

Abstract
The question of whether some cases of interstitial cystitis may have an infectious etiology has been debated for some time. Previous studies have looked for the presence of certain specific viruses, but generally did not use the types of sensitive and unbiased approaches that are currently available. As part of the MAPP (Multidisciplinary Approach to the Study of Chronic Pelvic Pain) Research Network, we examined urine specimens from interstitial cystitis patients who provided specimens over time and also reported various symptoms at the time of urine collection. We first performed next generation sequencing to look for the presence of viruses in urines, and detected two human polyomaviruses that are known to be excreted into urine, BKPyV and JCPyV. We were especially interested in BKPyV because it is a known cause of another bladder disease, hemorrhagic cystitis, in bone marrow transplant recipients. Further analysis of individual samples indicate a trend toward higher excretion of polyomaviruses in patients experiencing increased symptoms.
Keywords: polyomavirus, interstitial cystitis, virus discovery, PCR, NGS
Introduction
Interstitial Cystitis/Bladder Pain Syndrome (IC/BPS) is a painful condition associated with frequent and/or urgent urination that affects an estimated 1-3% of the US population [1, 2]. Current treatments have been developed empirically but, in the absence of a clear understanding of what causes the disease, are suboptimal [3]. Some of the histological findings in the bladders of IC patients, such as focal inflammatory infiltrates and the presence of vacuoles in epithelial cells [4], are suggestive of a viral infection. In the present study, we undertook a pilot study of viruses present in the urine of IC patients. After a preliminary global analysis of urine via NGS, we focused our attention on the polyomavirus family for several reasons. First, the only viral genomes detected with confidence in our initial screening belonged to the polyomaviruses JCPyV and BKPyV. In addition, the known biology of polyomaviruses indicates their suitability as candidates to cause or influence IC/BPS : a) the best studied polyomavirus, Simian Virus 40, was originally discovered by its ability to cause vacuolization in cells in culture [5], and it is known that other polyomaviruses cause this same morphological alteration; b) at least two human polyomaviruses, JCPyV and BKPyV, establish lifelong persistent infections in the urinary tract, and are periodically excreted in the absence of overt clinical symptoms [6] and, c) BKPyV in particular can cause hemorrhagic cystitis (HC) in bone marrow and hematopoietic stem cell transplant recipients, demonstrating that it has the ability to cause disease in the urothelium [7]. While a limited number of studies in the past looked for various viruses in IC patients and reported negative results, they used techniques that easily could have missed important pathogens and would not have identified unknown viruses [e.g., 4, 8–10]. There are, however, sporadic case reports of BKPyV in IC specimens [11, 12], and one more recent manuscript claimed to have detected BKPyV in IC urines at a higher frequency than in healthy urines [13].
The Multidisciplinary Approach to the Study of Chronic Pelvic Pain (MAPP) Research Network is an NIDDK-funded study that seeks to better understand the causes of IC [3, 14]. The MAPP network has collected a series of urine samples and symptom severity measurements from participants at six discovery sites in the US. These specimens include longitudinal collections from individual participants whose pain varies over time. With the advent of newer, DNA amplification-based and high-throughput technologies, we wished to undertake a more careful search for BKPyV and JCPyV in these carefully curated participant specimens, to determine if there truly is a link between either virus and IC. In the present study, we examined 50 urine specimens, 5 of which were collected from each of ten participants, for the presence of these two viruses and evaluate the association of virus presence with symptom severity. Our data indicate that the frequency of detection of the two viruses did not differ from reported frequencies in the general population. In participants observed to have both detectable and undetectable levels of virus across their five samples, however, pain and urinary symptom scores were often higher, on average, when viruses were detectable. In addition, we found no evidence for other known viruses by next generation sequencing (NGS).
Materials and Methods
Participant specimens.
Urine samples were collected as part of the MAPP protocol as described [14], A total of 50 deidentified samples taken from ten selected participants were sent to the University of Michigan and the University of Pittsburgh by the MAPP Tissue Analysis and Technology Core at the University of Colorado. The ten (5 female and 5 male) participants were selected based on availability of VB2 urine samples from baseline, 6 month, and 12 month clinic visits, with at least one of those 3 samples corresponding with the timing of the reporting of a flare (self-reported symptom exacerbation), and also a flare and nonflare sample collected via home collection kit, for a total of 5 specimens per participant. Participants were also chosen to have pain and urinary symptom severity scores in the top half of participants for at least one among the baseline, 6 month, and 12 month visits, and exhibit moderate to high variability in symptoms over the 1 year follow up period. All specimen collection was approved by the relevant IRBs at the six discovery sites at which collection occurred. Blinded specimens were sent to the laboratories performing the PCR and NGS analysis.
DNA extraction.
250 ul of each urine sample were aliquoted into a microfuge tube. 35 ul of proteinase K digestion buffer (25 mMTris, pH 8.0; 25 mM EDTA; 10 mM DTT, 1% SDS) and 1 ul proteinase K(Qiagen #19131) was added to each sample and incubated at 50°C for 15 min. The samples were heated to 70°C for 10 min to inactivate the enzyme. The DNA was then cleaned up using the QiaQuick spin PCR clean up kit (Qiagen #28106) and ethanol precipitated.
PCR analysis.
