

Abstract
Ascertaining the ionizing radiation (IR)-induced bystander response and its preceding molecular regulation would increase our understanding of the mechanism of acute and delayed radiobiological effects. Recent evidence clearly prompted that radiation-induced nuclear factor kappa B (NF-κB) would play a key role in bystander responses in nontargeted cells. Accordingly, we investigated the orchestration of NF-κB signaling after IR in a nontargeted distant organ. Heart tissues from C57/BL6 mice either mock irradiated or exposed (limited to lower abdomen 1 cm diameter) to single-dose IR (SDR: 2 or 10 Gy) or fractionated IR (FIR, 2 Gy per day for 5 days) were examined for onset of abscopal NF-κB signal transduction, translated activity, downstream functional signaling and associated DNA damage. Radiation significantly induced NF-κB DNA binding activity in nontargeted heart. Transcriptional profiling showed that 51, 46 and 26 of 88 genes were significantly upregulated after 2 Gy, 10 Gy and FIR. Of these genes, 22 showed dose- and fractionation-independent upregulation. Immunohistochemistry revealed a robust increase in p65 and cMyc expression in distant heart after SDR and FIR. Immunoblotting revealed increased phosphorylation of p38 after 2 Gy and extracellular signal-regulated kinases 1/2 after 10 Gy in nontargeted heart. In addition, IR exposure significantly enhanced DNA fragmentation in nontargeted heart. Together, these data clearly indicated an induced abscopal response in distant organ after clinically relevant IR doses. More importantly, the results imply that orchestration of NF-κB signal transduction in nontargeted tissues may serve as an effector and could play a key role in induced abscopal responses.
Keywords: ionizing radiation, in vivo abscopal effect, NF-κB, gene expression, fractionated dose
INTRODUCTION
The American Cancer Society estimates a total of 1 638 910 new cancer cases in the United States for 2012,1 and to that end, nearly two-thirds of all cancer patients will receive radiation therapy (RT) as part of their treatment plan.2 RT is used in curative, palliative and prophylactic treatment plans, and is delivered through external beam, internal placement or systemic administration, depending on the type of cancer and treatment goals.3 Despite their benefits and increased survival rates, an important limiting factor is the risk of treatment-related deleterious effects to normal tissues, including secondary cancers that determine the likelihood of cure. Importantly, the collective incidence of second primary malignancies in patients receiving RT is estimated to be as high as 20%.4 Moreover, even low doses of ionizing radiation (IR) being used in diagnostic procedures (X-ray or computed tomography) can increase the risk of development of IR-induced cancers. The potential risks of IR are being debated given their impact on the health of current and future generations.
IR exposure causes the cells to produce signals capable of altering the survival and response of nonirradiated cells.5 Termed as the bystander effect, these signals could be transmitted by direct gap junction and media-soluble factors through a range of signal transduction pathways.5 Many in vitro studies have reported that IR exposure results in increased expression of death receptors and ligands, cytokines and reactive oxygen species. Bystander effect could be manifested in various forms such as genomic instability, apoptosis, cell cycle delay, micronucleus formation, mutations and change in gene expression.5 Evidence of bystander effects has also been found in humans in the form of RT-induced abscopal ‘out-of-exposure field’ effects6 coordinating a complex interplay involving organs, tissues and cells. Studies have shown that IR could elicit molecular/cellular responses in nontargeted cells through the release of soluble clastogenic factors into circulating blood.7 Consequently, clastogenic activity has been reported in the plasma of patients receiving RT and individuals accidentally exposed to IR.8 The in vitro and in vivo9 generation of such factors after IR have also been reported after whole-body irradiation. These factors have been shown to be capable of inducing chromosome damage in cultured cells.10 Moreover, the existence of bystander effects in somatic organs was confirmed using whole-animal models wherein IR exposure of one side of the animal’s body caused profound epigenetic changes in the shielded bystander parts.11 Although the nature of bystander signal in vivo remains to be elucidated in detail, it can reach different tissues within the same organism and exert distinct tissue-specific responses with the nature and magnitude of effects differing with tissue. As the bystander signal transducers/clastogenic factors are extensively investigated both in vitro and in vivo, in this study, we focused on dissecting out the onset of molecular switch, nuclear factor kappa B (NF-κB) and its orchestration that instigates radioresponse in nontargeted tissue. NF-κB is a member of the c-rel proto-oncogene family found within the promoter and enhancer region of a wide variety of cellular genes. Upon activation, NF-κB can stimulate various targeted late-response genes, including those responsible for cell cycle, proliferation, cell growth, differentiation, metastasis, inflammation, apoptosis, adhesion, angiogenesis and oncogenesis. We recently demonstrated that IR selectively and significantly induced NF-κB in a number of tumor models and that inhibition of NF-κB enhances radiosensitivity.12 We observed that radiation-mediated bystander signal transducers/clastogenic factors target NF-κB signaling pathway transcriptome, resulting in NF-κB activation in nontargeted tissues and subsequent abscopal response, here in this case DNA fragmentation.
