

Abstract
Inhibitory control is crucial for regulating emotions and may also enable memory control. However, evidence for their shared neurobiological correlates is limited. Here, we report meta-analyses of neuroimaging studies on emotion regulation, or memory control and link neural commonalities to transcriptional commonalities using the Allen Human Brain Atlas (AHBA). Based on 95 functional magnetic resonance imaging studies, we reveal a role of the right inferior parietal lobule embedded in a frontal–parietal–insular network during emotion regulation and memory control, which is similarly recruited during response inhibition. These co-activation patterns also overlap with the networks associated with ‘inhibition’, ‘cognitive control’ and ‘working memory’ when consulting the Neurosynth. Using the AHBA, we demonstrate that emotion regulation- and memory control-related brain activity patterns are associated with transcriptional profiles of a specific set of ‘inhibition-related’ genes. Gene ontology enrichment analysis of these ‘inhibition-related’ genes reveal associations with the neuronal transmission and risk for major psychiatric disorders as well as seizures and alcoholic dependence. In summary, this study identified a neural network and a set of genes associated with inhibitory control across emotion regulation and memory control. These findings facilitate our understanding of the neurobiological correlates of inhibitory control and may contribute to the development of brain stimulation and pharmacological interventions.
Keywords: emotion regulation, memory control, inhibitory control, gene expression, transcriptional network
Introduction
One of the most influential cognitive theories of emotion regulation proposes that it is primarily supported by inhibitory control (Ochsner and Gross, 2005; Gross and Thompson, 2007). A potentially related cognitive process, memory control, is defined as an ability to actively reduce the accessibility of memories. Memory control is also thought to be supported by inhibitory control (Anderson and Green, 2001; Anderson, 2004; Anderson and Hanslmayr, 2014). Recently, Engen and Anderson (2018) summarized the conceptual link between emotion regulation and memory control and raised the question of whether there is a common neurobiological mechanism supporting these two processes. Here, we set out to empirically demonstrate the neurobiological commonalities between the two processes by first analyzing functional magnetic resonance imaging (fMRI) data and then linking neuroimaging findings with postmortem gene expression data.
The idea that inhibitory control, as the fundamental cognitive process, supports other higher-level processes such as emotion regulation and memory control is supported by behavioral and neural evidence. Behaviorally, a positive correlation between emotion regulation and memory control performances has been found (Depue et al., 2016). Human neuroimaging results showed overlapping recruitment of right superior medial frontal gyrus and right inferior frontal gyrus in both emotion regulation (Ochsner et al., 2002; Buhle et al., 2014; Kohn et al., 2014) and memory control (Anderson, 2004; Guo et al., 2018). Beyond these frontal regions, the parietal cortex is another candidate region that may play a critical role in both emotion regulation and memory control. Starting from the idea of multiple-demand system in the brain (Duncan, 2010), both the evidence from task fMRI (Fedorenko et al., 2013) and resting-state fMRI (Dosenbach et al., 2007; Power et al., 2011) suggest that a fronto-parietal network supports the initiation of top-down control and control adjustment in response to task goals and feedbacks.
No previous formal meta-analysis of human neuroimaging studies has investigated the neural commonalities between emotion regulation and memory control to pinpoint the overlap in inhibitory control, although common neural correlates between emotion regulation or memory control with response to inhibition tasks (e.g. stop-signal and go/no-go task) have been investigated in two recent meta-analyses (Guo et al., 2018; Langner et al., 2018). Using activation likelihood estimation (ALE), we aim to demonstrate that emotion regulation and memory control evoke activation of similar brain regions with a spatial pattern that is similar to the activation of typical response inhibition paradigms, including stop-signal and go/no-go tasks. Moreover, beyond overlapping regional brain activity, we also are interested in overlapping co-activation patterns of associated brain regions. We used meta-analytic connectivity modeling (MACM) to test whether these brain regions act as an interconnected functional network. We expected to find a similar set of co-activated brain regions that are associated with both emotion regulation and memory control, defining a core ‘inhibition-related’ network.
Identifying an underlying neural network is relevant for potential brain stimulation interventions including but not limited to transcranial magnetic stimulation (TMS) and transcranial direct current stimulation (tDCS). However, further dissection of molecular underpinnings is necessary to expand our understanding of inhibition control, which in turn may pave the way for pharmacological interventions. Therefore, we reasoned whether emotion regulation and memory control are not only related to a common neural network defined by activity and connectivity, but also by associated similarities in spatial transcriptional profiles. Conventional imaging genetic methods including candidate gene methods and genome-wide association approaches cannot reveal the relationship between localized gene transcription and task-related brain activity, even though they already showed the association between multiple common gene variants and neuroimaging measures (Hariri et al., 2002; Elliott et al., 2018). Thus, to explore the relationship between spatial transcriptional profiles and emotion regulation or memory control-related brain activity, we adopted a recently developed approach to associate spatial maps of gene expression in postmortem brain tissues with brain activation measures (Gorgolewski et al., 2014; Kong et al., 2017). Spatial pattern analysis of gene expression maps of the Allen Human Brain Atlas (AHBA) together with neuroimaging revealed fundamental features of transcriptional regulation (see review by Fornito et al., 2019) and related disruptions in brain disorders (Romme et al., 2017; Grothe et al., 2018; McColgan et al., 2018; Romero-Garcia et al., 2019). However, the correspondence between spatial transcriptional profiles and neural functionality has not yet provided full insight into how genetic correlates are linked to core cognitive abilities (e.g. inhibitory control in this study). Based on the assumption that spatial transcriptional profiles not only co-vary with the connectional architecture but also support the task-evoked, synchronous brain activity (Gorgolewski et al., 2014; Kong et al., 2017; Berto et al., 2018; Shine et al., 2019), we expected to find similar spatial transcriptional profiles between emotion regulation, memory control and response inhibition.
To investigate the neural and transcriptional commonality of emotion regulation and memory control, we combined task fMRI data, neuroimaging meta-analytic approaches and postmortem gene expression data. First, we used the ALE method to generate the task activation map for each paradigm of interest (e.g. emotion regulation or memory control) based on 95 published task fMRI studies with in total 1995 healthy participants (Figure 1A). Then, we used the brain regions identified by the ALE as seed regions and estimated the meta-analytic connectivity map (or co-activation map), using information from BrainMap (Figure 1B). And to explore the behavioral relevance of these findings, we examined the associations of these co-activation patterns in a data-driven way via Neurosynth with behavioral domains (Figure 1C). Next, we calculated the spatial association between task activation maps and human gene expression maps to identify their common spatial transcriptional profiles (Figure 1D). Finally, we performed a systematic and integrative analysis of the resulting gene list to gain an in-depth understanding of putative biological functions and disease associations of the identified ‘inhibition-related’ gene set (Figure 1E).
Fig. 1.
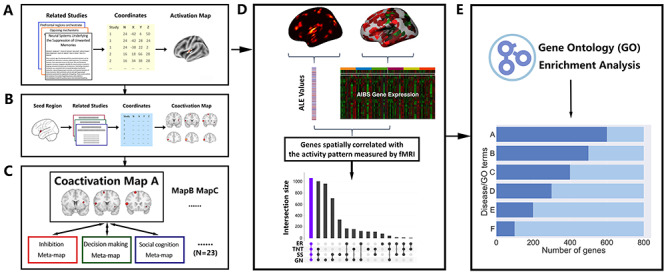
Schematic of the pipeline for the investigation of neural and transcriptional commonality. (A) ALE meta-analyses of functional MRI studies. MRI coordinates of reported brain activations from relevant studies were used to generate the activation map for each task of interest. (B) MACM of seed regions resulting from the ALE analyses was conducted to search co-activated regions over all studies in BrainMap. Co-activation maps were estimated via ALE. (C) To generate behavioral profiles of co-activation maps, we used Neurosynth to acquire a list of meta-maps, including the behavioral terms from low-level to high-level cognitive processes (total number of terms = 23). Next, each co-activation map was compared to all meta-maps (e.g. inhibition, decision-making and social cognition) to compute the similarity index for each meta-map. (D) The activation patterns were associated with gene expression maps from the AHBA. Each gene expression map was vectorized based on the expression measures within the three-dimensional maps. The ALE values at the corresponding brain regions on the activation map were extracted and vectorized. To identify the genes whose spatial patterns are similar to a specific activation map, the similarity between all vector pairs were quantified. ‘Inhibitory-related’ genes were defined by identifying genes of which the expression patterns are correlated with the activation maps of four paradigms (ER, emotion regulation; TNT, think/no-think; SS, stop-signal; GN, go/no-go). (E) ‘Inhibition-related’ genes were associated with biological functions (GO terms) and disease terms for the interpretation using gene ontology enrichment analysis (GOEA).
Materials and methods
Literature searches, selection and coordinates extraction
In total, we performed literature searches for four task paradigms (think/no-think, emotion regulation, stop-signal, and go/no-go) and one network [‘default mode network’ (DMN)]. To avoid biases, we used the following inclusion criteria during the search:
(i) We only included data from studies on healthy adults with no prior report of neurological, medical or psychiatric disorders in the current meta-analysis, while results of patients or specific sub-group effects (e.g. gender differences) were not included. Articles including patients were only selected if they reported results for a control group separately, and only the control group was included here.
(ii) Only neuroimaging studies, which used whole-brain fMRI and reported coordinates for brain activation or deactivation in standard anatomical reference space [Talairach/Tournoux; Montreal Neurological Institute (MNI)] were considered. Coordinates originally published in Talairach space were converted to MNI space using the algorithm implemented in GingerALE 3.0.2 (http://www.brainmap.org/ale/).
