

Abstract
Time-of-day dependent fluctuations in exercise performance have been documented across different sports and seem to affect both endurance and resistance modes of exercise. Most of the studies published to date have shown that the performance in short-duration maximal exercises (i.e. less than 1 min - e.g. sprints, jumps, isometric contractions) exhibits diurnal fluctuations, peaking between 16:00 and 20:00 h. However, the time-of-day effects on short duration exercise performance may be minimized by the following factors: (1) short exposures to moderately warm and humid environments; (2) active warm-up protocols; (3) intermittent fasting conditions; (4) warming-up while listening to music; or (5) prolonged periods of training at a specific time of day. This suggests that short-duration maximal exercise performance throughout the day is controlled not only by body temperature, hormone levels, motivation and mood state but also by a versatile circadian system within skeletal muscle. The time of day at which short-duration maximal exercise is conducted represents an important variable for training prescription. However, the literature available to date lacks a specific review on this subject. Therefore, the present review aims to (1) elucidate time-of-day specific effects on short-duration maximal exercise performance and (2) discuss strategies to promote better performance in short-duration maximal exercises at different times of the day.
Subject terms: Physiology, Health care
Introduction
Circadian rhythms are responsible for temporal regulation of numerous physiological phenomena in the human body. At a molecular level, circadian rhythms are defined as a function of clock gene expression levels over a 24-hour period1. Clock gene expression levels throughout the day set up the pace of sleep-wake cycles, hunger, hormone production, body temperature, as well as other physiological functions2. In mammals, biological rhythms are driven by a central pacemaker located in the suprachiasmatic nuclei (SCN) of the hypothalamus. It consists of approximately 20 000 neurons that exhibit independent rhythms of firing rate and gene expression3. The core clock system of the SCN works as a self-sustained transcriptional/translational feedback loop, involving a set of four integral proteins that act as activators or repressors within the system4. In addition, there is a group of kinases and phosphatases that regulate their localization and stability5. Apart from the core clock system in the SCN, circadian clocks and clock-controlled output genes (CCGs) are also present in peripheral tissues such as the liver, heart, kidney, pancreas, adipose tissue and skeletal muscle6–12. In this sense, the main synchronizer of the internal clock is solar light13, but it is known now that other non-photic stimuli such as feeding, social contact or physical exercise do also modulate the transcriptional activity of clock genes14–17. Likewise, internal clocks also play important roles on human behavior, accounting for variations in resting levels of neuromuscular, sensory-motor and cognitive performance throughout the day18–21. Recently, high-throughput transcriptomic and metabolomic analyses in mice have demonstrated that the time of day is a crucial factor to amplify the effect of exercise on systemic energy homeostasis and metabolic pathways within skeletal muscle22. In addition, exercise capacity exhibit diurnal fluctuations in mice and humans between the early and late part of their active phase23, which suggests that the time of day is a major modifier of exercise capacity.
Time-of-day dependent fluctuations in exercise performance have been documented across different sports and seem to affect both endurance and resistance modes of exercise24–30. Whereas differences in endurance exercise performance might be as large as 26%25, strength performance may vary as much as 41% throughout the day30. Short-duration maximal exercises (i.e. less than 1 min - e.g. all-out sprints, maximal jumps or isometric contractions) comprise both modes of exercise and are characterized by higher locomotor requirements31 and anaerobic energy contributions32 than longer exercises. Thus, they represent a robust model to explore time-of-day effects on the musculoskeletal system. Previous studies have shown that performance in short-duration maximal exercises exhibits time-of-day dependent fluctuations with amplitudes up to 29.4% between the morning and evening hours24,33–45. Considering the large variation in short-duration maximal exercise performance throughout the day, it is clear that the time of day at which our athletes train or compete is not trivial. However, the literature available to date lacks a specific review on this subject. Therefore, the present review aims to (1) elucidate time-of-day specific effects on short-duration maximal exercise performance and (2) discuss strategies to promote better performance in short-duration maximal exercises at different times of the day.
Methods
The present article provides an up-to-date review of the literature about the effects of time of day on short-duration maximal exercise performance. Articles were searched via three online databases (PubMed, PubMed Central and Google Scholar; 1924–2019). The literature search strategy included a combination of free terms using the Boolean operators “AND” and “OR”. The free terms used in the search were: time-of-day, circadian, chronotype, strength, neuromuscular, resistance, endurance, aerobic, anaerobic, short-duration, performance, isometric, isokinetic, dynamic, morning, evening, afternoon and night. The full search term strategy that was used for each scientific database is outlined in the supplementary material.
The studies retrieved from the databases that fit the inclusion criteria were imported to EndNote Web Software (Thomson Reuters, New York, USA) where duplicated articles were identified and excluded. Then, the titles and abstracts of the remaining articles were reviewed. Articles were excluded at this initial screening phase if they did not mention any specific effect of time of day on exercise performance. If this information could not be ascertained from the title or abstract, the article’s full text was reviewed in the next screening phase to determine whether it fitted the eligibility criteria. After the initial title/abstract screening process, the full texts of all the remaining articles were assessed to select those that fitted the inclusion criteria. The inclusion criteria that were applied to original articles included: (1) the study explored the effects of time of day (over a 24-hour period) on one or more aspects of exercise performance; (2) the study assessed time-of-day effects on isometric, isokinetic and dynamic short-duration maximal (i.e. less than 1 min) exercises (3) the study assessed the effects of either acute or chronic interventions on time-of-day dependent fluctuations in exercise performance; (4) the study assessed either direct (i.e. peak and mean power, isokinetic peak torque, total work, jump height) or indirect (i.e. RPE, neuromuscular efficiency, markers or muscle injury) features of exercise performance; (5) the study was published in indexed journals and was available in English. As a result, a total of 66 original articles were selected out of 545 initial results. A flow diagram of the article selection process is also outlined in the Fig. 1.
Figure 1.
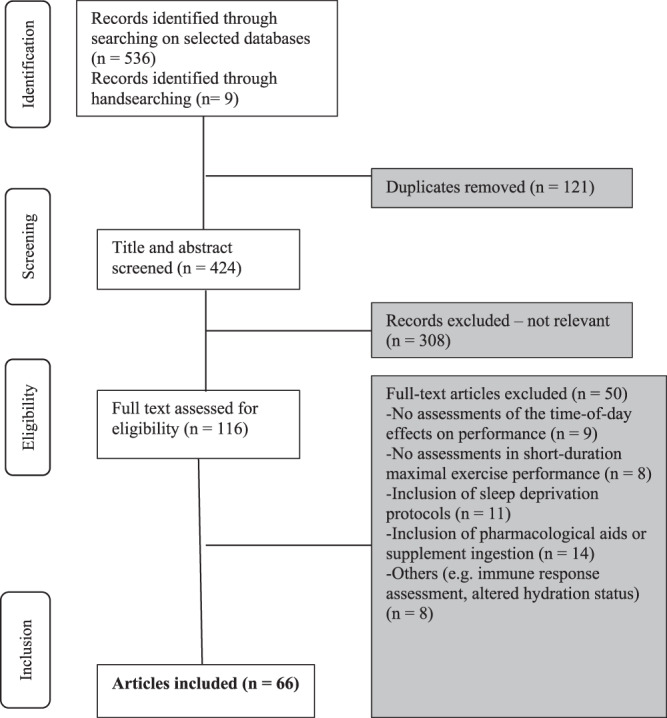
Flow diagram of the article selection process.
Results
A total of 66 articles were selected and then divided into dynamic and/or isometric/isokinetic short-duration maximal exercises. Forty-four articles assessed time-of-day effects on dynamic short-duration maximal exercise performance, including swimming, tennis, jumping, cycling, sprinting and resistance exercises (Table 1). On the other hand, 32 articles assessed time-of-day effects on isometric and isokinetic exercise performance, including electrically induced contractions, reflex contractions, maximal and submaximal voluntary isometric contractions, isokinetic leg contractions and magnetic stimulation of the cortex (Table 2). Most articles compared time-of-day effects on short-duration maximal exercise performance under acute exercise conditions; however, 11 articles assessed time-of-day effects on short-duration maximal exercise performance after a period of training (from 5 to 10 weeks) (Tables 1 and 2).
Table 1.
Summary of the literature about time-of-day effects on dynamic short-duration maximal exercise performance.
| Author(s) | Date | Protocol | Training period | Number of participants | Characteristics of the participants | Main Results (% change of peak performance time vs. other assessment time-points) | Time-of-day effect? | Peak performance time | Assessment Time-points |
|---|---|---|---|---|---|---|---|---|---|
| Aloui et al.85 | 2013 | Repeated Sprint Ability1 test | — | n = 12 males | Recreationally trained soccer players |
↑ 5.4% in Ppeak during the first sprint ↔ time-of-day dependent variations in Ppeak after 2 and 4 weeks of intermittent-fasting3 conditions |
Yes | 17:00–19:00 h | 07:00–09:00 and 17:00–19:00 h |
| Arnett47 | 2002 | All-out swim trials | — |
n = 6 males n = 4 females |
Competitive young swimmers |
↑ 0.9% in all-out (100-yd) swimming performance ↔ time-of-day variation in body temperature, but not evening superiority in all-out (100-yd) swimming performance when high-volume4 warm up conditions were imposed |
Yes | 17:00 h | 06:15 and 17:00 h |
| Atkinson and Speirs37 | 1998 | Tennis services | — |
n = 3 males n = 3 females |
Competitive young adult tennis players |
↑ 3.9% in first serves speed (but lower serve accuracy) ↔ second serves speed (nor serve accuracy) throughout the day |
Yes | 18:00 h | 09:00, 14:00 and 18:00 h |
| Baxter and Reilly48 | 1983 | All-out swim trials | — |
n = 4 males n = 10 females |
Competitive young swimmers | ↑ 3.5% in 100 m swimming performance | Yes | 17:00 h | 06:30, 09:00, 13:30, 17:00 and 22:00 h |
| Belkhir et al.86 | 2019 | 5-m shuttle run test | — | n = 12 males | Competitive soccer players |
↑ 3.6% in total distance and 13.1% in higher distance during the 5-m shuttle run test under warm-up without music conditions ↑ 3.5–6.9% in anaerobic performance at 07:00 h under neutral and self-selected music warm-up conditions ↑ 3.6–5.3% in anaerobic performance at 17:00 h under neutral and self-selected music warm-up conditions |
Yes | 17:00 h | 07:00 and 17:00 h |
| Bernard et al.38 | 1998 | Force-Velocity and Multi-Jump tests | — | n = 23 males | Physical Education students |
↑ 3.5% in maximal anaerobic power for cycling throughout the day ↑ 5–6.6% in maximal anaerobic power for jumping throughout the day |
Yes | 14:00 and 18:00 h | 09:00, 14:00 and 18:00 h |
| Blonc et al.46 | 2010 | Squat Jump, Countermovement Vertical Jump and Cycle sprints2 tests | — |
n = 12 males n = 4 females |
Physical Education students | ↔ SJ, CMJ and Cycle sprints performances throughout the day | No | — | 07:00–09:00 and 17:00–19:00 h |
| Castaingts et al.55 | 2004 | Drop Jump test | — | n = 11 males | N/A |
↑ 10.9% in jump height and 15.7% in NME (mechanical performances/EMG recordings) ↔ reflex excitability throughout the day |
Yes | 18:00 h | 06:00 and 18:00 h |
| Chtourou et al.13 | 2012 | Wingate anaerobic test | — | n = 12 males | Physical Education students |
↑ 5.3% in Ppeak and 3.3% in Pmean ↓ 48.9% time-of-day dependent variations in Ppeak after a music-coupled warm-up5 ↔ time-of-day dependent variations in Pmean after a music-coupled warm-up5 |
Yes | 17:00 h | 07:00 and 17:00 h |