Detection of polyoma large T antigen products was achieved with BKTAg-Forward (5’-AAGGAAAGGCTGGATTCTGA) and BKTAg-Reverse (5’-GTGATTGGGATTCAGTGCT) primers by amplification with the following parameters: 2 min at 50°C; 10 min at 95°C; and 40 cycles of denaturation at 95°C for 15 sec and annealing and extension at 58°C for 30 sec. To amplify specific JCPyV or BKPyV products the default amplification conditions were as follows: 2 min at 94°C followed by 35 cycles of 94°C for 30 sec, appropriate annealing (see below) for 30 sec and 72°C for 30 sec; and a final extension step at 72°C for 2-5 min. Specific PCR reactions were performed using the following sets of primers and annealing conditions: 1) JCV.F1 (5’-CTGATAAAGGTGGGGACGAAGAC) and JCV.R1 (5’-CCATTCATCTGTTCCATAGGTTGG) amplified a 143bp JCPyV LT product after 52°C annealing; 2) JCV.VP1.F3 (5’-TGCTCCTCAATGGATGTTGCC) and JCV.VP1.B4 (5’-GGGGTCCTTCCTTTCTCCTTTTC) amplified a JCPyV VP1 113 nt product after 53°C annealing; 3) BKVT.F (5’ GGGCGACGAGGATAAAATGAAG) and BKVT.R (5’ TTCTGTTCCATAGGTTGGCACC) yielded a 127bp BKPyV LT product after 52°C annealing and, 4) BKVP1 ,R1 (5’-CAGTTACAGCCTCCCACATCAGT AG) and BKVP1.R1 (5’-GCCCAGAGAGAAAAATGCTTCC) generated a 108bp BKPyV VP1 product after 53°C annealing._The reaction products were electrophoresed on 1% agarose gels and the bands visualized by 0.005% gel red stain. Negative control reactions were performed on both water that was taken through the extraction procedure, and water placed directly into the PCR reaction. Positive control reactions used plasmid templates containing complete genomes of JCPyV or BKPyV.
Nucleic acid obtention from urine pools for NGS:
Five different pools were generated by combining 200 ul each of 10 different randomly selected urine samples. Both DNA and RNA were extracted from each pool. For DNA extraction, urine samples were digested with proteinase K and processed with the DNeasy Blood/Tissue kit (Qiagen) according to manufacturer’s instructions. RNA was extracted from each pool using Trizol LS Reagent according to the manufacturer’s instructions. In each case, the material was then amplified by a non-bias random-primer protocol as described by Cantalupo et al. [15], and the resulting samples were submitted to the Genomics Research Core of the University of Pittsburgh (https://www.genetics.pitt.edu/) for library construction and lllumina sequencing. The NGS sequencing data has been deposited to NCBI and is available at: https://www.ncbi.nlm.nih.aov/bioproiect/PRJNA603336.
Viral discovery.
Pickaxe [16] was used to detect viral sequences in each urine pool. Non-host reads were aligned by Pickaxe to the ViralRefSeq database (https://www.ncbi.nlm.nih.gov/genome/viruses/) using Bowtie2. Metrics in Table 1 were calculated from the Bowtie2 alignment output. Sequence coverage (Seqcov) was defined as the percentage of the genome covered by at least one read. AvgDepth is the total aligned bases divided by the length of the virus. AvgMAPQ, avgScore, and avgEditDist is the average MAPQ (Bowtie2 map quality), Score (Bowtie2 AS) and Edit Distance (Bowtie2 NM) of the alignments. Integrated Genome Viewer (http://www.broadinstitute.org/igv/; version 2.3) was used to visualize alignments to JCPyV and BKPyV reference genomes.
TABLE 1.
NGS Analysis of Pooled Urine Specimens
| RNA | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Poola | Accession | Virus | Alignments | Seqcov | avgDepth | avgMAPQ | avgScore | avgEditDist | Length | GI |
| U1 | NC_001699.1 | JC polyomavirus | 13 | 14.4 | 0.2 | 33.9 | −8.2 | 1.7 | 5130 | 9628642 |
| U4 | NC_001699.1 | JC polyomavirus | 106 | 29.8 | 1.5 | 38.3 | −4.8 | 1.0 | 5130 | 9628642 |
| U5 | NC_001699.1 | JC polyomavirus | 98 | 17.9 | 1.4 | 37.8 | −6.7 | 1.4 | 5130 | 9628642 |
| U5 | NC_001501.1 | Moloney murine leukemia virus | 54 | 3.6 | 0.5 | 9.6 | −10.1 | 1.9 | 8332 | 9626958 |
| U5 | NC_001502.1 | Moloney murine sarcoma virus | 16 | 3.9 | 0.2 | 1.0 | −3.4 | 0.7 | 5833 | 9626962 |
| DNA | ||||||||||
| Pool | Accession | Virus | Alignments | Seqcov | avgDepth | avgMAPQ | avgScore | avgEditDist | Length | GI |
| U1 | NC_001699.1 | JC polyomavirus | 743 | 20.0 | 10.8 | 37.0 | −2.4 | 0.5 | 5130 | 9628642 |
| U1 | NC_001802.1 | Human immunodeficiency virus 1 | 404 | 5.1 | 3.2 | 33.4 | −7.8 | 1.6 | 9181 | 9629357 |
| U1 | NC_027532.1 | Pan troglodytes troglodytes polyomavirus 1 isolate F514.1 | 13 | 1.6 | 0.2 | 1.0 | −0.4 | 0.1 | 5159 | 897364561 |
| U3 | NC_001699.1 | JC polyomavirus | 1598 | 76.2 | 23.1 | 36.8 | −4.7 | 1.0 | 5130 | 9628642 |
