

Abstract
Despite substantial improvements in therapeutic strategies, cardiovascular disease (CVD) is still among the leading causes of mortality and morbidity worldwide. Exosomes, extracellular vesicles with a lipid bilayer membrane of endosomal origin, have been the focus of a large body of research in CVD. Exosomes not only serve as carriers for signal molecules responsible for intercellular and interorgan communication underlying CVD pathophysiology but also are bioactive agents which are partly responsible for the therapeutic effect of stem cell therapy of CVD. We here review recent insights gained into the role of exosomes in apoptosis, hypertrophy, angiogenesis, fibrosis, and inflammation in CVD pathophysiology and progression and the application and mechanisms of exosomes as therapeutic agents for CVD.
1. Introduction
Cardiovascular disease (CVD), with a global prevalence of 10.6%, imposes a large health and economic burden [1]. Numerous intra- and extracellular factors, biochemical complexes, and vesicles have been found to participate in regulating the pathophysiological progression of CVD, and studies have suggested that exosomes play important roles in it as well. Exosomes are a subgroup of extracellular vesicles surrounded by a lipid bilayer membrane of endosomal origin and ranging in size from 40 to 160 nm (average~100 nm) [2, 3]. The contents of exosomes include lipids, proteins, mitochondrial DNA, mRNAs, and noncoding RNAs, which are constantly changing both in quality and in quantity according to the microenvironment where the parent cells are [4, 5]. Exosomes are important in intercellular and interorgan communication by delivering signal molecules to recipient cells and regulating downstream signal pathways which have been associated with CVD progression [6, 7]. Moreover, exosomes released by progenitor cells are bioactive and are the key mediators of stem cell therapy of CVD, which simultaneously overcome some limitations of stem cell therapy [8–10].
In this review, we outline the role and function of exosomes in CVD pathophysiological progression and focus on their use to repair the cardiac injury.
2. Exosomes in Cardiovascular Pathophysiology
CVD progression entails a series of basic pathological changes, including cardiomyocyte apoptosis and hypertrophic growth, angiogenesis, cardiac fibrosis, and inflammation. Exosomes play a critical role in regulating CVD progression via transport and exchange of signal molecules [11, 12] (see Figure 1).
Figure 1.
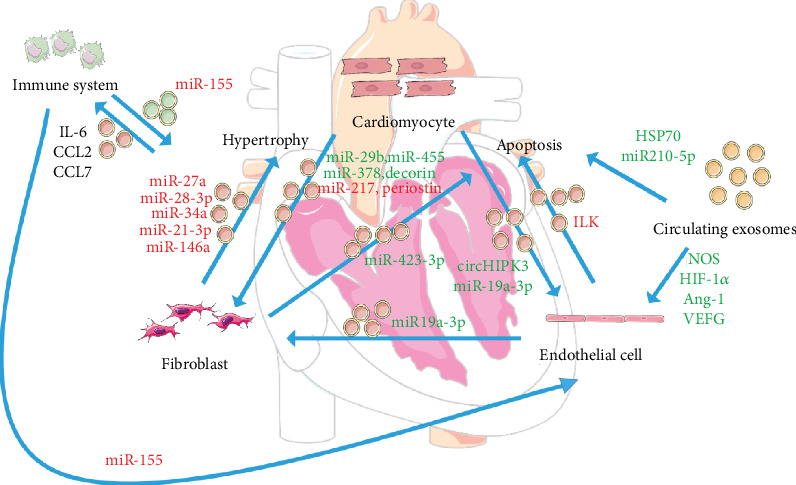
The roles of exosomes in the pathophysiology of cardiovascular diseases. Exosomes serve as the messengers exchanging bioinformation among cardiomyocytes, fibroblasts, endothelial cell, and the immune system. The molecules transported by exosomes regulate the hypertrophy, apoptosis, fibrosis, angiogenesis, and immune response in the recipient cells; those exerting adverse effects are depicted in red, while those depicted in green and black exert beneficial and neutral effects in the corresponding pathological process.
2.1. Apoptosis
The apoptosis of cardiomyocytes is a critical event in CVD progression, including myocardial infarction (MI) and heart failure. By transporting signal molecules between cardiomyocytes and other cells and organs, exosomes regulate the apoptosis of cardiomyocyte apoptosis. For example, exosomes released by cardiac fibroblasts rescue cardiomyocytes from ischemia-reperfusion injury. Mechanistically, miR-423-3p, which is remarkably enhanced in cardiac fibroblasts and exosomes, fulfills its cardioprotective effect by downregulating RAP2C in H9C2 cells and consequently reducing apoptosis [13]. The bioactive factors in the exosomes can also be proteins. For instance, circulating exosomes isolated from healthy volunteers and rats protect cardiomyocytes from apoptosis by the interaction between exosomal HSP70 and TLR4 on the cardiomyocytes and subsequent activation of the TLR4-ERK-p38MAPK-HSP27 pathway [14]. Even the exosomes secreted by cardiomyocyte itself, which is pretreated with lentivirus to overexpress HSP20, can reduce apoptosis via amplifying the AKT signaling pathway [15]. Exosomes also can be negative regulators of apoptosis. In systemic inflammation as mimicked by il-10 knockout, cardiac endothelial cell-derived exosomes aggravate cell apoptosis post infarction by increasing ILK in exosomes and activating the NF-κB pathway in recipient cells [16]. Moreover, the biological function of exosomes is highly dependent on the status quo of the cell type of origin. While exosomes derived from cardiomyocytes from healthy volunteers significantly promote proliferation and reduce apoptosis of neonatal rat cardiomyocytes in vitro, exosomes from patients with heart failure yield the opposite outcome, which may be secondary to decreased exosomal miR-21-5p which activates PTEN and downregulates Akt phosphorylation in cardiomyocytes. PDCD4, a pro-apoptotic protein, is also a target of miR-21-5p [17].
