 5 (extremely stressful—more than I can handle). The total score reflects the level of stress associated with day-to-day life in an urban environment. The scale is intended to identify chronic life stress as opposed to acute event stress. The scale’s reliability and validity have been established in African American and Latino groups (Sanders-Phillips, 1996), and Cronbach’s a in our sample was 0.87.
Gestational age.
The difference between date of last menstrual cycle and date of delivery, in days and obtained from medical records or maternal report, was used as a measure of gestational age.
Analytic strategy
Cortisol samples were nested within individuals, which were nested within Still Face paradigm and Strange Situation procedure visits. To account for the inherent correlation within individuals within time points, data were analyzed using Multilevel Mixed Effects Models in R (Bates, Maechler, & Bolker, 2012; Finch, Bolin, & Kelley, 2014; R Core Team, 2013; Pinheiro & Bates, 2000). A base model of linear and quadratic time predicting cortisol was established to index the change in cortisol from baseline within individuals. Intercepts and slopes varied across all models. In this framework, cortisol as the outcome of interest was predicted by time across the stressor and the intercept represents the putative baseline cortisol level for that individual:
This model predicts the within-individual growth curve of cortisol over the tasks. Between-individual and between-time point differences in within-individual trajectory were assessed using higher order nesting of the model (e.g., between-individual effects of race can load on a second level):
and these effects can vary by stress time point:
This allows for an overall model that can answer global questions about the impact of between-individual differences such as race in the entire sample. Next, each time point was examined independently as a two-level model to further decompose race effects. Known correlates of racial inequity such as SES, experiences of discrimination, racial socialization, and urban life stressors were added as main effects to these models to examine the impact they have on race. Gestational age and sex are covaried for in all models, and continuous variables were centered. Because inequities related to minority race are not theoretically predicted to manifest in majority White individuals and measures are not known to be valid and reliable in White individuals, investigation within race was limited to Black infants (Sen & Wasow, 2016).
Results
Overall model of cortisol across both the Still Face paradigm and the Strange Situation procedure
In an overall model of cortisol nested within individuals nested within visits, cortisol had an average log transformed, winsorized baseline value of 3.65. As time increased, cortisol increased across the stressors, β = 0.26, 95% confidence interval (CI) [0.10, 0.43], p < .001, until curving back toward baseline, β = −0.16, 95% CI [−0.46, −0.09], p < .001, in recovery. When cortisol was allowed to vary by visit and race, there was a Race × Visit interaction, which suggested that overall race effects differ by visit, β = −0.44, 95% CI [−0.83, −0.06], p = .025 (see Figure 1). This Race × Visit interaction is therefore decomposed below.
Figure 1.
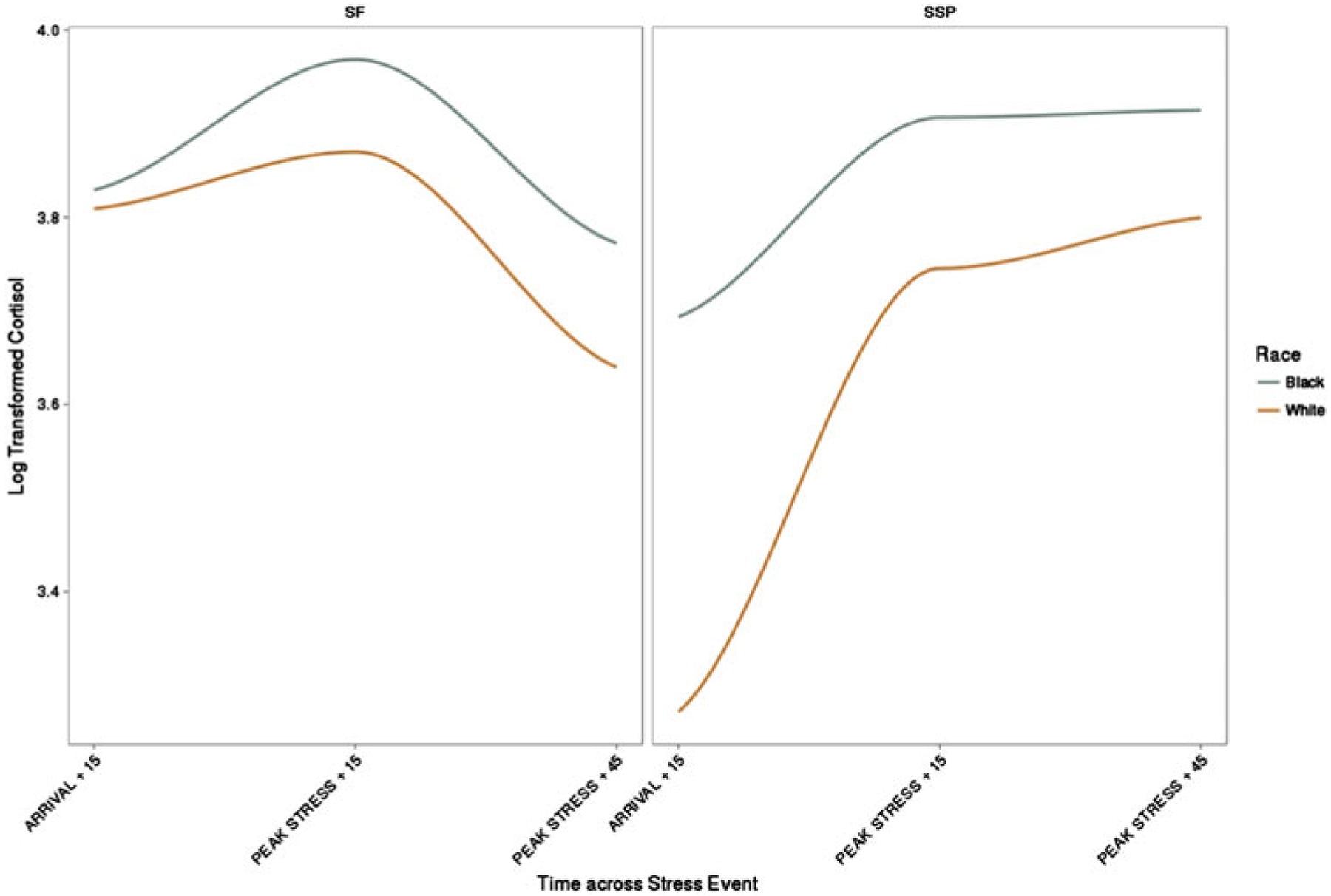
Cortisol differences by race across the 4-month Still Face procedure and the 12-month Strange Situation procedure.
Cortisol across the Still Face paradigm
At the Still Face paradigm, cortisol increased from baseline, β = 0.26, 95% CI [0.08, 0.44], p = .004, peaked, and curved downward in recovery, β = −0.16, 95% CI [−0.24, −0.08], p = .004. There was no effect of race on overall cortisol levels at this visit, β = −0.07, 95% CI [−0.29, 0.15], p = .004. Furthermore, race did not significantly moderate the trajectory of cortisol across the Still Face paradigm, β = −0.09, 95% [CI −0.45, 0.27], p = .63. The covariates infant sex and gestational age were not significant in this model.
