

Abstract
Background
Traumatic brain injury (TBI) occurs in as many as 64–74 million people worldwide each year and often results in one or more post-traumatic syndromes, including depression, cognitive, emotional, and behavioral deficits. TBI can also increase seizure susceptibility, as well as increase the incidence of epilepsy, a phenomenon known as post-traumatic epilepsy (PTE). Injury type and severity appear to partially predict PTE susceptibility. However, a complete mechanistic understanding of risk factors for PTE is incomplete.
Main body
From the earliest days of modern neuroscience, to the present day, accumulating evidence supports a significant role for neuroinflammation in the post-traumatic epileptogenic progression. Notably, substantial evidence indicates a role for astrocytes, microglia, chemokines, and cytokines in PTE progression. Although each of these mechanistic components is discussed in separate sections, it is highly likely that it is the totality of cellular and neuroinflammatory interactions that ultimately contribute to the epileptogenic progression following TBI.
Conclusion
This comprehensive review focuses on the neuroinflammatory milieu and explores putative mechanisms involved in the epileptogenic progression from TBI to increased seizure-susceptibility and the development of PTE.
Keywords: Traumatic brain injury, TBI, Astrocytes, Microglia, Cytokines, Chemokines, Epileptogenesis, Inflammation
Introduction
Traumatic brain injury (TBI) occurs in as many as 64-74 million people worldwide each year [1]. TBI severity ranges from mild to severe, and may cause post-traumatic syndromes, including depression, cognitive, emotional, and behavioral deficits. TBI may also cause post-traumatic seizures (PTS), increase seizure susceptibility and increase the incidence of epilepsy, a phenomenon known as post-traumatic epilepsy (PTE). Despite intensive research, biomarkers and treatments are lacking, as is a clear mechanistic understanding of the epileptogenic factors that may contribute to the onset of PTE.
While injury type and severity appear to partially predict PTE susceptibility, similar injuries in people do not always cause PTE [2]. Lesion location may influence the risk of PTE, as temporal lobe lesions following TBI are related to both a high incidence of early seizures and longitudinal development of PTE [3]. Penetrating lesions in motor areas and the parietal lobe are also associated with an increased risk of PTE [4]. Importantly, some risk factors have been suggested to be neuropathologically relevant in PTE development in humans, such as age, early seizures after TBI, and trauma severity [5].
Following TBI, a neuroinflammatory response is rapidly initiated and mounting evidence from human and animal studies support a pro-epileptogenic role of the neuroinflammatory response in the development of PTE [6]. In general, following TBI, there is a rapid release of inflammatory cytokines, chemokines and complement proteins. This immune response signals a variety of cellular mediators and also can initiate the acute phase response [7–13]. Following these signals, astrocytes and resident microglial cells are induced to become activated, proliferate, and migrate to the injury site [14, 15]. Peripheral immune cells are also described to infiltrate into the brain in response to TBI. Once this immune/neuroimmune response is activated to re-establish tissue homeostasis, these immune cells remove debris and identify potentially pathogenic signaling. Interestingly, while the most intense neuroinflammatory response occurs relatively early (within hours and days after the injury), a low-level of neuroinflammation often chronically persists [14, 16–20]. Both the acute/early and the chronic neuroinflammation have been implicated in epileptogenesis, and herein, the evidence for pro-epileptogenic contributions of neuroinflammation will be reviewed.
Some of the earliest neuropathological reports recognized that a progressive gliosis at the site of a brain injury was a major component of the development of an epileptogenic focus [4, 21]. Accumulating evidence continues to support glial scarring and other neuroinflammatory mechanisms in PTE. In 2004, the founding of the Journal of Neuroinflammation by Drs. Sue T. Griffin and Robert E. Mrak, provided a platform that sparked a reinvigorated focus on mechanisms of neuroinflammation in neuropathological disorders. To pay tribute to Dr. Mrak’s role in ushering in a new era of neuroscience, an overarching review of neuroinflammation following TBI is discussed, with a specific focus on neuroinflammatory mechanisms that can promote seizures, epileptogenesis and the development of PTE.
Etiology and incidence of PTE
The incidence of epilepsy is estimated to be approximately 0.5–2% of the general population. This incidence rate increases to approximately 5–7% in patients who experienced a precipitating head injury [22–24] and/or have been hospitalized for TBI [2, 25, 26]. A greater injury severity has been correlated with a higher PTE risk [27], and this risk increases up to 10-fold in military patients with penetrating head wounds. At the extreme, some estimates have suggested that the incidence of PTE is greater than 50% following severe penetrating head injuries [28–30]. Taken together, it is estimated that as many as 20% of symptomatic epilepsies are caused by TBI [31], and this population represents the largest known etiological cause of seizures and epilepsy.
Although early seizures that occur within a week of TBI can often be effectively managed by typical anti-seizure medications like levetiracetam and phenytoin [32], such treatments do not necessarily ameliorate the risk to develop PTE [33, 34]. Recurrent spontaneous seizures that define PTE are resistant to anti-epileptic treatments in about one-third of patients [34–36], and the side-effects from anti-epileptic drugs are found to be more severe in PTE patients [37]. Thus, understanding the pro-epileptogenic mechanisms of TBI is vital for the diagnosis and treatment of PTE and for improving quality-of-life measures in these patients.
Astrocytes and PTE: support cell, inflammatory mediator, or pathological nexus?
Transformative studies have re-defined the classical role of astrocytes in the brain. Astrocytes were initially considered to be primarily support cells [38], sub serving neuronal function and helping to maintain brain homeostasis. Although there was early recognition of the role of astrocytes in the response to injury [21], in the decades since, the extensive roles that astrocytes play in the pathogenic inflammatory response continues to be appreciated and explored [39]. Thus, it is abundantly clear, that far from their classification as merely support cells, astrocytes are actively and directly involved in multiple aspects of neuronal function.
Astrocytes are the most abundant cell type in the brain [40] and are now known to be involved in regulating ion homeostasis, maintaining blood-brain barrier function, metabolizing neurotransmitters, as well as providing nutrient and energy support for neuronal function. Astrocytes are key components in learning and memory, sleep, and other fundamental brain functions [41, 42] and are important components of the neuroinflammatory response.
Astrocytes play a key role in regulating neuronal activity, energizing neuronal metabolism by exchanging neuronal pyruvate for astrocytic lactate, and increasing NADH levels in neurons [43]. Astrocytes are active in neuronal information processing, and their processes envelop thousands of synapses to control neuronal activity through neurotransmitter uptake and release [41, 42]. Astrocytes also regulate the availability of glutamate and GABA in the synaptic cleft, thereby modulating synaptic transmission [44–46]. Thus, there are numerous mechanisms by which astrocytes might contribute to post-traumatic epileptogenesis. Here, we focus on those mechanisms that are related to neuroinflammation.
Astrocyte activation is a major cellular component of the neuroinflammatory response, and gliosis is commonly seen following TBI. Astrocytosis can also occur as part of the neuroinflammatory response. In post-mortem TBI human brains, a widespread astrocytosis is seen at the primary injury site, as well as at ipsilateral and contralateral brain regions that are distant from the initial injury site [47–51]. It is pertinent to note that in the post-mortem epileptic brain, it is not possible to delineate between seizure-induced gliosis and gliosis that might be pro-epileptogenic. The fact that similar patterns of gliosis are also observed in numerous animal models of TBI [14, 35, 52–57] provides an opportunity for the investigation of potential causal astrocytic mechanisms of epileptogenesis.
The astrocytic response to TBI results from neuronal cell death and axonal degeneration, as well as the associated rapid release of inflammatory complement system factors, cytokines, and chemokines from microglia, neurons, and the astrocytes themselves (Fig. 1). Regardless of the source, this release of cytokines may influence pathological functioning of the astrocytes, notably as it pertains to physiological signaling and epileptogenesis.
Fig. 1.
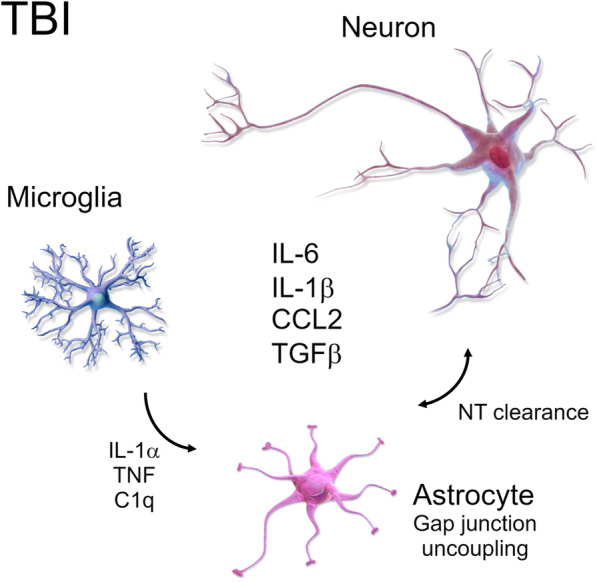
Microglia (purple, left), astrocytes (pink, bottom), and neurons (red, upper right) are activated and altered after TBI. Cytokines interleukin-1α, TNF, and complement component 1 subcomponent q (C1q) are secreted by activated microglia and can induce the A1 astrocyte phenotype. Astrocytes suffer gap junction uncoupling and have impaired neurotransmitter (NT) clearance and metabolic recycling from synapses. Cytokines interleukin-6, interleukin-1β, transforming growth factor beta (TGFβ), and chemokine CCL2 are secreted in high concentrations creating a neuroinflammatory milieu. Cells adapted from Blausen Medical Gallery [58]
In a series of important studies, Steinhauser and colleagues demonstrated that astrocytes are functionally changed in epileptic brains, such that they exhibit smaller K+ currents and lose the ability for gap junction coupling. These physiological changes were postulated to be a primary causative factor in the development of epilepsy [59–61]. Astrocyte activation leads to an increase in intracellular calcium concentration and results in the increase of glutamate release as a gliotransmitter [62, 63]. This glutamate release promotes neuronal excitotoxicity and increases the potential to generate seizures. This event appears to involve inflammation-associated alterations in receptor expression as well as dysfunctional gap junction coupling [63]. In TBI, one component of astrocyte activation is the uncoupling of astrocytic gap junctions [64, 65].
In addition to a role for astrocytic gap junction uncoupling in seizures, a role for astrocytic Cx43 hemichannels was observed for seizures in the pilocarpine model of epileptogenesis and the 6 HZ corneal kindling model [66]. Inhibiting the Cx43 hemichannel with GAP19, a selective hemichannel inhibitor that does not influence the gap junctions prevented their opening and decreased seizures [67]. This anti-convulsive effect was found to be mediated by D-serine because addition of exogenous D-serine prevented seizure inhibition by GAP19 [66]. Therefore, the TBI-induced neuroinflammatory response may interfere with the ability of astrocytes to effectively buffer ions throughout the astrocytic syncytium, and this dysfunction is likely related to issues with the astrocytic gap junctions (Fig. 1).