| U4 | NC_001699.1 | JC polyomavirus | 9781 | 75.7 | 141.9 | 38.4 | −4.9 | 1.0 | 5130 | 9628642 |
| U4 | NC_009225.1 | Torque teno midi virus 1 | 283 | 6.8 | 6.5 | 19.7 | −6.6 | 1.3 | 3245 | 134133206 |
| U4 | NC_007014.1 | Small anellovirus 2 | 221 | 7.4 | 6.3 | 16.1 | −6.7 | 1.4 | 2635 | 66391753 |
| U4 | NC_007013.1 | Small anellovirus 1 | 70 | 8.4 | 2.3 | 3.4 | −18.0 | 3.3 | 2249 | 66391749 |
| U4 | NC_028635.1 | Pan troglodytes verus polyomavirus 8 isolate Ch-Regina | 29 | 1.7 | 0.4 | 1.0 | −2.2 | 0.5 | 5163 | 960494790 |
| U4 | NC_027532.1 | Pan troglodytes troglodytes polyomavirus 1 isolate F514.1 | 25 | 1.7 | 0.4 | 1.0 | −3.3 | 0.7 | 5159 | 897364561 |
| U5 | NC_001538.1 | BK polyomavirus | 8842 | 30.5 | 127.9 | 36.9 | −2.5 | 0.5 | 5153 | 9627180 |
| U5 | NC_001699.1 | JC polyomavirus | 1391 | 6.9 | 20.1 | 33.6 | −10.1 | 2.1 | 5130 | 9628642 |
| U5 | NC_004107.1 | Woodchuck hepatitis virus | 123 | 4.0 | 2.7 | 15.0 | −23.2 | 4.5 | 3323 | 22256030 |
| U5 | NC_001802.1 | Human immunodeficiency virus 1 | 43 | 4.3 | 0.3 | 26.5 | −14.3 | 2.8 | 9181 | 9629357 |
- Pool – the urine sample pool identifier
- Accession - the Genbank accession number of the virus sequence in the Viral RefSeq database
- Virus - the virus name
- Alignments - total number of reads that aligned to the virus genome
- Seqcov - the % of the sequence covered by at least one read
- Avgdepth - the average depth of sequencing coverage across the genome
- avgMAPQ – the average MAPQ (map quality) of the read alignments
- avgScore – the average alignment score of the read alignments
- avgEditDistance – the average edit distance of the read alignments
- Length - length of the virus sequence in base pairs
- GI - the Genbank gi number of the virus sequence in the Viral RefSeq database
Symptom severity measures.
Two primary symptom scores, pain and urinary severity were assessed biweekly in the MAPP Epidemiology and Phenotyping study [17], Pain severity (0-28) is the sum of the pain subscore of the Genitourinary Pain index and item 4 of the Interstitial Cystitis Symptom Index. Urinary severity (0-25) is the sum of the GUPI Urinary subscore and items 1,2,3 and 5 of the ICSI [17], The presence or absence of a flare, defined as a perceived exacerbation of urological pain or urinary symtoms, was determined by item 11 of the Symptoms and Health Care Utilization Questionnaire (SYMQ-11). The present study included participant-reported symptom measures concurrent with provided urine specimens (baseline, 6months, 12 months, and at the time of home collection of flare and nonflare urine samples).
Statistical analysis.
The presence or absence of JCPyV and BKPyV for each sample was documented, and contingency tables were generated to compare the prevalence of JCPyV and BKPyV by flare status, both combined, and separately for in-clinic collected and home-collected specimens. Mean (sd) symptom severity scores were calculated by virus presence or absence within and across participants. Due to the small sample and limited within-patient variability of JCV status, we present descriptive analyses only.
Results
We first attempted to detect overall viral presence in the urine of participants by NGS. For DNA-Seq of the urine pools, we obtained a total of 436M pairs of 75bp reads. Reads were analyzed by Pickaxe for the detection of virus sequences. Deep sequencing of the DNA from 5 pools of urine samples (U1-U5, materials and methods) revealed the presence of JCPyV and BKPyV in urine pools. JCPyV DNA was detected in all urine pools except U2, and BKPyV DNA was only detected in one pool (U5). The number of alignments to JCPyV ranged from 743 to 9,781 and the sequence coverage ranged from 6.9% to 76.2% (Table 1). The alignments were visualized with IGV (Fig. 1). Coverage of JCPyV was quite even along the viral genome, particularly in the U3 and U4 pools (Fig. 1A). In contrast, BKPyV 8,842 alignments in pool U5 mapped to the early region of the virus (Fig. 1B). We also detected what appeared to be Pan troglodytes (chimpanzee) polyomavirus [18] in several samples. However, those reads align with 97 to 100% identity to JCPyV isolate GCN8 by BLASTN; therefore we conclude that Pan troglodytes PyV was not present in the urine samples. Finally, we detected several other human RNA viruses such as HIV1 in U1 and U5, and Anelloviruses in U4 (Table 1). However, gene expression from these viruses was not observed in the RNA-Seq experiment (see below).
Figure 1. JCV and BKV detection in urine pools (U1 – U5).