Cortisol across the Strange Situation procedure
At the Strange Situation procedure visit, cortisol increased from baseline, β = 0.43, 95% CI [0.24, 0.62], p < .001, but this responsivity was qualified by a quadratic effect, β = −0.12, 95% CI [−.21, −0.03], p =.008; Figure 1 shows that cortisol levels rose and then remained flat across the Strange Situation procedure. Race exhibited a main effect such that White infants had lower overall cortisol levels, β = −0.362, 95% CI [−0.62, −0.10], p = .008. When race was allowed to moderate change in cortisol across time, race continued to predict lower peak cortisol for White infants, β = −0.45, 95% CI [−0.73, −0.17], p = .002. Neither sex nor gestational age exhibited main effects on cortisol responsivity in this or any subsequent models described below. Table 3 demonstrates overall race and race by time interactions for the SSP procedure.
Table 3.
Coefficients of racial inequity at the Strange Situation procedure visit
| Predictor | Base | SES | EOD | RS | ULSS |
|---|---|---|---|---|---|
| Model coefficient | |||||
| Time | 0.293* | 0.382+ | 0.353* | 0.491* | 0.096 |
| Time2 | −0.071 | −0.140 | −0.115 | −0.188+ | −0.019 |
| Race | −0.450** | 0.061 | −0.249 | −0.368 | −0.520* |
| Time × Race | 0.339+ | 0.253 | 0.373 | 0.096 | −0.277 |
| Time2 × Race | −0.124 | −0.041 | −0.118 | 0.491* | 0.096 |
| RRI | 0.112* | 0.099** | 0.025 | 0.168 | |
| RRI × Race | −0.173* | −0.114 | −0.006 | 0.034 | |
| RRI × Time | −0.039 | −0.033 | −0.035 | 0.222 | |
| RRI × Race × Time | 0.039 | −0.238 | 0.110+ | −0.446 |
Note: RRI, the term for racial inequity loaded in the specific model. SES, socioeconomic status. EOD, experiences of discrimination. RS, racial socialization. ULSS, urban life stressors as measured by the Urban Life Stressors Scale. In the SES column, the RRI term represents the coefficient for SES and SES interactions; the same applies for EOD, RS, and ULSS.
p < .10.
p < .05.
p < .01.
Racial inequity models
To examine factors that could contribute to development of race differences that emerged by the Strange Situation procedure, a series of indices of racial inequity were added to the Strange Situation procedure race model, as both main effects and interaction terms with race. These models were subsequently run within Black infants, to move beyond contrasting between Black and White infants and avoid a deficit model.
SES.
When SES was added to the 12-month Strange Situation procedure model, no effect of SES or SES × Race interaction was found on cortisol reactivity to the Strange Situation procedure (see Table 2), but there was a significant a Race × SES interaction observed on overall cortisol levels, β = −0.17, 95% CI [−0.33, −0.02], p = .033. To the decompose SES × Race interaction, SES was analyzed within Black infants. SES was not significant in Black infants, β = 0.10, 95% CI [−0.02, 0.22], p = .094, but trended toward the direction that greater SES was associated with higher overall cortisol levels.
Experiences of discrimination.
The addition of experiences of discrimination to the 12-month Strange Situation procedure model resulted in the loss of a significant association with race, β = −0.25, 95% CI [−0.56, 0.06], p = .121, and a direct significant effect of experiences of discrimination, β = 0.10, 95% CI [0.03, 0.17], p = .007, illustrating that experiences of discrimination accounted for some of the variance previously accounted for by race in cortisol trajectory across the Strange Situation procedure. When examined within Black infants, higher maternal report of her own experiences of discrimination predicted higher cortisol, β = 0.101, 0.18], 95% CI [0.02, p = .014 (see Figure 2; Table 4).
Figure 2.
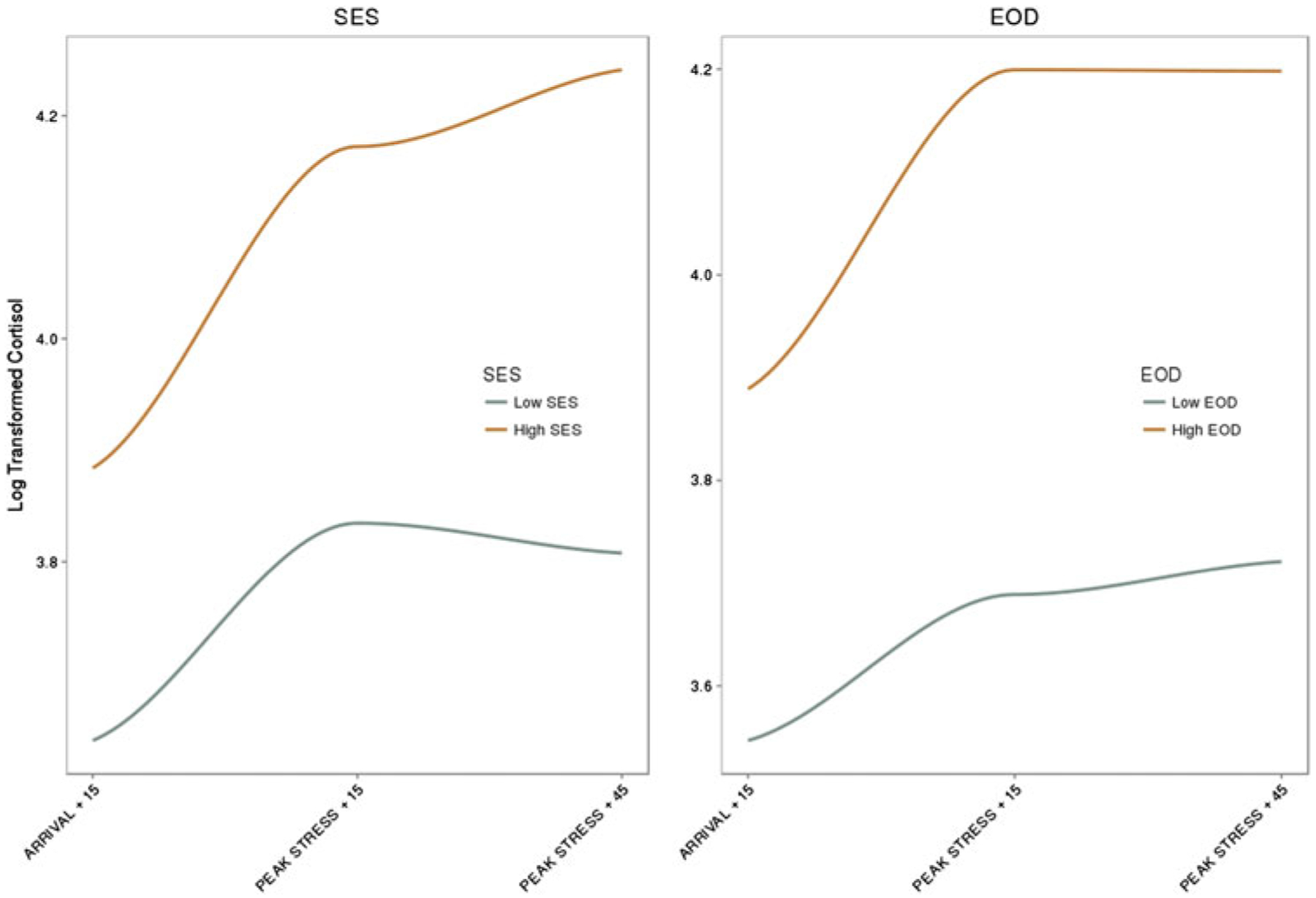
Cortisol differences by socioeconomic status and experiences of discrimination at the 12-month Strange Situation procedure.