| Chtourou et al.30 | 2012 | Wingate anaerobic test, Squat Jump and Countermovement Vertical Jump tests | 12 weeks of training + 2 weeks of tapering at a specific time of day | n = 31 males | Physical Education students |
↑ 3.3% in Ppeak and 3% in Pmean before the training period ↑ 7.2% in SJ and 5.9% in CMJ heights before the training period ↔ SJ, CMJ and Wingate test performance throughout the day after 12 weeks of training at a specific time of day ↑ 4–7.3% in anaerobic performance at 07:00 h after 12 weeks of training at a specific time of day ↑ 4–5.3% in anaerobic performance at 17:00 h after 12 weeks of training at a specific time of day |
Yes | 17:00 h | 07:00 and 17:00 h |
| Chtourou et al.50 | 2012 | Wingate anaerobic test | — | n = 10 males | Competitive young football players |
↑ 3.14% in Ppeak and 2.9% in Pmean ↔ time-of-day dependent variations in Ppeak and Pmean after 2 and 4 weeks of intermittent-fasting3 conditions |
Yes | 17:00 h | 07:00 and 17:00 h |
| Chtourou et al.24 | 2011 | Wingate anaerobic test | — | n = 22 males | Physical Education students |
↑ 2.6% in Ppeak and 2.3% in Pmean ↑ 4% in NME during the first (firsts 20 s) but not second (lasts 10 s) of the Wingate anaerobic test. |
Yes | 17:00 h | 07:00 and 17:00 h |
| Giacomoni, Billaut and Falgairette33 | 2006 | Repeated Sprint Ability6 test | — | n = 12 males | Physically active and healthy adults | ↔ biomechanical indices of neuromuscular performance (Ppeak, total mechanical work, peak pedaling rate) throughout the day | No | — | 08:00–10:00 and 17:00–19:00 h |
| Hammouda et al.58 | 2011 | Repeated Sprint Ability1 test | — | n = 12 males | Well-trained young football players |
↑ 5.4% in sprint 1 and 4.4% in sprint 2 Ppeak ↑ 1.6% in Ppeak sprint decrease ↑ 0.7% in RPE ↑ 12.3% in resting homocysteine levels and 17.6–35.4% in biological markers of muscle injury at 17:00–18:30 h ↓ 4.4–12.9% in biomarkers of antioxidant status’ resting levels at 17:00–18:30 h |
Yes | 17:00–18:30 h | 07:00–08:30 h and 17:00–18:30 h |
| Hammouda et al.27 | 2012 | Wingate anaerobic test | — | n = 15 males | Well-trained young football players | ↑ 3.6% in Ppeak and 2.8% in Pmean | Yes | 17:00–18:30 h | 07:00–08:30 h and 17:00–18:30 h |
| Hill et al.51 | 1992 | All-out cycle ergometer test | — |
n = 8 males n = 6 females |
N/A |
↑ 9.6% in total work in the afternoon compared to the morning ↑ 5.1% in anaerobic power in the afternoon compared to the morning |
Yes | N/A | N/A |
| Javierre et al.67 | 1996 | 80 m all-out sprint performance | — | n = 8 males | Competitive sprinters |
↑ sprint performance (faster sprint times) at 19:00 h ↑ 2.7–4.1% in sprint performance at 17:00 h when sleep/wake cycles and mealtimes were advanced for two hours (vs sprint performance at 17:00 h on a control day) ↑ 1.7–2.3% in sprint performance at 21:00 h when sleep/wake cycles and mealtimes were delayed for two hours (vs sprint performance at 21:00 h on a control day) |
Yes | 19:00 h | 09:00, 11:00, 13:00, 15:00, 17:00, 19:00, 21:00 and 23:00 h |
| Krčmárová et al.63 | 2018 | Dynamic leg press and seated-row 6-repetition maximum (6RM) and functional capacity7 tests | 12 weeks of training at a specific time of day | n = 31 females | Untrained healthy older adults | ↔ strength performance with time-of-day after 12 weeks of training at a specific time of day | No | — | 07:30 and 18:00 h |
| Küüsmaa et al.64 | 2016 | Dynamic leg press test of 1-repetition maximum (1RM) | 24 weeks of training at a specific time of day | n = 42 males | Untrained healthy youngsters | ↔ 1RM gains after 24 weeks of training at a specific time of day | No | — | 06:30–09:30 and 16:30–19:30 h |
| Lericollais et al.28 | 2009 | Wingate anaerobic test | — | n = 16 males | Competitive cyclists | ↑ 8.2% in Ppeak and 7.8% in Pmean | Yes | 18:00 h | 06:00 and 18:00 h |
| Lericollais et al.52 | 2011 | Wingate anaerobic test | — | n = 20 males | Physically active and healthy adults | ↑ 6.6% in Ppeak, 3.3% in Pmean30s and 2.7% in Pmean60s | Yes | 18:00 h | 06:00 and 18:00 h |
| López-Samanes et al.40 | 2017 | Serve velocity/accuracy, Countermovement Vertical Jump, Agility T-test8 and 10-m sprint tests | — | n = 13 males | Competitive tennis players |
↑ 4% in serve velocity/accuracy test ↑ 4.5% in CMJ ↑ 2.1% in Agility T-test ↑ 2.7% in 10-m RUN performances |
Yes | 16:30 h | 09:00 and 16:30 h |
| Melhim53 | 1993 | Wingate anaerobic test | — | n = 13 females | Physical Education students | ↑ 7% in Ppeak and 15–16% in Pmean | Yes | 15:00 and 21:00 h | 03:00, 09:00, 15:00 and 21:00 h |
| Pallarés et al.49 | 2014 | Bench Press, Countermovement Vertical Jump, Crank-arm (10 s Wingate test), and 25 m swimming freestyle tests | — |
n = 6 males n = 6 females |
Well-trained junior swimmers |
↑ 5.1% in bench press maximum strength and muscle power ↑ 1.7% in 25-m swimming performance ↑ 5.8% in CMJ height |
Yes | 18:00 h | 10:00 and 18:00 h |
| Pullinger et al.87 | 2019 | Handgrip strength, Bench Press and Back Squat tests | — | n = 10 males | Resistance trained young adults |
↑ 4.6% in right and. 6.5% in left handgrip strength ↑ 3.3% in bench press and 2.6% in back squat average force ↑ 13.6% in bench press and 11.5% in back squat peak velocity ↓ 17% in bench press and 14.1% in back squat time to peak velocity |
Yes | 17:30 h | 07:00 and 17:30 h |
| Pullinger et al.89 | 2018 | All-out sprint9 test | — | n = 12 males | Resistance trained young adults | ↑ 7.8–8.3% in total sprint distance, average mean and average velocity | Yes | 17:30 h | 07:00 and 17:30 h |
| Racinais, Blonc and Hue59 | 2005 | All-out sprint2 test | — | n = 8 males | Physical Education students |
↑ 4.5% in Pmax and 3.8% in Fmax ↑ 3.7% in Pmax and 3.1% in Vmax after active warm-up11 vs. passive12 warm-up conditions at any time of day |
Yes | 17:00–19:00 h | 07:00–09:00 and 17:00–19:00 h |
| Racinais, Hue and Blonc56 | 2004 | Squat-Jump, Countermovement Vertical Jump and all-out sprint2 tests | — | n = 12 males | Physical Education students |
↑ 3.1–3.7% in CMJ, SJ and cycling sprint performances ↔ muscular performance throughout the day when moderately warm and humid13 conditions were imposed |
Yes | 17:00–19:00 h | 07:00–09:00 and 17:00–19:00 h |
| Racinais et al.62 | 2004 | Countermovement Vertical Jump and Force-Velocity tests | — |
n = 15 males n = 8 females |
Physical Education students | ↔ maximal anaerobic power throughout the day when warm and humid14 conditions were imposed | No | — | 08:00, 13:00 and 17:00 h |
| Racinais et al.61 | 2005 | Repeated Sprint Ability1 test | — | n = 9 males | Physical Education students |
↑ 5.3% in Ppeak during the first sprint ↑ 12% in blood lactate concentration at the end of the RSA test |
Yes | 17:00–19:00 h | 07:00–09:00 and 17:00–19:00 h |
| Racinais et al.2 | 2006 | Countermovement Vertical Jumps, all-out2, and Isokinetic leg strength (knee flexors and extensors) tests | — | n = 9 males | Physically active and healthy adults | ↔ CMJ, sprint and isokinetic torque performances throughout the day when moderately warm and humid15 conditions were imposed | No | — | 07:00–09:00 and 17:00–19:00 h |
| Racinais et al.60 | 2009 | All-out sprint2 test | — | n = 7 males | Physical Education students |
↑ 12% in Pmax, 5% in Fmax, and 6% in Vmax ↔ Pmax, Fmax, and Vmax throughout the day when local pre-cooling16 or -heating17 conditions were imposed |
Yes | 17:00–19:00 h | 07:00–09:00 and 17:00–19:00 h |
| Robertson et al.88 | 2018 | Bench Press and Back Squat tests | — | n = 30 males | Resistance trained young adults |
↑ 1.9% in bench press and 2.5% in back squat average force ↑ 8.3% in bench press and 12.7% in back squat peak velocity ↓ 16.6% in bench press and 9.8% in back squat time to peak velocity |
Yes | 17:30 h | 07:00 and 17:30 h |
| Sedliak et al.41 | 2008 | Squat-Jump test | — | n = 32 males | Physically active and healthy adults10 | ↑ 4.6–5.6% in power output during the concentric phase of loaded SJ | Yes | 12:00–13:00, 17:00–18:00 and 20:30–21:30 h | 07:00–08:00, 12:00–13:00, 17:00–18:00 and 20:30–21:30 h |
| Sedliak et al.66 | 2008 | One-repetition maximum (1RM) half-squat test | 10 weeks of training at a specific time of day | n = 34 males | Physically active and healthy adults10 | ↔ Half-Squat 1RM peak torque after 10 weeks of training at a specific time of day | No | — | 09:00–16:00 h |
| Sedliak et al.65 | 2009 | One-repetition maximum (1RM) half-squat test | 10 weeks of training at a specific time of day | n = 24 males | Physically active and healthy10 | ↔ Half-Squat 1RM peak torque after 10 weeks of training at a specific time of day | No | — | 09:00–16:00 h |
| Souissi et al.35 | 2010 | Squat-Jump, Five-Jump and Wingate anaerobic tests | — | n = 20 males | Untrained healthy boys |
↑ 3.5% in SJ and 5% in Five-Jump test performances ↑ 5.5% in Ppeak and 6% in Pmean during the Wingate anaerobic test |
Yes | 14:00 and 18:00 h | 08:00, 14:00 and 18:00 h |
| Souissi et al.36 | 2012 | Squat-Jump, Countermovement Vertical Jump and Wingate anaerobic tests | 6 weeks of training at a specific time of day | n = 24 males | Untrained healthy boys |
↑ 11.5% in SJ and 10.7% in CMJ performances before the training period ↑ 6.3% in Ppeak and 5% in Pmean during the Wingate anaerobic test before the training period ↔ SJ, CMJ, Ppeak and Pmean throughout the day in the morning-training group after 6 weeks of training at a specific time of day |
Yes | 17:00 h | 07:00–08:00 and 17:00–18:00 h |
| Souissi et al.42 | 2010 | Wingate anaerobic tests | — | n = 12 males | Physical Education students |
↑ 6.8% in Ppeak and 4.1% in Pmean after 5-min active warm up conditions ↑ 3.7% in Ppeak and 1.6% in Pmean after 15-min active warm up conditions ↓ 45.5% time-of-day differences in Ppeak and 60.9% Pmean in the morning after longer (15-min) than shorter (5-min) active warm-ups |
Yes | 18:00 h | 08:00 and 18:00 h |
| Souissi et al.54 | 2002 | Wingate anaerobic test | 6 weeks of training at a specific time of day | n = 14 males | Physical Education students |
↑ 8.4% and 4.1% in Ppeak in the morning- and evening-training groups before the training period ↔ Ppeak throughout the day in the morning-training group after 6 weeks of training at a specific time of day |
Yes | 17:00–18:00 h | 07:00–08:00 and 17:00–18:00 h |
| Souissi et al.43 | 2004 | Force-Velocity and Wingate anaerobic tests | — | n = 19 males | Physical Education students | ↑ 7% in Pmax, 7.6% in Ppeak and 11.3% in Pmean | Yes | 17:10–18:00 h* | 02:00, 06:00, 10:00, 14:00, 18:00 and 22:00 h |
| Taylor et al.57 | 2011 | Countermovement Vertical Jump tests | — | n = 8 males | Recreationally trained adults18 |
↑ 6.4% in CMJ height (cm) ↔ time-of-day dependent variations in CMJ performance when extended warm-up19 conditions were imposed in the morning |
Yes | 16:00 h | 08:00 and 16:00 h |
| West et al.90 | 2014 | Countermovement Vertical Jump tests | — | n = 16 | Elite rugby union seven players | ↑ 3.1% in CMJ Ppeak | Yes | 17:00 h | 10:00 and 17:00 h |
| Zarrouk et al.44 | 2012 | Repeated Sprint Ability1 test | — | n = 12 males | Physical Education students |
↑ Total work, Ppeak and %Ppeak decrement during the first 3 sprints ↔ EMG throughout the day |
Yes | 18:00 h | 06:00 and 18:00 h |
RPE rating of perceived exertion, Pmax Maximal Power, Ppeak Peak Power, Pmean Mean Power, Fmax Maximal Force, Vmax Maximal Velocity, CMJ countermovement Vertical jump, SJ Squat Jump, MCV Maximal Voluntary Isometric Contraction, NME Neuromuscular efficiency, EMG Electromyographic activity, N/A Not available, 1 i.e. 5 × [6 s of maximal cycling sprint + 24 s of rest], 2 i.e. 3 × [7 s of maximal cycling sprint + 5 min of rest], 3 i.e. 15–16 h starvation/day; from ≈ 04:00 h till ≈ 19:00 h, 4 i.e. 200% of standard warm-up times, 5 i.e. 10-min warm up while listening high tempo music (>120 to 140 bpm) through headphones.
