

Abstract
Strain Y74T was an isolate from the sandy soil in the town of Huatugou, Qinghai–Tibet Plateau, China. An analysis of this strain's phenotypic, chemotaxonomic, and genomic characteristics established the relationship of the isolate with the genus Planococcus. Strain Y74T was able to grow between 4 and 42°C (with an optimum temperature of 28°C) at pH values of 6–8.5 and in 0%–7% (w/v) NaCl. The dominant quinones were MK‐8 and MK‐7. The polar lipids were diphosphatidylglycerol, phosphatidylethanolamine, phosphatidylglycerol, and an unknown phospholipid. The majority of the fatty acid content was anteiso‐C15:0 (28.8%) followed by C16:1 ω7c alcohol (20.9%) and iso‐C14:0 (13.4%). The 16S rRNA gene sequence similarity analysis demonstrated a stable branch formed by strain Y74T and Planococcus halotolerans SCU63T (99.66%). The digital DNA–DNA hybridization between these two strains was 57.2%. The G + C content in the DNA of Y74T was 44.5 mol%. In addition, the morphological, physiological, and chemotaxonomic pattern clearly differentiated the isolates from their known relatives. In conclusion, the strain Y74T (=JCM 32826T = CICC24461T) represents a novel member of the genus Planococcus, for which the name Planococcus antioxidans sp. nov. is proposed. Strain Y74T was found to have potent antioxidant activity via its hydrogen peroxide tolerance and its 1,1‐diphenyl‐2‐picrylhydrazyl (DPPH) radical‐scavenging activity. The DPPH radical‐scavenging activity was determined to be 40.2 ± 0.7%. The genomic analysis indicated that six peroxidases genes, one superoxide dismutase gene, and one dprA (DNA‐protecting protein) are present in the genome of Y74T.
Keywords: antioxidant, Planococcus antioxidans, polyphasic taxonomy, Qinghai–Tibetan Plateau
In this study, we reported a novel strain isolated from the desert soil in the Qinghai–Tibetan Plateau. According to an analysis of phenotypic, phylogenetic, and chemotaxonomic characteristics, strain Y74T was determined to be a new member within the genus Planococcus. Therefore, it was named as Planococcus antioxidans sp. nov. Y74T. Strain Y74T was found to have potent antioxidant activity via its hydrogen peroxide tolerance and its 1,1‐diphenyl‐2‐picrylhydrazyl (DPPH) radical‐scavenging activity.

1. INTRODUCTION
The accumulation of free radicals in living organisms can lead to many diseases, such as cancer and neurodegenerative diseases (Fischer & Maier, 2015; Lin & Beal, 2006). Thus, it may be possible to reduce and prevent these chronic diseases by decreasing the presence of free radicals and increasing the intake of antioxidants (Bonda et al., 2010; Fischer & Maier, 2015). Microorganisms are an abundant source of bioactive metabolites (Berdy, 2005; Velho‐Pereira, Parvatkar, & Furtado, 2015). Therefore, in order to prevent the toxic effects of free radicals, potent natural antioxidants have been an important target for researchers. Recently, exploring new taxa for new antioxidants has been one of the effective strategies employed in this search.
The Qinghai–Tibet Plateau is the highest plateau in the world, where the average altitude is above 4,500 m (Zhang et al., 2019). Because of the stressful conditions, such as low air temperatures, high UV radiation, and low atmospheric oxygen content, the organisms have had to adapt to survive on this plateau (Zhang et al., 2018; Zhang, Tang, et al., 2016). This environment is a potential source of genetic diversity and is an ideal place to search for antioxidant‐producing microbes (Zhang, Wu, et al., 2016).
The genus Planococcus was originally described by Migula (1895). There were 16 valid species in the genus Planococcus until recently: P. citreus (Migula, 1895), P. kocurii (Hao & Komagata, 1985), P. antarcticus (Reddy et al., 2002), P. maritimus (Yoon, Weiss, Kang, Oh, & Park, 2003), P. maitriensis (Alam, Singh, Dube, Reddy, & Shivaji, 2003), P. rifietoensis (Romano, Giordano, Lama, Nicolaus, & Gambacorta, 2003), P. columbae (Suresh, Mayilraj, Bhattacharya, & Chakrabarti, 2007), P. donghaensis (Choi et al., 2007), P. salinarum (Yoon, Kang, Lee, Oh, & Oh, 2010), P. halocryophilus (Mykytczuk, Wilhelm, & Whyte, 2012), P. plakortidis (Kaur et al., 2012), P. soli (Luo et al., 2014), P. faecalis (Kim, Kang, Yu, Kim, & Lee, 2015), P. ruber (Wang et al., 2017), P. salinus (Gan, Zhang, Tian, et al., 2018), and P. halotolerans (Gan, Zhang, Zhang, et al., 2018). Due to their phenotypic properties, menaquinone profiles, fatty acid composition and G + C content in the DNA, the species Planococcus mcmeekinii (Yoon et al., 2001), Planococcus okeanokoites (Yoon et al., 2001), Planococcus alkanoclasticum (Dai, Wang, Wang, Liu, & Zhou, 2005), Planococcus psychrophilum (Dai et al., 2005), and P. stackebrandtii (Jung, Kang, Oh, Yoon, & Kim, 2009) were reclassified to genus Planomicrobium, and Planococcus halophilus was cataloged to the genus Marinococcus (Hao, Kocur, & Komagata, 1984; Novitsky & Kushner, 1976). The known features of the genus Planococcus are that the species are Gram‐positive, aerobic, non‐spore‐forming and have cell shapes that include cocci, short rods, or rods (Gan, Zhang, Zhang, et al., 2018). The genera Planococcus and Planomicrobium are close phylogenetic neighbors. Dai et al. (2005) found that the specific difference in the 16S rRNA gene sequence between genus Planococcus and Planomicrobium at sites 183 and 190 (E. coli numbering) was that Planococcus contained the signature nucleotides T and A, whereas the Planomicrobium species contained C and G (Dai et al., 2005).
According to our research, a new Planococcus species strain, Y74T, was isolated from the desert soil in the Qinghai–Tibetan Plateau, China. Strain Y74T demonstrated a strong antioxidant activity, which has potential antioxidant applications.
2. MATERIALS AND METHODS
2.1. Bacteria isolation
The desert soil samples were obtained from the town of Huatugou, Qinghai province, China. Strains Y74T was isolated with modified 216 L agar medium (per liter distilled water: 1.0 g sodium acetate, 10.0 g tryptone, 2.0 g yeast extract, 0.5 g sodium citrate, 0.2 g ammonium nitrate, 0.5 g nutrient broth medium, 20.0 g agar, pH 7.6) and incubated for 7 days at 20°C, after which it was preserved at −80°C in 20% (v/v) glycerol (Wang, Wang, & Shao, 2010).
2.2. Genome sequencing and analysis
Genomic DNA was extracted with a bacterial genomic DNA extraction kit (Omega Bio‐tek, Inc.), according to the manufacturer's instructions, and the sequence was determined by the Illumina HiSeq 2000. The reads from the sequencing were assembled de novo using the Velvet 1.2.10 program. The genomes of the type strains that were similar to Y74T were retrieved from GenBank. The average nucleotide identity (ANI) and digital DNA–DNA hybridization (dDDH) were used to assess the degree of similarity of each pair. The ANI was calculated with the JSpeciesWS (Richter, Rossello‐Mora, Glockner, & Peplies, 2016). The ANI could be divided into ANIb and ANIm, depending on the BLASTN (Basic Local Alignment Search Tool) algorithm or the MUMMER ultra‐rapid aligning tool. The dDDH was computed by an online tool, GGDC 2.0: the results of this computation were obtained using the recommended formula 2 (Meier‐Kolthoff, Auch, Klenk, & Goker, 2013). The genome of strain Y74T was annotated using IMG Annotation Pipeline v.5.0.3 (Chen et al., 2019). The G + C content of the DNA of strain Y74T was deduced from the genomic data. Y74T horizontal gene transfer analysis by the method of Bertelli, Laird, and Williams (2017).