Table 4.
Coefficients of racial inequity within Black infants at the Strange Situation procedure visit
| RRI | ||||
|---|---|---|---|---|
| Predictor | SES | EOD | RS | ULSS |
| Model coefficient | ||||
| Time | 0.38+ | 0.35* | 0.49* | 0.09 |
| Time2 | −0.14 | −0.11 | −0.19+ | −0.02 |
| RRI | 0.10+ | 0.10* | 0.03 | 0.17 |
| RRI × Time | −0.04 | −0.03 | −0.03 | 0.22 |
| RRI × Time2 | 0.03 | 0.02 | 0.02 | −0.06 |
Note: RRI, the term for racial inequity loaded in the specific model. SES, socioeconomic status. EOD, experiences of discrimination. RS, racial socialization. ULSS, urban life stessors as measured by the Urban Life Stressors Scale. In the SES column, the RRI term represents the coefficient for SES within races; the same applies for EOD, RS, and ULSS.
p < .10.
p < .05.
Racial socialization.
When racial socialization was added to the 12-month Strange Situation procedure model, race was no longer a significant predictor of overall cortisol levels, β = −0.37, 95% CI [−0.81, 0.07], p = .102, and did not predict cortisol responsivity or recovery, linear β = −0.09, 95% CI [−0.71, 0.52], p = .766; quadratic β = 0.10, 95% CI [−0.19, 0.38], p = .516) Racial socialization was also not a significant predictor of overall cortisol or cortisol reactivity, levels β = 0.02, 95% CI [−0.01, 0.06], p = .208; linear β = −0.03, 95% CI [−0.09, 0.02], p = .226; quadratic β = 0.02, 95% CI [−0.01, 0.05], p = .134. The Racial Socialization × Race × Time, β = 0.11, 95% CI [−0.02, 0.24], p = .099, and Time2, β = −0.05, 95% CI [−0.11, 0.01], p = .086, interaction was not significant. When examined within Black infants, racial socialization was not significant for as a main effect, β = 0.03, 95% CI [−0.02, 0.07], p = .250, or as a cross-level interaction with time, β = −0.03, 95% CI [−0.09, 0.02], p = .223.
ULSS.
When urban life stressors were added to the 12-month Strange Situation procedure model, race remained significant, β = −0.52, 95% CI [−0.99, −0.06], p = .03. The ULSS was not significant as a predictor of cortisol levels or cortisol responsivity to the Strange Situation procedure, levels β = 0.17, 95% CI [−0.05, 0.39], p = .142; linear β = 0.22, 95% CI [−0.10, 0.55], p = .185; quadratic β = −0.06, 95% When examined within race, urban life stressors were not significant overall for Black infants, β = 0.17, 95% CI [−0.08, 0.42], p = .175, and were not significant as a time interaction with Black infants, β = 0.22, 95% CI [−0.10, 0.55], p = .183.
Discussion
This is the first study, to our knowledge, to examine race as a possible moderator of cortisol reactivity to an age-appropriate dyadic stressor before and after the expected development of the attachment relationship in infancy. Our results suggest that by 12 months of age, race differences in cortisol output are apparent across the Strange Situation procedure, with Black infants exhibiting overall higher cortisol output, whereas no race differences were observed across the Still Face procedure at 4 months of age. A core observation motivating this investigation was that, later in life, many of the diseases, including cardiovascular disease, obesity, diabetes, and stroke, associated with altered HPA function exhibit significant race disparities. (Kurian & Cardarelli, 2007; Wong et al., 2002). Our findings, when coupled with the broader literature, suggest that the earliest roots of race-related health disparities are emerging in the first year of life but, at least regarding cortisol reactivity, were not yet apparent by 4 months of age. This finding provides a critical window of opportunity for researchers aiming to reduce health disparities in stress-related disorders as it suggests that early preventative efforts, particularly those implemented in the first year of life, may mitigate the negative effects of maternal exposure to racism and biologically buffer the infant from future health risk.
Our results must be situated in a developmental and environmental context. We found elevated cortisol across the Strange Situation procedure for Black infants. Similar effects in the context of caregiving stress have been reported later in life such that Black women exhibit greater cortisol response to caregiving stress than White women (Wilcox, Bopp, Wilson, Fulk, & Hand, 2005). However, our results diverge from studies at later developmental time points. For example, Chong et al. (2008) found blunted cortisol reactivity to a psychosocial stressor in Black adolescents relative to White adolescents, and Hostinar et al. (2014) found Black adolescents had lower baseline cortisol at a psychosocial stressor than Whites. The lack of consensus in a clear pattern is likely a result of differences in developmental time point and choice of stressor. It is remarkable that, while reactive cortisol patterns may not be stable across development and context, clear cortisol patterns do manifest in infants as early as 4 months of age, and that race differences in cortisol levels emerge by 12 months. This suggests an early embedding of context, and the need for larger longitudinal studies to better delineate how individuals are changing across time and context.
To better understand the contributing factors underpinning racial differences, we used the frameworks suggested by Causadias, Telzer, and Lee (2017) and Doane et al. (2018). Racial inequity indices helped shed light on the racial differences observed in physiological processes during infancy. Specifically, we found that, at 12 months of age, the interaction between cortisol and race was moderated by maternal report of SES during pregnancy. This suggests that the impact of poverty and SES on the developing HPA axis may not be equivalent for Black and White infants. Furthermore, markers of SES and potentially larger structural factors (e.g., structural racism) that lead to community disinvestment have been linked to stress reactivity in children (Hackman, Betancourt, Brodsky, Hurt, & Farah, 2012). Neighborhood violence, often associated with concentrated poverty and disadvantage, has been shown to impact cortisol reactivity in an older, high-risk all Black sample (Theall, Shirtcliff, Dismukes, Wallace, & Drury, 2017). Although much of the current literature links inequalities in SES with potentiated racial inequality, our results suggest that SES as a component of racial inequality contributes to the development of physiological differences.
Urban life stressors did not influence race differences in cortisol patterns. However, experiences of discrimination and racial socialization resulted in changes to the relation between race and cortisol patterns. Experiences of discrimination in Black infants’ mothers accounted for a significant proportion of the variance in race differences in cortisol patterns, and higher maternal report of worry related to discrimination was associated with significantly higher cortisol levels providing the first data, to our knowledge, of a cross-generational impact of discrimination on infant physiology. This finding is in line with the framework suggested by Causadias (2013) and Doane et al. (2018), which suggest that race differences in physiological parameters manifest as a result of many different interacting pathways, of which racial discrimination often occupies a critical role. Cortisol output across the 12 month Strange Situation based on maternal report of discrimination further extends evidence provided by Korous et al. (2017), who found racial discrimination to have a small and inconsistent effect on cortisol output across studies in a meta-analysis of adolescent and adult cohorts. This is the first study to investigate this effect in infants. The fact that race was not significant when discrimination was included in the model highlights the role of racial discrimination in the relationship between race and physiological parameters such as cortisol, even across generations.