MATERIALS AND METHODS
Animals and irradiation experiments
All experiments conformed to American Physiological Society standards for Animal Care and were carried out in accordance with the guidelines laid down by the National Research Council and were approved by our institutional animal care and use committee (protocol 08-056X). The 23-day-old C57BL/6 (both male and female) mice (Charles River Laboratories, Wilmington, MA, USA), weighing 10–15 g, were acclimatized for at least 3 days before the study. Six animals were used per IR dose and animals were randomly allocated to each group.13–17 Animals were either mock irradiated or exposed to single-dose low-linear energy transfer γ-irradiation (SDR, 2/10 Gy) or fractionated irradiation (FIR, 2 Gy per day for 5 days (2 Gy × 5)) using gammacell 40 Exactor (Nordion International, Ontario, Canada) at a dose rate of 0.81 Gy min −1. A specially designed cerrobend shield was used to encase the body of the mice, exposing (1 cm diameter) only the lower abdomen (Figure 1). Dosimetry was measured using both thermoluminescent and radiochromic film dosimetry. The films revealed that the top and bottom surface of the mouse under the hole received 1.9 Gy and 2 Gy, respectively. The central axis of the mouse placed under the shielding’s hole received 1.7 Gy. Mock-irradiated animals were treated identically except that they were not subjected to IR. To assess the low dose ionizing radiation-dependent molecular alterations, heart tissue was harvested after a response time of 24 h. At the end of each experiment, animals were killed by inhalation of 5% isoflurane and rapid exsanguination via the arterial line. All efforts were made to minimize animal suffering.
Figure 1.

Photographs of specially designed cerrobend shield (22 cm diameter and 1.2 cm thickness) used to encase the body of the mice, exposing only 1 cm diameter of the lower abdomen.
Electrophoretic mobility shift assay (EMSA)
Nuclear protein extraction and EMSA were performed as described in our earlier study.12 For EMSA, a double-stranded oligonucleotide containing a tandem repeat of the consensus sequence of 5’-GGGGACTTTCC-3’ was end-labeled with [γ-32P]-ATP using T4 polynucleotide kinase. Binding reaction was performed by mixing nuclear extract (5 μg), 0.1 mg of poly(deoxyinosinic-deoxycytidylic) acid and 0.5 ng of [γ-32P]-ATP-labeled NF-κB-specific oligonucleotide probe in reaction buffer (10 mM Tris-Cl, pH 7.5, 100 mM NaCl, 1 mM dithiothreitol, 1 mM EDTA and 20% (v/v) glycerol). For competition assay, the nuclear extract was preincubated with unlabeled homologous NF-κB oligonucleotide followed by addition of [γ-32P]-ATP-labeled NF-κB probe. Super shift analysis was performed as described earlier.12
Quantitative PCR profiling of NF-κB signaling pathway molecules Total RNA extraction and NF-κB signaling pathway profiling were performed as described in our earlier studies.18 In brief, total RNA was extracted with RNA-Stat-60 reagent (Tel-Test, Friendswood, TX, USA) and the reverse transcription was carried out using the iScript complementary DNA synthesis kit (Bio-Rad). Complementary DNA samples, mixed with SYBR Green-based PCR super-mix (Bio-Rad, Hercules, CA, USA), were loaded in PCR plates equipped with a panel of 96 primer sets for a set of 88 NF-κB pathway genes (mouse NF-κB pathway profiler, www.realtimeprimers.com. ). We started with this highly selected quantitative PCR profiler, rather than an all-encompassing microarray, because this approach entails a well-characterized quantitative expression profile governing NF-κB signaling pathway and avoids genes that are not functionally characterized. As IR has been shown to activate NF-κB, a proto-oncogene found within the promoter/enhancer region of a wide variety of genes, we focused on dissecting out the onset of this molecular switch and its orchestration that instigates radioresponse in nontargeted tissue. Each profiling plate was also equipped with a RT-PCR-positive, genomic DNA and housekeeping (β-actin, GAPDH, Rpl13a, HPRT1, β2M) controls. The ΔΔct values were calculated by normalizing the gene expression levels to the internal housekeeping genes, compared between groups, and the relative expression level was expressed as fold change. When comparing each gene’s signal intensity between groups, we used an increase of ⩾2-fold in the experimental group as compared with mock-irradiated group to represent ‘stringent’ criteria for upregulation and an increase of <2-fold to represent ‘less stringent’ criteria.