2.3. Phylogenetic analysis
The closely related type strains of Y74T were obtained by comparing their 16S rRNA gene sequences, retrieved from the genome in the EzTaxon‐e database (Kim et al., 2012). Phylogenetic trees based on the 16S rRNA gene sequences were generated utilizing neighbor joining (Saitou & Nei, 1987), maximum parsimony (Tamura et al., 2011), and maximum likelihood (Felsenstein, 1981) algorithms in MEGA X (Kumar, Stecher, Li, Knyaz, & Tamura, 2018). The sequences were aligned with ClustalW (Larkin et al., 2007). The remaining parameters followed the model of Jukes and Cantor (Jukes & Cantor, 1969), and the bootstrap value was 1,000 re‐samplings (Felsenstein, 1985). A phylogenetic tree based on 25 housekeeping genes nucleotide sequences was generated using neighbor joining algorithms. The parameters were as same as for the phylogenetic tree based on 16S rRNA gene sequences. The sequences of 25 housekeeping genes were obtained from genomes of 19 type strain of genus Planococcus, Planomicrobium, and 1 outgroup strain (Lysinibacillus sphaericus IAM 13420T) after annotated using Rapid Annotations using Subsystems Technology (RAST) (Brettin et al., 2015). The sequences of 25 housekeeping genes were concatenated in the following order: CTP synthase, DNA primase, DNA‐directed RNA polymerase beta subunit, LSU ribosomal protein L11p, LSU ribosomal protein L13p, LSU ribosomal protein L16p, LSU ribosomal protein L20p, LSU ribosomal protein L27p, LSU ribosomal protein L3p, LSU ribosomal protein L4p, LSU ribosomal protein L5p, LSU ribosomal protein L6p, phosphoglycerate kinase, ribosome recycling factor, SSU ribosomal protein S10p, SSU ribosomal protein S11p, SSU ribosomal protein S13p, SSU ribosomal protein S2p, SSU ribosomal protein S3p, SSU ribosomal protein S5p, SSU ribosomal protein S9p, tmRNA‐binding protein SmpB, transcription termination protein NusA, translation elongation factor Ts, and translation initiation factor 3 (Gil, Silva, Pereto, & Moya, 2004).
2.4. Morphological and physiological analysis
Cell size and morphology were determined by scanning electron microscopy (JSM‐5600, JEOL) utilizing cells immobilized after gold sputtering for 60 s. For scanning electron microscopy, the strain Y74T was fixed on 4% glutaraldehyde for 8 hr. Subsequently, the cells were dehydrated in an ethanol series (15%, 30%, 50%, 70%, 80%, 90%, and 100%) for 10 min each. Colony color was evaluated on LB (Lysogeny Broth) agar (Oxoid).
Gram staining was tested using the Solarbio Gram staining kit. Growth temperatures (4, 10, 15, 20, 25, 30, 35, 40, 42, and 45°C) and NaCl concentrations (0%–10%, w/v, intervals of 0.5%) were determined on 216L medium. The pH range for growth was examined with strains cultured at 28°C in 216L medium, where the buffering system (KH2PO4/HCl, KH2PO4/K2HPO4, and K2HPO4/NaOH) was injected to adjust the pH value from 5 to 10 at 0.5 pH unit intervals. Oxidase activity was detected with 1% (w/v) tetramethyl‐p‐phenylenediamine. Starch and gelatin hydrolysis, nitrate reduction, catalase activity, methyl red, and Voges–Proskauer tests were performed according to the description of Kurup and Schmitt (1973). A carbohydrate utilization test was performed as described previously (Zhang et al., 2018). Additional enzyme activities were detected by API ZYM systems.
2.5. Chemotaxonomic analysis
For the chemotaxonomic analysis, cells were collected by centrifugation from strains cultured at 28°C in TSB medium (per liter distilled water: 17.0 g tryptone, 3.0 g soy peptone, 2.5 g D‐glucose, 5.0 g sodium chloride, 2.5 g monopotassium phosphate, pH 7.3) for 3 days and then washed twice with distilled water. The cell‐wall peptidoglycan was analyzed by the method of Schleifer and Kandler (1972). The whole‐cell sugars were analyzed by the methods of Lechevalier and Lechevalier (1970). The quinones and the polar lipids were analyzed by the method of Collins et al. and HPLC (Collins, Pirouz, Goodfellow, & Minnikin, 1977; Kroppenstedt, 1982) and by the method of Minnikin et al. (1984), respectively. The methylation, extraction, and analysis of the fatty acids were based on the methods of Sasser (1990) and identified in the TSBA 6.0 database of the Sherlock Microbial identification (MIDI) system (Kämpfer & Kroppenstedt, 1996).
2.6. Antioxidant activity analysis
The effect of hydrogen peroxide on the growth of strain Y74T was tested as follows: an inoculum of 100 μl of strain Y74T within the exponential growth phase (OD600 = 0.6) was mixed with 50 ml LB medium containing 0, 1, and 5 mM H2O2 and then incubated at 30°C for 48 hr. The cell concentration was monitored by spectrophotometer (absorbance at 600 nm). All experiments were performed in triplicate. The growth fitting curves of strain Y74T were drawn with Origin 2018 (logistics nonlinear fitting).
The 1,1‐diphenyl‐2‐picrylhydrazyl (DPPH) radical‐scavenging activity was tested in steps. First, an inoculum of 1 ml of strain Y74T within the exponential growth phase (OD600 = 1.0) was centrifuged at 5,300 g for 10 min, after which the supernatant was discarded, and the precipitate was resuspended with 500 μl PBS. This process was repeated three times. The resuspended precipitate was then mixed with 500 μl 0.4 mmol/L DPPH•ethanol (the control group used an equal volume of distilled water), after which the mixture was allowed to react in at low‐light area for 30 min at room temperature and subsequently centrifuged at 5,300 g for 10 min. The absorbance of the supernatant was measured with a spectrophotometer at 517 nm. The DPPH free radical‐scavenging rate was calculated as follows: scavenging activity (%) = [1 − (As − Ab)/Ac] × 100%, where Ab is the absorbance of the blank group, Ac is the absorbance of the control group, and As is the absorbance of the sample set.
3. RESULTS AND DISCUSSION
3.1. Phylogenetic analysis
The entirety of the 16S rRNA gene sequences was extracted from the genome of strain Y74T (1,512 bp, KU601236). The genome of strain Y74T was deposited at DDBJ/EMBL/GenBank with the accession number RCWH00000000.
Compared with the EzTaxon database, type strain Planococcus halotolerans SCU63T (99.66%), Planomicrobium okeanokoites IFO 12536T (98.43%), Planomicrobium flavidum ISL‐41T (98.30%), Planococcus maitriensis S1T (98.23%), and Planomicrobium mcmeekinii S23F2T (98.16%) were found to show high degrees of similarity with strain Y74T (Kim et al., 2012). The three 16S rRNA gene phylogenetic trees indicated that Y74T and Planococcus halotolerans SCU63T formed a stable clade (Figure 1). However, many adjacent clades were not stable. Therefore, a more stable phylogenetic tree was constructed, which was based on 25 concatenated housekeeping genes of strain Y74T and their related type strains (Figure 2). According to this phylogenetic tree, strain Y74T should be a member of the genus Planococcus. The exact position of Planomicrobium oekanokoites IFO12536T could not exactly be resolved within the analysis. The dDDH between strain Y74T and Planococcus halotolerans SCU63T, Planomicrobium okeanokoites IFO 12536T, or Planococcus maitriensis S1T were 57.2%, 30.5%, and 19.1%, respectively. The ANIb values between strain Y74T and Planococcus halotolerans SCU63T, Planomicrobium okeanokoites IFO 12536T, or Planococcus maitriensis S1T were 94.15%, 85.43%, and 72.19%, respectively, with ANIm values of 94.66%, 87.45%, and 83.50%, respectively (Table 1). These values were below the species demarcation threshold in prokaryotic species, the generally accepted species boundary for ANI and dDDH values were 95 ~ 96% and 70%, respectively (Chun et al., 2018; Kim, Oh, Park, & Chun, 2014; Meier‐Kolthoff et al., 2013). For all of these reasons, Planococcus antioxidans sp. nov. Y74T was designated as a novel species in the genus Planococcus.
Figure 1.

Neighbor joining phylogenetic tree, based on nearly complete 16S rRNA gene sequences, showing the relationships among strain Y74T and their related species. Numbers at nodes are bootstrap values based on 1,000 re‐samplings (only values above 50% are shown). Asterisks and hash marks indicate that the clades were also recovered in the maximum parsimony and maximum likelihood trees
Figure 2.

Neighbor joining phylogenetic tree based on 25 concatenated housekeeping genes of strain Y74T and their similar related type strains. Numbers at nodes are bootstrap values based on 1,000 re‐samplings (only values above 50% are shown)
Table 1.
The genome comparisons of strain Y74T and the related type species
| Species | 16S | dDDH | ANIb | ANIm |
|---|---|---|---|---|
| Planococcus halotolerans | 99.66 | 57.2 | 94.15 | 94.66 |
| Planomicrobium okeanokoites | 98.43 | 30.5 | 85.43 | 87.45 |
| Planococcus maitriensis | 98.23 | 19.1 | 72.19 | 83.50 |
| Planococcus citreus | 98.10 | 19.0 | 72.34 | 83.74 |
| Planococcus koreense | 98.10 | 19.6 | 73.37 | 83.96 |
| Planococcus salinarum | 98.03 | 24.9 | 80.89 | 84.83 |
3.2. Morphological and physiological characteristics
Strain Y74T was determined to be Gram‐positive, whose cellular shape was cocci, short rods, or rods, and whose colony color was all white (Figure 3). The growth temperature range of Y74T was 4–42°C (optimum temperature 30°C) with a pH range of 6–8.5 and a NaCl tolerance of up to 7% (w/v) (Table 2). Strain Y74T had a wide range of growth temperatures and a high salt tolerance, which was similar to other closely related type strains of Planococcus or Planomicrobium (Gan, Zhang, Zhang, et al., 2018; Jung et al., 2009). Strain Y74T could utilize D‐fructose, D‐galactose, D‐glucose, D‐lactose, or D‐maltose as sole carbon sources, weakly utilize D‐cellobiose, D‐lactose, D‐mannitol, D‐mannose, D‐melibiose, D‐raffinose, L‐rhamnose, D‐sorbitol, D‐trehalose, or myo‐inositol, and not utilize starch or sucrose (Table 2). However, Planococcus halotolerans SCU63T was able to utilize sucrose but could not utilize D‐sorbitol or melibiose (Gan, Zhang, Zhang, et al., 2018). There were some distinctions with other reference type strains. Strain Y74T showed positivity for catalase and for gelatin hydrolysis and negativity for oxidase, nitrate reduction, methyl red, and the Voges–Proskauer tests (Table 2). The result of the API ZYM test showed that strain Y74T was weakly positive for weak positive for α‐glucosidase, cystine arylamidase, esterase lipase, leucine arylamidase, naphthol‐AS‐BI‐phosphohydrolase, and valine arylamidase.