2.2. Hypertrophy
In response to TNF-α stimulation, cardiac fibroblasts secrete miR-27a-, miR-28-3p-, and miR-34a-enriched exosomes, which are taken up by cardiomyocytes and suppress the expression of Nfn2 and promote expression of cardiac hypertrophy-related genes such as ANP and β-MHC in cardiomyocytes [18]. miR-21-3p-enriched exosomes are also released by cardiac fibroblasts in the mouse model of cardiac hypertrophy, and exosomal miR-21-3p silences SORBS2 and PDLIM5 in cardiomyocytes resulting in cardiac hypertrophy [19]. Exosomes also intensify cardiac hypertrophy by promoting Ang II production and its receptor content in cardiomyocytes [20]. In a heart failure model, miR-146a is overexpressed in active myofibroblasts and secreted through exosomes. Exosomal miR-146a is taken up by cardiomyocyte leading to the dysfunction of SUMOylation [21].
2.3. Angiogenesis
The density of myocardial capillary is a critical pathological index of cardiac function and angiogenesis is of great importance in cardiac repair and regeneration post injury [5, 22]. In the MI microenvironment, proinflammatory M1-type macrophages have increased the expression of proinflammatory miRNAs, such as miR-155, and transport them to endothelial cells through exosomes. These proinflammatory exosomes inhibit Sirt1/AMPKα2/eNOS and RAC1/PAK2 pathways in endothelial cells which reduces angiogenic potential and aggravate myocardial injury [23]. Besides macrophage, cardiomyocytes in hypoxic environment release exosomes with significantly upregulated circHIPK3, which acts as an endogenous miR-29a sponge, abrogates the inhibition of IGF-1, and relieves oxidative stress-induced dysfunction in cardiac microvascular endothelial cells [24]. Exosomes derived from cardiac telocytes, which is a subgroup of interstitial Cajal-like cells, increase the proliferation, migration, and tube-formation of endothelial cells post MI [25]. Interestingly, a previous study suggests that pretreating the bilateral hindlimbs of rats with tourniquets could enhance the angiogenesis and alleviate the apoptosis of endothelial cells post MI, likely secondary to the proangiogenic contents of circulating exosomes, such as NOS, HIF-1α, Ang-1, and VEGF [26]. A novel therapy, low-energy shock wave therapy, also has been proven to enhance cardiac regeneration post MI by improving vascularization. The shock wave therapy can stimulate endothelial cells in the ischemic myocardium to release angiogenic exosomes, and miR-19a-3p is the effective cargo to promote endothelial tube formation and proliferation [27].
2.4. Cardiac Fibrosis
Cardiac fibroblasts are indispensable for normal myocardial physiology. However, pathologically activated cardiac fibroblasts deposit excess extracellular matrix, which negatively affects myocardial compliance and stiffness and cardiac function, leading to, for instance, heart failure with preserved ejection fraction [28–30]. Cardiac fibrosis can be modulated by different types of exosomes. Under exercise conditions, diabetic cardiomyocytes release exosomes with higher content of miR-29b and miR-455, which can bind to the 3′ region of MMP9, suppressing its expression and reducing cardiac fibrosis [31]. In the chronic heart failure model, cardiomyocyte-derived miR-217-containing exosomes target PTEN and aggravate cardiac fibrosis both in vivo and in vitro partly by promoting the proliferation of fibroblasts [32]. Mechanical stress increases the secretion of exosomal miR-378 from cardiomyocytes, and exosomal miR-378 inhibits excessive cardiac fibrosis by suppressing the p38 MAPK-Smad2/3 pathway [33]. Exosomes derived from macrophages are also involved in the pathological activation of cardiac fibroblasts. Exosomes derived from macrophages in a diabetic microenvironment exacerbate cardiac fibrosis, which is abrogated by human antigen R (HuR) knockdown (either in macrophages or in exosomes) [34]. Outsider intervention can also regulate the crosstalk inside a cardiac microenvironment and affect the process of fibrosis. Injection into the ischemic myocardium of exosomes derived from endothelial cells stimulated by shock wave therapy decreases myocardial fibrosis [27]. Simvastatin significantly attenuates collagen deposition and cardiac fibrosis in rats treated with Ang II, which appears secondary to simvastatin-mediated induction of decorin and reduction of periostin in Ang II-treated cardiomyocyte-derived exosomes [35].
2.5. Inflammation
The interaction between the immune and cardiovascular systems plays an important and complex role in CVD progression, with exosomes mediating the exchange of signals among cells and organs [36]. Inflammation promotes the secretion of miR-155-enriched exosomes by activated macrophages. Compared with the control group, the myocardium treated with miR-155-enriched exosomes exhibits markedly increased expression of proinflammatory cytokines, including IL-1β, IL-6, TNF-α, and CCL-2, likely secondary to the suppression of Socs1 [37]. Systematic inflammation is also reported to impair the beneficial effect of endothelial exosomes on cardiac repair post cardiac injury [16]. The “cardioimmune” regulation appears to be reciprocal, as myocardium-derived exosomes can also influence the localization and function of immune cells post cardiac injury. Post MI, cardiac exosomes are rapidly released to the interstitial space but also rapidly disappear, which is reported to be related to immune cell infiltration into the ischemic region. Migrating immune cells, mainly monocytes, engulf cardiac exosomes and increase the expression of IL-6 and chemokines CCL2 and CCL7, which shapes the inflammation post MI [38].