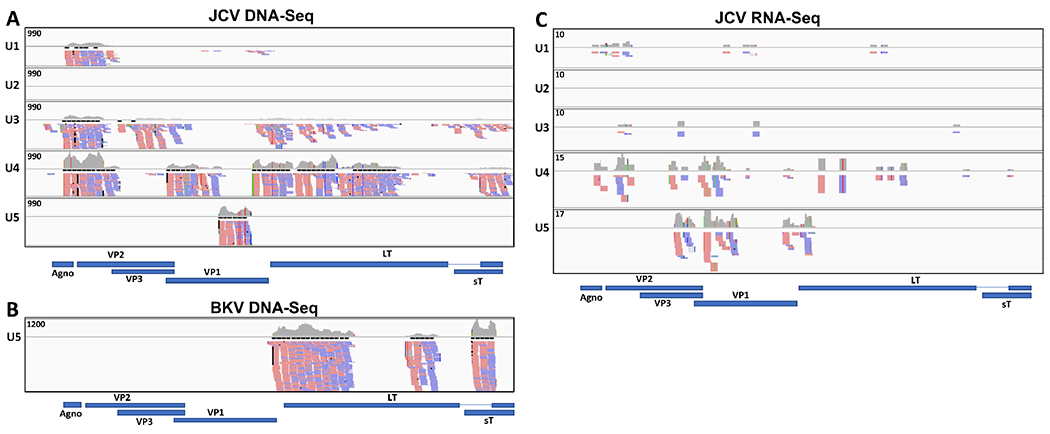
Coverage graphs across the whole genomes of JCV (NC_001699.1) and BKV (NC_001538.1) in the urine pools for; A) DNA-Seq JCPyV; B) DNA-seq BKPyV and C) RNA-Seq JCPyV. The top track in each urine pool (U1 – U5) displays the number of reads aligning to each base in the genome. Maximum Y axis values are indicated. Read alignments for forward (in red) and reverse orientation (in blue) are shown below the graph (not all alignments are shown). There were no alignments to JCV in pool U2 for neither DNA-Seq nor RNA-Seq, and alignments to BKV only in DNA-seq for urine pool U5. The gene annotations for each virus are shown below the coverage graphs.
For RNA-Seq of the urine pools, we obtained a total of 384M pairs of 75bp reads. Reads were analyzed by Pickaxe for the detection of virus sequences. The majority of deep sequencing reads of the RNA from 5 pools of urine samples (materials and methods) corresponded to JCPyV, but we found no evidence of BKPyV transcripts. JCPyV was detected in 4 out of 5 pools, and the reads mainly aligned to the late region of the virus, which encodes for the capsid components (Fig. 1C). The sequence coverage ranged from 14.4% to 29.8% (Table 1). We also detected two murine retroviruses in U5. The reads aligned to genomic region 2303 to 2524 on MMSV and 3370 to 3596 on MMLV; both regions overlap the pol or gag-pol gene of the virus. However, a BLAST search using several of these reads showed that they were 97 to 100% identical to the pAMS plasmid (AF010170.1). These results lead us to believe that a mouse retrovirus is not present in U5, but rather that the corresponding reads are the result of vector contamination during sequencing. Such contamination of deep sequencing data has been previously reported [19].
To confirm the presence of human polyomaviruses in specific urine samples, DNA was extracted and assayed for the presence of JCPyV and BKPyV genomes using PCR. Because of their DNA genomes, polyomaviruses are genetically stable except for one region of the genome called the non-coding control region (NCCR), which contains the two transcriptional promoters for expression of viral genes and the viral origin of DNA replication [20]. We therefore used primers that recognize the coding sequence for large T antigen, a highly conserved viral protein [21,22]. Furthermore, to ascertain detection of specific polyomaviruses, we designed additional primers amplifying large T antigen or VP1 sequences from either BKPyV or JCPyV (materials and methods). The BKPyV PCR was conducted in both the Imperiale and Pipas laboratories, while the JCPyV PCR was performed only in the latter location. Both labs were blinded to the identity of the specimens. After all the data were collected, the data were unmasked and the results were analyzed (Table 2).
Table 2.
PCR Analysis of Urine Specimens for JVPyV and BKPyV
| Barcode* | PID | Site id | sex | COLLECTION TYPE | FLARE | JCPyV | BKPyV |
|---|---|---|---|---|---|---|---|
| URI0029388 | 100964 | 401: U of Michigan | 1: Male | Male:IE:Urine | − | + | |
| URI0032512 | 100964 | 401: U of Michigan | 1: Male | Male:IE:Urine | − | − | |
| URI0033104 | 100964 | 401: U of Michigan | 1: Male | HomeCollection2:Urine | NonFlare | − | − |
| URI0033962 | 100964 | 401: U of Michigan | 1: Male | HomeCollection2:Urine | Flare | − | − |
| URI0072863 | 100964 | 401: U of Michigan | 1: Male | Male:IE:Urine | − | − | |
| URI0028966 | 100967 | 301: U of Iowa | 2: Female | HomeCollection2:Urine | NonFlare | + | − |
| URI0030202 | 100967 | 301: U of Iowa | 2: Female | Female:IE:Urine | + | − | |
| URI0038818 | 100967 | 301: U of Iowa | 2: Female | HomeCollection2:Urine | Flare | + | + |
| URI0043270 | 100967 | 301: U of Iowa | 2: Female | Female:IE:Urine | + | − | |
| URI0083129 | 100967 | 301: U of Iowa | 2: Female | Female:IE:Urine | + | − | |
| URI0033278 | 101059 | 501: U of Washington | 1: Male | Male:IE:Urine | − | − | |
| URI0036122 | 101059 | 501: U of Washington | 1: Male | HomeCollection2:Urine | NonFlare | − | − |
| URI0038980 | 101059 | 501: U of Washington | 1: Male | HomeCollection2:Urine | Flare | − | − |
| URI0069900 | 101059 | 501: U of Washington | 1: Male | Male:IE:Urine | − | − | |
| URI0078424 | 101059 | 501: U of Washington | 1: Male | Male:IE:Urine | − | − | |
| URI0036598 | 101146 | 301: U of Iowa | 2: Female | HomeCollection2:Urine | NonFlare | − | − |
| URI0037837 | 101146 | 301: U of Iowa | 2: Female | HomeCollection2:Urine | Flare | − | − |
| URI0040172 | 101146 | 301: U of Iowa | 2: Female | Female:IE:Urine | − | − | |
| URI0075747 | 101146 | 301: U of Iowa | 2: Female | Female:IE:Urine | − | − | |
