

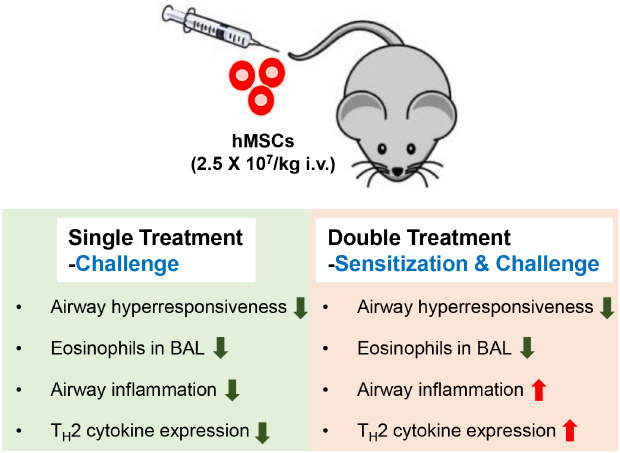
Abstract
Background
Studies in experimental models of allergic asthma have shown that mesenchymal stem cells (MSCs) have therapeutic potential for T-helper 2 (TH2) cell-mediated inflammation. However, the mechanisms underlying these therapeutic effects are not fully understood and their safety has not been confirmed.
Methods
Using a mouse model of experimental allergic asthma, we investigated the efficacy of human adipose-derived mesenchymal stem cells (hADSCs) or human bone marrow-derived mesenchymal stem cells (hBMSCs) according to treatment frequency and timing.
Results
Ovalbumin (OVA)-sensitized and -challenged mice exhibited airway hyperresponsiveness (AHR), airway inflammation, and significant increases in TH2 cytokine levels. Both double and single human mesenchymal stem cell (hMSC) treatments significantly decreased AHR and bronchoalveolar lavage fluid counts. In addition, single treatment with hMSCs showed significant attenuation of allergic airway inflammation. However, double treatment with hMSCs during OVA -sensitization and -challenge further increased inflammatory cell infiltration, and TH2 cytokine levels.
Conclusion
The results of treatment with hADSCs or hBMSCs suppresses AHR and airway inflammation. However, double hMSC treatment significantly induces eosinophilic airway inflammation and lung histological changes. Therefore, double hMSC treatment is ineffective against asthma and single injection frequency appears to be more important for the treatment of asthma. These results suggest that hMSC therapy can be used for treatment of asthma patients but that it should be used carefully.
Keywords: Human MSC, Frequency, Airway Inflammation, Asthma
Graphical Abstract
INTRODUCTION
Bronchial asthma is a chronic inflammatory disease characterized by airway hyperresponsiveness (AHR), airway inflammation, and airway structural changes.1,2 Airway inflammation and structural changes are associated with irreversible loss of lung function that tracks from childhood to adulthood.3 Although symptoms can be controlled in most patients using anti-inflammatory drugs and bronchodilators, they remain uncontrolled in 10% of patients.1,4,5 Therefore, there is a clear need for new drugs for the treatment of patients with uncontrolled asthma.
Mesenchymal stem cells (MSCs) are multipotent progenitor cells with the ability to self-renew and differentiate into multiple cell types.6 They can control inflammation by modulating T lymphocytes (T cells), natural killer (NK) cells, and dendritic cells (DCs),6,7,8 and can be isolated easily from different adult tissues, including adipose tissue (adipose-derived stem cells, ADSCs) and bone marrow (bone marrow-derived mesenchymal stem cells, BMSCs).9 Significant advances have been made in the use of stem cells to treat various diseases, such as acute myocardial infarction (AMI),10 liver cirrhosis,11 Crohn's disease, and graft-versus-host disease (GVHD).12
MSCs show anti-inflammatory effects in mouse models of asthma.13,14,15,16,17 These effects are associated with the induction of TGF-β,18,19 IFN-γ,16 recruitment of regulatory T cells,15,20,21 and modulation of the T-helper 1 (TH1) to T-helper 2 (TH2) cell balance.16 Similarly, human MSCs have been shown to ameliorate airway inflammation in mouse models of acute and chronic asthma.22 Taken together, these studies suggest that MSCs may play a role in ameliorating airway inflammation in allergic asthma. However, there have been no reports of clinical data regarding the efficacy of MSCs for the treatment of asthma.
The present study investigated the efficacy of human mesenchymal stem cells (hMSCs) in a mouse acute asthma model. We focused on the treatment frequency and timing of human ADSCs (hADSCs) and human BMSCs (hBMSCs) in a mouse model of acute asthma.
METHODS
Mice
Female, 6-week-old, BALB/c mice (Orient Bio, Seongnam, Korea) were used in all experiments. The mice were divided into the following groups: 1) control (CON, n = 5–6), 2) ovalbumin (OVA, n = 5–6) challenge, 3) OVA challenge + phosphate buffered saline (PBS) as vehicle (OVA + PBS, n = 5–8), 4) OVA challenge + hADSCs (OVA + hADSC, n = 5–8), and 5) OVA challenge + hBMSCs (OVA + hBMSC, n = 5–8).
Sensitization and antigen challenge protocol
The mice were immunized with 25 µg OVA (chicken egg albumin, grade V; Sigma-Aldrich, St. Louis, MO, USA) in 1 mg aluminum hydroxide (Sigma-Aldrich, Milwaukee, WI, USA) in 200 µL PBS according to our previous report.23 Mice were sacrificed 24 hours after the last OVA challenge. Lung tissues and bronchoalveolar lavage fluid (BALF) were collected.
