

Abstract
The section Pallasia is one of the components of the genus Allium subgenus Allium (Amaryllidaceae), and species relationship in this section is still not resolved very well, which hinders further evolutionary and adaptive studies. Here, the complete chloroplast genomes of five sect. Pallasia species were reported, and a comparative analysis was performed with other three related Allium species. The genome size of the eight species ranged from 151,672 bp to 153,339 bp in length, GC content changed from 36.7% to 36.8%, and 130 genes (except Allium pallasii), 37 tRNA, and 8 rRNA were identified in each genome. By analyzing the IR/LSC and IR/SSC boundary, A. pallasii exhibited differences compared with other seven species. Phylogenetic analysis achieved high supports in each branch, seven of the eight Allium species cluster into a group, and A. pallasii exhibit a close relationship with A. obliquum. Higher pairwise Ka/Ks ratios were found in A. schoenoprasoides compared to A. caeruleum and A. macrostemon while a lower value of Ka/Ks ratios was detected between A. caeruleum and A. macrostemon. This study will be a great contribution to the future phylogenetic and adaptive research in Allium.
1. Introduction
Genus Allium L. is one of the largest monocotyledonous genera, including about 950 species [1], and it is the only genus of Allioideae in the new APG IV [2]. This genus has a major center from the Mediterranean Basin to Central Asia and Pakistan and a second less pronounced one located in the western North America [3, 4]. Many important economical species, like onion, shallot, and scallion, were included in this genus, and some of them are regarded as vegetables or spices, even as ornamental plants like A. caeruleum [5]. Based on the phylogenetic study of ribosomal DNA-ITS sequences, the genus has been classified into 15 subgenera and 72 sections [6]. This research advanced that this genus Allium is a monophyletic group and evolved proceeded in three major clades. Species relationships in the first and second clades were almost clear, but those in the third clade were complex; in particular, species relationships among subgenera Rhizirideum, Allium, Cepa, Polyprason, and Reticulatobulbosa were still not well resolved [4, 7].
The section Pallasia is the largest section in the genus Allium subgenus Allium (Amaryllidaceae) in China. This section was firstly established by Friesen et al. with A. pallasia as the type species [6]. According to morphological study and molecular biological data, Li et al. divided the Chinese Allium species into 13 subgenera and 34 sections, and among them, A. delicatulum, A. eusperma, A. pallasii, A. glomeratum, A. schoenoprasoides, A. songpanicum, and A. tanguticum were classified into sect. Pallasia [4]. These species have a wide distribution and complex morphological diversity, in which A. schoenoprasoides grows in a grassy slope 3000 meters above the sea level, and A. pallasii grows in a dry slope with an altitude of 500 m [8]. Previous phylogenetic analysis showed that sect. Pallasia is not monophyletic, and parallel branches in phylogenetic trees made it difficult to understand the phylogenetic relationships of species in this section only using ITS and chloroplast gene fragment [4]. Polymorphism and homoplasious characters of these species also made it difficult to recognize phylogenetic position using the traditional classification method [4, 6, 7, 9].
The chloroplast is an important plastid, which provides necessary energy for growth by photosynthesis and plays vital roles in physiology and development of plant. It has a typical quadripartite circular DNA genome, mostly ranging from 120 to 160 kb, and includes a large single copy region (LSC), a small single copy region (SSC), and two inverted repeats (IRs) [10–12]. Cp genomes are haploid, and most of them are maternally inherited. High conservation of gene content and genome structure makes cp genome an important resource for reconstructing the phylogenetic relationships among plant groups [13–20], which may suggest that the cp genome could be better to solve the relationships between species [21–23]. Additionally, the cp genome encodes many chloroplast-specific components, like photosynthesis genes, which play fundamental roles in the life of plant [24–26]. A recent study showed that the adaptation of sunlight preferences was related to adaptation evolution of chloroplast genes [27, 28]. The differences of structural information in chloroplast genome may play an important role; previous studies suggested that the similarity of the IR region of some species reflected that they may have a common ancestor [29]. The comparative analysis of chloroplast genome can provide more information for phylogenetic research like highly divergent regions [28]. Other studies suggested that the selective pressure in chloroplast genomes has played a key role in Allium species adaptation and evolution [30, 31]. Although these studies have demonstrated that complete chloroplast genome has extraordinary advantages in phylogenetic analysis, the species in sect. Pallasia were not investigated, and the systematic relationships between these species are not clear until now.
In this study, we reported the complete chloroplast genomes of eight species (A. delicatulum, A. pallasii, A. schoenoprasoides, A. songpanicum, A. tanguticum from sect. Pallasia, and three related species A. caeruleum, A. teretifolium, and A. macrostemon) and performed comparative analyses. The objectives of this study were (1) to explore the overall structural pattern of the eight plastid genomes, (2) to solve the phylogenetic relationships between these species based on cp genome sequences, and (3) to analyze the adaptive evolution and selection pressures of Allium species based on chloroplast genomes.
2. Materials and Methods
2.1. DNA Extraction, Sequencing, and Annotation
The modified CTAB method [32] was used to extract genomic DNA from dried leaves of A. delicatulum, A. pallasii, A. schoenoprasoides, A. songpanicum, A. tanguticum, A. caeruleum, and A. teretifolium. Genomic data of these species were sequenced using an Illumina Hiseq 2500 platform by Biomarker Technologies, Inc. (Beijing, China). The collection information and Genebank accession numbers are in Table S1. Seven chloroplast sequences were completed with NOVOPlasty [33], a fast de novo assembler, and the seed sequence is the rbcL from the Allium cepa (KM088014); contig information is in Table S2. Contigs generated by NOVOPlasty were sorted and joined into a single draft sequence with A. cepa as the reference in the software Geneious [34]. Chloroplast genomes were annotated using PGA (Plastid Genome Annotator) [35] with two reference sequences A. cepa (KM088014) and Amborella trichopoda (AJ506156) which has the highest gene numbers among known gymnosperms and angiosperms; after that, sequences were adjusted using Geneious manually [34]. These plastid genome maps were generated using OGDRAW [36].
2.2. SSR Characterization
The MISA-web was used to find microsatellites (SSRs) in the eight Allium cp genomes [37]. The parameters are set to ten, five, four, three, three, and three for mononucleotide (mono-), dinucleotides (di-), trinucleotides (tri-), tetranucleotides (tetra-), pentanucleotide (penta-), and hexanucleotides (hexa-), respectively.