Immunoblotting
Total protein extraction and immunoblotting was performed as described earlier.19 For this study, the protein-transferred membranes were incubated with 2.0 ng of rabbit polyclonal anti-p38, extracellular signal-regulated kinase (ERK), pERK and p-p38 antibody. Group-wise comparisons were made using analysis of variance with Tukey’s post hoc comparison to determine the stringent significance in expression variation of different proteins in response to interventional procedures, here in this case multiple dose regimens of radiation.
Immunohistochemistry
Modulations in p65 and cMYC expression in distant-heart tissues were assessed by immunohistochemical staining as described earlier.18 All tissue processing and immunohistochemistry was performed in a blinded manner by our institutional tissue pathology core. To avoid variations in immunohistochemical staining across slides, a ‘tissue-macroarray’ was constructed.
DNA fragmentation
Fluorescein-FragEL DNA fragmentation (Oncogene Research Products, Boston, MA, USA) was performed as described in our earlier study.16
RESULTS
Radiation targets NF-κB signaling transcriptome in nontargeted tissue
To determine whether radiation-induced circulating clastogenic factors target the onset of NF-κB signaling in distant organ, heart tissues from animals that received either mock irradiation or exposed to radiation (SDR or FIR) were examined for transcriptional changes in 88 NF-κB signal transduction genes. Compared with mock-irradiated animals, radiation exposure robustly activated NF-κB signaling in nontargeted organ. To that end, we observed that 2 Gy, 10 Gy and 2 Gy FIR exposure resulted in the upregulation of 57, 59 and 44 NF-κB signaling pathway genes in nontargeted heart (Figure 2a). It is noteworthy to mention that 35 of these genes showed both dose- and fractionation-independent upregulation (Figure 2a). Applying stringent criteria (⩾2-fold) revealed a significant upregulation of 51, 46 and 26 NF-κB signaling pathway molecules after 2 Gy (Figure 2b), 10 Gy (Figure 2c) and FIR (Figure 2d) respectively. Pertinently, 22 of these > 2-fold upregulated genes (Figure 2b–d black bars) remained consistent across all three dose regimens investigated, demonstrating the definite onset of NF-κB signaling in nontargeted organ after radiation.
Figure 2.
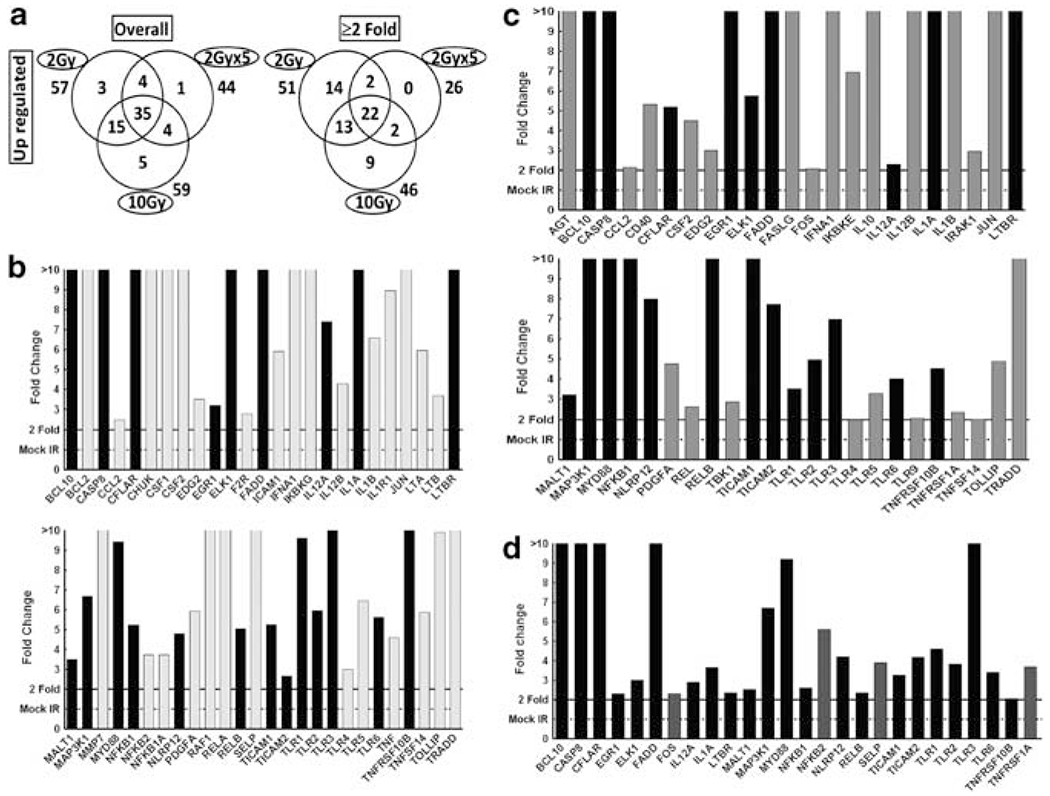
(a) Venn diagrams showing numbers of genes significantly upregulated, with or without application of stringent criteria (⩾2-fold) in nontargeted heart tissues of mice exposed to 2 Gy, 10 Gy or fractionated ionizing radiation (FIR; 2 Gy per day for 5 days (2 Gy × 5)). Numbers outside each circle indicate total upregulated genes in response to a specific dose, those inside greater part of the circles indicate selectively upregulated in that particular dose, those in two overlapping circles indicate upregulated commonly under both conditions and those in the center indicate upregulated in all three doses. (b-d) Histograms showing expression levels of nuclear factor kappa B (NF-κB) pathway molecules that were significantly (>2-fold) upregulated in nontargeted heart after (b) 2Gy, (c) 10 Gy, or (d) 2 Gy x 5. In all, 22 genes were commonly upregulated (highlighted as black bars) after all three dose and fractionation regimens of radiation.