Figure 3.

Scanning electron micrograph of strain Y74T (a, b) cultivated on LB medium
Table 2.
Comparison of the characteristics of strain Y74T and the closely related type species
| Characteristics | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| Cell shape | C, SR, R | C, SR | R | C, SR | C | C, SR | C, SR, R | C,SR,R |
| Cell length | 0.8–3.4 | 0.4–1.4 | 1.0–20 | 2.7–3.3 | 1–2.0 | 0.8–1.0 | 0.5–2.8 | 0.8–5.0 |
| Cell width | 0.8–1.1 | 0.4–0.6 | 0.4–0.8 | 0.4–0.8 | 1–2.0 | 0.8–1.0 | 0.4–0.8 | 0.4–0.8 |
| Gram stain | Positive | Positive | Positive to variable | Positive to variable | Positive | Positive | Positive to variable | Positive |
| Colony color | White | Moderate orange | Bright yellow‐bright orange | Light yellow | Orange | Yellow orange | Yellow orange | Pale yellow |
| Spore formation | − | − | − | − | − | − | − | − |
| NaCl (%)(w/v) | 0–7 | 15 | 0–7 | 0–14 | 0–12.5 | 0–10 | 0–7 | 0–13 |
| pH | 6–8.5 | 6.5–9.0 | 7.0 a | 6.0–8.0 b | 6.0–12 | 5.0–10 | 5.5–10 | 6–7.5 b |
| Temperature | 4–42 | 0–40 | 20−37 c | 4–37 | 0–30 | 10–45 | 4–38 | 4–38 |
| Catalase | + | + | + | + | + | N | + | + |
| Oxidase | − | + | W | + | − | − | − | + |
| Nitrate reduction | − | − | − | − | + | + | − | − |
| Gelatin hydrolysis | W | − | + | + | + | + | + | − |
| Utilization as carbon sources | ||||||||
| D‐cellobiose | W | + | − | − | − | − | + | − |
| D‐fructose | + | + | + | + | + | N | − | + |
| D‐galactose | + | + | − | − | − | N | − | − |
| D‐glucose | + | + | − | − | + | + | W | − |
| D‐lactose | W | N | − | − | − | − | + | − |
| D‐maltose | + | + | − | − | − | N | + | − |
| D‐mannitol | W | + | − | − | − | − | − | − |
| D‐mannose | W | + | − | − | − | − | − | − |
| D‐melibiose | W | − | − | − | + | − | + | − |
| D‐raffinose | W | N | − | − | + | − | − | − |
| D‐sorbitol | W | − | − | − | − | N | − | − |
| D‐trehalose | W | + | − | − | − | N | − | − |
| L‐rhamnose | W | N | − | − | − | − | − | − |
| myo‐inositol | W | + | − | − | − | N | − | − |
| Starch | − | − | − | − | − | − | − | − |
| Sucrose | − | + | − | − | + | − | − | − |
| Predominant menaquinone | MK‐8, MK‐7 | MK‐8, MK‐7 | MK‐8, MK‐7 | MK‐8,7 | MK‐7,MK‐8 | MK‐8 | MK‐8, MK‐7, MK‐6 | MK‐8, MK‐7 |
| GC content (mol%) | 44.5 | 44.6 | 46 | 45.9 | 39 | 34.8 | 47 | 48.3 |
Strains: 1, Y74T (data from this study); 2, Planococcus halotolerans SCU63T (Gan, Zhang, Zhang, et al., 2018); 3, Planomicrobium okeanokoites IFO 12536T (carbon source utilization and enzyme activity test data were from this study, other data were from Nakagawa, 1996); 4, Planomicrobium flavidum ISL‐41T (Jung, 2009); 5, Planococcus maitriensis S1T (Alam et al., 2003); 6, Planomicrobium chinense DX3‐12T (Dai et al., 2005); 7, Planomicrobium koreense JG07T (Yoon et al., 2001); 8, Planococcus salinarum DSM 23820T (Yoon et al., 2010).
C, cocci; SR, short rods; R, rods; +, positive; −, negative; W, weak positive; N, not determined.
The range of the growth temperatures was not reported.
The lower limit was the minimum value that the strain could grow in, and the upper limit was the maximum value of the optimal growth range.
The optimal growth range.
3.3. Chemotaxonomic characteristics
The whole‐cell hydrolysate of Y74T contained ribose. The peptidoglycan type was L‐Lys‐D‐Glu. The dominant quinone compounds of Y74T were MK‐8 (76%) and MK‐7 (24%) (Table 2). Table 2 shows that the predominant isoprenoid quinone compound of the type strains of Planococcus and Planomicrobium were all MK‐8 and MK‐7. The polar lipids of Y74T were diphosphatidylglycerol, phosphatidylethanolamine, phosphatidylglycerol, and an unknown phospholipid (Figure A1). The major fatty acids of Y74T were anteiso‐C15:0 (28.8%), C16:1 ω7c alcohol (20.9%), and iso‐C14:0 (13.4%) (Table 3). These results conformed to the characteristics of the Planococcus genus (Gan, Zhang, Tian, et al., 2018; Kim et al., 2015; Wang et al., 2017).
Table 3.
Cellular fatty acid composition of strain Y74T
| Fatty acid | Y74T |
|---|---|
| C16:0 | 1.04 |
| C18:0 | 1.14 |
| iso‐C14:0 | 13.90 |
| iso‐C15:0 | 3.54 |
| iso‐C16:0 | 9.62 |
| iso‐C17:0 | 2.77 |
| anteiso‐C15:0 | 28.81 |
| anteiso‐C17:0 | 2.81 |
| C16:1 ω11c | 3.02 |
| C17:0 10‐methyl | 1.03 |
| C16:1 ω7c alcohol | 20.88 |
| iso‐C17:1 ω10c | 3.05 |
| Sum In Feature 4 | 4.37 |
The amount of fatty acid was omitted when <1%. Sum In Feature 4: iso‐C17: 1 I and/or anteiso‐C17: 1 B.
3.4. Antioxidant characteristics
Strain Y74T showed potent antioxidant activity. The strain grew in LB medium supplemented with 1 or 5 mM hydrogen peroxide. The growth curves of Y74T in LB medium supplemented with 0, 1, and 5 mM hydrogen peroxide were similar. Strain Y74T reached a stable growth phase after approximately 18 hr under three conditions. However, the cell concentrations were significantly different, with the highest cell concentration observed in the LB medium with 0 mM hydrogen peroxide and the lowest cell concentration seen in the LB medium with 5 mM hydrogen peroxide (Figure 4). The DPPH radical‐scavenging activity was 40.2 ± 0.7%. There were many studies on antioxidants of genus Lactobacillus, which could produce antioxidants and had extensive application (Das & Goyal, 2015; Le & Cao, 2010; Lee et al., 2010). The strain Y74T had the significant resistant ability to hydrogen peroxide than many species of Lactobacillus, such as Lactobacillus plantarum DM5, Lactobacillus plantarum NRRL B‐4496, and Lactobacillus acidophilus NRRL B‐4495 (Das & Goyal, 2015). Simultaneously, the radical‐scavenging ability of DPPH of strain Y74T was higher than Lactobacillus plantarum NRRL B‐4496 but slightly lower than Lactobacillus acidophilus NRRL B‐4495 (Das & Goyal, 2015; Kaizu, Sasaki, Nakajima, & Suzuki, 1993). This indicates that the antioxidant activity of strain Y74T had reached a practical level. The species of Planococcus genus were always isolated from some extreme environments, such as the Arctic permafrost or the Antarctic (Mykytczuk et al., 2012; Reddy et al., 2002). Therefore, these strains have strong stress‐resistant abilities generally. Previous research found a new antioxidant in Planococcus (Shindo & Misawa, 2014). The species of Planococcus genus had the potential for antioxidant applications, but few studies were emphasized on this property.
Figure 4.