The roles and functions of exosomes in the cardiovascular system have also been characterized in the normal conditions. For example, a study revealed that both in the basal level and in the mild stress state, exosomal HSP60 is continuously released by cardiomyocytes [39]. However, the scarcity of such studies hinders us from figuring out the physiological role of exosomes.
3. Application of Exosomes in Cardiovascular Disease Treatment
Exosomes are deemed as the bioactive ingredient responsible for the beneficial effects of stem cell therapy in repairing cardiac injury. Exosomes as therapeutic agents can overcome some drawbacks of stem cell therapy, such as the low stem cell retention rate. We here summarize the therapeutic effects of exosomes and their underlying mechanisms (see Figure 2).
Figure 2.

Application of exosomes in the treatment of cardiovascular diseases. Multiple stem cells, namely, mesenchymal stromal cells, cardiosphere-derived cells, cardiac progenitor cells, induced pluripotent stem cells, and embryonic stem cells, secrete exosomes containing therapeutic molecules to modulate the macrophage polarization, cardiomyocyte apoptosis, angiogenesis, and other responses in cardiac injury.
3.1. Mesenchymal Stromal Cell-Derived Exosomes
Mesenchymal stromal cell- (MSC-) derived exosomes have become attractive candidates for cardiac injury repair. MSC-derived exosomes enhance the polarization of M1 macrophages to M2 macrophages which significantly alleviates the inflammation in the heart and reduces the infarct size. miR-182 knockout in exosomes or TLR4 knockout in macrophages attenuates the immunomodulation function of MSC-derived exosomes [40]. In terms of apoptosis, the content of miR-185 is dramatically increased in exosomes released from MSCs, which suppresses SOCS2 and rescues cardiomyocytes from apoptosis post infarction [41]. MSC-derived exosomal miR-19a also exhibit antiapoptosis effects, by targeting SOX6, activating AKT, and inhibiting JNK3/caspase-3 pathway in cardiomyocytes [42].
Despite the aforementioned salutary effects, the efficacy of exosomes isolated from original and nonartificial MSCs remains limited, which has triggered efforts to optimize and engineer MSCs. Exosomes obtained from MSCs overexpressing MIF show a better cardioprotective effect on reducing mitochondrial fragmentation and cardiomyocyte apoptosis compared to normal MSC-derived exosomes [43]; this is also the case for exosomes secreted by MSCs cultured in a medium containing extra MIF. Mechanically, the exosomal transfer of lncRNA-NEAT1 modulates miR-142-3p and FOXO1 in cardiomyocytes and ameliorates oxidative stress [44]. Interestingly, circulating exosomes isolated from serum post MI can also be a stimulant of MSCs. The serum exosomes, which are proved to mainly come from the kidney and ischemic myocardium, transfer miR-1956 to MSCs, downregulate Notch-1 pathway, and significantly enhance the pancreatic function of MSCs [45]. The efficacy of MSC-derived exosomes is also greatly influenced by the origin of MSCs. For instance, exosomes secreted by human fetal amniotic fluid stem cells (hAFSCs) possess higher cardioprotective potential than exosomes derived from traditional adult MSCs [46].
3.2. Cardiosphere-Derived Cell-Derived Exosomes
Exosomes released by cardiosphere-derived cells (CDCs) alleviate cardiac hypertrophy induced by Ang II and reduce myocardial infarct size post infarction. The Y RNA fragment EV-YF1, which is the maximum small RNA inside exosomes, can mimic such cardioprotective effect by altering the IL-10 expression [47, 48]. CDC-derived exosomes alleviate cardiac fibrosis, enhance tube formation of endothelial cells, and attenuate the cardiomyocyte apoptosis post MI [49–51]. Macrophage polarization is also regulated by CDC-derived exosomes after MI. Exosomal miR-181b hinders PKCδ expression and enhances M1 to M2 macrophage shift, which underlies the beneficial effect of CDC-derived exosomes [52]. Such a protective effect also has been documented in large animals such as pigs, further underscoring the therapeutic effect of CDC-derived exosomes following intravenous delivery in a myocardial infarction model [53]. Moreover, exosomes obtained from CDCs of neonatal rats or pediatric donors induce cardiac rejuvenation in old ones, as evidenced by the decreased cardiac hypertrophy and myocardial fibrosis, improved cardiac systolic and diastolic function, and enhanced exercise capability [54].
Like MSCs, exosomes obtained from engineered CDCs exhibit an improved protective effect. Hypoxic treatment, as the most widely used preconditioning method, increases the efficacy of CDC-derived exosomes by enriching proangiogenic miRs, such as miR-126, miR-130a, and miR-210, in exosomes [50]. The antiapoptotic effect of CDC-derived exosomes is also reinforced after hypoxic treatment [51]. Preclinical research has revealed that the decreased potency of CDC and its exosomes are associated with the inactivation of Wnt/β-catenin pathway, and β-catenin can efficiently reverse the therapeutic efficacy of the low potency CDCs [55]. To promote the cardiac tropism of CDC-derived exosomes, CDCs are engineered to connect a cardiomyocyte-specific binding peptide to the N-terminus of Lamp2b, which is a transmembrane protein on the exosomes. This engineered exosomes show better cardiomyocyte-specific uptake and enhanced protective effect [56].