Isolation of hADSCs and hBMSCs
hADSCs and hBMSCs were isolated from 17 adult donors with approval from the Institutional Review Board of our institution, and characterized as described previously.24 Briefly, the cells isolated from adipose tissue and bone marrow were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 20% fetal bovine serum (Gibco, Grand Island, NY, USA) at 37°C in a 5% CO2 humidified incubator. After 72 hours, nonadherent cells were removed and adherent cells were cultured for an additional 14 days. The adherent cells were retrieved using 0.25% trypsin (Gibco) and sorted using CD13+, CD90+, CD73+, CD105+, CD34−, CD45−, CD146−, and HLA− markers to obtain hADSCs (R&D Systems, Minneapolis, MN, USA). The sorted cells were enriched further by culturing.
Administration of hADSCs or hBMSCs
Vials of frozen hADSCs and hBMSCs were thawed and expanded according to the supplier's instructions. The hMSCs were used in all experiments between passages 3 and 4. Subsequently, 2.5 × 107/kg hADSCs and hBMSCs in 100 µL PBS were administered using an insulin syringe via the tail vein following last sensitization and OVA challenge. A single injection of hMSCs was performed during OVA challenge and double injection of hMSCs was performed during OVA sensitization and OVA challenge into the tail vein of the mice.
Measurement of AHR
Airway resistance (Rrs) to methacholine (Sigma-Aldrich) was measured by the FlexiVent system (SCIREQ, Montreal, QC, Canada) as reported previously.23 In brief, mice were exposed for 3 minutes to nebulized PBS. Baseline Rrs was established by PBS and Rrs was measured according to the increased concentration of nebulized methacholine (Sigma-Aldrich). Aerosonic ultrasonic nebulizer (DeVilbiss, Somerset, PA, USA) was used for nebulization.
Cell counts of bronchoalveolar lavage (BAL)
BALF was performed immediately after the measurement of AHR as previously reported.23 The BALF was cytospun at 2,000 rpm for 7 minutes onto microscope slides. It was stained with Diff-Quik (Sysmax, Kobe, Japan). The percentages of BALF macrophages, neutrophils, lymphocytes, and eosinophils were measured. Five hundred leukocytes were counted on randomly selected fields under a light microscope.
Enzyme-linked immunosorbent assay
The concentrations of interleukin (IL)-4, IL-5, and IL-13 in lung homogenate proteins were measured via enzyme-linked immunosorbent assay (ELISA) using commercial kits in accordance with the manufacturer's instructions (R&D Systems, Minneapolis, MN, USA).
Hematoxylin and eosin (H&E) staining
Lungs were fixed in 4% paraformaldehyde for 24 hours and then embedded in paraffin. Sections were cut at a thickness of 4 μm using a microtome, and deparaffinized tissue sections were subjected to H&E staining to examine airway inflammation.
Statistical analyses
Results are shown as the mean ± standard error of mean. Analysis of variance (ANOVA) with the nonparametric Kruskal-Wallis test, followed by post hoc Dunn's multiple comparison of means was used for the comparison between groups. Graph Pad Prism (Graph Pad, San Diego, CA, USA) statistical software package was utilized for the analyses. In all analyses, P < 0.05 was taken to indicate statistical significance.
Ethics statement
All procedure of animal research was provided in accordance with the Laboratory Animal Welfare Act, the Guide for the Care and Use of Laboratory Animals and the Guidelines and Policies for Rodent experiment provided by the Institutional Animal Care and Use Committee (IACUC) in School of Medicine, The Catholic University of Korea (IACUC Approval number: CUMC-2016-00654-01).
RESULTS
Double injection of hMSCs leads to greater improvement of AHR
To investigate the effects of the injection of hADSCs or hBMSCs on allergic asthma, we first measured AHR after the last OVA challenge (Fig. 1A). Rrs values of OVA challenge and in the PBS-treated OVA (OVA+PBS) mice showed significant increases in a methacholine dose-dependent manner compared to CON mice (P < 0.01). However, double administration of hADSCs or hBMSCs to mice given OVA significantly inhibited AHR to the same degree as in CON mice (P < 0.001) (Fig. 1B).
Fig. 1. Double injection of human mesenchymal stem cells inhibited airway hyperresponsiveness and airway inflammation. (A) Schematic diagram of the mouse model of acute asthma and treatment with hADSCs or hBMSCs. Mice were sensitized on days 0 and 7 by subcutaneous injection of OVA and challenged intranasally on days 21, 23, and 25 (black dots). Aliquots of 2.5 × 107/kg hADSCs or hBMSCs were injected via the tail vein on days 7 and 27 (blue triangles). Samples were harvested on day 29. (B) Rrs was used to measure airway hyperresponsiveness with increasing concentration of nebulized methacholine. *P < 0.05, **P < 0.01 compared to the CON group; ##P < 0.01, ###P < 0.001 compared to the OVA+PBS group. (C) BALF cells were isolated by centrifugation and stained with Diff-Quik. Total numbers of macrophages, eosinophils, lymphocytes, and neutrophils in the BALF were counted. ***P < 0.001 compared to the CON group; #P < 0.05, ###P < 0.001 compared to the OVA group.

hADSC = human adipose-derived mesenchymal stem cell, hBMSC = human bone marrow-derived mesenchymal stem cell, OVA = ovalbumin, Alum = aluminum, Rrs = airway resistance, CON = control, PBS = phosphate buffered saline, BAL = bronchoalveolar lavage, BALF = bronchoalveolar lavage fluid, S.c.= subcutaneous, I.n.= intranasal.