2.3. Phylogenetic Analyses
In order to clarify the phylogenetic relationship of species (A. delicatulum, A. pallasii, A. schoenoprasoides, A. songpanicum, A. tanguticum, A. caeruleum, A. teretifolium, and A. macrostemon), we download 22 cp genome sequences from Genebank, including 20 Allium species (Table S3), Agapanthus coddii (KX790363) from Agapanthoideae, and Narcissus poeticus (NC_039825) from Amaryllidoideae as the outgroups. All 30 cp genome sequences were used for phylogenetic analysis. Because molecular evolutionary rates were different in the whole cp genome, we built the phylogenetic trees based on the following three datasets: (1) complete chloroplast genomes, (2) the combined coding sequences, and (3) LSC regions. Firstly, all the sequences were aligned with MAFFT v. 7.1.2.9 [38] and adjusted with MEGA 7.0 [39] manually. Then, we used RAxML 8.2.8 [40] to perform the maximum likelihood (ML) analyses with the GTR+G model for 1000 replications. And Bayesian inference (BI) was performed in MrBayes v. 3.2.5 [41] under the GTR+G model. The Markov chain Monte Carlo (MCMC) algorithm was performed for 1,000,000 generations, trees were sampled every 1000 generations for each data partition, the first 20% of trees were discarded as burn-in, and the remaining trees were used to build a 50% majority-rule consensus tree.
2.4. Genome Comparative Analyses
The IR regions have been marked at the time of annotation using PGA [35]. IR/SSC and IR/LSC boundary analyses among the eight Allium species and the outgroups Agapanthus coddii and Narcissus poeticus were compared in Geneious [34].
Pairwise chloroplast genomic alignment among eight species was compared by mVISTA in the Shuffle-LAGAN mode [42], and Allium cepa (KM088014) was used as a reference.
The relative synonymous codon usage (RSCU) among the eight Allium species was calculated using DnaSP version 6 [43].
Combined CDS (coding sequences) of 42 photosynthesis genes were aligned using MAFFT v. 7.129 [38] for 30 cp genomes in phylogenetic analysis. Ka/Ks ratios were calculated using KaKs_Calculator 2.0 [44].
3. Results
3.1. Chloroplast Features of These Allium Species
The eight Allium complete cp genome sequences ranged from 151,672 bp (A. pallasii) to 153,339 bp (A. teretifolium) in length. All eight cp genomes showed a typical quadripartite structure (Figure 1), which consisted of a pair of IR regions (26,343–26,541 bp) separated by the LSC (80,924–82,539 bp) and SSC (17,600–18,064 bp) regions. The GC content is 36.7–36.8%, indicating nearly identical levels among the eight Allium cp genomes (Table 1). The total number of annotated genes varies between 128 and 130, including 83–85 protein-coding genes, eight rRNA genes, and 37 tRNA genes, respectively (Table 2). The length, GC content, and gene components of the eight species were included in Table 1.
Figure 1.
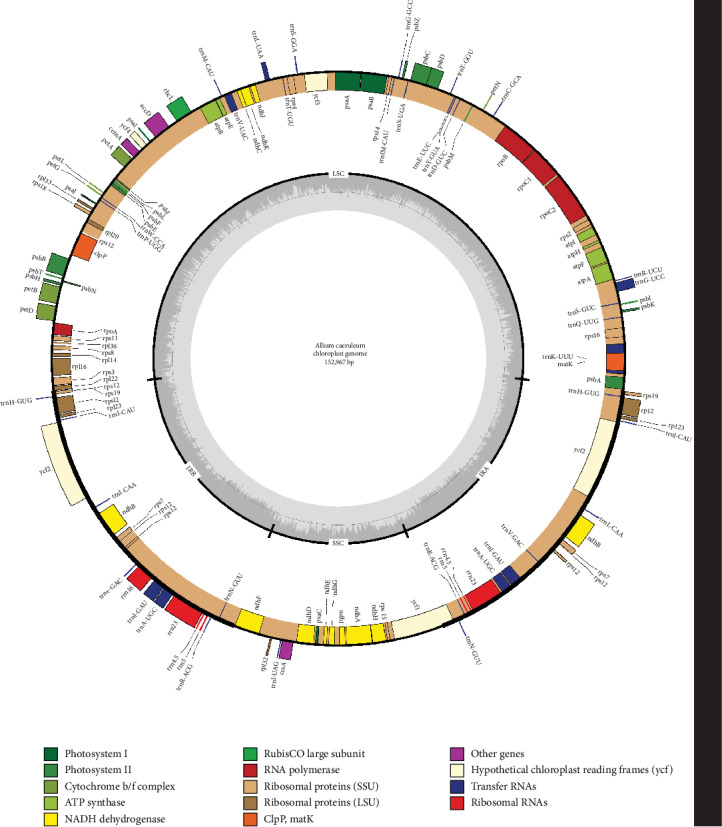
Gene map of Allium chloroplast genomes (represented by A. caeruleum). Genes outside the circle are transcribed clockwise, and genes shown on the inside of the circle are counterclockwise. Genes belonging to the functional group are color-coded. The darker gray in the inner corresponds to GC content, and the lighter gray corresponds to AT content. IR = inverted repeat; SSC = small single copy; LSC = large single copy.
Table 1.
The length and GC contents of the whole genome sequences, SSC, LSC, and IR regions and the number of the genes.
| Species | Length (bp) | GC contents (%) | Number of genes | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genome | SSC | LSC | IR | Genome | SSC | LSC | IR | Total | CDS | rRNA | tRNA | |
| A. caeruleum | 152967 | 18066 | 81884 | 26510 | 36.8 | 29.4 | 34.6 | 42.6 | 130 | 85 | 8 | 37 |
| A. delicatulum | 152984 | 17924 | 82046 | 26506 | 36.7 | 29.4 | 34.5 | 42.6 | 130 | 85 | 8 | 37 |
| A. macrostemon | 153158 | 17600 | 82700 | 26429 | 36.7 | 29.1 | 34.6 | 42.7 | 130 | 85 | 8 | 37 |
| A. pallasii | 151672 | 17665 | 80922 | 26541 | 36.6 | 29.1 | 34.4 | 42.5 | 128 | 83 | 8 | 37 |
| A. schoenoprasoides | 152729 | 18016 | 81678 | 26483 | 36.8 | 29.3 | 34.6 | 42.7 | 130 | 85 | 8 | 37 |
| A. songpanicum | 153247 | 18021 | 82537 | 26343 | 36.8 | 29.5 | 34.6 | 42.6 | 130 | 85 | 8 | 37 |
| A. tanguticum | 153024 | 17899 | 82123 | 26500 | 36.8 | 29.5 | 34.5 | 42.6 | 130 | 85 | 8 | 37 |
| A. teretifolium | 153340 | 17997 | 82281 | 26533 | 36.8 | 29.5 | 34.7 | 42.7 | 130 | 85 | 8 | 37 |
Abbreviations: CDS: protein-coding sequences/genes; LSC: large single-copy region; SSC: small single-copy region; IR: inverted repeat regions.
Table 2.