Discrimination is a multidimensional phenomenon and can act as a potent stressor, though current measures of discrimination are limited (Williams & Mohammed, 2009; Williams et al., 2003). The racial discrimination literature has begun to coalesce around the themes of daily hassles and life events (Williams & Mohammed, 2009). The broader stress literature can offer insights into future paths for the assessment of discrimination. Several decades of research (Cohen, Kessler, & Gordon, 1997) track how stress physiologists have grappled with questions surrounding objective versus perceived impact of stressors, developmental timing of stressors, the duration and/or chronicity of stressful events, and the distinction between stress and coping responses. Specifically for cortisol, the types of stress exposures appear to shape physiological responses, with uncontrollability and social evaluative threat emerging as particularly potent characteristics of stressors (Dickerson & Kemeny, 2004). Theoretical models posit that these contexts are stressful because they contain life-history relevant information about resource availability, extrinsic morbidity-mortality cues such as the presence of threats, and unpredictability or uncontrollability cues about the stability of the environment (Del Giudice, Ellis, & Shirtcliff, 2011). This literature increasingly focuses on whether or not a stressful event occurred (Anda et al., 2006) or whether the stress exposure was chronic and/or severe (McEwen, 2004).
We have employed the stress framework in our paper but acknowledge that this approach may inadvertently lead to a deficit model where, for example, being Black in the United States is contrasted with being a member of a majority group with presumably fewer stressors. Focusing on within-race analyses helps minimize the deficit model. We also explore acculturation factors that may broadly shape stress physiology in a protective manner. However, we also acknowledge race as a social categorization (American Anthropological Association, 1998; Jorde & Wooding, 2004) and the limitations that examining race entails. Bolstered by stereotypes and false myths about human differences and group behaviors perpetuated over centuries, racial stratification in societal treatment, opportunities, and access to power and resources in the United States remains. Therefore, examining the impact of race is key to understanding and eliminating racial disparities in health across the life course.
A stress framework distinguishes between objective and perceived stress, generally finding that objective stressors exert a stronger impact on stress physiology than subjective experiences (Cohen et al., 1997) and that individual differences in appraisal and coping may be fruitful targets for mitigating the impact of stress exposure (Taylor, 2010). Within racial/ ethnic minority populations, however, racial discrimination appears highly impactful. Discrimination largely revolves around power imbalances, and strategies geared toward racial socialization are employed when there is a perceived need for buffering the next generation from engendering feelings of powerlessness and inferiority based on these experiences. That race differences in HPA functioning in study after study persist even after accounting for objective and perceived stressors is a testament to the realities and complexities of what it means to be a racial/ethnic minority in the United States and supports the idea that inequalities tap into very real stressors that are unlikely to be mitigated through reappraisal or enhanced coping strategies. Our data suggests that these effects even span generations.
The physiological and biological information provided by measures like cortisol suggest that racial discrimination is impactful precisely because it is tapping into highly salient social cues. The stress-response system is calibrated by threats, including social evaluation and extrinsic morbidity-mortality cues, as well as unpredictable or uncontrollable contexts that inform the stability of the environment and availability of stress buffers (Del Giudice et al., 2011). Exposure to racial discrimination is encountered with unfortunate ubiquity for members of racial/ethnic minority groups. The stress-response system may be attuned and highly capable of differentiating subtle cues about social threats and availability of stress buffers (i.e., at distinguishing the ill-informed microaggressions from hostile power displays of intimidation).
Contrary to our expectations, maternal exposure to racial socialization did not influence race differences, suggesting that the buffering impact of racial socialization in other domains may not be transmitted across generations, or at least may not reflect changes in cortisol during the first year. It is also possible that increased racial socialization between grandparents and parents may index greater exposure to racism and discrimination in caregivers overall.
The articles included in this Special Issue may include biological measures, but do not espouse biological essentialism, which has been used to perpetuate racial hierarchies (Causadias et al., 2017; Hartigan, 2010). This view of race can lead to “justification for a racially inequitable status quo and for the continued social marginalization of historically disadvantaged groups” (Williams & Eberhardt, 2008). This essentialist view of behavior is neither acceptable nor ac curate. Human health and behavior is an interaction between biology and an individual’s environment that varies through time and across contexts, shifting and adapting continuously to the ever-changing world. The mitigation of racial differences with the inclusion of experiences of discrimination, a well-established correlate of social inequity, strongly contradicts the biological essentialist viewpoint and highlights the importance of viewing the individual as a contextually adaptive organism moving and changing across time (Causadias, 2013; Causadias et al., 2017; Del Giudice et al., 2011). Cultural context and biology are in concert, not at odds.
This study also provides novel information about the developing HPA axis given the targeted use of a dyadic stressor in which the social and emotional cues from the infant’s mother are ambiguous to the infant. This contrasts with much of the extant literature, which uses novelty or pain stimuli to induce a stress response which, as the infant develops, diminishes in intensity (Gunnar & Donzella, 2002; Gunnar et al., 2009; Loman, Gunnar, & Early Experience, Stress, and Neurobehavioral Development Center, 2010). In this study, we found cortisol reactivity to both the Still Face paradigm and the Strange Situation procedure, paradigms that are distressing to the infant through the disruption of the putative safe haven that is mom. This brief, experimental disruption is a cause for alarm for the infant and warrants a physiological response to mobilize all the resources of the infant to attend and respond to this situation. Mobilization of the infant’s stress-response systems ideally results in behavior patterns in both the infant and the mother that bring the dyad back in synchrony. The infant may be receptive to the mother’s contingent responses, creating a synchronous dyadic pattern that is a prerequisite for future infant self-regulation and socioemotional development (Feldman, 2012; Feldman & Eidelman, 2009).
In the first years of an infants’ life, much of the world around them is novel; they are constantly experiencing and integrating new information. Stress is partly characterized by novelty and unpredictability (Dickerson & Kemeny, 2004). To an infant newly experiencing the world, much will be novel and unpredictable. Stressors such as inoculation, hand restraint, or diaper change putatively tap into the stress of novelty and unpredictability. Despite this stress exposure, it may not be biologically adaptive to mount a dramatic stress response to novel, unpredictable, or even painful stimuli in a time period when much of the stimuli the infant encounters meet those criteria. The early caregiver-child relationship functions as a powerful buffer for infants and toddlers, protecting them from the continuous impact of novelty and unpredictability (Albers, Riksen-Walraven, Sweep, & de Weerth, 2008). The dyadic stressors used here may be more effective in eliciting HPA reactivity across infancy by involving the maternal caregiver as a source of stress and not allowing the infant to utilize the mother as a buffer. There is a proposed biological utility for the decreasing stress response to the environmental cues over the first years of life: to facilitate exploration without incurring the biological costs of a consistently hyperactive HPA axis.
Limitations
Several limitations to this study apply. Our findings are likely generalizable only to term infants. Table 1 showed that gestational period was shorter in Black infants, consistent with larger epidemiological studies, and no preterm infants were included in this analysis. However, our findings persist beyond the effects of gestational age, which, along with infant sex, was controlled for in all models. Although significant reactivity was observed to the Strange Situation procedure, we did not detect significant recovery. Although it is possible that this may be reflective of the developing HPA axis and the limited ability to recover in that time frame, an alternative explanation is that the remaining aspects of the visit continued to activate the HPA axis and recovery occurred at a later time point not captured with the collection timing and collecting more samples would have captured recovery. It is also possible that recovery is a function of attachment classification and that infants with a secure attachment relationship would exhibit the most robust recovery. Indices of racial inequity were chosen based on available data from our survey and are not exhaustive measures that meet the criteria of contributing to race-related differences in HPA activity or the broader social and cultural context of minorities in the United States. Due to power and sample size considerations, a formal test of mediation was not possible. Other potential covariates that are most often captured in daily diary format, or with older populations, such as sleep data, were not available for this sample given highly disordered sleep patterns of infants. It is also possible that differences between the Strange Situation and the Still Face procedures are due to the tasks themselves and not developmental considerations. We are limited to using tasks that are appropriate across development. Finally, the difference in availability of cortisol between the Still Face paradigm and the Strange Situation procedure samples is because some participants were not yet old enough to complete the Strange Situation procedure.