The neuroinflammatory response also induces other morphological and functional changes to astrocytes, and some of these mechanisms are related to seizures and epilepsy. For example, aquaporin-4 function and distribution is known to be altered within the neuroinflammatory environment. Aquaporin-4 was also found to be important for reducing post-traumatic seizure susceptibility in a PTZ second-hit challenge administered 1-month after TBI. In this study, aquaporin-4-/- mice had reduced latency to seizure onset and increased seizure severity [68], suggesting a role for astrocytic aquaporins in post-traumatic epileptogenesis. In other epilepsy models, mice lacking aquaporin-4 were found to be seizure-resistant to PTZ-induced seizures in the absence of a precipitating TBI [69], suggesting a potentially broader role for aquaporins in regulating seizure activity. In the context of TBI, it is possible that injury-induced alterations to aquaporin-4 dysfunction might be directly involved in promoting a pro-epileptogenic environment. Future studies are needed to better understand the role of astrocytes in this pathological process.
Another morphological change to astrocytes that is associated with the neuroinflammatory response is astrocyte hypertrophy. A series of studies have shown that hypertrophied astrocytes may play an important role in the development of pro-epileptogenic circuits after TBI that could promote the development of PTE. Shapiro and colleagues performed a series of studies examining the astrocytic, radial glial-like scaffold in the hippocampal dentate gyrus in the pilocarpine model of epileptogenesis [70–75]. Subsequently, these authors examined them in a model of neonatal hypoxia-induced epileptogenesis [76] and following TBI [77, 78]. In normal brains, these radial glial-like astrocytes send their radial processes through the granule cell layer providing a scaffold for the normal growth and integration of the granule cell apical dendrites [79–82]. Following pilocarpine, neonatal hypoxia, or TBI, these astrocytes were shown to be hypertrophied. In addition, they had altered their orientation, such that the radial glial cells preferentially extended their processes into the hilus instead of into the granule cell layer [70–74, 77]. Aberrantly sprouted basal dendrites from the granule cells grow along this ectopic glial scaffold into the hilus [70]. In addition to these ectopic granule cell basal dendrites after TBI, the mossy fiber axons of the granule cell are also induced to sprout [83–88]. Interestingly, the mossy fiber sprouting after TBI is most prominent within the dentate gyrus itself, which further primes the targeting of ectopic basal dendrites that become targeted for synaptogenesis, by the sprouted mossy fiber axons [72, 89]. This latter phenomenon of granule cell-to-granule cell connectivity has been termed a recurrent excitatory circuitry [79, 90]. This aberrant circuitry produces excitatory drive that can both promote and facilitate the spread of seizure activity [91, 92], and more recent studies further support the role of this aberrant circuitry in epileptogenesis [93–95], including the involvement of injury-induced atypical astrocytes [96]. Interestingly, a study also showed that during pilocarpine-induced epileptogenesis, the radial glial-like processes in the dentate gyrus upregulate the expression of CCR2. CCR2, along with its ligand CCL2, have been shown to act as chemotactic guidance cues for the migration of immature neurons [97–100]. Therefore, it is possible that these radial glial astrocytes provide an anatomical substrate and chemotactic cues for the aberrant growth of epileptogenic circuitry.
Microglia in PTE
Following TBI or other pro-epileptogenic stimuli, microglial cells become rapidly activated. This activation may persist for months or even years after the initial injury [101–106]. Microglial cells are the resident macrophages of the central nervous system (CNS), and their role in immune defense is widely accepted. During early development, microglia originate from primitive macrophages that migrate to developing neuroepithelium from the embryonic yolk sac and reside in the mature CNS throughout the lifespan [107, 108]. In addition to their immune role in the CNS, studies have indicated that microglial cells also play a pivotal role in neuronal proliferation, differentiation, and sculpting of synaptic connections [109–111].
Most studies of epileptogenesis following TBI do not distinguish resident microglial cells, from infiltrating macrophages that migrated to the brain in response to injury and blood-brain barrier breakdown. Circulating Ly-6C(hi) CCR2(+) monocytes are also recruited to lesioned areas [112]. In TBI studies, distinctions have rarely been made between resident microglial cells and infiltrating macrophages. Herein, the term microglial cells will be used to describe any macrophage in the brain after injury. When possible, differences between microglial cells and infiltrated macrophages will be noted.
Activation of microglial cells is also a common feature of TBI and epileptogenesis [113]. In resting state and normal conditions, microglia are highly dynamic cells that continually assess the microenvironment by extending and retracting processes with bulbous endings, throughout the brain parenchyma [114]. Microglial cells are homogenously distributed, and their processes are in close contact with astrocytes, neurons, and vessels. In response to activation cues, the microglial cells and their processes, orient, and migrate toward the injury site in order to isolate the injured tissue and phagocytose cellular debris [114, 115]. In addition, resident microglia or infiltrated macrophages may also act as antigen presenting cells [116].
The seizure-inducing role of monocytes and microglia is supported by studies showing that inhibition of microglial cells using minocycline or minocycline derivatives reduced post-traumatic seizures, epileptogenesis, and cognitive deficits [101, 117–119]. In a post-traumatic kindling model, rats with TBI were found to kindle faster and have more intense seizures than non-TBI rats [120, 121]. Targeting microglial cells by pretreatment with the toll-like receptor (TLR) antagonists, Pam3Cys, and monophosphoryl lipid A rendered rats less susceptible to kindling and more like kindled rats that did not undergo a prior TBI [122]. More specifically, antagonizing the toll-like receptors resulted in higher seizure thresholds, slower speed to kindling, and reduced duration of kindled seizures. Other studies outside of epileptogenesis have also postulated a role for microglial cells in hyperexcitability. Microglial cells activated by lipopolysaccharide or heat-killed Gram-negative bacteria induced hyperexcitability of cerebellar purkinje cells that was suppressed by inhibiting or depleting the microglia [123]. Another study showed that using minocycline to inhibit microglial activation in a repeated toluene inhalation model prevented neuronal hyperexcitability by ameliorating the loss of the slow calcium-dependent potassium current [124]. Finally, Devinsky et al. [125] reviewed the role of glia-induced hyperexcitability and concluded that microglial cells make a significant contribution to hyperexcitability, via direct and indirect (e.g., cytokine release) mechanisms. Therefore, there is some evidence for a direct role of microglial cells in inducing neuronal hyperexcitability, but more research is needed.
Interestingly, treatment with lipopolysaccharide (LPS) prior to TBI also reduced post-traumatic kindling susceptibility [126]. In these studies, the TLR antagonists were administered prior to TBI, suggesting that a priming effect on the microglial cells might be taking place. An alternative interpretation is that the microglial cells may exert a negative influence on epileptogenesis, but a precipitating immune insult such as toll-like receptor antagonism may prime the microglial cells, thus inhibiting the putative epileptogenic influence that they exert. It should be noted that these compounds exclusively target microglial cells, so other mechanisms of protection are probably not involved.
Microglia activation most likely occurs in response to various pro-inflammatory cytokines and chemokines and the release of danger-associated molecular patterns (DAMPs) by damaged cells [127]. Injury in the brain causes the release of the DAMP signal, high mobility group box 1 protein (HMGB1). Immune cells, neurons, and glia can also release HMGB1 in response to cytokine stimulation [128]. HMGB1 works through activating TLR 4, and it has been noted that mice with TLR4 mutation are resistant to seizures [129]. These data support a role for microglial signaling in PTE and suggest that targeting specific signaling components, such as toll-like receptors, MHCII, and other microglial-specific receptors, might be a viable therapeutic target.
Microglial cells might also wield a double-edged sword that can positively or negatively influence PTE development. Macrophage studies in vitro established the M1 (classical) and M2 (alternative) activation states [130], whereas in vivo macrophage cell populations present activation states as a continuum. The M1/M2 functional polarization may still have utility in defining cytokine secreting cellular profiles. Both M1 and M2 microglia are noted in damaged tissue [101], and their relative ratios may be related to outcomes. The M1 secreting profile could be more advantageous during the acute response to injury. However, M1 microglia have been shown to chronically persist after TBI, and these persistent pro-inflammatory microglia have been implicated in chronic neurological dysfunction following injury [131]. The appearance of M2 microglia likely occurs in response to elevated interleukin-4 (IL4) and IL13 to promote repair of tissue damage by matrix remodeling. In response to IL10, glucocorticoid, and transforming growth factor beta (TGFβ), microglia polarize to a deactivated M2c state and turn off inflammation [132]. However, it is not clear if, or how, this mechanism influences the chronic M1 microglial phenotype. Therefore, it appears as though a broad spectrum of microglial cells is involved in the inflammatory response to TBI. Future studies are needed to better define these microglial subsets and their roles in PTE.
Also, microglia interaction with astrocytes should be considered in PTE neuroinflammation. Cytokines, interleukin-1α (IL-1α), TNF, and complement component 1 subcomponent q (C1q) secreted by activated microglia can induce the A1 astrocyte phenotype [133] (Fig. 1). This phenotype was denominated in analogy to M1/M2 phenotypes observed in macrophages. A1 astrocytes are neurotoxic leading to neuronal death, synapse disassembly, and oligodendrocyte death.
Cytokine and chemokine contributions to PTE
One prominent consequence of TBI is the rapid and prolonged release of inflammatory cytokines and chemokines [134–136]. The list of, and role for, inflammatory proteins seems to expand at an almost exponential rate. Of the numerous cytokines known to be released in response to TBI, a multitude of studies demonstrate increased tumor necrosis factor (TNF), TGFβ, IL-1β, IL-6, and IL-10, among other cytokines, that are consistently found to be elevated after TBI. The TBI-induced release of cytokines and chemokines may directly and indirectly increase regional hyperexcitability of neurons and contribute to seizures (Fig. 1). While it is likely that many of the inflammatory proteins play a role in epileptogenesis, a paucity of research limits discussion only to those which have been explicitly explored for seizure-inducing mechanisms.
The CSF-serum ratio of IL-1β is elevated in TBI patients who are susceptible for developing epilepsy [137]. It has been reported that transgenic mice that overexpress IL-1β and TNF have decreased seizure threshold [138]. Consistent with this notion, treatment with an IL-1β receptor antagonist results in decreased seizure susceptibility in young mice [139] and promotes M1-type microglial cytokine and chemokine release after TBI [140]. IL-1β has been reported to increase NMDA-mediated Ca+ current in the pyramidal neurons through cell surface type 1 IL-1R (IL-1R1), co-localized on pyramidal cell dendrites, and to concurrently decrease the seizure threshold [141, 142]. Thus, IL-1β may be an important component in PTE. IL-10 can be used as treatment to inhibit IL-1β secretion as demonstrated in epileptic mouse experimental animals [143]. Numerous CNS and non-CNS cell types are capable of releasing IL-1β, and any one or more of these cells can contribute to pathology.
Emerging role of TGFβ in seizures and acquired epilepsies
TGFβ has been implicated in the development of excitatory synaptogenesis and PTE following TBI. TGFβ signaling has been shown to trigger seizures, neuronal hyperexcitability, and epileptogenesis [144, 145]. TGFβ expression is increased in cortex and hippocampus after TBI [35], and administration of LY-364947, a TGFβ type 1 receptor inhibitor, significantly reduced the duration and severity of post-traumatic seizures in the second-hit pentylenetetrazole (PTZ) challenge [35]. Incubating cortical slices with TGFβ induced epileptiform activity in slices, and this activity was blocked by inhibiting TGFβ receptors [146], suggesting an important role for TGFβ signaling in epileptogenesis. Transcriptome analysis also supports a role for TGFβ signaling in the epileptogenic transcriptional response, and this response can be blocked by inhibiting TGFβ receptor signaling [146]. Other studies found that blocking astrocytic TGFβ R1 activation prevented the development of epilepsy in the pilocarpine model of epileptogenesis [144]. Specifically, astrocytic TGFβ signaling was found to induce excitatory synaptogenesis that preceded the development of seizures, and these effects were blocked by inhibition of TGFβ signaling [145]. Therefore, TGFβ causes hyperexcitability and seizures, and blocking TGFβ signaling prevents the development of acquired epilepsies. This phenomenon may be directly mediated via astrocytic TGFβ.
