

Abstract
Loci associated with longevity are likely to harbor genes coding for key players of molecular pathways involved in a lifelong decreased mortality and decreased/compressed morbidity. However, identifying such loci is challenging. One of the most plausible reasons is the uncertainty in defining long‐lived cases with the heritable longevity trait among long‐living phenocopies. To avoid phenocopies, family selection scores have been constructed, but these have not yet been adopted as state of the art in longevity research. Here, we aim to identify individuals with the heritable longevity trait by using current insights and a novel family score based on these insights. We use a unique dataset connecting living study participants to their deceased ancestors covering 37,825 persons from 1,326 five‐generational families, living between 1788 and 2019. Our main finding suggests that longevity is transmitted for at least two subsequent generations only when at least 20% of all relatives are long‐lived. This proves the importance of family data to avoid phenocopies in genetic studies.
Keywords: aging, family tree, genetics, inheritance, longevity
In this study we show that human longevity is transmitted for at least 2 subsequent generations only when at least 20% of a person’s relatives are long‐lived. The analyses are based on a unique dataset, covering 37,825 persons from 1,326 five‐generational families, in which living study participants are connected to their deceased ancestors. Our findings highlight the importance of using survival information of relatives/ancestors to avoid long‐lived phenocopies in genetic studies and aim to stimulate cohort studies to extend their data with such ancestral information.
1. INTRODUCTION
In contrast to the low heritability of human lifespan (Herskind et al., 1996; Kaplanis et al., 2018; Ruby et al., 2018; van den Berg, Beekman, Smith, Janssens, & Slagboom, 2017), human longevity is strongly heritable as illustrated by the familial clustering of survival into extreme ages (Berg, Rodríguez‐Girondo, et al., 2018; Dutta et al., 2013; Gudmundsson et al., 2000; Houde, Tremblay, & Vézina, 2008; Jarry, Gagnon, & Bourbeau, 2013; Pedersen et al., 2017; Perls et al., 2002; Schoenmaker et al., 2006; Sebastiani, Nussbaum, Andersen, Black, & Perls, 2016; Terry et al., 2004; van den Berg et al., 2019). Identifying loci associated with longevity is important because they likely to harbor genes coding for molecular pathways involved in a lifelong decreased mortality (Berg, Rodríguez‐Girondo, et al., 2018; van den Berg et al., 2019), decreased morbidity (Dutta et al., 2013; Terry et al., 2004), and compression of morbidity toward the end of the lifespan (Andersen, Sebastiani, Dworkis, Feldman, & Perls, 2012; Christensen, McGue, Petersen, Jeune, & Vaupel, 2008; Christensen et al., 2013). Currently, genome‐wide linkage and association studies (GWAS) identified a limited number of loci promoting longevity (Broer et al., 2015; Deelen et al., 2014; Flachsbart et al., 2009; Joshi et al., 2017; Partridge, Deelen, & Slagboom, 2018; Pilling et al., 2017; Sebastiani et al., 2017; Shadyab & LaCroix, 2015; Slagboom, Berg, & Deelen, 2018; Willcox et al., 2008), for example, the APOE and FOXO3A genes (more details can be found in current review papers [Partridge et al., 2018; Shadyab & LaCroix, 2015; Slagboom et al., 2018]). However, many of the identified loci could not be replicated in independent studies as yet. In addition, the largest and most recent longevity GWAS, based on cases belonging to the top 10% oldest survivors, again only replicated association of the APOE locus (Deelen et al., 2019).
One of the main reasons for the limited success of longevity genetic studies (Broer et al., 2015; Deelen et al., 2014, 2019; Flachsbart et al., 2016; Sebastiani et al., 2017; Willcox et al., 2008; Zeng et al., 2016) is the uncertainty in defining the heritable longevity trait itself (Berg et al., 2019; van den Berg et al., 2017). Given the increased life expectancy of the past 200 years due to nongenetic factors (improved hygiene, nutrition, and medication), there are likely many phenocopies among the long‐lived cases selected for our genetic studies (Oeppen & Vaupel, 2002; Vaupel, 1998). The presence of phenocopies is illustrated by the increase in centenarians in the United States between 1994 and 2012 from 1 in 10,000 to 1 in 5,000 (Sebastiani & Perls, 2012). To avoid phenocopies, family selection scores, such as the Family Longevity Selection Score (FLoSS) and the Family Excess Longevity (FEL) score, have been constructed (Kerber, Brien, Smith, & Cawthon, 2001; Sebastiani et al., 2009). The use of such scores is substantiated by novel studies which showed that including family history information can provide valuable information about an individual's genetic liability for a trait and is likely to increase the power to detect genetic loci (Gordon, 2005; Hujoel, Gazal, Loh, Patterson, & Alkes, 2019; Liu, Erlich, & Pickrell, 2017). The scores focus, in different ways, on selecting multiple family members with the same trait (Arbeeva et al., 2018; Berg, Rodríguez‐Girondo, et al., 2018; Kerber et al., 2001; Rozing et al., 2010; Sebastiani et al., 2009), and usually focus on a single group of relatives, such as parents (Berg, Rodríguez‐Girondo, et al., 2018; Rozing et al., 2010) or siblings (Sebastiani et al., 2009) of cases.
As the definition of heritable longevity was not yet established, the construction and application of the family selection scores have not yet been adopted as state of the art in longevity research. As such, the majority of genealogical (Deluty, Atzmon, Crandall, Barzilai, & Milman, 2015; Dutta et al., 2013; Gudmundsson et al., 2000; Houde et al., 2008; Jarry et al., 2013; Perls et al., 2002; Sebastiani et al., 2016; Willcox, Willcox, He, Curb, & Suzuki, 2006) and genetic studies (Broer et al., 2015; Deelen et al., 2014, 2019; Flachsbart et al., 2016; Sebastiani et al., 2017; Willcox et al., 2008; Zeng et al., 2016) focus only on single, and thus including sporadic, long‐lived individuals (singletons), with some exceptions focusing, for example, on parental age (Joshi et al., 2017; Pilling et al., 2017) or multiple siblings (Broer et al., 2015; Schoenmaker et al., 2006). In previous work, we showed that longevity defined as top 10% survivors or more extreme is transmitted to subsequent generations (van den Berg et al., 2019). With this, a consistent definition of longevity was provided that is also adopted in the largest longevity GWAS up to now (Deelen et al., 2019). In addition, we showed that every additional long‐lived relative independently contributes to the survival advantage of study participants, according to their genetic distance (van den Berg et al., 2019). As such, there is room to incorporate these novel insights into family selection scores to gain knowledge about the extent that longevity needs to cluster in families in order to include individuals with the heritable longevity trait and increase the power of genetic studies.
Here, we aim to establish the proportion of ancestral blood relatives that should be long‐lived (sex‐specific top 10% survivors of their birth cohort or more extreme) in order to observe a survival advantage in their descendants and incorporate these insights into a novel family score to define cases with the heritable longevity trait for inclusion in genetic studies. For our analyses, we use the data available in the Historical Sample of the Netherlands (HSN) for the period between 1860 and 1875 which is based on Dutch citizens (Berg, Dijk, et al., 2018; Mandemakers, 2000, 2010). We primarily identify cases who died beyond 80 years (N = 884, on average top 10% survivors of their birth cohort), allowing us to select on more extreme ages at death, and controls who died between 40 and 59 years (N = 442, 40th‐60th survival percentile) and who represent the range of average mortality from birth at that time. We extend this filial (F) 1 generation data with a parental and three descendant generations of individuallife course and mortality data and refer to the data as the HSN case/control dataset. We subsequently exclude groups with high rates of missing mortality information and where the majority was still alive (Figure S4). This study covers 37,825 persons from 1,326 three‐generational families (F1–F3) and contains F1 index persons (IPs), two consecutive generations of descendants (F2–F3), and two generations of spouses (F2–F3; Table 1). The dataset is unique in that it covers multiple generations and connects alive persons to at least two generations of deceased ancestors.
Table 1.
Overview study sample for groups in all generations based on the proband and F3 perspective
| Role | Number | Deceased (%) | Alive (%) | Female (%) | Range birth cohort | Mean age (sd) | Median age (sd) | missing_age (%) |
|---|---|---|---|---|---|---|---|---|
| Cases (original approach) | ||||||||
| F1 IPs | 884 | 884 (100) | 0 (0) | 422 (50) | 1860–1875 | 85.79 (4.59) | 84.99 (4.95) | 0 (0) |
| F2 descendants | 4,916 | 4,405 (90) | 11 (1) | 2,435 (50) | 1879–1941 | 63.04 (31.11) | 75.51 (17.72) | 500 (9) |
| F2 spouses | 3,899 | 1,500 (38) | 16 (1) | 1504 (38) | 1873–1934 | 76.2 (15.09) | 78.78 (12.83) | 2,383 (61) |
| F3 descendants | 9,910 | 4,869 (49) | 4,146 (42) | 4,733 (48) | 1901–1973 | 70.35 (19.54) | 74.77 (11.38) | 895 (9) |
| F3 spouses | 3,431 | 1,289 (38) | 792 (23) | 1963 (57) | 1900–1959 | 77.14 (11.31) | 79.25 (10.1) | 1,350 (39) |
| F4 descendants a | 9,001 | 746 (8) | 7,172 (80) | 3,937 (44) | 1922–1995 | 57.7 (10.68) | 58.21 (9) | 1,083 (12) |
| Controls (original approach) | ||||||||
| F1 IPs | 442 | 442 (100) | 0 (0) | 214 (48) | 1860–1875 | 51.71 (5.71) | 52.88 (6.21) | 0 (0) |
| F2 descendants | 2,488 | 2,202 (89) | 1 (<1) | 1,217 (49) | 1881–1925 | 58.17 (32.49) | 71.72 (21.37) | 285 (11) |
| F2 spouses | 1,877 | 690 (37) | 7 (<1) | 734 (39) | 1875–1935 | 76.02 (14.77) | 78.34 (13.76) | 1,180 (63) |
| F3 descendants | 4,761 | 2,540 (53) | 1813 (38) | 2,265 (48) | 1904–1966 | 69.39 (20.38) | 74.49 (11.36) | 408 (9) |
| F3 spouses | 1,778 | 721 (41) | 376 (21) | 972 (55) | 1893–1965 | 76.54 (11.5) | 78.66 (10.47) | 681 (38) |
| F4 descendants a | 4,710 | 387 (8) | 3,744 (80) | 2099 (45) | 1871–1992 | 57.72 (11.17) | 58.37 (9.35) | 579 (12) |
| F3 perspective (combined approach) | ||||||||
| F3 descendants | 14,671 | 7,409 (51) | 5,959 (41) | 6,998 (48) | 1901–1973 | 70.03 (19.82) | 74.68 (11.38) | 1,303 (8) |
| F3 spouses | 5,209 | 2,010 (38) | 1,168 (22) | 2,935 (55) | 1893–1965 | 76.93 (11.38) | 79.07 (10.24) | 2,031 (40) |
| F2 parents | 9,728 | 6,139 (63) | 23 (1) | 4,137 (43) | 1873–1935 | 76.8 (13.4) | 78.9 (12.31) | 3,566 (36) |
| F2 aunts and uncles | 7,036 | 6,382 (91) | 10 (1) | 3,456 (49) | 1879–1941 | 61.81 (31.47) | 74.4 (18.67) | 644 (8) |
| F1 grandparents | 1,181 | 1,181 (100) | 0 (0) | 560 (47) | 1860–1875 | 74.88 (16.6) | 81.94 (9.72) | 0 (0) |
The Cases and Controls rows provide an overview of the groups of persons from the original case/control perspective of the data, described as the first study phase (original approach). The F3 perspective rows provide an overview of the groups of persons from the perspective of F3 descendants, described as the second study phase (combined approach). Mean and missing age refer to an unknown age at death or an unknown age at last observation. For the F0 and F1 groups, we assume everyone is dead because the birth cohorts date back further than 120 years. From the F2 generations, we requested Personal Records Data indicating if a person was still alive or not and if not, what the date of death was. The F1 IPs are the focal persons in the pedigrees as they are selected to be 80 years or older (cases) or to have died between 40 and 59 years (controls).