RPE rating of perceived exertion, Pmax Maximal Power, Ppeak Peak Power, Pmean Mean Power, Pmean30s Mean Power at 30s, Pmean60s Mean Power at 60s, Fmax Maximal Force, Vmax Maximal Velocity, CMJ countermovement Vertical jump, 10-m RUN 10-m sprint, SJ Squat Jump, MCV Maximal Voluntary Isometric Contraction, NME Neuromuscular efficiency, EMG Electromyographic activity, N/A Not available, 1 i.e. 5 × [6 s of maximal cycling sprint + 24 s of rest], 6 i.e. 10 ×[6 s of maximal cycling sprint + 30 s of rest]), 3 i.e. 15–16 h starvation/day; from ≈ 04:00 h till ≈ 19:00 h.
RPE rating of perceived exertion, Pmax Maximal Power, Ppeak Peak Power, Pmean Mean Power, Pmean30s Mean Power at 30s, Pmean60s Mean Power at 60s, Fmax Maximal Force, Vmax Maximal Velocity, CMJ countermovement Vertical jump, 10-m RUN 10-m sprint, SJ Squat Jump, MCV Maximal Voluntary Isometric Contraction, NME Neuromuscular efficiency, EMG Electromyographic activity, 7 i.e. 30-second chair stands and arm curl test, Timed Up and Go, 8 i.e. all-out running test with lateral and front-to-back movements, 9 i.e. 3 ×[3 s of maximal running sprint + 30 s of rest].
RPE rating of perceived exertion, Pmax Maximal Power, Ppeak Peak Power, Pmean Mean Power, Fmax Maximal Force, Vmax Maximal Velocity, CMJ countermovement Vertical jump, SJ Squat Jump, MCV Maximal Voluntary Isometric Contraction, NME Neuromuscular efficiency, EMG Electromyographic activity, 1 i.e. 5 ×[6 s of maximal cycling sprint + 24 s of rest], 2 i.e.3 × [7 s of maximal cycling sprint + 5 min of rest], 10 i.e. no medication within the last14 days, a non-smoker, regular sleep pattern with sleep duration ranging from 6 to 9 h per night and regular physical activity not more than once a week, 11 i.e. 12 min of pedaling at 50% of V˙O2max interspersed with three brief accelerations of 5 s, 12 i.e. 3 min of pedaling at 70 rpm at 50% of V˙O2max, 13. i.e. 60 min of resting in a room at 29 °C, 70% relative humidity, 14 i.e. 60 min of resting in a room at 28.1 °C, 62.6% relative humidity, 15 i.e. 60 min of resting in a room at 24 °C, 70% relative humidity, 16 i.e. 30 min exposure to a cold bath at 16 °C, 17 i.e. 30 min exposure to a bath at 38 °C.
RPE rating of perceived exertion, Pmax Maximal Power, Ppeak Peak Power, Pmean Mean Power, Fmax Maximal Force, Vmax Maximal Velocity, CMJ countermovement Vertical jump, SJ Squat Jump, MCV Maximal Voluntary Isometric Contraction, NME Neuromuscular efficiency, EMG Electromyographic activity, 1 i.e. 5 ×[6 s of maximal cycling sprint + 24 s of rest], 10 i.e. no medication within the last14 days, a non-smoker, regular sleep pattern with sleep duration ranging from 6 to 9 h per night and regular physical activity not more than once a week,18 i.e. a minimum of 6 months resistance training history 19 i.e. 20 min general warm-up on a stationary bike at 150–200 W prior to completion of the control warm-up, resulting in a whole-body temperature increase of 0.3 ± 0.2 °C, *estimated acrophase by cosinor analysis.
Table 2.
Summary of the literature about time-of-day effects on isometric and isokinetic exercise performance.
| Author(s) | Date | Protocol | Training period | Number of participants | Characteristics of the participants | Main Results (% change of peak performance time vs. other assessment time-points) | Time-of-day effect? | Peak performance time | Assessment Time-points |
|---|---|---|---|---|---|---|---|---|---|
| Aloui et al.85 | 2013 | Maximal voluntary isometric knee extensions | — | n = 12 males | Recreationally trained soccer players |
↑ 8.9% in MVC peak torque ↔ time-of-day dependent variations in MVC peak torque after 2 and 4 weeks of intermittent-fasting2 conditions |
Yes | 17:00–19:00 h | 07:00–09:00 and 17:00–19:00 h |
| Callard et al.80 | 2000 | Maximal voluntary isometric knee extensions | — | n = 6 males | Competitive cyclists |
↑ 6% in MVC peak torque under resting conditions ↑ 7.8% in MVC peak torque under 24-h cycling3 conditions ↑ EMG activity at 13:00, 17:00 and 21:00 h |
Yes | 19:10–19:30 h* | 00:01, 05:00, 09:00, 13:00, 17:00 and 21:00 h |
| Castaingts et al.55 | 2004 | Electrically induced, reflex and maximal and submaximal voluntary isometric contractions | — | n = 11 males | N/A |
↑ 17.6% in NME (in electrically induced contraction conditions) ↔ NME (in maximal voluntary isometric contraction conditions) throughout the day ↔ Reflex excitability throughout the day |
Yes | 18:00 h | 06:00 and 18:00 h |
| Chtourou et al.30 | 2012 | Maximal voluntary isometric knee extensions | 12 weeks of training + 2 weeks of tapering at a specific time of day | n = 31males | Physical Education students |
↑ 10.8% in MVC peak torque before the training period ↔ MVC peak torque throughout the day in the morning-training group after 12 weeks of training + 2 weeks of tapering at a specific time of day ↔ magnitude of MVC strength gains after 12 weeks of training + 2 weeks of tapering at a specific time of day |
Yes | 17:00 h | 07:00 and 17:00 h |
| Edwards et al.76 | 2013 | Handgrip strength, isokinetic leg strength1, and maximal voluntary isometric contractions | — | n = 10 males | Physically active and healthy adults |
↑ 3.3% in grip strength performance ↑ 20.9% in isokinetic knee flexion peak torque and 18.2% in Ppeak at 1.05 rad.s(-1) ↑ 10.8% in isokinetic knee extension peak torque at 1.05 rad.s(-1) and 9.4% in isokinetic knee extension peak torque at 4.19 rad.s(-1) ↔ time-of-day dependent variations in strength and power performances even when active4 or passive5 warm-up conditions were imposed in the morning |
Yes | 17:30 h | 07:30 and 17:30 h |
| Gauthier et al.77 | 1996 | Maximal and submaximal voluntary isometric contractions at 90° of elbow flexors | — |
n = 7 males n = 6 females |
Physical Education students |
↑ 3.94% in isometric elbow flexion peak torque at 90° ↑ NME slope throughout the day |
Yes | 18:00 h | 06:00, 09:00, 12:00, 15:00, 18:00, 21:00 and 00:00 h |
| Giacomoni, Billaut and Falgairette33 | 2006 | Maximal voluntary isometric knee extensions | — | n = 12 males | Physically active and healthy adults |
↑ 2.1% in 5-min post-test EMG ↑ 14% in 5-min post-test NME (faster short-term recovery patterns of neuromuscular function) |
Yes | 08:00–10:00 h | 08:00–10:00 and 17:00–19:00 h |
| Gueldich et al.78 | 2017 | Maximal voluntary isometric knee extensions | 5 weeks of training | n = 20 males | Physical Education students |
↑ 3.6% and 4.3% in MVC peak torque (in the morning- and evening-training groups, respectively) before the training period ↔ EMG throughout the day ↓ 86% time-of-day dependent variations in MVC peak torque in the morning-training group after 5 weeks of training at a specific time of day |
Yes | 17:00 h | 07:00 and 17:00 h |
| Guette, Godin and Martin79 | 2005 | Plantar flexion of soleus muscle under voluntary and evoked conditions | — | n = 12 males | Physical Education students | ↓ 4.9% in MVC peak torque and 18.8% in associated soleus EMG in the evening | Yes | 06:00–08:00 h | 06:00–08:00 and 17:00–19:00 h |
| Guette, Gondin and Martin26 | 2005 | Electrically evoked and maximal voluntary isometric contractions | — | n = 10 males | Physical Education students | ↑ 3.3% in MVC peak torque of the quadriceps and semi-tendinous muscles | Yes | 18:18 h* | 06:00, 10:00, 14:00, 18:00 and 22:00 h |
| Guette et al.39 | 2006 | Percutaneous electrical stimuli and maximal voluntary isometric contraction of the plantar flexors | — | n = 11 males | Physical Education students | ↓ 7% in MVC peak torque and 21% in associated soleus EMG in the evening | Yes | 06:00–08:00 h | 06:00–08:00 and 17:00–19:00 h |
| Kuusmaa, Sedliak and Hakkinen68 | 2015 | Maximal bilateral isometric leg press, maximal unilateral isometric knee extension and maximal voluntary activation level during unilateral isometric knee extension | — | n = 72 males | Physically active and healthy adults |
↑ 4.4% in MVCLP and 4.3% in MVCKE peak torque at 18:00 h ↑ 10.8% in MVCLP and 5.7% in MVCKE peak torque at 07:30 h in morning-type individuals ↑ 16.1% in MVCLP, 13.5% in MVCKE, 6.2% in MVCVA peak torque and VA% at 18:00 h in evening-type individuals ↔ MVCVA peak torque and VA% throughout the day ↔ EMGLP and EMGVA throughout the day |
Yes | - | 07:30 and 18:00 h |
| Küüsmaa-Schildt et al.69 | 2017 | Maximal voluntary isometric knee extensions coupled with EMG recordings | 24 weeks of training | n = 51 males | Physically active and healthy adults | ↔ MVC peak torque, Ppeak and VA% throughout the day after 24 weeks of training at a specific time of day | No | - | 06:30–09:30 and 16:30–19:30 h |
| Lappalainen et al.81 | 2009 | Isokinetic leg strength (knee extension at 120°/s) | — | n = 26 males | Untrained healthy adults | ↑ isokinetic peak torque at 120°/s and total work | Yes | 16:30 h | 08:00 and 16:30 h |
| Martin et al.29 | 1999 | Electrically evoked and maximal voluntary isometric contractions | — |
n = 12 males n = 1 female |
Healthy adults |
↑ 8.9% in MVC peak torque ↑ 9.2% in tetanic force, 18% in maximum rate of tension development and 32% in relaxation of the twitch |
Yes | 18:00 h | 07:00 and 18:00 h |
| Nicolas et al.82 | 2005 | Isokinetic leg strength6 | — | n = 12 males | Physically active and healthy adults |
↑ 7.7% in isokinetic peak torque ↓ 3.4–5.1% in NME of vastus lateralis, vastus medialis and rectus femoris muscles in the evening ↔ EMG throughout the day |
Yes | 18:00 h | 06:00 and 18:00 h |
| Nicolas et al.70 | 2007 | Isokinetic and isometric leg strength7 | — | n = 10 males | Physically active and healthy adults | ↑ 4–8.8% in MVC peak torque and isokinetic peak torque at 60, 240, 0 and −60°/s | Yes | 18:00 h | 06:00 and 18:00 h |
| Nicolas et al.34 | 2008 | Maximal voluntary isometric knee extensions | — | n = 11 males | Competitive cyclists | ↑ 6.7% in MVC peak torque and 6.8% in NME | Yes | 18:00 h | 06:00 and 18:00 h |
| Pearson and Onambele83 | 2005 | Isokinetic leg strength8 coupled with EMG recordings and theta and patella tendon stiffness recordings | — | n = 13 males | N/A |
↑ 29.4% in isokinetic knee extension peak torque at 70° ↑ 8% and 35% in vastus lateralis pennation angle in relaxed and peak contracted conditions, respectively ↓ 40% in tendon stiffness in the evening |
Yes | 17:45 h | 07:45 and 17:45 h |
| Robinson et al.72 | 2013 | Handgrip strength, isokinetic leg strength9 and maximal voluntary isometric contractions | — | n = 10 males | Physically active and healthy adults |
↑ 4.3% and 7.6% in left and right handgrip strength, respectively ↑ 10.3% in MVC peak torque ↑ 12.6–16.3% in isokinetic knee flexion and extension peak torque and power at 1.05 rad.s(-1) and 8.6% in isokinetic knee extension peak torque at 4.19 rad.s(-1) ↔ time-of-day dependent variations in strength and power performances when pre-cooling11 conditions were imposed in the evening |