IL-6 elevation is associated with epileptogenesis
TBI increases the level of IL-6 in the peripheral blood [147], and it has been reported that the serum level of IL-6 positively correlates with the severity of TBI [20]. Sustained elevation of IL-6 is associated with increased odds for detrimental overall outcomes in the first year following TBI [19]. IL-6 is also thought to play a critical role in seizure development, and elevated IL-6 has been observed in patients with temporal lobe epilepsy [148], pediatric epilepsy [149], and electrical status epilepticus in sleep (ESES) [150]. Consistent with the notion that elevated IL-6 is associated with seizures in patients with ESES, immunomodulation that reduced IL-6 levels also improved electroencephalographic seizure activity [150]. IL-6 gene polymorphisms have also been implicated in pediatric epilepsy and febrile seizure cases [149, 151–153]. Transgenic mice that expressed elevated astrocytic IL-6 were more susceptible to NMDA-induced seizures and sub-threshold doses of kainic acid [154], with the latter causing mice to have severe tonic-clonic seizures and death. IL-6 administration in rodents has been shown to increase seizure severity and decrease seizure threshold [155]. Moreover, a potential role for IL-6 in epileptogenesis has been reported in cases of subarachnoid hemorrhage, where IL-6 was significantly elevated in patients that developed seizures [156]. Taken together, IL-6 appears to have numerous roles in neuronal homeostasis, and its elevation is clearly associated with increased-seizure susceptibility and epilepsy.
Conclusion: a neuroinflammatory role in PTE
PTE is the most common type of symptomatic epilepsy. We described inflammatory components of cellular and molecular mechanisms in the CNS that can contribute to the epileptogenic progression following TBI. While the innate inflammatory response to an injury is relatively consistent, the combined effects that contribute to PTE are highly variable and notably appear to depend on timing, location, and an individual immune response. In this context, diagnostic tools that consider multi-modal variables will likely need to be developed. It is also important to recognize that adaptive immune components are also likely to be playing a role in the post-traumatic epileptogenic progression, and such variables require further investigation.
Considering that PTE is difficult to treat, being more resistant to first and second-line anti-epileptic treatments, there is hope that therapies which target specific inflammatory components after TBI may ultimately yield meaningful diagnostic tools and effective therapeutic strategies. When thinking about these possibilities, it is important to recognize the pioneers who have helped to pave the way for these lines of research. To this end, we wish to thank Dr. Mrak for his contributions to the field of neuroinflammation, and we dedicate this article to his founding, success, and leadership of the Journal of Neuroinflammation.
Acknowledgements
The authors would like to thank Gabriella Hollingsworth for providing feedback on a preliminary version of this review. Charles E. Ribak provided helpful comments to the manuscript.
Abbreviations
- CNS
Central nervous system
- C1q
Complement component 1 subcomponent q
- DAMPs
Danger associated molecular patterns
- GFAP
Glial fibrillary acidic protein
- HMGB1
High mobility group box 1 protein
- IL
Interleukin
- LPS
Lipopolysaccharide
- MHC
Major histocompatibility complex
- MBP
Myelin basic protein
- PTS
Post-traumatic seizures
- PTE
Post-traumatic epilepsy
- PTZ
Pentylenetetrazole
- TBI
Traumatic brain injury
- TGFβ
Transforming growth factor beta
- TNF
Tumor necrosis factor
- TLR-4
Toll-like receptor-4
Authors’ contributions
SM: contributed to the conceptual design and creation of this work, notably the section on astrocytes. GA: contributed to the conceptual design and creation of this work, notably the sections on microglia, cytokines, and chemokines. KM: contributed to the conceptual design and creation of this work, notably the sections on TBI etiology and microglia. KO: contributed to the conceptual design and creation of this work, notably the sections on TBI etiology, astrocytes, and microglia. LS: contributed to the overall conceptual design and creation of this work. The authors read and approved the final manuscript.
Funding
Not applicable
Availability of data and materials
Not applicable
Ethics approval and consent to participate
No human subjects or tissue
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Sanjib Mukherjee and Gabriel M. Arisi contributed equally to this work.
Contributor Information
Gabriel M. Arisi, Email: arisi@unifesp.br
Lee A. Shapiro, Email: lshapiro@medicine.tamhsc.edu
References
- 1.Dewan MC, Rattani A, Gupta S, Baticulon RE, Hung YC, Punchak M, Agrawal A, Adeleye AO, Shrime MG, Rubiano AM, Rosenfeld JV, Park KB. Estimating the global incidence of traumatic brain injury. J Neurosurg. 2018:1-18. Epub 2018/04/28. doi: 10.3171/2017.10.JNS17352. PubMed PMID: 29701556. [DOI] [PubMed]
- 2.Frey LC. Epidemiology of posttraumatic epilepsy: a critical review. Epilepsia. 2003;44 Suppl 10:11-17. PubMed PMID: 14511389. [DOI] [PubMed]
- 3.Tubi MA, Lutkenhoff E, Blanco MB, McArthur D, Villablanca P, Ellingson B, Diaz-Arrastia R, Van Ness P, Real C, Shrestha V, Engel J, Jr., Vespa PM. Early seizures and temporal lobe trauma predict post-traumatic epilepsy: a longitudinal study. Neurobiol Dis. 2019;123:115-21. Epub 2018/06/04. doi: 10.1016/j.nbd.2018.05.014. PubMed PMID: 29859872; PMCID: PMC6274611. [DOI] [PMC free article] [PubMed]
- 4.Russell WR, Whitty CW. Studies in traumatic epilepsy. I. Factors influencing the incidence of epilepsy after brain wounds. J Neurol Neurosurg Psychiatry. 1952;15(2):93-8. Epub 1952/05/01. doi: 10.1136/jnnp.15.2.93. PubMed PMID: 14928078; PMCID: PMC497164. [DOI] [PMC free article] [PubMed]
- 5.DeGrauw X, Thurman D, Xu L, Kancherla V, DeGrauw T. Epidemiology of traumatic brain injury-associated epilepsy and early use of anti-epilepsy drugs: an analysis of insurance claims data, 2004-2014. Epilepsy Res. 2018;146:41-9. Epub 2018/08/03. doi: 10.1016/j.eplepsyres.2018.07.012. PubMed PMID: 30071385; PMCID: PMC6547364. [DOI] [PMC free article] [PubMed]
- 6.Dadas A, Janigro D. Breakdown of blood brain barrier as a mechanism of post-traumatic epilepsy. Neurobiol Dis. 2019;123:20-26. Epub 2018/07/22. doi: 10.1016/j.nbd.2018.06.022. PubMed PMID: 30030025. [DOI] [PMC free article] [PubMed]
- 7.Kossmann T, Hans VH, Imhof HG, Stocker R, Grob P, Trentz O, Morganti-Kossmann C. Intrathecal and serum interleukin-6 and the acute-phase response in patients with severe traumatic brain injuries. Shock. 1995;4(5):311-317. Epub 1995/11/01. doi: 10.1097/00024382-199511000-00001. PubMed PMID: 8595516. [DOI] [PubMed]
- 8.Yang S, Ma Y, Liu Y, Que H, Zhu C, Liu S. Elevated serum haptoglobin after traumatic brain injury is synthesized mainly in liver. J Neurosci Res. 2013;91(2):230-239. Epub 2012/11/23. doi: 10.1002/jnr.23159. PubMed PMID: 23172820. [DOI] [PubMed]
- 9.Nizamutdinov D, DeMorrow S, McMillin M, Kain J, Mukherjee S, Zeitouni S, Frampton G, Bricker PC, Hurst J, Shapiro LA. Hepatic alterations are accompanied by changes to bile acid transporter-expressing neurons in the hypothalamus after traumatic brain injury. Sci Rep. 2017;7:40112. Epub 2017/01/21. doi: 10.1038/srep40112. PubMed PMID: 28106051; PMCID: PMC5247752. [DOI] [PMC free article] [PubMed]
- 10.Ondruschka B, Schuch S, Pohlers D, Franke H, Dressler J. Acute phase response after fatal traumatic brain injury. Int J Legal Med. 2018;132(2):531-539. Epub 2018/01/08. doi: 10.1007/s00414-017-1768-2. PubMed PMID: 29306988. [DOI] [PubMed]
- 11.Hazelton I, Yates A, Dale A, Roodselaar J, Akbar N, Ruitenberg MJ, Anthony DC, Couch Y. Exacerbation of acute traumatic brain injury by circulating extracellular vesicles. J Neurotrauma. 2018;35(4):639-651. Epub 2017/11/19. doi: 10.1089/neu.2017.5049. PubMed PMID: 29149810. [DOI] [PubMed]
- 12.Boone DR, Weisz HA, Willey HE, Torres KEO, Falduto MT, Sinha M, Spratt H, Bolding IJ, Johnson KM, Parsley MA, DeWitt DS, Prough DS, Hellmich HL. Traumatic brain injury induces long-lasting changes in immune and regenerative signaling. PLoS One. 2019;14(4):e0214741. Epub 2019/04/04. doi: 10.1371/journal.pone.0214741. PubMed PMID: 30943276; PMCID: PMC6447179 not alter the authors' adherence to PLOS ONE policies on sharing data and materials. [DOI] [PMC free article] [PubMed]
- 13.Soriano S, Moffet B, Wicker E, Villapol S. Serum amyloid A is expressed in the brain after traumatic brain injury in a sex-dependent manner. Cell Mol Neurobiol. 2020. Epub 2020/02/16. doi: 10.1007/s10571-020-00808-3. PubMed PMID: 32060858. [DOI] [PMC free article] [PubMed]
- 14.Mukherjee S, Zeitouni S, Cavarsan CF, Shapiro LA. Increased seizure susceptibility in mice 30 days after fluid percussion injury. Front Neurol. 2013;4:28. Epub 2013/03/23. doi: 10.3389/fneur.2013.00028. PubMed PMID: 23519723; PMCID: PMC3604640. [DOI] [PMC free article] [PubMed]
- 15.Karve IP, Taylor JM, Crack PJ. The contribution of astrocytes and microglia to traumatic brain injury. Br J Pharmacol. 2016;173(4):692-702. Epub 2015/03/11. doi: 10.1111/bph.13125. PubMed PMID: 25752446; PMCID: PMC4742296. [DOI] [PMC free article] [PubMed]
- 16.Morganti-Kossmann MC, Satgunaseelan L, Bye N, Kossmann T. Modulation of immune response by head injury. Injury. 2007;38(12):1392-1400. Epub 2007/12/01. doi: 10.1016/j.injury.2007.10.005. PubMed PMID: 18048036. [DOI] [PubMed]
- 17.Witcher KG, Bray CE, Dziabis JE, McKim DB, Benner BN, Rowe RK, Kokiko-Cochran ON, Popovich PG, Lifshitz J, Eiferman DS, Godbout JP. Traumatic brain injury-induced neuronal damage in the somatosensory cortex causes formation of rod-shaped microglia that promote astrogliosis and persistent neuroinflammation. Glia. 2018;66(12):2719-2736. Epub 2018/11/01. doi: 10.1002/glia.23523. PubMed PMID: 30378170. [DOI] [PMC free article] [PubMed]
- 18.Wofford KL, Harris JP, Browne KD, Brown DP, Grovola MR, Mietus CJ, Wolf JA, Duda JE, Putt ME, Spiller KL, Cullen DK. Rapid neuroinflammatory response localized to injured neurons after diffuse traumatic brain injury in swine. Exp Neurol. 2017;290:85-94. Epub 2017/01/14. doi: 10.1016/j.expneurol.2017.01.004. PubMed PMID: 28081963; PMCID: PMC5529036. [DOI] [PMC free article] [PubMed]
- 19.Kumar RG, Diamond ML, Boles JA, Berger RP, Tisherman SA, Kochanek PM, Wagner AK. Acute CSF interleukin-6 trajectories after TBI: associations with neuroinflammation, polytrauma, and outcome. Brain Behav Immun. 2015;45:253-262. Epub 2015/01/04. doi: 10.1016/j.bbi.2014.12.021. PubMed PMID: 25555531. [DOI] [PubMed]
- 20.Helmy A, Carpenter KL, Menon DK, Pickard JD, Hutchinson PJ. The cytokine response to human traumatic brain injury: temporal profiles and evidence for cerebral parenchymal production. J Cereb Blood Flow Metab. 2011;31(2):658-70. Epub 2010/08/19. doi: 10.1038/jcbfm.2010.142. PubMed PMID: 20717122; PMCID: PMC3049520. [DOI] [PMC free article] [PubMed]
- 21.Penfield W. The physiology of epilepsy. Adv Neurol. 1975;8:1-9. Epub 1975/01/01. PubMed PMID: 804229. [PubMed]
- 22.Annegers J. In: The epidemiology of epilepsy. In: The treatment of epilepsy: principles and practice. 3. Wyllie E, editor. Philadelphia: Lippincott Williams; 2001. [Google Scholar]
- 23.Garga N, DH L. Posttraumatic Epilepsy: A major problem in desperate need of major advances. Epilepsy Currents. 2006;6(1):1–5. doi: 10.1111/j.1535-7511.2005.00083.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mani J, Barry E. Posttraumatic epilepsy. In Wyllie E, Gupta A, Lachhwani DK The treatment of epilepsy: principles and practice Hagerstown, MD: Lippincott Williams & Wilkins 2006:521-4.