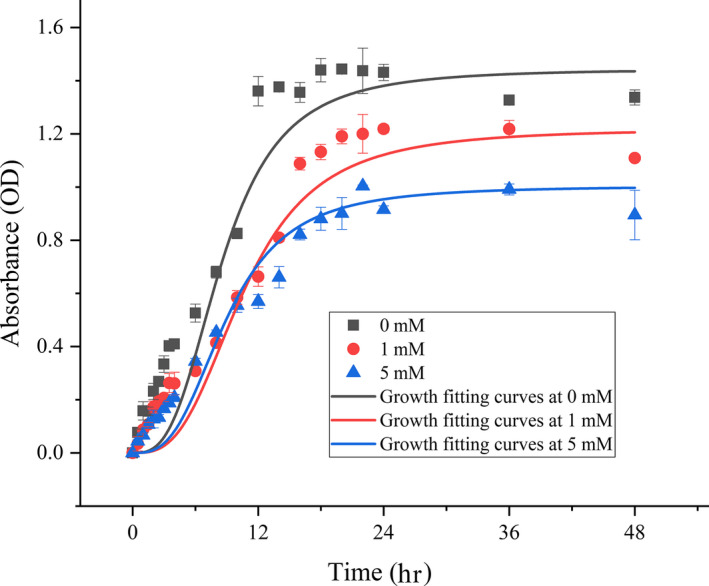
The growth curves of strain Y74T cultivated on LB medium with 0, 1, and 5 mM H2O2
3.5. Genome properties
The draft genome of strain Y74T was 3,672,033 bp (Table 4). The G + C content of the DNA of Y74T was 44.5 mol%. A total of 3,831 genes were detected in strain Y74T, 3,668 of which were protein‐coding genes. There were 967 protein‐coding genes containing enzymes. The number of protein‐coding genes with a function prediction was 2,913. The number of protein‐coding genes with COGs was 3,006, which accounted for 78.47% of all genes. The genome of strain Y74T contained 106 RNA genes, including 67 tRNA genes, 10 5S rRNA genes, 7 16S rRNA genes, and 12 23S rRNA genes (Table 4). The copy number of the rRNA operons in prokaryotic organisms is generally thought to be related to growth rates (Klappenbach, Dunbar, & Schmidt, 2000). It could be suggested that strain Y74T could grow rapidly at lower temperatures. Multiple copy numbers of key genes could increase the radiation resistance of the bacteria as well (Slade & Radman, 2011). There are six peroxidase genes in the genome of Y74T (Table A1), including two glutathione peroxidases, one catalase family peroxidase, one heme‐dependent peroxidase, one thioredoxin‐dependent thiol peroxidase, and one thiol peroxidase. Peroxidases are enzymes that catalyze the oxidation of substrates by hydrogen peroxide as an electron acceptor (Welinder, 1992). This property may explain why the strain can grow in a medium containing hydrogen peroxide. Genes with antioxidant abilities, DNA‐protecting protein (DprA), and superoxide dismutase were also found in the genome of Y74T.
Table 4.
Summary of Planococcus antioxidans Y74T
| Feature | Genome | |
|---|---|---|
| Value | % of total | |
| Size (bp) | 3,672,033 | 100 |
| Coding region (bp) | 3,150,934 | 85.81 |
| Total genes | 3,831 | 100 |
| RNA genes | 106 | 2.77 |
| tRNA | 67 | 1.75 |
| 5S rRNA | 10 | 0.26 |
| 16S rRNA | 7 | 0.18 |
| 23S rRNA | 12 | 0.31 |
| Other RNA genes | 10 | 0.26 |
| Protein‐coding genes | 3,668 | 95.75 |
| Protein‐coding genes with function prediction | 2,913 | 76.04 |
| Protein‐coding genes with enzymes | 967 | 25.24 |
| Protein‐coding genes coding signal peptides | 178 | 4.65 |
| Protein‐coding genes coding transmembrane proteins | 987 | 25.76 |
| Protein‐coding genes with COGs | 3,006 | 78.47 |
In addition to the core proteins and other orthologs presenting in the organism, some nonortholog proteins were found in the strain of Y74T. Therefore, the gene cluster of strain Y74T was analyzed by the method of Bertelli et al. (2017). The results showed that multiple horizontal gene transfer events were found in the Y74T genome. There were 28 gene islands be found in the genome, which contained 395 genes ranging from 4,000 to 700,000 bp, including 162 unclear functional genes of them were annotated as hypothetical proteins; 16 genes were predicted to be recombinase and phage‐associated proteins (Figure 5, Table A2). Based on the gene function analysis on gene island, most of the genes were involved in metabolism, signal transduction, and DNA repair.
Figure 5.

Characteristics and position of the predicted genomic islands found in the draft genome sequence of strain Y74T. The genomic islands show that several horizontal gene transfer events have occurred in strain Y74T. The red in the circles represents the prediction from integrating three different methods (IslandPath‐DIMOB, SIGI‐HMM, and IslandPick); the orange represents the prediction result using IslandPath‐DIMOB; the dark blue represents the prediction result using SIGI‐HMM
4. CONCLUSIONS
According to an analysis of phenotypic, phylogenetic, and chemotaxonomic characteristics, strain Y74T was determined to be a new member within the genus Planococcus. Therefore, it was named as Planococcus antioxidans sp. nov. Y74T. Strain Y74T was found to have potent antioxidant activity via its hydrogen peroxide tolerance and its DPPH radical‐scavenging activity.
4.1. Description of Planococcus antioxidans sp. nov
Planococcus antioxidans (an.ti.o'xi.dans. gr. pref. anti, against; N.L. v. oxidare, to oxidize; N.L. part. adj. antioxidans, referring to the characteristic of this strain).
The colonies on LB agar (Oxoid) are circular, smooth, and white. The cells are aerobic, Gram‐stain‐positive, non‐spore‐forming, and occur as cocci, short rods, or rods (0.8–1.1 × 0.8–3.4 μm). Growth occurs between 4 and 42°C (optimum temperature 28°C), pH values of 6–8.5 and 0%–7% (w/v) NaCl. Strain Y74T utilizes D‐fructose, D‐galactose, D‐glucose, D‐lactose, or D‐maltose as sole carbon sources, weakly utilizes D‐cellobiose, D‐lactose, D‐mannitol, D‐mannose, D‐melibiose, D‐raffinose, L‐rhamnose, D‐sorbitol, D‐trehalose, or myo‐inositol, and does not utilize starch or sucrose. Strain Y74T shows positively for catalase and for gelatin hydrolysis, and negativity for oxidase, nitrate reduction, methyl red, and the Voges–Proskauer tests. In assays with the API ZYM system, strain Y74T is weakly positive for α‐glucosidase, cystine arylamidase, esterase lipase, leucine arylamidase, naphthol‐AS‐BI‐phosphohydrolase, and valine arylamidase, and negative for α‐chymotrypsin, α‐fucosidase, α‐galactosidase, α‐mannosidase, acid phosphatase, alkaline phosphatase, β‐galactosidase, β‐glucosidase, β‐glucuronidase, esterase, lipase, N‐acetyl‐β‐glucosaminidase, and trypsin. The cell wall contains ribose. The peptidoglycan type was L‐Lys‐D‐Glu. The dominant quinones are MK‐8 and MK‐7. The polar lipids are diphosphatidylglycerol, phosphatidylethanolamine, phosphatidylglycerol, and an unknown phospholipid. The majority of the fatty acid is anteiso‐C15:0 (28.8%), followed by C16:1 ω7c alcohol (20.9%) and iso‐C14:0 (13.4%).
The type strain, Y74T (=JCM 32826T = CICC24461T), was isolated from the sandy soil in the town of Huatugou, Qinghai province, China. The G + C content of the DNA of strain Y74T is 44.5 mol%.
CONFLICT OF INTEREST
None declared.
AUTHOR CONTRIBUTION
Binglin Zhang: Conceptualization‐Equal, Software‐Equal, Writing‐review & editing‐Equal; Ruiqi Yang: Investigation‐Equal, Resources‐Equal; Gaosen Zhang: Methodology‐Equal, Resources‐Equal; Yang Liu: Formal analysis‐Equal; Dongming Zhang: Formal analysis‐Equal, Software‐Equal; Wei Zhang: Methodology‐Equal; Tuo Chen: Conceptualization‐Equal, Writing‐review & editing‐Equal; Guangxiu Liu: Conceptualization‐Equal, Writing‐review & editing‐Equal.
ETHICS STATEMENT
None required.
ACKNOWLEDGMENTS
The research was funded by the Bureau of International Cooperation, Chinese Academy of Sciences (131B62KYSB20160014), and the National Natural Science Foundation of China (No. 41801045, 31570498), CAS "Light of West China" Program.
Appendix A.
Figure A1.

Polar lipid profiles of strain Y74T separated by two‐dimensional thin‐layer chromatography and detected with molybdatophosphoric acid (a), molybdenum blue (b), ninhydrin (c) and alpha‐naphthol (d). The solvent systems used were as following: Direction 1 was chloroform/methyl alcohol/H2O (65/25/4, by vol.), Direction 2 was chloroform/ acetic acid/methyl alcohol/H2O (80/15/12/4, by vol.). Abbreviations: DPG, diphosphatidylglycerol; PE, phosphatidylethanolamine; PG, phosphatidylglycerol; UPL, unidentified phospholipid
Table A1.