Indicates that the group is excluded for this study, and sd refers to standard deviation. Please note that the F3 perspective (second study phase, combined approach) is used to identify a new group of cases and controls based on the LRC score and are referred to as family cases and family controls.
2. RESULTS
2.1. Outline
We analyzed the data across multiple steps (Figure S5) in two phases. In the first phase, we used standardized mortality ratios (SMRs) to investigate the intergenerational transmission of longevity for cases (died beyond 80 years) and controls (died between 40 and 59 years) as defined in the original approach (Figure 1a), focusing on the F1 IPs and two generations of descendants.
Figure 1.
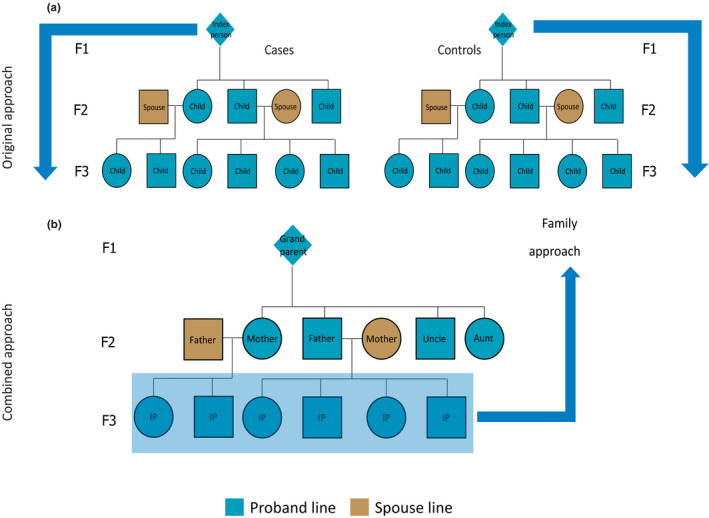
Pedigree overview of the data structure. This figure illustrates the two approaches: (1) the original approach and (2) the combined approach. The original approach refers to the case and control group based on the F1 IPs where cases died at 80 years or older and controls died between 40 and 59 years (panel a). Panel b shows a pedigree of the data from the perspective of F3 children (combined approach). The combined approach refers to the dataset where we combined the cases and controls from the original approach and constructed a new case and control group in the F3 descendants. To this end, F3 descendants with ≥30% long‐lived ancestors were labeled as family cases and those without long‐lived ancestors as family controls. F3 spouses were left out of this figure, but this group was used to confirm a genetic enrichment in the F3 descendants
In the second phase of our study (the combined approach), we combined original cases and controls and their descendants into one combined group and focused on the survival of the F3 descendants in relation to their F2 and F1 ancestral family members (Figure 1b). First, we constructed the Longevity Relatives Count (LRC) score. We used the LRC score to investigate the proportion of long‐lived (top 10% survivors of their birth cohort) F1 and F2 ancestors required for F3 descendants to express a survival advantage compared with members of the same birth cohort and sex (family method, Figure 1b). On the basis of these observations, we defined a new case and control group in F3, where we labeled F3 descendants with ≥30% long‐lived ancestors as family cases and those without long‐lived ancestors as family controls. Subsequently, these F3 family cases and controls were compared for their survival, that of their spouses (to investigate environmental influences), and for survival differences with the F3 descendants, selected to have at least one (singleton) long‐lived ancestor or at least one average‐lived ancestor. This means that they could have more than one long‐ or average‐lived ancestor but we actively selected for the presence of only one such ancestor. Figure S3a provides a conceptual overview of this selection. To this end, we selected either F3 descendants with at least one top 10% grandparent, at least one top 10% parent, or with grandparents who died between 40 and 59 years (their children (parents) resembled the general population). In a final step, we focused on the F3 descendants with at least one long‐lived parent and calculated LRC scores within this F3 group to determine whether parents transmitted their longevity more frequently if they were part of a long‐lived (LRC ≥ 0.30) family (Figure 1b). The analysis steps are summarized in Figure S5, and an overview of the available data per group and generation is shown in Table 1.
2.2. Longevity is transmitted in the case group and not in the control group
Focusing on the original approach (Figure 1a), we determined to what extent longevity is transmitted in the original case and the control group by estimating SMRs per generation for all cases and controls separately. Table 2 shows that F1 cases had a similar survival pattern to birth cohort members of the same sex, indicating that they resemble a representative group of random Dutch persons aged ≥80 years and born between 1860 and 1875. The SMR for the descendants of the cases (F2 case descendants) was 0.87 (95% CI = 0.84–0.89), indicating 13% less deaths than expected based on individuals from a similar birth cohort and sex. From here, we refer to this as 13% excess survival (or, if appropriate, excess mortality) compared with the general population. The descendants of controls (F2 control descendants) had a similar survival pattern to the general population (SMR = 1.01 [95% CI = 0.96–1.05]). The spouses of the F2 case and control descendants surprisingly also showed a pattern of excess survival (SMRcase_F2spouses = 0.89 [95% CI = 0.85–0.94] and SMRcontrol_F2spouses = 0.9 [95% CI = 0.83–0.97]). Next, we observed 14% (95% CI = 11%–16%) excess survival compared with the general population for F3 descendants of the F1 cases, whereas F3 control descendants resembled the general population (SMR = 0.96 [95% CI = 0.93–1.00]) just as observed in the F2 generation. The spouses of both F3 groups resembled the general population (SMRcase_F3spouses = 1.00 [95% CI = 0.95–1.05] and SMRcontrol_F3spouses = 1.07 [95% CI = 0.99–1.15]). We conclude that two descendant generations of cases, who belong on average to the top 10% survivors, have 13%–14% excess survival compared with the general populations and that the descendants of controls resemble the general population.
Table 2.
Standardized mortality ratios for original case and control group individuals
| Role | Case group SMRs | Number (N) | Control group SMRs | Number (N) | Adjustment for left truncation |
|---|---|---|---|---|---|
| F1 IPs | 1.06 (0.99–1.13) | 884 | NA | NA | 80 years |
| F2 descendants | 0.87 (0.84–0.89) | 4,416 | 1.01 (0.96–1.05) | 2,203 | No adjustment |
| F2 spouses | 0.89 (0.85–0.94) | 1,516 | 0.9 (0.83–0.97) | 697 | 20 years |
| F3 descendants | 0.86 (0.84–0.89) | 9,015 | 0.96 (0.93–1.00) | 4,353 | No adjustment |
| F3 spouses | 1.00 (0.95–1.05) | 2,081 | 1.07 (0.99–1.15) | 1,097 | 20 years |
Original cases (F1 IPs) died at 80 years or older, and original controls (F1 IPs) died between 50 and 69 years. If persons could not die before a specific age due to direct or indirect selection, due to, for example, that all persons in a group were selected to have a child, an adjustment for right truncation was applied so that a fair comparison could be made with their birth cohort members. An SMR for F1 control IPs could not be estimated due to a combination of left truncation and right truncation in the data. The life tables can only be adjusted for right or left truncation, but not a combination between the two.
To explore to what extent the survival of F2 and F3 descendants depends on the extremity of the longevity of their parents, we calculated SMRs for F2 and F3 case and control descendants with increasing parental longevity (e.g., a parent belonged to the top 10%, 5%, or 1% survivors). We observed that the SMR decreased in descendants when defining parental longevity in terms of more extreme survival percentiles. This was the case for descendants of both the IP cases and controls although the effects were stronger in the descendants of the cases, especially in F3, since this group is now selected to have long‐lived parents and grandparents (Table S1). This illustrates that selection on single long‐lived persons belonging on average to the top 10% survivors, as we did for the IP selection, leads only to a modest transmission of longevity in two generations (max 14%). Likely, the control group includes misclassified persons of which the descendants do live longer, whereas the case group includes long‐lived persons that do not transmit longevity to their descendants (potentially these are phenocopies). Such misclassification can jeopardize genetic studies immensely. To be able to evaluate living persons as potential carriers of the heritable longevity trait in genetic studies, we constructed and validated a familial longevity score.