- 25.Rasmussen MK. [Vaccination against yellow fever]. Ugeskr Laeger. 1992;154(4):215-216. Epub 1992/01/20. PubMed PMID: 1736453. [PubMed]
- 26.Hauser WA, Annegers JF, Kurland LT. Incidence of epilepsy and unprovoked seizures in Rochester, Minnesota: 1935-1984. Epilepsia. 1993;34(3):453-468. Epub 1993/05/01. PubMed PMID: 8504780. [DOI] [PubMed]
- 27.Annegers JF, Hauser WA, Coan SP, Rocca WA. A population-based study of seizures after traumatic brain injuries. N Engl J Med. 1998;338(1):20-24. Epub 1998/01/01. doi: 10.1056/NEJM199801013380104. PubMed PMID: 9414327. [DOI] [PubMed]
- 28.Salazar AM, Jabbari B, Vance SC, Grafman J, Amin D, Dillon JD. Epilepsy after penetrating head injury. I. Clinical correlates: a report of the Vietnam Head Injury Study. Neurology. 1985;35(10):1406-1414. Epub 1985/10/01. doi: 10.1212/wnl.35.10.1406. PubMed PMID: 3929158. [DOI] [PubMed]
- 29.Caveness WF. Epilepsy, a product of trauma in our time. Epilepsia. 1976;17(2):207-215. Epub 1976/06/01. PubMed PMID: 820549. [DOI] [PubMed]
- 30.Caveness WF, Walker AE, Ascroft PB. Incidence of posttraumatic epilepsy in Korean veterans as compared with those from World War I and World War II. J Neurosurg. 1962;19:122-129. Epub 1962/02/01. doi: 10.3171/jns.1962.19.2.0122. PubMed PMID: 13877492. [DOI] [PubMed]
- 31.Agrawal A, Timothy J, Pandit L, Manju M. Post-traumatic epilepsy: an overview. Clin Neurol Neurosurg. 2006;108(5):433-439. Epub 2005/10/18. doi: 10.1016/j.clineuro.2005.09.001. PubMed PMID: 16225987. [DOI] [PubMed]
- 32.Temkin NR, Dikmen SS, Wilensky AJ, Keihm J, Chabal S, Winn HR. A randomized, double-blind study of phenytoin for the prevention of post-traumatic seizures. N Engl J Med. 1990;323(8):497-502. Epub 1990/08/23. doi: 10.1056/NEJM199008233230801. PubMed PMID: 2115976. [DOI] [PubMed]
- 33.Beghi E. Overview of studies to prevent posttraumatic epilepsy. Epilepsia. 2003;44(s10):21-26. Epub 2003/09/27. PubMed PMID: 14511391. [DOI] [PubMed]
- 34.Teasell R, Bayona N, Lippert C, Villamere J, Hellings C. Post-traumatic seizure disorder following acquired brain injury. Brain Inj. 2007;21(2):201-214. Epub 2007/03/17. doi: 10.1080/02699050701201854. PubMed PMID: 17364531. [DOI] [PubMed]
- 35.Wang F, Wang X, Shapiro LA, Cotrina ML, Liu W, Wang EW, Gu S, Wang W, He X, Nedergaard M, Huang JH. NKCC1 up-regulation contributes to early post-traumatic seizures and increased post-traumatic seizure susceptibility. Brain Struct Funct. 2017;222(3):1543-56. Epub 2016/09/03. doi: 10.1007/s00429-016-1292-z. PubMed PMID: 27586142; PMCID: PMC5368191. [DOI] [PMC free article] [PubMed]
- 36.Irimia A, Van Horn JD. Epileptogenic focus localization in treatment-resistant post-traumatic epilepsy. J Clin Neurosci. 2015;22(4):627-31. Epub 2014/12/30. doi: 10.1016/j.jocn.2014.09.019. PubMed PMID: 25542591; PMCID: PMC4380645. [DOI] [PMC free article] [PubMed]
- 37.Mootz W, Schondorf J, Hubers H. [Fluorescence microscopy demonstration of histamine in the mast cells of the human tonsil]. Z Laryngol Rhinol Otol. 1971;50(2):140-145. Epub 1971/02/01. PubMed PMID: 5542591. [PubMed]
- 38.Kettenmann H, Verkhratsky A. Neuroglia: the 150 years after. Trends Neurosci. 2008;31(12):653-659. Epub 2008/10/24. doi: 10.1016/j.tins.2008.09.003. PubMed PMID: 18945498. [DOI] [PubMed]
- 39.Verkhratsky A, Ho MS, Vardjan N, Zorec R, Parpura V. General pathophysiology of Astroglia. Adv Exp Med Biol. 2019;1175:149-179. Epub 2019/10/05. doi: 10.1007/978-981-13-9913-8_7. PubMed PMID: 31583588. [DOI] [PMC free article] [PubMed]
- 40.Oberheim NA, Takano T, Han X, He W, Lin JH, Wang F, Xu Q, Wyatt JD, Pilcher W, Ojemann JG, Ransom BR, Goldman SA, Nedergaard M. Uniquely hominid features of adult human astrocytes. J Neurosci. 2009;29(10):3276-87. Epub 2009/03/13. doi: 10.1523/JNEUROSCI.4707-08.2009. PubMed PMID: 19279265; PMCID: PMC2819812. [DOI] [PMC free article] [PubMed]
- 41.Halassa MM, Haydon PG. Integrated brain circuits: astrocytic networks modulate neuronal activity and behavior. Annu Rev Physiol. 2010;72:335-55. Epub 2010/02/13. doi: 10.1146/annurev-physiol-021909-135843. PubMed PMID: 20148679; PMCID: PMC3117429. [DOI] [PMC free article] [PubMed]
- 42.Araque A, Carmignoto G, Haydon PG, Oliet SH, Robitaille R, Volterra A. Gliotransmitters travel in time and space. Neuron. 2014;81(4):728-39. Epub 2014/02/25. doi: 10.1016/j.neuron.2014.02.007. PubMed PMID: 24559669; PMCID: PMC4107238. [DOI] [PMC free article] [PubMed]
- 43.Weber B, Barros LF. The Astrocyte: powerhouse and recycling center. Cold Spring Harb Perspect Biol. 2015;7(12). Epub 2015/02/15. doi: 10.1101/cshperspect.a020396. PubMed PMID: 25680832; PMCID: PMC4665076. [DOI] [PMC free article] [PubMed]
- 44.Ye ZC, Sontheimer H. Cytokine modulation of glial glutamate uptake: a possible involvement of nitric oxide. Neuroreport. 1996;7(13):2181-2185. Epub 1996/09/02. doi: 10.1097/00001756-199609020-00025. PubMed PMID: 8930985. [DOI] [PubMed]
- 45.Zhu G, Okada M, Yoshida S, Mori F, Ueno S, Wakabayashi K, Kaneko S. Effects of interleukin-1beta on hippocampal glutamate and GABA releases associated with Ca2+-induced Ca2+ releasing systems. Epilepsy Res. 2006;71(2-3):107-116. Epub 2006/06/30. doi: 10.1016/j.eplepsyres.2006.05.017. PubMed PMID: 16806825. [DOI] [PubMed]
- 46.Hu S, Sheng WS, Ehrlich LC, Peterson PK, Chao CC. Cytokine effects on glutamate uptake by human astrocytes. Neuroimmunomodulation. 2000;7(3):153-159. Epub 2000/04/08. doi: 10.1159/000026433. PubMed PMID: 10754403. [DOI] [PubMed]
- 47.Payan H, Toga M, Berard-Badier M. The pathology of post-traumatic epilepsies. Epilepsia. 1970;11(1):81-94. Epub 1970/03/01. doi: 10.1111/j.1528-1157.1970.tb03869.x. PubMed PMID: 4987163. [DOI] [PubMed]
- 48.Oehmichen M, Walter T, Meissner C, Friedrich HJ. Time course of cortical hemorrhages after closed traumatic brain injury: statistical analysis of posttraumatic histomorphological alterations. J Neurotrauma. 2003;20(1):87-103. Epub 2003/03/05. doi: 10.1089/08977150360517218. PubMed PMID: 12614591. [DOI] [PubMed]
- 49.Pelinka LE, Kroepfl A, Leixnering M, Buchinger W, Raabe A, Redl H. GFAP versus S100B in serum after traumatic brain injury: relationship to brain damage and outcome. J Neurotrauma. 2004;21(11):1553-1561. Epub 2005/02/03. doi: 10.1089/neu.2004.21.1553. PubMed PMID: 15684648. [DOI] [PubMed]
- 50.Swartz BE, Houser CR, Tomiyasu U, Walsh GO, DeSalles A, Rich JR, Delgado-Escueta A. Hippocampal cell loss in posttraumatic human epilepsy. Epilepsia. 2006;47(8):1373-1382. Epub 2006/08/23. doi: 10.1111/j.1528-1167.2006.00602.x. PubMed PMID: 16922884. [DOI] [PubMed]
- 51.van Landeghem FK, Weiss T, Oehmichen M, von Deimling A. Decreased expression of glutamate transporters in astrocytes after human traumatic brain injury. J Neurotrauma. 2006;23(10):1518-1528. Epub 2006/10/06. doi: 10.1089/neu.2006.23.1518. PubMed PMID: 17020486. [DOI] [PubMed]
- 52.Brooks DM, Patel SA, Wohlgehagen ED, Semmens EO, Pearce A, Sorich EA, Rau TF. Multiple mild traumatic brain injury in the rat produces persistent pathological alterations in the brain. Exp Neurol. 2017;297:62-72. Epub 2017/08/02. doi: 10.1016/j.expneurol.2017.07.015. PubMed PMID: 28756201. [DOI] [PubMed]
- 53.Domowicz M, Wadlington NL, Henry JG, Diaz K, Munoz MJ, Schwartz NB. Glial cell responses in a murine multifactorial perinatal brain injury model. Brain Res. 2018;1681:52-63. Epub 2017/12/25. doi: 10.1016/j.brainres.2017.12.020. PubMed PMID: 29274879; PMCID: PMC5780221. [DOI] [PMC free article] [PubMed]
- 54.Bye N, Carron S, Han X, Agyapomaa D, Ng SY, Yan E, Rosenfeld JV, Morganti-Kossmann MC. Neurogenesis and glial proliferation are stimulated following diffuse traumatic brain injury in adult rats. J Neurosci Res. 2011;89(7):986-1000. Epub 2011/04/14. doi: 10.1002/jnr.22635. PubMed PMID: 21488090. [DOI] [PubMed]
- 55.Budde MD, Janes L, Gold E, Turtzo LC, Frank JA. The contribution of gliosis to diffusion tensor anisotropy and tractography following traumatic brain injury: validation in the rat using Fourier analysis of stained tissue sections. Brain. 2011;134(Pt 8):2248-60. Epub 2011/07/19. doi: 10.1093/brain/awr161. PubMed PMID: 21764818; PMCID: PMC3155707. [DOI] [PMC free article] [PubMed]