Airway inflammation is attenuated by hMSC double injection in BALF cells
Airway inflammation is another important feature of allergic asthma. To examine the effects of double injection of hMSCs in the mouse model of asthma, total and differential cell counts in BALF were determined. However, we were unable to count the total number of BALF cells in hADSC double injection mice, as there were few viable cells. The treatment seems to adversely affect the viability of BALF cells, so BAL studies were limited to counting only hBMSCs. OVA-challenged mice and OVA+PBS mice showed significantly increased total cell numbers and eosinophils levels compared to CON mice (P < 0.001). However, double treatment with hBMSCs markedly reduced infiltration of inflammatory cells in the airway according to the absolute cell count in BALF compared to OVA+PBS mice (total cells, P < 0.001; eosinophils, P < 0.05) (Fig. 1C). These findings indicate that double hMSC treatment inhibited the recruitment of eosinophils into the airway and BALF and reduced AHR in asthmatic mice.
Double injection of hMSCs further increases lung inflammation
To investigate the extent of inflammatory cell infiltration, we examined histological changes in OVA-induced asthmatic lung tissues. OVA-induced mice exhibited marked infiltration of inflammatory cells into peribronchial and perivascular lesions in the lung tissue. Interestingly, hADSC or hBMSC double injected mice showed further increases in inflammatory cell infiltration compared to OVA+PBS mice (Fig. 2A). Moreover, expanded images clearly showed that OVA challenge induced inflammation around peribronchial (Fig. 2B) and perivascular (Fig. 2C) lesions. Strikingly, hADSC or hBMSC double injection markedly increased the accumulation of inflammatory cells. Furthermore, the results of H&E staining showed that hMSC double treatment induced goblet cell hyperplasia compared to the OVA+PBS group and smooth muscle hyperplasia (Fig. 2B). Histological analyses of tissue inflammation in lungs of mice showed that human MSCs significantly alter tissue inflammation (Fig. 2D). Specifically, perivascular-associated inflammation was robustly induced in hADSCs and hBMSCs treatment. These results suggest that double injection of hMSCs significantly decreased total cell number and eosinophils in BALF, but exacerbated lung pathology, such as lung inflammation.
Fig. 2. Double injection of hMSCs led to more severe airway inflammation. (A–C) Representative photomicrographs of hematoxylin and eosin-stained lung sections. (A) The lungs in the mouse model of acute asthma showing airways and blood vessels surrounded by alveoli (×100). (B) Airway (×400). Black arrows indicate smooth muscle, and black stars indicate goblet cell metaplasia. (C) Blood vessel (×400). (D) Histological quantification of peribronchial and perivascular inflammation in the lung. ***P < 0.001 compared to the CON group; #P < 0.05 compared to the OVA+PBS group.

CON = control, OVA = ovalbumin, PBS = phosphate buffered saline, hADSC = human adipose-derived mesenchymal stem cell, hBMSC = human bone marrow-derived mesenchymal stem cell, Br = bronchus, Bm = basement membrane, Bl = blood vessel, Eo = eosinophil, Ep = epithelium.
Double injection of hMSCs increases TH2 cytokine production in BALF
To understand the mechanisms underlying hADSC- or hBMSC-mediated stimulation of lung inflammation, we assayed key mediators of asthma in our model. TH2 cytokines are important for the pathophysiological features of allergic asthma.25 We examined the levels of TH2 cytokines in the BALF of mice. As expected, levels of IL-4, IL-5, and IL-13 were significantly increased in OVA-challenged mice compared to CON mice (P < 0.05). Interestingly, the levels of each were increased to a greater extent in hADSC- or hBMSC-double treated mice compared to PBS-treated OVA mice (Fig. 3). These observations indicate that treatment with hMSCs induced airway inflammation by promoting IL-4, IL-5, and IL-13 secretion.
Fig. 3. Double injection of human mesenchymal stem cells exacerbated levels of T-helper 2 cytokine. Bronchoalveolar lavage fluid was obtained from the mouse model of acute asthma with and without treatment with hADSCs or hBMSCs and the levels of IL-4, IL-5, and IL-13 were measured.

IL = interleukin, CON = control, OVA = ovalbumin, PBS = phosphate buffered saline, hADSC = human adipose-derived mesenchymal stem cell, hBMSC = human bone marrow-derived mesenchymal stem cell.
*P < 0.05 compared to the CON group; #P < 0.05, ##P < 0.01, ###P < 0.001 compared to the OVA+PBS group.
Single injection of hMSCs also reduces AHR
The optimal treatment timing with hMSCs for asthma patients has not been established. Therefore, we investigated the treatment timing-dependent effects on features of asthma in a mouse model of OVA-induced asthma. Mice were sensitized with OVA/aluminum hydroxide and then challenged three times with intranasal OVA. A single injection of hMSCs was performed after the last challenge (Fig. 4A). We first checked the AHR. OVA-challenged mice showed increased AHR to methacholine compared to CON mice. Single treatment with hADSCs and hBMSCs clearly showed significant inhibition of the increase in resistance induced by OVA (Fig. 4B). There were no significant differences in these results between animals given double or single hMSC injections (Figs. 1B and 4B). These results indicate that injection frequency is not important with regard to attenuation of AHR by hMSCs in this mouse model of acute asthma.
Fig. 4. Single injection of human mesenchymal stem cells reduced airway hyperresponsiveness and airway inflammation. (A) Schematic diagram of the mouse model of acute asthma and treatment with hADSCs or hBMSCs. Mice were sensitized on days 0 and 7 by subcutaneous injection of OVA and challenged intranasally on days 21, 23, and 25 (black dots). Aliquots of 2.5 × 107/kg hADSCs or hBMSCs were injected via the tail vein on day 27 (blue triangles). Samples were harvested on day 29. (B) Rrs was used to measure airway hyperresponsiveness with increasing concentration of nebulized methacholine. *P < 0.05, **P < 0.01, ***P < 0.001 compared to the CON group; #P < 0.05, ##P < 0.01, ###P < 0.001 compared to the OVA+PBS group. (C) BAL cells were isolated by centrifugation and stained with Diff-Quik. Total numbers of macrophages, eosinophils, lymphocytes, and neutrophils in the BAL fluid were counted. *P < 0.05, **P < 0.01, ***P < 0.001 compared to the CON group; #P < 0.05, ##P < 0.01, ###P < 0.001 compared to the OBA+PBS group.