Yes | 17:30 h | 07:30 and 17:30 h |
| Racinais et al.71 | 2005 | Maximal and submaximal voluntary isometric knee extensions | — | n = 11 males | Physical Education students |
↑ 12% in MVC peak torque and 25.4% in muscle contractility ↔ time-of-day dependent variations in strength and power performances in moderately warm and humid12 compared with neutral13 conditions |
Yes | 17:00–19:00 | 07:00–09:00 and 17:00–19:00 h |
| Sedliak et al.41 | 2008 | Maximal and submaximal voluntary isometric knee extensions | — | n = 32 males | Physically active and healthy adults10 | ↑ 2.4–8.7% in MVC peak torque at 120°/s throughout the day | Yes | 12:00–13:00, 17:00–18:00 and 20:30–21:30 h | 07:00–08:00, 12:00–13:00, 17:00–18:00 and 20:30–21:30 h |
| Sedliak et al.66 | 2008 | Maximal voluntary isometric knee extensions | 10 weeks of training at a specific time of day | n = 34 males | Physically active and healthy adults10 | ↔ magnitude of MVC strength gains after 10 weeks of training at a specific time of day | No | — | 09:00–16:00 h |
| Sedliak et al.73 | 2007 | Maximal voluntary isometric knee extensions | 10 weeks of training at a specific time of day | n = 38 males | Physically active and healthy adults10 |
↑ 3.3–9.2% in MVC peak torque throughout the day before the training period ↔ time-of-day dependent variations in MVC peak torque in the morning-training group after 10 weeks of training at a specific time of day |
Yes | 12:00, 17:00 and 20:30 h | 07:00, 12:00, 17:00 and 20:30 h |
| Sedliak et al.65 | 2009 | Maximal voluntary isometric knee extensions | 10 weeks of time-of-day-specific resistance training | n = 24 males | Physically active and healthy adults10 | ↔ magnitude of MVC strength gains after 10 weeks of training at a specific time of day | No | — | 09:00–16:00 h |
| Sedliak et al.74 | 2018 | Maximal voluntary isometric knee extensions | 11 weeks of training at a specific time of day | n = 25 males | Physically active and healthy adults10 | ↔ magnitude of MVC strength gains after 11 weeks of training at a specific time of day | No | — | 07:30–08:30 and 16:00–17:00 h |
| Souissi et al.35 | 2010 | Handgrip strength | — | n = 20 males | Untrained healthy boys | ↑ 5.9% in handgrip strength throughout the day | Yes | 14:00 and 18:00 h | 08:00, 14:00 and 18:00 h |
| Souissi et al.36 | 2012 | Maximal voluntary isometric knee extensions | 6 weeks of training at a specific time of day | n = 24 males | Untrained healthy boys |
↑ 8.4% in MVC peak torque before the training period. ↔ time-of-day dependent variations in MVC peak torque in the morning-training group after 6 weeks of training at a specific time of day |
Yes | 17:00–18:00 h | 07:00–08:00 and 17:00–18:00 h |
| Souissi et al.54 | 2002 | Isokinetic leg strength at six angular velocities (1.05, 2.10, 3.14, 4.19, 5.24 and 6.29 rad.s(-1)) | 6 weeks of training at a specific time of day | n = 14 males | Physical Education students |
↑ isokinetic knee extension peak torque at 17:00–18:00 h before the training period. ↔ time-of-day dependent variations in isokinetic knee extension peak torque in the morning-training group after 6 weeks of training at a specific time of day ↑ 38.6% in absolute levels of isometric knee extension peak torque at 07:00–08:00 h in the morning-training group after 6 weeks of training at a specific time of day ↑ 21.2% in absolute levels of isometric knee extension peak torque at 17:00–18:00 h in the evening-training group after 6 weeks of training at a specific time of day |
Yes | — | 07:00–08:00 and 17:00–18:00 h |
| Tamm et al.75 | 2009 | Magnetic stimulation of the cortex, electrical stimulation of the tibial nerve and maximal isometric contractions of the triceps surae muscles | — |
n = 16 males n = 7 females |
Untrained healthy adults |
↑ cortical excitability at 09:00 h in morning-type individuals ↑ cortical excitability at 21:00 h in evening type-individuals ↔ spinal excitability throughout the day in morning- nor evening-type individuals ↔ MVC peak torque nor EMG throughout the day in morning-type individuals. ↑ 13% in MVC peak torque and 23% in EMG throughout the day in evening-type individuals |
Yes | — | 09:00, 13:00, 17:00 and 21:00 h |
| Wyse et al.84 | 1994 | Isokinetic leg strength14 | — | n = 9 males | Collegiate sportsmen adults | ↑ 5–12% in isokinetic peak torque of knee flexors and extensors at 1.05 and 3.14 rad.s(-1) | Yes | 18:00–19:30 h | 08:00–09:00, 13:00–14:00 and 18:00–19:30 h |
| Zbidi et al.45 | 2016 | Maximal voluntary isometric contractions of the elbow flexors and extensors | 6 weeks of training at a specific time of day | n = 20 males | Physical Education students |
↑ 5.9% and 6.5% in MVF and MRFD, respectively, before the training period ↔ time-of-day dependent variations in MVF and MRFD in the morning-training group after 6 weeks of training at a specific time of day |
Yes | 17:00–18:00 h | 07:00–08:00 and 17:00–18:00 h |
EMG electromyographic activity, MCV maximal voluntary contraction, NME Neuromuscular efficiency (mechanical performances/EMG recordings), N/A Not available, 1 i.e. knee flexion and extension at 1.05 and 4.19 rad.s(-1) through a 90° range of motion 2 i.e. 15–16 h starvation/day; from ≈ 04:00 h till ≈ 19:00 h, 3 i.e. 24-h cycling on an indoor trainer with minimal resting periods at a submaximal work rate (paced speed set at 70% of the subject’s maximal aerobic speed, corresponding approximately to 50% of their maximal aerobic power) 4i.e. 20–40 min of pedaling on a cycle ergometer at 150 W, 5i.e. 45–65 min of resting in a room at 35 °C, 45% relative humidity, *estimated acrophase by cosinor analysis.
EMG electromyographic activity, MCV maximal voluntary contraction, NME Neuromuscular efficiency (mechanical performances/EMG recordings), MVCLP maximal bilateral isometric leg press, EMGLP myoelectric activity during maximal bilateral isometric leg press, MVCKE maximal unilateral isometric knee extension, MVCVA maximal voluntary activation level unilateral isometric knee extension, EMGVA myoelectric activity during maximal voluntary activation level unilateral isometric knee extension, VA% voluntary activation percentage of the quadriceps muscles, *estimated acrophase by cosinor analysis.
EMG electromyographic activity, MCV maximal voluntary contraction, NME Neuromuscular efficiency (mechanical performances/EMG recordings), N/A Not available, 6i.e. knee extension at 2.09 rad.s(-1) through a 90° range of motion, 7 i.e. knee flexion at 240°/s, 60°/s, 0°/s, −60°/s, 8i.e. knee flexion and extension at 90°/s, 80°/s, 70°/s, 50°/s and 30°/s, 9 i.e. knee flexion and extension at 1.05 and 4.19 rad.s(-1) through a 90° range of motion, 10 i.e. no medication within the last14 days, a non-smoker, regular sleep pattern with sleep duration ranging from 6 to 9 h per night and regular physical activity not more than once a week, 111 i.e. immersion in cold water (16.5 °C) before testing to lower rectal temperature to morning values, 12i.e. 60 min of resting in a room at 29.5 °C, 74% relative humidity, 13 i.e. 60 min of resting in a room at 20.5 °C, 67% relative humidity.
EMG electromyographic activity, MCV maximal voluntary contraction, 10i.e. no medication within the last14 days, a non-smoker, regular sleep pattern with sleep duration ranging from 6 to 9 h per night and regular physical activity not more than once a week.
MVF maximal voluntary force, MRFD maximal rate of force development, 14i.e. knee flexion and extension at 1.05 and 3.14 rad.s(-1) through a 90° range of motion.
Discussion
Effects of time of day on dynamic short-duration maximal exercise performance
In order to explore the influence of time of day on dynamic short-duration maximal exercise performance, several studies have assessed the presence of time-of-day dependent fluctuations in both continuous and intermittent exercises. Dynamic short-duration maximal exercise performance seems to oscillate consistently throughout the day, peaking in the afternoon (i.e. between 16:00 and 20:00 h) with amplitudes ranging from 1.7 to 17.5% (Table 1). Except for two studies33,46, better short-duration maximal exercise performances were found in the afternoon when single bouts of exercise were performed under neutral climate conditions. Short-duration maximal exercises that are influenced by the time of day include all-out swimming trials47–49, tennis services37,40, all-out cycling13,23,24,27,28,35,36,38,42,43,50–54, maximal jumps2,23,36,38,40–42,46,49,55–57, repeated sprint ability2,33,44,46,56,58–62, one repetition maximum (1RM) assessments63–66 as well as other force-velocity based tests35,40,49,54,62,63,67.
Neuromuscular differences in dynamic short-duration maximal exercise performance throughout the day has been explored using electromyographic (EMG) activity recordings. Castaingts et al.55 analyzed variations in force and EMG activity of skeletal muscles throughout the day as well as the ratio between these parameters (i.e. force/EMG activity) during a natural movement (i.e. drop jump). Such relationship between the force and EMG activity is called neuromuscular efficiency (NME). In this study, they observed a higher jump height and NME in the evening than in the morning, which indicates that the process of storage-release of potential energy in muscle elastic elements is improved in the evening, without a parallel increase in motor unit activation55. A further analysis of time-of-day fluctuations in power and EMG activity of vastus lateralis, rectus femoris, vastus medialis and biceps femoris muscles recorded in a repeated sprint exercise protocol (5 × [6 s of maximal cycling sprint + 30 s of rest]) showed that total work, percentage of peak power decrement and peak power were higher in the evening than in the morning, although it was not accompanied by a time-of-day effect on EMG activity levels44. Similarly, Chtourou et al.24 recorded EMG activity changes during a Wingate anaerobic test and showed that power output and NME were higher in the evening during the first phase of the test (i.e. first 20 s), where peripheral mechanisms of muscle contraction have the main role in exercise performance. Yet, they were independent of the time of day during the second phase of the test (last 10 s), where central mechanisms of muscle contraction have a higher role in exercise performance. Thus, since most studies have shown that changes in muscle function throughout the day are not accompanied by changes in EMG activity levels, it has been suggested that adaptations at the muscle fibre level rather than changes of the neural drive, motor unit properties, and/or muscle membrane properties are more likely to cause time-of-day dependent variations in dynamic short-duration maximal exercise performance66.
