

Abstract
Endometrial cancer is one of the most prevalent tumors of the female reproductive system causing serious health effects to women worldwide. Although numerous studies, including analysis of gene expression profile and cellular microenvironment have been reported in this field, pathogenesis of this disease remains unclear. In this study, we performed a system bioinformatics analysis of endometrial cancer using the Gene Expression Omnibus (GEO) datasets (GSE17025, GSE63678, and GSE115810) to identify the core genes. In addition, exosomes derived from endometrial cancer cells were also isolated and identified. First, we analyzed the differentially expressed genes (DEGs) between endometrial cancer tissues and normal tissues in clinic samples. We found that HAND2-AS1, PEG3, OGN, SFRP4, and OSR2 were co-expressed across all 3 datasets. Pathways analysis showed that several pathways associated with endometrial cancer, including “p53 signaling pathway”, “Glutathione metabolism”, “Cell cycle”, and etc. Next, we selected DEGs with highly significant fold change and co-expressed across the 3 datasets and validated them in the TCGA database using Gene Expression Profiling Interactive Analysis (GEPIA). Finally, we performed a survival analysis and identified four genes (TOP2A, ASPM, EFEMP1, and FOXL2) that play key roles in endometrial cancer. We found up-regulation of TOP2A and ASPM in endometrial cancer tissues or cells, while EFEMP1 and FOXL2 were down-regulated. Furthermore, we isolated exosomes from the culturing supernatants of endometrial cancer cells (Ishikawa and HEC-1-A) and found that miR-133a, which regulates expression of FOXL2, were present in exosomes and that they could be delivered to normal endometrial cells. The common DEGs, pathways, and exosomal miRNAs identified in this study might play an important role in progression as well as diagnosis of endometrial cancer. In conclusion, our results provide insights into the pathogenesis and risk assessment of endometrial cancer. Even so, further studies are required to elucidate on the precise mechanism of action of these genes in endometrial cancer.
Subject terms: Cancer, Cell biology, Biomarkers
Introduction
Endometrial cancer, a gynecologic malignancy tumor ranked behind breast, lung, and colorectal cancers1, greatly affects women’s health around the world. In the recent past, incidence and the rate of this disease has increased due to lack of effective diagnosis before menopause2. Meanwhile, patient age at disease onset has also decreased due to changes in lifestyles3. In general, endometrial cancer is classified into two types based on pathological and histological characteristics2. Type I commonly known as endometrioid adenocarcinoma is the major type of endometrial cancer4. Type II is the second category of endometrial cancer, and have a higher prevalence and poor prognosis5. Early diagnosis is required for effective disease assessment and treatment, but numerous studies are now focusing on specific molecular alterations that aid in the development of endometrial cancers6,7.
Initiation and progression of tumors is associated with abnormal expression of genes and the cancer microenvironment8. Numerous processes and factors have been shown to medicate cell-cell and cell-microenvironment communication. Most importantly, tumor cells secrete molecules to the extracellular microenvironment, and these contribute to cancer progression. In addition, these molecules are regarded as biomarkers for diagnosis of diseases. Recent studies have demonstrated that exosomes, with the size range from 40 to 200 nm, which contain cargos including proteins, lipids, and nucleic acids9, transfer signaling molecules to cells via the extracellular environment. This is a novel mode of cellular communication that could influence the physiological conditions and behaviors of the recipient cells, and also connect with a variety of diseases process10–12. miRNAs, a group of small non-coding RNAs that regulate expression of endogenous genes by targeting to 3’ untranslated region (3' UTR)13, are mainly present in exosomes and play essential roles in occurrence of multiple tumor types14,15. Exosomal miRNAs mediate regulation of cancer milieu, containing cancer metastasis and progression16. In addition, exosomes also serve as a source of diagnostic and prognostic biomarkers for endometrial cancer12,17. Integrative research on endometrial cancer is still lacking with most studies only focusing on gene expression or exosomal cargos of the disease18–20.
Gene chips analysis is a useful tool for identifying the factors and genes involved in endometrial cancer21. Previous studies, particularly those focusing on screening differentially expressed genes, have reported identification of several targets and elucidated the roles played by specific biological process and pathways in endometrial cancer22,23. In this study, an integrative bioinformatics analysis was performed on endometrial cancer and normal endometrial tissues. Using the Gene Expression Omnibus (GEO) database, several key differentially expressed genes (DEGs) were identified, and their biological processes and pathways found to be closely associated with endometrial cancer. In addition, we found that endometrial cancer cells-derived exosomal miRNA targets the core DEG. These exosomes also could be delivered to normal cells.
Results
Identification of DEGs in endometrial cancer
We used basic packages implemented in R to identify potential DEGs across the three data sets. Based on a cut-off criteria of adjust P-value <0.05 and |logFC | >1.0, we identified a total of 2346, 340, and 233 genes in GSE17025, GSE63678 and GSE115810, respectively. The top 10 DEGs with the largest fold changes in the three GEO data sets are shown in Tables 1–3, while the top 100 DEGs in each dataset, represented by heat maps are shown in Fig. 1b–d. After expurgating duplicate genes and expression values which lacked specific gene symbols, we combined the top 100 DEGs in GSE17025, GSE63678 and GSE115810 to create a Venn diagram. Five genes were found to be co-expressed across all three data sets, including HAND2-AS1, PEG3, OGN, SFRP4, and OSR2. All these genes were, however, down-regulated in endometrial cancer tissues (Fig. 2). Three genes, including FOSB, FOS and LTBP4, were co-expressed in GSE17025 and GSE115810. In addition, a total of 10 genes, including CFB, RBM47, WT1, ADAMTS5, TSPYL5, HTR2B, ANK2, FOXL2, CXCL12 and EFEMP1, were co-expressed in GSE63678 and GSE115810. On the other hand, 13 genes, including BCHE, LMOD1, TPX2, CXCL9, LIPG, TFAP2A, DLGAP5, MMP12, CEP55, RRM2, MELK, TOP2A and ASPM were co-expressed in GSE17025 and GSE63678,.
Table 2.