hADSC = human adipose-derived mesenchymal stem cell, hBMSC = human bone marrow-derived mesenchymal stem cell, OVA = ovalbumin, Alum = aluminum, Rrs = airway resistance, CON = control, PBS = phosphate buffered saline, BAL = bronchoalveolar lavage, S.c= subcutaneous, I.n.= intranasal.
Single hMSC injection is more effective for reducing airway inflammation
To confirm the effects of hMSCs on airway inflammation, we performed differential cell counts and measurement of TH2 cytokines in BALF. As shown in Fig. 4C, total cell counts and the percentage of eosinophils in OVA-challenged mice were significantly increased compared to CON mice (P < 0.001). However, there were robust decreases in both hADSC- and hBMSC-treated mice compared to PBS-treated mice given OVA (P < 0.001 and P < 0.01, respectively) (Fig. 4C). Next, we examined the levels of TH2 cytokines in BLAF to further define the antiinflammatory effects. A single injection of hADSCs and hBMSCs resulted in significant decreases in levels of IL-4, IL-5, and IL-13 in the BALF (Fig. 5). These data strongly suggest that single injection of hMSCs attenuated airway inflammation.
Fig. 5. Single injection with human mesenchymal stem cells attenuated levels of T-helper 2 cytokine. Bronchoalveolar lavage fluid was obtained from the mouse model of acute asthma with and without hADSC or hBMSC treatment and levels of IL-4, IL-5, and IL-13 were measured.

IL = interleukin, CON = control, OVA = ovalbumin, PBS = phosphate buffered saline, hADSC = human adipose-derived mesenchymal stem cell, hBMSC = human bone marrow-derived mesenchymal stem cell.
**P < 0.01, ***P < 0.001 compared to the CON group; #P < 0.05, ##P < 0.01, ###P < 0.001 compared to the OVA or OVA+PBS group.
Single hMSC injection inhibits eosinophilic lung inflammation
We examined whether a single injection of hMSCs can suppress OVA-induced lung inflammation in a mouse model of acute asthma. In histological examinations, OVA-sensitized and challenged mice showed extensive infiltration of inflammatory cells and substantial changes in the peribronchial layers. Single injection of hADSCs and hBMSCs markedly reduced the lung inflammation induced by OVA sensitization and challenge (Fig. 6A). Also, OVA-sensitization/challenge caused dense peribronchial (Fig. 6B) and perivascular (Fig. 6C) accumulation of inflammatory cells. However, these changes were minimal in single-treated hMSC mice with OVA-sensitization/challenge (Fig. 6B and C). As for the peribronchial and perivascular inflammation, the degree of the accumulation of inflammatory cells was assessed as inflammation score (Fig. 6D). Thus, a single injection of hMSCs can protect against lung inflammation.
Fig. 6. Single injection with human mesenchymal stem cells reduced eosinophil infiltration and lung inflammation. (A) Paraffin-embedded lung tissue sections were stained with hematoxylin and eosin (original magnification: ×200). (B) Airway (× 400). Black arrows indicate smooth muscle, and black stars indicate goblet cell metaplasia. (C) Blood vessel (× 400). (D) Histological quantification of peribronchial and perivascular inflammation in the lung.

CON = control, OVA = ovalbumin, PBS = phosphate buffered saline, hADSC = human adipose-derived mesenchymal stem cell, hBMSC = human bone marrow-derived mesenchymal stem cell, Br = bronchus, Bm = basement membrane, Bl = blood vessel, Eo = eosinophil, Ep = epithelium.
***P < 0.001 compared to the CON group; ###P < 0.001 compared to the OVA+PBS group.
DISCUSSION
Asthma is a complex disease with various features, including airway inflammation and AHR. The optimal treatment strategy should relieve symptoms, attenuate inflammation, and prevent remodeling. The need for new treatment options for patients with asthma has prompted research in various fields of study. We assessed the effects of hMSCs in attenuating airway inflammation and AHR. We found that hMSCs suppressed AHR and airway inflammation according to the timing and frequency of injection.