Radiation induces NF-κB DNA-binding activity in nontargeted tissue
To delineate whether radiation-targeted NF-κB signaling translates to NF-κB activation in nontargeted tissue, we examined the modulations in NF-κB DNA-binding activity. We observed a significant induction of NF-κB DNA-binding activity in nontargeted tissue after all radiation dose regimens investigated (Figure 3a). Relatively, FIR exposure resulted in a robust NF-κB activation. Specific binding of NF-κB to its sequence-specific oligonucleotide was confirmed with competition binding assay (Figure 3a). Moreover, supershift assay revealed that the major subunits are p50 and p65. Together, these results demonstrate that radiation exposure stabilize NF-κB activation in distant nontargeted tissue.
Figure 3.
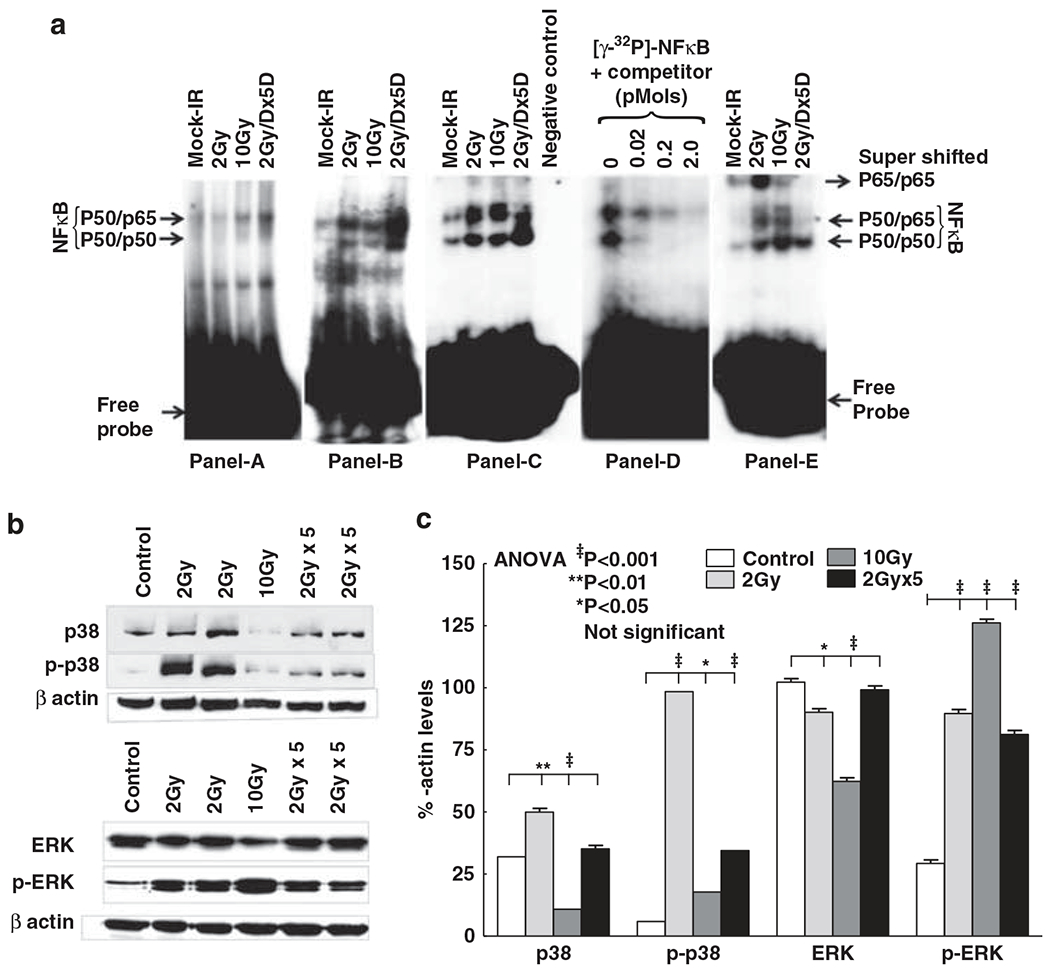
(a) Representative electrophoretic mobility shift assay (EMSA) autoradiograms (A–C) showing nuclear factor kappa B (NF-κB) DNA-binding activity in nontargeted heart tissues of mice exposed to 2 Gy, 10 Gy or fractionated ionizing radiation (FIR; 2 Gy per day for 5 days (2 Gy × 5D)). Specificity of NF-κB DNA-binding activity D). Nuclear protein was incubated in the absence (lane 1) or presence (lanes 2, 3 and 4) of increased concentrations of homologous unlabeled competitor and probed with [γ-32P]-ATP-labeled NF-κB-specific oligonucleotide. Identification of NF-κB subunits by supershift assay (E). The addition of antibodies directed against potential components of NF-κB complex resulted in supershift when antibody of p65 was used. (b) Immunoblots showing total and phosphorylated p38 and extracellular signal-regulated kinase-1/2 (ERK1/2) expression in nontargeted heart tissues of mice exposed to 2 Gy, 10 Gy or FIR (2 Gy × 5). (c) Histograms showing densitometry analysis of total and phosphorylated p38 and ERK1/2. The experiments were repeated at least three times and the group-wise comparisons were made using analysis of variance (ANOVA).