Conclusions and future directions
It is important for future studies to track the long-term implications of higher or lower cortisol levels beginning at this early age. It may be that the expected patterns that underlie resilience or risk differ by race and/or the environmental context in which the infant grows and develops. Studies in both typically developing and high-risk samples side-by-side that are adequately powered to test both between and within race are needed. Longitudinal studies that examine these trajectories in communities, and ideally large epidemiological samples, are needed to determine what pathways are adaptive and/or characteristic of resilience.
Although we cannot say that one pattern of stress reactivity or another is more adaptive, we can say that Black and White infants do not have the same HPA levels across a dyadic stressor at 12 months of age, and that this is informed by experiences such as racial discrimination within Black families. Our data suggest that in order to minimize disparities in health trajectories across the life span that are related to the stress-response systems, intervention strategies need to be employed as early as possible. Given the key relevance of the early parent-child relationship in shaping the stress-response systems, including the HPA axis, early interventions that focus on strengthening the biological buffering capacity of the attachment relationship represents an underutilized and likely powerful approach to addressing persistent health disparities.
References
- Adam EK, Quinn ME, Tavernier R, McQuillan MT, Dahlke KA, & Gilbert KE (2017). Diurnal cortisol slopes and mental and physical health outcomes: A systematic review and meta-analysis. Psychoneuroendocrinology, 83, 25–41. doi: 10.1016/j.psyneuen.2017.05.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ainsworth MD, Blehar MC, Waters E, & Wall SN (2015). Patterns of attachment: A psychological study of the Strange Situation. New York: Psychology Press. [Google Scholar]
- Albers EM, Riksen-Walraven JM, Sweep FCGJ, & de Weerth C (2008). Maternal behavior predicts infant cortisol recovery from a mild everyday stressor. Journal of Child Psychology and Psychiatry, and Allied Disciplines, 49, 97–103. doi: 10.1111/j.1469-7610.2007.01818.x [DOI] [PubMed] [Google Scholar]
- American Anthropological Association. (1998). AAA statement on race. Retrieved from http://www.americananthro.org/ConnectWithAAA/Content.aspx?ItemNumber=2583
- Anda RF, Felitti VJ, Bremner JD, Walker JD, Whitfield C, Perry BD, & Giles WH (2006). The enduring effects of abuse and related adverse experiences in childhood: A convergence of evidence from neurobiology and epidemiology. European Archives of Psychiatry and Clinical Neuroscience, 256, 174–186. doi: 10.1007/s00406-005-0624-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Attar BK, Guerra NG, & Tolan PH (1994). Neighborhood disadvantage, stressful life events and adjustments in urban elementary-school children. Journal of Clinical Child Psychology, 23, 391–400. doi: 10.1207/s15374424jccp2304_5 [DOI] [Google Scholar]
- Barr DA (2014). Health disparities in the United States: Social class, race, ethnicity, and health. Baltimore, MD: Johns Hopkins University Press. [Google Scholar]
- Bates D, Maechler M, & Bolker B (2012). lme4: linear mixed-effects models using S4 classes. R package version 0.999375–42. 2011.
- Black LL, Johnson R, & VanHoose L (2015). The relationship between perceived racism/discrimination and health among Black American women: A review of the literature from 2003 to 2013. Journal of Racial and Ethnic Health Disparities, 2, 11–20. doi: 10.1007/s40615-014-0043-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowlby J (2008). A secure base: Parent-child attachment and healthy human development. New York: Basic Books. [Google Scholar]
- Braveman PA, Cubbin C, Egerter S, Chideya S, Marchi KS, Metzler M, & Posner S (2005). Socioeconomic status in health research: One size does not fit all. Journal of the American Medical Association, 294, 2879–2888. doi: 10.1001/jama.294.22.2879 [DOI] [PubMed] [Google Scholar]
- Broderick J, Brott T, Kothari R, Miller R, Khoury J, Pancioli A, … Shukla R (1998). The greater Cincinnati/northern Kentucky stroke study. Stroke: A Journal of Cerebral Circulation, 29, 415–421. doi: 10.1016/j.psyneuen.2005.02.010 [DOI] [PubMed] [Google Scholar]
- Brondolo E (2015). Racial and ethnic disparities in health: Examining the contexts that shape resilience and risk. Psychosomatic Medicine, 77, 2–5. doi: 10.1097/PSY.0000000000000149 [DOI] [PubMed] [Google Scholar]
- Brown DL (2008). African American resiliency: Examining racial socialization and social support as protective factors. Journal of Black Psychology, 34, 32–48. doi: 10.1177/0095798407310538 [DOI] [Google Scholar]
- Burke HM, Davis MC, Otte C, & Mohr DC (2005). Depression and cortisol responses to psychological stress: A meta-analysis. Psychoneuroendocrinology, 30, 846–856. [DOI] [PubMed] [Google Scholar]
- Case A, & Deaton A (2015). Rising morbidity and mortality in midlife among white non-Hispanic Americans in the 21st century. Proceedings of the National Academy of Sciences of the United States of America, 112, 15078–15083. doi: 10.1073/pnas.1518393112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Causadias JM (2013). A roadmap for the integration of culture into developmental psychopathology. Development and Psychopathology, 25(4, Pt. 2), 1375–1398. doi: 10.1017/S0954579413000679 [DOI] [PubMed] [Google Scholar]
- Causadias JM, Telzer EH, & Lee RM (2017). Culture and biology interplay: An introduction. Cultural Diversity & Ethnic Minority Psychology, 23, 1–4. doi: 10.1037/cdp0000121 [DOI] [PubMed] [Google Scholar]
- Chong RY, Uhart M, McCaul ME, Johnson E, & Wand GS (2008). Whites have a more robust hypothalamic-pituitary-adrenal axis response to a psychological stressor than blacks. Psychoneuroendocrinology, 33, 246–254. doi: 10.1016/j.psyneuen.2007.10.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark R, Anderson NB, Clark VR, & Williams DR (1999). Racism as a stressor for African Americans: A biopsychosocial model. American Psychologist, 54, 805–816. [DOI] [PubMed] [Google Scholar]
- Cohen S, Kessler RC, & Gordon LU (1997). Measuring stress: A guide for health and social scientists. Oxford: Oxford University Press. [Google Scholar]
- Cohen S, Schwartz JE, Epel E, Kirschbaum C, Sidney S, & Seeman T (2006). Socioeconomic status, race, and diurnal cortisol decline in the Coronary Artery Risk Development in Young Adults (CARDIA) Study. Psychosomatic Medicine, 68, 41–50. doi: 10.1097/01.psy.0000195967.51768.ea [DOI] [PubMed] [Google Scholar]