2.3. Constructing the Longevity Relatives Count score
We now look at the HSN data from a different perspective, the combined approach (Figure 1b). In the combined approach, we consider the F3 generation as the focal point of the pedigree, instead of the F1 generation, as was the case in the original approach. To identify individuals with the heritable longevity trait, we constructed the LRC score.
where i refers to the F3 descendants for whom the score is built. k is an index referring to each ancestral blood relative (from here: ancestors) of person i who are used to construct the score. Ni refers to the total number of ancestors of person i (Figure 1b), Pk is the sex and birth year‐specific survival percentile, based on life tables, of ancestor k, and I(Pk ≥ 0.9) indicates whether ancestor k belongs to the top 10% survivors. is the weighted total number of ancestors of F3 descendant i. The relationship coefficients are used as weights wk . The LRC score indicates the proportion of ancestors that has become long‐lived. For example, an LRC of 0.5 indicates 50% long‐lived ancestors (see methods for a more detailed and general description of the LRC score).
2.4. Longevity is transmitted when at least 20% of all ancestors are long‐lived
To determine what proportion of long‐lived ancestors could be associated with the survival of F3 descendants, we calculated LRC scores for all F3 descendants and subsequently defined nine mutually exclusive LRC groups (g) of F3 descendants: LRC_g1 = 0, LRC_g2 = [>0 & <0.1], LRC_g3 = [≥0.1 & <0.2], LRC_g4 = [≥0.2 & <0.3], LRC_g5 = [≥0.3 & <0.4], LRC_g6 = [≥0.4 & <0.5], LRC_g7 = [≥0.5 & <0.6], LRC_g8 = [≥0.6 & <0.7], and LRC_g9 = [≥0.7 & ≥1.0]. For each group of F3 descendants, we explored whether they have a survival benefit compared with the general population by estimating SMRs (Figure 2). F3 descendants without any long‐lived ancestors (LRC score of 0) had a survival pattern that resembled the general population (SMR = 0.97 [95% CI = 0.93–1.01]). Similarly, we observed a survival pattern that resembled the general population for F3 descendants with up to 20% long‐lived ancestors (groups 2 and 3, SMR = 0.97 [95% CI = 0.91–1.04] and SMR = 0.95 [95% CI = 0.91–1.00], respectively). This shows that the long‐lived ancestors of group 2 and group 3 F3 descendants were likely phenocopies instead of genetically enriched long‐lived persons. We observed a pattern of excess survival for F3 descendants with more than 20% long‐lived ancestors. The weakest significant effect was observed for group 3, with an SMR of 0.84 (95% CI = 0.80–0.89) which is comparable to the excess survival of the F3 descendants of the singleton F1 cases in the original approach (first part of the results). The strongest significant effect was observed for group 8, with an SMR of 0.56 (95% CI = 0.45–0.69). Hence, the higher the degree of long‐lived ancestors, the lower the SMR. This indicates that the more long‐lived ancestors an F3 descendant has, the higher the level of excess survival of these F3 descendants is compared to the general population, and the more likely that genetic effects drive the transmission of longevity.
Figure 2.
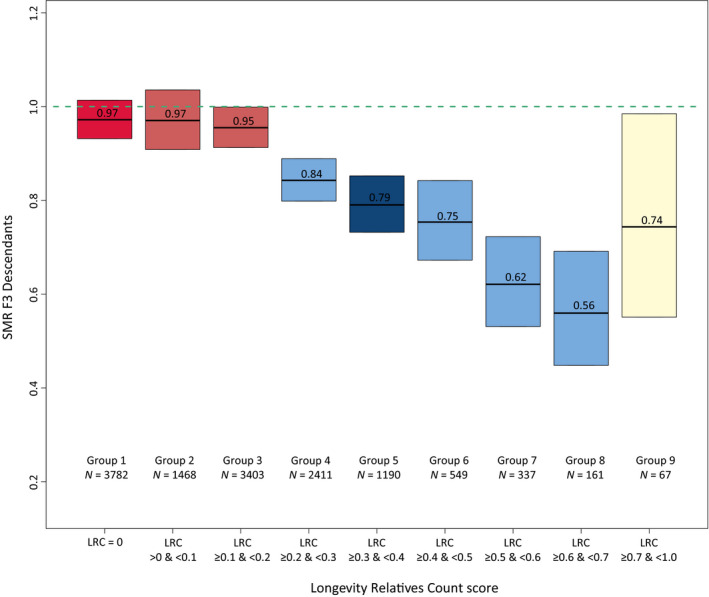
LRC score in mutually exclusive F3 descendant groups. The figure shows standardized mortality ratios for all F3 descendants without missing age (at death or last observation) information. Please note that the group sizes are smaller than mentioned in the text as those in the text represent the full group sizes, including the persons with missing age information. The F3 descendants are grouped into mutually exclusive groups based on the LRC score. The LRC score represents the family approach as illustrated in Figure 1b. The dark red color of group 1 represents F3 descendants without any long‐lived (top 10%) ancestors and is denoted as family controls. The light red represents F3 descendants who had more than 0 and less than 20% long‐lived ancestors. The light blue colors represent the F3 descendants with 20% or more long‐lived ancestors. The dark blue color represents our cutoff point for the family case definition. Hence, all F3 descendants with 30% or more long‐lived ancestors were considered family cases. The beige color of group 9 shows that this bar represents all F3 ancestors with more than 70% long‐lived ancestors as their sample size was very low, we grouped them into one group
Using the LRC score family method, we defined a new case and control group in the F3 generation, which is based on the presence or absence of longevity among the ancestors of the F3 generation and potential excess survival or mortality in the F3 generation itself (Figure 1b). The F3 family controls include all F3 descendants without any long‐lived ancestors (LRC score of 0, N = 4,166). To define the F3 family cases, we chose an LRC cutoff based on a tradeoff between the size and the uncertainty, given by the sample size, of the SMR. The F3 family cases include all F3 descendants with at least 30% long‐lived ancestors (LRC score ≥0.30 [N = 2,526]). Even if F3 family cases are not long‐lived themselves, their survival reflects the presence of longevity of their ancestors, which is transmitted by their parents. Similarly, F3 controls reflect the absence of longevity of their ancestors. Figure S1 shows the variation in lifespan of the F3 family case and control descendants. F3 descendants with more than 0% and up to 20% long‐lived ancestors (LRC score >0 and <0.2) did not express excess survival (N = 5,340). The F3 descendants with an LRC score ≥0.2 and <0.30 showed some excess survival compared with the general population, but the size of the SMR was considered too low to enter our family case definition. Hence, we denoted them as nonclassified (N = 2,639).
2.5. Strong survival advantage and genetic enrichment for F3 family cases
To validate the LRC score, we investigate survival differences, measured as age at death or last observation, between the F3 family cases and controls and used a Cox‐type random effects (frailty) regression model to adjust for within‐family relations of the F3 descendants. Figure 3 and Table 3a show that F3 cases have a 25% (95% CI = 18%–31%) lower hazard of dying than F3 controls, even after adjustment for sibship size, birth year, and sex. The difference between the cases and controls became increasingly more pronounced when confining the cases to a higher proportion of long‐lived ancestors, for example, an LRC score of 0.40, 0.50, or 0.60, reflecting 40%, 50%, or 60% long‐lived ancestors (Figure S2). The strongest effect was observed for those with an LRC score ≥0.60 (hazard ratio [HR] of 0.62 [95% CI = 0.50–0.77]). The mortality pattern for the spouses of these F3 cases resembled that of the F3 controls (HR = 0.94 [95% CI = 0.82–1.07]; Table 3b) and the general population (SMR = 0.92 [95% CI = 0.83–1.02]). The survival of the spouses, equal to the F3 controls and the general population, in addition to the absence of effects of environmental covariate adjustment, indicates that environmental factors were likely of limited influence to the observed survival benefit of the F3 cases as defined by our novel family‐based definition. Hence, the observed survival benefit of F3 cases likely represents a genetic longevity component.
Figure 3.
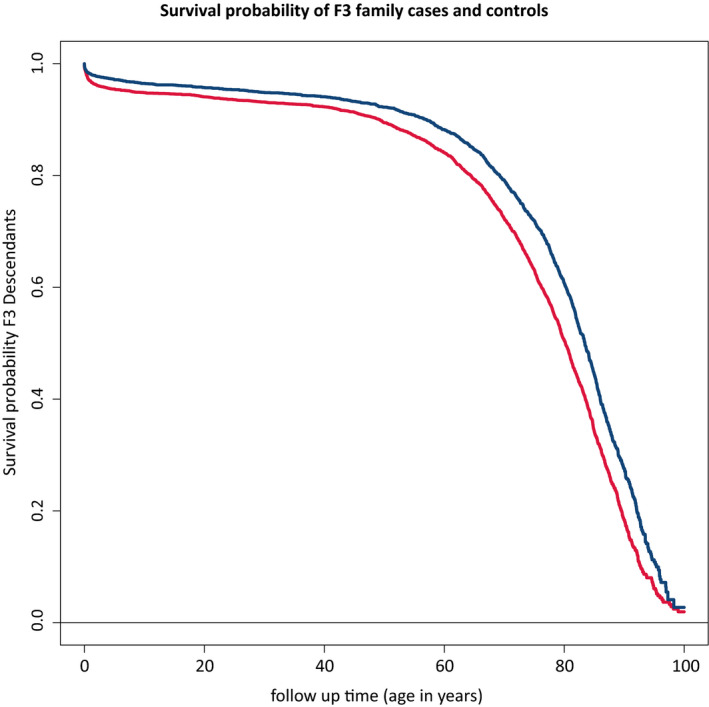
Survival differences between family‐based cases and their spouses. This figure shows the survival curve for the difference in survival between the F3 family cases and controls. The figure is connected to Table 3a which shows the hazard ratios corresponding to the difference between the two curves. Blue color represents the cases, and red color represents the controls
Table 3.