(iii) Only studies reporting results of general linear model contrasts were included, while studies focusing on functional connectivity, structural, resting-state or brain-behavior correlations were excluded.
(iv) Only studies reporting whole-brain analyses were included, while studies based on partial coverage or employing only region-of-interest analyses were excluded.
Detailed search and extraction procedures were as follows for each paradigm:
Think/no-think studies
A step-wise procedure was used to search articles, published before February 2020, using functional MRI to investigate brain activity during think/no-think paradigm. First, we used standard search in PubMed and ISI Web of Science to perform the search. More specifically, we used the combination of the following key words during the search: ‘memory regulation’, ‘memory control’, ‘memory suppression regulation’, ‘memory inhibition’, ‘think/no-think’, ‘fMRI’, ‘neuroimaging’, ‘functional magnetic resonance imaging’ or ‘functional MRI’. At the same time, we carefully exclude studies using the ‘directed forgetting’ paradigm, which targets at the memory control during the encoding. Next, two lab members compared the search results with a recent review article (Anderson and Hanslmayr, 2014) to find additional relevant studies. The same two lab members independently extracted the coordinates and other essential information (e.g. sample size, type of stimulus) extraction based on the identified think/no-think literature and then cross-validated the coordinates. In summary, this search and inclusion/exclusion criteria led to 15 think/no-think studies (491 subjects and 256 foci).
Emotion regulation studies
We used the databases of previously published meta-analyses on emotion regulation (Kohn et al., 2014; Morawetz et al., 2017). We used the key words: ‘emotion regulation’, ‘affective regulation’, ‘implicit emotion regulation’, ‘explicit emotion regulation’, ‘interpersonal emotion regulation’, ‘extrinsic emotion regulation, ‘intrinsic emotion regulation’, ‘reappraisal’, ‘suppression’, ‘distraction’, ‘detachment’, ‘labelling’, ‘affective labelling’, ‘reinterpretation’, ‘rumination’, ‘fMRI’, ‘neuroimaging’, ‘functional magnetic resonance imaging’ or ‘functional MRI’. In the case that a study did not report the contrast of interest for this meta-analysis, the corresponding authors were contacted and asked to provide more information on their data. The term ‘experiment’ refers to any single contrast analysis, while the term ‘study’ refers to a scientific publication, usually reporting several contrasts, i.e. experiments. This search and the employed inclusion/exclusion criteria led to a total inclusion of 107 studies from peer-reviewed journals on 31 July 2017 (385 experiments, 3204 participants).
Each experiment was manually coded by the authors of the previous meta-analysis (C.M. and N.K) with terms that described the experimental design with respect to contrast, stimulus type utilized, emotion regulation strategy, goal of the strategy, valence of the stimuli, tactics of the strategy and the task nature. To achieve a more appropriate comparison between think/no-think and emotion regulation, we restricted inclusion to ER studies that used the ‘suppression’ or ‘distraction’ strategy. This led to a similar amount of studies compared to memory control, and most crucially suppression or distraction of emotions is conceptually also closer to the process of suppressing memories. These criteria led to the inclusion of 15 emotion regulation studies (387 subjects and 165 foci).
Go/no-go and stop-signal studies
A similar procedure was used to search published whole-brain functional MRI studies using the ‘go/no-go’ and ‘stop-signal’ paradigm. To confirm the completeness of our search, we compared our results with used studies in a recent meta-analysis of motor inhibitory and memory control (Guo et al., 2018). Again, two lab members (W.L and N.P) performed the coordinates and study information extraction for the go/no-go and stop-signal studies.
Default mode network (task-negative network) identification
We also performed a coordinated-based meta-analysis to identify the DMN. Instead of manually searching related studies and extracting coordinates, we used the BrainMap database (Fox and Lancaster, 2002; Laird et al., 2005) to find peak coordinates of task-independent deactivation reported in neuroimaging literature. This method was used before by Laird et al. (2009b) to identify the core regions in DMN. More specifically, we searched the BrainMap for all contrasts that were labeled as ‘deactivation’ and ‘low-level control’ during submission. ‘Deactivation’ refers to contrasts in which stronger signal was observed during a baseline condition than during task condition (e.g. control task); ‘low-level control’ are conditions in which either fixation or resting was defined as the baseline. Our search was further limited to ‘normal mapping’, which means that participants who are diagnosed with disease or disorders were excluded. In total, 105 studies (1588 foci) matched our search criteria and were used in the following analyses.
Activation likelihood estimation analyses
The ALE analyses were based on the revised ALE algorithm (Eickhoff et al., 2009) in GingerALE 2.3. Firstly, two separate meta-analyses were conducted for the think/no-think (contrast, no-think vs think) and emotion regulation (contrast, regulation vs view) tasks using cluster-level inference (FWE cluster-level correction P < 0.05, uncorrected cluster-forming threshold P < 0.001, threshold permutations = 1000). Secondly, contrast analyses (Eickhoff et al., 2011) were conducted between the think/no-think and emotion regulation tasks. In these contrast analyses, the thresholded activation maps from the two separate analyses, as well as the pooled results from both tasks, were used as inputs. Conjunction and contrast maps between the conditions were given as output. For the output images, the same cluster-level threshold correction was used (FWE cluster-level correction P < 0.05, uncorrected cluster-forming threshold P < 0.001, threshold permutations = 1000).
Additionally, meta-analyses of published fMRI studies using the stop-signal and go/no-go paradigms were also performed. The same software (GingerALE 2.3) and threshold (cluster-level P < 0.05) was used to perform the analysis. Due to the unbalanced number of studies and subjects included, no conjunction or contrast analyses were performed for four tasks to identify the overlap.
Co-activation analyses using BrainMap
We conducted the MACM analyses on the regions from the ALE meta-analysis. More specifically, for each ROI, we used the BrainMap database (Laird et al., 2009a, 2011) to search for experiments that also activated the particular ROI. Next, we retrieved all foci reported in the identified experiments. Finally, ALE analyses were performed over these foci to identify regions of significant convergence. Sequentially, raw co-activation maps were corrected for multiple comparisons [voxel-wise false discovery rate (FDR) < 0.05. All clusters sizes >200 mm3].
Functional profiles of the co-activation maps
To assess associated functional terms of the co-activation maps generated by MACM, we used the Neurosynth meta-analytic database (www.neurosynth.org) (Yarkoni et al., 2011). We followed the methodology used in a previous study to assess topic terms associated with the principal connectivity gradient in the human brain (Margulies et al., 2016). More specifically, we conducted a term-based meta-analysis for the same list of Neurosynth topic terms as Margulies and colleagues did. This list covered well-studied functional terms from low-level cognition (e.g. visual perception and auditory) to high-level cognition (e.g. language and rewarding). Sequentially, we examined the association between these term-based activation maps with our four sets of co-activation maps (emotion regulation, think/no-think, stop-signal, go/no-go). For each co-activation map, a spatial similarity index (r statistic) between each co-activation map and each meta-map of the functional term was provided. The terms were then ordered based on the average correlation for interpretation and visualization.
Activation–gene expression association analysis
We used a recently developed activation–gene expression association analysis to link the task-related brain activity to gene expression in postmortem human brains. This analysis can identify a list of associated genes based on the MRI-space statistical map(s). This analysis presupposes that if certain genes are associated with the cognitive task of interest, then spatial distributions of their expression values and task-related activation pattern measured by functional MRI should be similar.
The Allen Human Brain Atlas (http://www.brainmap.org) was used in the gene expression decoding analysis. The atlas provided genome-wide microarray-based gene expression data based on six postmortem human brains (gene expression level of over 62 000 gene probes for around 1000 sampling sites across the whole brain were recorded) (Hawrylycz et al., 2012). Additionally, structural brain imaging data of each donor was collected and provided, which enable users to visualize gene expression in its naive space and perform the registration to the standard MRI MNI space.
Previous studies used slightly different statistical methods to associate task-independent MRI-based brain measures (e.g. cortical thickness, functional or structural networks) with the gene expression data (Richiardi et al., 2015; Wang et al., 2015; Seidlitz et al., 2018). We used the method developed by Gorgolewski and colleagues implemented in the alleninfo tool (Gorgolewski et al., 2014) (https://github.com/chrisfilo/alleninf). This method was originally designed for the association analysis between voxel-wise statistical maps and gene expression maps. This method was also by default implemented in Neurovault (Gorgolewski et al., 2015) (https://neurovault.org/). The method has two important features: (i) nonlinear coregistration of the donor’s brain with MNI space was allowed and (ii) the ability to use a random-effects model makes it possible to generalize the results to the whole population. The activation–gene expression association analysis works as follows: (i) data from gene probes were aggregated for each gene, resulting in 20 787 gene expression maps. (ii) For each gene expression map, MNI coordinates of each sampling location (the locations in which brain tissues were analyzed for the gene expression data) were extracted to draw a spherical ROI (r = 4 mm). We used these ROIs to extract the average values of the ALE statistical map within each ROI. Next, the gene expression and meta-analysis vectors were correlated. (iii) This extraction and correlation procedure was repeated for each gene expression map to quantify the spatial pattern similarity between the statistical maps and gene expression map. (iv) Different thresholds (i.e. 500, 1000, 1500, 2000) were implemented to generate the significantly associated gene list(s) among the 20 787 genes. We mainly presented results under the threshold of 2000 most significant genes, but results under other thresholds can be found in the Supplementary Material. Since the negative correlation between brain measures and gene expression is difficult to explain, we only considered the positively correlated genes. Additionally, because of fundamental differences in gene expression between cortical and subcortical regions, we only performed our analyses within the cortical regions.