Top 10 differentially expressed genes in GSE63678.
| Rank | Probe set ID | Gene symbol | Description | Regulation |
|---|---|---|---|---|
| 1 | 209242_at | PEG3 | paternally expressed 3 | Down-regulation |
| 2 | 219791_s_at | HAND2-AS1 | HAND2 antisense RNA 1 | Down-regulation |
| 3 | 206067_s_at | WT1 | WT1 transcription factor | Down-regulation |
| 4 | 205433_at | BCHE | butyrylcholinesterase | Down-regulation |
| 5 | 218730_s_at | OGN | osteoglycin | Down-regulation |
| 6 | 204653_at | TFAP2A | transcription factor AP-2 alpha | Up-regulation |
| 7 | 201839_s_at | EPCAM | epithelial cell adhesion molecule | Up-regulation |
| 8 | 201596_x_at | KRT18 | keratin 18 | Up-regulation |
| 9 | 219148_at | PBK | PDZ binding kinase | Up-regulation |
| 10 | 204580_at | MMP12 | matrix metallopeptidase 12 | Up-regulation |
Table 1.
Top 10 differentially expressed genes in GSE17025.
| Rank | Probe set ID | Gene symbol | Description | Regulation |
|---|---|---|---|---|
| 1 | 202768_at | FOSB | FBJ osteosarcoma oncogene B | Down-regulation |
| 2 | 206622_at | TRH | thyrotropin releasing hormone | Down-regulation |
| 3 | 219791_s_at | HAND2-AS1 | HAND2 antisense RNA 1 | Down-regulation |
| 4 | 206228_at | PAX2 | paired box 2 | Down-regulation |
| 5 | 209242_at | PEG3 | paternally expressed 3 | Down-regulation |
| 6 | 201291_s_at | TOP2A | DNA topoisomerase II alpha | Up-regulation |
| 7 | 1553655_at | CDC20B | cell division cycle 20B | Up-regulation |
| 8 | 219918_s_at | ASPM | abnormal spindle microtubule assembly | Up-regulation |
| 9 | 202018_s_at | LTF | lactotransferrin | Up-regulation |
| 10 | 205476_at | CCL20 | C-C motif chemokine ligand 20 | Up-regulation |
Table 3.
Top 10 differentially expressed genes in GSE115810.
| Rank | Probe set ID | Gene symbol | Description | Regulation |
|---|---|---|---|---|
| 1 | 204051_s_at | SFRP4 | secreted frizzled related protein 4 | Down-regulation |
| 2 | 219791_s_at | HAND2-AS1 | HAND2 antisense RNA 1 | Down-regulation |
| 3 | 220102_at | FOXL2 | forkhead box L2 | Down-regulation |
| 4 | 201842_s_at | EFEMP1 | EGF containing fibulin extracellular matrix protein 1 | Down-regulation |
| 5 | 206637_at | P2RY14 | purinergic receptor P2Y14 | Down-regulation |
| 6 | 201195_s_at | SLC7A5 | solute carrier family 7 member 5 | Up-regulation |
| 7 | 202357_s_at | CFB | complement factor B | Up-regulation |
| 8 | 209875_s_at | SPP1 | secreted phosphoprotein 1 | Up-regulation |
| 9 | 204846_at | CP | ceruloplasmin | Up-regulation |
| 10 | 204623_at | TFF3 | trefoil factor 3 | Up-regulation |
Figure 1.

(a) Flow diagram of the study design. GO, gene ontology. KEGG, Kyoto Encyclopedia of Genes and Genomes. (b–d) Heat maps showed the DEGs between normal endometrial tissues and endometrial cancer tissue in three GEO datasets. Heat map for potential DEGs in GSE17025 (containing 12 normal endometrial tissues and 79 tumor tissues) (b) GSE63678 (containing 5 normal endometrial tissues and 7 tumor tissues) (c) GSE115810 (containing 3 normal endometrial tissues and 24 tumor tissues) (d).
Figure 2.
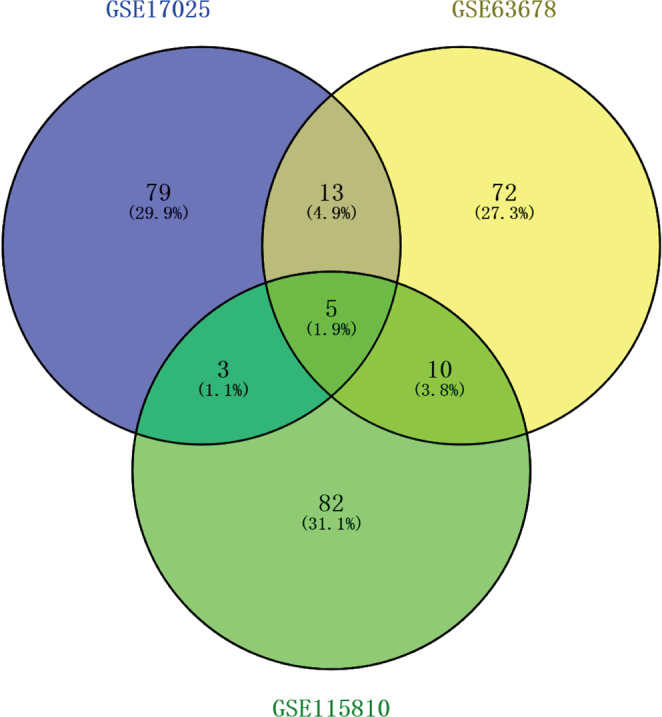
The Venn diagram shows the top 100 DEGs and co-expressed genes among GSE17025, GSE63678 and GSE115810.
Biological process analysis of DEGs
To gain more insights into the function of the DEGs, we performed a GO analysis. Sorting the transcripts by false discovery rate (FDR) revealed that there are 15, 20 and 10 GO terms of biological processes in GSE17025, GSE63678 and GSE115810, respectively (Fig. 3a–c). There was no common biological processes were found across the 3 datasets. Nevertheless, we found 6 common biological processes including “ cell division “, “mitotic nuclear division”, “ cell proliferation “, “ sister chromatid cohesion “, “ chromosome segregation “, and “mitotic sister chromatid segregation” in GSE63678 and GSE17025. Other biological processes, such as “epithelial cell differentiation”, “apoptotic process “, “extracellular matrix organization”, “ cell development “, “blood vessel development” and “ extracellular structure organization “, were also identified very vital in each dataset.
Figure 3.

The significant biological functions in terms of GO analysis related to DEGs in GSE17025 (a), GSE63678 (b) and GSE115810 (c).
Signaling pathway enrichment analysis
The potential pathways regulated by the DEGs in endometrial cancer were explored using KEGG analysis, We found that specific pathways were enriched in each dataset (Fig. 4a–c). Bubble plots from this analysis presented the most significantly enriched pathways (Fig. 4a–c). Among them, “Pathways in cancer” was the common pathway in the three datasets. The top 10 pathways are as outlined in Table 4. Of these, 7 pathways, including those involved in p53 signaling pathway, Glutathione metabolism, Cell cycle, ECM-receptor interaction, PI3K-Akt signaling pathway, Focal adhesion and MAPK signaling were notable because of their association with cancer progression. Particularly, these pathways were implicated in processes such as cell proliferation, cell migration and invasion. Pathways involved in HIF-1 signaling pathway, protein digestion and absorption as well as FoxO signaling pathway were also other interesting pathways following pairwise comparisons.