- Del Giudice M, Ellis BJ, & Shirtcliff EA (2011). The Adaptive Calibration Model of stress responsivity. Neuroscience and Biobehavioral Reviews, 35, 1562–1592. doi: 10.1016/j.neubiorev.2010.11.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeSantis AS, Adam EK, Doane LD, Mineka S, Zinbarg RE, & Craske MG (2007). Racial/ethnic differences in cortisol diurnal rhythms in a community sample of adolescents. Journal of Adolescent Health, 41, 3–13. doi: 10.1016/j.jadohealth.2007.03.006 [DOI] [PubMed] [Google Scholar]
- Dickerson SS, & Kemeny ME (2004). Acute stressors and cortisol responses: A theoretical integration and synthesis of laboratory research. Psychological Bulletin, 130, 355–391. doi: 10.1037/0033-2909.130.3.355 [DOI] [PubMed] [Google Scholar]
- Doane LD, Sladek MR, & Adam EK (2018). An introduction to cultural neurobiology: Evidence from physiological stress systems In Causadias JM, Telzer EH, & Gonzales NA (Eds.), Handbook of culture and biology (Vol. 1, pp. 227–254). New York: Wiley. [Google Scholar]
- Epel E, Lapidus R, McEwen B, & Brownell K (2001). Stress may add bite to appetite in women: A laboratory study of stress-induced cortisol and eating behavior. Psychoneuroendocrinology, 26, 37–49. [DOI] [PubMed] [Google Scholar]
- Epel ES, McEwen B, Seeman T, Matthews K, Castellazzo G, Brownell KD, … Ickovics JR (2000). Stress and body shape: Stress-induced cortisol secretion is consistently greater among women with central fat. Psychosomatic Medicine, 62, 623–632. [DOI] [PubMed] [Google Scholar]
- Federal Reserve. (2018). Recent trends in wealth-holding by race and ethnicity: Evidence from the Survey of Consumer Finances. Retrieved April 19, 2018, from https://www.federalreserve.gov/econres/notes/feds-notes/recent-trends-in-wealth-holding-by-race-and-ethnicity-evidence-from-the-survey-of-consumer-finances-20170927.htm
- Feldman R (2012). Parent-infant synchrony: A biobehavioral model of mutual influences in the formation of affiliative bonds. Monographs of the Society for Research in Child Development, 77, 42–51. doi: 10.1111/j.1540-5834.2011.00660.x [DOI] [Google Scholar]
- Feldman R, & Eidelman AI (2009). Biological and environmental initial conditions shape the trajectories of cognitive and social-emotional development across the first years of life. Developmental Science, 12, 194–200. doi: 10.1111/j.1467-7687.2008.00761.x [DOI] [PubMed] [Google Scholar]
- Finch WH, Bolin JE, & Kelley K (2016). Multilevel modeling using R. Crc Press. [Google Scholar]
- Fischer AR, & Shaw CM (1999). African Americans’ mental health and perceptions of racist discrimination: The moderating effects of racial socialization experiences and self-esteem. Journal of Counseling Psychology, 46, 395. doi: 10.1037/0022-0167.46.3.395 [DOI] [Google Scholar]
- Gluck ME, Geliebter A, & Lorence M (2004). Cortisol stress response is positively correlated with central obesity in obese women with binge eating disorder (BED) before and after cognitive-behavioral treatment. Annals of the New York Academy of Sciences, 1032, 202–207. doi: 10.1196/annals.1314.021 [DOI] [PubMed] [Google Scholar]
- Goldmann E, Aiello A, Uddin M, Delva J, Koenen K, Gant LM, & Galea S (2011). Pervasive exposure to violence and posttraumatic stress disorder in a predominantly African American urban community: The Detroit Neighborhood Health Study. Journal of Traumatic Stress, 24, 747–751. doi: 10.1002/jts.20705 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray SAO, Jones CW, Theall KP, Glackin E, & Drury SS (2017). Thinking across generations: Unique contributions of maternal early life and prenatal stress to infant physiology. Journal of the American Academy of Child & Adolescent Psychiatry, 56, 922–929. doi: 10.1016/j.jaac.2017.09.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gunnar MR, Brodersen L, Krueger K, & Rigatuso J (1996). Dampening of adrenocortical responses during infancy: Normative changes and individual differences. Child Development, 67, 877–889. doi: 10.1111/j.1467-8624.1996.tb01770.x [DOI] [PubMed] [Google Scholar]
- Gunnar MR, Brodersen L, Nachmias M, Buss K, & Rigatuso J (1996). Stress reactivity and attachment security. Developmental Psychobiology, 29, 191–204. doi: 10.1002/(SICI)1098-2302(199604)29:3,191::AID-DEV1.3.0.CO;2-M [DOI] [PubMed] [Google Scholar]
- Gunnar MR, & Donzella B (2002). Social regulation of the cortisol levels in early human development. Psychoneuroendocrinology, 27, 199–220. doi: 10.1016/S0306-4530(01)00045-2 [DOI] [PubMed] [Google Scholar]
- Gunnar MR, Talge NM, & Herrera A (2009). Stressor paradigms in developmental studies: What does and does not work to produce mean increases in salivary cortisol. Psychoneuroendocrinology, 34, 953–967. doi: 10.1016/j.psyneuen.2009.02.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hackman DA, Betancourt LM, Brodsky NL, Hurt H, & Farah MJ (2012). Neighborhood disadvantage and adolescent stress reactivity. Frontiers in Human Neuroscience, 6, 277. doi: 10.3389/fnhum.2012.00277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hajat A, Diez-Roux A, Franklin TG, Seeman T, Shrager S, Ranjit N, … Kirschbaum C (2010). Socioeconomic and race/ethnic differences in daily salivary cortisol profiles: The multi-ethnic study of atherosclerosis. Psychoneuroendocrinology, 35, 932–943. doi: 10.1016/j.psyneuen.2009.12.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haley DW, & Stansbury K (2003). Infant stress and parent responsiveness: Regulation of physiology and behavior during still-face and reunion. Child Development, 74, 1534–1546. doi: 10.1111/1467-8624.00621 [DOI] [PubMed] [Google Scholar]
- Hamer M, O’Donnell K, Lahiri A, & Steptoe A (2010). Salivary cortisol responses to mental stress are associated with coronary artery calcification in healthy men and women. European Heart Journal, 31, 424–429. doi: 10.1093/eurheartj/ehp386 [DOI] [PubMed] [Google Scholar]
- Harrell CJP, Burford TI, Cage BN, Nelson TM, Shearon S, Thompson A, & Green S (2011). Multiple pathways linking racism to health outcomes. Du Bois Review, 8, 143–157. doi: 10.1017/S1742058X11000178 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartigan J (2010). Race in the 21st century: Ethnographic approaches. Oxford: Oxford University Press. [Google Scholar]
- Hasson BR, Apovian C, & Istfan N (2015). Racial/ethnic differences in insulin resistance and beta cell function: Relationship to racial disparities in Type 2 diabetes among African Americans versus Caucasians. Current Obesity Reports, 4, 241–249. doi: 10.1007/s13679-015-0150-2 [DOI] [PubMed] [Google Scholar]
- Hertsgaard L, Gunnar M, Erickson MF, & Nachmias M (1995). Adrenocortical responses to the Strange Situation in infants with disorganized/disoriented attachment relationships. Child Development, 66, 1100. doi: 10.2307/1131801 [DOI] [PubMed] [Google Scholar]