| URI0081171 | 101146 | 301: U of Iowa | 2: Female | Female:IE:Urine | − | − | |
| URI0038951 | 101303 | 601: WashU St. Louis | 1: Male | HomeCollection2:Urine | NonFlare | − | − |
| URI0040319 | 101303 | 601: WashU St. Louis | 1: Male | HomeCollection2:Urine | Flare | − | − |
| URI0066936 | 101303 | 601: WashU St. Louis | 1: Male | Male:IE:Urine | − | − | |
| URI0076065 | 101303 | 601: WashU St. Louis | 1: Male | Male:IE:Urine | − | + | |
| URI0088334 | 101303 | 601: WashU St. Louis | 1: Male | Male:IE:Urine | − | − | |
| URI0048464 | 101490 | 301: U of Iowa | 1: Male | HomeCollection2:Urine | NonFlare | − | + |
| URI0048505 | 101490 | 301: U of Iowa | 1: Male | HomeCollection2:Urine | Flare | − | − |
| URI0056485 | 101490 | 301: U of Iowa | 1: Male | Male:IE:Urine | − | − | |
| URI0078528 | 101490 | 301: U of Iowa | 1: Male | Male:IE:Urine | − | − | |
| URI0094469 | 101490 | 301: U of Iowa | 1: Male | Male:IE:Urine | − | − | |
| URI0062016 | 101759 | 201: UCLA | 2: Female | HomeCollection2:Urine | NonFlare | − | + |
| URI0067024 | 101759 | 201: UCLA | 2: Female | HomeCollection2:Urine | Flare | − | − |
| URI0072698 | 101759 | 201: UCLA | 2: Female | Female:IE:Urine | − | + | |
| URI0092080 | 101759 | 201: UCLA | 2: Female | Female:IE:Urine | − | − | |
| URI0098859 | 101759 | 201: UCLA | 2: Female | Female:IE:Urine | − | − | |
| URI0072540 | 101873 | 401: U of Michigan | 2: Female | HomeCollection2:Urine | NonFlare | − | + |
| URI0072881 | 101873 | 401: U of Michigan | 2: Female | HomeCollection2:Urine | Flare | − | + |
| URI0092029 | 101873 | 401: U of Michigan | 2: Female | Female:IE:Urine | − | + | |
| URI0098883 | 101873 | 401: U of Michigan | 2: Female | Female:IE:Urine | − | + | |
| URI0099114 | 101873 | 401: U of Michigan | 2: Female | Female:IE:Urine | − | − | |
| URI0072536 | 101913 | 401: U of Michigan | 2: Female | HomeCollection2:Urine | NonFlare | + | − |
| URI0078538 | 101913 | 401: U of Michigan | 2: Female | HomeCollection2:Urine | Flare | − | − |
| URI0092008 | 101913 | 401: U of Michigan | 2: Female | Female:IE:Urine | + | − | |
| URI0098775 | 101913 | 401: U of Michigan | 2: Female | Female:IE:Urine | + | − | |
| URI0099117 | 101913 | 401: U of Michigan | 2: Female | Female:IE:Urine | − | − | |
| URI0072548 | 101925 | 101: Northwestern U | 1: Male | HomeCollection2:Urine | Flare | + | − |
| URI0072962 | 101925 | 101: Northwestern U | 1: Male | HomeCollection2:Urine | NonFlare | + | − |
| URI0094240 | 101925 | 101: Northwestern U | 1: Male | Male:IE:Urine | + | − | |
| URI0098346 | 101925 | 101: Northwestern U | 1: Male | Male:IE:Urine | − | − | |
| URI0099163 | 101925 | 101: Northwestern U | 1: Male | Male:IE:Urine | + | + |
Explanation of columns. Barcode = specimen number; PID = patient ID; SitelD = site at which specimen was collected; Sex = sex of patient; Collection type = home if collected by patient at home, otherwise the specimen was collected during a clinic visit; Flare = patient’s description of symptoms at time of home collection; JCV, BKV = results of assay
Overall, VP1 products from BKPyV DNA were detected in 7/10 (70%) participants, albeit not in all urine samples from the same individual (e.g. products were detected in only one out of the five corresponding samples from five participants). VP1 products from JCPyV DNA were detected in 3/10 (30%) participants but the prevalence in samples from the same individual was considerably higher. In particular, one participant (100967) had JCPyV in her urine in all the specimens, one (101925) was positive for JCPyV in 4/5 of the specimens and one (101913) showed JCPyV presence in 3/5 specimens. In contrast, only one participant (101873) showed consistent shedding of BKPyV (4/5 samples). In all, 12/50 and 11/50 urine specimens were positive for JCPyV and BKPyV, respectively.
Across all 50 samples, mean pain severity scores were 19.9 and 14.2 in urine samples with JCPyV present versus absent, respectively. Similarly, the mean urinary severity score was 17.0 when JCPyV was present and 13.2 in the absence of JCPyV. Mean pain severity scores were 16.6 and 15.3 for BKPyV present versus absent; mean urinary scores were 12.82 when BKPyV was present and 14.5 in the absence of BKPyV. In 2 of 10 participants who were both undetectable and detectable for JCPyV across different samples collected at different times, within-participant mean pain and urinary scores were higher when JCPyV was detectable. Of 7 participants who were undetectable and detectable for BKPyV across the 5 samples collected, 6 of 7 had higher pain scores with BKPyV detectable as compared with BKPyV undetectable, and 5 of 7 had higher urinary scores when BKPyV was detectable versus undetectable (Figure 2).