Radiation exerts p38 and ERK phosphorylation in nontargeted tissue
To substantiate that induced radioresponse in nontargeted tissues involves de novo synthesis of NF-κB, we investigated the activation of p38 and ERK1/2 that mediate both NF-κB activation and transactivation. The 2 Gy SDR exerted constitutive p38 increase and exhibited robust p38 phosphorylation in nontargeted (Figures 3b and c). Both 10 Gy SDR and 2 Gy FIR did not result in increased p38 levels. However, we found a significant increase in p38 phosphorylation after 10 Gy SDR and FIR. In addition, we observed a significant increase in ERK1/2 phosphorylation after all doses of radiation in nontargeted heart (Figures 3b and c). These data imply that distant organ radiation (both SDR and FIR) employs ERK1/2 signal transduction at least to an extent for NF-κB maintenance in nontargeted tissue. It is pertinent that 2 Gy SDR may additionally involve p38 signal transduction for NF-κB activation/maintenance.
Radiation increases p65 and cMYC in nontargeted tissue
To corroborate the onset/activation of NF-κB and the alteration of NF-κB downstream oncogenic cMYC in this setting, we analyzed the cellular expression of p65 and cMYC (Supplementary Figure 1A). Immunohistochemistry revealed that both SDR and FIR increased p65 in nontargeted heart with a maximal increase after FIR (Figure 4a). These data correlated well in accordance with our EMSA data. Furthermore, we observed a marked cMYC increase in nontargeted tissue after 2 Gy, 10 Gy SDR or 2 Gy FIR (Figure 4a).
Figure 4.
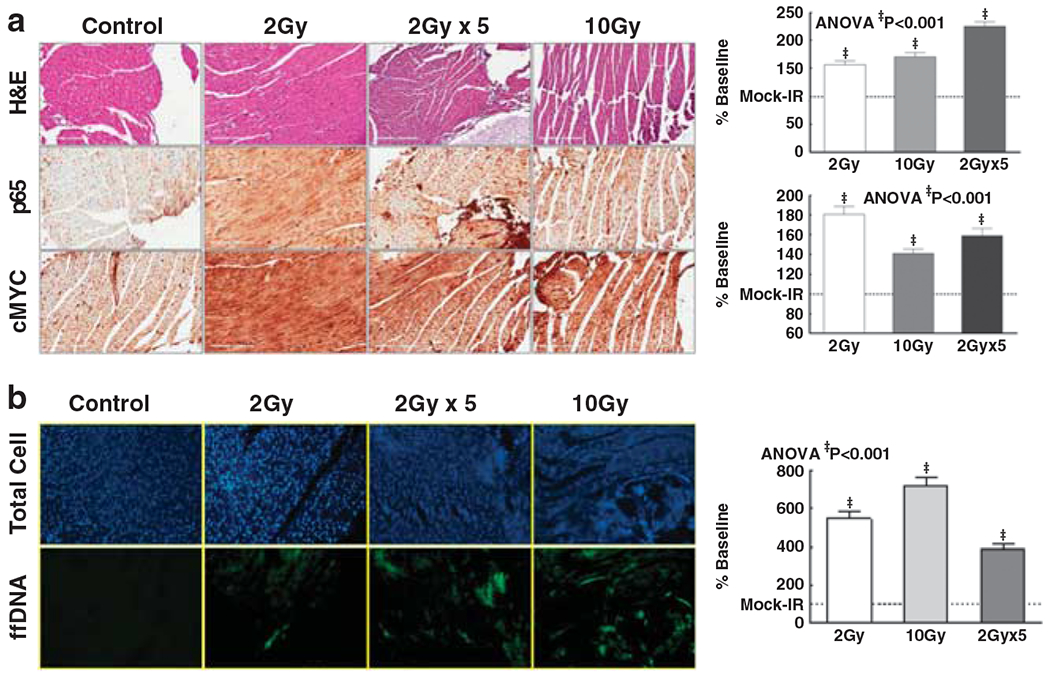
(a) Representative photomicrographs (× 40 magnifications) of hematoxylin and eosin (H&E) staining, p65 and cMYC levels in nontargeted heart in mice exposed to 2 Gy, 10 Gy or 2 Gy × 5 (2 Gy per day for 5 days). Histograms showing immunohistochemical-intensity of p65 and cMyc in nontargeted tissue. (b) Representative photomicrographs of total cell population (4’,6-diamidino-2-phenylindole (DAPI)) and DNA fragmentation in nontargeted heart in mice exposed to 2 Gy, 10 Gy or 2 Gy × 5. Each tissue section was analyzed for fluorescence intensity in at least 10 areas. Histograms showing fluorescein intensity in nontargeted heart tissue in mice exposed to varying doses of low-abdomen-focused radiation.