Antioxidant related genes of strain Y74T
| Gene | Protein | Length | DNA sequence |
|---|---|---|---|
| cds_RLQ92685 | DNA‐protecting protein DprA | 906 | ATGGATTCATTGTTTGAACAAAGACTGATGGCATTGCATTATGTATACCCTCAACCACTCAACCGCATAAAACGGCTGATGATTGACGATTCGAATCTTGAACATTTGGAAAGCAGGCCAGCCTGGGAAATCAGCCAATTACTCGGCATAAAGCCGGAAGCGGCGATATCACTGAAGGATGCTTATAGAAAATCACTGAACAACCCCTATTCTGAAACTTATGAAAAACATAAGATAATCCCCATATCTTATAACCATCCCAATTATCCACAAAGTCTATTTCATCTGATCGACCCACCTGTAATTCTTTACGCCAAAGGAAAAATAGAGTACTTGCTGAATGAAGATCGGATAGCTGTGATAGGTGCCCGTAAGGCTTCTGTTTATTCACAGAAAGCTATGGATCTTATACTTCCTGATCTTGTTGCAGCGGGCTTTATCGTGGTAAGCGGCTTGGCAAAAGGGGCAGATGCAATGGCTCACCGGACAGCAATCGATTGCGGCGGCAAAACGATTGCTGTTACCGGCAGCGGCTTTTTGCATCCGTATCCGAAAGAGAATGATGAATTGAATATTATAATAGAAGAAACTCAACTCGCAATTACAGAATATCCGCCATATATGCAGCCGAAACGCTGGAATTTCCCTATGCGGAACCGCATTATAAGCGGCTTGGCAAAAGGGGTACTGGTAACGGAAGCGGAAGTGAAAAGCGGCACGCTCAGCACGATTGAACATGCCCTGGAACACGGCAAGGATATTTTTGCGGTACCAGGGAGTATCTGTTCACCTCTGTCAGCCGGGCCGCATAAACTGATTTTTGAAGGTGCAAAACCGGTCTGGAATGGGCTTCAAGTGCTGGAGGAATACCGTGAAATTAGGGCTTTAAATAAGTCGATAAAATGA |
| cds_RLQ92091 | Superoxide dismutase | 609 | ATGGCTTATGAATTACCGGAACTACCTTACGCGTATGACGCACTGGAACCACACATCGACAAAGAGACGATGAATATTCACCACACAAAACACCATAATACTTACGTAACCAATGTTAACGCTGCCCTGGAAGGCCACGAAGATCTTTCTTCAAAATCTGTAGAAGAACTGATTTCTGACTTGAACGCTGTGCCTGAAGATATTCGTACAGCTGTACGCAATAACGGCGGTGGACACGCAAACCACTCATTATTCTGGCAATTATTGACTCCAAACGGCACTGGCGCTCCATCAGGTGCACTTGCGGAAGCAATCGACAGCAAGTTCGGCAGCTTTGACGAATTCAAAACGAAATTCGAAGCAGCCGGTAAAACACGCTTCGGTTCAGGCTGGGCTTGGCTTGTTGTATCTAATGGTGAATTGGAAGTAACTTCAACTGCCAACCAGGATTCTCCATTGATGGACGGCAAAACGCCAATTCTTGGAGTAGACGTTTGGGAGCATGCTTACTACTTGAAATACCAGAACAAACGCCCTGACTATTTGGCTGCTTTCTGGAACGTAGTAAACTGGGACGAAGTTTCAAAACGCTATGAAGCTGCAAAATAA |
| cds_RLQ91554 | Glutathione peroxidase | 477 | ATGAGTATTTATGAATTTTCGGCCAGAAAGTCCGATGGCAGCATTTATCCGTTAAGTGAATACGAAGGGAAAACGATGTTGATTGTCAATACTGCTACGAAATGCGGGTTGCGTGATCAATTCGATGGACTGGAGAAGTTGTACCAGAAGTATGAAGATGACGGACTTGTCGTCCTTGGCTTTCCTTCCGATCAGTTCGGGCAGGAACTTGATGGGGCGAAAGAAGCGGAGGAATCCTGCCGGATGACTTACGGAGTTTCGTTCCCAATGCATGACCTGGTCAAAGTTAACGGAAAAAATGCCGATCCTCTTTTTAAATATCTAACTGAAAACAGCAAGGGAGTCCTTGGCAGCAGCATCAAGTGGAATTTCACAAAATTTCTCATCAACAAAGAGGGAAAGCTGGTTGCACGGTTTTCGCCAAAAGATAAACCTGAAAAATTTGAAGAAGAAATTAAACAATACTTAACGAACTGA |
| cds_RLQ90872 | Catalase family peroxidase | 927 | ATGGCGAAGGAAAAGCTTGCGGAAACCGCAGTCAATAAAATCGAGAAAGTGTTTGGGGAACATAAAACTTATAGACGTGCGCATTCAAGAGGAACGGGATATGAAGCCCTATTCACAGCAAACGGCGAAGGGCAGAATTGGACCGTCGCGCAGCATCTCCGGGAAGGGACGACCAAAGCTGTGGTGCGATTTTCGCACAGTTCTCCAGATCCATTTTGGACGGATAATTTATCACCGGTAAAAGGAATGGCTGTGCAATTCCAGTTGCCGGATGGCCAAGTGATGAACAGTGTCGGCGTAACTTCCCCGATATTCTTTTCGCGCACTCCGGAAGTGTTTACGGAAATGCTGGATATCGCGAAATCGTTTAAAAAGGGCAAGCCCCGGCTGCGGGATCTCATCAAATTGTTCATCAAATATCCCGAAAGCCGAGCAGCAATCCGCATCATCCGGAAAATGCAGAGTCCGGCTAGTTTCGCGACCGGTCTCTACCATTCCATTCACGCATTTTACCTGGTTAACGGTACTGGCCAGCGCGTACCCGTCAAATTCCAGTGGCATCCGGAAGCGGGCGTGGAGTCGTTGAATCCGGTGGAGGCTGCGTCAGTGAAAAAAGGAGATTTTGAGGAAGAGCTTGAAGAACGCGTCTTGAGCGGAGAGACGGCTTTCCGTCTGATGGCAGTGATTGGGGATGCGGATGACCCTGTAGATGACCCGACGAAAGACTGGGCTAAAGATAGAAAGAAAGTCGAATTGGGGCGCCTGGTGCTGAAAGAACAGACTGACGAAGCGGAAGGGTTGCTGTATGATCCAACCATCCTGGCAGAAGGCGTCGAATGCACGGATGATCCCATCCTGCATTTCCGCAACCCGGCATATGCTATTTCTTATATGCGGAGAGAAGGGGAGAAGCAAAAAGAAAGTTGA |
| cds_RLQ90525 | Heme‐dependent peroxidase | 750 | ATGAATGAAGCAGCAATCACTTTAGACGGCTGGTACGTCCTCCACGATTTCCGTTCGATGGACTGGGTATCATGGAAAATGCTTGAAGACGAAGAACGCCAATTCGCAATCGACGAATATCAGGCATTCATGGACAAAGTAAACCAGGCCGATGAAAATAAAACCGGTGCACACGCATTGTATTCAATTATTGGCCAAAAAGCTGACTTGATGCTGATGCTATTGCGCGAAACTATGGACGAATTGCGTGAACTTGAAACGGAATACAATAAGCTGACATTGGTCGCTTACACGGTTCCGACTTACTCTTACGTATCTGTAGTGGAACTTTCCAACTATCTTGCAGGTAAATCAGAAGAAGATCCATACCAGAACCCGCATGTCCGCGCGCGTCTGTATCCGGAGCTTCAGCGTTCGCAGTACATCTGCTTCTACCCGATGGACAAGCGCCGCGACGGCAACGACAACTGGTACATGCTGCCGATGGACGAGCGCAAAGATTTGATGCTGTCACACGGCAAAATCGGCCGCAGCTACGCAGGCAAAGTAAAACAGATCATTTCCGGCTCTGTCGGCTTTGATGATTACGAATGGGGCGTAACCTTGTTCGCAGATGACGTTCTGCAGTTTAAAAAACTGATCTATGAAATGCGTTTTGACGAAGTCAGCGCGCGTTACGCTGAATTCGGTTCGTTCTACGTCGGCACTCGCCTTGATAAAGAAAGAATCGTTAAGTTTTTGGAAGTTTAA |