Mortality difference between family cases and controls and their spouses
| A | B | |||||
|---|---|---|---|---|---|---|
| N (mean) | HR (95% CI) | p‐value | N (mean) | HR (95% CI) | p‐value | |
| Family‐based case/control group | ||||||
| Control group (ref) | 3,714 (0.62) | 3,714 (0.50) | ||||
| Case group | 2,282 (0.38) | 0.75 (0.69–0.82) | 1.75e−10 | 2,282 (0.30) | 0.74 (0.68–0.80) | 4.08e−12 |
| Spouses of cases | 541 (0.07) | 0.94 (0.82–1.07) | 3.44e−01 | |||
| Spouses of controls | 937 (0.13) | 1.12 (1.00–1.25) | 4.07e−02 | |||
| Birth year | 5,996 (1933) | 0.99 (0.98–0.99) | 1.99e−05 | 7,474 (1932) | 0.98 (0.98–0.99) | 1.39e−12 |
| Sex | ||||||
| Males (ref) | 3,133 (0.52) | 3,364 (0.45) | ||||
| Females | 2,863 (0.48) | 0.56 (0.52–0.61) | <1.00e−15 | 4,110 (0.55) | 0.49 (0.46–0.53) | <1.00e−15 |
| Sibship size | ||||||
| Small—1–2 sibs (ref) | 1,531 (0.26) | |||||
| Medium—3–5 sibs | 1,770 (0.30) | 1.17 (1.04–1.32) | 8.51e−03 | |||
| Large—6–8 sibs | 927 (0.15) | 1.22 (1.04–1.43) | 1.21e−02 | |||
| Exceptional—9–15 sibs | 441 (0.07) | 1.36 (1.09–1.68) | 5.84e−03 | |||
| Single child—0 sibs | 1,327 (0.22) | 1.81 (1.62–2.02) | <1.00e−15 | |||
“A” corresponds to the CH curves of panel A of Figure 3. Means represent a mean for a continuous variable and a proportion for a categorical variable. When the p‐value was lower than 1.00e−15, we indicated the p‐value as <1.00*10–15. CI = confidence interval. F3 descendants with relatives who were still alive and had no last moment of observation ≥100 years were removed to assure an equal comparison between cases and controls. In “B”, the spouses of cases and controls are adjusted for the fact that they could not die before the birth of at least their first child (left truncation). We adjusted for this left truncation by entering the spouses of cases and controls in the model based on the first observed death in the groups (cases: 30 years and controls: 25 years). In model A, no adjustment for left truncation was necessary. In both models, we adjusted for right censoring by including a censoring indicator in the Cox model.
2.6. Family cases live longer than those with one long‐lived parent or grandparent
Next, we test whether the F3 descendants with 30% long‐lived ancestors (the family cases) have a stronger survival advantage than F3 descendants with at least one long‐lived (top 10%) parent or grandparent. We actively selected this group of F3 descendants to have one long‐lived parent or grandparent, meaning that other ancestors could also be long‐lived but there was no active selection on the presence of their longevity (Figure S3a,b), hence the designation “at least” for this group. Subsequently, we test whether F3 descendants without long‐lived ancestors (the family controls) had a similar survival pattern to the F3 descendants with parents resembling the general population (those with a grandparent who died between 40 and 59 years). Table 4 shows that we observed 14% (95% CI = 11%–17%) excess survival compared with the general population for F3 descendants with at least one long‐lived grandparent (F1). When identifying F3 descendants with at least one long‐lived parent (F2), we observed 16% (95% CI = 8%–24%) excess survival compared with the general population. Using the family method at 30% long‐lived family members to identify F3 family cases, we observed 26% (95% CI = 22%–30%) excess survival compared with the general population and this increased to 38% (95% CI = 31%–45%) when applying a 50% threshold to the family method. For the identification of controls, both methods seem to perform equally well, with almost identical SMRs of around one. This indicates that the F3 controls, whether defined by having no long‐lived ancestors or by grandparents dying between 40 and 50 years, have a similar survival pattern to the general population. We conclude that, at least for cases, the family method provides a better contrast in excess survival compared with the general population and seems to better represent the heritable longevity trait.
Table 4.
Standardized mortality ratio for different F3 descendant groups
| Group | SMR | N |
|---|---|---|
| Cases | ||
| F3 descendant with at least one long‐lived grandparent | 0.86 (95% CI = 0.83–0.89) | 4,986 |
| F3 descendant with at least one long‐lived parent | 0.84 (95% CI = 0.76–0.92) | 852 |
| F3 descendant with ≥30% long‐lived ancestors (LRC ≥ 30%) | 0.74 (95% CI = 0.70–0.78) | 2,304 |
| F3 descendant with ≥50% long‐lived ancestors (LRC ≥ 50%) | 0.62 (95% CI = 0.55–0.96) | 565 |
| Controls | ||
| F3 descendant with grandparent who died between 40 and 59 years | 0.96 (95% CI = 0.93–1.00) | 4,353 |
| F3 descendant with no long‐lived ancestors (LRC = 0) | 0.97 (95% CI = 0.93–1.01) | 3,782 |
Long‐lived is defined as belonging to the top 10% survivors of their birth cohort. Note that the group size (N) reflects only those with a known age at death as this was necessary to estimate a standardized mortality ratio.
Since the F3 descendants with ≥30% long‐lived ancestors have a stronger survival advantage than those with at least one long‐lived parent, it is possible to get an indication of how many F3 descendants did not appear to have a survival advantage compared with the general population, even though at least one parent was long‐lived. This is relevant in view of case definitions used in large genetic studies into longevity. Figures 4 and S3 show that 919 F3 descendants had a long‐lived parent. Out of those 919 F3 descendants, 247 (27%) had more than 0% but less than 20% long‐lived ancestors (LRC > 0 and <0.20) and thus as a group had an SMR that resembled the general population (Figure S3d). The other 672 (73%) had exactly or more than 20% long‐lived ancestors (LRC ≥ 0.20) and thus, as a group, showed excess survival compared with the general population (Figure S3b,c). These results suggest that if living persons are selected as case in genetic studies on the basis of one long‐lived parent, 27% of these persons is unlikely to be a carrier of the longevity trait. Persons defined as 30% long‐lived ancestors, on the other hand would be potential carriers.
Figure 4.
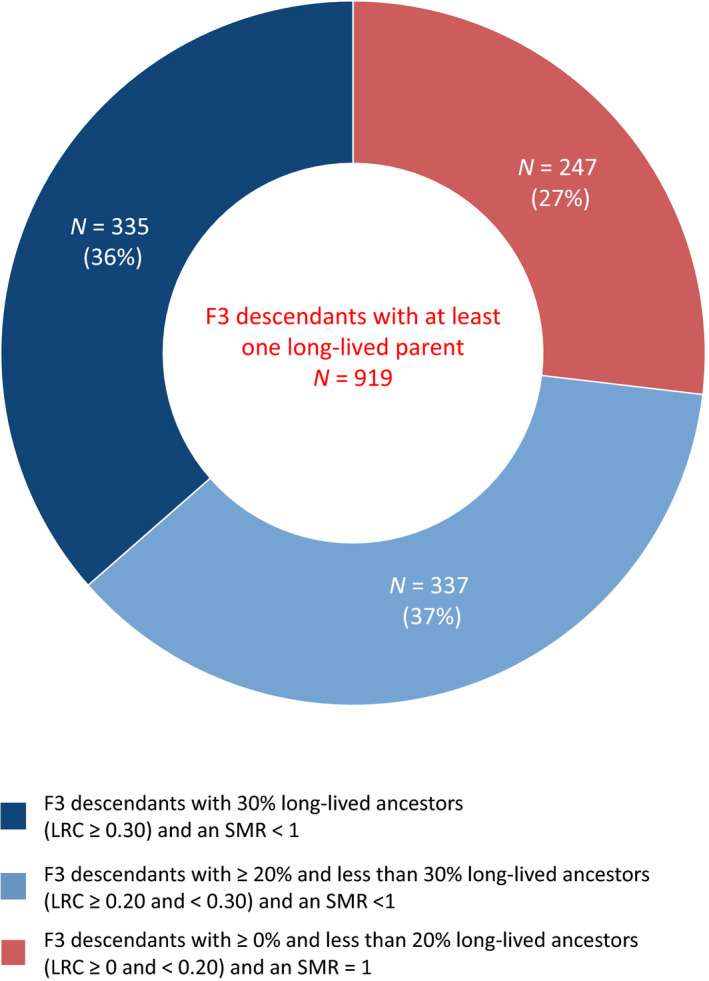
LRC score for F3 descendants with at least one long‐lived parent. This center of this doughnut figure shows all F3 descendants (N = 919) with at least one long‐lived (top 10%) parent, ignoring the rest of the ancestors. Thus, at least means that they could have more than 1 long‐lived ancestor but we actively selected for the presence of only 1 such ancestor. The edges of the doughnut illustrate the number and proportion of these 919 F3 descendants with at least one long‐lived parent who had (1) 30% or more long‐lived ancestors (LRC ≥ 0.30) and excess survival compared with the general population (SMR < 1), N = 335 (36%), (2) between 20% and 30% long‐lived ancestors (LRC ≥ 0.20 and <0.30) and excess survival compared with the general population (SMR < 1), N = 337 (37%), and (3) between 0% and 20% long‐lived ancestors (LRC > 0.20 and <0.20) and a similar survival pattern to the general population (SMR ~ 1), N = 247(27%)
3. DISCUSSION
Human longevity is heritable and clusters in specific families. Studying the familial clustering of longevity in these families is important to improve our understanding of genetic factors promoting longevity and healthy aging. The main observations supporting this are as follows: (a) In the original approach, we observed 14% excess survival of the cases compared with their birth cohort for two subsequent generations (F2–F3), while in the controls no such benefit was observed; (b) in the combined approach, the excess survival of the F3 cases compared with the general population was 26%–38% depending on the proportion of long‐lived family members being 30%–50%, and these estimates strongly overlap to the survival difference between the F3 family cases and controls based on the Cox models; (c) no excess survival as compared to the birth cohort and general population was observed for F3 controls, spouses of cases or controls, and neither for F3 cases with up to 20% long‐lived ancestors. The analyses in the HSN case/control dataset provide strong evidence that longevity is transmitted for at least two subsequent generations and only when at least 20% of all ancestors are long‐lived. Moreover, the family cases seem to be genetically enriched for longevity while the controls resemble the general population. Finally, 27% of the F3 descendants showed a survival pattern similar to the general population even though they had at least one long‐lived parent.