Radiation increases DNA damage in nontargeted tissue
The functional response on the nontargeted heart after radiation was assessed by measuring DNA damage (Supplementary Figure 1B). DNA fragmentation analysis revealed a significant (P<0.001) increase of DNA damage in nontargeted tissue after 2/10Gy SDR and FIR (Figure 4b). Relatively, distant-organ DNA fragmentation was robust after 10 Gy SDR.
DISCUSSION
Since the first measurement of clonogenic cell survival of HeLa cells exposed to X-rays in 1955,20 and up to the early 1990s, radiobiological research has mainly focused on cell death and survival with an approach from the target-cell hypothesis. It was believed that the direct killing of parenchymal and vascular endothelial cells successively led to organ failure.21 In addition, the time interval between exposure and manifestation of late normal tissue effects was considered to be ‘silent’, without any signs of tissue damage. For acute normal tissue damage, the target-cell hypothesis is still viable; however, it is not as reliable for modeling late normal tissue damage. The discovery of early radiation-induced cytokine cascades in 1995 was a major step toward new concepts in radiobiology.22 These findings, together with the identification of the bystander effect and studies of late side effects such as radiation-induced fibrosis, sequentially lead to a paradigm shift where the basal mechanisms of the cellular and molecular response to IR now receive much focus.22 Now, the cellular response to IR may be seen as an orchestrated reaction where cell loss and gene expression both play important roles.
The present study represents a multifaceted and extensive approach to investigate the onset and translation of signaling that effects radioresponse in nontargeted tissue. A broad range of absorbed doses (2 Gy/10 Gy SDR and 2 Gy × 5 FIR) were used in order to make the study valid for clinical practices. The results show that the nontargeted heart responds to distant radiation in an absorbed dose-independent manner but with few differentially expressed genes. The most striking inference was the onset of a large group of NF-κB pathway transcriptomes commonly across all radiation doses, which implies that circulating clastogenic factors after radiation initiate NF-κB signal transduction in nontargeted organs. To the best of our knowledge, no studies to date (in vivo, ex vivo or in vitro) have investigated the comprehensive, yet functionally specific, abscopal signaling response over SDR and FIR radiation exposure.
NF-κB, the transcriptional regulator involved in cellular responses to a variety of stimuli including radiation, is initiated by the signal-induced degradation of IκB proteins. It has been suggested that irradiated cells secrete soluble factors including interleukin-8, transforming growth factor-β, tumor necrosis factor-a and free radicals as bystander effect messengers.12 The increased secretion of these soluble factors after direct IR interaction triggers Ikka degradation-mediated NF-κB activation.23 To that end, recently we have demonstrated that the occurrence of NF-κB/tumor necrosis factor-α feedback in response to radiation persistently maintains radiation-induced NF-κB activation.12 New to science, the results of the current study demonstrate that radiation (both SDR and FIR) robustly activates NF-κB DNA-binding activity in abscopal out-of-field organ. Furthermore, activation of ERK1/2 or p38 mitogen-activated protein kinases observed in nontargeted tissue has been identified as key players in bystander responses. Global transcript profiling study confirmed the importance of NF-κB at the level of transcription, wherein genes regulated by NF-κB such as IL8, BCL2A1 and PTGS2 were expressed nearly equally in both bystander and irradiated cells.24 More importantly, the results revealed an increased DNA damage in nontargeted tissue after lower-abdomen-focused radiation. Pathophysiological aspects of radiation-associated distant organ response/function include systemic inflammatory response and consequences of cell loss because of radiation damage. Nonetheless, we observed a significant increase in NF-κB activation and signaling pathway transcription, and we also observed a consistent increase in DNA damage after 2 Gy, 10 Gy and 2 Gy × 5 distant radiation exposure. Although NF-κB is known to impede apoptosis, growing evidence suggests that it may also potentiate cell death depending upon the nature of the apoptotic stimulus. Indeed, it can be both an activator and repressor of its target genes depending upon the manner in which it is induced.25 Hence it becomes essential to understand the molecular mechanisms and variables involved in deciding how a particular cell type is going to counter NF-κB-mediated signaling. On the other hand, the results of the present study identified a profound increase in proto-oncogene cMyc in the nontargeted heart after both SDR and FIR radiation exposure. NF-κB has been showed to transactivate a number of genes pertaining to cell growth, carcinogenesis, proliferation and so on. With respect to carcinogenesis, its target genes include Myc along with another 158 genes that have NF-kB-binding sites in their promoters.26 A number of earlier studies have shown the increase of cMyc expression after radiation in target cells/tissues. However, to our knowledge, this is the first report showing the increased levels of cMyc in nontargeted organ after irradiation.