| cds_RLQ82443 | Thioredoxin‐dependent thiol peroxidase | 477 | TTGACAACATTAGAAGGTTTGCATGCACCGGATTTTACATTGAAAAATGAAAACGGCGAAACAGTTTCTTTGGAGGATTTTGCCGGCAAAAAATACGTAGTGCTTTATTTTTACCCGAAAGATATGACACCGGGCTGCACTACACAGGCCTGCGATTTCCGGGATGCAGAGAAGGATTTTTCCGAATTGGGAGCAGTCATTCTTGGCGTTAGCGCAGACTCTGAAAAACAGCACAGTAAATTTATCAGCAAACACGGTTTGCCATTCTCTTTATTGGTTGACGAAGATCATAAAGTTTCTGAGGCATACGGTGTGTGGGTGGAGAAGAAGATGTACGGAAAAGAATTTATGGGGATTGAACGCTCTACATTTTTAATCGACCCAACCGGGACTGTCGTAAAAGAATGGCGAAAAGTCAAAGTGAAAGACCATATCCAGGAAGTCCTTGAAACGGTCAAAGAGCTCAGCCAAGCATAG |
| cds_RLQ81587 | Glutathione peroxidase | 477 | ATGGTTTCGGTTTATGATTACAAAGTTAAAAATTTGCAGGGAGAAATGGAATCACTTGAAAAGTTTAAAGGGAATGCGCTGGTAATAGTCAATACAGCAAGCAAATGTGGATTGACTCCTCAATTTGAAGACCTTCAAAAACTCTATGAAAAGTATTCCAGTAAAGATTTTCAAATCCTCGGTTTTCCAAGTTCTCAATTTAATAATCAGGAATTTGAAAATCAGGAAGAAACGATGGAATTCTGCCAGATGAATTATGGTGTAACATTTCCTATGTTTGCAAAAACAGATGTCAAAGGAGCAGATGCAGCTCCACTATTTACCTATCTGACTTCAAAGCATGAAAATCTAGAGGCTGAAGAAATTGCATGGAACTTTGCGAAGTTTTTAATAGATAAAGAAGGACATGTCATTAAAAGATATTCCCCTCGATCTTCGCCACTGGAAATTGAAGAAGACCTGAAGACTATTCTATAA |
| cds_RLQ91872 | Thiol peroxidase | 507 | ATGGTACAAGTTACATTTCATGAAAATCCTGTTACCTTACCAAACAAAGAAGTCAAAGTTGGAGACCAAATCCCGAATTTCACAGTACTTGACAATGACTTGAACCCTGTAACTGCACAGGATACAGCAGGTAAAGTCAGATTATTCACAGTGTCACCATCCTTGGATACAAGTGTTTGTTCAGATCAGGCGAAACGTTTCAGCGAAGAAGCTTCATCAATGGGAGACGAAGTTGCAATCTACTATGTAACCTGCGATTTGCCTTTCGCACAAAAGCGTTGGGTTGAAGTTAATGAAGCCAAAAATCTCACTACTCTTTCCGATCACCGTGATCTTTCTTTCGGTGAAGCGTTCGGAGTAACGATGCAGGAACTGCGTTTGCTGGCCCGTTCCATATTTGTAGTGGATGAAAACGATAAAGTGACTTATGTGGAATATGTTCCTGAGGGAACGAATCATCCGAATTACGATAAAGCGATTGAAGCAGTAAAAGAACTGACAAAATAA |
Table A2.
Characteristics of the genomic islands found in the genome of strain Y74T
| GI | Length | Total no. of gene | hypothetical proteins | Integrase/ phage | Predicted function |
|---|---|---|---|---|---|
| 1 | 4,681 | 6 | 2 | 0 | Metabolism/ Transcription factors |
| 2 | 6,367 | 8 | 2 | 0 | |
| 3 | 7,602 | 6 | 3 | 0 | General function prediction only/ Metabolism |
| 4 | 4,374 | 3 | 2 | 0 | Metabolism |
| 5 | 5,080 | 6 | 4 | 0 | |
| 6 | 6,976 | 5 | 3 | 0 | Replication and repair |
| 7 | 21,818 | 19 | 7 | 1 | Genetic information processing /Transcription factors/Cell processing/ Nucleotide metabolism |
| 8 | 12,222 | 10 | 6 | 1 | Transcription factors/DNA repair |
| 9 | 14,623 | 20 | 14 | 0 | Secretion system/DNA repair |
| 10 | 11,232 | 14 | 11 | 0 | |
| 11 | 16,787 | 17 | 4 | 2 | Transporters/Carbohydrate metabolism/ Transferases/Two‐component system |
| 12 | 42,958 | 29 | 4 | 2 | Two‐component system/ Metabolism/Transcription factors |
| 13 | 10,266 | 7 | 1 | 0 | Metabolism/DNA repair |
| 14 | 4,777 | 7 | 1 | 0 | |
| 15 | 4,674 | 5 | 3 | 0 | |
| 16 | 11,402 | 9 | 4 | 0 | |
| 17 | 9,706 | 11 | 4 | 1 | Metabolism/Transcription factors/ Transferases /Signal transduction |
| 18 | 53,470 | 60 | 25 | 0 | |
| 19 | 4,656 | 6 | 0 | 0 | Metabolism/Membrane transport |
| 20 | 75,201 | 74 | 30 | 2 | Secretion system/Metabolism/Transcription factors/Membrane transport |
| 21 | 11,374 | 13 | 10 | 1 | Genetic information processing |
| 22 | 13,041 | 16 | 6 | 1 | Metabolism/Transcription factors |
| 23 | 5,465 | 9 | 4 | 0 | Transcription factors/Genetic information processing |
| 24 | 5,113 | 6 | 1 | 0 | Transcription factors/Transferases |
| 25 | 6,724 | 10 | 3 | 0 | |
| 26 | 4,021 | 6 | 3 | 0 | |
| 27 | 6,818 | 8 | 4 | 2 | Transcription factors |
| 28 | 10,911 | 5 | 1 | 3 | |
| Total | 392,339 | 395 | 162 | 16 |
Zhang B, Yang R, Zhang G, et al. Characteristics of Planococcus antioxidans sp. nov., an antioxidant‐producing strain isolated from the desert soil in the Qinghai–Tibetan Plateau. MicrobiologyOpen. 2020;9:e1028 10.1002/mbo3.1028
Binglin Zhang and Ruiqi Yang contributed equally to this work.
Contributor Information
Tuo Chen, Email: chentuo@lzb.ac.cn.
Guangxiu Liu, Email: liugx@lzb.ac.cn.
DATA AVAILABILITY STATEMENT
All data are provided in full in the results section of this paper apart from the DNA sequences. The 16S rRNA gene sequence of strain Y74T is available in the GenBank database under accession number KU601236. The genome of strain Y74T is deposited in GenBank with accession number RCWH00000000. The type strain Y74T is deposited in the Japan Collection of Microorganisms (https://www.jcm.riken.jp/cgi‐bin/jcm/jcm_number?JCM=32826) and the China Center of Industrial Culture Collection (http://sales.china‐cicc.org/category.php?id=1&sh=jd&keywords=24461).
REFERENCES
- Alam, S. I. , Singh, L. , Dube, S. , Reddy, G. S. N. , & Shivaji, S. (2003). Psychrophilic Planococcus maitriensis sp. nov from Antarctica. Systematic and Applied Microbiology, 26, 505–510. 10.1078/072320203770865792 [DOI] [PubMed] [Google Scholar]
- Berdy, J. (2005). Bioactive microbial metabolites. Journal of Antibiotics, 58, 1–26. [DOI] [PubMed] [Google Scholar]
- Bertelli, C. , Laird, M. R. , Williams, K. P. , Simon Fraser University Research Computing Group , Lau, B. Y. , Hoad, G. H. , … Brinkman, F. S. (2017). IslandViewer 4: Expanded prediction of genomic islands for larger‐scale datasets. Nucleic Acids Research, 45(W1), W30–W35. 10.1093/nar/gkx343 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonda, D. J. , Wang, X. L. , Perry, G. , Nunomura, A. , Tabaton, M. , Zhu, X. W. , & Smith, M. A. (2010). Oxidative stress in Alzheimer disease: A possibility for prevention. Neuropharmacology, 59, 290–294. 10.1016/j.neuropharm.2010.04.005 [DOI] [PubMed] [Google Scholar]
- Brettin, T. , Davis, J. J. , Disz, T. , Edwards, R. A. , Gerdes, S. , Olsen, G. J. , … Xia, F. (2015). RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Scientific Reports, 5, 8365 10.1038/srep08365 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, I. M. A. , Chu, K. , Palaniappan, K. , Pillay, M. , Ratner, A. , Huang, J. H. , … Kyrpides, N. C. (2019). IMG/M vol 5.0: An integrated data management and comparative analysis system for microbial genomes and microbiomes. Nucleic Acids Research, 47, D666–D677. 10.1093/nar/gky901 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi, J. H. , Im, W. T. , Liu, Q. M. , Yoo, J. S. , Shin, J. H. , Rhee, S. K. , & Roh, D. H. (2007). Planococcus donghaensis sp. nov., a starch‐degrading bacterium isolated from the East Sea, South Korea. International Journal of Systematic and Evolutionary Microbiology, 57, 2645–2650. 10.1099/ijs.0.65036-0 [DOI] [PubMed] [Google Scholar]
- Chun, J. , Oren, A. , Ventosa, A. , Christensen, H. , Arahal, D. R. , da Costa, M. S. , … Trujillo, M. E. (2018). Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. International Journal of Systematic and Evolutionary Microbiology, 68(1), 461–466. 10.1099/ijsem.0.002516 [DOI] [PubMed] [Google Scholar]
- Collins, M. D. , Pirouz, T. , Goodfellow, M. , & Minnikin, D. E. (1977). Distribution of menaquinones in actinomycetes and corynebacteria. Journal of General Microbiology, 100, 221–230. 10.1099/00221287-100-2-221 [DOI] [PubMed] [Google Scholar]
- Dai, X. , Wang, Y. N. , Wang, B. J. , Liu, S. J. , & Zhou, Y. G. (2005). Planomicrobium chinense sp. nov., isolated from coastal sediment, and transfer of Planococcus psychrophilus and Planococcus alkanoclasticus to Planomicrobium as Planomicrobium psychrophilum comb. nov and Planomoicrobium alkanoclasticum comb. nov. International Journal of Systematic and Evolutionary Microbiology, 55, 699–702. 10.1099/ijs.0.63340-0 [DOI] [PubMed] [Google Scholar]
- Das, D. , & Goyal, A. (2015). Antioxidant activity and γ‐aminobutyric acid (GABA) producing ability of probiotic Lactobacillus plantarum DM5 isolated from Marcha of Sikkim. LWT‐food Science and Technology, 61(1), 263–268. 10.1016/j.lwt.2014.11.013 [DOI] [Google Scholar]
- Felsenstein, J. (1981). Evolutionary trees from DNA sequences ‐ a maximum‐likelihood approach. Journal of Molecular Evolution, 17, 368–376. [DOI] [PubMed] [Google Scholar]
- Felsenstein, J. (1985). Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39, 783–791. 10.1111/j.1558-5646.1985.tb00420.x [DOI] [PubMed] [Google Scholar]
- Fischer, R. , & Maier, O. (2015). Interrelation of oxidative stress and inflammation in neurodegenerative disease: Role of TNF. Oxidative Medicine and Cellular Longevity, 2015, 1–18. 10.1155/2015/610813 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gan, L. Z. , Zhang, H. M. , Tian, J. W. , Li, X. G. , Long, X. F. , Zhang, Y. Q. , … Tian, Y. Q. (2018). Planococcus salinus sp. nov., a moderately halophilic bacterium isolated from a saline‐alkali soil. International Journal of Systematic and Evolutionary Microbiology, 68, 589–595. 10.1099/ijsem.0.002548 [DOI] [PubMed] [Google Scholar]
- Gan, L. Z. , Zhang, Y. , Zhang, L. L. , Li, X. G. , Wang, Z. K. , He, L. L. , … Tian, Y. Q. (2018). Planococcus halotolerans sp nov., isolated from a saline soil sample in China. International Journal of Systematic and Evolutionary Microbiology, 68, 3500–3505. 10.1099/ijsem.0.003019 [DOI] [PubMed] [Google Scholar]
- Gil, R. , Silva, F. J. , Pereto, J. , & Moya, A. (2004). Determination of the core of a minimal bacterial gene set. Microbiology Molecular Biology Reviews, 68, 518–537. 10.1128/MMBR.68.3.518-537.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hao, M. V. , Kocur, M. , & Komagata, K. (1984). Marinococcus Gen‐Nov, a new genus for motile Cocci with Meso‐Diaminopimelic acid in the cell‐wall ‐ and Marinococcus‐Albus Sp‐Nov and Marinococcus‐Halophilus (Novitsky and Kushner) Comb‐Nov. Journal of General and Applied Microbiology, 30, 449–459. 10.2323/jgam.30.449 [DOI] [Google Scholar]
- Hao, M. V. , & Komagata, K. (1985). A new species of Planococcus, Planococcus‐Kocurii isolated from fish, frozen foods, and fish curing brine. Journal of General and Applied Microbiology, 31, 441–455. 10.2323/jgam.31.441 [DOI] [Google Scholar]
- Jukes, T. H. , & Cantor, C. R. (1969). Evolution of protein molecules. Mammalian Protein Metabolism, 3, 21–132. [Google Scholar]
- Jung, Y. T. , Kang, S. J. , Oh, T. K. , Yoon, J. H. , & Kim, B. H. (2009). Planomicrobium flavidum sp. nov., isolated from a marine solar saltern, and transfer of Planococcus stackebrandtii Mayilraj et al 2005 to the genus Planomicrobium as Planomicrobium stackebrandtii comb. nov. International Journal of Systematic and Evolutionary Microbiology, 59, 2929–2933. 10.1099/ijs.0.009191-0 [DOI] [PubMed] [Google Scholar]
- Kaizu, H. , Sasaki, M. , Nakajima, H. , & Suzuki, Y. (1993). Effect of antioxidative lactic acid bacteria on rats fed a diet deficient in vitamin E. Journal of Dairy Science, 76, 2493–2499. 10.3168/jds.S0022-0302(93)77584-0 [DOI] [PubMed] [Google Scholar]
- Kämpfer, P. , & Kroppenstedt, R. M. (1996). Numerical analysis of fatty acid patterns of coryneform bacteria and related taxa. Canadian Journal of Microbiology, 42, 989–1005. 10.1139/m96-128 [DOI] [Google Scholar]
- Kaur, L. , Das, A. P. , Acharya, M. , Klenk, H. P. , Sree, A. , & Mayilraj, S. (2012). Planococcus plakortidis sp. nov., isolated from the marine sponge Plakortis simplex (Schulze). International Journal of Systematic and Evolutionary Microbiology, 62, 883–889. 10.1099/ijs.0.029967-0 [DOI] [PubMed] [Google Scholar]
- Kim, J. H. , Kang, H. J. , Yu, B. J. , Kim, S. C. , & Lee, P. C. (2015). Planococcus faecalis sp. nov., a carotenoid‐producing species isolated from stools of Antarctic penguins. International Journal of Systematic and Evolutionary Microbiology, 65, 3373–3378. 10.1099/ijsem.0.000423 [DOI] [PubMed] [Google Scholar]
- Kim, M. , Oh, H. S. , Park, S. C. , & Chun, J. (2014). Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. International Journal of Systematic and Evolutionary Microbiology, 64, 346–351. 10.1099/ijs.0.059774-0 [DOI] [PubMed] [Google Scholar]
- Kim, O. S. , Cho, Y. J. , Lee, K. , Yoon, S. H. , Kim, M. , Na, H. , … Chun, J. (2012). Introducing EzTaxon‐e: A prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species. International Journal of Systematic and Evolutionary Microbiology, 62, 716–721. 10.1099/ijs.0.038075-0 [DOI] [PubMed] [Google Scholar]
- Klappenbach, J. A. , Dunbar, J. M. , & Schmidt, T. M. (2000). rRNA operon copy number reflects ecological strategies of bacteria. Applied and Environmental Microbiology, 66, 1328–1333. 10.1128/AEM.66.4.1328-1333.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kroppenstedt, R. M. (1982). Separation of bacterial menaquinones by HPLC using reverse phase (RP18) and a silver loaded ion exchanger as stationary phases. Journal of Liquid Chromatography, 5, 2359–2367. 10.1080/01483918208067640 [DOI] [Google Scholar]
- Kumar, S. , Stecher, G. , Li, M. , Knyaz, C. , & Tamura, K. (2018). MEGA X: Molecular evolutionary genetics analysis across computing platforms. Molecular Biology and Evolution, 35, 1547–1549. 10.1093/molbev/msy096 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurup, P. V. , & Schmitt, J. A. (1973). Numerical taxonomy of Nocardia. Canadian Journal of Microbiology, 19, 1035–1048. 10.1139/m73-164 [DOI] [PubMed] [Google Scholar]
- Larkin, M. A. , Blackshields, G. , Brown, N. P. , Chenna, R. , McGettigan, P. A. , McWilliam, H. , … Higgins, D. G. (2007). Clustal W and Clustal X version 2.0. Bioinformatics, 23, 2947–2948. 10.1093/bioinformatics/btm404 [DOI] [PubMed] [Google Scholar]