Previous family studies, usually focusing on two generations and single individuals, showed that siblings and children of long‐lived persons lived longer than first‐degree ancestors of non‐long‐lived persons or population controls (Berg, Rodríguez‐Girondo, et al., 2018; Deluty et al., 2015; Dutta et al., 2013; Gudmundsson et al., 2000; Houde et al., 2008; Jarry et al., 2013; Kemkes‐Grottenthaler, 2004; Perls et al., 2002; Schoenmaker et al., 2006; Sebastiani et al., 2016; Terry et al., 2004; Willcox et al., 2006). This knowledge about the familial clustering of longevity was utilized to construct longevity ranking scores such as the Family Mortality History Score (FMHS; Rozing et al., 2010), the est(SE) which subsequently was developed into the FLOSS (Arbeeva et al., 2018; Sebastiani et al., 2009), the Longevity Family Score (LFS) which is an adaptation to the est(SE) and the FMHS (Berg, Rodríguez‐Girondo, et al., 2018), and finally a method was developed to rank individuals by the survival of their ancestors, the FEL score (Kerber et al., 2001). The FMHS, FLOSS, and LFS all resemble excess survival of a family (FMHS focus on parents and FLOSS and LFS focus on siblings) compared with the general population. The FEL score focuses on excess survival, defined as the difference between a person's attained and expected age, derived from an accelerated failure time model. This excess survival was estimated for ancestors, and from this, a score was created for individuals.
We developed a novel tool based on mapping the longevity of a person's ancestors, the LRC score. Benefits of the LRC score compared with the other scores are that: (a) it is based on scientific evidence for the cutoff (top 10% survivors of their birth cohort) of when longevity becomes heritable. Thus, the score has as a benefit that it captures the heritable longevity component, and (b) it can be constructed using reference life tables of any study participants’ country. Hence, it deals with unobserved measurements that associate with the increase in life expectancy of the past 200 years. It also ensures that study populations with relatively young persons cannot be denoted as long‐lived; (c) it is always on a 0–1 scale, resembling a proportion, regardless of whether different reference population life tables or study cohorts are used. This ensures possibilities for international comparisons; (d) it is based on a mean (resulting in the proportion property) which ensures that the score does not have a preference toward large families and that the score can easily be based on a single generation of relatives or multiple generations, providing good flexibility. (e) It incorporates the latest insights of longevity research as it was shown that every additional long‐lived family member independently contributes to the longevity of study participants according to their genetic distance (van den Berg et al., 2019). The LRC takes this into account by applying weights that correspond to these genetic distances.
The LRC score can be used to select carriers of the heritable longevity trait (cases) and controls who resemble the general population. Another interesting group, which we did not address in this article, is composed of persons without any long‐lived ancestors who themselves are long‐lived. It may be interesting to study environmental factors contributing to a long and healthy life in this group. Here, we used the LRC score to construct a novel family case and control group and observed a survival advantage for F3 case descendants, even when their parents were not necessarily long‐lived, supporting the idea that a beneficial genetic component was transmitted. Likewise, the increase in the LRC score ≥20% associated with an increase in survival advantage for F3 descendants. This indicates that every additional ancestor contributes to the survival advantage of F3 descendants and confirms our previous findings in the LINKing System for historical demography (LINKS) data and the Utah Population Database (van den Berg et al., 2019). This additive pattern is not readily expected if the observations are due to nongenetic factors, such as wealth, that cluster in families. The fact that none of the environmental confounders (sex, birth year, and sibship size) affected the survival differences between the family cases and controls provided additional evidence for the transmission of a genetic component. A final indication for the genetic enrichment of the family cases is based on the observed mortality pattern for the spouses of the family cases and controls which resembled the family controls themselves and the general population.
We observed that F3 descendants with at least one long‐lived parent had less excess survival than a subset of these F3 descendants who had at least 30% long‐lived ancestors and this difference increased when at least 50% of their ancestors were long‐lived. These results indicate that some parents were long‐lived but might not have transmitted their longevity to the subsequent F3 generation. In fact, 27% of the F3 descendants with at least one long‐lived parent did not have an LRC ≥ 0.20 and, as a group, did not express excess survival. Hence, the parents of these 27% F3 descendants were sporadically long‐lived as they did not transmit their longevity. Thus, genetic studies may benefit from a case definition, where cases are long‐lived and have at least 30% long‐lived ancestors, as current genetic studies, based on long‐lived cases, often not include ancestral longevity in their case selection. Even though our data did not allow for an exact misclassification analysis, studies showed that the level of phenotypic misclassification in case and control annotation has a strong inhibiting effect on the power to identify variants in genetic association studies, including GWAS (Barral, Haynes, Stone, & Gordon, 2006; Bross, 1954; Buyske, Yang, Matise, & Gordon, 2009; Edwards, Haynes, Levenstien, Finch, & Gordon, 2005; Gordon, 2005; Gordon, Haynes, Yang, Kramer, & Finch, 2007; Ji, Yang, Haynes, Finch, & Gordon, 2006; Platz, De Marzo, & Giovannucci, 2004; Rekaya, Smith, Hay, Farhat, & Aggrey, 2016; Royall, Chiodo, & Polk, 2004). Moreover, it was shown that the power to identify genetic variants decreases at an equal rate to the level of misclassification (Gordon, 2005). For example, a study with 95% power to detect an association based on a sample of 100 cases and controls when there are no phenotypic errors may actually have only 75% power when 20% of the cases are misclassified as controls and vice versa (Gordon, 2005). Interestingly, when known, methods exist to adjust for the level of phenotypic misclassification (Barral et al., 2006; Edwards et al., 2005; Gordon et al., 2007; Rekaya et al., 2016; Smith, Hay, Farhat, & Rekaya, 2013), providing opportunities for specific application in genetic longevity research.
Due to the nature of the HSN data, we could not use the mortality data for the parents (F0), siblings (F1), and spouses (F1) of the F1 IPs. Mortality data were less incomplete for the F2 and F3 spouses (Table 1a), but there were still a relatively large number of missing mortality data. Thus, for future studies with this dataset it might be interesting to extend the mortality information for these groups. The missing mortality data for the F2 spouses may also have led to the pattern of excess survival for the spouses of both F2 cases and controls, as the final data for these groups covered an overrepresentation of relatively high ages at death for those spouses. Furthermore, life course data were only present for persons with an identified personal card (PC) or personal list (PL; details in the methods section). Consequently, socioeconomic status and religion was only available for a small part (around 15%) of the F3 descendants with an unequal share of availability between men and women. This led to the exclusion of these environmental factors from our analyses. Even though we could not adjust our models for socioeconomic status and religion, it is known from other studies that those factors are not influencing the association between parental longevity and offspring survival (Mourits et al., 2019; van den Berg et al., 2019). Similarly, previous studies showed only a minor (You, Danan, & Yi, 2010) or no (Gavrilov & Gavrilova, 2015; van den Berg et al., 2019) influence of early and mid‐life environmental covariates, such as farm ownership, parental literacy, parental and own occupation, and birth intervals, on the association between parental longevity and offspring survival. We, however, cannot completely rule out that other, unobserved nongenetic familial effects may affect our results. The observed excess survival of F2 case and control group spouses in the original approach seems to be an exception, as we observed a survival advantage for both groups. This is likely a form of ascertainment bias because mortality data for this group were difficult to obtain in the Dutch Personal Records Database (PRD), leading to an overrepresentation of high ages at death. These observations add to the mixed results about whether spouses married to a long‐lived person have a survival advantage themselves (Berg, Rodríguez‐Girondo, et al., 2018; Jarry et al., 2013; Montesanto et al., 2011; Pedersen et al., 2017; Schoenmaker et al., 2006; van den Berg et al., 2019).
Our results have two important implications. First, existing studies based on living study participants who have not yet reached the ages to express longevity, but have ancestral survival data, such as UK Biobank, can now better distinguish cases by incorporating a liability based on the LRC score. Second, new studies would obtain a maximum power to identify loci that promote survival to the highest ages in the population when cases are included with at least 30% (LRC ≥ 0.30) ancestors who belong at least to the top 10% survivors of their birth cohort and are themselves among the 10% longest lived. As such, this strategy invites existing study cohorts of long‐lived study participants to extend their data with ancestral mortality information. In the past, it was difficult to obtain such ancestral information, but currently it is much more feasible to do so, as population‐scale family tree data are becoming increasingly available (Erlich, Shor, Pe’er, & Carmi, 2018; Kaplanis et al., 2018; van den Berg et al., 2019). More extreme selections can be made on the survival percentile by, for example, focusing on the top 5% or 1% survivors, and/or on the proportion of long‐lived family members, for example, 50%. However, this is not strictly necessary and might unnecessarily lead to limited sample sizes (van den Berg et al., 2019). In addition, controls without any ancestors living to the top 10% survivors of their birth cohort should be included, as their mortality pattern resembles that of the general population. Finally, for future research it may be interesting to study the environmental factors causing the longevity in those individuals who were long‐lived but had no long‐lived ancestors. If our proposed method is consistently applied across studies, the comparative nature of longevity studies may improve and facilitate the discovery of novel genetic variants.