The described association analysis takes MRI statistical map(s) as input(s) and will output a list of significantly associated genes. To investigate the common transcriptional signatures (associated genes) of emotion regulation and memory control, we used the unthresholded statistical maps from the ALE analyses to identify the task-associated gene list via the association algorithm. Next, the common gene list was generated by overlapping the emotion regulation-associated gene list and memory control-associated gene list. To further investigate whether the two gene lists significantly overlapped with each other, we generated the null distribution of the number of overlapping genes by creating two gene lists (with the same size as the real gene lists) from the 20 787 genes. To control for autocorrelation, we generated 5000 lists in this way, and for each we estimated the overall spatial similarity between identified genes (i.e. emotion regulation-related and memory control-related genes) for each donor (Supplementary Table S3). Identified genes demonstrated modest spatial correlations (mean = 0.063, s.d. = 0.27) across different thresholds and donors. Based on these correlation values, 1718 out of 5000 pairs of randomly generated gene lists with a similarity ranging from 0.05 to 0.07 and the standard deviation ranging from 0.25 to 0.29 were further selected. The significance of overlapping genes was estimated by comparing the real number of overlapping genes with the number of overlapping genes within these 1718 pairs.
An alternative method (i.e. ‘permutated’ statistical maps) was also used to evaluate the significance of the overlap further. We permutated the spatial distribution of the fMRI statistical maps used in the activation–gene expression association analysis for 100 times. Voxel-wise statistical values that were stored at the real maps (i.e. emotion regulation or memory control map) were relocated to other locations, testing for spatial specificity of the original task-related spatial patterns, but containing all voxel-wise values. It is notable that this relocation procedure was restricted by the functional-defined brain parcellation (Schaefer et al., 2018). In this way, intrinsic connectivity associations that might underlie functional networks and brain parcels are retained. These ‘permutated’ statistical maps were used for the activation–gene expression association analysis to generate 100 pairs of gene lists. Before the overlapping calculation, we also calculated overall spatial similarity with each pair. Seventy-two out of 100 pairs of these gene lists had a similarity ranging from 0.05 to 0.07 (mean = 0.06) with a standard deviation ranging from 0.25 to 0.29 (s.d. = 0.27). These results were used as the null distribution to estimate the P-value of the overlap. To identify the ‘inhibition-related’ genes, we first used the same activation–gene expression association analysis to identify the stop-signal and go/no-go-associated gene list and then defined the overlap between four gene lists (emotion regulation, think/no-think, stop-signal and go/no-go) as ‘inhibitory-related’ genes.
We aim to improve the specificity of our activation–gene expression association analysis and safeguard the possibility that these identified genes may only support the general functional brain network architecture instead of particular cognitive functions. To rule out this possibility, we performed a dedicated control analysis: (i) An unthresholded DMN ALE map was used as a comparison with four inhibitory control tasks because control processes hardly take place in the DMN and the DMN has been associated with homeostasis and undirected thought or mind-wandering. (ii) A pair of ‘permutated’ statistical maps with comparable spatial similarity level (mean ranging from 0.05 to 0.07, standard deviation ranging from 0.25 to 0.29) was also used to calculate the transcriptional overlap with four inhibitory control tasks.
We used a range of different numbers of genes (x from 1 to 2000, with step size 10) as thresholds to identify the top x most similar genes. We calculated three kinds of overlap under different thresholds: the first one is ‘within control’ overlap, which is the overlap in gene expression association between two inhibitory tasks (e.g. think/no-think and emotion regulation or stop-signal and go/no-go); the second one is ‘inhibitory and DMN’ overlap, which is the transcriptional overlap between one of the control-related tasks and DMN (e.g. think/no-think and DMN or emotion regulation and DMN). The third one is the ‘inhibition and permutation’ overlap, which is the overlap of associated gene lists between one of the actual statistical maps (i.e. emotion regulation or memory control) and one of the permutated statistical maps. Finally, we averaged the percentage of each kind of overlap for a certain threshold x and compared the average percentages across all thresholds.
Gene ontology enrichment analysis
The gene ontology (GO) is a widely used bioinformatics tool to interpret the complex gene list based on the knowledge regarding functions of genes and gene products (Ashburner et al., 2000; Huang et al., 2009). To systematically investigate the biological meaning of the ‘inhibitory- related’ genes, we use GO to perform a binary version of the overrepresentation test for biological processes (BP), molecular function (MF) and cellular component (CC). We did not use additional parameters to restrict the selection of GO categories. Currently, experimental findings from over 140 000 published papers are represented as over 600 000 experimentally supported GO annotations in the GO knowledgebase. For the input gene list, ‘gene ontology enrichment analysis’ can identify relevant groups of genes that function together and associate and statistically test the relationship between these groups and GO annotations. In this way, it can reduce a big list of genes to a much smaller number of biological functions (BP, MF or CC) and make the input list more easily comprehendible. This method has been applied successfully to understand the output of many gene expression studies, including the study that also used the Allen Human Brain Atlas (Richiardi et al., 2015). More specifically, we used GOATOOLS (Ashburner et al., 2000) to perform the GO analyses based on ontologies and associations downloaded on 15 November 2018. All of the significant GO items were corrected by FDR correction (P < 0.05). We further identified the frequently seen words within all the significant GO items by counting the frequency of the words after removal of not meaningful words (e.g. ‘of’ and ‘the’).
Disease-related gene set enrichment
Although GO analysis can provide insights into the biological functions (BP, MF or CC) of the overlapped gene list, the approach does not provide sufficient information to identify disease associations of the gene list. We leveraged ToppGene (Chen et al., 2009) to explore the gene-disease associations. The ToppGene platform can cluster groups of genes together according to their disease associations and perform a statistical test as well as the multiple testing error corrections. The latest version of DisGeNET included associations between 17 549 genes and 24 166 diseases, disorders or abnormal human phenotypes. In this study, our analysis was based on one of the sub-databases of DisGeNET, the DisGeNET Curated (expert-curated associations obtained from UniProt and CTD human datasets). We did not use additional parameters to restrict the selection of disease items and performed a binary version of the overrepresentation analysis. All of the significant disease items were FDR-corrected (P < 0.05).
Data and code availability
All the data (excluding neuroimaging data) is stored in the Open Science Framework (OSF). Open access data includes study summary, extracted coordinates for ALE, significantly associated genes for each task paradigm and overlapped gene list(s) (OSF link: https://doi.org/10.17605/OSF.IO/6WZ2J). Neurovault was used to store all the neuroimaging data (e.g. results of ALE from Figure 2, MACM from Figure 3 and ‘Top Genes’ expression maps from Figure 4) and provide 3D visualization of all the statistical maps (Neurovault link: https://neurovault.org/collections/4845/). Functional-defined brain parcellation can be found in https://github.com/ThomasYeoLab/CBIG/tree/master/stable_projects/brain_parcellation/Schaefer2018_LocalGlobal. Other research data supporting reported findings are available from the authors upon request.
Fig. 2.
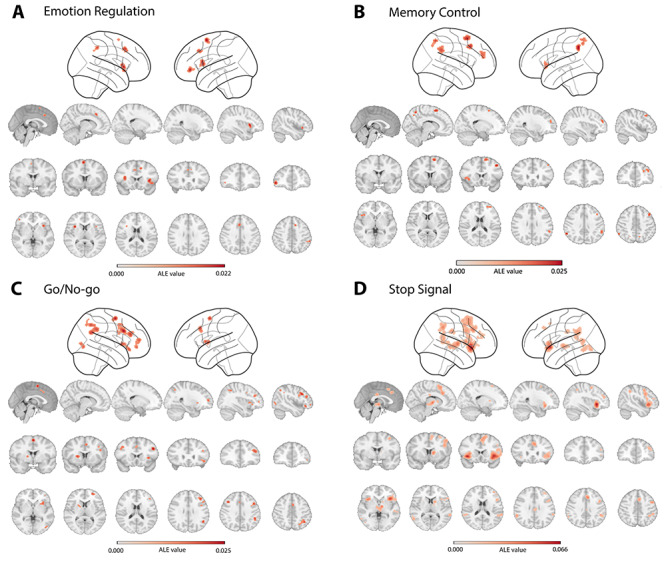
Brain activity underlying four inhibitory control tasks revealed by ALE meta-analyses. (A) Brain regions significantly more activated in the ‘regulation’ compared to the ‘baseline’ condition during the emotion regulation task. (B) Brain regions significantly more activated in the ‘control’ compared to the ‘non-control’ condition during the memory control task. (C) Brain regions significantly more activated in the ‘no-go’ compared to the ‘go’ condition during the go/no-go task. (D) Brain regions significantly more activated in the ‘stop’ compared to the ‘go’ condition during the stop-signal task.
Fig. 3.