3.3. Cardiac Progenitor Cell-Derived Exosomes
Cardiac progenitor cells (CPCs) are also emerging as a promising candidate in stem cell-based therapy for cardiac repair. CPC-derived exosomes effectively attenuate cardiomyocyte apoptosis induced by oxidative stress in vitro in an exosomal miR-21-dependent manner, which targets the gene PDCD4 in cardiomyocytes [57]. In vivo, reduced apoptosis and improved ejection fraction are observed in the infarcted hearts injected with exosomes released by CPCs, which has been associated with increased miR-210 in exosomes and decreased ephrin A3 and PTP1b in cardiomyocytes [58]. Moreover, tube formation in endothelial cells is promoted by CPC-derived exosomes because the miR-132 in exosomes inhibits the RasGAP-p120 [58]. Exosomes from CPCs perform better in alleviating ischemic cardiac injury than those from bone marrow-derived MSCs. The beneficial effects are associated with the activation of PAPP-A, the release of IGF-1 in exosomes, and the enhancement of intracellular Akt and ERK1/2 [59].
As for expanding the efficacy of CPC-derived exosomes, ticagrelor appears to be an effective stimulant, which can improve both the quantity and quality of CPC-derived exosomes. Ticagrelor enhances the mitotic activity of CPCs and thus increases the number of the exosome level. Meanwhile, exosomes from CPC precondition with ticagrelor show improved antiapoptotic activity through the activation of the ERK1/2 pathway [60].
3.4. Other Stem Cell-Derived Exosomes
Exosomes secreted by induced pluripotent stem (iPS) cells protect the heart against multiple stress [61]. iPS cell-derived exosomes enhance the autophagic flux by inhibiting mTOR pathway and promoting cardiomyocyte survival both in vitro and in vivo [62]. Cardioprotective miR-21 and miR-210 are transported to cardiomyocytes by exosomes obtained from iPS cells, and they suppress caspase 3/7 and improve the function of an ischemic myocardium [63].
Embryonic stem cell-derived exosomes are also effective in relieving cardiac injury [64]. For example, cardiomyocyte pyroptosis induced by doxorubicin can be greatly attenuated by the treatment of embryonic stem cell-derived exosomes, which can decrease the inflammation in the injured myocardium by blocking caspase-1-dependent cell death [65]. Cardiac injury induced by doxorubicin is also ameliorated by the increased number of M2 macrophages and levels of anti-inflammatory cytokines, which is mediated by embryonic stem cell-derived exosomes [66]. Besides promoting neovascularization and cardiomyocyte survival in infarcted hearts, exosomes from embryonic stem cells augment the survival and proliferation of CPCs and promote endogenous repair, which is mediated by the exosomal miR-294 [67].
3.5. Nonstem Cell-Derived Exosomes
Cardiac injury also can be alleviated by the exosomes derived from nonstem cells. For example, upon hypoxia stress, cardiac fibroblasts would release miR-423-3p-enriched exosomes and reduce apoptosis of cardiomyocytes [13]. Cardiomyocyte-derived exosomes can reduce the cardiac fibrosis caused by mechanical stress through expanding the content of miR-378 [33]. The response partially belongs to the body's self-repair system, and the targeted activation of the process can be the new treatment strategy of cardiovascular diseases.
3.6. Exosome Delivery Method
Exosomes are most commonly delivered via the intravenous route and have been shown to be effective in many animal models and in patients with graft versus host disease [68, 69]. Exosomes also alleviate cardiac injury when administered via an intracoronary route, intramyocardial injection, and cardiac patch [53, 70]. The huge benefits of exosome-based therapy have fueled exploration of more clinically feasible and effective exosome delivery methods, which will greatly promote the clinical applications of exosomes [71, 72].
4. Conclusion
Exosomes play critical and important roles in the regulation of physiological and pathophysiological processes, recognition and diagnosis, and treatment of CVD [11]. In this review, we focus on the regulation of pathophysiological process by exosomes and the therapeutic potential, which is of great importance in understanding the mechanism of CVDs and exploring therapeutic approaches. However, there remain many hurdles, including tropism and pharmacokinetics of exosomes, to surmount for their clinical application. Research to further assess the contents of exosomes and the signals exchanged by exosomes will continue, as will efforts to optimize efficacy and delivery methods for potential clinical applications.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 81900338, 31771371, and 31970654), Shaanxi Natural Science Basic Research Program (2020JQ-455), and Eagle Program from the Fourth Military Medical University (No. 015210).
Abbreviations
- CVD:
Cardiovascular disease
- MI:
Myocardial infarction
- RAP2C:
Ras-related protein Rap-2c
- ILK:
Integrin-linked kinase
- NF-κB:
Nuclear factor κB
- Nfn2:
Nuclear factor erythroid 2-related factor 2
- SORBS2:
Sorbin and SH3 domain-containing protein 2
- PDLIM5:
PDZ and LIM domain 5
- Ang II:
Angiotensin II
- Sirt1:
Sirtuin 1
- AMPK:
Protein kinase AMP-activated catalytic subunit
- eNOS:
Endothelial nitric oxide synthase
- RAC1:
Rac family small GTPase 1
- PAK2:
p21 (RAC1)-activated kinase 2
- MMP9:
Matrix Metalloprotease 9
- MSCs:
Mesenchymal stromal cells
- TLR4:
Toll-like receptor 4
- SOCS2:
Suppressor of cytokine signaling 2
- MIF:
Macrophage migration inhibitory factor
- FOXO1:
Forkhead class O1
- CDCs:
Cardiosphere-derived cells
- PKCδ:
Protein kinase Cδ
- CPCs:
Cardiac progenitor cells
- PDCD4:
Programmed cell death 4
- PAPP-A:
Pregnancy-associated plasma protein-A
- IGF-1:
Insulin-like growth factor-1
- iPS cells:
Induced pluripotent stem cells
- mTOR:
Mammalian target of rapamycin
- HSP60:
Heat shock protein 60.