- Hostinar CE, McQuillan MT, Mirous HJ, Grant KE, & Adam EK (2014). Cortisol responses to a group public speaking task for adolescents: Variations by age, gender, and race. Psychoneuroendocrinology, 50, 155–166. doi: 10.1016/j.psyneuen.2014.08.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hughes D, & Chen L (1997). When and what parents tell children about race: An examination of race-related socialization among African American families. Applied Developmental Science, 1, 200–214. doi: 10.1207/s1532480xads0104_4 [DOI] [Google Scholar]
- Hughes D, Rodriguez J, Smith EP, Johnson DJ, Stevenson HC, & Spicer P (2006). Parents’ ethnic-racial socialization practices: A review of research and directions for future study. Developmental Psychology, 42, 747–770. doi: 10.1037/0012-1649.42.5.747 [DOI] [PubMed] [Google Scholar]
- Iceland J, & Wilkes R (2006). Does socioeconomic status matter? Race, class, and residential segregation. Social Problems, 53, 248–273. doi: 10.1525/sp.2006.53.2.248 [DOI] [Google Scholar]
- Jaffee KD, Liu GC, Canty-Mitchell J, Qi RA, Austin J, & Swigonski N (2005). Race, urban community stressors, and behavioral and emotional problems of children with special health care needs. Psychiatric Services, 56, 63–69. doi: 10.1176/appi.ps.56.1.63 [DOI] [PubMed] [Google Scholar]
- Jansen J, Beijers R, Riksen-Walraven M, & de Weerth C (2010). Cortisol reactivity in young infants. Psychoneuroendocrinology, 35, 329–338. doi: 10.1016/j.psyneuen.2009.07.008 [DOI] [PubMed] [Google Scholar]
- Jones CP (2000). Levels of racism: A theoretic framework and a gardener’s tale. American Journal of Public Health, 90, 1212–1215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jorde LB, & Wooding SP (2004). Genetic variation, classification, and “race.” Nature Genetics, 11, S28–S33. [DOI] [PubMed] [Google Scholar]
- Kochanek KD, Murphy SL, & Xu J (2015). Deaths: Final data for 2011. National Vital Statistics Reports, 63, 3 Retrieved from https://www.cdc.gov/nchs/data/nvsr/nvsr63/nvsr63_03.pdf [PubMed] [Google Scholar]
- Korous KM, Causadias JM, & Casper DM (2017). Racial discrimination and cortisol output: A meta-analysis. Social Science & Medicine, 193, 90–100. doi: 10.1016/j.socscimed.2017.09.042 [DOI] [PubMed] [Google Scholar]
- Kressin NR, Raymond KL, & Manze M (2008). Perceptions of race/ethnicity-based discrimination: A review of measures and evaluation of their usefulness for the health care setting. Journal of Health Care for the Poor and Underserved, 19, 697–730. doi: 10.1353/hpu.0.0041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krieger N (1990). Racial and gender discrimination: Risk factors for high blood pressure? Social Science & Medicine, 30, 1273–1281. [DOI] [PubMed] [Google Scholar]
- Krieger N, Smith K, Naishadham D, Hartman C, & Barbeau EM (2005). Experiences of discrimination: Validity and reliability of a self-report measure for population health research on racism and health. Social Science & Medicine, 61, 1576–1596. doi: 10.1016/j.socscimed.2005.03.006 [DOI] [PubMed] [Google Scholar]
- Kurian AK, & Cardarelli KM (2007). Racial and ethnic differences in cardiovascular disease risk factors: A systematic review. Ethnicity & Disease, 17, 143–152. [PubMed] [Google Scholar]
- Levy DJ, Heissel JA, Richeson JA, & Adam EK (2016). Psychological and biological responses to race-based social stress as pathways to disparities in educational outcomes. American Psychologist, 71, 455–473. doi: 10.1037/a0040322 [DOI] [PubMed] [Google Scholar]
- Lewis M, & Ramsay D (2005). Infant emotional and cortisol responses to goal blockage. Child Development, 76(2), 518–530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis M, Ramsay DS, & Kawakami K (1993). Differences between Japanese infants and Caucasian American infants in behavioral and cortisol response to inoculation. Child Development, 64, 1722–1731. doi: 10.2307/1131465 [DOI] [PubMed] [Google Scholar]
- Loman MM, Gunnar MR, & Early Experience, Stress, and Neurobehavioral Development Center. (2010). Early experience and the development of stress reactivity and regulation in children. Neuroscience and Biobehavioral Reviews, 34, 867–876. doi: 10.1016/j.neubiorev.2009.05.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lupien SJ, King S, Meaney MJ, & McEwen BS (2001). Can poverty get under your skin? Basal cortisol levels and cognitive function in children from low and high socioeconomic status. Development and Psychopathology, 13, 653–676. doi: 10.1017/S0954579401003133 [DOI] [PubMed] [Google Scholar]
- Maniam J, Antoniadis C, & Morris MJ (2014). Early-life stress, HPA axis adaptation, and mechanisms contributing to later health outcomes. Frontiers in Endocrinology, 5, 73. doi: 10.3389/fendo.2014.00073 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez-Torteya C, Muzik M, McGinnis EW, Rosenblum KL, Bocknek EL, Beeghly M, … Abelson JL (2015). Longitudinal examination of infant baseline and reactivity cortisol from ages 7 to 16 months. Developmental Psychobiology, 57, 356–364. doi: 10.1002/dev.21296 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McEwen BS (2004). Protection and damage from acute and chronic stress: Allostasis and allostatic overload and relevance to the pathophysiology of psychiatric disorders. Annals of the New York Academy of Sciences, 1032, 1–7. doi: 10.1196/annals.1314.001 [DOI] [PubMed] [Google Scholar]
- Merritt MM, & Harrell JP (1998). The relationships of somatic symptoms with coping strategies, life satisfaction and daily hassles. Psychosomatic Medicine, 60, 114. [Google Scholar]
- Miller DB (1999). Racial socialization and racial identity: Can they promote resiliency for African American adolescents? Adolescence, 34, 493–501. [PubMed] [Google Scholar]
- Miller DB, & MacIntosh R (1999). Promoting resilience in urban African American adolescents: Racial socialization and identity as protective factors. Social Work Research, 23, 159–169. doi: 10.1093/swr/23.3.159 [DOI] [Google Scholar]
- Miller GE, Chen E, & Zhou ES (2007). If it goes up, must it come down? Chronic stress and the hypothalamic-pituitary-adrenocortical axis in humans. Psychological Bulletin, 133, 25–45. doi: 10.1037/0033-2909.133.1.25 [DOI] [PubMed] [Google Scholar]
- Needham BL, Smith JA, Zhao W, Wang X, Mukherjee B, Kardia SLR, … Diez Roux AV (2015). Life course socioeconomic status and DNA methylation in genes related to stress reactivity and inflammation: The multi-ethnic study of atherosclerosis. Epigenetics, 10, 958–969. doi: 10.1080/15592294.2015.1085139 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norris FH (1992). Epidemiology of trauma: Frequency and impact of different potentially traumatic events on different demographic groups. Journal of Consulting and Clinical Psychology, 60, 409–418. doi: 10.1037/0022-006X.60.3.409 [DOI] [PubMed] [Google Scholar]