Figure 2.
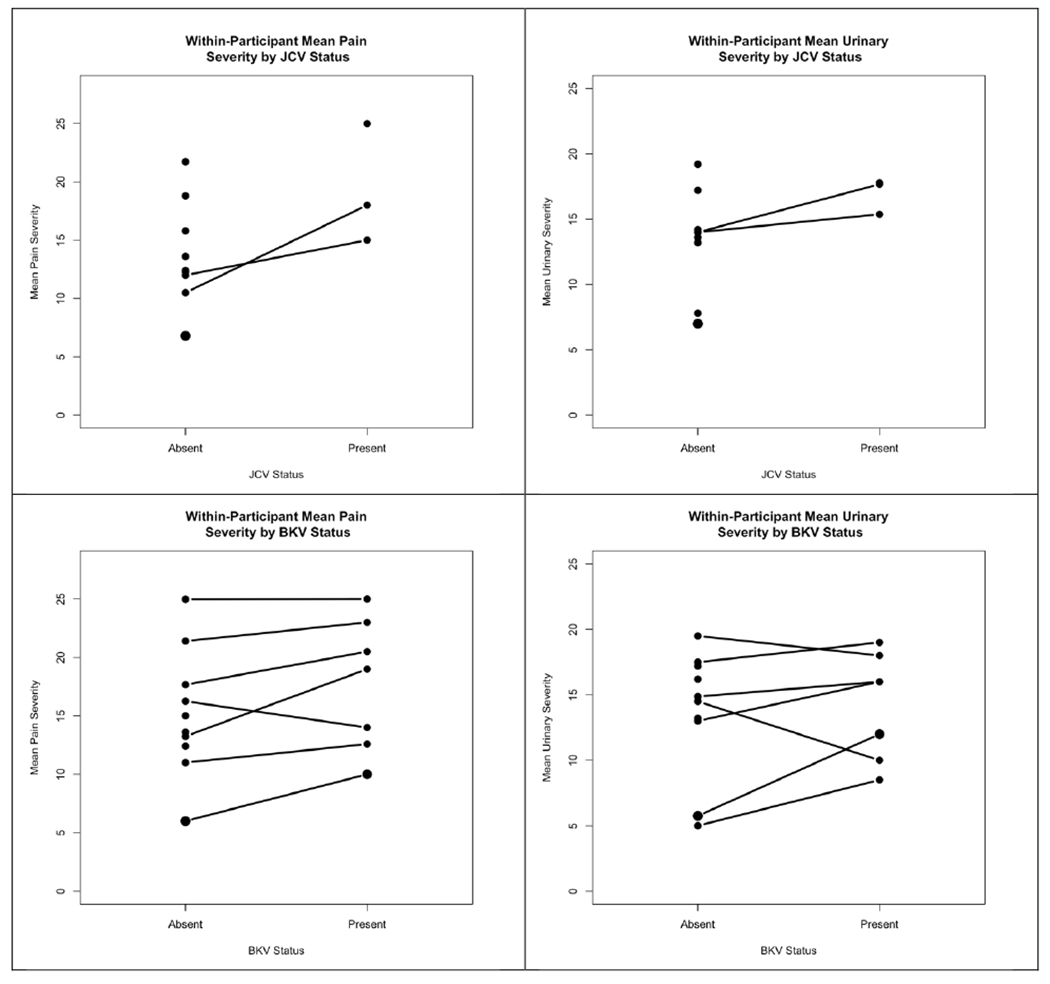
Within participant mean pain and urinary symptom severity scores when JCPyV and BKPyV are present versus absent in the urine. A line connects the mean symptom score for the same participant across urine samples with a virus present or absent. A dot with no line indicates that a participant’s samples were all either positive or all negative for the respective virus.
JCPyV was detected in 5 of 20 (25%) ‘flare’ samples, identified as flare either by home collection kit or SYMQ-11, and 7 of 30 (23.3%) nonflare samples. BKPyV was detected in 6 (30%) of 20 flare samples and 5 of 30 (16.7%) nonflare samples.
Discussion
Histological studies of specimens isolated from IC patients suggested that some IC cases may have a viral etiology. Such evidence includes inflammatory infiltrates and vacuolization of cells [4], Initial attempts to find viral sequences in these specimens were unsuccessful, but more recent studies suggested a possible role for BKPyV in IC [11–13], The advent of improved viral detection techniques, along with the availability of a large number of carefully curated participant specimens from the MAPP network, prompted us to re-evaluate the role of viruses, and polyomaviruses in particular, in IC. In this study, we assayed fifty participant specimens for the presence of viral sequences. All virological assays on specimens were blindly performed to avoid any bias in the analyses.
For our NGS studies, we pooled urine specimens and looked for both DNA and RNA sequences. JCPyV DNA was detected in four out of the five pools. JCPyV coverage ranged from 6.9% to 76.2% and was evenly distributed along both the early and late regions of the viral genome. The same four pools that contained JCPyV DNA also contained JCPyV RNA. Most of the RNA reads aligned to the late region of the viral genome, which encodes the capsid proteins (14.4% to 29.8% genome coverage). The presence of late transcripts in urine indicates that the viral life cycle was being completed in infected cells, strongly suggesting that productively infected cells were present in these samples. In contrast to JCPyV, BKPyV DNA reads were only detected in one of five pools and all mapped to the early viral region (30.5% of genome covered). Furthermore, no BKPyV sequences were found in our RNA-seq analysis. These results are consistent with the presence of BKPyV virions in these samples, most likely shed from infected urinary tract cells.