4. METHODS
4.1. Historical sample of the Netherlands
The HSN Dataset Life Courses, Release 2010.01, is based on a sample of birth certificates and contains complete life course information for 37,137 Dutch individuals (IPs) born in and between 1850 and 1922 (Berg, Dijk, et al., 2018; Mandemakers, 2000, 2010). These 37,137 persons were subsequently identified in the Dutch population registers and followed in the registers throughout their entire life course (Berg, Dijk, et al., 2018; Mandemakers, 2002, 2010). The database includes information about the IPs’ household, including their siblings, parents, and children, occupation at several points in time and religion. Households were only followed as long as the IP was present in that household meaning that information on kin was only partly covered (Berg, Dijk, et al., 2018; Mandemakers, 2002). For this study, we selected 884 IPs who died at 80 years or beyond (case group) and 442 IPs who died between 40 and 59 years (control group), representing 1,326 disjoint families. IPs from both groups were born between 1860 and 1875. The case group was defined so that we would obtain a sample with overrepresentation of long‐lived individuals. This was interesting since it would potentially allow to select on more extreme ages at death and still guarantee numbers reasonably large. The control group was selected to represent the mortality pattern of the general population of that time as best as possible. We based this selection on the full HSN where we calculated the range of average age at death from birth for the birth cohorts 1860–1875. We used age at death from birth as it is known that the survival advantage for relatives of long‐lived individuals starts already from birth (Berg, Rodríguez‐Girondo, et al., 2018; Perls et al., 2002). With this, we ensured a fair comparison between the case and control groups. Individuals from both groups were selected to have an available date of birth, date of death, and at least one child should be identified. In conclusion, we identified 1,326 IPs (cases and controls), their F0 parents (N = 2,652), F1 siblings (N = 5,179), F2 descendants (N = 7,404), and F1 spouses (N = 1,409), covering three filial generations (F0–F2) spanning from 1788 to 1941 (Table 1; Figure 1a). The underlying data for this specific study were released as Kees Mandemakers and Cor Munnik, HSN. Project Genes, Germs and Resources. Dataset LongLives. Release 2016.01.
4.2. Extending the HSN study
For this study, we extended the pedigrees until we identified the living descendants for all 1,326 families. From the population registers, we know the names of all F2 descendants and we subsequently identified the F2 descendants on PCs and PLs which were obtained from the Dutch central bureau of genealogy. These PLs and PCs were, respectively, introduced in 1939 and 1994 as the individualized and subsequently digitized form of the population register (Berg, Dijk, et al., 2018). The cards contain similar information to the population registers and because of privacy legislation could only be obtained for deceased persons, 1 year after they passed away (https://cbg.nl/bronnen/cbg‐verzamelingen/persoons kaarten‐en‐lijsten). Hence, from these cards we obtained similar life course and mortality information for the F2 descendants as for the F1 IPs and we obtained the names of their descendants (F3). We repeated this procedure until no cards could be obtained anymore, which was at the F3 generation. Thus, the F4 generation was not identified on the PCs of PLs anymore. In conclusion, we identified and obtained information for the F2 descendants, F2 spouses, F3 descendants, F3 spouses, and F4 descendants (Table 1; Figure 1a). We will refer to this database as the HSN case/control database.
4.3. Obtaining information for the living descendants
In a final step, we obtained as much mortality information as possible for the relatives of the identified persons and we obtained addresses, as contact information for the living descendants. This information was obtained through the PRD which is managed by Dutch governmental service for identity information. https://www.government.nl/topics/personal‐data/personal‐records‐database‐brp. The PRD contains PL information on all Dutch citizens (alive and death), and PC information is continuously added. We were granted permission (permission number: 2016‐0000364875) to obtain the date of death, date of last observation, current living address, and identifying information such as names of a person's father and mother to double check whether the person identified in the PRD was identical to the person in our HSN case/control database. Using the PRD, we were able to obtain addresses for F3 and F4 descendants and additional mortality information for F2 descendants, F2 spouses, F3 descendants, F3 spouses, and F4 descendants (Table 1; Figure 1a). The final database covers 57,337 persons from 1,326 five‐generational families (F0–F4) and contains F1 IPs, their parents (F0), siblings (F1), spouses (F1), and three consecutive generations of descendants (F2–F4) and spouses (F2–F4), connecting deceased persons to their living descendants.
4.4. Exclusion criteria and study population
Due to the nature of the source data, there is a high rate of missing mortality information for F0 parents, F1 spouses, and F1 siblings, which we therefore excluded from analyses. We further excluded F4 descendants because 92% is still alive (Table 1; Figure 1b). The final study population covers 37,825 persons from 1,326 three‐generational families (F1–F3) and contains F1 IPs, 2 consecutive generations of descendants (F2–F3), and two generations of spouses (F2–F3).
4.5. Statistical analyses
Statistical analyses were conducted using R version 3.4.1 (R Core Team, 2016). We reported 95% confidence intervals (CIs) and considered p‐values statistically significant at the 5% level (α = 0.05).
4.6. Life tables
In the Netherlands, population‐based cohort life tables are available from 1850 until 2019 (Carolina, Uijvenhoven, & van der Laan, 2009; Van Der Meulen, 2012). These life tables contain, for each birth year and sex, an estimate of the hazard of dying between ages x and x + n (hx) based on yearly intervals (n = 1) up to 99 years of age. Conditional cumulative hazards (Hx) and survival probabilities (Sx) can be derived using these hazards. In turn, we can determine to which sex and birth year‐based survival percentile each person of our study belonged to. For example, a person was born in 1876, was a female, and died at age 92. According to the life table information, this person belonged to the top three percent survivors of her birth cohort, meaning that only three percent of the women born in 1876 reached a higher age. We used the life tables to calculate the birth cohort and sex‐specific survival percentiles for all persons in the HSN case/control study. This approach prevents against the effects of secular mortality trends over the last centuries and enables comparisons across study populations (Sebastiani et al., 2016; van den Berg et al., 2017). Figure S6 shows the ages at death corresponding to the top 10, 5, and 1 percent survivors of their birth cohorts for the period 1850–1935.
4.7. Standardized mortality ratios
To indicate excess mortality or excess survival of groups, such as F2 case or control group descendants in the HSN case/control study compared with Dutch birth cohort members of the same sex, we used SMRs. An SMR is estimated by dividing the observed number of deaths by the expected number of deaths. The expected number of deaths is given by the sum of all individual cumulative hazards based on the birth cohort and sex‐specific life tables of the Dutch population. An SMR between 1 and 0 indicates excess survival, an SMR of 1 indicates that the study population shows a similar survival to the reference population, and an SMR above 1 indicates excess mortality. The SMR can be estimated conditional on the specific age at which an individual starts to be observed in the study (correction for left truncation). This was necessary to avoid selection bias if individuals in a study population were not at risk of dying before a specific age of entry.
where dt = dead status (1 = dead, 0 = alive), Ht 0 i = sex and birth year‐specific cumulative hazard based on life table, ti = timing, referring to age at death or last observation, t 0 i = liftable age conditioning, for example, from birth (t 0 i = 0), and N = group sample size. Exact CIs were derived (Ulm, 1990) and compared to bootstrap CIs for family data (Berg, Rodríguez‐Girondo, et al., 2018). Both methods provided identical CIs, and thus, to reduce the amount of computational time necessary to estimate bootstrap CIs, we estimated exact CIs.
where LL is the lower CI limit and UL is the upper CI limit. is the (100*αth) chi‐square centile with v = 2d degrees of freedom, d is the number of observed deaths, and e is the number of expected deaths.
4.8. Longevity Relatives Count score
Based on the results of a recent study which shows that longevity is heritable beyond the 10% survivors of their birth cohort and that multiple family members, such as parents and/or aunts and uncles, should belong to the top 10% survivors (van den Berg et al., 2019), we constructed a novel score that summarizes the familial history of longevity, the LRC score.
where i refers to the persons for whom the score is built. k is an index referring to each relative of person i who are used to construct the score. Ni refers to the total number of relatives of person i, Pk is the sex and birth year‐specific survival percentile based on life tables of relative k, and I(Pk ≥ 0.9) indicates whether relative k belongs to the top Z% survivors, for example, the top 10% survivors (Z = top 10%). is the weighted total number of relatives of person i. The relationship coefficients are used as weights wk . For example, persons share on average 50% of their nuclear DNA with their parents and siblings and this is 25% for aunts, uncles, or grandparents. Hence, in the LRC, each parent and sibling contributes 0.5 to the score, while each aunt, uncle, or grandparent contributes only 0.25. This is consistent with a previous study of us, which shows that more distant long‐lived relatives associate significantly but less strong with a person's survival than a close long‐lived relative (van den Berg et al., 2019). The higher the score, the higher the familial aggregation level of longevity. For example, a score of 0.5 indicates that 50% of a person's relatives were long‐lived. We utilized the LRC score to map the proportion of long‐lived ancestors for all F3 descendants, select cases with the heritable longevity trait and controls resembling the general population, and compare the survival advantage of F3 descendants who had at least one long‐lived parent to those who had at least 30% long‐lived descendants. The LRC scores were based on all identified relatives of F3 descendants with sufficient data quality (Figures S4 and S5).
4.9. Survival analysis (Cox‐type random effects regression model)
To investigate the extent of a survival difference between the family F3 case and control group, we use a Cox‐type random effects model:
where tij is the age at death for person j in family i. λ0(tij) refers to the baseline hazard, which is left unspecified in a Cox‐type model. β is the vector of regression coefficients for the main effects of interest (Z). γ is a vector of regression coefficients for the effects of covariates and possible confounders (X). ui > 0 refers to an unobserved random effect (frailty). In all Cox models, we adjust for sibship size, birth year, and sex.
CONFLICT OF INTEREST
The authors declare no competing interests.
AUTHOR CONTRIBUTIONS
Niels van den Berg is the study investigator and was responsible for initiating the study, data management, data analyses, writing the manuscript, and finalizing it. Angelique Janssens and P. Eline Slagboom are the study principal investigators who conceived and obtained funding for the project, which this study is a part of. P. Eline Slagboom and Marian Beekman provided overall project coordination and supervision. Mar Rodriguez‐Girondo provided overall statistical analyses coordination and supervision. Kees Mandemakers is the head of the HSN and provided access and support to the HSN data, and supervised the new data collection for this study.
CODE AVAILABILITY
The scripts containing the code for data preprocessing and data analyses can be freely downloaded at https://git.lumc.nl/molepi/PUBLIC/LRCscore.
Supporting information
Supplementary Material
van den Berg N, Rodríguez‐Girondo M, Mandemakers K, Janssens AAPO, Beekman M, Slagboom PE. Longevity Relatives Count score identifies heritable longevity carriers and suggests case improvement in genetic studies. Aging Cell. 2020;19:e13139 10.1111/acel.13139
DATA AVAILABILITY STATEMENT
The data used for this study will be made freely available at the Data Archiving and Networked Services (DANS) repository. The available data are recoded and trimmed in accordance with Dutch and international privacy legislation. An instruction file on how to obtain the data can be found at https://git.lumc.nl/molepi/PUBLIC/LRCscore.