Figure 4.

Pathways in terms of KEGG pathway analysis related to DEGs in GSE17025 (a), GSE63678 (b) and GSE115810 (c).
Table 4.
| Rank | GSE17025 | GSE63678 | GSE115810 |
|---|---|---|---|
| 1 | Cell cycle | Cell cycle | ECM-receptor interaction |
| 2 | Leukocyte transendothelial migration | p53 signaling pathway | PI3K-Akt signaling pathway |
| 3 | PPAR signaling pathway | Oocyte meiosis | Focal adhesion |
| 4 | Pathways in cancer | Proteoglycans in cancer | Pathways in cancer |
| 5 | Glutathione metabolism | Progesterone-mediated oocyte maturation | Melanoma |
| 6 | p53 signaling pathway | Melanoma | Protein digestion and absorption |
| 7 | ECM-receptor interaction | Mineral absorption | Cell adhesion molecules (CAMs) |
| 8 | Bile secretion | Biosynthesis of antibiotics | MAPK signaling pathway |
| 9 | Progesterone-mediated oocyte maturation | Prostate cancer | Amoebiasis |
| 10 | Chemokine signaling pathway | Pathways in cancer | Complement and coagulation cascades |
Validation of DEGs in TCGA database
To further validate and determine the core genes highly associate with endometrial cancer from these DEGs, eight down-regulated and five up-regulated genes that showed the most significant fold changes (top10) between endometrial cancer and normal tissue in GSE17025, GSE63678 and GSE115810, were selected. Meanwhile, these 13 genes are derived from DEGs that overlap across the three data sets (Fig. 2). These included HAND2-AS1, PEG3, OGN, SFRP4, TFAP2A, MMP12, TOP2A, ASPM, FOSB, CFB, WT1, FOXL2 and EFEMP1. Hypothesis test analysis proves that these 13 genes have statistical significance (data not shown). Next, we explored the Gene Expression Profiling Interactive Analysis (GEPIA) database for identification and retrieval of the expression profiles of these DEGs in both endometrial cancer and normal tissues. We did not find any significant differences in FOSB expression in the GEPIA database between the treatments. However, expression profiles of the remaining twelve DEGs were consistent with our analysis results in GEO database. Particularly, expression of TFAP2A, MMP12, TOP2A, ASPM and CFB were higher in endometrial cancer relative to normal tissues. On the other hand, HAND2-AS1, PEG3, OGN, SFRP4, WT1, FOXL2 and EFEMP1 were significantly lower in endometrial cancer tissues than in normal tissues (Fig. 5).
Figure 5.

The GEPIA was used to verify the DEGs between normal endometrial tissues and endometrial cancer tissues in the TCGA database.
Survival analysis of core genes
We then used prognostic information of the 12 core genes, available in the free online TIMER database (Tumor IMmune Estimation Resource) (https://cistrome.shinyapps.io/timer/), to perform a survival analysis. Aberrant expression levels of four genes out of 12 core DEGs showed a significant (P < 0.05)association with poor prognosis of endometrial cancer, and this included two up-regulated (TOP2A and ASPM) and two down-regulated (FOXL2 and EFEMP1) genes (Fig. 6a–d). This indicated that the four genes might play important roles in the progression of endometrial cancer.
Figure 6.

Survival analysis of the core genes (a) TOP2A, (b) ASPM, (c) FOXL2, (d) EFEMP1.
Validation of core genes expression in endometrial cancer cells
Given the lack of clinical samples, we only used three cell lines in this study to validate the results of endometrial cancer from bioinformatics analyses. Ishikawa and HEC-1-A cell lines were derived from endometrioid adenocarcinoma, while 4003 was the normal endometrial cells and was used as a control. Real-time qPCR assay was employed to quantify expression levels of 4(TOP2A, ASPM, EFEMP1, and FOXL2) core DEGs across the three types of cells. Results showed a significant down-regulation of EFEMP1 and FOXL2 in endometrial cancer cells (Fig. 7c,d). A comparison with normal cells showed that TOP2A was highly expressed in endometrial cancer cells (Fig. 7a), which was consistent with the resultsfrom bioinformatics analysis above. However, we noted that ASPM was only significantly expressed in HEC-1-A cells (Fig. 7b).
Figure 7.

The validation of four core DEGs in endometrial cell lines. (a) TOP2A. (b) ASPM, (c) FOXL2, (d) EFEMP1.
Exosomes secretion by endometrial cancer cells
Cancer progression is always related to the microenvironment. Recently, exosomes have been found to modulate cancer cell communication as a novel way and almost all types of cancer cells can release exosomes into their microenvironment10. In this study, we hypothesized that endometrial cancer cells could secrete exosomes and pass them on to recipient cells, thereby promoting cancer development. Consequently, we isolated exosomes from the culture supernatants of Ishikawa and HEC-1-A cells (Fig. 8a). The concentration and particle distribution of exosomes were first characterized using NTA analysis, and this revealed average sizes of 127.2 nm and 146.9 nm for Ishikawa and HEC-1-A, respectively (Fig. 8b). Results from Western blotting analysis showed presence of CD63, an exosome marker, in tumor cell-derived exosomes. Conversely, no endoplasmic reticulum protein CANX was detected in the exosomes (Fig. 8c). TEM results showed that the vesicles from endometrial cancer cells were either round or oval, with a complete lipid bilayer membrane structure (Fig. 8d,e). Previous studies have demonstrated presence of miRNAs in exosomes, which are the main source of circulating miRNAs24, and play a vital role in cancer development25. In this study, we observed that FOXL2 was down-regulated in endometrial cancer tissues and was significantly associated with endometrial cancer. Previous studies have shown that FOXL2 is regulated by miR-133a26 (Fig. 8f). Based on this, we detected the expression of miR-133a in all three cell lines. Compared with normal endometrial cancer cells, we found that miR-133a was highly expressed in endometrial cancer cells (Fig. 8g). At the same time, we also detected miR-133a expression in Ishikawa and HEC-1-A-derived exosomes (Fig. 8h). In addition, exosomes derived from endometrial cancer cells could be taken up by normal endometrial cells (Figure S), indicating that endometrial cancer cells release exosomes, which might be then transported to normal endometrial cells.
Figure 8.