Our NGS bioinformatics software, Pickaxe [16], has the ability to detect viral sequences in clinical specimens. However, other than BKPyV or JCPyV we did not find any significant evidence of the presence of other viruses in urine from IC participants. Although we detected some DNA reads corresponding to human viruses such as HIV1 and Anelloviruses, the genome coverage was low in all cases (≤ 8.4%), in contrast with coverage observed for BKPyV (30.5%) and JCPyV (6.9% to 76.2%). Furthermore, gene expression from these viruses was not observed in the RNA-Seq experiment.
We followed up the NGS experiments with PCR analysis of viral DNA in the individual specimens, which confirmed and extended the NGS findings. For BKPyV, we found viral DNA in one out of five specimens from five out of the ten participants we analyzed. Detection in these cases did not appear to correlate with symptoms. Only one participant had consistent shedding of BKPyV, which was detected in 4/5 of this participant’s urines. With JCPyV, the prevalence among participants was lower, with DNA detected in only 3/10 participants, but in these participants the virus was detected in 5/5, 4/5, and 3/5 urine specimens. Again, there was no obvious correlation with flares in these participants. As a whole, 12/50 and 11/50 specimens contained JCPyV and BKPyV DNA, respectively. We cannot distinguish whether this DNA was present in viral particles or was free DNA that may have been released from lysed infected cells. Our ability to detect both BKPyV and JCPyV DNA indicate that both viruses are present in infected urinary tract cells. However, the RNA-seq results support that only JCPyV replication was occurring in these participants.
Another group evaluated the presence of BK and JC polyomaviruses in the blood and urine of 400 healthy (mostly Caucasian) donors of different ages and sex [23], Although the seroprevalence was fairly high for BK (82% average) and JC (58% average), the presence of viral DNA in urine was much lower (7% BK, 19% JC). In comparison, our study found a little higher polyomavirus presence in urine of IC participants: a) We analyzed one home collection flare sample from each IC participant, of which 30% showed polyoma presence (1 BK, 1 JC, one both); b) 6 out of the 10 non-flare home-collection samples available (one per participant) contained polyoma sequences (60%, 3 BK, 3 JC).
We cannot explain what appears to be a difference between our results and those of a pilot study by Van der Aa et al [13], who used real time PCR and found BKPyV sequences in 11/15 (73.3%) of urines taken at flare from IC patients, while none of the control patients with benign urological disorders appeared to contain the virus. In our hands, the frequency of BKPyV detection is lower and more similar to previously reported frequencies in the general population (e.g., [24]). One possible explanation for this discrepancy resides in the primers used to detect polyomaviruses in both studies. While we used primers specific for BKPyV and JCPyV, those used by Van der Aa et al are not indicated in the manuscript. In addition, it is unclear whether the assay they used is specific for BKPyV detection because they state that it “gives quantitative results and is considered negative when no polyomavirus [our emphasis] DNA is detected.” Thus, the authors might have detected a wider range of polyomaviruses, although this is inconsistent with our NGS analyses. Our analysis indicates a possible role of BKPyV in symptom fluctuations in IC, with worse symptoms observed when BKPyV is detectable in urine. A larger study is needed, however to confirm our findings. Limited within-patient variability for JCPyV limits our conclusions regarding its potential role in the treated natural history of IC. As a whole, these results suggest that additional studies with greater numbers of specimens are warranted to determine if there is truly an association between either virus and some or all of the parameters of IC.
Acknowledgements
This work was performed as part of an ancillary award to MJI and JMP from the Multidisciplinary Approach to the Study of Chronic Pelvic Pain research network (MAPP), funded by the NIDDK (DK82370, DK82342, DK82315, DK82344, DK82325, DK82345, DK82333, and DK82316).
Footnotes
Publisher's Disclaimer: This Author Accepted Manuscript is a PDF file of an unedited peer-reviewed manuscript that has been accepted for publication but has not been copyedited or corrected. The official version of record that is published in the journal is kept up to date and so may therefore differ from this version.
Conflict of Interest
None of the authors has a conflict to disclose.
Research Involving Human Participants/Informed Consent
All subjects in this study were enrolled at one of six MAPP participating centers, each of whose IRB’s approved the protocol. The subjects were consented at the time of enrollment at each center.