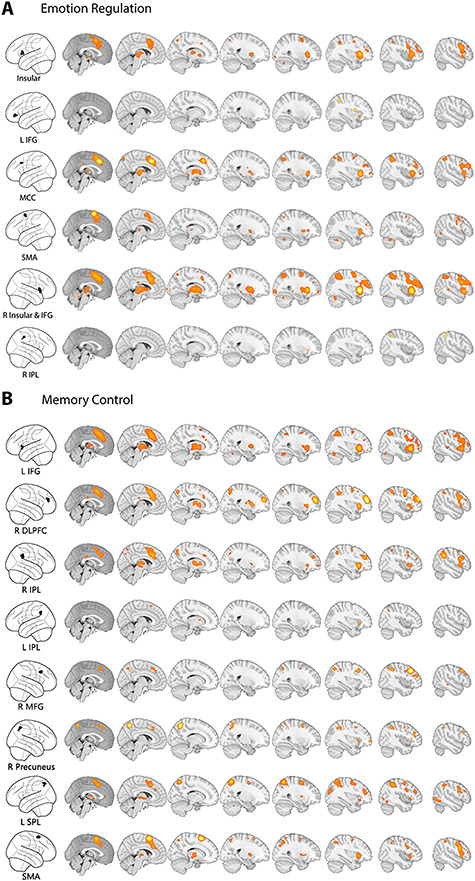
Co-activation maps of emotion regulation and MACM. (A) Co-activation maps for regions of interest (ROIs) resulting from ALE analysis of the emotion regulation. (B) Co-activation maps for ROIs resulting from ALE analysis of the memory control. ALE-based ROIs are projected onto glass brains (left column), and co-activation patterns are rendered on an MNI template second to ninth column (right columns).
Fig. 4.
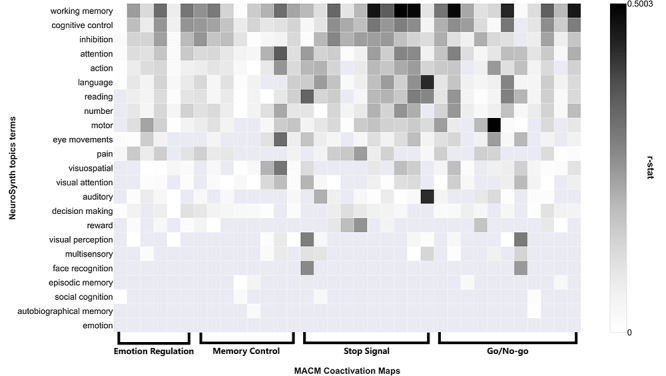
Behavioral profiles of co-activation maps across four inhibitory control tasks. Spatial similarity between Neurosynth meta-maps and co-activation maps across 23 topic terms. Terms are ordered by the mean r-values across the row(s) (all co-activation maps). ‘Working memory’, ‘cognitive control’ and ‘inhibition’ are located at the top, suggesting the common stronger association. Domain-specific cognitive functions (e.g. autobiographical memory, emotion) are located at the bottom, suggesting a limited association.
For the neuroimaging meta-analysis, two software (GingerALE and Sleuth) (http://www.brainmap.org/software.html) were used. The Neurosynth Tool (https://github.com/neurosynth/neurosynth), a Python package that covered most of the functions on the Neurosynth (http://www.neurosynth.org/) website, was used in functional profile analyses and co-activation analyses. Activation–gene expression association analyses were based on alleninfo tool (https://github.com/chrisfilo/alleninf) and web application of gene decoder within the Neurovault (https://neurovault.org/). Nilearn (https://nilearn.github.io/) was used to load, manipulate and visualize MRI statistical maps.
GOATOOLS (https://github.com/tanghaibao/goatools), a Python library, was used for gene ontology analyses and related visualization. The ontology was downloaded from the gene ontology website (http://geneontology.org/ontology/), and the association data were downloaded from the National Center for Biotechnology Information (ftp://ftp.ncbi.nlm.nih.gov/gene/DATA/). ToppGene Suite (https://toppgene.cchmc.org/) was used for disease-related gene set enrichment. The gene-disease association database can be downloaded from http://www.disgenet.org/.
Anaconda (https://www.anaconda.com/) Python 3.6 version for Win10 was used as the platform for all the programming and statistical analyses. Custom Python scripts were written to perform all analyses described based on the mentioned Python packages and are released via the OSF.
Results
Regional brain activity associated with emotion regulation and memory control
Taking data from 95 published studies including in total 1995 subjects, we used 15 emotion regulation studies to represent emotion regulation and 15 think/no-think studies to represent memory control. It is noteworthy that we explicitly only focus on emotion regulation studies using the ‘suppression’ or ‘distraction’ as the regulation techniques rather than the more common ‘cognitive reappraisal’ because of the conceptual relation with memory control and equal statistical power between contrasted conditions. Furthermore, 27 go/no-go studies and 38 stop-signal studies were used to represent response inhibition. (A list of studies and coordinates are available via the Open Science Framework; Search and inclusion criteria in Materials and Methods.)
Regional brain activity of emotion regulation
The meta-analysis of the emotion regulation studies revealed six brain regions that are active during ‘regulation’ compared to a ‘passive viewing’ condition (FWE cluster-level correction P < 0.05, uncorrected P < 0.001, threshold permutations = 1000). Emotion regulation task consistently led to activation in right insula/inferior frontal gyrus (IFG), left IFG, left insula, middle cingulate gyrus, right inferior parietal lobule (IPL) and left supplementary motor area (SMA) (Table 1, Figure 2A).
Table 1.
Significant activated clusters during emotion regulation and think/no-think task
| Task | Brain region | Hemisphere | MNI coordinates | Cluster size | Peak voxel value |
|---|---|---|---|---|---|
| Emotion regulation | Insula/IFG | R | 36 16 6 | 209 | 0.02056 |
| IFG | L | −46 44 −4 | 138 | 0.02051 | |
| Insula | L | −34 16 12 | 156 | 0.02089 | |
| Middle cingulate gyrus | L/R | 10 22 44 | 215 | 0.0189 | |
| Inferior parietal lobule | R | 60–42 46 | 90 | 0.01688 | |
| SMA | L | −4 6 62 | 110 | 0.0217 | |
| Memory control | IFG/insula | L | −36 20 –4 | 115 | 0.017 |
| DLPFC | R | 36 48 24 | 221 | 0.018 | |
| Supramarginal gyrus/IPL | R | 56 −44 32 | 167 | 0.02 | |
| Supramarginal gyrus/IPL | L | −58 −52 38 | 125 | 0.025 | |
| Middle frontal gyrus | R | 42 20 42 | 126 | 0.022 | |
| Precuneus | R | 8 –56 54 | 140 | 0.021 | |
| Precuneus/SPL | L | −16 −64 56 | 114 | 0.017 | |
| SMA | R | 12 12 60 | 190 | 0.023 |
SPL, superior parietal lobule.
Regional brain activity of memory control
Memory control studies revealed five brain regions during the ‘no-think’ condition compared to the ‘think’ condition (FWE cluster-level correction P < 0.05, uncorrected P < 0.001, threshold permutations = 1000): left insula/IFG, right dorsolateral prefrontal cortex (DLPFC), right middle frontal gyrus, bilateral IPL/supramarginal gyrus, bilateral precuneus and SMA (Table 1, Figure 2B).
Regional brain activity of response inhibition
We used the same meta-analytical approach for all go/no-go and stop-signal studies and found similar significant clusters during the ‘control’ condition compared to ‘baseline’ condition (no-go vs go; stop vs go) in the insula, IFG, middle cingulate gyrus, SMA, IPL and DLPFC (Figure 2C and D; Supplementary Tables S1 and S2).
Analyses of convergence and divergence
To examine the spatial convergence and divergence between emotion regulation and memory control activations, we performed formal conjunction and contrast analysis (FWE cluster-level correction P < 0.05, threshold permutations = 1000). The conjunction and contrast analysis did not find any reliable clusters after correction. Informal overlap analysis of two thresholded maps (i.e. emotion regulation and memory control) revealed a shared cluster of right IPL (MNI:60/−42/42; BA40).
Co-activation maps of regions associated with emotion regulation and memory control
To gain deeper insight into the co-activation profiles of brain regions associated with emotion regulation and memory control, we used a large database of fMRI studies (BrainMap: http://www.brainmap.org/) and applied MACM. This approach reveals brain regions that are consistently activated together with the seed regions resulting from the ALE meta-analyses. To control for specific modeling methods and sizes of seed regions, we also performed similar co-activation analyses using Neurosynth (http://neurosynth.org/) based on peak voxel coordinates of each seed region instead of the entire clusters. The two approaches yielded similar co-activation maps for each ROI. In the main test, we only report results from the BrainMap analysis (Results from the Neurosynth can be found in Supplementary Figures S1 and S2).
In total, we analyzed six ROIs from the meta-analysis of emotion regulation studies. MACM analyses for the left IFG and right IPL revealed co-activation patterns in the direct vicinity of the seed regions. Co-activation analyses of other seed regions, including the bilateral insula, SMA, MACC revealed co-activation between these ROIs and with the cerebellum, IFG, IPL, thalamus and DLPFC (Figure 3A). Similarly, we analyzed eight seed regions from the meta-analysis of memory control studies. ROIs tended to be co-activated with each other. Only the MACM analysis for the left IPL ROI identified modest co-activation patterns (Figure 3B). In addition, we investigated the co-activation profiles of response inhibition tasks. The co-activation patterns of 11 ROIs from go/no-go meta-analysis and 10 ROIs from stop-signal meta-analysis were estimated using the same method. We found a brain network including bilateral DLPFC, insular, IPL, thalamus, SMA and MACC across the co-activation patterns of ROIs from response inhibition.
Behavioral profiles of the co-activation maps
To identify the cognition domains most strongly associated with these co-activation maps, we created the behavioral profiles of each co-activation maps using the Neurosynth database.
First, we quantified the behavioral profiles of co-activation maps of emotion regulation and memory control. As expected, these two sets of co-activation maps were both strongly characterized by terms such as ‘inhibition’, ‘cognitive control’ and ‘working memory’. Then, the behavioral profiles of stop-signal and go/no-go co-activation maps were estimated in the same way. Finally, we investigated the commonalities of behavioral characteristics of all co-activation maps across four paradigms and revealed that these co-activation maps have the highest correlations to terms including ‘inhibition’, ‘cognitive control’ and ‘working memory’, compared to other items (Figure 4).
Transcriptional signatures underlying the emotion regulation and memory control
Next, to understand the transcriptional correlates that may be associated with brain activity elicited by each task, we combined the AHBA, a brain-wide atlas of gene expression of postmortem brains and spatial pattern correlation. More specifically, we aimed to identify the genes for which the spatial transcriptional patterns are similar to the spatial pattern of brain activity during a given task.
Common transcriptional correlates of brain activity
We used the activation–gene expression association analysis to identify two lists of genes whose spatial patterns correlate with emotion regulation-related brain activity patterns or memory control-related brain activity patterns separately. The top 10 genes with the highest spatial similarities are represented in Figure 5A, and their expression patterns in the brain are depicted in Figure 5B (Complete gene lists in the OSF repository). We found a substantial overlap (Figure 5A) between the two identified gene lists for the emotion regulation and memory control activation networks: there are in total 1212 genes (60.6% of all correlated genes) whose expression pattern correlated with the brain activity pattern of both tasks. We evaluated the significance of the overlap by generating a set of null distributions of the overlapping genes under the restriction of spatial similarity. Specifically, two gene lists with identical sizes (list1 = 2531; list2 = 1529) were randomly selected from all the genes (N = 20 787), and then overlapping genes between the two random lists were identified. This procedure was repeated 5000 times. 1718 out of 5000 pairs of randomly sampled genes demonstrated a similar level of spatial similarity as our gene lists of interests. We found that the amount of overlapping genes between the memory control and the emotion regulation was significantly larger than the number of overlapping genes within these 1718 randomly sampled overlapping gene lists across different thresholds (threshold = 2000, real overlapping = 1212, random overlapping = 193.28 ± 12.3, P < 0.001) (results at different thresholds are presented in Supplementary Table S4). Furthermore, the significance of the overlap was estimated by generating 100 pairs of ‘permutated’ statistical maps. 72 out of 100 pairs of genes that were associated with these ‘permutated’ maps were used for the estimation because they showed a comparable level of spatial similarity across genes. The amount of real overlapping genes was significantly higher than the number of overlapping genes within these 72 pairs of genes across different thresholds (threshold = 2000, real overlapping = 1212, permutated overlapping = 667.3 ± 147.39, P < 0.01) (Full results in Supplementary Table S5).
Fig. 5.
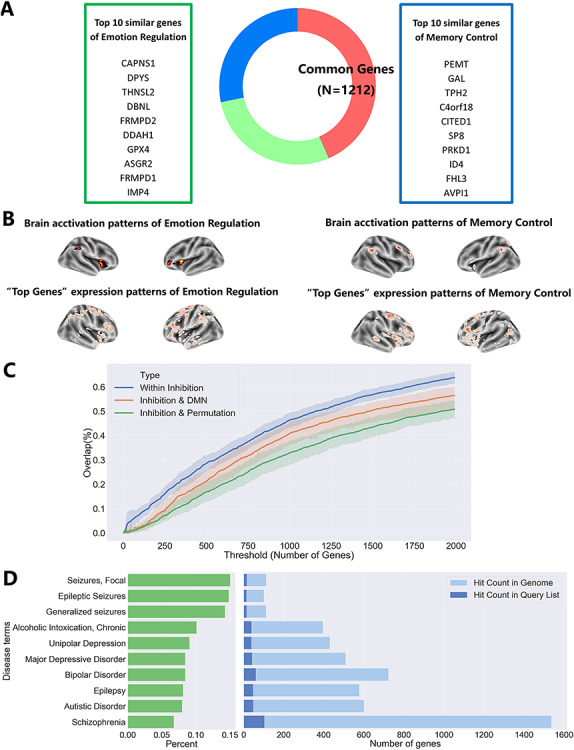
Common transcriptional profiles revealed by activation–gene expression association analysis. (A) Transcriptional patterns of 1212 genes correlated with both the emotion regulation- and memory control-related brain activity patterns. (B) Visualization and comparison of brain activation patterns and expression patterns of ‘Top Genes’; ‘Top Genes’ are 10 genes with the most similar expression patterns as the brain activation. (C) The gene lists of two inhibitory control tasks were more overlap with each other (within inhibition) compared to one of the inhibitory control and DMN (inhibitory and DMN) or one of the real inhibitory control and one of permutated inhibition inhibitory control (inhibition and permutation). (D) Disease associations of ‘inhibitory-related’ genes. ‘Inhibition-related’ genes are associated with genetic risks for depression, bipolar disorder, autism spectrum disorders, schizophrenia, seizures/epilepsy and chronic alcoholic intoxication. Terms are ordered by the percentage (left green) of hit counts in the query list to hit count in the genome. The hit count in the query list is the number of genes belonging to ‘inhibitory-related’ genes (right dark blue). The hit count in the genome is the total number of genes associated with the risk for certain disease terms based on the DisGeNET (right light blue).
Specificity of ‘inhibition-related’ genes
To test the specificity of the ‘inhibition-related’ genes vs genes related to neuronal activity and brain function in general, we first generated, also using an ALE meta-analysis, the activation map of ‘default mode network’ or ‘task-negative network’ (see Materials and Methods; Supplementary Figure S3). Next, we identified a list of genes associated with the DMN network and ‘permutated’ maps emotion regulation and memory control via the AHBA and the same activation–gene expression association analysis. Given the fact that the DMN has been associated with homeostasis and undirected thought or mind-wandering, we expected that DMN-associated genes are different from inhibition-associated genes. To test this, we calculated the number of overlapping genes between all possible combinations of two out of the seven reported gene lists (memory control, emotion regulation, stop-signal and go/no-go, DMN, ‘permutated’ memory control, ‘permutated’ emotion regulation) across different thresholds. As depicted in Figure 5C, genes associated with inhibitory control tasks demonstrate more overlap between each other than the associated genes between the inhibitory control tasks and the DMN (mean within inhibition =41.24%, mean inhibition and DMN =35.18%, t = 3.44, P = 0.0006). They also showed more overlap compared to associated genes between inhibitory control tasks and ‘permutated’ inhibitory control maps (mean within inhibition =41.24%, mean inhibition and permutation =29.53%, t = 6.97, P = 1.26 × 10−11).
Biological functions and disease associations of the ‘inhibition-related’ genes
Using GO (Ashburner et al., 2000), a widely used literature-based gene-to-function annotation analysis, we generated a list of biological functions related to the ‘inhibition-related’ genes. Furthermore, due to the common finding of impairment in inhibitory control in neurological and psychiatric disorders, we explored the human disease association of the identified gene list.
Enrichment for biological functions
Gene ontology enrichment analysis (GOEA) of 779 ‘inhibition-related’ genes identified 5 related BP, which are associated with information communications between neurons (Supplementary Table S6). More specifically, 19 genes within the list are associated with the neuropeptide signaling pathway, 30 genes with the chemical synaptic transmission, 17 genes with the potassium ion transmembrane transport, 38 with the cell adhesion and 60 with signal transduction.
Enrichment for human diseases
Additionally, we used ToppGene (https://toppgene.cchmc.org/) to perform the gene list enrichment analysis of 708 ‘inhibition-related’ genes for human diseases. Using DisGeNET (www.disgenet.org), a comprehensive database on the relationship between human diseases and genes, we found significant associations between our gene list and 36 disease terms (Supplementary Table S7). Among these diseases, seizures/epilepsy, chronic alcoholic intoxication, depression, bipolar disorder, autism spectrum disorders and schizophrenia were ranked top 10 of the list (i.e. most significant associations) (Figure 5D).
We performed a preliminary investigation of the unique disease associations for inhibition, but not DMN. First, we calculated the differences between ‘inhibition-related’ genes and DMN-related genes under four different thresholds and included the results into the ToppGene. Only under the threshold of 1500 genes, the algorithm identified a significant association between 103 ‘inhibition-unique’ genes and the risk for schizophrenia. In other words, variance in these genes, which are associated with inhibition, but not DMN, are reported to be associated with an increased risk for schizophrenia. We also performed the disease association analysis for ‘inhibition-related’ and DMN-related genes separately. These two sets of genes were associated with similar brain disorders (e.g. schizophrenia, autistic disorder, unipolar depression, alcoholic intoxication) (see OSF folder). One important difference between the two sets of genes is that ‘DMN-related’ genes are uniquely associated with Alzheimer’s disease and age-related memory disorders (Supplementary Figure S4), which is consistent with our knowledge regarding the relationship between DMN, aging and memory.
Discussion
Inhibitory control is a fundamental cognitive function supporting other processes like emotion regulation and response inhibition. Here, we provided neurobiological evidence from human neuroimaging and transcriptional mapping to support the concept that there is one generic neural network of inhibitory control with a set of ‘inhibition-related’ genes modulating not only response inhibition but also emotion regulation and memory control. Our meta-analysis of 95 neuroimaging studies revealed a common role of the right IPL and related regions in a frontal–parietal–insular network during emotion regulation and memory control. These co-activation patterns were also similar to the meta-analysis results of response inhibition tasks and ‘inhibition-related’ network reported in the literature. Additionally, we used the Allen Human Brain Atlas as an avenue to link this neural network to common transcriptional profiles and identified ‘inhibition-related’ genes, which are associated with the neuronal transmission, and risk for major psychiatric disorders as well as seizures and alcoholic dependency.
The idea that inhibitory control is the underlying core cognitive function in emotion regulation was already suggested before (Ochsner et al., 2002; Ochsner and Gross, 2005; Schmeichel et al., 2008; Wager et al., 2008; McRae et al., 2012). Similarly, it was also suggested that inhibitory control plays a fundamental role in memory control (Levy and Anderson, 2002). A unified theory proposed a central role of inhibitory control in various psychological domains (e.g. motor inhibition, emotional response and memory retrieval) depending on external task requirements (Aron et al., 2004, 2014; Depue et al., 2016). Engen and Anderson (2018) recently reviewed behavioral and neuroimaging studies in this field and proposed the conceptual link between emotion regulation and memory control. However, there has been little empirical support for this link. Our multimodal analysis provides rich evidence beyond neuroimaging supporting a conceptual link and suggests that inhibitory control, as well as its underlying neural and transcriptional correlates, modulates both emotion regulation and memory control.
Brain activation patterns of emotion regulation and memory control found here are in accordance with previous meta-analyses (emotion regulation: e.g. Buhle et al., 2014; Kohn et al., 2014; memory control: e.g. Guo et al., 2018). We found one overlapping region between emotion regulation and memory control, the right IPL. Due to the central role of inhibitory control in both tasks, only one overlapping brain region seems surprising at first glance. However, MACM revealed that other regions (including IFG, insula, preSMA/MACC, IPL) that lacked significantly overlapping activations across emotion regulation and memory control form a tightly integrated network. Our results suggest that although these regions do not overlap strictly, they belong to the same functional network. Our behavioral profile analysis corroborated this interpretation: both co-activation maps of emotion regulation and memory control, as well as stop-signal and go/no-go paradigms, have comparable behavioral profiles and were characterized by terms like ‘inhibition,’ ‘cognitive control’ and ‘working memory’.
We have demonstrated the neural commonality of emotion regulation and memory control. Next, we proceeded to investigate if transcriptional profiles overlap with activity patterns in a similar way. Critically, this study adopted an imaging genetic approach to investigate the common transcriptional signatures across neural networks of emotion regulation and memory control. Activation–gene expression association analysis revealed a largely overlapping gene list whose expression patterns were similar to the activation patterns. Furthermore, we identified a list of ‘inhibition-related’ genes and characterized their biological function and disease associations. ‘Inhibition-related’ genes were primarily associated with the neuropeptide signaling pathway, chemical synaptic transmission, the potassium ion transmembrane transport and cell adhesion. One common feature of these biological functions is that they are critical for information communications between neurons. Together with our neuroimaging finding of frontal–parietal–insular network, these genes may act as molecular correlates underlying synchronous brain activity during inhibitory control. ‘Inhibition-related’ genes were also associated with risks for several psychiatric disorders (e.g. depression, bipolar disorder, schizophrenia and autism), seizures and alcoholic dependence.
Recently, neuroimaging (Goodkind et al., 2015; Sha et al., 2019) and genetic studies (Cross-Disorder Group of the Psychiatric Genomics Consortium., 2013; Anttila et al., 2018; Schork et al., 2019) collectively demonstrated the potential common biological roots across psychiatric disorders. However, common phenotypes across disorders are less well understood. Thus, the National Institute of Mental Health’s Research Domain Criteria (NIMH’s RDoC) (https://www.nimh.nih.gov/research/research-funded-by-nimh/rdoc/index.shtml) summarized several domains of phenotypes, where inhibitory control is a central aspect. Here, our results highlighted the critical role of inhibitory control and its biological underpinning across psychiatric disorders. Firstly, dysfunctional inhibitory control (e.g. impaired response inhibition, lack of emotion regulation and compromised memory control) is evident across different psychiatric disorders (Magee and Zinbarg, 2007; Price and Mohlman, 2007; Tull et al., 2007; Amstadter, 2008; Falconer et al., 2008; Ehring and Quack, 2010; Erk et al., 2010; Joormann and Gotlib, 2010; Lipszyc and Schachar, 2010; Catarino et al., 2015; Yang et al., 2016; Sacchet et al., 2017). Also, inhibitory control deficits can be further linked to some disorder-specific symptoms [e.g. lack of inhibition of negative thoughts (or rumination) in depression, lack of fear control in anxiety and failure to avoid retrieval of traumatic memories in PTSD]. Secondly, our results suggest an overlap between brain regions (frontal–parietal–insular network) involved in inhibitory control and regions whose structural abnormalities were observed consistently in a variety of psychiatric diagnoses (Goodkind et al., 2015). Thirdly, ‘inhibition-related’ genes, which we identified by spatial transcriptional profiles that overlap with activation patterns of inhibitory control tasks, were also associated with the risks for a variety of psychiatric disorders. Furthermore, although brain disorders such as epilepsy and alcoholic dependence may involve different neurobiological underlying correlates compared to psychiatric disorders, impairment of inhibitory control seems to also be a critical behavioral aspect of epilepsy (Helmstaedter, 2001; Elger et al., 2004) and alcoholic dependence (Lawrence et al., 2009; Courtney et al., 2013; Papachristou et al., 2013).
Our study has two limitations that should be mentioned. First, our neural commonality analyses were based on fMRI studies only. However, overlap in fMRI activation or co-activation patterns lacks temporal information of the underlying cognitive processes. Electroencephalography or magnetoencephalography in humans could provide further confirmatory evidence for the idea of a common cognitive process. Recently, Castiglione and colleagues reported that memory control elicited an electrophysiological signature, increased right frontal beta, which was seen in the stop-signal task (Castiglione et al., 2019). Follow-up electrophysiological studies or even a meta-analysis of them might confirm the idea of a common electrophysiological signature. Second, the current activation–gene expression association analysis is still preliminary (e.g. low sample size and spatial resolution of the postmortem data) and without the possibility of testing the specific relationship between expression maps and cognitive function. For example, although our preliminary results demonstrated that ‘inhibition-specific’ genes are associated with the risk for schizophrenia, while DMN-related genes are more closely linked to Alzheimer’s disease, we also identified considerable transcriptional overlaps between ‘inhibition-related’ genes and DMN-related genes. The latter may suggested that ‘inhibition-related’ genes, as defined in our study may include both ‘inhibition-specific’ genes and other genes supporting general brain function. However, it is challenging to separate them using methods currently available. Nevertheless, the method already showed great potential when helping to understand basic molecular principles of both the structural and functional connectome (see review by Fornito et al., 2019), and it identified molecular mechanisms underlying changes in brain structure or function associated with brain disorders [e.g. autism spectrum disorder (Romero-Garcia et al., 2019), Huntington’s disease (McColgan et al., 2018), schizophrenia (Romme et al., 2017) and Alzheimer’s disease (Grothe et al., 2018)]. Results from these studies are consistent with the genetics of neuropsychiatric disorders using conventional methods like genome-wide association studies or animal models. Taken together, although preliminary, neuroimaging-gene expression association analysis has demonstrated its potential to bridge brain structure–function associations and to reveal its underlying molecular processes. To detect more specific associations between spatial transcriptional profiles and neuroimaging data, large sets of postmortem gene expression data with higher spatial resolution need to be collected. Also, more dedicated analytical methods need to be developed and validated (Arnatkevic̆iūtė et al., 2019) with new methods that may better control for the effects of domain–general genes that are supporting brain function in general and the bias in gene set enrichment analyses of brain-wide gene expression data, probably induced by the gene–gene co-expression or autocorrelation (Fulcher et al., 2020).
In summary, our multimodal analysis identified a frontal–parietal–insular neural network and a set of genes associated with inhibitory control across emotion regulation, memory control and response inhibition. The integrative approach established here bridges between cognitive, neural and molecular correlates of inhibitory control and can be used to study other higher-level cognitive processes. Our findings may deepen our understanding of emotion regulation and memory control in health and pave the way for better emotion regulation and memory control by targeting the core inhibitory-related network or related molecular targets in patients with such deficit at issue.
Supplementary Material
Acknowledgements
We thank Prof. Hans van Bokhoven for comments on the early results of this study, Dr Yuhua Guo for providing brain activation coordinates of unpublished studies, Dr Dace Apšvalka for providing valuable feedback on our preprint and Dr Xiangzhen Kong for suggestions on our revision.
Funding
W.L. is funded by the Ph.D. scholarship (201606990020) of the Chinese Scholarship Council.
Conflict of interest
All authors declare that they have no competing interests.
Author contributions
The experiment was designed by W.L., G.F. and N.K. Data were collected by W.L., N.P. and N.K. Analyses were performed by W.L. and W.L. N.P. wrote the manuscript. W.L, G.F. and N.K. revised and edited the manuscript. All authors provided essential feedback on the manuscript at different stages, contributed substantially to, and approved the final version of the manuscript.
References
- Amstadter A. (2008). Emotion regulation and anxiety disorders. Journal of Anxiety Disorders, 22, 211–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson M.C. (2004). Neural systems underlying the suppression of unwanted memories. Science, 303, 232–5. [DOI] [PubMed] [Google Scholar]
- Anderson M.C., Green C. (2001). Suppressing unwanted memories by executive control. Nature, 410, 366–9. [DOI] [PubMed] [Google Scholar]
- Anderson M.C., Hanslmayr S. (2014). Neural mechanisms of motivated forgetting. Trends in Cognitive Sciences, 18, 279–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anttila V., Bulik-Sullivan B., Finucane H.K., et al. (2018). Analysis of shared heritability in common disorders of the brain. Science, 360, eaap8757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnatkevic̆iūtė A., Fulcher B.D., Fornito A. (2019). A practical guide to linking brain-wide gene expression and neuroimaging data. NeuroImage, 189, 353–67. [DOI] [PubMed] [Google Scholar]
- Aron A.R., Robbins T.W., Poldrack R.A. (2004). Inhibition and the right inferior frontal cortex. Trends in Cognitive Sciences, 8, 170–7. [DOI] [PubMed] [Google Scholar]
- Aron A.R., Robbins T.W., Poldrack R.A. (2014). Inhibition and the right inferior frontal cortex: one decade on. Trends in Cognitive Sciences, 18, 177–85. [DOI] [PubMed] [Google Scholar]
- Ashburner M., Ball C.A., Blake J.A., Botstein D., Butler H., Cherry J.M. (2000). Gene ontology: tool for the unification of biology. Nature Genetics, 25, 25–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berto S., Wang G.-Z., Germi J., Lega B.C., Konopka G. (2018). Human genomic signatures of brain oscillations during memory encoding. Cerebral Cortex, 28, 1733–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buhle J.T., Silvers J.A., Wager T.D., et al. (2014). Cognitive reappraisal of emotion: a meta-analysis of human neuroimaging studies. Cerebral Cortex, 24, 2981–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castiglione A., Wagner J., Anderson M., Aron A. (2019). Preventing a thought from coming to mind elicits increased right frontal beta just as stopping action does. Cerebral Cortex, 29, 2160–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Catarino A., Küpper C.S., Werner-Seidler A., Dalgleish T., Anderson M.C. (2015). Failing to forget: inhibitory-control deficits compromise memory suppression in posttraumatic stress disorder. Psychological Science, 26, 604–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen J., Bardes E.E., Aronow B.J., Jegga A.G. (2009). ToppGene suite for gene list enrichment analysis and candidate gene prioritization. Nucleic Acids Research, 37, W305–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Courtney K.E., Ghahremani D.G., Ray L.A. (2013). Fronto-striatal functional connectivity during response inhibition in alcohol dependence. Addiction Biology, 18, 593–604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cross-Disorder Group of the Psychiatric Genomics Consortium (2013). Identification of risk loci with shared effects on five major psychiatric disorders: a genome-wide analysis. The Lancet, 381, 1371–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Depue B.E., Orr J.M., Smolker H.R., Naaz F., Banich M.T. (2016). The Organization of Right Prefrontal Networks reveals common mechanisms of inhibitory regulation across cognitive, emotional, and motor processes. Cerebral Cortex, 26, 1634–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dosenbach N.U.F., Fair D.A., Miezin F.M., et al. (2007). Distinct brain networks for adaptive and stable task control in humans. Proceedings of the National Academy of Sciences, 104, 11073–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duncan J. (2010). The multiple-demand (MD) system of the primate brain: mental programs for intelligent behaviour. Trends in Cognitive Sciences, 14, 172–9. [DOI] [PubMed] [Google Scholar]
- Ehring T., Quack D. (2010). Emotion regulation difficulties in trauma survivors: the role of trauma type and PTSD symptom severity. Behavior Therapy, 41, 587–98. [DOI] [PubMed] [Google Scholar]
- Eickhoff S.B., Laird A.R., Grefkes C., Wang L.E., Zilles K., Fox P.T. (2009). Coordinate-based activation likelihood estimation meta-analysis of neuroimaging data: a random-effects approach based on empirical estimates of spatial uncertainty. Human Brain Mapping, 30, 2907–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eickhoff S.B., Bzdok D., Laird A.R., et al. (2011). Co-activation patterns distinguish cortical modules, their connectivity and functional differentiation. NeuroImage, 57, 938–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elger C.E., Helmstaedter C., Kurthen M. (2004). Chronic epilepsy and cognition. The Lancet Neurology, 3, 663–72. [DOI] [PubMed] [Google Scholar]
- Elliott L.T., Sharp K., Alfaro-Almagro F., et al. (2018). Genome-wide association studies of brain imaging phenotypes in UK Biobank. Nature, 562, 210–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Engen H.G., Anderson M.C. (2018). Memory control: a fundamental mechanism of emotion regulation. Trends in Cognitive Sciences, 22, 982–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erk S., Mikschl A., Stier S., et al. (2010). Acute and sustained effects of cognitive emotion regulation in major depression. Journal of Neuroscience, 30, 15726–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falconer E., Bryant R., Felmingham K.L., et al. (2008). The neural networks of inhibitory control in posttraumatic stress disorder. Journal of Psychiatry & Neuroscience, 33, 413. [PMC free article] [PubMed] [Google Scholar]
- Fedorenko E., Duncan J., Kanwisher N. (2013). Broad domain generality in focal regions of frontal and parietal cortex. Proceedings of the National Academy of Sciences, 110, 16616–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fornito A., Arnatkevičiūtė A., Fulcher B.D. (2019). Bridging the gap between connectome and transcriptome. Trends in Cognitive Sciences, 23, 34–50. [DOI] [PubMed] [Google Scholar]
- Fox P.T., Lancaster J.L. (2002). Mapping context and content: the BrainMap model. Nature Reviews Neuroscience, 3, 319–21. [DOI] [PubMed] [Google Scholar]
- Fulcher B.D., Arnatkeviciute A., Fornito A. (2020). Overcoming bias in gene-set enrichment analyses of brain-wide transcriptomic data. bioRxiv. [Google Scholar]
- Goodkind M., Eickhoff S.B., Oathes D.J., et al. (2015). Identification of a common neurobiological substrate for mental illness. JAMA Psychiatry, 72, 305–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gorgolewski K., Fox A., Chang L., et al. (2014). Tight Fitting Genes: Finding Relations Between Statistical Maps and Gene Expression Patterns, Hamburg: Organization for Human Brain Mapping. [Google Scholar]
- Gorgolewski K.J., Varoquaux G., Rivera G., et al. (2015). NeuroVault.org: a web-based repository for collecting and sharing unthresholded statistical maps of the human brain. Frontiers in Neuroinformatics, 9, 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gross J.J., Thompson R.A. (2007). Emotion Regulation: Conceptual Foundations. In: Gross J. J., editor. Handbook of Emotion Regulation. New York: Guilford Press, pp. 3–24. [Google Scholar]
- Grothe M.J., Sepulcre J., Gonzalez-Escamilla G., et al. (2018). Molecular properties underlying regional vulnerability to Alzheimer’s disease pathology. Brain, 141, 2755–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo Y., Schmitz T.W., Mur M., Ferreira C.S., Anderson M.C. (2018). A supramodal role of the basal ganglia in memory and motor inhibition: meta-analytic evidence. Neuropsychologia, 108, 117–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hariri A.R., Mattay V.S., Tessitore A., et al. (2002). Serotonin transporter genetic variation and the response of the human amygdala. Science, 297, 400–3. [DOI] [PubMed] [Google Scholar]
- Hawrylycz M.J., Lein E.S., Guillozet-Bongaarts A.L., et al. (2012). An anatomically comprehensive atlas of the adult human brain transcriptome. Nature, 489, 391–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helmstaedter C. (2001). Behavioral aspects of frontal lobe epilepsy. Epilepsy & Behavior, 2, 384–95. [DOI] [PubMed] [Google Scholar]
- Huang D.W., Sherman B.T., Lempicki R.A. (2009). Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nature Protocols, 4, 44–57. [DOI] [PubMed] [Google Scholar]
- Joormann J., Gotlib I.H. (2010). Emotion regulation in depression: relation to cognitive inhibition. Cognition and Emotion, 24, 281–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kohn N., Eickhoff S.B., Scheller M., Laird A.R., Fox P.T., Habel U. (2014). Neural network of cognitive emotion regulation—an ALE meta-analysis and MACM analysis. NeuroImage, 87, 345–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kong X., Song Y., Zhen Z., Liu J. (2017). Genetic variation in S100B modulates neural processing of visual scenes in Han Chinese. Cerebral Cortex, 27, 1326–36. [DOI] [PubMed] [Google Scholar]
- Laird, Eickhoff S.B., Kurth F., et al. (2009a). ALE meta-analysis workflows via the BrainMap database: progress towards a probabilistic functional brain atlas. Frontiers in Neuroinformatics, 3, 23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laird A.R., Lancaster J.J., Fox P.T. (2005). Brainmap. Neuroinformatics, 3, 65–77. [DOI] [PubMed] [Google Scholar]
- Laird A.R., Eickhoff S.B., Li K., Robin D.A., Glahn D.C., Fox P.T. (2009b). Investigating the functional heterogeneity of the default mode network using coordinate-based meta-analytic modeling. Journal of Neuroscience, 29, 14496–505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laird A.R., Eickhoff S.B., Fox P.M., et al. (2011). The BrainMap strategy for standardization, sharing, and meta-analysis of neuroimaging data. BMC Research Notes, 4, 349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Langner R., Leiberg S., Hoffstaedter F., Eickhoff S.B. (2018). Towards a human self-regulation system: common and distinct neural signatures of emotional and behavioural control. Neuroscience & Biobehavioral Reviews, 90, 400–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lawrence A.J., Luty J., Bogdan N.A., Sahakian B.J., Clark L. (2009). Impulsivity and response inhibition in alcohol dependence and problem gambling. Psychopharmacology, 207, 163–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levy B.J., Anderson M.C. (2002). Inhibitory processes and the control of memory retrieval. Trends in Cognitive Sciences, 6, 299–305. [DOI] [PubMed] [Google Scholar]
- Lipszyc J., Schachar R. (2010). Inhibitory control and psychopathology: a meta-analysis of studies using the stop signal task. Journal of the International Neuropsychological Society, 16, 1064–76. [DOI] [PubMed] [Google Scholar]
- Magee J.C., Zinbarg R.E. (2007). Suppressing and focusing on a negative memory in social anxiety: effects on unwanted thoughts and mood. Behaviour Research and Therapy, 45, 2836–49. [DOI] [PubMed] [Google Scholar]
- Margulies D.S., Ghosh S.S., Goulas A., et al. (2016). Situating the default-mode network along a principal gradient of macroscale cortical organization. Proceedings of the National Academy of Sciences, 113, 12574–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McColgan P., Gregory S., Seunarine K.K., et al. (2018). Brain regions showing white matter loss in Huntington’s disease are enriched for synaptic and metabolic genes. Biological Psychiatry, 83, 456–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McRae K., Jacobs S.E., Ray R.D., Johnd O.P., Gross J.J. (2012). Individual differences in reappraisal ability: links to reappraisal frequency, well-being, and cognitive control. Journal of Research in Personality, 46, 2–7. [Google Scholar]
- Morawetz C., Bode S., Derntl B., Heekeren H.R. (2017). The effect of strategies, goals and stimulus material on the neural mechanisms of emotion regulation: a meta-analysis of fMRI studies. Neuroscience & Biobehavioral Reviews, 72, 111–28. [DOI] [PubMed] [Google Scholar]
- Ochsner K.N., Gross J.J. (2005). The cognitive control of emotion. Trends in Cognitive Sciences, 9, 242–9. [DOI] [PubMed] [Google Scholar]
- Ochsner K.N., Bunge S.A., Gross J.J., Gabrieli J.D.E. (2002). Rethinking feelings: an FMRI study of the cognitive regulation of emotion. Journal of Cognitive Neuroscience, 14, 1215–29. [DOI] [PubMed] [Google Scholar]
- Papachristou H., Nederkoorn C., Havermans R., Bongers P., Beunen S., Jansen A. (2013). Higher levels of trait impulsiveness and a less effective response inhibition are linked to more intense cue-elicited craving for alcohol in alcohol-dependent patients. Psychopharmacology, 228, 641–9. [DOI] [PubMed] [Google Scholar]
- Power J.D., Cohen A.L., Nelson S.M., et al. (2011). Functional network organization of the human brain. Neuron, 72, 665–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price R.B., Mohlman J. (2007). Inhibitory control and symptom severity in late life generalized anxiety disorder. Behaviour Research and Therapy, 45, 2628–39. [DOI] [PubMed] [Google Scholar]
- Richiardi J., Altmann A., Milazzo A.-C., et al. (2015). Correlated gene expression supports synchronous activity in brain networks. Science, 348, 1241–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Romero-Garcia R., Warrier V., Bullmore E.T., Baron-Cohen S., Bethlehem R.A.I. (2019). Synaptic and transcriptionally downregulated genes are associated with cortical thickness differences in autism. Molecular Psychiatry, 24, 1053–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Romme I.A.C., Reus M.A., Ophoff R.A., Kahn R.S., Heuvel M.P. (2017). Connectome disconnectivity and cortical gene expression in patients with schizophrenia. Biological Psychiatry, 81, 495–502. [DOI] [PubMed] [Google Scholar]
- Sacchet M.D., Levy B.J., Hamilton J.P., et al. (2017). Cognitive and neural consequences of memory suppression in major depressive disorder. Cognitive, Affective, & Behavioral Neuroscience, 17, 77–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaefer A., Kong R., Gordon E.M., et al. (2018). Local-global parcellation of the human cerebral cortex from intrinsic functional connectivity MRI. Cerebral Cortex, 28, 3095–114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmeichel B.J., Volokhov R.N., Demaree H.A. (2008). Working memory capacity and the self-regulation of emotional expression and experience. Journal of Personality and Social Psychology, 95, 1526. [DOI] [PubMed] [Google Scholar]
- Schork A.J., Won H., Appadurai V., et al. (2019). A genome-wide association study of shared risk across psychiatric disorders implicates gene regulation during fetal neurodevelopment. Nature Neuroscience, 22, 353–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seidlitz J., Váša F., Shinn M., et al. (2018). Morphometric similarity networks detect microscale cortical organization and predict inter-individual cognitive variation. Neuron, 97, 231–247.e7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sha Z., Wager T.D., Mechelli A., He Y. (2019). Common dysfunction of large-scale neurocognitive networks across psychiatric disorders. Biological Psychiatry, 85, 379–88. [DOI] [PubMed] [Google Scholar]
- Shine J.M., Breakspear M., Bell P.T., et al. (2019). Human cognition involves the dynamic integration of neural activity and neuromodulatory systems. Nature Neuroscience, 22, 289–96. [DOI] [PubMed] [Google Scholar]
- Tull M.T., Barrett H.M., McMillan E.S., Roemer L. (2007). A preliminary investigation of the relationship between emotion regulation difficulties and posttraumatic stress symptoms. Behavior Therapy, 38, 303–13. [DOI] [PubMed] [Google Scholar]
- Wager T.D., Davidson M.L., Hughes B.L., Lindquist M.A., Ochsner K.N. (2008). Prefrontal-subcortical pathways mediating successful emotion regulation. Neuron, 59, 1037–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang G.-Z., Belgard T.G., Mao D., et al. (2015). Correspondence between resting-state activity and brain gene expression. Neuron, 88, 659–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang W., Chen Q., Liu P., et al. (2016). Abnormal brain activation during directed forgetting of negative memory in depressed patients. Journal of Affective Disorders, 190, 880–8. [DOI] [PubMed] [Google Scholar]
- Yarkoni T., Poldrack R.A., Nichols T.E., Van Essen D.C., Wager T.D. (2011). Large-scale automated synthesis of human functional neuroimaging data. Nature Methods, 8, 665–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All the data (excluding neuroimaging data) is stored in the Open Science Framework (OSF). Open access data includes study summary, extracted coordinates for ALE, significantly associated genes for each task paradigm and overlapped gene list(s) (OSF link: https://doi.org/10.17605/OSF.IO/6WZ2J). Neurovault was used to store all the neuroimaging data (e.g. results of ALE from Figure 2, MACM from Figure 3 and ‘Top Genes’ expression maps from Figure 4) and provide 3D visualization of all the statistical maps (Neurovault link: https://neurovault.org/collections/4845/). Functional-defined brain parcellation can be found in https://github.com/ThomasYeoLab/CBIG/tree/master/stable_projects/brain_parcellation/Schaefer2018_LocalGlobal. Other research data supporting reported findings are available from the authors upon request.
Fig. 2.
Brain activity underlying four inhibitory control tasks revealed by ALE meta-analyses. (A) Brain regions significantly more activated in the ‘regulation’ compared to the ‘baseline’ condition during the emotion regulation task. (B) Brain regions significantly more activated in the ‘control’ compared to the ‘non-control’ condition during the memory control task. (C) Brain regions significantly more activated in the ‘no-go’ compared to the ‘go’ condition during the go/no-go task. (D) Brain regions significantly more activated in the ‘stop’ compared to the ‘go’ condition during the stop-signal task.
Fig. 3.
Co-activation maps of emotion regulation and MACM. (A) Co-activation maps for regions of interest (ROIs) resulting from ALE analysis of the emotion regulation. (B) Co-activation maps for ROIs resulting from ALE analysis of the memory control. ALE-based ROIs are projected onto glass brains (left column), and co-activation patterns are rendered on an MNI template second to ninth column (right columns).
Fig. 4.
Behavioral profiles of co-activation maps across four inhibitory control tasks. Spatial similarity between Neurosynth meta-maps and co-activation maps across 23 topic terms. Terms are ordered by the mean r-values across the row(s) (all co-activation maps). ‘Working memory’, ‘cognitive control’ and ‘inhibition’ are located at the top, suggesting the common stronger association. Domain-specific cognitive functions (e.g. autobiographical memory, emotion) are located at the bottom, suggesting a limited association.
For the neuroimaging meta-analysis, two software (GingerALE and Sleuth) (http://www.brainmap.org/software.html) were used. The Neurosynth Tool (https://github.com/neurosynth/neurosynth), a Python package that covered most of the functions on the Neurosynth (http://www.neurosynth.org/) website, was used in functional profile analyses and co-activation analyses. Activation–gene expression association analyses were based on alleninfo tool (https://github.com/chrisfilo/alleninf) and web application of gene decoder within the Neurovault (https://neurovault.org/). Nilearn (https://nilearn.github.io/) was used to load, manipulate and visualize MRI statistical maps.
GOATOOLS (https://github.com/tanghaibao/goatools), a Python library, was used for gene ontology analyses and related visualization. The ontology was downloaded from the gene ontology website (http://geneontology.org/ontology/), and the association data were downloaded from the National Center for Biotechnology Information (ftp://ftp.ncbi.nlm.nih.gov/gene/DATA/). ToppGene Suite (https://toppgene.cchmc.org/) was used for disease-related gene set enrichment. The gene-disease association database can be downloaded from http://www.disgenet.org/.
Anaconda (https://www.anaconda.com/) Python 3.6 version for Win10 was used as the platform for all the programming and statistical analyses. Custom Python scripts were written to perform all analyses described based on the mentioned Python packages and are released via the OSF.