In summary, the present study for the first time delineates that the clinical doses of radiation significantly initiate the onset of NF-κB signal transduction and subsequent NF-κB activation in nontargeted distant organ. To that note, identifying the activation of a large subset (22 genes) of transcripts pertaining to NF-κB signaling pathway after all dose regimens of radiation signify its direct and definite involvement in flow-through molecular orchestration in nontargeted tissue. Moreover, this study affirms DNA damage and proto-oncogene (cMyc) expression in nontargeted heart tissue after radiation. However, the NF-κB-dependent downstream targeting and cause-effective functional responses including inflammation and damage in nontargeted tissue remain to be explored and are currently under investigation in our laboratory.
Supplementary Material
ACKNOWLEDGEMENTS
We were supported by Presbyterian Health Foundation, American Cancer Society (Grant ACS-IRG-05-066-01) and National Institutes of Health (NIH-COBRE-1P20GM103639-01) to N Aravindan, and the National Institutes of Health (Grant R01 CA112175) and US Department of Energy Grant DE-FG02-03ER63449 to M Natarajan. We acknowledge Dr Salahuddin Ahmad and Mr Daniel A Johnson, Division of Radiation Physics, Department of Radiation Oncology, OUHSC, for their help in radiation dosimetry studies.
Footnotes
CONFLICT OF INTEREST
The authors declare no conflict of interest.
REFERENCES
- 1.American Cancer Society. Cancer Facts & Figures 2012. Atlanta: American Cancer Society, 2012. [Google Scholar]
- 2.American Society of Radiation Oncology. Fast facts about radiation therapy. 2012. Fairfax, VA: American Society of Radiation Oncology (ASTRO). https://www.astro.org/News-and-Media/Media-Resources/FAQs/Fast-Facts-About-Radiation-Therapy/Index.aspx. [Google Scholar]
- 3.Berkey FJ. Managing the adverse effects of radiation therapy. Am Fam Physician. [Review] 2010; 82: 381–388394. [PubMed] [Google Scholar]
- 4.Tubiana M Can we reduce the incidence of second primary malignancies occurring after radiotherapy? A critical review. Radiother Oncol 2009; 91: 4–15, discussion 1–3. [DOI] [PubMed] [Google Scholar]
- 5.Mothersill C, Seymour CB. Cell-cell contact during gamma irradiation is not required to induce a bystander effect in normal human keratinocytes: evidence for release during irradiation of a signal controlling survival into the medium. Radiat Res 1998; 149: 256–262. [PubMed] [Google Scholar]
- 6.Sgouros G, Knox SJ, Joiner MC, Morgan WF, Kassis AI. MIRD continuing education: bystander and low dose-rate effects: are these relevant to radionuclide therapy? J Nucl Med 2007; 48: 1683–1691. [DOI] [PubMed] [Google Scholar]
- 7.Parsons WB Jr, Watkins CH, Pease GL, Childs DS Jr.. Changes in sternal marrow following roentgen-ray therapy to the spleen in chronic granulocytic leukemia. Cancer 1954; 7: 179–189. [DOI] [PubMed] [Google Scholar]
- 8.Marozik P, Mothersill C, Seymour CB, Mosse I, Melnov S. Bystander effects induced by serum from survivors of the Chernobyl accident. Exp Hematol 2007; 35(4 Suppl 1): 55–63. [DOI] [PubMed] [Google Scholar]
- 9.Mothersill C, Smith RW, Agnihotri N, Seymour CB. Characterization of a radiation-induced stress response communicated in vivo between zebrafish. Environ Sci Technol 2007; 41: 3382–3387. [DOI] [PubMed] [Google Scholar]
- 10.Emerit I, Oganesian N, Sarkisian T, Arutyunyan R, Pogosian A, Asrian K et al. Clastogenic factors in the plasma of Chernobyl accident recovery workers: anticlastogenic effect of Ginkgo biloba extract. Radiat Res 1995; 144: 198–205. [PubMed] [Google Scholar]