References Cited
- 1.Berry SH, Elliott MN, Suttorp M, Bogart LM, Stoto MA, Eggers P, et al. Prevalence of symptoms of bladder pain syndrome/interstitial cystitis among adult females in the United States. J Urol. 2011;186(2):540–4. doi: 10.1016/j.juro.2011.03.132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Link CL, Pulliam SJ, Hanno PM, Hall SA, Eggers PW, Kusek JW, et al. Prevalence and psychosocial correlates of symptoms suggestive of painful bladder syndrome: results from the Boston area community health survey. J Urol. 2008;180(2):599–606. doi: 10.1016/j.juro.2008.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Clemens JQ, Mullins C, Kusek JW, Kirkali Z, Mayer EA, Rodriguez LV, et al. The MAPP research network: a novel study of urologic chronic pelvic pain syndromes. BMC Urol. 2014;14:57 Epub 2014/08/03. doi: 10.1186/1471-2490-14-57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Fall M, Johansson SL, Vahlne A. A clinicopathological and virological study of interstitial cystitis. J Urol. 1985;133(5):771–3. [DOI] [PubMed] [Google Scholar]
- 5.Sweet BH, Hilleman MR. The vacuolating virus, S.V. 40. Proc Soc Exp Biol Med. 1960;105:420–7. [DOI] [PubMed] [Google Scholar]
- 6.DeCaprio JA, Imperiale MJ, Major EO. Polyomaviruses In: Knipe DM, Howley PM, Cohen JI, Griffin DE, Lamb RA, Martin MA, et al. , editors. Fields Virology. 6th ed. Philadelphia: Lippincott Williams & Wilkins; 2013. [Google Scholar]
- 7.Dropulic LK, Jones RJ. Polyomavirus BK infection in blood and marrow transplant recipients. Bone marrow transplantation. 2008;41 (1):11–8. doi: 10.1038/sj.bmt.1705886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Al-Hadithi HN, Williams H, Hart CA, Frazer M, Adams EJ, Richmond DH, et al. Absence of bacterial and viral DNA in bladder biopsies from patients with interstitial cystitis/chronic pelvic pain syndrome. J Urol. 2005;174(1):151–4. doi: 10.1097/01.ju.0000161605.14804.a9. [DOI] [PubMed] [Google Scholar]
- 9.Hanash KA, Pool TL. Interstitial and hemorrhagic cystitis: viral, bacterial and fungal studies. J Urol. 1970;104(5):705–6. [DOI] [PubMed] [Google Scholar]
- 10.Hukkanen V, Haarala M, Nurmi M, Klemi P, Kiilholma P. Viruses and interstitial cystitis: adenovirus genomes cannot be demonstrated in urinary bladder biopsies. Urol Res. 1996;24(4):235–8. [DOI] [PubMed] [Google Scholar]
- 11.Fioriti D, Penta M, Mischitelli M, Degener AM, Pierangeli A, Gentile V, et al. Interstitial cystitis and infectious agents. Int J Immunopathol Pharmacol. 2005;18(4):799–804. [DOI] [PubMed] [Google Scholar]
- 12.Eisen DP, Fraser IR, Sung LM, Finlay M, Bowden S, O’Connell H. Decreased viral load and symptoms of polyomavirus-associated chronic interstitial cystitis after intravesical cidofovir treatment. Clin Infect Dis. 2009;48(9):e86–8. doi: 10.1086/597827. [DOI] [PubMed] [Google Scholar]
- 13.Van der Aa F, Beckley I, de Ridder D. Polyomavirus BK--a potential new therapeutic target for painful bladder syndrome/interstitial cystitis? Medical hypotheses. 2014;83(3):317–20. doi: 10.1016/j.mehy.2014.06.004. [DOI] [PubMed] [Google Scholar]
- 14.Landis JR, Williams DA, Lucia MS, Clauw DJ, Naliboff BD, Robinson NA, et al. The MAPP research network: design, patient characterization and operations. BMC Urol. 2014;14:58 Epub 2014/08/03. doi: 10.1186/1471-2490-14-58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cantalupo PG, Calgua B, Zhao G, Hundesa A, Wier AD, Katz JP, et al. Raw sewage harbors diverse viral populations. MBio. 2011;2(5). Epub 2011/10/06. doi: 10.1128/mBio.00180-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cantalupo PG, Katz JP, Pipas JM. Viral sequences in human cancer. Virology. 2018;513:208–16. Epub 2017/11/07. doi: 10.1016/j.virol.2017.10.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Griffith JW, Stephens-Shields AJ, Hou X, Naliboff BD, Pontari M, Edwards TC, et al. Pain and Urinary Symptoms Should Not be Combined into a Single Score: Psychometric Findings from the MAPP Research Network. J Urol. 2016;195(4 Pt 1):949–54. Epub 2015/11/21. doi: 10.1016/j.juro.2015.11.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Johne R, Enderlein D, Nieper H, Muller H. Novel polyomavirus detected in the feces of a chimpanzee by nested broad-spectrum PCR. J Virol. 2005;79(6):3883–7. Epub 2005/02/26. doi: 10.1128/JVI.79.6.3883-3887.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cantalupo PG, Pipas JM. Detecting viral sequences in NGS data. Current opinion in virology. 2019;39:41–8. Epub 2019/08/30. doi: 10.1016/j.coviro.2019.07.010. [DOI] [PubMed] [Google Scholar]
- 20.Imperiale MJ. The human polyomaviruses, BKV and JCV: molecular pathogenesis of acute disease and potential role in cancer. Virology. 2000;267(1):1–7. Epub 2000/01/29. doi: 10.1006/viro.1999.0092. [DOI] [PubMed] [Google Scholar]
- 21.Broekema NM, Imperiale MJ. miRNA regulation of BK polyomavirus replication during early infection. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(20):8200–5. doi: 10.1073/pnas.1301907110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Jiang M, Zhao L, Gamez M, Imperiale MJ. Roles of ATM and ATR-mediated DNA damage responses during lytic BK polyomavirus infection. PLoS Pathog. 2012;8(8):e1002898 Epub 2012/09/07. doi: 10.1371/journal.ppat.1002898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Egli A, Infanti L, Dumoulin A, Buser A, Samaridis J, Stebler C, et al. Prevalence of polyomavirus BK and JC infection and replication in 400 healthy blood donors. The Journal of infectious diseases. 2009;199(6):837–46. [DOI] [PubMed] [Google Scholar]
- 24.Kling CL, Wright AT, Katz SE, McClure GB, Gardner JS, Williams JT, et al. Dynamics of urinary polyomavirus shedding in healthy adult women. Journal of medical virology. 2012;84(9):1459–63. doi: 10.1002/jmv.23319. [DOI] [PMC free article] [PubMed] [Google Scholar]