- 56.Kasturi BS, Stein DG. Traumatic brain injury causes long-term reduction in serum growth hormone and persistent astrocytosis in the cortico-hypothalamo-pituitary axis of adult male rats. J Neurotrauma. 2009;26(8):1315-1324. Epub 2009/03/26. doi: 10.1089/neu.2008-0751. PubMed PMID: 19317601; PMCID: PMC2850256. [DOI] [PMC free article] [PubMed]
- 57.Carbonell WS, Grady MS. Regional and temporal characterization of neuronal, glial, and axonal response after traumatic brain injury in the mouse. Acta Neuropathol. 1999;98(4):396-406. Epub 1999/09/29. doi: 10.1007/s004010051100. PubMed PMID: 10502046. [DOI] [PubMed]
- 58.Medical gallery of Blausen Medical 2014. WikiJournal of Medicine. 2014;1(2). doi: 10.15347/wjm/2014.010.
- 59.Hinterkeuser S, Schroder W, Hager G, Seifert G, Blumcke I, Elger CE, Schramm J, Steinhauser C. Astrocytes in the hippocampus of patients with temporal lobe epilepsy display changes in potassium conductances. Eur J Neurosci. 2000;12(6):2087-2096. Epub 2000/07/11. PubMed PMID: 10886348. [DOI] [PubMed]
- 60.Schroder W, Hinterkeuser S, Seifert G, Schramm J, Jabs R, Wilkin GP, Steinhauser C. Functional and molecular properties of human astrocytes in acute hippocampal slices obtained from patients with temporal lobe epilepsy. Epilepsia. 2000;41 Suppl 6:S181-S184. Epub 2000/09/22. PubMed PMID: 10999541. [DOI] [PubMed]
- 61.Bedner P, Dupper A, Huttmann K, Muller J, Herde MK, Dublin P, Deshpande T, Schramm J, Haussler U, Haas CA, Henneberger C, Theis M, Steinhauser C. Astrocyte uncoupling as a cause of human temporal lobe epilepsy. Brain. 2015;138(Pt 5):1208-22. Epub 2015/03/15. doi: 10.1093/brain/awv067. PubMed PMID: 25765328; PMCID: PMC5963418. [DOI] [PMC free article] [PubMed]
- 62.Ding S, Fellin T, Zhu Y, Lee SY, Auberson YP, Meaney DF, Coulter DA, Carmignoto G, Haydon PG. Enhanced astrocytic Ca2+ signals contribute to neuronal excitotoxicity after status epilepticus. J Neurosci. 2007;27(40):10674-84. Epub 2007/10/05. doi: 10.1523/JNEUROSCI.2001-07.2007. PubMed PMID: 17913901; PMCID: PMC2917229. [DOI] [PMC free article] [PubMed]
- 63.Vargas-Sanchez K, Mogilevskaya M, Rodriguez-Perez J, Rubiano MG, Javela JJ, Gonzalez-Reyes RE. Astroglial role in the pathophysiology of status epilepticus: an overview. Oncotarget. 2018;9(42):26954-76. Epub 2018/06/22. doi: 10.18632/oncotarget.25485. PubMed PMID: 29928494; PMCID: PMC6003549. [DOI] [PMC free article] [PubMed]
- 64.Sun L, Gao J, Zhao M, Cui J, Li Y, Yang X, Jing X, Wu Z. A novel cognitive impairment mechanism that astrocytic p-connexin 43 promotes neuronic autophagy via activation of P2X7R and down-regulation of GLT-1 expression in the hippocampus following traumatic brain injury in rats. Behav Brain Res. 2015;291:315-324. Epub 2015/06/03. doi: 10.1016/j.bbr.2015.05.049. PubMed PMID: 26031379. [DOI] [PubMed]
- 65.Homkajorn B, Sims NR, Muyderman H. Connexin 43 regulates astrocytic migration and proliferation in response to injury. Neurosci Lett. 2010;486(3):197-201. Epub 2010/09/28. doi: 10.1016/j.neulet.2010.09.051. PubMed PMID: 20869426. [DOI] [PubMed]
- 66.Walrave L, Pierre A, Albertini G, Aourz N, De Bundel D, Van Eeckhaut A, Vinken M, Giaume C, Leybaert L, Smolders I. Inhibition of astroglial connexin43 hemichannels with TAT-Gap19 exerts anticonvulsant effects in rodents. Glia. 2018;66(8):1788-1804. Epub 2018/04/24. doi: 10.1002/glia.23341. PubMed PMID: 29683209. [DOI] [PubMed]
- 67.Abudara V, Bechberger J, Freitas-Andrade M, De Bock M, Wang N, Bultynck G, Naus CC, Leybaert L, Giaume C. The connexin43 mimetic peptide Gap19 inhibits hemichannels without altering gap junctional communication in astrocytes. Front Cell Neurosci. 2014;8:306. Epub 2014/11/07. doi: 10.3389/fncel.2014.00306. PubMed PMID: 25374505; PMCID: PMC4204617. [DOI] [PMC free article] [PubMed]
- 68.Lu DC, Zador Z, Yao J, Fazlollahi F, Manley GT. Aquaporin-4 reduces post-traumatic seizure susceptibility by promoting astrocytic glial scar formation in mice. J Neurotrauma. 2011. Epub 2011/09/24. doi: 10.1089/neu.2011.2114. PubMed PMID: 21939392. [DOI] [PMC free article] [PubMed]
- 69.Binder DK, Oshio K, Ma T, Verkman AS, Manley GT. Increased seizure threshold in mice lacking aquaporin-4 water channels. Neuroreport. 2004;15(2):259-262. Epub 2004/04/13. PubMed PMID: 15076748. [DOI] [PubMed]
- 70.Shapiro LA, Korn MJ, Ribak CE. Newly generated dentate granule cells from epileptic rats exhibit elongated hilar basal dendrites that align along GFAP-immunolabeled processes. Neuroscience. 2005;136(3):823-831. Epub 2005/12/14. doi: 10.1016/j.neuroscience.2005.03.059. PubMed PMID: 16344154. [DOI] [PubMed]
- 71.Shapiro LA, Ribak CE. Newly born dentate granule neurons after pilocarpine-induced epilepsy have hilar basal dendrites with immature synapses. Epilepsy Res. 2006;69(1):53-66. Epub 2006/02/17. doi: 10.1016/j.eplepsyres.2005.12.003. PubMed PMID: 16480853. [DOI] [PubMed]
- 72.Shapiro LA, Figueroa-Aragon S, Ribak CE. Newly generated granule cells show rapid neuroplastic changes in the adult rat dentate gyrus during the first five days following pilocarpine-induced seizures. Eur J Neurosci. 2007;26(3):583-592. Epub 2007/08/10. doi: 10.1111/j.1460-9568.2007.05662.x. PubMed PMID: 17686039. [DOI] [PubMed]
- 73.Shapiro LA, Wang L, Ribak CE. Rapid astrocyte and microglial activation following pilocarpine-induced seizures in rats. Epilepsia. 2008;49 Suppl 2:33-41. Epub 2008/03/08. doi: 10.1111/j.1528-1167.2008.01491.x. PubMed PMID: 18226170. [DOI] [PubMed]
- 74.Shapiro LA, Ribak CE, Jessberger S. Structural changes for adult-born dentate granule cells after status epilepticus. Epilepsia. 2008;49 Suppl 5:13-18. Epub 2008/07/17. doi: 10.1111/j.1528-1167.2008.01633.x. PubMed PMID: 18522596. [DOI] [PubMed]
- 75.Ribak CE, Shapiro LA, Yan XX, Dashtipour K, Nadler JV, Obenaus A, Spigelman I, Buckmaster PS. Seizure-induced formation of basal dendrites on granule cells of the rodent dentate gyrus. In: th, Noebels JL, Avoli M, Rogawski MA, Olsen RW, Delgado-Escueta AV, editors. Jasper’s basic mechanisms of the epilepsies. Bethesda (MD) 2012. [PubMed]
- 76.Sanchez RM, Ribak CE, Shapiro LA. Synaptic connections of hilar basal dendrites of dentate granule cells in a neonatal hypoxia model of epilepsy. Epilepsia. 2012;53 Suppl 1:98-108. Epub 2012/05/25. doi: 10.1111/j.1528-1167.2012.03481.x. PubMed PMID: 22612814. [DOI] [PubMed]
- 77.Robinson C, Apgar C, Shapiro LA. Astrocyte hypertrophy contributes to aberrant neurogenesis after traumatic brain injury. Neural Plast. 2016;2016:1347987. Epub 2016/06/09. doi: 10.1155/2016/1347987. PubMed PMID: 27274873; PMCID: PMC4870378. [DOI] [PMC free article] [PubMed]
- 78.Shapiro LA. Altered hippocampal neurogenesis during the first 7 days after a fluid percussion traumatic brain injury. Cell Transplant. 2017;26(7):1314-8. Epub 2017/09/22. doi: 10.1177/0963689717714099. PubMed PMID: 28933222; PMCID: PMC5657729. [DOI] [PMC free article] [PubMed]
- 79.Shapiro LA, Ribak CE. Integration of newly born dentate granule cells into adult brains: hypotheses based on normal and epileptic rodents. Brain Res Brain Res Rev. 2005;48(1):43-56. Epub 2005/02/15. doi: 10.1016/j.brainresrev.2004.08.003. PubMed PMID: 15708627. [DOI] [PubMed]
- 80.Shapiro LA, Korn MJ, Shan Z, Ribak CE. GFAP-expressing radial glia-like cell bodies are involved in a one-to-one relationship with doublecortin-immunolabeled newborn neurons in the adult dentate gyrus. Brain Res. 2005;1040(1-2):81-91. Epub 2005/04/05. doi: 10.1016/j.brainres.2005.01.098. PubMed PMID: 15804429. [DOI] [PubMed]
- 81.Shapiro LA, Upadhyaya P, Ribak CE. Spatiotemporal profile of dendritic outgrowth from newly born granule cells in the adult rat dentate gyrus. Brain Res. 2007;1149:30-37. Epub 2006/08/15. doi: 10.1016/j.brainres.2006.07.032. PubMed PMID: 16904657. [DOI] [PubMed]
- 82.Ribak CE, Shapiro LA. Dendritic development of newly generated neurons in the adult brain. Brain Res Rev. 2007;55(2):390-4. Epub 2007/01/24. doi: 10.1016/j.brainresrev.2006.12.005. PubMed PMID: 17239443; PMCID: PMC2072906. [DOI] [PMC free article] [PubMed]