Endometrial cancer cells secret exosomal miRNAs. (a) Procedure for isolating exosomes from Ishikawa and HEC-1-A cell culture supernatant using ultracentrifugation. (b) NTA analysis of exosomes size distribution and concentration. (c) Western blot analysis of exosome protein marker (CD63). CANX is a negative control. (d-e) The structure of exosomes were observed using TEM. (f) The relationship between miR-133a and FOXL2. (g) The expression of miR-133a in endometrial cancer cells. (h) The expression of miR-133a in endometrial cancer cell-derived exosomes.
Discussion
Endometrial cancer occurs due to abnormal expression of many cancer-related genes27, many of which are associated with the susceptibility and development of endometrial cancer22,23. Numerous molecular approaches have been performed to identify biomarkers for endometrial cancer28. Nonetheless, most of them only focused on a single genetic aspect, limiting the reliability of the biomarkers29. In the present study, we used three GEO datasets to perform an integrative analysis with a view of identifying DEGs between normal and cancerous endometrial tissues. The results were validated using the TCGA database. Key biological processes and pathways based on the DEGs between endometrial cancer and normal tissues were also identified. Particularly, we focused on the common DEGs across the three GEO datasets as well as those that were significantly expressed in each dataset. Moreover, we identified exosomes derived from endometrial cancer cells.
Activation of cell proliferation process and p53 signaling pathways have been previously associated with cancers development30,31. In this study, GO and KEGG analysis showed that DEGs are involved in progression of endometrial cancer through these biological processes and pathways. A total of 12 DEGs, includingHAND2-AS1, PEG3, OGN, SFRP4, TFAP2A, MMP12, TOP2A, ASPM, CFB, WT1, FOXL2, and EFEMP1, were identified as candidate genes for endometrial cancer. Furthermore, a cross comparison of common DEGs across the three datasets revealed that TOP2A, ASPM, FOXL2 and EFEMP1 were potential biomarkers for distinguishing endometrial cancer, and this was significantly associated with the survival ratios among patients. Further validation in endometrial cancer cells showed that indeed TOP2A, ASPM, FOXL2 and EFEMP1 were significantly regulated. Particularly, TOP2A was significantly up-regulated in endometrial cancer tissues and cells, consistent with Lapinska-Szumczyk et al.32. These results further indicated that high levels of TOP2A can lead to EC progression. Previous studies have reported that aberrant expression of TOP2A is related to the stage of endometrial cancer and its progression33. Functionally, TOP2A encodes a DNA topoisomerase that controls and alters the topologic states of DNA during transcription34, and is involved in processes such as chromosome condensation, DNA transcription and replication35. High levels of TOP2A have also been detected in other types of cancers, including ovarian cancer36 and prostate cancer37. Based on these results, therefore, we hypothesized that abnormal expression of TOP2A could be a positive tumor metastasis marker and a poor biomarker for prognosis. In addition, inhibition of TOP2A has been found to mediate a decrease in migration, proliferation or invasion of cells38. These results might lead to development of a new treatment strategy for endometrial cancer. Abnormal spindle-like microcephaly-associated (ASPM), which encodes the ASPM protein, plays an essential role in mitotic spindle function during cell replication39. ASPM is associated with cell cycle and mediates the tightly coordinated Ubiquitin-Cyclin E-Retinoblastoma-E2F bistable-signaling pathway to control restriction point progression and stem cell maintenance40. Previous studies have demonstrated that overexpression of the ASPM gene is associated with progression and poor outcomes in various types of cancer, such as bladder cancer41, prostate cancer42, and breast cancer43. However, none of these studies have elucidated the relationship between ASPM and endometrial cancer. In this study, our results first showed that high levels of ASPM were related to endometrial cancer. When combined with the function of the gene, ASPM is expected to become a new molecule for detection and treatment of this disease although its mechanism in endometrial cancer is yet to be studied.
In addition to the above findings on DEGs, EFEMP1 and FOXL2 were found to be down-regulated in endometrial cancer tissues. The epidermal growth factor-containing fibulin-like extracellular matrix protein 1(EFEMP1), functions as an oncogene or a suppressor, depending on the types of tumor. With previous studies showing that EFEMP1 is a new candidate tumor suppressor gene in endometrial carcinoma44. Our findings showed that EFEMP1 was down-regulated in endometrial cancer tissues, as well as endometrial cancer cell lines. Functionally, EFEMP1 inhibits migration of tumor cells by regulating MMP2 and MMP9 via ERK1/2 activity45,46. The findings on EFEMP1 in endometrial cancer revealed that the gene could inhibit tumor growth and invasion both in vitro and in vivo consistent with what has been reported in other types of tumors44. A separate studies found that EFEMP1 was elevated in highly invasive ovarian cancer, compared with the low invasive types47. Combined, these studies suggest that EFEMP1 plays different roles in the development of different types of carcinoma. FOXL2 on its part encodes a fork head transcription factor. Although many RNA-sequencing studies have been performed to understand the genetic regulation of endometrial cancer, the relationship between FOXL2 and the disease is still lacking48. In the current study, analysis on GSE115810 showed that FOXL2 was one of the top10 DEGs found to be down-regulated during the development of endometrial cancer. On the other hand, we observed a significantly low FOXL2 expression in endometrial cancer relative to normal cells. A related study showed that that FOXL2 could suppress cells proliferation and enhance cells apoptosis in cervical squamous cancer, mainly by decreasing Ki67 expression49. Various conclusions have been drawn regarding the mechanism of action of FOXL2 in other types of tumors50. However, its function remains unclear in endometrial cancer thereby necessitating the need for further clarification using molecular experiments.
A vast array of researches have demonstrated that almost all types of cells can release exosomes that mediate the cell-cell communication51, especially tumor cells52. Exosomal cargos, particularly those associated with development of cancer, have been considered the best biomarkers for non-invasive diagnostic53. Furthermore, studies have revealed that miRNAs are mainly packaged in exosomes. In our study, results from the GO analysis showed that the DEGs were associated with extracellular exosomes. Here, we assumed that the progression of endometrial cancer was mediated by exosomal miRNAs derived from cancer cells. Meanwhile, we observed a down-regulation of FOXL2 in endometrial cancer cells while the expression of miR-133a, which targets FOXL254, was high in the cancerous cells. We found that endometrial cancer cells could secrete exosomes, which contained miR-133a. Exosomal miR-133a may regulate the down-regulation of FOXL2 in endometrial cancer tissues. Furthermore, the progression of endometrial cancer was not only achieved by malignant epithelial cells, but also through involvement of stromal cells. Previous research evidence indicated that extracellular vesicles mediate he communication between tumors and stroma55. In the current study, it was found that exosomes derived from endometrial cancer cells could be taken up by stromal cells, indicating that exosomal miRNAs may contribute to the progression of endometrial cancer.