Contributor Information
Yan Li, Email: profleeyan@163.com.
Mingming Zhang, Email: winterzhang3@163.com.
Conflicts of Interest
The authors declare that there is no conflict of interest regarding the publication of this article.
Authors' Contributions
Dong Guo and Yuerong Xu contributed equally to this work.
References
- 1.Virani S. S., Alonso A., Benjamin E. J., et al. Heart disease and stroke statistics-2020 update: a report from the American Heart Association. Circulation. 2020;141(9):e139–e596. doi: 10.1161/CIR.0000000000000757. [DOI] [PubMed] [Google Scholar]
- 2.Théry C., Witwer K. W., Aikawa E., et al. Minimal information for studies of extracellular vesicles 2018 (MISEV 2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. Journal of Extracellular Vesicles. 2018;7(1):p. 1535750. doi: 10.1080/20013078.2018.1535750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kalluri R., LeBleu V. S. The biology function and biomedical applications of exosomes. Science. 2020;367(6478):p. eaau6977. doi: 10.1126/science.aau6977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Yáñez-Mó M., Siljander P. R. M., Andreu Z., et al. Biological properties of extracellular vesicles and their physiological functions. Journal of Extracellular Vesicles. 2015;4(1):p. 27066. doi: 10.3402/jev.v4.27066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bei Y., Das S., Rodosthenous R. S., et al. Extracellular vesicles in cardiovascular theranostics. Theranostics. 2017;7(17):4168–4182. doi: 10.7150/thno.21274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Waldenström A., Ronquist G. Role of exosomes in myocardial remodeling. Circulation Research. 2014;114(2):315–324. doi: 10.1161/CIRCRESAHA.114.300584. [DOI] [PubMed] [Google Scholar]
- 7.Meldolesi J. Exosomes and ectosomes in intercellular communication. Current Biology. 2018;28(8):R435–R444. doi: 10.1016/j.cub.2018.01.059. [DOI] [PubMed] [Google Scholar]
- 8.Ibrahim A. . G.-E., Cheng K., Marbán E. Exosomes as critical agents of cardiac regeneration triggered by cell therapy. Stem Cell Reports. 2014;2(5):606–619. doi: 10.1016/j.stemcr.2014.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kervadec A., Bellamy V., el Harane N., et al. Cardiovascular progenitor–derived extracellular vesicles recapitulate the beneficial effects of their parent cells in the treatment of chronic heart failure. The Journal of Heart and Lung Transplantation. 2016;35(6):795–807. doi: 10.1016/j.healun.2016.01.013. [DOI] [PubMed] [Google Scholar]
- 10.Marbán E. The secret life of exosomes: what bees can teach us about next-generation therapeutics. Journal of the American College of Cardiology. 2018;71(2):193–200. doi: 10.1016/j.jacc.2017.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Barile L., Moccetti T., Marbán E., Vassalli G. Roles of exosomes in cardioprotection. European Heart Journal. 2017;38(18):1372–1379. doi: 10.1093/eurheartj/ehw304. [DOI] [PubMed] [Google Scholar]
- 12.Sahoo S., Losordo D. W. Exosomes and cardiac repair after myocardial infarction. Circulation Research. 2014;114(2):333–344. doi: 10.1161/CIRCRESAHA.114.300639. [DOI] [PubMed] [Google Scholar]
- 13.Luo H., Li X., Li T., et al. microRNA-423-3p exosomes derived from cardiac fibroblasts mediates the cardioprotective effects of ischaemic post-conditioning. Cardiovascular Research. 2019;115(7):1189–1204. doi: 10.1093/cvr/cvy231. [DOI] [PubMed] [Google Scholar]
- 14.Vicencio J. M., Yellon D. M., Sivaraman V., et al. Plasma exosomes protect the myocardium from ischemia-reperfusion injury. Journal of the American College of Cardiology. 2015;65(15):1525–1536. doi: 10.1016/j.jacc.2015.02.026. [DOI] [PubMed] [Google Scholar]
- 15.Yu D. W., Ge P. P., Liu A. L., Yu X. Y., Liu T. T. HSP20-mediated cardiomyocyte exosomes improve cardiac function in mice with myocardial infarction by activating Akt signaling pathway. European Review for Medical and Pharmacological Sciences. 2019;23(11):4873–4881. doi: 10.26355/eurrev_201906_18075. [DOI] [PubMed] [Google Scholar]
- 16.Yue Y., Wang C., Benedict C., et al. Interleukin-10 deficiency alters endothelial progenitor cell-derived exosome reparative effect on myocardial repair via integrin-linked kinase enrichment. Circulation Research. 2020;126(3):315–329. doi: 10.1161/CIRCRESAHA.119.315829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Qiao L., Hu S., Liu S., et al. microRNA-21-5p dysregulation in exosomes derived from heart failure patients impairs regenerative potential. The Journal of Clinical Investigation. 2019;129(6):2237–2250. doi: 10.1172/JCI123135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tian C., Gao L., Zimmerman M. C., Zucker I. H. Myocardial infarction-induced microRNA-enriched exosomes contribute to cardiac Nrf2 dysregulation in chronic heart failure. American Journal of Physiology. Heart and Circulatory Physiology. 2018;314(5):H928–H939. doi: 10.1152/ajpheart.00602.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bang C., Batkai S., Dangwal S., et al. Cardiac fibroblast-derived microRNA passenger strand-enriched exosomes mediate cardiomyocyte hypertrophy. The Journal of Clinical Investigation. 