- 83.Villasana LE, Kim KN, Westbrook GL, Schnell E. Functional integration of adult-born hippocampal neurons after traumatic brain injury(1,2,3). eNeuro. 2015;2(5). Epub 2015/10/20. doi: 10.1523/ENEURO.0056-15.2015. PubMed PMID: 26478908; PMCID: PMC4603252. [DOI] [PMC free article] [PubMed]
- 84.Golarai G, Greenwood AC, Feeney DM, Connor JA. Physiological and structural evidence for hippocampal involvement in persistent seizure susceptibility after traumatic brain injury. J Neurosci. 2001;21(21):8523-37. Epub 2001/10/19. PubMed PMID: 11606641; PMCID: PMC6762822. [DOI] [PMC free article] [PubMed]
- 85.Guo D, Zeng L, Brody DL, Wong M. Rapamycin attenuates the development of posttraumatic epilepsy in a mouse model of traumatic brain injury. PLoS One. 2013;8(5):e64078. Epub 2013/05/22. doi: 10.1371/journal.pone.0064078. PubMed PMID: 23691153; PMCID: PMC3653881. [DOI] [PMC free article] [PubMed]
- 86.Kelly KM, Miller ER, Lepsveridze E, Kharlamov EA, McHedlishvili Z. Posttraumatic seizures and epilepsy in adult rats after controlled cortical impact. Epilepsy Res. 2015;117:104-116. Epub 2015/10/04. doi: 10.1016/j.eplepsyres.2015.09.009. PubMed PMID: 26432760. [DOI] [PubMed]
- 87.Pugh P, Adlaf E, Zhao CS, Markwardt S, Gavin C, Wadiche J, Overstreet-Wadiche L. Enhanced integration of newborn neurons after neonatal insults. Front Neurosci. 2011;5:45. Epub 2011/04/15. doi: 10.3389/fnins.2011.00045. PubMed PMID: 21490706; PMCID: PMC3070953. [DOI] [PMC free article] [PubMed]
- 88.Villasana LE, Westbrook GL, Schnell E. Neurologic impairment following closed head injury predicts post-traumatic neurogenesis. Exp Neurol. 2014;261:156-62. Epub 2014/05/28. doi: 10.1016/j.expneurol.2014.05.016. PubMed PMID: 24861442; PMCID: PMC4194237. [DOI] [PMC free article] [PubMed]
- 89.Ribak CE, Shapiro LA. Ultrastructure and synaptic connectivity of cell types in the adult rat dentate gyrus. Prog Brain Res. 2007;163:155-166. Epub 2007/09/04. doi: 10.1016/S0079-6123(07)63009-X. PubMed PMID: 17765717. [DOI] [PubMed]
- 90.Ribak CE, Tran PH, Spigelman I, Okazaki MM, Nadler JV. Status epilepticus-induced hilar basal dendrites on rodent granule cells contribute to recurrent excitatory circuitry. J Comp Neurol. 2000;428(2):240-253. Epub 2000/11/07. doi: 10.1002/1096-9861(20001211)428:2<240::aid-cne4>3.0.co;2-q. PubMed PMID: 11064364. [DOI] [PubMed]
- 91.Scharfman HE, Sollas AE, Berger RE, Goodman JH, Pierce JP. Perforant path activation of ectopic granule cells that are born after pilocarpine-induced seizures. Neuroscience. 2003;121(4):1017-1029. Epub 2003/10/29. doi: 10.1016/s0306-4522(03)00481-0. PubMed PMID: 14580952. [DOI] [PubMed]
- 92.Morgan RJ, Soltesz I. Nonrandom connectivity of the epileptic dentate gyrus predicts a major role for neuronal hubs in seizures. Proc Natl Acad Sci U S A. 2008;105(16):6179-84. Epub 2008/04/01. doi: 10.1073/pnas.0801372105. PubMed PMID: 18375756; PMCID: PMC2299224. [DOI] [PMC free article] [PubMed]
- 93.Ngwenya LB, Danzer SC. Impact of traumatic brain injury on neurogenesis. Front Neurosci. 2018;12:1014. Epub 2019/01/29. doi: 10.3389/fnins.2018.01014. PubMed PMID: 30686980; PMCID: PMC6333744. [DOI] [PMC free article] [PubMed]
- 94.Danzer SC. Contributions of adult-generated granule cells to hippocampal pathology in temporal lobe epilepsy: a neuronal bestiary. Brain Plast. 2018;3(2):169-81. Epub 2018/08/29. doi: 10.3233/BPL-170056. PubMed PMID: 30151341; PMCID: PMC6091048. [DOI] [PMC free article] [PubMed]
- 95.Danzer SC. Adult neurogenesis in the development of epilepsy. Epilepsy Curr. 2019;19(5):316-20. Epub 2019/08/15. doi: 10.1177/1535759719868186. PubMed PMID: 31409149; PMCID: PMC6864561. [DOI] [PMC free article] [PubMed]
- 96.Danzer SC. A Hit, a Hit-a very palpable hit: mild TBI and the development of epilepsy. Epilepsy Curr. 2019;19(4):261-3. Epub 2019/06/19. doi: 10.1177/1535759719854758. PubMed PMID: 31208231; PMCID: PMC6891830. [DOI] [PMC free article] [PubMed]
- 97.Arisi GM, Foresti ML, Katki K, Shapiro LA. Increased CCL2, CCL3, CCL5, and IL-1beta cytokine concentration in piriform cortex, hippocampus, and neocortex after pilocarpine-induced seizures. J Neuroinflammation. 2015;12:129. Epub 2015/07/03. doi: 10.1186/s12974-015-0347-z. PubMed PMID: 26133170; PMCID: PMC4509848. [DOI] [PMC free article] [PubMed]
- 98.Chang GQ, Karatayev O, Boorgu D, Leibowitz SF. Third ventricular injection of CCL2 in rat embryo stimulates CCL2/CCR2 neuroimmune system in neuroepithelial radial glia progenitor cells: relation to sexually dimorphic, stimulatory effects on peptide neurons in lateral hypothalamus. Neuroscience. 2020. Epub 2020/01/27. doi: 10.1016/j.neuroscience.2020.01.020. PubMed PMID: 31982472. [DOI] [PMC free article] [PubMed]
- 99.Liu XS, Zhang ZG, Zhang RL, Gregg SR, Wang L, Yier T, Chopp M. Chemokine ligand 2 (CCL2) induces migration and differentiation of subventricular zone cells after stroke. J Neurosci Res. 2007;85(10):2120-2125. Epub 2007/05/19. doi: 10.1002/jnr.21359. PubMed PMID: 17510981. [DOI] [PubMed]
- 100.Foresti ML, Arisi GM, Katki K, Montanez A, Sanchez RM, Shapiro LA. Chemokine CCL2 and its receptor CCR2 are increased in the hippocampus following pilocarpine-induced status epilepticus. J Neuroinflammation. 2009;6:40. Epub 2009/12/26. doi: 10.1186/1742-2094-6-40. PubMed PMID: 20034406; PMCID: PMC2804573. [DOI] [PMC free article] [PubMed]
- 101.Loane DJ, Kumar A. Microglia in the TBI brain: The good, the bad, and the dysregulated. Exp Neurol. 2016;275 Pt 3:316-27. Epub 2015/09/08. doi: 10.1016/j.expneurol.2015.08.018. PubMed PMID: 26342753; PMCID: PMC4689601. [DOI] [PMC free article] [PubMed]
- 102.Kim CC, Nakamura MC, Hsieh CL. Brain trauma elicits non-canonical macrophage activation states. J Neuroinflammation. 2016;13(1):117. Epub 2016/05/26. doi: 10.1186/s12974-016-0581-z. PubMed PMID: 27220367; PMCID: PMC4879757. [DOI] [PMC free article] [PubMed]
- 103.Loane DJ, Kumar A, Stoica BA, Cabatbat R, Faden AI. Progressive neurodegeneration after experimental brain trauma: association with chronic microglial activation. J Neuropathol Exp Neurol. 2014;73(1):14-29. Epub 2013/12/18. doi: 10.1097/NEN.0000000000000021. PubMed PMID: 24335533; PMCID: PMC4267248. [DOI] [PMC free article] [PubMed]
- 104.Beschorner R, Nguyen TD, Gozalan F, Pedal I, Mattern R, Schluesener HJ, Meyermann R, Schwab JM. CD14 expression by activated parenchymal microglia/macrophages and infiltrating monocytes following human traumatic brain injury. Acta Neuropathol. 2002;103(6):541-549. Epub 2002/05/16. doi: 10.1007/s00401-001-0503-7. PubMed PMID: 12012085. [DOI] [PubMed]
- 105.Engel S, Schluesener H, Mittelbronn M, Seid K, Adjodah D, Wehner HD, Meyermann R. Dynamics of microglial activation after human traumatic brain injury are revealed by delayed expression of macrophage-related proteins MRP8 and MRP14. Acta Neuropathol. 2000;100(3):313-322. Epub 2000/08/31. doi: 10.1007/s004019900172. PubMed PMID: 10965802. [DOI] [PubMed]
- 106.Ramlackhansingh AF, Brooks DJ, Greenwood RJ, Bose SK, Turkheimer FE, Kinnunen KM, Gentleman S, Heckemann RA, Gunanayagam K, Gelosa G, Sharp DJ. Inflammation after trauma: microglial activation and traumatic brain injury. Ann Neurol. 2011;70(3):374-383. Epub 2011/06/29. doi: 10.1002/ana.22455. PubMed PMID: 21710619. [DOI] [PubMed]
- 107.Ginhoux F, Lim S, Hoeffel G, Low D, Huber T. Origin and differentiation of microglia. Front Cell Neurosci. 2013;7:45. Epub 2013/04/26. doi: 10.3389/fncel.2013.00045. PubMed PMID: 23616747; PMCID: PMC3627983. [DOI] [PMC free article] [PubMed]
- 108.Ginhoux F, Prinz M. Origin of microglia: current concepts and past controversies. Cold Spring Harb Perspect Biol. 2015;7(8):a020537. Epub 2015/07/03. doi: 10.1101/cshperspect.a020537. PubMed PMID: 26134003; PMCID: PMC4526747. [DOI] [PMC free article] [PubMed]
- 109.Schafer DP, Lehrman EK, Kautzman AG, Koyama R, Mardinly AR, Yamasaki R, Ransohoff RM, Greenberg ME, Barres BA, Stevens B. Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron. 2012;74(4):691-705. Epub 2012/05/29. doi: 10.1016/j.neuron.2012.03.026. PubMed PMID: 22632727; PMCID: PMC3528177. [DOI] [PMC free article] [PubMed]