REFERENCES
- Andersen, S. L. , Sebastiani, P. , Dworkis, D. A. , Feldman, L. , & Perls, T. T. (2012). Health span approximates life span among many supercentenarians: Compression of morbidity at the approximate limit of life span. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 67A(4), 395–405. 10.1093/gerona/glr223 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arbeeva, L. S. , Hanson, H. A. , Arbeev, K. G. , Kulminski, A. M. , Stallard, E. , Ukraintseva, S. V. , … Yashin, A. I. (2018). How well does the family longevity selection score work: A validation test using the Utah population database. Frontiers in Public Health, 6, 1‐7. 10.3389/fpubh.2018.00277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barral, S. , Haynes, C. , Stone, M. , & Gordon, D. (2006). LRTae: Improving statistical power for genetic association with case/control data when phenotype and/or genotype misclassification errors are present. BMC Genetics, 7, Article number 24. 10.1186/1471-2156-7-24 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broer, L. , Buchman, A. S. , Deelen, J. , Evans, D. S. , Faul, J. D. , Lunetta, K. L. , … Murabito, J. M. (2015). GWAS of longevity in CHARGE consortium confirms APOE and FOXO3 candidacy. Journals of Gerontology ‐ Series A Biological Sciences and Medical Sciences, 70(1), 110–118. 10.1093/gerona/glu166 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bross, I. (1954). Misclassification in 2 X 2 tables. Biometrics, 10(4), 478‐486. 10.2307/3001619 [DOI] [Google Scholar]
- Buyske, S. , Yang, G. , Matise, T. C. , & Gordon, D. (2009). When a case is not a case: Effects of phenotype misclassification on power and sample size requirements for the transmission disequilibrium test with affected child trios. Human Heredity, 67(4), 287–292. 10.1159/000194981 [DOI] [PubMed] [Google Scholar]
- Carolina, T. , Uijvenhoven, L. , & van der Laan, J. (2009). Overlevingstafels en longitudinale analyse. CBS, 10010, 1–25. Retrieved from https://www.cbs.nl/nl‐nl/onze‐diensten/methoden/statistische‐methoden/output/output/overlevingstafels‐en‐longitudinale‐analyse‐survival‐analyse‐duurmodellen. [Google Scholar]
- Christensen, K. , McGue, M. , Petersen, I. , Jeune, B. , & Vaupel, J. W. (2008). Exceptional longevity does not result in excessive levels of disability. Proceedings of the National Academy of Sciences, 105(36), 13274–13279. 10.1073/pnas.0804931105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christensen, K. , Thinggaard, M. , Oksuzyan, A. , Steenstrup, T. , Andersen‐Ranberg, K. , Jeune, B. , … Vaupel, J. W. (2013). Physical and cognitive functioning of people older than 90 years: A comparison of two Danish cohorts born 10 years apart. The Lancet, 382(9903), 1507–1513. 10.1016/S0140-6736(13)60777-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deelen, J. , Beekman, M. , Uh, H.‐W. , Broer, L. , Ayers, K. L. , Tan, Q. , … Slagboom, P. E. (2014). Genome‐wide association meta‐analysis of human longevity identifies a novel locus conferring survival beyond 90 years of age. Human Molecular Genetics, 23(16), 4420–4432. 10.1093/hmg/ddu139 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deelen, J. , Evans, D. S. , Arking, D. E. , Tesi, N. , Nygaard, M. , Liu, X. , … Murabito, J. M. (2019). A meta‐analysis of genome‐wide association studies identifies multiple longevity genes. Nature Communications, 10(1), Article number 3669. 10.1038/s41467-019-11558-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deluty, J. A. , Atzmon, G. , Crandall, J. , Barzilai, N. , & Milman, S. (2015). The influence of gender on inheritance of exceptional longevity. Aging, 7(6), 412–418. 10.18632/aging.100763 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dutta, A. , Henley, W. , Robine, J. M. , Langa, K. M. , Wallace, R. B. , & Melzer, D. (2013). Longer lived parents: Protective associations with cancer incidence and overall mortality. Journals of Gerontology ‐ Series A Biological Sciences and Medical Sciences, 68(11), 1409–1418. 10.1093/gerona/glt061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edwards, B. J. , Haynes, C. , Levenstien, M. A. , Finch, S. J. , & Gordon, D. (2005). Power and sample size calculations in the presence of phenotype errors for case/control genetic association studies. BMC Genetics, 6, 18. 10.1186/1471-2156-6-18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erlich, Y. , Shor, T. , Pe’er, I. , & Carmi, S. (2018). Identity inference of genomic data using long‐range familial searches. Science, 362(6415), 690–694. 10.1126/science.aau4832 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flachsbart, F. , Caliebe, A. , Kleindorp, R. , Blanche, H. , von Eller‐Eberstein, H. , Nikolaus, S. , … Nebel, A. (2009). Association of FOXO3A variation with human longevity confirmed in German centenarians. Proceedings of the National Academy of Sciences, 106(8), 2700–2705. 10.1073/pnas.0809594106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flachsbart, F. , Ellinghaus, D. , Gentschew, L. , Heinsen, F.‐A. , Caliebe, A. , Christiansen, L. , … Nebel, A. (2016). Immunochip analysis identifies association of the RAD50/IL13 region with human longevity. Aging Cell, 15(3), 585–588. 10.1111/acel.12471 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gavrilov, L. A. , & Gavrilova, N. S. (2015). Predictors of exceptional longevity: Effects of early‐life and midlife conditions, and familial longevity. North American Actuarial Journal, 19(3), 174–186. 10.1080/10920277.2015.1018390 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gordon, D. (2005). Factors affecting statistical power in the detection of genetic association. Journal of Clinical Investigation, 115(6), 1408–1418. 10.1172/JCI24756 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gordon, D. , Haynes, C. , Yang, Y. , Kramer, P. L. , & Finch, S. J. (2007). Linear trend tests for case–control genetic association that incorporate random phenotype and genotype misclassification error. Genetic Epidemiology, 31(8), 853–870. 10.1002/gepi.20246 [DOI] [PubMed] [Google Scholar]
- Gudmundsson, H. , Gudbjartsson, D. F. , Kong, A. , Gudbjartsson, H. , Frigge, M. , Gulcher, J. R. , & Stefánsson, K. (2000). Inheritance of human longevity in Iceland. European Journal of Human Genetics, 8(10), 743–749. 10.1038/sj.ejhg.5200527 [DOI] [PubMed] [Google Scholar]
- Herskind, A. M. , McGue, M. , Holm, N. V. , Sørensen, T. I. A. , Harvald, B. , & Vaupel, J. W. (1996). The heritability of human longevity: A population‐based study of 2872 Danish twin pairs born 1870–1900. Human Genetics, 97(3), 319–323. 10.1007/BF02185763 [DOI] [PubMed] [Google Scholar]
- Houde, L. , Tremblay, M. , & Vézina, H. (2008). Intergenerational and genealogical approaches for the study of longevity in the Saguenay‐Lac‐St‐Jean population. Human Nature, 19(1), 70–86. 10.1007/s12110-008-9031-7 [DOI] [PubMed] [Google Scholar]
- Hujoel, M. L. A. , Gazal, S. , Loh, P. , Patterson, N. , & Alkes, L. (2019). Combining case‐control status and family history of disease increases association power, pp. 1–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarry, V. , Gagnon, A. , & Bourbeau, R. (2013). Survival advantage of siblings and spouses of centenarians in 20th‐century Quebec. Canadian Studies in Population, 39(3–4), 67‐78. 10.25336/P65C9R [DOI] [Google Scholar]
- Ji, F. , Yang, Y. , Haynes, C. , Finch, S. J. , & Gordon, D. (2006). Computing asymptotic power and sample size for case‐control genetic association studies in the presence of phenotype and/or genotype misclassification errors. Statistical Applications in Genetics and Molecular Biology, 4(1). 10.2202/1544-6115.1184 [DOI] [PubMed] [Google Scholar]
- Joshi, P. K. , Pirastu, N. , Kentistou, K. A. , Fischer, K. , Hofer, E. , Schraut, K. E. , … Wilson, J. F. (2017). Genome‐wide meta‐analysis associates HLA‐DQA1/DRB1 and LPA and lifestyle factors with human longevity. Nature Communications, 8(1), 1–13. 10.1038/s41467-017-00934-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaplanis, J. , Gordon, A. , Shor, T. , Weissbrod, O. , Geiger, D. , Wahl, M. , … Erlich, Y. (2018). Quantitative analysis of population‐scale family trees with millions of relatives. Science, 360(6385), 171–175. 10.1126/science.aam9309 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kemkes‐Grottenthaler, A. (2004). Parental effects on offspring longevity—Evidence from 17th to 19th century reproductive histories. Annals of Human Biology, 31(2), 139–158. 10.1080/03014460410001663407 [DOI] [PubMed] [Google Scholar]
- Kerber, R. A. , Brien, E. O. , Smith, K. R. , & Cawthon, R. M. (2001). Familial excess longevity in Utah genealogies. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 56(3), 130–139. 10.1093/gerona/56.3.B130 [DOI] [PubMed] [Google Scholar]
- Liu, J. Z. , Erlich, Y. , & Pickrell, J. K. (2017). Case–control association mapping by proxy using family history of disease. Nature Genetics, 49(3), 325–331. 10.1038/ng.3766 [DOI] [PubMed] [Google Scholar]
- Mandemakers, K. (2000). Historical sample of the Netherlands In Hall P. K., McCaa R., & Thorvaldsen G. (Eds.), Handbook of international historical microdata for population research (pp. 149–177). Minneapolis, MN: Minnesota Population Cente; [Google Scholar]
- Mandemakers, K. (2002). Building life courses datasets from population registers by the historical sample of the Netherlands In History and computing (vol. 14(1‐2), pp. 87–107). Edinburgh, UK: Edinburgh University Press; 10.3366/hac.2002.14.1-2.87 [DOI] [Google Scholar]
- Mandemakers, K. (2010). https://socialhistory.org/en/hsn/hsn‐releases.