2014;124(5):2136–2146. doi: 10.1172/JCI70577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lyu L., Wang H., Li B., et al. A critical role of cardiac fibroblast-derived exosomes in activating renin angiotensin system in cardiomyocytes. Journal of Molecular and Cellular Cardiology. 2015;89(Part B):268–279. doi: 10.1016/j.yjmcc.2015.10.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Oh J. G., Watanabe S., Lee A., et al. miR-146a suppresses SUMO1 expression and induces cardiac dysfunction in maladaptive hypertrophy. Circulation Research. 2018;123(6):673–685. doi: 10.1161/CIRCRESAHA.118.312751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Silvestre J.-S., Smadja D. M., Lévy B. I. Postischemic revascularization: from cellular and molecular mechanisms to clinical applications. Physiological Reviews. 2013;93(4):1743–1802. doi: 10.1152/physrev.00006.2013. [DOI] [PubMed] [Google Scholar]
- 23.Liu S., Chen J., Shi J., et al. M1-like macrophage-derived exosomes suppress angiogenesis and exacerbate cardiac dysfunction in a myocardial infarction microenvironment. Basic Research in Cardiology. 2020;115(2):p. 22. doi: 10.1007/s00395-020-0781-7. [DOI] [PubMed] [Google Scholar]
- 24.Wang Y., Zhao R., Liu W., et al. Exosomal circHIPK3 Released from Hypoxia-Pretreated Cardiomyocytes Regulates Oxidative Damage in Cardiac Microvascular Endothelial Cells via the miR-29a/IGF-1 Pathway. Oxidative medicine and cellular longevity. 2019;2019, article 7954657:28. doi: 10.1155/2019/7954657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Yang J., Li Y., Xue F., Liu W., Zhang S. Exosomes derived from cardiac telocytes exert positive effects on endothelial cells. American Journal of Translational Research. 2017;9(12):5375–5387. [PMC free article] [PubMed] [Google Scholar]
- 26.Chen Q., Huang M., Wu J., Jiang Q., Zheng X. Exosomes isolated from the plasma of remote ischemic conditioning rats improved cardiac function and angiogenesis after myocardial infarction through targeting Hsp 70. Aging. 2020;12(4):3682–3693. doi: 10.18632/aging.102837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gollmann-Tepeköylü C., Pölzl L., Graber M., et al. miR-19a-3p containing exosomes improve function of ischaemic myocardium upon shock wave therapy. Cardiovascular Research. 2020;116(6):1226–1236. doi: 10.1093/cvr/cvz209. [DOI] [PubMed] [Google Scholar]
- 28.Aghajanian H., Kimura T., Rurik J. G., et al. Targeting cardiac fibrosis with engineered T cells. Nature. 2019;573(7774):430–433. doi: 10.1038/s41586-019-1546-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lam C. S. P., Voors A. A., de Boer R. A., Solomon S. D., van Veldhuisen D. J. Heart failure with preserved ejection fraction: from mechanisms to therapies. European Heart Journal. 2018;39(30):2780–2792. doi: 10.1093/eurheartj/ehy301. [DOI] [PubMed] [Google Scholar]
- 30.Kong P., Christia P., Frangogiannis N. G. The pathogenesis of cardiac fibrosis. Cellular and Molecular Life Sciences. 2014;71(4):549–574. doi: 10.1007/s00018-013-1349-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chaturvedi P., Kalani A., Medina I., Familtseva A., Tyagi S. C. Cardiosome mediated regulation of MMP9 in diabetic heart: role of mir29b and mir455 in exercise. Journal of Cellular and Molecular Medicine. 2015;19(9):2153–2161. doi: 10.1111/jcmm.12589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Nie X., Fan J., Li H., et al. miR-217 promotes cardiac hypertrophy and dysfunction by targeting PTEN. Molecular Therapy - Nucleic Acids. 2018;12:254–266. doi: 10.1016/j.omtn.2018.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yuan J., Liu H., Gao W., et al. MicroRNA-378 suppresses myocardial fibrosis through a paracrine mechanism at the early stage of cardiac hypertrophy following mechanical stress. Theranostics. 2018;8(9):2565–2582. doi: 10.7150/thno.22878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Govindappa P. K., Patil M., Garikipati V. N. S., et al. Targeting exosome-associated human antigen R attenuates fibrosis and inflammation in diabetic heart. The FASEB Journal. 2020;34(2):2238–2251. doi: 10.1096/fj.201901995R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kuo H.-F., Hsieh C.-C., Wang S.-C., et al. Simvastatin attenuates cardiac fibrosis via regulation of cardiomyocyte-derived exosome secretion. Journal of Clinical Medicine. 2019;8(6):p. 794. doi: 10.3390/jcm8060794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wu R., Gao W., Yao K., Ge J. Roles of exosomes derived from immune cells in cardiovascular diseases. Frontiers in Immunology. 2019;10:p. 648. doi: 10.3389/fimmu.2019.00648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wang C., Zhang C., Liu L., et al. Macrophage-derived mir-155-containing exosomes suppress fibroblast proliferation and promote fibroblast inflammation during cardiac injury. Molecular Therapy. 2017;25(1):192–204. doi: 10.1016/j.ymthe.2016.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Loyer X., Zlatanova I., Devue C., et al. Intra-cardiac release of extracellular vesicles shapes inflammation following myocardial infarction. Circulation Research. 2018;123(1):100–106. doi: 10.1161/CIRCRESAHA.117.311326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gupta S., Knowlton A. A. HSP60 trafficking in adult cardiac myocytes: role of the exosomal pathway. American Journal of Physiology-Heart and Circulatory Physiology. 2007;292(6):H3052–H3056. doi: 10.1152/ajpheart.01355.2006. [DOI] [PubMed] [Google Scholar]