- Le, H. , & Cao, Y. (2010). Lactic acid bacterial cell factories for gamma‐aminobutyric acid. Amino Acids, 39, 1107–1116. 10.1007/s00726-010-0582-7 [DOI] [PubMed] [Google Scholar]
- Lechevalier, M. P. , & Lechevalier, H. (1970). Chemical composition as a criterion in the classification of aerobic actinomycetes. International Journal of Systematic Bacteriology, 20, 435–443. 10.1099/00207713-20-4-435 [DOI] [Google Scholar]
- Lee, B. J. , Kim, J. S. , Kang, Y. M. , Lim, J. H. , Kim, Y. M. , Lee, M. S. , … Je, J. Y. (2010). Antioxidant activity and γ‐aminobutyric acid (GABA) content in sea tangle fermented by Lactobacillus brevis BJ20 isolated from traditional fermented foods. Food Chemistry, 122, 271–276. 10.1016/j.foodchem.2010.02.071 [DOI] [Google Scholar]
- Lin, M. T. , & Beal, M. F. (2006). Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature, 443, 787–795. 10.1038/nature05292 [DOI] [PubMed] [Google Scholar]
- Luo, X. N. , Zhang, J. L. , Li, D. , Xin, Y. H. , Xin, D. , & Fan, L. (2014). Planomicrobium soli sp. nov., isolated from soil. International Journal of Systematic and Evolutionary Microbiology, 64, 2700–2705. 10.1099/ijs.0.055426-0 [DOI] [PubMed] [Google Scholar]
- Meier‐Kolthoff, J. P. , Auch, A. F. , Klenk, H. P. , & Goker, M. (2013). Genome sequence‐based species delimitation with confidence intervals and improved distance functions. BMC Bioinformatics, 14, 60 10.1186/1471-2105-14-60 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Migula, W. (1895). Über ein neues System der Bakterien. [Google Scholar]
- Minnikin, D. E. , O'Donnell, A. G. , Goodfellow, M. , Alderson, G. , Athalye, M. , Schaal, A. , & Parlett, J. H. (1984). An integrated procedure for the extraction of bacterial isoprenoid quinones and polar lipids. Journal of Microbiological Methods, 2, 233–241. 10.1016/0167-7012(84)90018-6 [DOI] [Google Scholar]
- Mykytczuk, N. C. S. , Wilhelm, R. C. , & Whyte, L. G. (2012). Planococcus halocryophilus sp. nov., an extreme sub‐zero species from high Arctic permafrost. International Journal of Systematic and Evolutionary Microbiology, 62, 1937–1944. 10.1099/ijs.0.035782-0 [DOI] [PubMed] [Google Scholar]
- Novitsky, T. J. , & Kushner, D. J. (1976). Planococcus halophilus sp. nov, a Facultatively Halophilic coccus. International Journal of Systematic Bacteriology, 26, 53–57. 10.1099/00207713-26-1-53 [DOI] [Google Scholar]
- Reddy, G. S. N. , Prakash, J. S. S. , Vairamani, M. , Prabhakar, S. , Matsumoto, G. I. , & Shivaji, S. (2002). Planococcus antarcticus and Planococcus psychrophilus spp. nov isolated from cyanobacterial mat samples collected from ponds in Antarctica. Extremophiles, 6, 253–261. 10.1007/s00792-001-0250-7 [DOI] [PubMed] [Google Scholar]
- Richter, M. , Rossello‐Mora, R. , Glockner, F. O. , & Peplies, J. (2016). JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics, 32, 929–931. 10.1093/bioinformatics/btv681 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Romano, I. , Giordano, A. , Lama, L. , Nicolaus, B. , & Gambacorta, A. (2003). Planococcus rifietensis sp. nov, isolated from algal mat collected from a sulfurous spring in Campania (Italy). Systematic and Applied Microbiology, 26, 357–366. 10.1078/072320203322497383 [DOI] [PubMed] [Google Scholar]
- Saitou, N. , & Nei, M. (1987). The neighbor‐joining method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, 4, 406–425. 10.1093/oxfordjournals.molbev.a040454 [DOI] [PubMed] [Google Scholar]
- Sasser, M. (1990). Identification of bacteria by gas chromatography of cellular fatty acids. [Google Scholar]
- Schleifer, K. H. , & Kandler, O. (1972). Peptidoglycan types of bacterial cell walls and their taxonomic implications. Bacteriological Reviews, 36, 407–477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shindo, K. , & Misawa, N. (2014). New and rare carotenoids isolated from marine bacteria and their antioxidant activities. Marine Drugs, 12, 1690–1698. 10.3390/md12031690 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slade, D. , & Radman, M. (2011). Oxidative stress resistance in Deinococcus radiodurans . Microbiology Molecular Biology Review, 75(1), 133–191. 10.1128/MMBR.00015-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suresh, K. , Mayilraj, S. , Bhattacharya, A. , & Chakrabarti, T. (2007). Planococcus columbae sp. nov., isolated from pigeon faeces. International Journal of Systematic and Evolutionary Microbiology, 57, 1266–1271. 10.1099/ijs.0.64742-0 [DOI] [PubMed] [Google Scholar]
- Tamura, K. , Peterson, D. , Peterson, N. , Stecher, G. , Nei, M. , & Kumar, S. (2011). MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution, 28(10), 2731–2739. 10.1093/molbev/msr121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Velho‐Pereira, S. , Parvatkar, P. , & Furtado, I. J. (2015). Evaluation of antioxidant producing potential of halophilic bacterial bionts from marine invertebrates. Indian Journal of Pharmaceutical Sciences, 77, 183–189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, W. P. , Wang, L. P. , & Shao, Z. Z. (2010). Diversity and abundance of oil‐degrading bacteria and alkane hydroxylase (alkB) genes in the subtropical seawater of Xiamen Island. Microbial Ecology, 60(2), 429–439. 10.1007/s00248-010-9724-4 [DOI] [PubMed] [Google Scholar]
- Wang, X. , Wang, Z. , Zhao, X. Q. , Huang, X. , Zhou, Y. , & Li, W. J. (2017). Planococcus ruber sp. nov., isolated from a polluted farmland soil sample. International Journal of Systematic and Evolutionary Microbiology, 67, 2549–2554. 10.1099/ijsem.0.001960 [DOI] [PubMed] [Google Scholar]
- Welinder, K. G. (1992). Superfamily of plant, fungal and bacterial peroxidases. Current Opinion in Structural Biolog, 2(3), 388–393. 10.1016/0959-440X(92)90230-5 [DOI] [Google Scholar]
- Yoon, J. H. , Kang, S. J. , Lee, S. Y. , Oh, K. H. , & Oh, T. K. (2010). Planococcus salinarum sp. nov., isolated from a marine solar saltern, and emended description of the genus Planococcus . International Journal of Systematic and Evolutionary Microbiology, 60, 754–758. 10.1099/ijs.0.013136-0 [DOI] [PubMed] [Google Scholar]
- Yoon, J. H. , Lee, K. C. , Kang, S. S. , Kho, Y. H. , Lee, E. S. , Park, Y. H. , & Kang, K. H. (2001). Planomicrobium koreense gen. nov., sp nov., a bacterium isolated from the Korean traditional fermented seafood jeotgal, and transfer of Planococcus okeanokoites (Nakagawa et al 1996) and Planococcus mcmeekinii (Junge et al 1998) to the genus Planomicrobium . International Journal of Systematic and Evolutionary Microbiology, 51, 1511–1520. 10.1099/00207713-51-4-1511 [DOI] [PubMed] [Google Scholar]
- Yoon, J. H. , Weiss, N. , Kang, K. H. , Oh, T. K. , & Park, Y. H. (2003). Planococcus maritimus sp. nov., isolated from sea water of a tidal flat in Korea. International Journal of Systematic and Evolutionary Microbiology, 53, 2013–2017. 10.1099/ijs.0.02557-0 [DOI] [PubMed] [Google Scholar]
- Zhang, B. , Tang, S. , Chen, X. , Zhang, G. , Zhang, W. , Chen, T. , … Dyson, P. (2018). Streptomyces qaidamensis sp. nov., isolated from sand in the Qaidam Basin, China. The Journal of Antibiotics, 70, 880–886. 10.1038/s41429-018-0080-9 [DOI] [PubMed] [Google Scholar]
- Zhang, B. , Tang, S. , Chen, X. , Zhang, L. , Zhang, G. , Zhang, W. , … Dyson, P. (2016a). Streptomyces lacrimifluminis sp. nov., a novel actinobacterium that produces antibacterial compounds, isolated from soil. International Journal of Systematic and Evolutionary Microbiology, 66, 4981–4986. 10.1099/ijsem.0.001456 [DOI] [PubMed] [Google Scholar]
- Zhang, B. , Tang, S. , Yang, R. , Chen, X. , Zhang, D. , Zhang, W. , … Dyson, P. (2019). Streptomyces dangxiongensis sp. nov., isolated from soil of Qinghai‐Tibet. Plateau. International Journal of Systematic and Evolutionary Microbiology, 69(9), 2729–2734. 10.1099/ijsem.0.003550 [DOI] [PubMed] [Google Scholar]
- Zhang, B. , Wu, X. , Zhang, G. , Li, S. , Zhang, W. , Chen, X. , … Chen, T. (2016b). The diversity and biogeography of the communities of Actinobacteria in the forelands of glaciers at a continental scale. Environmental Research Letters, 11, 054012 10.1088/1748-9326/11/5/054012 [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data are provided in full in the results section of this paper apart from the DNA sequences. The 16S rRNA gene sequence of strain Y74T is available in the GenBank database under accession number KU601236. The genome of strain Y74T is deposited in GenBank with accession number RCWH00000000. The type strain Y74T is deposited in the Japan Collection of Microorganisms (https://www.jcm.riken.jp/cgi‐bin/jcm/jcm_number?JCM=32826) and the China Center of Industrial Culture Collection (http://sales.china‐cicc.org/category.php?id=1&sh=jd&keywords=24461).