In summary, four core genes (TOP2A, ASPM, EFEMP1 and FOXL2) and several interesting pathways involved in endometrial cancer were identified. Analysis of qRT-PCR assay revealed that expression of mRNA for TOP2A and ASPM are up-regulated in endometrial cancer cells, whereas those of EFEMP1 and FOXL2 are down-regulated. Currently, there are no studies on the role of ASPM and FOXL2 in endometrial cancer. Based on the findings of this study, we propose that multiple molecules can be effectively used to diagnose endometrial cancer. Accordingly, further studies into the functions of these genes can guide development of treatment strategies for EC. In addition, exosomes secreted by endometrial cancer cells, contain miRNAs that regulate FOXL2 and can be transported to normal stromal cells. These results open up new frontiers for the development of approaches for detection and treatment of endometrial cancer. However, further studies are required to elucidate the function and underlying mechanisms of action of these potential biomarkers in the progression of endometrial cancer.
Materials and Methods
Study design
The aim of this study was to identify core genes involved in the development of endometrial cancer, using bioinformatics analysis on GEO datasets. A flow diagram of the study design is shown in Fig. 1a. Initially, three GEO datasets (GSE17025, GSE63678 and GSE115810) were reasonably searched in published databases, and then bioinformatics tools used to screen DEGs in the datasets. Analyses on the functions of GO and KEGG pathways were also performed on the DEGs. Next, Genes co-expressed in the three datasets were identified and verified in TCGA database. Survival analyses were performed to further identify core genes related to endometrial cancer. Finally, significant co-expressed genes were verified using endometrial cell lines and miRNAs in exosomes from supernatants of endometrial cancer cell culture evaluated.
Gene expression omnibus datasets
We retrieved endometrial cancer gene expression array datasets by the GO (https://www.ncbi.nlm.nih. gov/gds) which is a public repository at the National Center of Biotechnology Information for storing high-throughput gene expression datasets56. The terms “endometrial cancer” and “Homo sapiens” and “gene expression array” were used to search the potential datasets. Finally, three datasets (GSE17025, GSE63678 and GSE115810) were found suit for our study. Those three datasets contain endometrioid adenocarcinoma tissues and normal endometrial tissues, and the three mRNA chips were sequenced from the same chip company. The detail information of datasets as follows. GSE17025 contained 12 normal endometrial tissues and 79 endometrioid cancer samples22. Platform information: GPL570 [HG-U133_Plus_2] Affymetrix Human Genome U133 Plus 2.0 Array. GSE63678 contained 5 normal endometrial tissues and 7 tumor tissues57. Platform information: GPL571 [HG-U133A_2] Affymetrix Human Genome U133A 2.0 Array. GSE115810 contained 3 normal endometrial tissues and 24 tumor tissues58. Platform information: GPL96 [HG-U133A] Affymetrix Human Genome U133A Array. Those three datasets were used for further analysis.
Identification of differentially expressed genes
Expression matrixes and platform information were downloaded from GEO database. Then, the datasets were normalized before analysis. R basic (version 3.6.0, https://www.rproject.org/) and Bioconductor packages (available online: http://www.bioconductor.org/) were applied to screen the DEGs59. The analysis was performed using limma package at cut off values |log FC | > 1.0 and Adjust P value value <0.05 in these three datasets. Based on the DEGs in each GEO dataset, gene ontology (GO) was used to analyze in terms of biological functions. Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis was employed for pathway analysis. The top 10 analysis results were shown. The GO and KEGG analysis were performed by an online tool DAVID (https://david.ncifcrf.gov/home.jsp)60. We used the latest updated data from this site. FDR < 0.05 was considered to indicate a statistically significant difference. VENNY2.1 (https://bioinfogp.cnb.csic.es/tools/venny/index.html) was used to compare the DEGs in three datasets.
The gene expression in TCGA database and survival analysis
In order to further validate the expression of co-expressed DEGs, the GEPIA (Gene Expression Profiling Interactive Analysis)61 database (http://gepia.cancer-pku.cn/) were employed, which based on the TCGA database. Furthermore, we analyzed the correlation between gene expression and survival conditions using TIMER (Tumor IMmune Estimation Resource) (https://cistrome.shinyapps.io/timer/)62.
Cell culture
Ishikawa, HEC-1-A and 4003 cell lines were purchased from cell bank of Chinese Academy of Science (Shanghai, China). Ishikawa and 4003 cells lines were cultured in DMEM (Gibco) culture medium supplemented with 10% FBS. while HEC-1-A cell lines were cultured in McCOY’s 5 A (SIGMA) supplemented with 10% FBS. The cultures were maintained n at 37 °C, under 5% CO2. The two types of endometrial cancer cell lines were derived from adenocarcinoma63,64.
Isolation and identification of exosomes from endometrial cancer cells culture medium
To purify exosomes from tumor cell culture medium, cells were cultured in medium supplemented with 10% exo-free FBS and cell culture supernatants were collected 24–48 h after culture. A standard differential centrifugation protocol was used to isolate exosomes. Briefly, cell cultures were centrifuged at 500 g for 10 min to remove dead cells, followed by a second centrifugation at 2,000 g for 20 min to remove cell debris. Micro and macro vesicles were depleted by centrifugation at 10,000 g for 45 min. Collected supernatants were then centrifuged at 120,000 g for 1.5 h at 4 °C. The pelleted exosomes were resuspended in PBS and washed again, and re-ultracentrifuged at 120,000 g for 1.5 h to collect exosomes.
To study the characteristics of exosomes, transmission electron microscopy was performed. Summarily, exosomes were suspended in PBS and placed onto formvar carbon-coated coper grids at room temperature for 1 min. The exosomes were then stained by 2% uranyl acetate at room temperature for 1 min. and the grids were dried. The stained exosomes were visualized under a TecnaiG2 Spirit120KV transmission electron microscope operating at 120 kV (Thermo FEI).
Particel sizes distribution and concentration of exosomes were examined by Nanoparticle Tracking Analysis (NTA) analysis using a ZetaView PMX 100 (Particle Metrix, Meerbusch, Germany). For each condition analysis, at least three independent experiments were performed.