- 110.Walton NM, Sutter BM, Laywell ED, Levkoff LH, Kearns SM, Marshall GP, 2nd, Scheffler B, Steindler DA. Microglia instruct subventricular zone neurogenesis. Glia. 2006;54(8):815-825. Epub 2006/09/16. doi: 10.1002/glia.20419. PubMed PMID: 16977605. [DOI] [PubMed]
- 111.Morgan SC, Taylor DL, Pocock JM. Microglia release activators of neuronal proliferation mediated by activation of mitogen-activated protein kinase, phosphatidylinositol-3-kinase/Akt and delta-Notch signalling cascades. J Neurochem. 2004;90(1):89-101. Epub 2004/06/17. doi: 10.1111/j.1471-4159.2004.02461.x. PubMed PMID: 15198670. [DOI] [PubMed]
- 112.Mildner A, Schmidt H, Nitsche M, Merkler D, Hanisch UK, Mack M, Heikenwalder M, Bruck W, Priller J, Prinz M. Microglia in the adult brain arise from Ly-6ChiCCR2+ monocytes only under defined host conditions. Nat Neurosci. 2007;10(12):1544-1553. Epub 2007/11/21. doi: 10.1038/nn2015. PubMed PMID: 18026096. [DOI] [PubMed]
- 113.Klein P, Dingledine R, Aronica E, Bernard C, Blumcke I, Boison D, Brodie MJ, Brooks-Kayal AR, Engel J, Jr., Forcelli PA, Hirsch LJ, Kaminski RM, Klitgaard H, Kobow K, Lowenstein DH, Pearl PL, Pitkanen A, Puhakka N, Rogawski MA, Schmidt D, Sillanpaa M, Sloviter RS, Steinhauser C, Vezzani A, Walker MC, Loscher W. Commonalities in epileptogenic processes from different acute brain insults: do they translate? Epilepsia. 2018;59(1):37-66. Epub 2017/12/17. doi: 10.1111/epi.13965. PubMed PMID: 29247482; PMCID: PMC5993212. [DOI] [PMC free article] [PubMed]
- 114.Nimmerjahn A, Kirchhoff F, Helmchen F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science. 2005;308(5726):1314-1318. Epub 2005/04/16. doi: 10.1126/science.1110647. PubMed PMID: 15831717. [DOI] [PubMed]
- 115.Eyo UB, Dailey ME. Microglia: key elements in neural development, plasticity, and pathology. J Neuroimmune Pharmacol. 2013;8(3):494-509. Epub 2013/01/29. doi: 10.1007/s11481-013-9434-z. PubMed PMID: 23354784; PMCID: PMC3657325. [DOI] [PMC free article] [PubMed]
- 116.Town T, Nikolic V, Tan J. The microglial “activation” continuum: from innate to adaptive responses. J Neuroinflammation. 2005;2:24. Epub 2005/11/02. doi: 10.1186/1742-2094-2-24. PubMed PMID: 16259628; PMCID: PMC1298325. [DOI] [PMC free article] [PubMed]
- 117.Chhor V, Moretti R, Le Charpentier T, Sigaut S, Lebon S, Schwendimann L, Ore MV, Zuiani C, Milan V, Josserand J, Vontell R, Pansiot J, Degos V, Ikonomidou C, Titomanlio L, Hagberg H, Gressens P, Fleiss B. Role of microglia in a mouse model of paediatric traumatic brain injury. Brain Behav Immun. 2017;63:197-209. Epub 2016/11/08. doi: 10.1016/j.bbi.2016.11.001. PubMed PMID: 27818218; PMCID: PMC5441571. [DOI] [PMC free article] [PubMed]
- 118.Somera-Molina KC, Robin B, Somera CA, Anderson C, Stine C, Koh S, Behanna HA, Van Eldik LJ, Watterson DM, Wainwright MS. Glial activation links early-life seizures and long-term neurologic dysfunction: evidence using a small molecule inhibitor of proinflammatory cytokine upregulation. Epilepsia. 2007;48(9):1785-1800. Epub 2007/05/25. doi: 10.1111/j.1528-1167.2007.01135.x. PubMed PMID: 17521344. [DOI] [PubMed]
- 119.Lloyd E, Somera-Molina K, Van Eldik LJ, Watterson DM, Wainwright MS. Suppression of acute proinflammatory cytokine and chemokine upregulation by post-injury administration of a novel small molecule improves long-term neurologic outcome in a mouse model of traumatic brain injury. J Neuroinflammation. 2008;5:28. Epub 2008/07/02. doi: 10.1186/1742-2094-5-28. PubMed PMID: 18590543; PMCID: PMC2483713. [DOI] [PMC free article] [PubMed]
- 120.Hesam S, Khoshkholgh-Sima B, Pourbadie HG, Babapour V, Zendedel M, Sayyah M. Monophosphoryl lipid A and Pam3Cys prevent the increase in seizure susceptibility and epileptogenesis in rats undergoing traumatic brain injury. Neurochem Res. 2018;43(10):1978-1985. Epub 2018/09/03. doi: 10.1007/s11064-018-2619-3. PubMed PMID: 30173323. [DOI] [PubMed]
- 121.Eslami M, Ghanbari E, Sayyah M, Etemadi F, Choopani S, Soleimani M, Amiri Z, Hadjighassem M. Traumatic brain injury accelerates kindling epileptogenesis in rats. Neurol Res. 2016;38(3):269-274. Epub 2015/09/01. doi: 10.1179/1743132815Y.0000000086. PubMed PMID: 26315855. [DOI] [PubMed]
- 122.Hesam S, Khoshkholgh-Sima B, Pourbadie HG. Babapour V, Zendedel M, Sayyah M. Monophosphoryl lipid A and Pam3Cys prevent the increase in seizure susceptibility and epileptogenesis in rats undergoing traumatic brain injury. Neurochem Res. 2018;43(10):1978-85. Epub 2018/09/01. 10.1007/s11064-018-2619-3. PubMed PMID: 30173323. [DOI] [PubMed]
- 123.Yamamoto M, Kim M, Imai H, Itakura Y, Ohtsuki G. Microglia-triggered plasticity of intrinsic excitability modulates psychomotor behaviors in acute cerebellar inflammation. Cell Rep. 2019;28(11):2923-2938 e8. Epub 2019/09/12. doi: 10.1016/j.celrep.2019.07.078. PubMed PMID: 31509752. [DOI] [PubMed]
- 124.Cruz SL, Armenta-Resendiz M, Carranza-Aguilar CJ, Galvan EJ. Minocycline prevents neuronal hyperexcitability and neuroinflammation in medial prefrontal cortex, as well as memory impairment caused by repeated toluene inhalation in adolescent rats. Toxicol Appl Pharmacol. 2020;395:114980. Epub 2020/04/03. doi: 10.1016/j.taap.2020.114980. PubMed PMID: 32234516. [DOI] [PubMed]
- 125.Devinsky O, Vezzani A, Najjar S, De Lanerolle NC, Rogawski MA. Glia and epilepsy: excitability and inflammation. Trends Neurosci. 2013;36(3):174-184. Epub 2013/01/10. doi: 10.1016/j.tins.2012.11.008. PubMed PMID: 23298414. [DOI] [PubMed]
- 126.Eslami M, Sayyah M, Soleimani M, Alizadeh L, Hadjighassem M. Lipopolysaccharide preconditioning prevents acceleration of kindling epileptogenesis induced by traumatic brain injury. J Neuroimmunol. 2015;289:143-151. Epub 2015/12/01. doi: 10.1016/j.jneuroim.2015.11.003. PubMed PMID: 26616884. [DOI] [PubMed]
- 127.Kigerl KA, de Rivero Vaccari JP, Dietrich WD, Popovich PG, Keane RW. Pattern recognition receptors and central nervous system repair. Exp Neurol. 2014;258:5-16. Epub 2014/07/16. doi: 10.1016/j.expneurol.2014.01.001. PubMed PMID: 25017883; PMCID: PMC4974939. [DOI] [PMC free article] [PubMed]
- 128.Yang H, Antoine DJ, Andersson U, Tracey KJ. The many faces of HMGB1: molecular structure-functional activity in inflammation, apoptosis, and chemotaxis. J Leukoc Biol. 2013;93(6):865-73. Epub 2013/03/01. doi: 10.1189/jlb.1212662. PubMed PMID: 23446148; PMCID: PMC4051189. [DOI] [PMC free article] [PubMed]
- 129.Maroso M, Balosso S, Ravizza T, Liu J, Aronica E, Iyer AM, Rossetti C, Molteni M, Casalgrandi M, Manfredi AA, Bianchi ME, Vezzani A. Toll-like receptor 4 and high-mobility group box-1 are involved in ictogenesis and can be targeted to reduce seizures. Nat Med. 2010;16(4):413-419. Epub 2010/03/30. doi: 10.1038/nm.2127. PubMed PMID: 20348922. [DOI] [PubMed]
- 130.Morganti JM, Riparip LK, Rosi S. Call off the dog(ma): M1/M2 polarization is concurrent following traumatic brain injury. PLoS One. 2016;11(1):e0148001. Epub 2016/01/26. doi: 10.1371/journal.pone.0148001. PubMed PMID: 26808663; PMCID: PMC4726527. [DOI] [PMC free article] [PubMed]
- 131.Kumar A, Barrett JP, Alvarez-Croda DM, Stoica BA, Faden AI, Loane DJ. NOX2 drives M1-like microglial/macrophage activation and neurodegeneration following experimental traumatic brain injury. Brain Behav Immun. 2016;58:291-309. Epub 2016/08/02. doi: 10.1016/j.bbi.2016.07.158. PubMed PMID: 27477920; PMCID: PMC5067217. [DOI] [PMC free article] [PubMed]
- 132.Cherry JD, Olschowka JA, O'Banion MK. Neuroinflammation and M2 microglia: the good, the bad, and the inflamed. J Neuroinflammation. 2014;11:98. Epub 2014/06/04. doi: 10.1186/1742-2094-11-98. PubMed PMID: 24889886; PMCID: PMC4060849. [DOI] [PMC free article] [PubMed]
- 133.Liddelow SA, Guttenplan KA, Clarke LE, Bennett FC, Bohlen CJ, Schirmer L, Bennett ML, Munch AE, Chung WS, Peterson TC, Wilton DK, Frouin A, Napier BA, Panicker N, Kumar M, Buckwalter MS, Rowitch DH, Dawson VL, Dawson TM, Stevens B, Barres BA. Neurotoxic reactive astrocytes are induced by activated microglia. Nature. 2017;541(7638):481-7. Epub 2017/01/19. doi: 10.1038/nature21029. PubMed PMID: 28099414; PMCID: PMC5404890. [DOI] [PMC free article] [PubMed]