- 40.Zhao J., Li X., Hu J., et al. Mesenchymal stromal cell-derived exosomes attenuate myocardial ischaemia-reperfusion injury through miR-182-regulated macrophage polarization. Cardiovascular Research. 2019;115(7):1205–1216. doi: 10.1093/cvr/cvz040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Li Y., Zhou J., Zhang O., et al. Bone marrow mesenchymal stem cells-derived exosomal microRNA-185 represses ventricular remolding of mice with myocardial infarction by inhibiting SOCS2. International Immunopharmacology. 2020;80:p. 106156. doi: 10.1016/j.intimp.2019.106156. [DOI] [PubMed] [Google Scholar]
- 42.Huang L., Yang L., Ding Y., Jiang X., Xia Z., You Z. Human umbilical cord mesenchymal stem cells-derived exosomes transfers microRNA-19a to protect cardiomyocytes from acute myocardial infarction by targeting SOX6. Cell Cycle. 2020;19(3):339–353. doi: 10.1080/15384101.2019.1711305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Liu X., Li X., Zhu W., et al. Exosomes from mesenchymal stem cells overexpressing MIF enhance myocardial repair. Journal of Cellular Physiology. 2020 doi: 10.1002/jcp.29456. [DOI] [PubMed] [Google Scholar]
- 44.Chen H., Xia W., Hou M. LncRNA-NEAT1 from the competing endogenous RNA network promotes cardioprotective efficacy of mesenchymal stem cell-derived exosomes induced by macrophage migration inhibitory factor via the miR-142-3p/FOXO1 signaling pathway. Stem Cell Research & Therapy. 2020;11(1):p. 31. doi: 10.1186/s13287-020-1556-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Gao L., Mei S., Zhang S., et al. Cardio-renal exosomes in myocardial infarction serum regulate proangiogenic paracrine signaling in adipose mesenchymal stem cells. Theranostics. 2020;10(3):1060–1073. doi: 10.7150/thno.37678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Takov K., He Z., Johnston H. E., et al. Small extracellular vesicles secreted from human amniotic fluid mesenchymal stromal cells possess cardioprotective and promigratory potential. Basic Research in Cardiology. 2020;115(3):p. 26. doi: 10.1007/s00395-020-0785-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Cambier L., Giani J. F., Liu W., et al. Angiotensin II-induced end-organ damage in mice is attenuated by human exosomes and by an exosomal Y RNA fragment. Hypertension. 2018;72(2):370–380. doi: 10.1161/HYPERTENSIONAHA.118.11239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Cambier L., Couto G., Ibrahim A., et al. Y RNA fragment in extracellular vesicles confers cardioprotection via modulation of IL-10 expression and secretion. EMBO Molecular Medicine. 2017;9(3):337–352. doi: 10.15252/emmm.201606924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Tseliou E., Fouad J., Reich H., et al. Fibroblasts rendered antifibrotic, antiapoptotic, and angiogenic by priming with cardiosphere-derived extracellular membrane vesicles. Journal of the American College of Cardiology. 2015;66(6):599–611. doi: 10.1016/j.jacc.2015.05.068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Namazi H., Mohit E., Namazi I., et al. Exosomes secreted by hypoxic cardiosphere-derived cells enhance tube formation and increase pro-angiogenic miRNA. Journal of Cellular Biochemistry. 2018;119(5):4150–4160. doi: 10.1002/jcb.26621. [DOI] [PubMed] [Google Scholar]
- 51.Namazi H., Namazi I., Ghiasi P., et al. Exosomes secreted by normoxic and hypoxic cardiosphere-derived cells have anti-apoptotic effect. Iranian Journal of Pharmaceutical Research. 2018;17(1):377–385. [PMC free article] [PubMed] [Google Scholar]
- 52.de Couto G., Gallet R., Cambier L., et al. Exosomal microRNA transfer into macrophages mediates cellular postconditioning. Circulation. 2017;136(2):200–214. doi: 10.1161/CIRCULATIONAHA.116.024590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Gallet R., Dawkins J., Valle J., et al. Exosomes secreted by cardiosphere-derived cells reduce scarring, attenuate adverse remodelling, and improve function in acute and chronic porcine myocardial infarction. European Heart Journal. 2017;38(3):201–211. doi: 10.1093/eurheartj/ehw240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Grigorian-Shamagian L., Liu W., Fereydooni S., et al. Cardiac and systemic rejuvenation after cardiosphere-derived cell therapy in senescent rats. European Heart Journal. 2017;38(39):2957–2967. doi: 10.1093/eurheartj/ehx454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ibrahim A. G. E., Li C., Rogers R., et al. Augmenting canonical Wnt signalling in therapeutically inert cells converts them into therapeutically potent exosome factories. Nature biomedical engineering. 