A western blot analysis was performed to detect exosomal surface protein markers. The exosomal proteins were separated using 10% SDS PAGE, and then transferred onto the polyvinylidene difluoride membranes (Millipore). The membranes were blocked with QuickBlockTM western buffer (Beyotime, China) for 20 min, and incubated with primary antibody(diluted with manufacturer’s instructions) at 4 °C for overnight, followed by a 2 h incubation with HRP-conjugated secondary antibodies at room temperature. The membranes were detected with ECL reagents (Beyotime, China). CD63 was exosome protein marker. CANX, an endoplasmic reticulum protein, was used as the negative control.
Exosomes tracking analysis
Here, 50 μg of exosomes from supernatants of cultured endometrial cancer cells were incubated for 15 min at room temperature with 20 μM DiI dye containing 5% BSA, added to avert over staining. Exosomes were diluted with PBS and then centrifuged at 120,000 g for 1.5 h at 4 °C. The labeled exosomes were suspended in PBS and collected by ultracentrifugation at 120,000 g for 1.5 h. Next, the labeled exosomes were added to endometrial stroma cell culture medium and incubated for 12 h. The cells were fixed with 4% paraformaldehyde for 20 min, permeabilized with 0.1% Triton X-100 for 15 min, then incubated for 50 min with a FITC-conjugated phalloidin (Solarbio, China) to stain F-actin. DAPI was used to label cell nuclei (Solarbio, China). The respective images were visualized by a confocal microscopy and captured using ZEN lite software (https://www.zeiss.com.cn/microscopy/products/microscope-software/zen.html#inpagetabs-5).
RNA extraction and qRT-PCR analysis
Total RNA was isolated using the Trizol reagent (TIANGEN, China), then reverse-transcribed to cDNA using the first strand cDNA systhesis kit (TIANGEN, China). To analyze mRNA expression, the SuperReal PreMix Color (SYBR Green) qRT-PCR kit (TIANGEN, China) was used, with GAPDH as an internal amplification control. We isolated cellular and exsomal miRNAs using miRNeasy Mini Kit (QIAGEN, Germany), and performed cDNA synthesis using miRcute Plus miRNA First-Strand cDNA Kit (TIANGEN, China). miRNA expression was examined using miRcute Plus miRNA qPCR Kit (SYBR Green) (TIANGEN, China). To detect cellular miRNAs, U6 was used as a standard control. To compare the expression of miRNAs between cell and exosomes, equivalent synthetic external control cel-39–3p was added to the system during the reverse transcription step. Primer sequences used during PCR analysis are outlined in Table S1.
Statistical analyses
Data were presented as mean ± standard errors of the means. A student’s t-test was used to reveal any significant differences between two groups, while One-way analysis of variance (ANOVA) was performed to compare multiple (>2) means. Statistical analyses were performed using SPSS 22.0 software. P-values ≤ 0.05 were considered to be significant. All experiments were performed using at least 3 independent replicates.
Data availibility
The data that support the findings of this study are available in GEO at https://www.ncbi.nlm.nih.gov/geo/. These data were derived from the following resources available in the public domain: GSE17025, GSE63678 and GSE115810. The authors declare that there are no conflicts of interests. The corresponding author is responsible for submitting a competing interest’s statement on behalf of all authors of the paper.
Supplementary information
Acknowledgements
This research was supported by grants from Department of Science Technology of Zhejiang Province (2016C02054-8), Department of Science Technology of Tongxiang City (201901002), Zhejiang Team Science and Technology Commissioner Project (Tongxiang) and Leading Innovation Team Project of South Taihu Elite Program.
Author contributions
T.Q., and S.S. performed the experiments and Drafted manuscript; Analyzed data: T.Q., S.S., and H.H.P.; Interpreted results of experiments: T.Q., S.S., and W.Z.G; Prepared figures: T.Q.; Cell culturing: L.J.J., C.D.R., and T.Q.; Western blot: H.H.P, T.Q., and S.S.; Edited and revised manuscript: T.Q., W,Z,G., S.S., and H.H.P; Conception and design of research: T.Q., W.Z.G.; Approved final version of manuscript: S.S., T.Q., F.Q.F., H.H.P., L.J.J., C.D.R., and W.Z.G.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Shuang Shi and Qiang Tan.
Contributor Information
Qiang Tan, Email: tq19952010@126.com.
Zhengguang Wang, Email: wzhguang68@zju.edu.cn.
Supplementary information
is available for this paper at 10.1038/s41598-020-66872-3.
References
- 1.Braun MM, Overbeek-Wager EA, Grumbo RJ. Diagnosis and Management of Endometrial Cancer. Am. Fam. Physician. 2016;93:468–474. [PubMed] [Google Scholar]
- 2.Sorosky JI. Endometrial Cancer. Obstet. Gynecol. 2012;120:383–397. doi: 10.1097/AOG.0b013e3182605bf1. [DOI] [PubMed] [Google Scholar]
- 3.Zhang Y, et al. The association between metabolic abnormality and endometrial cancer: A large case-control study in China. Gynecol. Oncol. 2010;117:41–46. doi: 10.1016/j.ygyno.2009.12.029. [DOI] [PubMed] [Google Scholar]
- 4.Murali R, Soslow RA, Weigelt B. Classification of endometrial carcinoma: more than two types. Lancet Oncol. 2014;15:E268–E278. doi: 10.1016/S1470-2045(13)70591-6. [DOI] [PubMed] [Google Scholar]
- 5.Mhawech-Fauceglia, P. et al. Gene Expression Profiles in Stage I Uterine Serous Carcinoma in Comparison to Grade 3 and Grade 1 Stage I Endometrioid Adenocarcinoma. PLoS One6 (2011). [DOI] [PMC free article] [PubMed]
- 6.Huo, X. et al. Identification of prognosis markers for endometrial cancer by integrated analysis of DNA methylation and RNA-Seq data. Sci. Rep. 9 (2019). [DOI] [PMC free article] [PubMed]
- 7.Lee, C.J. et al. Crosstalk between SOX2 and cytokine signaling in endometrial carcinoma. Sci. Rep. 8 (2018). [DOI] [PMC free article] [PubMed]
- 8.Levine RL, et al. PTEN Mutations and Microsatellite Instability in Complex Atypical Hyperplasia, a Precursor Lesion to Uterine Endometrioid Carcinoma. Cancer Res. 1998;58:3254. [PubMed] [Google Scholar]
- 9.Milane L, Singh A, Mattheolabakis G, Suresh M, Amiji MM. Exosome mediated communication within the tumor microenvironment. J. Controlled Release. 2015;219:278–294. doi: 10.1016/j.jconrel.2015.06.029. [DOI] [PubMed] [Google Scholar]
- 10.Tkach M, Thery C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell. 2016;164:1226–1232. doi: 10.1016/j.cell.2016.01.043. [DOI] [PubMed] [Google Scholar]
- 11.Colombo M, Raposo G, Thery C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annual Review of Cell and Developmental Biology, 2014;30:255–289. doi: 10.1146/annurev-cellbio-101512-122326. [DOI] [PubMed] [Google Scholar]
- 12.EL Andaloussi S, Maeger I, Breakefield XO, Wood MJA. Extracellular vesicles: biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 2013;12:348–358. doi: 10.1038/nrd3978. [DOI] [PubMed] [Google Scholar]
- 13.Bartel DP. MicroRNAs: Target Recognition and Regulatory Functions. Cell. 2009;136:215–233. doi: 10.1016/j.cell.2009.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Fang, T. et al. Tumor-derived exosomal miR-1247-3p induces cancer-associated fibroblast activation to foster lung metastasis of liver cancer. Nat. Commun. 9 (2018). [DOI] [PMC free article] [PubMed]
- 15.Liu X, et al. Exosomal transfer of miR-501 confers doxorubicin resistance and tumorigenesis via targeting of BLID in gastric cancer. Cancer Lett. 2019;459:122–134. doi: 10.1016/j.canlet.2019.05.035. [DOI] [PubMed] [Google Scholar]
- 16.Peinado H, et al. Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET. Nat. Med. 2012;18:883–891. doi: 10.1038/nm.2753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Srivastava, A. et al. A Non-invasive Liquid Biopsy Screening of Urine-Derived Exosomes for miRNAs as Biomarkers in Endometrial Cancer Patients. The AAPS Journal. 20 (2018). [DOI] [PubMed]
- 18.Lander, E.S. & Park, P.J.J.N. Integrated genomic characterization of endometrial carcinoma. Nature. [DOI] [PMC free article] [PubMed]
- 19.Liu L, Chen F, Xiu A, Du B, Xie WJAPJoCPA. Identification of Key Candidate Genes and Pathways in Endometrial Cancer by Integrated Bioinformatical Analysis. Asian Pacific Journal of Cancer Prevention Apjcp. 2018;19:969–975. doi: 10.22034/APJCP.2018.19.4.969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Akhil S, et al. A Non-invasive Liquid Biopsy Screening of Urine-Derived Exosomes for miRNAs as Biomarkers in Endometrial Cancer Patients. Aaps Journal. 2018;20:82. doi: 10.1208/s12248-018-0220-y. [DOI] [PubMed] [Google Scholar]
- 21.Martinez-Garcia E, et al. Advances in endometrial cancer protein biomarkers for use in the clinic. Expert Rev. Proteomics. 2018;15:81–99. doi: 10.1080/14789450.2018.1410061. [DOI] [PubMed] [Google Scholar]
- 22.Day RS, et al. Identifier mapping performance for integrating transcriptomics and proteomics experimental results. BMC Bioinformatics. 2011;12:213. doi: 10.1186/1471-2105-12-213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Pappa KI, et al. Profiling of Discrete Gynecological Cancers Reveals Novel Transcriptional Modules and Common Features Shared by Other Cancer Types and Embryonic Stem Cells. PLoS One. 2015;10:e0142229. doi: 10.1371/journal.pone.0142229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Benmoussa, A. et al. Commercial Dairy Cow Milk microRNAs Resist Digestion under Simulated Gastrointestinal Tract Conditions. J. Nutr. 2016:jn.116.237651. [DOI] [PubMed]
- 25.Shurtleff, M.J., M, T.-D.M., V, K.K., Sayaka, R. & Elife, S.R.J. Y-box protein 1 is required to sort microRNAs into exosomes in cells and in a cell-free reaction. eLife. 5.(2016). [DOI] [PMC free article] [PubMed]
- 26.Yueqiu et al. microRNA133a targets Foxl2 and promotes differentiation of C2C12 into myogenic progenitor cells. DNA Cell Biol.2015. [DOI] [PMC free article] [PubMed]
- 27.Hanahan D, Weinberg RA. Hallmarks of Cancer: The Next Generation. Cell. 2011;144:646–674. doi: 10.1016/j.cell.2011.02.013. [DOI] [PubMed] [Google Scholar]
- 28.Arend RC, Jones BA, Martinez A, Goodfellow P. Endometrial cancer: Molecular markers and management of advanced stage disease. Gynecol. Oncol. 2018;150:569–580. doi: 10.1016/j.ygyno.2018.05.015. [DOI] [PubMed] [Google Scholar]
- 29.Di Cello, A. et al. DJ-1 is a reliable serum biomarker for discriminating high-risk endometrial cancer. Tumor Biology39 (2017). [DOI] [PubMed]
- 30.Stegh AH. Targeting the p53 signaling pathway in cancer therapy - the promises, challenges and perils. Expert Opin. Ther. Targets. 2012;16:67–83. doi: 10.1517/14728222.2011.643299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zhang Y, et al. Prognostic factors and genes associated with endometrial cancer based on gene expression profiling by bioinformatics analysis. Arch. Gynecol. Obstet. 2016;293:1287–1295. doi: 10.1007/s00404-015-3886-8. [DOI] [PubMed] [Google Scholar]
- 32.Lapinska-Szumczyk S, et al. HER2-Positive Endometrial Cancer Subtype Carries Poor Prognosis. Clinical and Translational Science. 2014;7:482–488. doi: 10.1111/cts.12207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Supernat A, et al. Tumor Heterogeneity at Protein Level as an Independent Prognostic Factor in Endometrial Cancer. Transl. Oncol. 2014;7:613–619. doi: 10.1016/j.tranon.2014.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Claire B, et al. Post-translational modifications in DNA topoisomerase 2α highlight the role of a eukaryote-specific residue in the ATPase domain. Sci Rep. 2018;8:10673. doi: 10.1038/s41598-018-28936-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Chen T, Sun Y, Ji P, Kopetz S, Zhang W. Topoisomerase IIalpha in chromosome instability and personalized cancer therapy. Oncogene. 2015;34:4019–4031. doi: 10.1038/onc.2014.332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Erriquez J, et al. TOP2A gene copy gain predicts response of epithelial ovarian cancers to pegylated liposomal doxorubicin TOP2A as marker of response to PLD in ovarian cancer. Gynecol. Oncol. 2015;138:627–633. doi: 10.1016/j.ygyno.2015.06.025. [DOI] [PubMed] [Google Scholar]