Other studies have also suggested the existence of peripheral mechanisms which might explain diurnal oscillations in dynamic short-duration maximal exercise performance. For instance, Hammouda et al.58 observed that the neuromuscular performance during a repeated sprint ability test (5 × [6 s of maximal cycling sprint + 24 s of rest]) was higher in the evening and it was accompanied not only by higher levels of biological markers of muscle injury but also a lower antioxidant status at this time of the day. Furthermore, Racinais et al.61 assessed time-of-day differences on repeated sprint exercise performance and found a better performance and higher blood lactate concentrations in the evening than in the morning. Although the physiological basis of these time-of-day dependent oscillations has not been elucidated, the evidence suggests the existence of a muscle specific mechanism that accounts for time-of-day dependent fluctuations in neuromuscular performance, whose activity is at least partially independent of the central nervous system.
Effects of time of day on isometric and isokinetic exercise performance
The performance in isometric and isokinetic exercises fluctuates throughout the day over a wide variety of muscles, with amplitudes ranging from 3 to 29.4% (Table 2). Greater amplitudes in isometric and isokinetic exercise performance were found at around 17:00–19:00 h. These observations included assessments of maximal voluntary isometric contractions26,29,30,33,34,36,39,41,45,55,65,66,68–80, isokinetic leg contractions54,70,72,76,81–84, handgrip strength35,72,76 and electrically evoked contractions26,29,39,55,75,79.
The time of day influences isometric and isokinetic exercise performance in both lower and upper extremities. Regarding lower extremities, Guette, Gondin and Martin26 found a significant time-of-day effect on maximal voluntary muscle contraction (MVC) peak torque of the quadriceps muscles on the dominant and non-dominant leg, with the highest values occurring at 18:00 h. Regarding upper extremities, Gauthier et al.77 observed a time-of-day dependent rhythm in elbow flexor torque, whose acrophase was reached at around 18:00 h. This was accompanied by a time-of-day dependent rhythm in biceps muscle EMG activity. Thus, it is clear that upper and lower limb muscle contractility in isometric and isokinetic actions is affected by the time of day. In this sense, most of the studies have shown that when the performance is assessed at a peripheral level by means of peak power, mean power or total work, then a typical peak in performance is found in the evening29,30,34–36,41,45,54,61,68,70,72,73,76–79,81–85.
However, the ability to generate force in isometric and isokinetic exercises depends not only on peripheral but also on central mechanisms of muscle contraction. Central mechanisms include central nervous system command, alertness and motivation, being them all normally assessed by electromyographic activity recording of skeletal muscles68. When muscle performance is assessed at central level, higher morning values77,79, higher evening values80 or no differences throughout the day26,29,66,82 are observed. Such differences in EMG activity throughout the day observed among different studies are difficult to be explained. However, many researchers suggest that EMG activity recordings can be affected by the muscle groups examined. Strikingly, time-of-day dependent fluctuations in maximal isometric and isokinetic performance seem to be different between fast and slow muscles of the lower limbs. In this sense, a decrease in MVC peak torque of triceps surae muscles and soleus muscle EMG activity was observed in the evening in comparison to the morning. According to the authors, this is probably due to a higher fatigue state of the slow motor units as the day progresses39,79. It is worth noting that methodological factors associated with EMG activity recordings might also account for differences in central mechanisms of muscle contraction throughout the day.
Finally, some studies have suggested that diurnal fluctuations in short-duration maximal exercise performance may be partially controlled by the individual circadian typology/chronotype. Thus, inter-individual differences related to the chronotype might generate opposite responses in central and peripheral mechanisms of muscle contraction throughout the day. Chronotype is an individual’s characteristic pattern that reflects preferences towards morningness or eveningness, and it is usually evaluated using self-assessment questionnaires. The effect of the chronotype on isometric and dynamic exercise performance has not been extensively studied, but some studies have shown the importance of the individual typology on isometric exercise performance. For instance, Kuusmaa et al.68 showed that morning-type (M-type) individuals exhibited lower force values in the evening whereas evening-type (E-type) individuals exhibited lower force values in the morning, despite the fact that EMG activities where not different throughout the day in either group. Tamm et al.75 reported that E-type individuals showed parallel increases in cortical and spinal excitability over the day, which were reflected on an increased triceps surae EMG activity and MVC torque at 21:00 h. However, M-type individuals exhibited the highest cortical excitability at 09:00 h but the highest spinal excitability at 21:00 h, without significant differences in triceps surae EMG activity or torque produced during MVCs over the day. Such discrepancies have led some scientists to the conclusion that EMG activity and MVC torque could increase, remain constant or even decrease throughout the day, thus reflecting an orchestrated response between peripheral and central mechanisms in the control of skeletal muscle contractile properties55.
General mechanisms of time-of-day effects on short-duration maximal exercise performance
Performances in short-duration maximal exercises follow a time-of-day dependent rhythmicity, peaking in the afternoon at around 16:00–20:00 h. This effect was observed in children and adults35,66 and among different sports disciplines40,49,50. Diurnal amplitudes in short-duration maximal exercise performance range from 1.7 to 29.4% depending on the muscle feature, muscle group and assessment method. According to the scientific literature published to date, it is clear that a better short-duration maximal exercise performance is achieved in the evening than in the morning in both dynamic13,24,27,28,30,35–38,40–44,47–61,67,85–90 and isometric/isokinetic26,29,30,33–36,39,41,45,54,55,68,70–73,75–85 exercise modes. However, these differences may be blunted in warm and humid environments56,62. This also happens when appropriate warm-up protocols are used57,59 or if music is listened to during warm-ups13,86. The same effect may take place if intermittent fasting conditions are imposed50,85 or if regular exercise training is performed at a specific time of day30,36,45,54,63–66,69,73,74,78.
Effects of temperature
Since short-duration maximal exercise performance fluctuates in parallel with body temperature56,62, it has therefore been suggested that body temperature affects the contractile properties, viscosity and conduction velocity of action potentials in skeletal muscle as well as the extensibility of connective tissue91. In fact, Racinais and Oksa92 have found a positive and linear relationship between performance and muscle temperature, where performance improves from 2 to 5% with a 1 °C increase in muscle temperature. Hence, several research groups have been devoted to studying the effects of getting exposed to warm or cold environments on short-duration maximal exercise performance at different times of the day. In this sense, it was observed that a 60-min exposure to a moderately warm and humid environment (i.e. 28.1–29.5 °C, 62.6–74% relative humidity) blunts the diurnal variation in muscle function that is observed in neutral climate conditions (i.e. 20–20.5 °C, 57–67% relative humidity)56,62,71. Conversely, immersion in cold water (i.e. 16–16.5 °C) before performance testing in the evening has shown to reduce the force and muscle power towards morning values60,72. This is achieved through a “passive effect” of thermal conditions on muscle temperature92. In addition to the passive effect of climate conditions on body temperature, time-of-day dependent variations in muscle performance might also diminish or even disappear when extended warm-up protocols are used (i.e. 20 min cycling on a stationary bike at 150–200 W prior to the completion of the regular warm-up)57. Diurnal fluctuations in performance also disappear under active (i.e. 12 min of pedaling at 50% of VO2max interspersed with three brief accelerations of 5 s) but not passive (i.e. 3 min of pedaling at 70 rpm at 50% of VO2max) warm-up conditions59. The same can be stated when longer active warm-ups (i.e. 15 min of pedaling at 50% of maximal power output) are compared with shorter ones (i.e. 5 min of pedaling at 50% of maximal power output)42. The effectiveness of these protocols to increase performance in the morning towards evening values is due to an “active effect” on muscle temperature92. Thus, increases in muscle temperature might help athletes to minimize diurnal dependent variations in short-duration maximal exercise performance. This can be achieved by means of short exposures to warm and humid environments as well as by the inclusion of active warm-up protocols (e.g. 12–15 min pedaling at minimum intensities of 50% VO2max and interspersed with sprint exercises) before exercising.
Finally, although there is strong evidence to support that time-of-day dependent fluctuations in short-duration maximal exercise are dependent on body temperature, it is clear that there are additional factors which might affect such fluctuations. In this sense, it should be noted that in moderately warm and humid climate conditions better performances are achieved in the morning, when body temperature is at its lowest. However, this is not observed in the evening56,62,71. In this regard, it seems that the effects of the time of day on body temperature and those of warm and humid climate conditions do not interact to potentiate exercise performance in the evening. According to some authors, this is due to the similar effects of both variables on neuromuscular efficiency59, although there is no data to support this hypothesis. On the other hand, it has been observed that an increase in rectal morning temperature towards evening values does not increase muscle strength levels in the morning towards levels found in the evening76,87,89. This might suggest that a specific mechanism within the muscle is orchestrating the effects of time of day on short-duration maximal exercise performance. It is also worth mentioning that when passive exposure to hot environments increases the central temperature towards values close to 39 °C, there is a reduction in the neural drive and the maximal voluntary activation of muscles. This will negatively affect power production and short-duration maximal exercise performance92.
Effects of music
In addition to peripheral factors, it has been suggested that central factors related to alertness, motivation and mood might affect short-duration maximal exercise performance at different times of the day13,28,33,86,93,94. It has been well established that an auditory stimulus, such as music, is effective in increasing levels of arousal, reducing levels of perceived exertion and facilitating a better motor coordination during short-duration maximal exercises94–98. In this regard, Chtourou et al.13 observed higher performances in the Wingate anaerobic test when it was performed after warm-up with music [10-min warm-up while listening high tempo music (>120–140 bpm) through headphones] in comparison to warm-up without music conditions. Furthermore, listening to music during the warm-up period blunted time-of-day dependent variations in muscle power output. This effect was achieved due to a greater increase in performance in the morning than in the evening. Interestingly, a recent study carried out by the same research group showed that this beneficial effect of music on short-duration maximal exercise performance can be achieved when the 10-min warm-up is performed either under neutral (i.e. non-selected) or self-selected motivational music conditions86. In the same study, a greater effect of music on short-duration maximal exercise performance in the morning than in the evening was reported. According to the authors, the ergogenic effect of music could be related to better individual perceptions of self-esteem and sense of confidence in the morning, where the level of motivation seems to be lower among individuals. Thus, listening to music during the warm-up period could be an effective strategy to improve short-duration maximal exercise performance, especially in the morning hours.
Effects of testosterone and cortisol
Short-duration maximal exercises produce an acute increase in both serum testosterone and cortisol levels, affecting the anabolic and catabolic status in skeletal muscle99. In addition, testosterone and cortisol are under circadian regulation, both peaking in the morning hours100. This has raised the question of whether the exercise-induced hormonal response can be influenced by the time of day. In this regard, Bird and Tarpenning101 found that a single session of heavy resistance exercise produced a lower cortisol response when performed in the evening compared with the morning, without affecting the diurnal rhythmicity of testosterone. This suppression in the cortisol response after resistance training in the evening hours was also observed by Burley et al.102. The authors proposed that this reduction in the testosterone/cortisol ratio leads to a reduced catabolic environment which could favor muscle adaptations to resistance exercise in the evening hours. Interestingly, Sedliak et al.73 observed that a 10-week training period at a specific time of day gradually reduced morning but not evening cortisol levels. The authors attributed these changes to a lower anticipatory stress and a better accommodation to early waking rather than any chronic change in cortisol diurnal rhythmicity. However, this period of training at a specific time of day was enough to blunt the time-of-day effects on strength performance in these individuals. Moreover, the absolute increase in maximum strength at the end of the training period was similar in all individuals regardless the time of day at which training was conducted.
It is worth mentioning that, although some studies have observed a relationship between hormone levels, short-duration maximal exercise performance and the time of day, there are other ones that have not so63,64,74. For instance, Kūūsma et al.64 observed no changes in the typical diurnal variations in resting serum testosterone and cortisol concentrations after a 24-week period of training at a specific time of day. According to the authors, the effects of a temporary phase shift caused after a prolonged period of training might explain these results. Furthermore, Sedliak et al.74 observed that resting cortisol levels declined independently of the time of day after 11 weeks of training at a specific time of day in untrained young males. However, they did not find changes in resting testosterone levels after the training period in such individuals. Likewise, no significant changes were observed in resting testosterone levels in a group of healthy older women after 12 weeks of training at a specific time of day63. In summary, whereas resistance exercise training periods at a specific time of day do not seem to affect the circadian pattern of testosterone secretion, their effect on cortisol secretion diurnal pattern is more controversial. Therefore, further research is required to elucidate the role of the time of day on the exercise-induced hormonal response.
Effects of circadian systems
Recently, a large-scale transcriptomic analysis has revealed the existence of rhythmic and CLOCK-driven pathways in human skeletal muscle that affect ~8% of muscle genes103. This analysis showed that the transcription of human skeletal muscle clock genes was distributed into two phases of transcript accumulation at 04:00 and 16:00 h. The afternoon peak was enriched in genes related to muscle contraction and mitochondrial activity whereas the early morning peak was enriched in genes associated with inflammation and the immune response103. In addition, the use of genomic techniques has led to the identification of metabolic pathways regulated by clock genes within skeletal muscle, including those involved in glucose uptake, lipid metabolism and myokine secretion104–106. Dyar et al.104 observed that skeletal muscles of mice with a specific ablation of the core clock gene Bmal1 present an impaired insulin-stimulated glucose uptake due to (1) reduced protein levels of the insulin-dependent glucose transporter GLUT4 and the Rab-GTPase TBC1D1 involved in GLUT4 translocation to the plasma membrane and (2) a decrease in the activity of the glycolytic enzyme PDH. Furthermore, a high-resolution microarray analysis in a muscle-specific Bmal1 knockout mice model revealed a temporal separation of genes involved in carbohydrates and fatty acids use and storage over a period or circadian time of 24 h (CT 1 to 24)107. In this sense, circadian expression of genes involved in carbohydrate catabolism such as hexokinase-2 (Hk2; which catalyzes the first step of glycolysis) and pyruvate dehydrogenase phosphatase (Pdp1; which activates PDH) peak in the late inactive and early active phases of the day (CT 10–12). Moreover, the pyruvate dehydrogenase kinase gene (Pdk4; which inhibits PDH) peaks in the middle of the inactive phase of the day (CT 6). This circadian pattern of gene expression might promote an increase in the glycolytic flux during the active phase of the day (CT 12–24)107. On the other hand, genes involved in fatty acid uptake and β-oxidation such as acyl-carnitine translocase (Slc25a20; which transfers fatty acids into the mitochondrial matrix) and malonyl-CoA decarboxylase (Mlycd; which promotes β-oxidation by reducing malonyl-CoA levels) reach their peak expression in the middle of the inactive phase of the day (CT 7.5). Moreover, the gene that encodes for the nuclear PPARG co-activator 1 beta (Ppargc1b) which stimulates the activity of several transcription factors involved in mitochondrial biogenesis, fatty acid uptake and β-oxidation, also peaks in the middle of the inactive phase of the day (CT 7)107. Altogether, these results could suggest that there is a shift in the use of substrates from lipids to carbohydrates. This metabolic shift, which depends on the circadian gene expression in skeletal muscle, goes from the inactive (CT 1–12) to the active (CT 12–24) phase of the day. This circadian pattern of metabolic gene expression in skeletal muscle could explain the greater performance in short-duration maximal exercises observed in the evening hours (i.e. between 16:00 and 20:00 h). In this sense, a greater activation of the glycolytic pathway in the active phase of the day would promote greater performances in anaerobic exercises in the evening hours.
In spite of the typical temporary peak observed in the evening hours, many studies have demonstrated that regular training at a specific time of the day is able to blunt the diurnal fluctuations in maximal exercise performance30,36,45,54,63–66,69,73,74,78. The mechanisms responsible for the adaptations of training at a specific time of day in short-duration maximal exercise performance remain unknown. However, Sedliak et al.66 observed that the EMG activity during a maximal voluntary contraction of the knee extensors did not show any time-of-day specific adaptation after a 10-week training period at a specific time of day. This led the authors to suggest that peripheral rather than neural adaptations are the main source of temporal specificity in strength training. This could be caused by (1) an attenuation in training adaptations throughout the 10-week training period or (2) individual chronotype-related differences in responsiveness to training at a specific time of day. Likewise, intermittent fasting conditions, such as those imposed during the Ramadan period (i.e. 15–16 h starvation/day; from ≈ 04:00 h until ≈ 19:00 h) have shown to exert the same effect, minimizing the time-of-day dependent variations in exercise performance50,85. Chtourou et al.50 have suggested that Ramadan might act directly on the circadian rhythm of anaerobic performance by means of (1) inducing a phase advance or delay in the rhythm or (2) reducing the amplitude of the rhythm of the anaerobic power development. Despite the fact that none of these hypotheses have been tested to date, some of them have raised the question as whether circadian systems could account for time-of-day specific training adaptations.
There is evidence that scheduled exercise is able to cause phase shifts in the circadian system within skeletal muscle in mice15,16. Using a PER2::LUC circadian reporter mouse model, Wolff and Esser15 showed that four weeks of either voluntary or involuntary exercise for 2 hours/day were able to shift the phase of locomotor and molecular rhythms in skeletal muscle towards an earlier time of day. Furthermore, Edgar and Dement16 used two different wheel restriction schedules to determine whether exercise duration was an important determinant of feedback activity to the mouse circadian system. In this study, they found that more than 75% of total wheel activity occurred in the initial 2 hours over a 6-h or 12-h period of wheel availability. This led the authors to suggest that exercise intensity might be a primary determinant of the exercise-dependent phase shift strength. Thus, the ability of exercise to provide temporal feedback to the circadian system could explain why time-of-day effects on short-maximal exercise performance are blunted after a training period at a specific time of day. In humans, Zambon et al.17 found that the expression of circadian clock genes was affected in quadriceps muscles at 6 and 18 h after having performed a single session of one-leg resistance exercise. In this study, resistance exercise shifted the expression patterns of diurnal-regulated genes through two mechanisms: (1) by upregulating genes that are normally repressed in the morning; and (2) by downregulating genes that are normally activated in the morning. Among them, it was observed an upregulation of the gene that encodes for a regulatory subunit of protein phosphatase-1 (Ppp1r5), which is normally repressed in the morning. Protein phosphatase-1 is responsible for activating glycogen synthase while also inhibiting glycogen breakdown. Moreover, it was observed a downregulation of the gene that encodes for the mitochondrial uncoupling protein 3 (Ucp3), which is upregulated in the morning. Uncoupling protein 3 stimulates oxidative phosphorylation by creating proton leaks across the inner mitochondrial membrane. These data show that exercise represents an important circadian time cue and changes the phase of the molecular clock, specifically in peripheral tissues. Altogether, these results suggest that skeletal muscle molecular clocks might account for the adaptations of training at a specific time of day, suggesting a link between molecular clocks in skeletal muscle and exercise performance throughout the day.
Practical applications
Based on the studies summarized in this manuscript, it is clear that the time of day at which short-duration maximal exercise is conducted is an important variable for training prescription. However, to date, there are no guidelines to help athletes or coaches to achieve optimal performances or avoid deleterious effects of time of day on short-duration maximal exercise performance. Accordingly, some general remarks in this respect include: (1) under neutral climate conditions (i.e. 20–20.5 °C, 57–67% relative humidity), better short-duration maximal exercise performances are achieved in the afternoon (i.e. between 16:00 and 20:00 h) compared with the morning; (2) time-of-day deleterious effects on short-duration maximal exercise performance in the morning may be minimized after: (a) a 10-min warm-up period while listening to neutral or self-selected high-tempo music (>120–140 bpm) through headphones; (b) a 60-min exposure to warm and humid climate conditions (i.e. 28.1–29.5 °C, 62.6–74% relative humidity); (c) active warm-up protocols (e.g. 12–15 min pedaling at minimum intensities of 50% VO2max and interspersed with 5-s sprint exercises); (d) 2–4 weeks of intermittent fasting conditions of 15–16 h starvation/day from ≈04:00 h till ≈19:00 h (e) a training period of at least 5 weeks performed in the morning.
Conclusion
In summary, this review shows that, under neutral climate conditions, short duration maximal exercise performance is affected by the time of day, peaking between 16:00 and 20:00 h. However, a similar performance may be achieved in the morning hours if exercise is conducted after: (1) short exposures to moderately warm and humid environments; (2) active warm-up protocols; (3) intermittent fasting conditions; (4) warming-up while listening to music; (5) prolonged periods of training at a specific time of day. This suggests that time-of-day dependent fluctuations in short-duration maximal exercise performance are controlled not only by body temperature, hormone levels, motivation or mood states but also by a versatile circadian system within skeletal muscle.
Supplementary information
Acknowledgements
Ministério da Educação, Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) e Fundação a Pesquisa do Distrito Federal (FAPDF). We thank Diego López for the grammar and spelling correction of the manuscript.
Author contributions
G.G.M. carried out the review and wrote the paper, R.S.M.N., D.A.V. and E.V. conceptualized the study, C.F. and E.V. acted as additional reviewers and assisted in the review of the final manuscript. G.G.M. design of the review, drafting the work, revising it critically for important intellectual content. R.S.M.N. drafting the work and search for articles in the data bases, revising it critically for important intellectual content. D.A.V. Search for articles in the data bases and revising it critically for important intellectual content. C.F. Revising it critically for important intellectual content and language revising. E.V. Conception and design of the work, interpretation of data for the work, drafting the work and revising it critically for important intellectual content.
Data availability
The datasets generated during and/or analyzed during the current study are available in the MEDLINE and Google Scholar repositories, https://www.ncbi.nlm.nih.gov/pubmed/, https://scholar.google.com/, respectively.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
is available for this paper at 10.1038/s41598-020-66342-w.
References
- 1.Liu AC, Lewis WG, Kay SA. Mammalian circadian signaling networks and therapeutic targets. Nature Chemical Biology. 2007;3(10):630–639. doi: 10.1038/nchembio.2007.37. [DOI] [PubMed] [Google Scholar]
- 2.Racinais S, et al. Effect of an acute hot and dry exposure in moderately warm and humid environment on muscle performance at different times of day. Int J Sports Med. 2006;27(1):49–54. doi: 10.1055/s-2005-837503. [DOI] [PubMed] [Google Scholar]
- 3.Mohawk JA, Takahashi JS. Cell autonomy and synchrony of suprachiasmatic nucleus circadian oscillators. Trends Neurosci. 2011;34(7):349–58. doi: 10.1016/j.tins.2011.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Partch CL, Green CB, Takahashi JS. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014;24(2):90–9. doi: 10.1016/j.tcb.2013.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kimberly HC, Joseph ST. Circadian clock genes and the transcriptional architecture of the clock mechanism. Journal of Molecular Endocrinology. 2019;63(4):R93–R102. doi: 10.1530/JME-19-0153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kennaway DJ, et al. Metabolic homeostasis in mice with disrupted Clock gene expression in peripheral tissues. Am J Physiol Regul Integr Comp Physiol. 2007;293(4):R1528–37. doi: 10.1152/ajpregu.00018.2007. [DOI] [PubMed] [Google Scholar]
- 7.Kohsaka A, et al. High-fat diet disrupts behavioral and molecular circadian rhythms in mice. Cell Metab. 2007;6(5):414–21. doi: 10.1016/j.cmet.2007.09.006. [DOI] [PubMed] [Google Scholar]
- 8.Vieira E, Burris TP, Quesada I. Clock genes, pancreatic function, and diabetes. Trends Mol Med. 2014;20(12):685–93. doi: 10.1016/j.molmed.2014.10.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Vieira E, et al. The clock gene Rev-erbalpha regulates pancreatic beta-cell function: modulation by leptin and high-fat diet. Endocrinology. 2012;153(2):592–601. doi: 10.1210/en.2011-1595. [DOI] [PubMed] [Google Scholar]
- 10.Vieira E, et al. Involvement of the clock gene Rev-erb alpha in the regulation of glucagon secretion in pancreatic alpha-cells. PLoS One. 2013;8(7):e69939. doi: 10.1371/journal.pone.0069939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Balsalobre A, Damiola F, Schibler U. A serum shock induces circadian gene expression in mammalian tissue culture cells. Cell. 1998;93(6):929–37. doi: 10.1016/s0092-8674(00)81199-x. [DOI] [PubMed] [Google Scholar]
- 12.Ko CH, Takahashi JS. Molecular components of the mammalian circadian clock. Hum Mol Genet. 2006;15(Spec No 2):R271–7. doi: 10.1093/hmg/ddl207. [DOI] [PubMed] [Google Scholar]
- 13.Chtourou H, et al. Listening to music affects diurnal variation in muscle power output. Int J Sports Med. 2012;33(1):43–7. doi: 10.1055/s-0031-1284398. [DOI] [PubMed] [Google Scholar]
- 14.Monk TH. Enhancing circadian zeitgebers. Sleep. 2010;33(4):421–2. doi: 10.1093/sleep/33.4.421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wolff G, Esser KA. Scheduled exercise phase shifts the circadian clock in skeletal muscle. Med Sci Sports Exerc. 2012;44(9):1663–70. doi: 10.1249/MSS.0b013e318255cf4c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Edgar DM, Dement WC. Regularly scheduled voluntary exercise synchronizes the mouse circadian clock. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology. 1991;261(4):R928–R933. doi: 10.1152/ajpregu.1991.261.4.R928. [DOI] [PubMed] [Google Scholar]
- 17.Zambon AC, et al. Time- and exercise-dependent gene regulation in human skeletal muscle. Genome biology. 2003;4(10):R61–R61. doi: 10.1186/gb-2003-4-10-r61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Manfredini R, et al. Circadian rhythms, athletic performance, and jet lag. Br J Sports Med. 1998;32(2):101–6. doi: 10.1136/bjsm.32.2.101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Winget CM, DeRoshia CW, Holley DC. Circadian rhythms and athletic performance. Med Sci Sports Exerc. 1985;17(5):498–516. [PubMed] [Google Scholar]
- 20.Tahara Y, Shibata S. Entrainment of the mouse circadian clock: Effects of stress, exercise, and nutrition. Free Radic Biol Med. 2018;119:129–138. doi: 10.1016/j.freeradbiomed.2017.12.026. [DOI] [PubMed] [Google Scholar]
- 21.Tahara Y, Aoyama S, Shibata S. The mammalian circadian clock and its entrainment by stress and exercise. J Physiol Sci. 2017;67(1):1–10. doi: 10.1007/s12576-016-0450-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Sato, S. et al. Time of Exercise Specifies the Impact on Muscle Metabolic Pathways and Systemic Energy Homeostasis. Cell Metab (2019). [DOI] [PubMed]
- 23.Ezagouri, S. et al. Physiological and Molecular Dissection of Daily Variance in Exercise Capacity. Cell Metab, (2019). [DOI] [PubMed]
- 24.Chtourou H, et al. Diurnal variation in Wingate-test performance and associated electromyographic parameters. Chronobiol Int. 2011;28(8):706–13. doi: 10.3109/07420528.2011.596295. [DOI] [PubMed] [Google Scholar]
- 25.Facer-Childs E, Brandstaetter R. The impact of circadian phenotype and time since awakening on diurnal performance in athletes. Curr Biol. 2015;25(4):518–22. doi: 10.1016/j.cub.2014.12.036. [DOI] [PubMed] [Google Scholar]
- 26.Guette M, Gondin J, Martin A. Time-of-day effect on the torque and neuromuscular properties of dominant and non-dominant quadriceps femoris. Chronobiol Int. 2005;22(3):541–58. doi: 10.1081/CBI-200062407. [DOI] [PubMed] [Google Scholar]
- 27.Hammouda O, et al. High intensity exercise affects diurnal variation of some biological markers in trained subjects. Int J Sports Med. 2012;33(11):886–91. doi: 10.1055/s-0032-1301887. [DOI] [PubMed] [Google Scholar]
- 28.Lericollais R, et al. Time-of-day effects on fatigue during a sustained anaerobic test in well-trained cyclists. Chronobiol Int. 2009;26(8):1622–35. doi: 10.3109/07420520903534492. [DOI] [PubMed] [Google Scholar]
- 29.Martin A, et al. Effect of time of day on force variation in a human muscle. Muscle Nerve. 1999;22(10):1380–7. doi: 10.1002/(sici)1097-4598(199910)22:10<1380::aid-mus7>3.0.co;2-u. [DOI] [PubMed] [Google Scholar]
- 30.Chtourou H, et al. The effect of training at the same time of day and tapering period on the diurnal variation of short exercise performances. J Strength Cond Res. 2012;26(3):697–708. doi: 10.1519/JSC.0b013e3182281c87. [DOI] [PubMed] [Google Scholar]
- 31.Buchheit, M. & Laursen, P. High-Intensity Interval Training, Solutions to the Programming Puzzle: Part II: Anaerobic Energy, Neuromuscular Load and Practical Applications. Sports medicine (Auckland, N.Z.), 43 (2013). [DOI] [PubMed]
- 32.Gastin PB. Energy system interaction and relative contribution during maximal exercise. Sports Med. 2001;31(10):725–41. doi: 10.2165/00007256-200131100-00003. [DOI] [PubMed] [Google Scholar]
- 33.Giacomoni M, Billaut F, Falgairette G. Effects of the time of day on repeated all-out cycle performance and short-term recovery patterns. Int J Sports Med. 2006;27(6):468–74. doi: 10.1055/s-2005-865822. [DOI] [PubMed] [Google Scholar]
- 34.Nicolas, A. et al. Effect of time-of-day on neuromuscular properties of knee extensors after a short exhaustive cycling exercise. Isokinetics and exercise science, 16 (2008).
- 35.Souissi H, et al. Time-of-Day Effects on Short-Term Exercise Performances in 10- to 11-Year-Old Boys. Pediatric Exercise Science. 2010;22(4):613–623. doi: 10.1123/pes.22.4.613. [DOI] [PubMed] [Google Scholar]
- 36.Souissi H, et al. The effect of training at a specific time-of-day on the diurnal variations of short-term exercise performances in 10- to 11-year-old boys. Pediatr Exerc Sci. 2012;24(1):84–99. doi: 10.1123/pes.24.1.84. [DOI] [PubMed] [Google Scholar]
- 37.Atkinson G, Speirs L. Diurnal Variation in Tennis Service. Perceptual and Motor Skills. 1998;86(3_suppl):1335–1338. doi: 10.2466/pms.1998.86.3c.1335. [DOI] [PubMed] [Google Scholar]
- 38.Bernard T, et al. Time-of-day effects in maximal anaerobic leg exercise. Eur J Appl Physiol Occup Physiol. 1998;77(1-2):133–8. doi: 10.1007/s004210050311. [DOI] [PubMed] [Google Scholar]
- 39.Guette M, et al. Plantar flexion torque as a function of time of day. Int J Sports Med. 2006;27(3):171–7. doi: 10.1055/s-2005-837618. [DOI] [PubMed] [Google Scholar]
- 40.López-Samanes Á, et al. Circadian rhythm effect on physical tennis performance in trained male players. Journal of Sports Sciences. 2017;35(21):2121–2128. doi: 10.1080/02640414.2016.1258481. [DOI] [PubMed] [Google Scholar]
- 41.Sedliak M, et al. Diurnal variation in maximal and submaximal strength, power and neural activation of leg extensors in men: multiple sampling across two consecutive days. Int J Sports Med. 2008;29(3):217–24.. doi: 10.1055/s-2007-965125. [DOI] [PubMed] [Google Scholar]
- 42.Souissi N, et al. Diurnal variation in Wingate test performances: influence of active warm-up. Chronobiol Int. 2010;27(3):640–52. doi: 10.3109/07420528.2010.483157. [DOI] [PubMed] [Google Scholar]
- 43.Souissi N, et al. Circadian rhythms in two types of anaerobic cycle leg exercise: force-velocity and 30-s Wingate tests. Int J Sports Med. 2004;25(1):14–9. doi: 10.1055/s-2003-45226. [DOI] [PubMed] [Google Scholar]
- 44.Zarrouk N, et al. Time of day effects on repeated sprint ability. Int J Sports Med. 2012;33(12):975–80.. doi: 10.1055/s-0032-1312626. [DOI] [PubMed] [Google Scholar]
- 45.Zbidi S, et al. Diurnal Rhythm of Muscular Strength Depends on Temporal Specificity of Self-Resistance Training. J Strength Cond Res. 2016;30(3):717–24. doi: 10.1519/JSC.0000000000001144. [DOI] [PubMed] [Google Scholar]
- 46.Blonc S, et al. Effects of 5 weeks of training at the same time of day on the diurnal variations of maximal muscle power performance. J Strength Cond Res. 2010;24(1):23–9. doi: 10.1519/JSC.0b013e3181b295d6. [DOI] [PubMed] [Google Scholar]
- 47.Arnett MG. Effects of prolonged and reduced warm-ups on diurnal variation in body temperature and swim performance. Journal of strength and conditioning research. 2002;16(2):256–261. [PubMed] [Google Scholar]
- 48.Baxter C, Reilly T. Influence of time of day on all-out swimming. British journal of sports medicine. 1983;17(2):122–127. doi: 10.1136/bjsm.17.2.122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Pallarés, J. et al. Circadian rhythm effects on neuromuscular and sprint swimming performance. Vol. 45. 2014.
- 50.Chtourou H, et al. The effect of time-of-day and Ramadan fasting on anaerobic performances. Int J Sports Med. 2012;33(2):142–7. doi: 10.1055/s-0031-1286251. [DOI] [PubMed] [Google Scholar]
- 51.Hill DW, et al. Effect of time of day on aerobic and anaerobic responses to high-intensity exercise. Can J Sport Sci. 1992;17(4):316–9. [PubMed] [Google Scholar]
- 52.Lericollais R, et al. Diurnal evolution of cycling biomechanical parameters during a 60-s Wingate test. Scand J Med Sci Sports. 2011;21(6):e106–14. doi: 10.1111/j.1600-0838.2010.01172.x. [DOI] [PubMed] [Google Scholar]
- 53.Melhim AF. Investigation of circadian rhythms in peak power and mean power of female physical education students. Int J Sports Med. 1993;14(6):303–6. doi: 10.1055/s-2007-1021182. [DOI] [PubMed] [Google Scholar]
- 54.Souissi N, et al. Effects of regular training at the same time of day on diurnal fluctuations in muscular performance. J Sports Sci. 2002;20(11):929–37. doi: 10.1080/026404102320761813. [DOI] [PubMed] [Google Scholar]
- 55.Castaingts V, et al. Neuromuscular efficiency of the triceps surae in induced and voluntary contractions: morning and evening evaluations. Chronobiol Int. 2004;21(4-5):631–43. doi: 10.1081/cbi-120039207. [DOI] [PubMed] [Google Scholar]
- 56.Racinais S, Hue O, Blonc S. Time-of-day effects on anaerobic muscular power in a moderately warm environment. Chronobiol Int. 2004;21(3):485–95. doi: 10.1081/cbi-120038632. [DOI] [PubMed] [Google Scholar]
- 57.Taylor K, et al. Warm-up affects diurnal variation in power output. Int J Sports Med. 2011;32(3):185–9. doi: 10.1055/s-0030-1268437. [DOI] [PubMed] [Google Scholar]
- 58.Hammouda O, et al. Diurnal variations of plasma homocysteine, total antioxidant status, and biological markers of muscle injury during repeated sprint: effect on performance and muscle fatigue–a pilot study. Chronobiol Int. 2011;28(10):958–67. doi: 10.3109/07420528.2011.613683. [DOI] [PubMed] [Google Scholar]
- 59.Racinais S, Blonc S, Hue O. Effects of active warm-up and diurnal increase in temperature on muscular power. Med Sci Sports Exerc. 2005;37(12):2134–9. doi: 10.1249/01.mss.0000179099.81706.11. [DOI] [PubMed] [Google Scholar]
- 60.Racinais S, et al. Does the diurnal increase in central temperature interact with pre-cooling or passive warm-up of the leg? J Sci Med Sport. 2009;12(1):97–100. doi: 10.1016/j.jsams.2007.09.008. [DOI] [PubMed] [Google Scholar]
- 61.Racinais S, et al. Morning versus evening power output and repeated-sprint ability. Chronobiol Int. 2005;22(6):1029–39. doi: 10.1080/07420520500397918. [DOI] [PubMed] [Google Scholar]
- 62.Racinais S, et al. Time-of-day effects in maximal anaerobic leg exercise in tropical environment: a first approach. Int J Sports Med. 2004;25(3):186–90. doi: 10.1055/s-2003-45258. [DOI] [PubMed] [Google Scholar]
- 63.Krčmárová B, et al. The effects of 12-week progressive strength training on strength, functional capacity, metabolic biomarkers, and serum hormone concentrations in healthy older women: morning versus evening training. Chronobiology International. 2018;35(11):1490–1502. doi: 10.1080/07420528.2018.1493490. [DOI] [PubMed] [Google Scholar]
- 64.Kuusmaa M, et al. Effects of morning versus evening combined strength and endurance training on physical performance, muscle hypertrophy, and serum hormone concentrations. Appl Physiol Nutr Metab. 2016;41(12):1285–1294. doi: 10.1139/apnm-2016-0271. [DOI] [PubMed] [Google Scholar]
- 65.Sedliak M, et al. Effect of time-of-day-specific strength training on muscular hypertrophy in men. J Strength Cond Res. 2009;23(9):2451–7. doi: 10.1519/JSC.0b013e3181bb7388. [DOI] [PubMed] [Google Scholar]
- 66.Sedliak M, et al. Effect of time-of-day-specific strength training on maximum strength and EMG activity of the leg extensors in men. J Sports Sci. 2008;26(10):1005–14. doi: 10.1080/02640410801930150. [DOI] [PubMed] [Google Scholar]
- 67.Javierre C, et al. Influence of sleep and meal schedules on performance peaks in competitive sprinters. Int J Sports Med. 1996;17(6):404–8. doi: 10.1055/s-2007-972869. [DOI] [PubMed] [Google Scholar]
- 68.Kuusmaa M, Sedliak M, Hakkinen K. Effects of time-of-day on neuromuscular function in untrained men: Specific responses of high morning performers and high evening performers. Chronobiol Int. 2015;32(8):1115–24. doi: 10.3109/07420528.2015.1065269. [DOI] [PubMed] [Google Scholar]
- 69.Kuusmaa-Schildt M, et al. Neuromuscular Adaptations to Combined Strength and Endurance Training: Order and Time-of-Day. Int J Sports Med. 2017;38(9):707–716. doi: 10.1055/s-0043-101376. [DOI] [PubMed] [Google Scholar]
- 70.Nicolas A, et al. Effect of circadian rhythm of neuromuscular properties on muscle fatigue during concentric and eccentric isokinetic actions. Isokinetics and Exercise Science. 2007;15:117–129. [Google Scholar]
- 71.Racinais S, et al. Time of day influences the environmental effects on muscle force and contractility. Med Sci Sports Exerc. 2005;37(2):256–61. doi: 10.1249/01.mss.0000149885.82163.9f. [DOI] [PubMed] [Google Scholar]
- 72.Robinson WR, et al. Does lowering evening rectal temperature to morning levels offset the diurnal variation in muscle force production? Chronobiol Int. 2013;30(8):998–1010. doi: 10.3109/07420528.2013.793197. [DOI] [PubMed] [Google Scholar]
- 73.Sedliak M, et al. Effect of time-of-day-specific strength training on serum hormone concentrations and isometric strength in men. Chronobiol Int. 2007;24(6):1159–77. doi: 10.1080/07420520701800686. [DOI] [PubMed] [Google Scholar]
- 74.Sedliak M, et al. Morphological, molecular and hormonal adaptations to early morning versus afternoon resistance training. Chronobiol Int. 2018;35(4):450–464. doi: 10.1080/07420528.2017.1411360. [DOI] [PubMed] [Google Scholar]
- 75.Tamm AS, et al. Chronotype influences diurnal variations in the excitability of the human motor cortex and the ability to generate torque during a maximum voluntary contraction. J Biol Rhythms. 2009;24(3):211–24. doi: 10.1177/0748730409334135. [DOI] [PubMed] [Google Scholar]
- 76.Edwards BJ, et al. Does raising morning rectal temperature to evening levels offset the diurnal variation in muscle force production? Chronobiol Int. 2013;30(4):486–501. doi: 10.3109/07420528.2012.741174. [DOI] [PubMed] [Google Scholar]
- 77.Gauthier A, et al. Diurnal rhythm of the muscular performance of elbow flexors during isometric contractions. Chronobiol Int. 1996;13(2):135–46. doi: 10.3109/07420529609037077. [DOI] [PubMed] [Google Scholar]
- 78.Gueldich H, et al. Electrostimulation Training Effects on diurnal Fluctuations of Neuromuscular Performance. Int J Sports Med. 2017;38(1):41–47. doi: 10.1055/s-0042-115033. [DOI] [PubMed] [Google Scholar]
- 79.Guette M, Gondin J, Martin A. Morning to evening changes in the electrical and mechanical properties of human soleus motor units activated by H reflex and M wave. Eur J Appl Physiol. 2005;95(4):377–81. doi: 10.1007/s00421-005-0023-6. [DOI] [PubMed] [Google Scholar]
- 80.Callard D, et al. Circadian rhythms in human muscular efficiency: continuous physical exercise versus continuous rest. A crossover study. Chronobiol Int. 2000;17(5):693–704. doi: 10.1081/cbi-100101075. [DOI] [PubMed] [Google Scholar]
- 81.Lappalainen Z, et al. Time-of-day effects during acute isokinetic exhaustive eccentric exercise: Serum leptin response. Isokinetics and Exercise Science. 2009;17:19–25. [Google Scholar]
- 82.Nicolas A, et al. Time-of-day effects on myoelectric and mechanical properties of muscle during maximal and prolonged isokinetic exercise. Chronobiol Int. 2005;22(6):997–1011. doi: 10.1080/07420520500397892. [DOI] [PubMed] [Google Scholar]
- 83.Pearson SJ, Onambele GN. Acute changes in knee-extensors torque, fiber pennation, and tendon characteristics. Chronobiol Int. 2005;22(6):1013–27. doi: 10.1080/07420520500397900. [DOI] [PubMed] [Google Scholar]
- 84.Wyse JP, Mercer TH, Gleeson NP. Time-of-day dependence of isokinetic leg strength and associated interday variability. Br J Sports Med. 1994;28(3):167–70. doi: 10.1136/bjsm.28.3.167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Aloui A, et al. Effects of Ramadan on the diurnal variations of repeated-sprint performances. Int J Sports Physiol Perform. 2013;8(3):254–62. doi: 10.1123/ijspp.8.3.254. [DOI] [PubMed] [Google Scholar]
- 86.Belkhir Y, et al. Listening to neutral or self-selected motivational music during warm-up to improve short-term maximal performance in soccer players: Effect of time of day. Physiol Behav. 2019;204:168–173. doi: 10.1016/j.physbeh.2019.02.033. [DOI] [PubMed] [Google Scholar]
- 87.Pullinger S, et al. Effects of an active warm-up on variation in bench press and back squat (upper and lower body measures) Chronobiol Int. 2019;36(3):392–406. doi: 10.1080/07420528.2018.1552596. [DOI] [PubMed] [Google Scholar]
- 88.Robertson CM, et al. Is the diurnal variation in muscle force output detected/detectable when multi-joint movements are analysed using the musclelab force-velocity encoder? Chronobiol Int. 2018;35(10):1391–1401. doi: 10.1080/07420528.2018.1485685. [DOI] [PubMed] [Google Scholar]
- 89.Pullinger SA, et al. Diurnal variation in repeated sprint performance cannot be offset when rectal and muscle temperatures are at optimal levels (38.5 degrees C) Chronobiol Int. 2018;35(8):1054–1065. doi: 10.1080/07420528.2018.1454938. [DOI] [PubMed] [Google Scholar]
- 90.West DJ, et al. The influence of the time of day on core temperature and lower body power output in elite rugby union sevens players. J Strength Cond Res. 2014;28(6):1524–8. doi: 10.1519/JSC.0000000000000301. [DOI] [PubMed] [Google Scholar]
- 91.Shephard RJ. Sleep, Biorhythms and Human Performance. Sports Medicine. 1984;1(1):11–37. [Google Scholar]
- 92.Racinais S, Oksa J. Temperature and neuromuscular function. Scandinavian Journal of Medicine & Science in Sports. 2010;20(s3):1–18.. doi: 10.1111/j.1600-0838.2010.01204.x. [DOI] [PubMed] [Google Scholar]
- 93.Reilly T, Down A. Investigation of circadian rhythms in anaerobic power and capacity of the legs. J Sports Med Phys Fitness. 1992;32(4):343–7. [PubMed] [Google Scholar]
- 94.Chtourou H, Hmida C, Souissi N. Effect of music on short-term maximal performance: sprinters vs. long distance runners. Sport Sciences for Health. 2017;13(1):213–216. [Google Scholar]
- 95.Simpson SD, Karageorghis CI. The effects of synchronous music on 400-m sprint performance. J Sports Sci. 2006;24(10):1095–102. doi: 10.1080/02640410500432789. [DOI] [PubMed] [Google Scholar]
- 96.Yamashita S, et al. Effects of music during exercise on RPE, heart rate and the autonomic nervous system. J Sports Med Phys Fitness. 2006;46(3):425–30. [PubMed] [Google Scholar]
- 97.Eliakim M, et al. The effect of music during warm-up on consecutive anaerobic performance in elite adolescent volleyball players. Int J Sports Med. 2007;28(4):321–5. doi: 10.1055/s-2006-924360. [DOI] [PubMed] [Google Scholar]
- 98.Pujol TJ, Langenfeld ME. Influence of music on Wingate Anaerobic Test performance. Percept Mot Skills. 1999;88(1):292–6. doi: 10.2466/pms.1999.88.1.292. [DOI] [PubMed] [Google Scholar]
- 99.Kraemer WJ, Ratamess NA. Hormonal responses and adaptations to resistance exercise and training. Sports Med. 2005;35(4):339–61. doi: 10.2165/00007256-200535040-00004. [DOI] [PubMed] [Google Scholar]
- 100.Piro C, et al. Circadian rhythm of plasma testosterone, cortisol and gonadotropins in normal male subjects. Journal of Steroid Biochemistry. 1973;4(3):321–329. doi: 10.1016/0022-4731(73)90056-3. [DOI] [PubMed] [Google Scholar]
- 101.Bird SP, Tarpenning KM. Influence of circadian time structure on acute hormonal responses to a single bout of heavy-resistance exercise in weight-trained men. Chronobiol Int. 2004;21(1):131–46. doi: 10.1081/cbi-120027987. [DOI] [PubMed] [Google Scholar]
- 102.Burley SD, et al. The Differential Hormonal Milieu of Morning versus Evening May Have an Impact on Muscle Hypertrophic Potential. PLoS One. 2016;11(9):e0161500. doi: 10.1371/journal.pone.0161500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Perrin L, et al. Transcriptomic analyses reveal rhythmic and CLOCK-driven pathways in human skeletal muscle. eLife. 2018;7:e34114. doi: 10.7554/eLife.34114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Dyar KA, et al. Muscle insulin sensitivity and glucose metabolism are controlled by the intrinsic muscle clock. Molecular Metabolism. 2014;3(1):29–41. doi: 10.1016/j.molmet.2013.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Perrin L, et al. Human skeletal myotubes display a cell-autonomous circadian clock implicated in basal myokine secretion. Molecular Metabolism. 2015;4(11):834–845. doi: 10.1016/j.molmet.2015.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.van Moorsel D, et al. Demonstration of a day-night rhythm in human skeletal muscle oxidative capacity. Molecular metabolism. 2016;5(8):635–645. doi: 10.1016/j.molmet.2016.06.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Hodge BA, et al. The endogenous molecular clock orchestrates the temporal separation of substrate metabolism in skeletal muscle. Skeletal Muscle. 2015;5(1):17. doi: 10.1186/s13395-015-0039-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available in the MEDLINE and Google Scholar repositories, https://www.ncbi.nlm.nih.gov/pubmed/, https://scholar.google.com/, respectively.