2019;3(9):695–705. doi: 10.1038/s41551-019-0448-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Mentkowski K. I., Lang J. K. Exosomes engineered to express a cardiomyocyte binding peptide demonstrate improved cardiac retention in vivo. Scientific Reports. 2019;9(1):p. 10041. doi: 10.1038/s41598-019-46407-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Xiao J., Pan Y., Li X. H., et al. Cardiac progenitor cell-derived exosomes prevent cardiomyocytes apoptosis through exosomal miR-21 by targeting PDCD4. Cell Death & Disease. 2016;7(6):e2277–e2277. doi: 10.1038/cddis.2016.181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Barile L., Lionetti V., Cervio E., et al. Extracellular vesicles from human cardiac progenitor cells inhibit cardiomyocyte apoptosis and improve cardiac function after myocardial infarction. Cardiovascular Research. 2014;103(4):530–541. doi: 10.1093/cvr/cvu167. [DOI] [PubMed] [Google Scholar]
- 59.Barile L., Cervio E., Lionetti V., et al. Cardioprotection by cardiac progenitor cell-secreted exosomes: role of pregnancy-associated plasma protein-A. Cardiovascular Research. 2018;114(7):992–1005. doi: 10.1093/cvr/cvy055. [DOI] [PubMed] [Google Scholar]
- 60.Casieri V., Matteucci M., Pasanisi E. M., et al. Ticagrelor enhances release of anti-hypoxic cardiac progenitor cell-derived exosomes through increasing cell proliferation in vitro. Scientific Reports. 2020;10(1):p. 2494. doi: 10.1038/s41598-020-59225-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Fan C., Zhang E., Joshi J., Yang J., Zhang J., Zhu W. Utilization of human induced pluripotent stem cells for cardiac repair. Frontiers in Cell and Developmental biology. 2020;8:p. 36. doi: 10.3389/fcell.2020.00036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Santoso M. R., Ikeda G., Tada Y., et al. Exosomes from induced pluripotent stem cell-derived cardiomyocytes promote autophagy for myocardial repair. Journal of the American Heart Association. 2020;9(6, article e014345) doi: 10.1161/JAHA.119.014345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wang Y., Zhang L., Li Y., et al. Exosomes/microvesicles from induced pluripotent stem cells deliver cardioprotective miRNAs and prevent cardiomyocyte apoptosis in the ischemic myocardium. International Journal of Cardiology. 2015;192:61–69. doi: 10.1016/j.ijcard.2015.05.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Song Y.-H., Shao L., Zhang Y., et al. Exosomes derived from embryonic stem cells as potential treatment for cardiovascular diseases. Advances in Experimental Medicine and Biology. 2017;998:187–206. doi: 10.1007/978-981-10-4397-0_13. [DOI] [PubMed] [Google Scholar]
- 65.Tavakoli Dargani Z., Singla D. K. Embryonic stem cell-derived exosomes inhibit doxorubicin-induced TLR4-NLRP3-mediated cell death-pyroptosis. American Journal of Physiology. Heart and Circulatory Physiology. 2019;317(2):H460–H471. doi: 10.1152/ajpheart.00056.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Singla D. K., Johnson T. A., Tavakoli Dargani Z. Exosome treatment enhances anti-inflammatory M2 macrophages and reduces inflammation-induced pyroptosis in doxorubicin-induced cardiomyopathy. Cells. 2019;8(10):p. 1224. doi: 10.3390/cells8101224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Khan M., Nickoloff E., Abramova T., et al. Embryonic stem cell-derived exosomes promote endogenous repair mechanisms and enhance cardiac function following myocardial infarction. Circulation Research. 2015;117(1):52–64. doi: 10.1161/CIRCRESAHA.117.305990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Vandergriff A. C., de Andrade J. B. M., Tang J., et al. Intravenous cardiac stem cell-derived exosomes ameliorate cardiac dysfunction in doxorubicin induced dilated cardiomyopathy. Stem Cells International. 2015;2015:8. doi: 10.1155/2015/960926.960926 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Kordelas L., Rebmann V., Ludwig A. K., et al. MSC-derived exosomes: a novel tool to treat therapy-refractory graft-versus-host disease. Leukemia. 2014;28(4):970–973. doi: 10.1038/leu.2014.41. [DOI] [PubMed] [Google Scholar]
- 70.Huang K., Ozpinar E. W., Su T., et al. An off-the-shelf artificial cardiac patch improves cardiac repair after myocardial infarction in rats and pigs. Science Translational Medicine. 2020;12(538):p. eaat9683. doi: 10.1126/scitranslmed.aat9683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Gao L., Gregorich Z. R., Zhu W., et al. Large cardiac muscle patches engineered from human induced-pluripotent stem cell-derived cardiac cells improve recovery from myocardial infarction in swine. Circulation. 2018;137(16):1712–1730. doi: 10.1161/CIRCULATIONAHA.117.030785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Williams A. M., Wu Z., Bhatti U. F., et al. Early single-dose exosome treatment improves neurologic outcomes in a 7-day swine model of traumatic brain injury and hemorrhagic shock. Journal of Trauma and Acute Care Surgery. 2020:p. 1. doi: 10.1097/ta.0000000000002698. [DOI] [PubMed] [Google Scholar]