- Montesanto, A. , Latorre, V. , Giordano, M. , Martino, C. , Domma, F. , & Passarino, G. (2011). The genetic component of human longevity: Analysis of the survival advantage of parents and siblings of Italian nonagenarians. European Journal of Human Genetics, 19(8), 882–886. 10.1038/ejhg.2011.40 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mourits, R. J. , van den Berg, N. , Rodriguez‐Girondo, M. , Mandemakers, K. , Slagboom, P. E. , Beekman, M. , & Janssens, A. A. P. O. (2019). Intergenerational transmission of longevity is not affected by other familial factors: Evidence from 16,905 Dutch families from Zeeland, 1812–1962. BioRxiv, 1–40. 10.1101/781500 [DOI] [Google Scholar]
- Oeppen, J. , & Vaupel, J. W. (2002). Broken limits to life expectancy. Science, 296(5570), 1029–1031. [DOI] [PubMed] [Google Scholar]
- Partridge, L. , Deelen, J. , & Slagboom, P. E. (2018). Facing up to the global challenges of ageing. Nature, 561(7721), 45–56. 10.1038/s41586-018-0457-8 [DOI] [PubMed] [Google Scholar]
- Pedersen, J. K. , Elo, I. T. , Schupf, N. , Perls, T. T. , Stallard, E. , Yashin, A. I. , & Christensen, K. (2017). The survival of spouses marrying into longevity‐enriched families. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 72(1), 109–114. 10.1093/gerona/glw159 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perls, T. T. , Wilmoth, J. , Levenson, R. , Drinkwater, M. , Cohen, M. , Bogan, H. , … Puca, A. (2002). Life‐long sustained mortality advantage of siblings of centenarians. Proceedings of the National Academy of Sciences, 99(12), 8442–8447. 10.1073/pnas.122587599 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pilling, L. C. , Kuo, C.‐L. , Sicinski, K. , Tamosauskaite, J. , Kuchel, G. A. , Harries, L. W. , … Melzer, D. (2017). Human longevity: 25 genetic loci associated in 389,166 UK biobank participants. Aging, 9(12), 2504–2520. 10.18632/aging.101334 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Platz, E. A. , De Marzo, A. M. , & Giovannucci, E. (2004). Prostate cancer association studies: Pitfalls and solutions to cancer misclassification in the PSA era. Journal of Cellular Biochemistry, 91(3), 553–571. 10.1002/jcb.10700 [DOI] [PubMed] [Google Scholar]
- R Core Team . (2016). R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. [Google Scholar]
- Rekaya, R. , Smith, S. , Hay, E. H. , Farhat, N. , & Aggrey, S. (2016). Analysis of binary responses with outcome‐specific misclassification probability in genome‐wide association studies. The Application of Clinical Genetics, 9, 169–177. 10.2147/TACG.S122250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Royall, D. R. , Chiodo, L. K. , & Polk, M. J. (2004). Misclassification is likely in the assessment of mild cognitive impairment. Neuroepidemiology, 23(4), 185–191. 10.1159/000078504 [DOI] [PubMed] [Google Scholar]
- Rozing, M. P. , Houwing‐Duistermaat, J. J. , Slagboom, P. E. , Beekman, M. , Frölich, M. , de Craen, A. J. M. , … van Heemst, D. (2010). Familial longevity is associated with decreased thyroid function. Journal of Clinical Endocrinology and Metabolism, 95(11), 4979–4984. 10.1210/jc.2010-0875 [DOI] [PubMed] [Google Scholar]
- Ruby, J. G. , Wright, K. M. , Rand, K. A. , Kermany, A. , Noto, K. , Curtis, D. , … Ball, C. (2018). Estimates of the heritability of human longevity are substantially inflated due to assortative mating. Genetics, 210(3), 1109–1124. 10.1534/genetics.118.301613 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schoenmaker, M. , de Craen, A. J. M. , de Meijer, P. H. E. M. , Beekman, M. , Blauw, G. J. , Slagboom, P. E. , & Westendorp, R. G. J. (2006). Evidence of genetic enrichment for exceptional survival using a family approach: The Leiden Longevity Study. European Journal of Human Genetics, 14(1), 79–84. 10.1038/sj.ejhg.5201508 [DOI] [PubMed] [Google Scholar]
- Sebastiani, P. , Gurinovich, A. , Bae, H. , Andersen, S. , Malovini, A. , Atzmon, G. , … Perls, T. T. (2017). Four genome‐wide association studies identify new extreme longevity variants. The Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 72(11), 1453–1464. 10.1093/gerona/glx027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sebastiani, P. , Hadley, E. C. , Province, M. , Christensen, K. , Rossi, W. , Perls, T. T. , & Ash, A. S. (2009). A family longevity selection score: Ranking sibships by their longevity, size, and availability for study. American Journal of Epidemiology, 170(12), 1555–1562. 10.1093/aje/kwp309 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sebastiani, P. , Nussbaum, L. , Andersen, S. L. , Black, M. J. , & Perls, T. T. (2016). Increasing sibling relative risk of survival to older and older ages and the importance of precise definitions of “Aging”, “Life Span”, and “Longevity”. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 71(3), 340–346. 10.1093/gerona/glv020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sebastiani, P. , & Perls, T. T. (2012). The genetics of extreme longevity: Lessons from the New England Centenarian Study. Frontiers in Genetics, 3, 1–7. 10.3389/fgene.2012.00277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shadyab, A. H. , & LaCroix, A. Z. (2015). Genetic factors associated with longevity: A review of recent findings. Ageing Research Reviews, 19, 1–7. 10.1016/j.arr.2014.10.005 [DOI] [PubMed] [Google Scholar]
- Slagboom, P. E. , van den Berg, N. , & Deelen, J. (2018). Phenome and genome based studies into human ageing and longevity: An overview. Biochimica Et Biophysica Acta (BBA) ‐ Molecular Basis of Disease, 1864(9), 2742–2751. 10.1016/j.bbadis.2017.09.017 [DOI] [PubMed] [Google Scholar]
- Smith, S. , Hay, E. , Farhat, N. , & Rekaya, R. (2013). Genome wide association studies in presence of misclassified binary responses. BMC Genetics, 14(1), Article number 124. 10.1186/1471-2156-14-124 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terry, D. F. , Wilcox, M. A. , McCormick, M. A. , Pennington, J. Y. , Schoenhofen, E. A. , Andersen, S. L. , & Perls, T. T. (2004). Lower all‐cause, cardiovascular, and cancer mortality in centenarians’ offspring. Journal of the American Geriatrics Society, 52(12), 2074–2076. 10.1111/j.1532-5415.2004.52561.x [DOI] [PubMed] [Google Scholar]
- Ulm, K. (1990). Simple method to calculate the confidence interval of a standardized mortality ratio (SMR). American Journal of Epidemiology, 131(2), 373–375. 10.1134/S1063783415100224 [DOI] [PubMed] [Google Scholar]
- van den Berg, N. , Beekman, M. , Smith, K. R. , Janssens, A. , & Slagboom, P. E. (2017). Historical demography and longevity genetics: Back to the future. Ageing Research Reviews, 38, 28–39. 10.1016/j.arr.2017.06.005 [DOI] [PubMed] [Google Scholar]
- van den Berg, N. , Rodríguez‐Girondo, M. , de Craen, A. J. M. , Houwing‐Duistermaat, J. J. , Beekman, M. , & Slagboom, P. E. (2018). Longevity around the turn of the 20th century: Life‐long sustained survival advantage for parents of today’s nonagenarians. The Journals of Gerontology: Series A, 73(10), 1295–1302. 10.1093/gerona/gly049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van den Berg, N. , Rodríguez‐Girondo, M. , van Dijk, I. K. , Mourits, R. J. , Mandemakers, K. , Janssens, A. A. P. O. , … Slagboom, P. E. (2019). Longevity defined as top 10% survivors and beyond is transmitted as a quantitative genetic trait. Nature Communications, 10(1), Article number 35. 10.1038/s41467-018-07925-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van den Berg, N. , van Dijk, I. K. , Mourits, R. J. , Slagboom, P. E. , Janssens, A. A. P. O. , & Mandemakers, K. (2020). Families in Comparison: An individual‐level comparison of life course and family reconstructions between population and vital event registers. Population Studies. 10.1080/00324728.2020.1718186 [DOI] [PubMed] [Google Scholar]
- Van Der Meulen, A. (2012). Life tables and Survival analysis. Retrieved from https://www.cbs.nl/en‐gb/our‐services/methods/statistical‐methods/output/output/life‐tables [Google Scholar]
- Vaupel, J. W. (1998). Biodemographic trajectories of longevity. Science, 280(5365), 855–860. 10.1126/science.280.5365.855 [DOI] [PubMed] [Google Scholar]
- Willcox, B. J. , Donlon, T. A. , He, Q. , Chen, R. , Grove, J. S. , Yano, K. , … Curb, J. D. (2008). FOXO3A genotype is strongly associated with human longevity. Proceedings of the National Academy of Sciences, 105(37), 13987–13992. 10.1073/pnas.0801030105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willcox, B. J. , Willcox, D. C. , He, Q. , Curb, J. D. , & Suzuki, M. (2006). Siblings of Okinawan centenarians share lifelong mortality advantages. The Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 61(4), 345–354. 10.1093/gerona/61.4.345 [DOI] [PubMed] [Google Scholar]
- You, D. , Danan, G. U. , & Yi, Z. (2010). Familial transmission of human longevity among the oldest‐old in China. Journal of Applied Gerontology, 29(3), 308–332. 10.1177/0733464809340154 [DOI] [Google Scholar]
- Zeng, Y. I. , Nie, C. , Min, J. , Liu, X. , Li, M. , Chen, H. , … Vaupel, J. W. (2016). Novel loci and pathways significantly associated with longevity. Scientific Reports, 6(1), Article number 21243. 10.1038/srep21243 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Material
Data Availability Statement
The data used for this study will be made freely available at the Data Archiving and Networked Services (DANS) repository. The available data are recoded and trimmed in accordance with Dutch and international privacy legislation. An instruction file on how to obtain the data can be found at https://git.lumc.nl/molepi/PUBLIC/LRCscore.