Gene contents in eight Allium species.
| Category | Group | Name |
|---|---|---|
| Self-replication | Large subunit of ribosome (LSU) | rpl2(2), rpl14, rpl16, rpl20, rpl22, rpl23(2), rpl32, rpl33, rpl36 |
| Small subunit of ribosome (SSU) | rps2 ∗, rps3, rps4, rps7(2), rps8, rps11, rps12(2), rps14, rps15, rps16∗, rps18, rps19(2) | |
| DNA-dependent RNA polymerase | rpoA, rpoB, rpoC1, rpoC2 | |
| Ribosomal RNA | rrn4.5 (2), rrna5(2), rrn16(2), rrn23(2) | |
| Transfer RNAs (tRNA) | trnA-UGC(2), trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnfM-CAU, trnG-GCC, trnG-UCC, trnH-GUG(2), trnI-CAU(2), trnI-GAU(2), trnK-UUU, trnL-CAA(2), trnL-UAA, trnL-UAG, trnM-CAU, trnN-GUU(2), trnP-UGG, trnQ-UUG, trnR-ACG(2), trnR-UCU, trnS-GCU, trnS-GGA, trnS-UGA, trnT-GGU, trnT-UGU, trnV-GAC(2), trnV-UAC, trnW-CCA, trnY-GUA | |
| Photosynthesis | Photosystem I | psaA, psaB, psaC, psaI, psaJ |
| Photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ | |
| Subunits of NADH-dehydrogenase | ndhA, ndhB(2), ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK | |
| Subunits of cytochrome b/f complex | petA, petB, petD, petG, petL, petN | |
| Subunits of ATP synthase | atpA, atpB, atpE, atpF, atpH, atpI | |
| Large subunit of rubisco | rbcL | |
| Translation initiation factor | infA | |
| Other genes | ATP-dependent protease subunit p gene | clpP |
| Maturase | matK | |
| Envelope membrane protein | cemA | |
| Subunit of acetyl-CoA-carboxylase | accD | |
| C-type cytochrome synthesis gene | ccsA | |
| Hypothetical chloroplast reading frames (ycf) | ycf1(2), ycf2(2), ycf3, ycf4 |
Note: the gene names rps2 and rps16 with ∗ show the missing gene in A. pallasii.
3.2. SSR Analysis of These Species and Codon Usage
We used MISA-web to find microsatellites (SSRs) in the eight cp genomes, and six types of perfect SSRs were found. In total, 99, 96, 95, 79, 98, 91, 95, and 91 SSRs were detected in A. delicatulum, A. macrostemon, A. pallasii, A. schoenoprasoides, A. songpanicum, A. tanguticum, A. caeruleum, and A. teretifolium. Only A. delicatulum and A. tanguticum have hexanucleotide repeats, which is special in these species. The largest group of SSRs was mononucleotide repeats, ranging from 52 to 67 in these eight species, making up more than half of SSRs (Figure 2(a)). Most SSRs are located in the LSC region (Figure 2(b)), which is identical with previous research studies [28]. The relative synonymous codon usage (RSCU) was calculated in Table S4.
Figure 2.
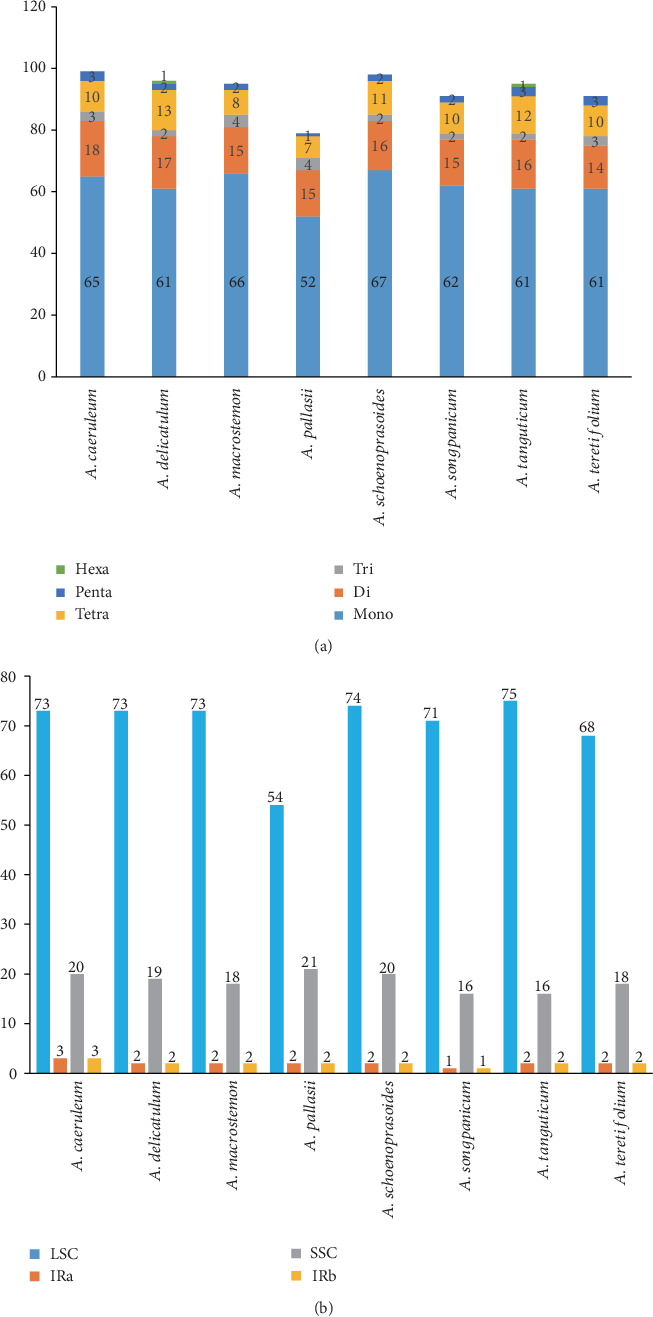
(a) Microsatellite loci in the eight chloroplast genomes for mono-, di-, tri-, tetra-, penta-, and hexanucleotides. (b) Number of SSRs in the LSC, IR, and SSC regions in eight Allium chloroplast genome sequences.
3.3. IR/SSC and IR/LSC Boundary
The expansion and contraction of the IR region in chloroplast are the main reason for the change of chloroplast genome size [45–47]. The expansion of the IRs to rps19 or rpl22 (Figure 3), which has been described by previous studies, was also found in our results. The ndhF gene flanked the junction between SSC and IRb in A. pallasii, Agapanthus coddii, and Narcissus poeticus while 22 bp–49 bp was located in the IRa region, but the ndhF gene clusters with a length of 18–50 bp away from the SSC/IRa boundary in the other seven Allium species. The psbA gene also has different positions in these species; the length from psbA gene to the LSC/IRb boundary in A. pallasii (627 bp) was much longer than that in other species (67–127 bp). There are also some differences in positions of rpl22 gene, 36 bp-49 bp was located in the IRa region in eight Allium species, but rpl22 gene was almost located in the LSC region in Agapanthus coddii and Narcissus poeticus.
Figure 3.
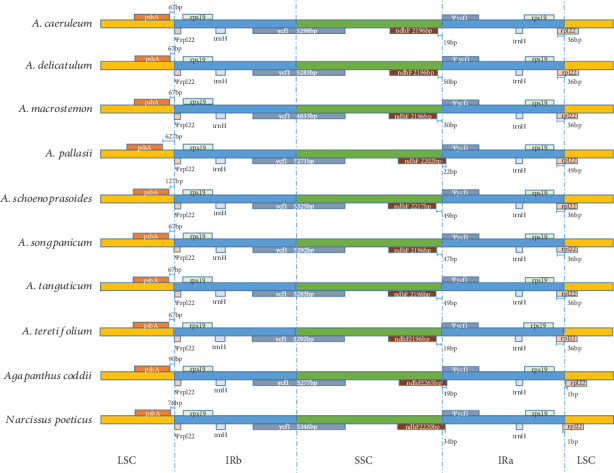
Comparison of LSC, SSC, and IR border regions among eight Allium cp genomes. Colored boxes for genes represent the gene position.
3.4. Nucleotide Diversity and Sequence Identity Plot
The mVISTA software was used to compare the complete chloroplast genome of these eight species; the annotation of A. cepa (KM088014) was used as a reference. The alignment revealed a high sequence similarity across eight Allium plastid genomes, which showed that the genomes were highly conserved (Figure 4). Highly divergent regions among eight Allium chloroplast genomes were mainly located in the intergenic spacers, including trnK-UUU–trnQ-UUG, trnS-GCU–trnR-UCU, rpoB–psbD, rps4–trnL-UAA, petA–rpl20, and ndhF–ndhD, but some protein-coding regions also have distributions like ycf1.
Figure 4.
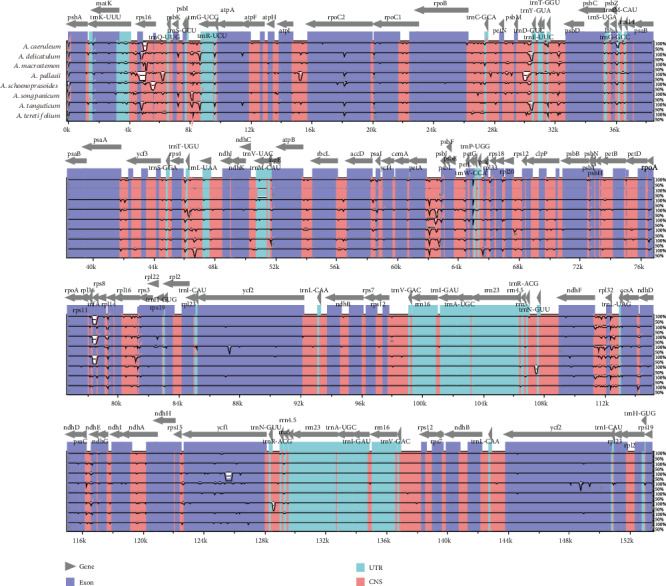
Visualization alignment of eight Allium cp genomes. VISTA-based identity plot showing sequence identity among eight Allium species using A. cepa as a reference.
3.5. Phylogenetic Analysis
The chloroplast genomes have showed great potential in reconstructing the phylogenetic relationships among plant groups [17, 48–51]. A phylogenetic tree of 28 Allium species and their outgroups (Narcissus poeticus and Agapanthus coddii) was constructed in this study. For Bayesian inference (BI) and maximum likelihood (ML) of three datasets, the posterior probabilities and bootstrap values were very high for each lineage, and only a clade had a relatively low posterior (Figure 5) (Figure S1). Both the maximum likelihood (ML) and BI phylogenetic results strongly supported that A. macrostemon, A. caeruleum, and A. schoenoprasoides were the closest species in the phylogenetic trees based on three datasets, which is the same as the previous study based on ITS sequences. These 3 species and A. platyspathum formed the sister groups to the clade of A. delicatulum and A. tanguticum. These 6 species and A. teretifolium proposed a sister relationship to the clade formed by the species from sect. Daghestanica (A. chrysanthum, A. rude, A. xichuanense, A. chrysocephalum, A. maowenense, and A. herderianum) and sect. Cepa (A. fistulosum, A. altaicum, and A. cepa). And A. chinense, A. songpanicum, A. pskemense, and A. oschaninii had a close relationship.
Figure 5.
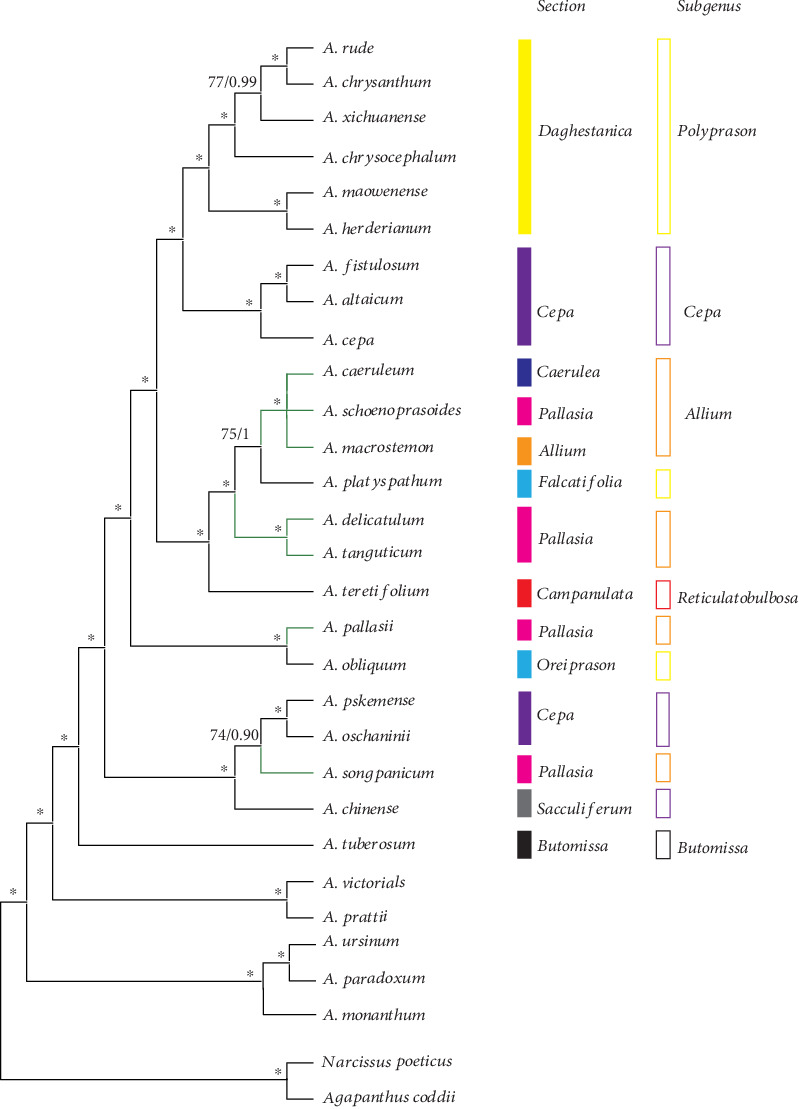
The phylogenetic relationships of these eight Allium species with other 22 related species based on whole genome sequences. Tree constructed by Bayesian inference (BI) and maximum likelihood (ML) with the posterior probabilities of BI and the bootstrap values of ML above the branches, respectively. ∗ represents maximum support in all two analyses.
3.6. The Ka/Ks Ratios of Species Pairwise
The pairwise Ka/Ks ratios of photosynthesis gene single-CDS of each species pair were calculated (Figure 6). Higher pairwise Ka/Ks ratios were observed between A. schoenoprasoides and A. macrostemon, A. schoenoprasoides, and A. caeruleum rather than that between A. macrostemon and A. caeruleum. A. delicatulum and A. tanguticum also had distinctive Ka/Ks ratios. There were higher pairwise Ka/Ks ratios in the clade formed by A. chinense, A. songpanicum, A. pskemense, and A. oschaninii.
Figure 6.
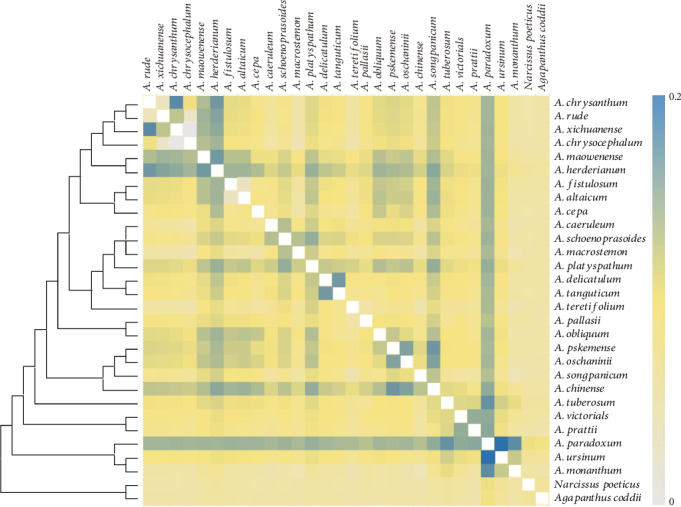
Pairwise Ka/Ks ratios in Allium (Allioideae) and their outgroups. This heat map shows pairwise Ka/Ks ratios between every sequence in the multigene nucleotide alignment.
4. Discussion
4.1. The Intergenetic Analysis of Species in Sect. Pallasia and Related Species
In this study, a phylogenetic tree with high support values was obtained by analyzing 30 cp genomes from Allium species. Three species (A. caeruleum, A. schoenoprasoides, and A. macrostemon) show well support in the cp genome phylogenetic tree, which had been found in the previous studies [4, 52], these three species all have distinctive common-possessed bulbs and special fistulose leaves, and these might demonstrate their close relationships [8, 53].
A. tanguticum and A. delicatulum used to be positioned into sect. Pallasia [4]; however, our phylogenetic analysis results show that A. tanguticum and A. delicatulum do not exhibit close relationship with the type species of sect. Pallasia (A. pallasii); phylogenetic results strongly support that these two species are closely clustered with A. platyspathum, A. caeruleum, A. schoenoprasoides, and A. macrostemon. But our results show that A. platyspathum from subgenus Polyprason sect. Falcatifolia has a close affinity with species in subgenus Allium, which is not consistent with the classification based on morphology. A. songpanicum used to be positioned into sect. Pallasia based on morphology (Li et al., 2010); it has a similar bulb like A. pallasii. However, our results show that A. songpanicum is clustered with A. chinense, A. pskemense, and A. oschaninii, and they present a monophyletic clade. A. pallasii used to be an isolated clade, and it was the type species of the sect. Pallasia in subgenus Allium, but it shows close relationship with A. obliquum from sect. Oreiprason in subgenus Polyprason [4, 6, 9], and it has far relationships with species in subgenus Allium. Therefore, the phylogenetic inference of A. pallasii is still unclear; more Allium species in Central Asia is needed to solve the phylogenetic position of it.
Homoplasious characters of these species may make it difficult to recognize their relationship from morphological characteristics [4, 6, 9]. However, the phylogenetic inference of these results remained as the ambiguous species relationship in the third evolutionary lineage of Allium; more individuals and morphological data are needed to further explore phylogenetic relationships of this complex group.
4.2. Comparative Analysis of Eight Chloroplast Genomes
By analyzing genome size, GC contents, and gene numbers, high genome conservation was detected. In all chloroplast genomes, IR regions had the highest GC content; this may attribute to the presence of eight rRNA sequences in these regions (Table 1). The rps16 gene and rps2 gene were annotated as pseudogenes in A. pallasii, but they were annotated as protein-coding genes in the other seven cp genomes; it was reported that rps16 was a pseudogene in A. obliquum and A. sativum, and the rps2 was a pseudogene in more species [54]. These studies may be helpful to systematically understand the gene number, gene order, and chloroplast genome structure of Allium species.
The change in position of the IR/LSC and IR/SSC boundary may be caused by contraction or expansion of the IR region, and it is common in most of angiosperms [24, 55, 56]. The IR/SSC and IR/LSC boundary regions in these Allium species showed similar characteristics, excepting A. pallasii, which exhibited differences compared with other seven species. For example, the length from psbA gene to the LSC/IRb boundary in A. pallasii (627 bp) was much longer than that in other species (67–127 bp). Other genes, such as rpl22, ycf1, and ndhF, also had some differences in length and position. The differences in A. pallasii were consistent with our phylogenetic analysis; these seem to reflect far phylogenetic relationships between A. pallasii and other species in our study. The LSC-IR borders of eight Allium species were different from those of Agapanthus coddii and Narcissus poeticus by showing expansions, which may help in the prevention of gene loss-and-gain events [57]; these expansion or contraction of IR into LSC/SSC regions can often be observed in angiosperm plastomes [20, 57, 58]. The expansion and contraction of the IR region are related to the mutational hotspots; the highly divergent region ycf1 is located in the SSC/IR boundary.
SSRs, thought to be the results of slipped strand mispairing during DNA replication, which are 16 bp repeating sequences in the chloroplast genome, have been always used as the molecular markers because of their high variability [18, 51, 59, 60]. The majority of these SSRs consisted of mono- and dinucleotide repeats, and most SSRs are located in the LSC region; the same results have appeared in previous research studies [28]. This uneven distribution of SSRs suggests that the difference of SSR numbers between A. pallasii and other species may be related to its short LSC region. For the wide use of SSRs in population studies, our new research could be used for studies on population genetics of this genus. Taking A. cepa as the reference, the mVISTA software was used to compare the complete chloroplast genome of these eight species. Seven highly divergent regions were selected for their greater nucleotide diversity, which might be more suitable sequences for developing potential molecular markers and species identification.
4.3. The Adaptation Evolution of Allium Species in Sect. Pallasia and Related Species
The pairwise Ka/Ks ratios are widely used as an effective way to detect positive selection or adaptive evolution in species, and the adaptation evolution of chloroplast genes to diverse ecological habitats of sunlight preferences has been reported in a recent study [27]. The Ka/Ks analysis of chloroplast genome genes in Allium species has also been reported, which suggested that Allium species may have undergone some selective forces in the evolutionary process [30].
Here, 42 photosynthesis genes of these 30 Allium species were concatenated, and higher pairwise Ka/Ks ratios were found in A. schoenoprasoides compared to A. caeruleum and A. macrostemon while a lower value of Ka/Ks ratios was detected between A. caeruleum and A. macrostemon, which may be caused by their different altitudes. A. caeruleum and A. macrostemon usually live in grass with an elevation of 500 m [5, 8], while A. schoenoprasoides lives in mountain pastures 3000 m above the sea level. A similar Ka/Ks ratio is also obtained between A. tanguticum (living in 3000 m) and A. delicatulum (living in 1500 m). The adaptation to different altitudes may lead to these higher pairwise Ka/Ks ratios. The Ka/Ks value difference among species is accompanied by great altitude difference; this seemed to reflect that the adaptive evolution of photosynthesis genes was mainly related to altitude or temperature, which has been identified in the previous study [61].
5. Conclusions
Here, we sequenced, assembled, and annotated eight chloroplast genomes of Allium with high-throughput sequencing technology. All eight cp genomes showed a typical quadripartite structure in length, gene content, gene order, and GC content similar among these sequences. Seven highly divergent regions were selected for their greater nucleotide diversity, which can be used to develop useful markers for future phylogenetic analysis. The maximum likelihood and BI phylogenetic results showed that A. caeruleum, A. schoenoprasoides, and A. macrostemon have close relationships with high support values.
Ka/Ks analysis indicated that the adaptation to different altitudes may lead to these higher pairwise Ka/Ks ratios. In conclusion, our results not only will be valuable to understanding the relationship between these eight Allium species but also provide useful cp genome resources for Allium phylogenetic study.
Acknowledgments
We thank Xin Yang, Juan Li, and Chang Peng in Sichuan University for their help in the collection of specimens. This work was supported by the National Natural Science Foundation of China (Grant Nos. 31872647 and 31570198), the Chinese Ministry of Science and Technology through the “National Science and Technology Infrastructure Platform” project (Grant No. 2005DKA21403-JK), and the fourth national survey of traditional Chinese medicine resources (Grant No. 2019PC002).
Data Availability
All data are openly available in Genebank, and the accession numbers have been showed in Supplementary Materials Table S1.
Conflicts of Interest
The authors declare no conflict of interest.
Authors' Contributions
For resources, most species are collected by F.-M.X., and A. songpanicum was collected by C.X.; writing and formal analysis were handled by F.-M.X.; review and editing were worked by D.-F.X.; methodology was secured by Y.Y. and D.-F.X.; supervision was conducted by X.-J.H.; and conceptualization was made by F.-M.X., Y.Y., S.-D.Z., and X.-J.H. All authors have read and agreed to the published version of the manuscript.
Supplementary Materials
Table S1: the accession number and collection locations of genomes in this study; genome of Allium macrostemon was sequenced in our previous study. Table S2: contig information generated by NOVOPlasty. Table S3: the accession number of the other species used in the phylogenetic analysis. Table S4: RSCU value in eight Allium chloroplast genome CDS. Figure S1: the phylogenetic relationships of these eight Allium species with other 22 related species based on combined coding sequences (A) and LSC regions (B). Tree constructed by Bayesian inference (BI) and maximum likelihood (ML) with the posterior probabilities of BI and the bootstrap values of ML above the branches, respectively. ∗ represent maximum support in all two analyses. Figure S2: pairwise Ka/Ks ratios of rbcL gene in Allium (Allioideae) and their outgroups (A). Pairwise Ka/Ks ratios of clpP gene in Allium (Allioideae) and their outgroups (B).
References
- 1.Hauenschild F., Favre A., Schnitzler J., Michalak I., Freiberg M., Muellner-Riehl A. N. Spatio-temporal evolution of _Allium_ L. in the Qinghai -Tibet-Plateau region: Immigration and _in situ_ radiation. Plant Diversity. 2017;39(4):167–179. doi: 10.1016/j.pld.2017.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society. 2016;181(1):1–20. doi: 10.1111/boj.12385. [DOI] [Google Scholar]
- 3.Herden T., Hanelt P., Friesen N. Phylogeny of _Allium_ L. subgenus _Anguinum_ (G. Don. ex W.D.J. Koch) N. Friesen (Amaryllidaceae) Molecular Phylogenetics and Evolution. 2016;95:79–93. doi: 10.1016/j.ympev.2015.11.004. [DOI] [PubMed] [Google Scholar]
- 4.Li Q. Q., Zhou S. D., He X. J., Yu Y., Zhang Y. C., Wei X. Q. Phylogeny and biogeography of Allium (Amaryllidaceae: Allieae) based on nuclear ribosomal internal transcribed spacer and chloroplast rps 16 sequences, focusing on the inclusion of species endemic to China. Annals of Botany. 2010;106(5):709–733. doi: 10.1093/aob/mcq177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Block E. Garlic and Other Alliums: The Lore and the Science. Choice Reviews Online. 2010;47(10):p. 47-5636. doi: 10.5860/choice.47-5636. [DOI] [Google Scholar]
- 6.Friesen N., Fritsch R., Blattner F. Phylogeny and new intrageneric classification of Allium (Alliaceae) based on nuclear ribosomal DNA ITS sequences. Aliso. 2006;22(1):372–395. doi: 10.5642/aliso.20062201.31. [DOI] [Google Scholar]
- 7.He X. J., Ge S., Xu J. M., Hong D. Y. Phylogeny of Chinese Allium (Liliaceae) using PCR-RFLP analysis. Science in China Series C-Life Sciences. 2000;43(5):454–463. [Google Scholar]
- 8.Xu J. M. R. V. K. Flora of China. 2013;24 [Google Scholar]
- 9.Fritsch R. M., Blattner F. R., Gurushidze M. New classification of Allium L. subg. Melanocrommyum (WEBB & BERTHEL.) ROUY (Alliaceae) based on molecular and morphological characters. Phyton-Annales Rei Botanicae. 2010;49(2):145–220. [Google Scholar]
- 10.Howe C. J., Barbrook A. C., Koumandou V. L., Nisbet R. E. R., Symington H. A., Wightman T. F. Evolution of the chloroplast genome. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences. 2003;358(1429):99–106. doi: 10.1098/rstb.2002.1176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Raven J. A., Allen J. F. Genomics and chloroplast evolution: what did cyanobacteria do for plants? Genome Biology. 2003;4(3) doi: 10.1186/gb-2003-4-3-209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sugiura M. The chloroplast genome. Plant Molecular Biology. 1992;19(1):149–168. doi: 10.1007/BF00015612. [DOI] [PubMed] [Google Scholar]
- 13.Asaf S., Khan A. L., Khan A. R., et al. Complete chloroplast genome of Nicotiana otophora and its comparison with related species. Frontiers in Plant Science. 2016;7 doi: 10.3389/fpls.2016.00843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Carbonell-Caballero J., Alonso R., Ibanez V., Terol J., Talon M., Dopazo J. A phylogenetic analysis of 34 chloroplast genomes elucidates the relationships between wild and domestic species within the genus Citrus. Molecular Biology and Evolution. 2015;32(8):2015–2035. doi: 10.1093/molbev/msv082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chaney L., Mangelson R., Ramaraj T., Jellen E. N., Maughan P. J. The complete chloroplast genome sequences for FourAmaranthusSpecies (Amaranthaceae) Applications in Plant Sciences. 2016;4(9) doi: 10.3732/apps.1600063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chen Q., Wu X., Zhang D. Phylogenetic analysis ofFritillaria cirrhosaD. Don and its closely related species based on complete chloroplast genomes. Peerj. 2019;7:p. e7480. doi: 10.7717/peerj.7480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Du Y.-p., Bi Y., Yang F.-p., et al. Complete chloroplast genome sequences of _Lilium_ : insights into evolutionary dynamics and phylogenetic analyses. Scientific Reports. 2017;7(1):p. 5751. doi: 10.1038/s41598-017-06210-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hu Y., Woeste K. E., Zhao P. Completion of the chloroplast genomes of five Chinese Juglans and their contribution to chloroplast phylogeny. Frontiers in Plant Science. 2017;7 doi: 10.3389/fpls.2016.01955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yang Y., Zhou T., Duan D., Yang J., Feng L., Zhao G. Comparative analysis of the complete chloroplast genomes of five Quercus species. Frontiers in Plant Science. 2016;7 doi: 10.3389/fpls.2016.00959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhai W., Duan X., Zhang R., et al. Chloroplast genomic data provide new and robust insights into the phylogeny and evolution of the Ranunculaceae. Molecular Phylogenetics and Evolution. 2019;135:12–21. doi: 10.1016/j.ympev.2019.02.024. [DOI] [PubMed] [Google Scholar]
- 21.Mariac C., Scarcelli N., Pouzadou J., et al. Cost-effective enrichment hybridization capture of chloroplast genomes at deep multiplexing levels for population genetics and phylogeography studies. Molecular Ecology Resources. 2014;14(6):1103–1113. doi: 10.1111/1755-0998.12258. [DOI] [PubMed] [Google Scholar]
- 22.Martin W., Deusch O., Stawski N., Grunheit N., Goremykin V. Chloroplast genome phylogenetics: why we need independent approaches to plant molecular evolution. Trends in Plant Science. 2005;10(5):203–209. doi: 10.1016/j.tplants.2005.03.007. [DOI] [PubMed] [Google Scholar]
- 23.Yuan C., Zhong W., Mou F., et al. The complete chloroplast genome sequence and phylogenetic analysis of Chuanminshen (Chuanminshenviolaceum Sheh et Shan) Physiology and Molecular Biology of Plants. 2017;23(1):35–41. doi: 10.1007/s12298-016-0395-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Davis J. I., Soreng R. J. Migration of endpoints of two genes relative to boundaries between regions of the plastid genome in the grass family (Poaceae) American Journal of Botany. 2010;97(5):874–892. doi: 10.3732/ajb.0900228. [DOI] [PubMed] [Google Scholar]
- 25.Dyall S. D., Brown M. T., Johnson P. J. Ancient invasions: from endosymbionts to organelles. Science. 2004;304(5668):253–257. doi: 10.1126/science.1094884. [DOI] [PubMed] [Google Scholar]
- 26.Wicke S., Schneeweiss G. M., dePamphilis C. W., Müller K. F., Quandt D. The evolution of the plastid chromosome in land plants: gene content, gene order, gene function. Plant Molecular Biology. 2011;76(3-5):273–297. doi: 10.1007/s11103-011-9762-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gao L. Z., Liu Y. L., Zhang D., et al. Evolution of _Oryza_ chloroplast genomes promoted adaptation to diverse ecological habitats. Communications Biology. 2019;2(1) doi: 10.1038/s42003-019-0531-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yang X., Xie D. F., Chen J. P., Zhou S. D., Yu Y., He X. J. Comparative analysis of the complete chloroplast genomes in Allium subgenus Cyathophora (Amaryllidaceae): phylogenetic relationship and adaptive evolution. Biomed Research International. 2020;2020:17. doi: 10.1155/2020/1732586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Li P., Lu R. S., Xu W. Q., et al. Comparative genomics and phylogenomics of East Asian tulips (Amana, Liliaceae) Frontiers in Plant Science. 2017;8 doi: 10.3389/fpls.2017.00451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Xie D.-F., Yu H.-X., Price M., et al. Phylogeny of Chinese Allium species in section Daghestanica and adaptive evolution of Allium (Amaryllidaceae, Allioideae) species revealed by the chloroplast complete genome. Frontiers in Plant Science. 2019;10 doi: 10.3389/fpls.2019.00460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Omelchenko D. O., Krinitsina A. A., Belenikin M. S., et al. Complete plastome sequencing of _Allium paradoxum_ reveals unusual rearrangements and the loss of the _ndh_ genes as compared to _Allium ursinum_ and other onions. Gene. 2020;726:p. 144154. doi: 10.1016/j.gene.2019.144154. [DOI] [PubMed] [Google Scholar]
- 32.Doyle J. L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin. 1987;19:11–15. [Google Scholar]
- 33.Dierckxsens N., Mardulyn P., Smits G. NOVOPlasty:de novoassembly of organelle genomes from whole genome data. Nucleic Acids Research. 2017;45(4):p. gkw955. doi: 10.1093/nar/gkw955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kearse M., Moir R., Wilson A., et al. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 2012;28(12):1647–1649. doi: 10.1093/bioinformatics/bts199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Qu X. J., Moore M. J., Li D. Z., Yi T. S. PGA: a software package for rapid, accurate, and flexible batch annotation of plastomes. Plant Methods. 2019;15(1) doi: 10.1186/s13007-019-0435-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lohse M., Drechsel O., Kahlau S., Bock R. OrganellarGenomeDRAW-a suite of tools for generating physical maps of plastid and mitochondrial genomes and visualizing expression data sets. Nucleic Acids Research. 2013;41(W1):W575–W581. doi: 10.1093/nar/gkt289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Beier S., Thiel T., Munch T., Scholz U., Mascher M. MISA-web: a web server for microsatellite prediction. Bioinformatics. 2017;33(16):2583–2585. doi: 10.1093/bioinformatics/btx198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Katoh K., Standley D. M. MAFFT Multiple Sequence Alignment Software Version 7: improvements in performance and usability. Molecular Biology and Evolution. 2013;30(4):772–780. doi: 10.1093/molbev/mst010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kumar S., Stecher G., Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Molecular Biology and Evolution. 2016;33(7):1870–1874. doi: 10.1093/molbev/msw054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Stamatakis A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014;30(9):1312–1313. doi: 10.1093/bioinformatics/btu033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ronquist F., Teslenko M., van der Mark P., et al. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology. 2012;61(3):539–542. doi: 10.1093/sysbio/sys029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Frazer K. A., Pachter L., Poliakov A., Rubin E. M., Dubchak I. VISTA: computational tools for comparative genomics. Nucleic Acids Research. 2004;32(Web Server):W273–W279. doi: 10.1093/nar/gkh458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Rozas J., Ferrer-Mata A., Sánchez-DelBarrio J. C., et al. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Molecular Biology and Evolution. 2017;34(12):3299–3302. doi: 10.1093/molbev/msx248. [DOI] [PubMed] [Google Scholar]
- 44.Wang D., Zhang Y., Zhang Z., Zhu J., Yu J. KaKs_Calculator 2.0: a toolkit incorporating gamma-series methods and sliding window strategies. Genomics, proteomics & bioinformatics. 2010;8(1):77–80. doi: 10.1016/s1672-0229(10)60008-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Cavalier-Smith T. Chloroplast evolution: secondary symbiogenesis and multiple losses. Current Biology. 2002;12(2):R62–R64. doi: 10.1016/s0960-9822(01)00675-3. [DOI] [PubMed] [Google Scholar]
- 46.Kolodner R., Tewari K. K. Inverted repeats in chloroplast DNA from higher plants. Proceedings of the National Academy of Sciences of the United States of America. 1979;76(1):41–45. doi: 10.1073/pnas.76.1.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wang R. J., Cheng C. L., Chang C. C., Wu C. L., Su T. M., Chaw S. M. Dynamics and evolution of the inverted repeat-large single copy junctions in the chloroplast genomes of monocots. Bmc Evolutionary Biology. 2008;8(1):p. 36. doi: 10.1186/1471-2148-8-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hong S.-Y., Cheon K.-S., Yoo K.-O., et al. Complete chloroplast genome sequences and comparative analysis of Chenopodium quinoa and C. album. Frontiers in Plant Science. 2017;8 doi: 10.3389/fpls.2017.01696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Song Y., Chen Y., Lv J., et al. Development of chloroplast genomic resources for Oryza species discrimination. Frontiers in Plant Science. 2017;8 doi: 10.3389/fpls.2017.01854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wu Y., Liu F., Yang D. G., et al. Comparative chloroplast genomics of Gossypium species: insights into repeat sequence variations and phylogeny. Frontiers in Plant Science. 2018;9 doi: 10.3389/fpls.2018.00376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Xie D. F., Yu Y., Deng Y. Q., et al. Comparative analysis of the chloroplast genomes of the Chinese endemic Genus Urophysa and their contribution to chloroplast phylogeny and adaptive evolution. International Journal of Molecular Sciences. 2018;19(7):p. 1847. doi: 10.3390/ijms19071847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Huang D., Qin-Qin L. I., Zhou S., Xing-Jin H. E. Systematic position of Allium macrostemon based on nr DNA ITS and cp DNA trn L-F sequence data. Plant Diversity & Resources. 2015 [Google Scholar]
- 53.Choi H. J., Giussani L. M., Jang C. G., Oh B. U., Cota-Sanchez J. H. Systematics of disjunct northeastern Asian and northern North AmericanAllium(Amaryllidaceae) Botany. 2012;90(6):491–508. doi: 10.1139/b2012-031. [DOI] [Google Scholar]
- 54.Huo Y., Gao L., Liu B., et al. Complete chloroplast genome sequences of four _Allium_ species: comparative and phylogenetic analyses. Scientific Reports. 2019;9(1):p. 12250. doi: 10.1038/s41598-019-48708-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Li R., Ma P. F., Wen J., Yi T. S. Complete sequencing of five Araliaceae chloroplast genomes and the phylogenetic implications. Plos One. 2013;8(10):p. e78568. doi: 10.1371/journal.pone.0078568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Yi X., Gao L., Wang B., Su Y. J., Wang T. The complete chloroplast genome sequence of Cephalotaxus oliveri (Cephalotaxaceae): evolutionary comparison of Cephalotaxus chloroplast DNAs and insights into the loss of inverted repeat copies in gymnosperms. Genome Biology and Evolution. 2013;5(4):688–698. doi: 10.1093/gbe/evt042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ye W. Q., Yap Z. Y., Li P., Comes H. P., Qiu Y. X. Plastome organization, genome-based phylogeny and evolution of plastid genes in Podophylloideae (Berberidaceae) Molecular Phylogenetics and Evolution. 2018;127:978–987. doi: 10.1016/j.ympev.2018.07.001. [DOI] [PubMed] [Google Scholar]
- 58.Huang J., Yu Y., Liu Y. M., Xie D. F., He X. J., Zhou S. D. Comparative chloroplast genomics of Fritillaria (Liliaceae), inferences for phylogenetic relationships between Fritillaria and Lilium and plastome evolution. Plants-Basel. 2020;9(2):p. 133. doi: 10.3390/plants9020133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Bryan G. J., McNicoll J., Ramsay G., Meyer R. C., De Jong W. S. Polymorphic simple sequence repeat markers in chloroplast genomes of Solanaceous plants. Theoretical and Applied Genetics. 1999;99(5):859–867. doi: 10.1007/s001220051306. [DOI] [Google Scholar]
- 60.de Cesare M., Hodkinson T. R., Barth S. Chloroplast DNA markers (cpSSRs, SNPs) for Miscanthus, Saccharum and related grasses (Panicoideae, Poaceae) Molecular Breeding. 2010;26(3):539–544. doi: 10.1007/s11032-010-9451-z. [DOI] [Google Scholar]
- 61.Munné-Bosch S., Cotado A., Morales M., Fleta-Soriano E., Villellas J., Garcia M. B. Adaptation of the Long-Lived Monocarpic Perennial, Saxifraga longifolia to High Altitude. Plant Physiology. 2016;172:00877.2016–00877.2775. doi: 10.1104/pp.16.00877. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1: the accession number and collection locations of genomes in this study; genome of Allium macrostemon was sequenced in our previous study. Table S2: contig information generated by NOVOPlasty. Table S3: the accession number of the other species used in the phylogenetic analysis. Table S4: RSCU value in eight Allium chloroplast genome CDS. Figure S1: the phylogenetic relationships of these eight Allium species with other 22 related species based on combined coding sequences (A) and LSC regions (B). Tree constructed by Bayesian inference (BI) and maximum likelihood (ML) with the posterior probabilities of BI and the bootstrap values of ML above the branches, respectively. ∗ represent maximum support in all two analyses. Figure S2: pairwise Ka/Ks ratios of rbcL gene in Allium (Allioideae) and their outgroups (A). Pairwise Ka/Ks ratios of clpP gene in Allium (Allioideae) and their outgroups (B).
Data Availability Statement
All data are openly available in Genebank, and the accession numbers have been showed in Supplementary Materials Table S1.