- 11.Koturbash I, Loree J, Kutanzi K, Koganow C, Pogribny I, Kovalchuk O. In vivo bystander effect: cranial X-irradiation leads to elevated DNA damage, altered cellular proliferation and apoptosis, and increased p53 levels in shielded spleen. Int J Radiat Oncol Biol Phys 2008; 70: 554–562. [DOI] [PubMed] [Google Scholar]
- 12.Veeraraghavan J, Natarajan M, Aravindan S, Herman TS, Aravindan N. Radiation-triggered tumor necrosis factor (TNF) alpha-NFkappaB cross-signaling favors survival advantage in human neuroblastoma cells. J Biol Chem 2011; 286: 21588–21600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Aravindan N, Aravindan S, Riedel BJ, Weng HR, Shaw AD. Furosemide prevents apoptosis and associated gene expression in a rat model of surgical ischemic acute renal failure. Ren Fail 2007; 29: 399–407. [DOI] [PubMed] [Google Scholar]
- 14.Aravindan N, Aravindan S, Shanmugasundaram K, Shaw AD. Periods of systemic partial hypoxia induces apoptosis and inflammation in rat skeletal muscle. Mol Cell Biochem 2007; 302: 51–58. [DOI] [PubMed] [Google Scholar]
- 15.Aravindan N,Cata JP, Dougherty PM, Shaw AD. Effect off enoldopamon ischemia/reperfusion-induced apoptosis. Ren Fail 2006; 28: 337–344. [DOI] [PubMed] [Google Scholar]
- 16.Aravindan N, Cata JP, Hoffman L, Dougherty PM, Riedel BJ, Price KJ et al. Effects of isoflurane, pentobarbital, and urethane on apoptosis and apoptotic signal transduction in rat kidney. Acta Anaesthesiol Scand 2006; 50: 1229–1237. [DOI] [PubMed] [Google Scholar]
- 17.Aravindan N, Natarajan M, Shaw AD. Fenoldopam inhibits nuclear translocation of nuclear factor kappa B in a rat model of surgical ischemic acute renal failure. J Cardiothorac Vasc Anesth 2006; 20: 179–186. [DOI] [PubMed] [Google Scholar]
- 18.Veeraraghavan J, Natarajan M, Herman TS, Aravindan N. Low-dose gamma-radiation-induced oxidative stress response in mouse brain and gut: regulation by NFkappaB-MnSOD cross-signaling. Mutat Res 2011; 718: 44–55. [DOI] [PubMed] [Google Scholar]
- 19.Natarajan M, Aravindan N, Meltz ML, Herman TS. Post-translational modification of I-kappa B alpha activates NF-kappa B in human monocytes exposed to 56Fe ions. Radiat Environ Biophys 2002; 41: 139–144. [DOI] [PubMed] [Google Scholar]
- 20.Puck TT, Marcus PI. A rapid method for viable cell titration and clone production with Hela cells in tissue culture: the use of X-irradiated cells to supply conditioning factors. Proc Natl Acad Sci USA 1955; 41: 432–437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Stewart FA, Dorr W. Milestones in normal tissue radiation biology over the past 50 years: from clonogenic cell survival to cytokine networks and back to stem cell recovery. Int J Radiat Biol. [Review] 2009; 85: 574–586. [DOI] [PubMed] [Google Scholar]
- 22.Rubin P, Johnston CJ, Williams JP, McDonald S, Finkelstein JN. A perpetual cascade of cytokines postirradiation leads to pulmonary fibrosis. Int J Radiat Oncol Biol Phys 1995; 33: 99–109. [DOI] [PubMed] [Google Scholar]
- 23.Lee SJ, Dimtchev A, Lavin MF, Dritschilo A, Jung M. A novel ionizing radiation-induced signaling pathway that activates the transcription factor NF-kappaB. Oncogene 1998; 17: 1821–1826. [DOI] [PubMed] [Google Scholar]
- 24.Ghandhi SA, Yaghoubian B, Amundson SA. Global gene expression analyses of bystander and alpha particle irradiated normal human lung fibroblasts: synchronous and differential responses. BMC Med Genomics 2008; 1 : 63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kaltschmidt B, Kaltschmidt C, Hofmann TG, Hehner SP, Droge W, Schmitz ML. The pro- or anti-apoptotic function of NF-kappaB is determined by the nature of the apoptotic stimulus. Eur J Biochem 2000; 267: 3828–3835. [DOI] [PubMed] [Google Scholar]
- 26.Pahl HL. Activators and target genes of Rel/NF-kappaB transcription factors. Oncogene 1999; 18: 6853–6866. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.