Cell-based therapies are attractive approaches for the treatment of asthma and other diseases. MSC administration attenuates airway inflammation and AHR.13,14,15,16,17,20,21,22 Similarly, we found that both single and double injection of hMSCs suppressed AHR. However, histological changes, production of TH2 cytokines, and inflammatory cell counts differed according to injection timing. Furthermore, early hMSC injection induced lung inflammation and production of TH2 cytokines compared to the OVA-challenged group. Interestingly, late hMSC injection significantly reduced airway inflammation, such as BALF cell counts, TH2 cytokine levels, and lung inflammation. Several studies have reported that MSCs attenuate airway inflammation in experimental asthma. Lee et al.26 revealed that rat BMSCs effectively reduced toluene diisocyante-induced airway inflammation, remodeling, and AHR. Additionally, Ou-Yang et al.27 and Abreu et al.28 demonstrated that mouse MSCs (mMSCs) can migrate to sites of inflammation and protect against allergic airway inflammation in asthmatic mice. Kavanagh et al.29 demonstrated that mBMSCs induce Tregs and reduce allergen-driven pathology. Sun et al.30 revealed that hBMSCs attenuate allergic-specific inflammation and pathological changes in OVA-induced asthmatic mice. These studies evaluated the effects of hBMSC administration before challenge on allergic inflammation. However, in our study, double injection of hMSCs were treated during OVA-sensitization and challenge. These different methods may be related to the differing effects on airway inflammation. The limitations of this study are the failure to clarify this point. Further studies are required to investigate the effect of hMSCs in attenuating airway inflammation according to the timing and frequency of injection. Recent findings have demonstrated that MSCs actively interact with components of the innate immune system and divide both antiinflammatory and proinflammatory effects via these interactions.31,32,33,34 These switching mechanisms depend on the production of soluble mediators, such as inducible nitric oxide synthase in mice and indoleamine 2,3-dioxygenase in humans. The concentrations of these mediators may be important to trigger the switch between proinflammatory and antiinflammatory MSCs. These concepts may help explain the observation that OVA-induced pulmonary responses differed according to the timing of hMSC injection. In our first model, hMSCs were injected during OVA sensitization and after the last challenge with OVA. In the period of sensitization, the inflammatory microenvironment of lung tissue in not yet fully activated. Early injection of hMSCs had a profound effect on disease progression in a mouse model of acute asthma. A few reports have demonstrated that hMSCs tend to shift the TH2-polarized immune response in encephalomyelitis and cultured immune cells in vitro.35,36 Our observations are consistent with these findings in that treatment with hMSCs may be either ineffective or could potentially exacerbate immune responses in a model of TH2-mediated inflammation, such as asthma. However, in our second experiment, hMSCs were only injected after the last OVA challenge when lung tissue was sufficiently characterized by an inflammatory environment. Therefore, late injection of hMSCs had antiinflammatory effects and attenuated airway inflammation as determined by examining BALF inflammatory cell numbers, TH2 cytokine production, and lung histology. We found that hMSCs can modulate airway inflammatory condition by microenvironment of lung tissue. Therefore, the timing of MSC treatment is very important.
Cell dose, cell injection frequency, and injection site are crucial in MSC therapy. Recent studies have shown that the survival of injected MSCs is dependent on cell dose and injection frequency in an animal model of critical limb ischemia.37 Our study provides further evidence that hMSCs can significantly reduce airway inflammation and AHR. Notably, single hMSC injection improved OVA-induced airway inflammation to a greater extent than double injection of hMSCs in our mouse model. These observations indicate that injection frequency must be taken into consideration in the application of MSC therapies to the treatment of asthma. The ability to use a single injection of hMSCs has the advantage of reducing the costs of therapy. However, the mechanisms underlying the beneficial effects of hMSCs on AHR without decreasing airway inflammation are unclear. AHR and airway inflammation may not always occur at the same time.38,39 This focused our attention on other effector functions of T cell cytokines in the pathophysiology of asthma. Airway overexpression of other cytokines, including IL-25,40 IL-33,41 and thymic stromal lymphopoietin,42 indirectly promotes asthma by stimulating production of TH2 cytokines. Ballantyne et al.43 reported that IL-25 promotes AHR independently of its stimulatory effects on TH2 cytokine secretion. Strikingly, neutralization of IL-25 only during OVA challenge specifically prevents AHR during ongoing TH2 inflammation in the lungs. Similarly, IL-12p40, a subunit of IL-12 and IL-23, is required to inhibit AHR and peribronchial fibrosis, but not eosinophilic inflammation, in a mouse model of chronic asthma.44 Consistent with these observations, hMSC treatment may suppress AHR by inducing IL-25 production or IL-12p40 expression. Also, we showed that double-treated hMSCs significantly exacerbated perivascular-associated inflammation (Fig. 2C and D). Delivery routes are essential for therapy efficiency and systemic delivery by infusion in the obvious goal for cell therapy. However, the mean size of suspended MSCs is larger than the size of pulmonary capillaries. Thus, IV-injected MSCs are trapped within the pulmonary capillaries, causing pulmonary alteration.45,46 In this study, we found a striking difference between single- and double-treated MSCs on lung inflammation effect. It seems that MSC size and treatment frequency were the major reason behind lung trapping. The size of hMSCs in six passages was after culture expansion in monolayer. The average diameter of hMSCs in six passages was small (17.9 μm) and large (30.4 μm).47,48 This size is about three to five times larger than pulmonary capillary (11.3 μm in human and 5.7 μm in mouse),49 probably increasing the difficulty of the cells to pass through capillaries. Similarly, several papers revealed that lung trapping of MSCs has been observed to cause severe lung damage in mice models.50,51 Current knowledge does not allow us to make definite conclusions whether trapping of MSCs in the lung according frequency is harmful or beneficial. Therefore, strategies to decrease the mismatch between MSCs and the capillary diameters are needed for cell-based therapy: the route of administration (intratracheal or systematic), the method for repeated treatment.
In summary, we demonstrated a beneficial effect of hMSCs on AHR and airway inflammation dependent on the timing of injection in a mouse model of acute asthma. A single injection of hMSCs inhibited allergic airway inflammation. Double hMSC treatment may have harmful effects in OVA-induced lung inflammation. Compared to PBS-treated OVA mice, hADSC or hBMSC double treated mice showed exacerbation of lung inflammation and higher levels of TH2 cytokines, such as IL-4, IL-5, and IL-13. These observations indicate that care is required for hMSC administration. Further studies regarding the timing of hMSC administration are required.
Footnotes
Funding: This research was supported by the National Research Foundation of Korea (NRF) grant funded by the Ministry of Science, ICT & Future Planning (NRF-2018R1C1B6003980) and grants (18172MFDS182 and 14172MFDS974) from Ministry of Food and Drug Safety of Korea in 2018.
Disclosure: The authors have no potential conflicts of interest to disclose.
- Conceptualization: Jung CK, Rhee CK.
- Data curation: Kang JY, Lee SY, Kim YK.
- Formal analysis: Hur J, Jeon S, Kim Y.
- Methodology: Hur J.
- Writing - original draft: Hur J.
- Writing - review & editing: Rhee CK.
References
- 1.Olin JT, Wechsler ME. Asthma: pathogenesis and novel drugs for treatment. BMJ. 2014;349:g5517. doi: 10.1136/bmj.g5517. [DOI] [PubMed] [Google Scholar]
- 2.Holgate ST, Arshad HS, Roberts GC, Howarth PH, Thurner P, Davies DE. A new look at the pathogenesis of asthma. Clin Sci (Lond) 2009;118(7):439–450. doi: 10.1042/CS20090474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Saglani S, Lloyd CM. Novel concepts in airway inflammation and remodelling in asthma. Eur Respir J. 2015;46(6):1796–1804. doi: 10.1183/13993003.01196-2014. [DOI] [PubMed] [Google Scholar]
- 4.Barnes PJ. Current therapies for asthma. Promise and limitations. Chest. 1997;111(2) Suppl:17S–26S. doi: 10.1378/chest.111.2_supplement.17s. [DOI] [PubMed] [Google Scholar]
- 5.Riccioni G, Di Ilio C, D'Orazio N. Review: Pharmacological treatment of airway remodeling: inhaled corticosteroids or antileukotrienes? Ann Clin Lab Sci. 2004;34(2):138–142. [PubMed] [Google Scholar]
- 6.Nauta AJ, Fibbe WE. Immunomodulatory properties of mesenchymal stromal cells. Blood. 2007;110(10):3499–3506. doi: 10.1182/blood-2007-02-069716. [DOI] [PubMed] [Google Scholar]
- 7.Le Blanc K, Ringdén O. Immunomodulation by mesenchymal stem cells and clinical experience. J Intern Med. 2007;262(5):509–525. doi: 10.1111/j.1365-2796.2007.01844.x. [DOI] [PubMed] [Google Scholar]
- 8.Rasmusson I. Immune modulation by mesenchymal stem cells. Exp Cell Res. 2006;312(12):2169–2179. doi: 10.1016/j.yexcr.2006.03.019. [DOI] [PubMed] [Google Scholar]
- 9.Nejad-Moghaddam A, Panahi Y, Abdollahpour Alitappeh M, Borna H, Shokrgozar MA, Ghanei M. Therapeutic potential of mesenchymal stem cells for the treatment of airway remodeling in pulmonary diseases. Iran J Allergy Asthma Immunol. 2015;14(6):552–568. [PubMed] [Google Scholar]
- 10.Kanelidis A, Premer C, Lopez JG, Balkan W, Hare JM. Route of delivery modulates the efficacy of mesenchymal stem cell therapy for myocardial infarction: a meta-analysis of preclinical studies and clinical trials. Circ Res. 2017;120(7):1139–1150. doi: 10.1161/CIRCRESAHA.116.309819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mohamadnejad M, Alimoghaddam K, Mohyeddin-Bonab M, Bagheri M, Bashtar M, Ghanaati H, et al. Phase 1 trial of autologous bone marrow mesenchymal stem cell transplantation in patients with decompensated liver cirrhosis. Arch Iran Med. 2007;10(4):459–466. [PubMed] [Google Scholar]
- 12.Le Blanc K, Rasmusson I, Sundberg B, Götherström C, Hassan M, Uzunel M, et al. Treatment of severe acute graft-versus-host disease with third party haploidentical mesenchymal stem cells. Lancet. 2004;363(9419):1439–1441. doi: 10.1016/S0140-6736(04)16104-7. [DOI] [PubMed] [Google Scholar]
- 13.Braza F, Dirou S, Forest V, Sauzeau V, Hassoun D, Chesné J, et al. Mesenchymal stem cells induce suppressive macrophages through phagocytosis in a mouse model of asthma. Stem Cells. 2016;34(7):1836–1845. doi: 10.1002/stem.2344. [DOI] [PubMed] [Google Scholar]
- 14.Mathias LJ, Khong SM, Spyroglou L, Payne NL, Siatskas C, Thorburn AN, et al. Alveolar macrophages are critical for the inhibition of allergic asthma by mesenchymal stromal cells. J Immunol. 2013;191(12):5914–5924. doi: 10.4049/jimmunol.1300667. [DOI] [PubMed] [Google Scholar]
- 15.Lathrop MJ, Brooks EM, Bonenfant NR, Sokocevic D, Borg ZD, Goodwin M, et al. Mesenchymal stromal cells mediate Aspergillus hyphal extract-induced allergic airway inflammation by inhibition of the Th17 signaling pathway. Stem Cells Transl Med. 2014;3(2):194–205. doi: 10.5966/sctm.2013-0061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Goodwin M, Sueblinvong V, Eisenhauer P, Ziats NP, LeClair L, Poynter ME, et al. Bone marrow-derived mesenchymal stromal cells inhibit Th2-mediated allergic airways inflammation in mice. Stem Cells. 2011;29(7):1137–1148. doi: 10.1002/stem.656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Firinci F, Karaman M, Baran Y, Bagriyanik A, Ayyildiz ZA, Kiray M, et al. Mesenchymal stem cells ameliorate the histopathological changes in a murine model of chronic asthma. Int Immunopharmacol. 2011;11(8):1120–1126. doi: 10.1016/j.intimp.2011.03.009. [DOI] [PubMed] [Google Scholar]
- 18.Gao P, Zhou Y, Xian L, Li C, Xu T, Plunkett B, et al. Functional effects of TGF-β1 on mesenchymal stem cell mobilization in cockroach allergen-induced asthma. J Immunol. 2014;192(10):4560–4570. doi: 10.4049/jimmunol.1303461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Nemeth K, Keane-Myers A, Brown JM, Metcalfe DD, Gorham JD, Bundoc VG, et al. Bone marrow stromal cells use TGF-beta to suppress allergic responses in a mouse model of ragweed-induced asthma. Proc Natl Acad Sci U S A. 2010;107(12):5652–5657. doi: 10.1073/pnas.0910720107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Cho KS, Park MK, Kang SA, Park HY, Hong SL, Park HK, et al. Adipose-derived stem cells ameliorate allergic airway inflammation by inducing regulatory T cells in a mouse model of asthma. Mediators Inflamm. 2014;2014:436476. doi: 10.1155/2014/436476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ogulur I, Gurhan G, Aksoy A, Duruksu G, Inci C, Filinte D, et al. Suppressive effect of compact bone-derived mesenchymal stem cells on chronic airway remodeling in murine model of asthma. Int Immunopharmacol. 2014;20(1):101–109. doi: 10.1016/j.intimp.2014.02.028. [DOI] [PubMed] [Google Scholar]
- 22.Bonfield TL, Koloze M, Lennon DP, Zuchowski B, Yang SE, Caplan AI. Human mesenchymal stem cells suppress chronic airway inflammation in the murine ovalbumin asthma model. Am J Physiol Lung Cell Mol Physiol. 2010;299(6):L760–70. doi: 10.1152/ajplung.00182.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hur J, Kang JY, Rhee CK, Kim YK, Lee SY. The leukotriene receptor antagonist pranlukast attenuates airway remodeling by suppressing TGF-β signaling. Pulm Pharmacol Ther. 2018;48:5–14. doi: 10.1016/j.pupt.2017.10.007. [DOI] [PubMed] [Google Scholar]
- 24.Yoon DS, Kim YH, Lee S, Lee KM, Park KH, Jang Y, et al. Interleukin-6 induces the lineage commitment of bone marrow-derived mesenchymal multipotent cells through down-regulation of Sox2 by osteogenic transcription factors. FASEB J. 2014;28(7):3273–3286. doi: 10.1096/fj.13-248567. [DOI] [PubMed] [Google Scholar]
- 25.Finkelman FD, Hogan SP, Hershey GK, Rothenberg ME, Wills-Karp M. Importance of cytokines in murine allergic airway disease and human asthma. J Immunol. 2010;184(4):1663–1674. doi: 10.4049/jimmunol.0902185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lee SH, Jang AS, Kwon JH, Park SK, Won JH, Park CS. Mesenchymal stem cell transfer suppresses airway remodeling in a toluene diisocyanate-induced murine asthma model. Allergy Asthma Immunol Res. 2011;3(3):205–211. doi: 10.4168/aair.2011.3.3.205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ou-Yang HF, Huang Y, Hu XB, Wu CG. Suppression of allergic airway inflammation in a mouse model of asthma by exogenous mesenchymal stem cells. Exp Biol Med (Maywood) 2011;236(12):1461–1467. doi: 10.1258/ebm.2011.011221. [DOI] [PubMed] [Google Scholar]
- 28.Abreu SC, Antunes MA, Xisto DG, Cruz FF, Branco VC, Bandeira E, et al. Bone marrow, adipose, and lung tissue-derived murine mesenchymal stromal cells release different mediators and differentially affect airway and lung parenchyma in experimental asthma. Stem Cells Transl Med. 2017;6(6):1557–1567. doi: 10.1002/sctm.16-0398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kavanagh H, Mahon BP. Allogeneic mesenchymal stem cells prevent allergic airway inflammation by inducing murine regulatory T cells. Allergy. 2011;66(4):523–531. doi: 10.1111/j.1398-9995.2010.02509.x. [DOI] [PubMed] [Google Scholar]
- 30.Sun YQ, Deng MX, He J, Zeng QX, Wen W, Wong DS, et al. Human pluripotent stem cell-derived mesenchymal stem cells prevent allergic airway inflammation in mice. Stem Cells. 2012;30(12):2692–2699. doi: 10.1002/stem.1241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Keating A. Mesenchymal stromal cells: new directions. Cell Stem Cell. 2012;10(6):709–716. doi: 10.1016/j.stem.2012.05.015. [DOI] [PubMed] [Google Scholar]
- 32.Le Blanc K, Mougiakakos D. Multipotent mesenchymal stromal cells and the innate immune system. Nat Rev Immunol. 2012;12(5):383–396. doi: 10.1038/nri3209. [DOI] [PubMed] [Google Scholar]
- 33.Prockop DJ, Oh JY. Medical therapies with adult stem/progenitor cells (MSCs): a backward journey from dramatic results in vivo to the cellular and molecular explanations. J Cell Biochem. 2012;113(5):1460–1469. doi: 10.1002/jcb.24046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Bernardo ME, Fibbe WE. Mesenchymal stromal cells: sensors and switchers of inflammation. Cell Stem Cell. 2013;13(4):392–402. doi: 10.1016/j.stem.2013.09.006. [DOI] [PubMed] [Google Scholar]
- 35.Bai L, Lennon DP, Eaton V, Maier K, Caplan AI, Miller SD, et al. Human bone marrow-derived mesenchymal stem cells induce Th2-polarized immune response and promote endogenous repair in animal models of multiple sclerosis. Glia. 2009;57(11):1192–1203. doi: 10.1002/glia.20841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Aggarwal S, Pittenger MF. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood. 2005;105(4):1815–1822. doi: 10.1182/blood-2004-04-1559. [DOI] [PubMed] [Google Scholar]
- 37.Kang WC, Oh PC, Lee K, Ahn T, Byun K. Increasing injection frequency enhances the survival of injected bone marrow derived mesenchymal stem cells in a critical limb ischemia animal model. Korean J Physiol Pharmacol. 2016;20(6):657–667. doi: 10.4196/kjpp.2016.20.6.657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Swedin L, Neimert-Andersson T, Hjoberg J, Jonasson S, van Hage M, Adner M, et al. Dissociation of airway inflammation and hyperresponsiveness by cyclooxygenase inhibition in allergen challenged mice. Eur Respir J. 2009;34(1):200–208. doi: 10.1183/09031936.00030908. [DOI] [PubMed] [Google Scholar]
- 39.Wilder JA, Collie DD, Wilson BS, Bice DE, Lyons CR, Lipscomb MF. Dissociation of airway hyperresponsiveness from immunoglobulin E and airway eosinophilia in a murine model of allergic asthma. Am J Respir Cell Mol Biol. 1999;20(6):1326–1334. doi: 10.1165/ajrcmb.20.6.3561. [DOI] [PubMed] [Google Scholar]
- 40.Sharkhuu T, Matthaei KI, Forbes E, Mahalingam S, Hogan SP, Hansbro PM, et al. Mechanism of interleukin-25 (IL-17E)-induced pulmonary inflammation and airways hyper-reactivity. Clin Exp Allergy. 2006;36(12):1575–1583. doi: 10.1111/j.1365-2222.2006.02595.x. [DOI] [PubMed] [Google Scholar]
- 41.Kondo Y, Yoshimoto T, Yasuda K, Futatsugi-Yumikura S, Morimoto M, Hayashi N, et al. Administration of IL-33 induces airway hyperresponsiveness and goblet cell hyperplasia in the lungs in the absence of adaptive immune system. Int Immunol. 2008;20(6):791–800. doi: 10.1093/intimm/dxn037. [DOI] [PubMed] [Google Scholar]
- 42.Zhou B, Headley MB, Aye T, Tocker J, Comeau MR, Ziegler SF. Reversal of thymic stromal lymphopoietin-induced airway inflammation through inhibition of Th2 responses. J Immunol. 2008;181(9):6557–6562. doi: 10.4049/jimmunol.181.9.6557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ballantyne SJ, Barlow JL, Jolin HE, Nath P, Williams AS, Chung KF, et al. Blocking IL-25 prevents airway hyperresponsiveness in allergic asthma. J Allergy Clin Immunol. 2007;120(6):1324–1331. doi: 10.1016/j.jaci.2007.07.051. [DOI] [PubMed] [Google Scholar]
- 44.Onari Y, Yokoyama A, Haruta Y, Nakashima T, Iwamoto H, Hattori N, et al. IL-12p40 is essential for the down-regulation of airway hyperresponsiveness in a mouse model of bronchial asthma with prolonged antigen exposure. Clin Exp Allergy. 2009;39(2):290–298. doi: 10.1111/j.1365-2222.2008.03131.x. [DOI] [PubMed] [Google Scholar]
- 45.Schrepfer S, Deuse T, Reichenspurner H, Fischbein MP, Robbins RC, Pelletier MP. Stem cell transplantation: the lung barrier. Transplant Proc. 2007;39(2):573–576. doi: 10.1016/j.transproceed.2006.12.019. [DOI] [PubMed] [Google Scholar]
- 46.Kang WJ, Kang HJ, Kim HS, Chung JK, Lee MC, Lee DS. Tissue distribution of 18F-FDG-labeled peripheral hematopoietic stem cells after intracoronary administration in patients with myocardial infarction. J Nucl Med. 2006;47(8):1295–1301. [PubMed] [Google Scholar]
- 47.Ge J, Guo L, Wang S, Zhang Y, Cai T, Zhao RC, et al. The size of mesenchymal stem cells is a significant cause of vascular obstructions and stroke. Stem Cell Rev Rep. 2014;10(2):295–303. doi: 10.1007/s12015-013-9492-x. [DOI] [PubMed] [Google Scholar]
- 48.Bartosh TJ, Ylöstalo JH, Mohammadipoor A, Bazhanov N, Coble K, Claypool K, et al. Aggregation of human mesenchymal stromal cells (MSCs) into 3D spheroids enhances their antiinflammatory properties. Proc Natl Acad Sci U S A. 2010;107(31):13724–13729. doi: 10.1073/pnas.1008117107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Häfeli UO, Saatchi K, Elischer P, Misri R, Bokharaei M, Labiris NR, et al. Lung perfusion imaging with monosized biodegradable microspheres. Biomacromolecules. 2010;11(3):561–567. doi: 10.1021/bm9010722. [DOI] [PubMed] [Google Scholar]
- 50.Anjos-Afonso F, Siapati EK, Bonnet D. In vivo contribution of murine mesenchymal stem cells into multiple cell-types under minimal damage conditions. J Cell Sci. 2004;117(Pt 23):5655–5664. doi: 10.1242/jcs.01488. [DOI] [PubMed] [Google Scholar]
- 51.Lee RH, Seo MJ, Pulin AA, Gregory CA, Ylostalo J, Prockop DJ. The CD34-like protein PODXL and alpha6-integrin (CD49f) identify early progenitor MSCs with increased clonogenicity and migration to infarcted heart in mice. Blood. 2009;113(4):816–826. doi: 10.1182/blood-2007-12-128702. [DOI] [PMC free article] [PubMed] [Google Scholar]