- Pascoe EA, & Smart Richman L (2009). Perceived discrimination and health: A meta-analytic review. Psychological Bulletin, 135, 531–554. doi: 10.1037/a0016059 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phillips AC, Ginty AT, & Hughes BM (2013). The other side of the coin: Blunted cardiovascular and cortisol reactivity are associated with negative health outcomes. International Journal of Psychophysiology, 90, 1–7. doi: 10.1016/j.ijpsycho.2013.02.002 [DOI] [PubMed] [Google Scholar]
- Pinheiro JC & Bates DM (2000). Mixed-effects models in S and S-PLUS Statistics and Computing Series. New York: Springer. [Google Scholar]
- R Core Team (2013). R: A language and environment for statistical computing R Foundation for Statistical Computing, Vienna, Austria: Retrievied from http://www.R-project.org/. [Google Scholar]
- Rodriguez J, Huntington-Moskos L, Johnson A, Williams S, Gulledge E, Feeley C, & Rice M (2016). Collecting biological measures for research with children and adolescents. Journal of Pediatric Health Care, 30, 279–283. doi: 10.1016/j.pedhc.2015.12.007 [DOI] [PubMed] [Google Scholar]
- Rosmond R, & Björntorp P (2000). The hypothalamic-pituitary-adrenal axis activity as a predictor of cardiovascular disease, type 2 diabetes and stroke. Journal of Internal Medicine, 247(2), 188–197. [DOI] [PubMed] [Google Scholar]
- Ross MG, & Desai M (2012). Developmental origins of adult health and disease In Gabbe SG, Niebyl JR, Simpson JL, Landon MB, & Galan HL (Eds.), Obstetrics: Normal and problem pregnancies (pp. 83–98). Philadelphia: Elsevier. [Google Scholar]
- Sanders-Phillips K (1996). Correlates of health promotion behaviors in low-income Black women and Latinas. American Journal of Preventive Medicine, 12, 450–458. doi: 10.1016/S0749-3797(18)30267-8 [DOI] [PubMed] [Google Scholar]
- Schoenbaum M, & Waidmann T (1997). Race, socioeconomic status, and health: Accounting for race differences in health. Journals of Gerontol ogy. Series B, Psychological Sciences and Social Sciences, 52, 61–73. doi: 10.1093/geronb/52B.Special_Issue.61 [DOI] [PubMed] [Google Scholar]
- Sen M, & Wasow O (2016). Race as a bundle of sticks: Designs that estimate effects of seemingly immutable characteristics. Annual Review of Political Science, 19, 499–522. doi: 10.1146/annurev-polisci-032015-010015 [DOI] [Google Scholar]
- Skinner ML, Shirtcliff EA, Haggerty KP, Coe CL, & Catalano RF (2011). Allostasis model facilitates understanding race differences in the diurnal cortisol rhythm. Development and Psychopathology, 23, 1167–1186. doi: 10.1017/S095457941100054X [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stevenson HC, & Arrington EG (2009). Racial/ethnic socialization mediates perceived racism and the racial identity of African American adolescents. Cultural Diversity & Ethnic Minority Psychology, 15, 125–136. doi: 10.1037/a0015500 [DOI] [PubMed] [Google Scholar]
- Susman EJ, Dorn LD, Inoff-Germain G, Nottelmann ED, & Chrousos GP (1997). Cortisol reactivity, distress behavior, and behavioral and psychological problems in young adolescents: A longitudinal perspective. Journal of Research on Adolescence, 7, 81–105. doi: 10.1207/s15327795jra0701_5 [DOI] [Google Scholar]
- Tarullo AR, & Gunnar MR (2006). Child maltreatment and the developing HPA axis. Hormones and Behavior, 50, 632–639. doi: 10.1016/j.yhbeh.2006.06.010 [DOI] [PubMed] [Google Scholar]
- Taylor SE (2010). Mechanisms linking early life stress to adult health outcomes. Proceedings of the National Academy of Sciences of the United States of America, 107, 8507–8512. doi: 10.1073/pnas.1003890107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theall KP, Shirtcliff EA, Dismukes AR, Wallace M, & Drury SS (2017). Association between neighborhood violence and biological stress in children. JAMA Pediatrics, 171, 53–60. doi: 10.1001/jamapediatrics.2016.2321 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tronick E, Als H, Adamson L, Wise S, & Brazelton TB (1978). The infant’s response to entrapment between contradictory messages in face-to-face interaction. Journal of the American Academy of Child Psychiatry, 17, 1–13. doi: 10.1016/S0002-7138(09)62273-1 [DOI] [PubMed] [Google Scholar]
- Valencia RR (2010). Dismantling contemporary deficit thinking: Educational thought and practice. London: Routledge. [Google Scholar]
- van Bakel HJA, & Riksen-Walraven JM (2004). Stress reactivity in 15-month-old infants: links with infant temperament, cognitive competence, and attachment security. Developmental Psychobiology, 44(3), 157–167. [DOI] [PubMed] [Google Scholar]
- Wagner J, Lampert R, Tennen H, & Feinn R (2015). Exposure to discrimination and heart rate variability reactivity to acute stress among women with diabetes. Stress and Health, 31, 255–262. doi: 10.1002/smi.2542 [DOI] [PubMed] [Google Scholar]
- Walker BR (2007). Glucocorticoids and cardiovascular disease. European Journal of Endocrinology / European Federation of Endocrine Societies, 157(5), 545–559. [DOI] [PubMed] [Google Scholar]
- Wilcox S, Bopp M, Wilson DK, Fulk LJ, & Hand GA (2005). Race differences in cardiovascular and cortisol responses to an interpersonal challenge in women who are family caregivers. Ethnicity & Disease, 15, 17–24. [PubMed] [Google Scholar]
- Williams DR (1999). Race, socioeconomic status, and health: The added effects of racism and discrimination. Annals of the New York Academy of Sciences, 896, 173–188. doi: 10.1111/j.1749-6632.1999.tb08114.x [DOI] [PubMed] [Google Scholar]
- Williams DR, & Mohammed SA (2009). Discrimination and racial disparities in health: Evidence and needed research. Journal of Behavioral Medicine, 32, 20–47. doi: 10.1007/s10865-008-9185-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams DR, Mohammed SA, Leavell J, & Collins C (2010). Race, socioeconomic status, and health: Complexities, ongoing challenges, and research opportunities. Annals of the New York Academy of Sciences, 1186, 69–101. doi: 10.1111/j.1749-6632.2009.05339.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams DR, Neighbors HW, & Jackson JS (2003). Racial/ethnic discrimination and health: Findings from community studies. American Journal of Public Health, 93, 200–208. doi: 10.2105/AJPH.93.2.200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams DR, Priest N, & Anderson NB (2016). Understanding associations among race, socioeconomic status, and health: Patterns and prospects. Health Psychology, 35, 407–411. doi: 10.1037/hea0000242 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams MJ, & Eberhardt JL (2008). Biological conceptions of race and the motivation to cross racial boundaries. Journal of Personality and Social Psychology, 94, 1033–1047. doi: 10.1037/0022-3514.94.6.1033 [DOI] [PubMed] [Google Scholar]
- Wong MD, Shapiro MF, Boscardin WJ, & Ettner SL (2002). Contribution of major diseases to disparities in mortality. New England Journal of Medicine, 347, 1585–1592. doi: 10.1056/NEJMsa012979 [DOI] [PubMed] [Google Scholar]