- 37.de Resende, M.F. et al. Prognostication of prostate cancer based on TOP2A protein and gene assessment: TOP2A in prostate cancer. J. Transl. Med. 11 (2013). [DOI] [PMC free article] [PubMed]
- 38.Pei, Y.F., Yin, X.-m. & Liu, X.-q.J.B.B.A. TOP2A induces malignant character of pancreatic cancer through activating β-catenin signaling pathway. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease. S0925443917303836 (2017). [DOI] [PubMed]
- 39.Kouprina N, et al. The microcephaly ASPM gene is expressed in proliferating tissues and encodes for a mitotic spindle protein. Hum. Mol. Genet. 2005;14:2155–2165. doi: 10.1093/hmg/ddi220. [DOI] [PubMed] [Google Scholar]
- 40.Capecchi, M.R. & Pozner, A. ASPM regulates symmetric stem cell division by tuning Cyclin E ubiquitination. Nat. Commun. 6 (2015). [DOI] [PMC free article] [PubMed]
- 41.Xu ZL, Zhang Q, Luh F, Jin BY, Liu XY. Overexpression of the ASPM gene is associated with aggressiveness and poor outcome in bladder cancer. Oncol. Lett. 2019;17:1865–1876. doi: 10.3892/ol.2018.9762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Xie JJ, et al. High expression of ASPM correlates with tumor progression and predicts poor outcome in patients with prostate cancer. Int. Urol. Nephrol. 2017;49:817–823. doi: 10.1007/s11255-017-1545-7. [DOI] [PubMed] [Google Scholar]
- 43.Tang, J.N. et al. Overexpression of ASPM, CDC20, and TTK Confer a Poorer Prognosis in Breast Cancer Identified by Gene Co-expression Network Analysis. Frontiers in Oncology9 (2019). [DOI] [PMC free article] [PubMed]
- 44.Yang, T.T. et al. Epigenetic Inactivation of EFEMP1 Is Associated with Tumor Suppressive Function in Endometrial Carcinoma. PLoS One8 (2013). [DOI] [PMC free article] [PubMed]
- 45.Dou CY, et al. EFEMP1 inhibits migration of hepatocellular carcinoma by regulating MMP2 and MMP9 via ERK1/2 activity. Oncol. Rep. 2016;35:3489–3495. doi: 10.3892/or.2016.4733. [DOI] [PubMed] [Google Scholar]
- 46.Lang Y, Meng J, Song X, Chen XJCJoLC. EFEMP1 Suppresses Growth and Invasion of Lung Cancer Cells by Downregulating Matrix Metalloproteinase-7 Expression. Chinese Journal of LungCancer. 2015;18:92–97. doi: 10.3779/j.issn.1009-3419.2015.02.08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Yin XX, et al. EFEMP1 promotes ovarian cancer cell growth, invasion and metastasis via activated the AKT pathway. Oncotarget. 2016;7:47938–47953. doi: 10.18632/oncotarget.10296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Stelloo E, et al. Refining prognosis and identifying targetable pathways for high-risk endometrial cancer; a TransPORTEC initiative. Mod. Pathol. 2015;28:836–844. doi: 10.1038/modpathol.2015.43. [DOI] [PubMed] [Google Scholar]
- 49.Liu XL, et al. FOXL2 suppresses proliferation, invasion and promotes apoptosis of cervical cancer cells. Int. J. Clin. Exp. Pathol. 2014;7:1534–1543. [PMC free article] [PubMed] [Google Scholar]
- 50.Dong JQ, et al. HMGA2-FOXL2 Axis Regulates Metastases and Epithelial-to-Mesenchymal Transition of Chemoresistant Gastric Cancer. Clin. Cancer. Res. 2017;23:3461–3473. doi: 10.1158/1078-0432.CCR-16-2180. [DOI] [PubMed] [Google Scholar]
- 51.Shao HL, et al. New Technologies for Analysis of Extracellular Vesicles. Chem. Rev. 2018;118:1917–1950. doi: 10.1021/acs.chemrev.7b00534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hon, K.W., Ab-Mutalib, N.S., Abdullah, N.M.A., Jamal, R. & Abu, N. Extracellular Vesicle-derived circular RNAs confers chemoresistance in Colorectal cancer. Sci. Rep. 9 (2019). [DOI] [PMC free article] [PubMed]
- 53.Thery C. CANCER Diagnosis by extracellular vesicles. Nature. 2015;523:161–162. doi: 10.1038/nature14626. [DOI] [PubMed] [Google Scholar]
- 54.Luo YQ, et al. microRNA133a Targets Foxl2 and Promotes Differentiation of C2C12 into Myogenic Progenitor Cells. DNA Cell Biol. 2015;34:29–36. doi: 10.1089/dna.2014.2522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kharaziha P, Ceder S, Li Q, Panaretakis T. Tumor cell-derived exosomes: A message in a bottle. Biochimica et Biophysica Acta. 2012;1826:103–111. doi: 10.1016/j.bbcan.2012.03.006. [DOI] [PubMed] [Google Scholar]
- 56.Clough E, Barrett TJMiMB. The Gene Expression Omnibus Database. Methods in Molecular Biology. 2016;1418:93. doi: 10.1007/978-1-4939-3578-9_5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Pappa, K.I. et al. Profiling of Discrete Gynecological Cancersop Reveals Novel Transcriptional Modules and Common Features Shared by Other Cancer Types and Embryonic Stem Cells. Plos One. 10, e0142229. [DOI] [PMC free article] [PubMed]
- 58.Hermyt, E., Grabarek, Z. N. & Kruszniewska-Rajs, B. C. Interplay between miRNAs and Genes Associated with Cell Proliferation in Endometrial Cancer. Int J Mol Sci6, 23 (2019). [DOI] [PMC free article] [PubMed]
- 59.Robinson MD, McCarthy DJ, Smyth GKJB. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics. 2010;26:139–140. doi: 10.1093/bioinformatics/btp616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Huang, D.W., Sherman, B.T. & Lempicki, R.A.J.N.P. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 4, 44-57. [DOI] [PubMed]
- 61.Tang ZF, Kang BX, Li CW, Chen TX, Zhang ZM. GEPIA2: an enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019;47:W556–W560. doi: 10.1093/nar/gkz430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Li TW, et al. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017;77:E108–E110. doi: 10.1158/0008-5472.CAN-17-0307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Satoh T, et al. Establishment of a new human endometrial adenocarcinoma cell line, Watanabe cells, containing estrogen receptor. Hum. Cell. 1995;8:195–201. [PubMed] [Google Scholar]
- 64.Kuramoto H, Tamura S, Notake Y. Establishment of a cell line of human endometrial adenocarcinom. American Journal of Obstetrics & Gynecology. 1972;114:1012. doi: 10.1016/0002-9378(72)90861-7. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.