- 134.Mukherjee S, Katki K, Arisi GM, Foresti ML, Shapiro LA. Early TBI-induced cytokine alterations are similarly detected by two distinct methods of multiplex assay. Front Mol Neurosci. 2011;4:21. Epub 2011/09/29. doi: 10.3389/fnmol.2011.00021. PubMed PMID: 21954376; PMCID: PMC3174383. [DOI] [PMC free article] [PubMed]
- 135.Morganti-Kossmann MC, Semple BD, Hellewell SC, Bye N, Ziebell JM. The complexity of neuroinflammation consequent to traumatic brain injury: from research evidence to potential treatments. Acta Neuropathol. 2019;137(5):731-755. Epub 2018/12/12. doi: 10.1007/s00401-018-1944-6. PubMed PMID: 30535946. [DOI] [PubMed]
- 136.Morganti-Kossman MC, Lenzlinger PM, Hans V, Stahel P, Csuka E, Ammann E, Stocker R, Trentz O, Kossmann T. Production of cytokines following brain injury: beneficial and deleterious for the damaged tissue. Mol Psychiatry. 1997;2(2):133-136. Epub 1997/03/01. PubMed PMID: 9106236. [DOI] [PubMed]
- 137.Webster KM, Sun M, Crack P, O'Brien TJ, Shultz SR, Semple BD. Inflammation in epileptogenesis after traumatic brain injury. J Neuroinflammation. 2017;14(1):10. Epub 2017/01/15. doi: 10.1186/s12974-016-0786-1. PubMed PMID: 28086980; PMCID: PMC5237206. [DOI] [PMC free article] [PubMed]
- 138.Viviani B, Gardoni F, Marinovich M. Cytokines and neuronal ion channels in health and disease. Int Rev Neurobiol. 2007;82:247-263. Epub 2007/08/07. doi: 10.1016/S0074-7742(07)82013-7. PubMed PMID: 17678965. [DOI] [PubMed]
- 139.Semple BD, O'Brien TJ, Gimlin K, Wright DK, Kim SE, Casillas-Espinosa PM, Webster KM, Petrou S, Noble-Haeusslein LJ. Interleukin-1 receptor in seizure susceptibility after traumatic injury to the pediatric brain. J Neurosci. 2017;37(33):7864-77. Epub 2017/07/21. doi: 10.1523/JNEUROSCI.0982-17.2017. PubMed PMID: 28724747; PMCID: PMC5559762. [DOI] [PMC free article] [PubMed]
- 140.Helmy A, Guilfoyle MR, Carpenter KLH, Pickard JD, Menon DK, Hutchinson PJ. Recombinant human interleukin-1 receptor antagonist promotes M1 microglia biased cytokines and chemokines following human traumatic brain injury. J Cereb Blood Flow Metab. 2016;36(8):1434-48. Epub 2015/12/15. doi: 10.1177/0271678X15620204. PubMed PMID: 26661249; PMCID: PMC4976751. [DOI] [PMC free article] [PubMed]
- 141.Bertani I, Iori V, Trusel M, Maroso M, Foray C, Mantovani S, Tonini R, Vezzani A, Chiesa R. Inhibition of IL-1beta signaling normalizes NMDA-dependent neurotransmission and reduces seizure susceptibility in a mouse model of Creutzfeldt-Jakob disease. J Neurosci. 2017;37(43):10278-10289. Epub 2017/09/20. doi: 10.1523/JNEUROSCI.1301-17.2017. PubMed PMID: 28924012; PMCID: PMC6596629. [DOI] [PMC free article] [PubMed]
- 142.Vezzani A, Friedman A, Dingledine RJ. The role of inflammation in epileptogenesis. Neuropharmacology. 2013;69:16-24. Epub 2012/04/24. doi: 10.1016/j.neuropharm.2012.04.004. PubMed PMID: 22521336; PMCID: PMC3447120. [DOI] [PMC free article] [PubMed]
- 143.Sun Y, Ma J, Li D, Li P, Zhou X, Li Y, He Z, Qin L, Liang L, Luo X. Interleukin-10 inhibits interleukin-1beta production and inflammasome activation of microglia in epileptic seizures. J Neuroinflammation. 2019;16(1):66. Epub 2019/03/30. doi: 10.1186/s12974-019-1452-1. PubMed PMID: 30922332; PMCID: PMC6437919. [DOI] [PMC free article] [PubMed]
- 144.Bar-Klein G, Cacheaux LP, Kamintsky L, Prager O, Weissberg I, Schoknecht K, Cheng P, Kim SY, Wood L, Heinemann U, Kaufer D, Friedman A. Losartan prevents acquired epilepsy via TGF-beta signaling suppression. Ann Neurol. 2014;75(6):864-875. Epub 2014/03/25. doi: 10.1002/ana.24147. PubMed PMID: 24659129; PMCID: PMC4077937. [DOI] [PMC free article] [PubMed]
- 145.Weissberg I, Wood L, Kamintsky L, Vazquez O, Milikovsky DZ, Alexander A, Oppenheim H, Ardizzone C, Becker A, Frigerio F, Vezzani A, Buckwalter MS, Huguenard JR, Friedman A, Kaufer D. Albumin induces excitatory synaptogenesis through astrocytic TGF-beta/ALK5 signaling in a model of acquired epilepsy following blood-brain barrier dysfunction. Neurobiol Dis. 2015;78:115-125. Epub 2015/04/04. doi: 10.1016/j.nbd.2015.02.029. PubMed PMID: 25836421; PMCID: PMC4426044. [DOI] [PMC free article] [PubMed]
- 146.Cacheaux LP, Ivens S, David Y, Lakhter AJ, Bar-Klein G, Shapira M, Heinemann U, Friedman A, Kaufer D. Transcriptome profiling reveals TGF-beta signaling involvement in epileptogenesis. J Neurosci. 2009;29(28):8927-8935. Epub 2009/07/17. doi: 10.1523/JNEUROSCI.0430-09.2009. PubMed PMID: 19605630; PMCID: PMC2875073. [DOI] [PMC free article] [PubMed]
- 147.Gill J, Motamedi V, Osier N, Dell K, Arcurio L, Carr W, Walker P, Ahlers S, Lopresti M, Yarnell A. Moderate blast exposure results in increased IL-6 and TNFalpha in peripheral blood. Brain Behav Immun. 2017;65:90-94. Epub 2017/02/25. doi: 10.1016/j.bbi.2017.02.015. PubMed PMID: 28232173; PMCID: PMC5537025. [DOI] [PMC free article] [PubMed]
- 148.Liimatainen S, Fallah M, Kharazmi E, Peltola M, Peltola J. Interleukin-6 levels are increased in temporal lobe epilepsy but not in extra-temporal lobe epilepsy. J Neurol. 2009;256(5):796-802. Epub 2009/03/03. doi: 10.1007/s00415-009-5021-x. PubMed PMID: 19252806. [DOI] [PubMed]
- 149.Ishikawa N, Kobayashi Y, Fujii Y, Kobayashi M. Increased interleukin-6 and high-sensitivity C-reactive protein levels in pediatric epilepsy patients with frequent, refractory generalized motor seizures. Seizure. 2015;25:136-140. Epub 2014/12/03. doi: 10.1016/j.seizure.2014.10.007. PubMed PMID: 25455727. [DOI] [PubMed]
- 150.van den Munckhof B, de Vries EE, Braun KP, Boss HM, Willemsen MA, van Royen-Kerkhof A, de Jager W, Jansen FE. Serum inflammatory mediators correlate with disease activity in electrical status epilepticus in sleep (ESES) syndrome. Epilepsia. 2016;57(2):e45-e50. Epub 2015/12/17. doi: 10.1111/epi.13274. PubMed PMID: 26666401. [DOI] [PubMed]
- 151.Azab SF, Abdalhady MA, Ali A, Amin EK, Sarhan DT, Elhindawy EM, Almalky MA, Elhewala AA, Salam MM, Hashem MI, Soliman AA, Akeel NE, Abdellatif SH, Ismail SM, Elsamad NA, Arafat MS, Rass AA, Basset MA. Interleukin-6 gene polymorphisms in Egyptian children with febrile seizures: a case-control study. Ital J Pediatr. 2016;42:31. Epub 2016/03/11. doi: 10.1186/s13052-016-0244-9. PubMed PMID: 26960986; PMCID: PMC4784363. [DOI] [PMC free article] [PubMed]
- 152.Shahrokhi A, Zare-Shahabadi A, Soltani S, Ashrafi MR, Zoghi S, Hosseini SA, Heidari M, Yaghmaei B, Pourakbari B, Rezaei N. Association of IL6 single nucleotide polymorphisms with febrile seizures. J Neurol Sci. 2014;342(1-2):25-28. Epub 2014/05/20. doi: 10.1016/j.jns.2014.04.003. PubMed PMID: 24834995. [DOI] [PubMed]
- 153.Chou IC, Lin WD, Wang CH, Tsai CH, Li TC, Tsai FJ. Interleukin (IL)-1beta, IL-1 receptor antagonist, IL-6, IL-8, IL-10, and tumor necrosis factor alpha gene polymorphisms in patients with febrile seizures. J Clin Lab Anal. 2010;24(3):154-159. Epub 2010/05/21. doi: 10.1002/jcla.20374. PubMed PMID: 20486195. [DOI] [PMC free article] [PubMed]
- 154.Samland H, Huitron-Resendiz S, Masliah E, Criado J, Henriksen SJ, Campbell IL. Profound increase in sensitivity to glutamatergic- but not cholinergic agonist-induced seizures in transgenic mice with astrocyte production of IL-6. J Neurosci Res. 2003;73(2):176-187. Epub 2003/07/02. doi: 10.1002/jnr.10635. PubMed PMID: 12836160. [DOI] [PubMed]
- 155.Kalueff AV, Lehtimaki KA, Ylinen A, Honkaniemi J, Peltola J. Intranasal administration of human IL-6 increases the severity of chemically induced seizures in rats. Neurosci Lett. 2004;365(2):106-110. Epub 2004/07/13. doi: 10.1016/j.neulet.2004.04.061. PubMed PMID: 15245788. [DOI] [PubMed]
- 156.Chaudhry SR, Stoffel-Wagner B, Kinfe TM, Guresir E, Vatter H, Dietrich D, Lamprecht A, Muhammad S. Elevated systemic IL-6 levels in patients with aneurysmal subarachnoid hemorrhage is an unspecific marker for post-SAH complications. Int J Mol Sci. 2017;18(12). Epub 2017/12/02. doi: 10.3390/ijms18122580. PubMed PMID: 29194369; PMCID: PMC5751183. [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable