

Abstract
Nutraceuticals had always been known for their therapeutic effects in ancient medicine and had been the primary healing remedy until the introduction of modern chemistry and pharmacology. However, their use has not been dismissed but actually is acquiring a new acclamation among the scientific community especially for their efficacy on the Central Nervous System (CNS). Molecular mechanisms of the most common neurodegenerative diseases are now being uncovered and along with that the molecules that drive the neurodegenerative processes. It is not surprising that some natural compounds can interact with those molecules and interfere with the pathological pathways halting the cascades that ultimately lead to neuronal cell death. The plant Ruta graveolens has gained increased attention in medicinal chemistry due to its beneficial role to treat a variety of human diseases and also because of the presence of a huge number of compounds belonging to different classes of natural products, including neuroactive compounds potentially able to promote neuroprotection. Among all the components of the plant extract, rutin – which is highly, if not the most, abundant – positively interacts with the neurophysiology of the CNS too, being particularly efficient against neurotoxicity. Rutin, has proven to be protective in a variety of experimental settings of neurodegeneration. Finally, it has been shown that the water extract of Ruta graveolens (RGWE) induces death of glioblastoma cells but not of neuronal cells. Moreover, it also fosters cell cycle re-entry and differentiation of neuronal cells. This peculiarity represents a promising tool to promote neural plasticity in pathological conditions.
Keywords: Ruta graveolens, Natural compounds, Neural plasticity, Neurodegeneration, Glioblastoma
Graphical abstract
Highlights
-
•
Traditional therapeutic use of the plant Ruta graveolens in a large variety of illnesses.
-
•
It has been used, together with its main component rutin to treat some neurodegenerative diseases.
-
•
It might own compounds able to foster plasticity in neural cells.
-
•
It is able to kill neural cancer cells but not neurons.
List of abbreviations
- CNS
Central Nervous System
- AD
Alzheimer’s Disease
- PD
Parkinson’s Disease
- HD
Huntington’s Disease
- RGWE
Ruta graveolens water extract
- MAO-B
monoamine oxidase B
- 6-OHDA
6-hydroxydopamine
- RO
reactive oxygen species
- PC12
pheochromocytoma clone12 cells
- AChE
acetylcholinesterase
- TREM
triggering receptor expressed on myeloid cells
- OXPHOS
oxidative phosphorylation
- COX
cyclooxygenase
- Htt
Huntingtin
- IGF1
Insulin-like growth factor 1
- NPCs
neural progenitor cells
- ERK 1/2
extracellular signal-regulated kinase 1 and 2
- HUVEC
Human Umbilical Vein Endothelial Cells
- Akt/PKB
Akt protein kinase B
- mTOR
mammalian target of rapamycin
- A1
A1 mes c-myc neural cells
- cAMP
cyclic adenosine monophosphate
1. Introduction
By definition nutraceuticals are bioactive natural substances with pharmaceutical properties that can positively interfere with disease-related processes.1 What confers to these natural products their therapeutic effects are the secondary metabolites which can promote or restore normal physiological conditions.2 Some of the phytochemicals are effective on the CNS, owning neurobiological activity that has been shown to re-establish normal functions in several neuropathological driven conditions.3 In the last decade a lot of effort has been spent by several research groups in understanding the molecular mechanisms behind the most common neurodegenerative disorders – as Alzheimer’s Disease (AD), Parkinson’s Disease (PD) and Huntington’s Disease (HD) – and in finding the molecules that might halt the disease progression. Although most of the causes are basically still obscure, some of the pathogenetic mechanisms that drive the neurodegeneration have been uncovered and along with those the altered physiological molecular pathways that ultimately lead to neuronal cell death.4 Considering the physiology of the CNS, it is generally known that once neurons are mature and fully differentiated they lose their ability to proliferate except in the two well-defined neurogenic areas.5 Thus, once they die as the brain ages or because of diseases, basically they cannot be replaced. Thus, it would be extremely useful in pathological conditions having compounds that can interfere with those processes. Ruta graveolens is endowed with potent anti-inflammatory effects due to the presence of bioactive compounds like flavonoids.6 Since inflammation is a hallmark of neurodegenerative diseases, Ruta graveolens has been considered as a potentially effective remedy in this field. In addition, analyses have further shown that Ruta graveolens contains a number of molecules known to exert broad effects on neural cell.7 Therefore, envisaging a role in promoting or restoring neural functions, which highlights the importance of investigating it as a potential therapeutic tool in neurodegenerative diseases.
2. Materials and methods
Searches were carried out in Pub Med to collect articles related to the subject. The main terms searched alone or with Boolean operators were the following: “ruta graveolens”; "ruta graveolens" and neur∗; "ruta graveolens" and neuron; "ruta graveolens" and Alzheimer; "ruta graveolens" and inflammation; "ruta graveolens" and traditional medicine; "ruta graveolens" and inflammation and neuron; rutin and neurodegeneration; rutin and Parkinson; rutin and Alzheimer; rutin and huntington; rutin and inflammation and neuron; and other related terms. Articles published from 1999 to 2020 were included in the review.
3. Results
3.1. Ruta graveolens: ethnobotany and traditional medicine
Ruta graveolens L., also known as rue, common rue or sadab, is a shrubby perennial plant that belongs to the family of rutacee. To the same family belong very common edible fruits such as the ones of the genus citrus including lemon, orange and lime. Ruta graveolens is a very resistant plant growing in almost any conditions. This plant is native of mediterranean area although it is now cultivated all over the world including Europe and also in many African, Asian and South American countries such as Ethiopia, China and Japan. The name Ruta originally comes from latin that borrowed it from ancient Greek. Ancient Greek also used the word peganon that is still used in modern Greek to mean ruta.8 Concerning the word graveolens, it also comes from the latin, and means strong-smelling due to the strong unpleasant odour emanating from the leaves. The flavour is very bitter, nevertheless it is used in ethnic cuisine such as in Ethiopia where it is also a coffee flavourant and in Italy where it is used to flavor grappa, an Italian type of brandy.9 Since ancient times, Ruta graveolens has been used and it still is in traditional medicine for its healing properties. According to traditional and folk medicine, Ruta graveolens is known to be effective on a long list of diseases when administered as fresh herb, infusion, decoctions, powder or oils. Among the most common illnesses cured by Ruta graveolens, there are rheumatic diseases, aching pain, eye problems, dermatitis and also multiple sclerosis. Finally, abortion and contraception are among the most widely and ancient prescriptions of Ruta graveolens, thus, according to Wood it “is probably the single most important remedy in Latin American folk medicine" to be used for abortion.10,11 Recently, a study carried out in zebrafish, a well-known animal model to study genetics, developmental biology and toxic compounds, has shown that Ruta graveolens administration exerts an effect on reproduction, namely a decrease in eggs production and fertilization likely due to a disruption of gonadal and/or thyroid hormones.12
3.2. Ruta graveolens in neurodegeneration
As mentioned above, Ruta graveolens, owns anti-inflammatory properties but has also been found to exert positive effects on the CNS. The neurobiological activity of Ruta graveolens, and of its important metabolite rutin, is not only confined to one specific action, but has a wide spectrum activity. Above all, some are particularly relevant because can interfere with pathological pathways involved in the onset and/or the development of neurodegenerative mechanisms such those occurring in PD, AD and HD.13, 14, 15, 16
3.2.1. Ruta graveolens in Parkinson’s disease
A study investigating the effects of Ruta graveolens in neurodegenerative diseases uncovered its ability to inhibit monoamine oxidase B (MAO-B).16
Monoamine oxidases are flavin adenine dinucleotide dependent enzymes that catalyze oxidative deamination of amines. In particular, MAO-B metabolizes the neurotransmitter dopamine whose decrease is the major culprit in PD, the latter being caused by the death of dopaminergic neurons in the substantia nigra pars compacta. Thus, MAO-B inhibitors, such as selegiline and reasagiline, play a pivotal role in the treatment of PD, increasing the basal level of both endogenous and exogenous derived dopamine that otherwise would be likely deaminated by MAO-B.17 Additionally, MAO-B does not only affect dopamine levels but also has proven to slow the progression of the disease.18 Indeed, preclinical studies including experiments in animal and cell culture, provide also evidence for a neuroprotective, neurotrophic and anti-apoptotic effect of these drugs. Such further effects might be due also to the decrease of the metabolites produced during amine turnover, such as reactive oxygen species (ROS), which promote neural damage.19 Interestingly, rutin, (3,3′,4′,5,7-pentahydroxyflavone-3-rhamnoglucoside), the main components of Ruta graveolens water extract, also displays MAO-B inhibiting ability, along with antioxidant, and scavenging properties.20,21
It is worth to pinpoint that rutin, discovered by Albert Szent-Györgyi in 1936 and commonly also referred to as vitamin P, is a glycoside of the flavonoid quercitin, particularly efficient against neurotoxicity.22, 23, 24
Rutin has been shown to exert beneficial effects when administered in a number of dopaminergic cellular models of PD. In particular, in human SH-SY5Y neuroblastoma cells it inhibits amylin-induced toxicity but, more importantly, it has proven to counteract the effects of 6-hydroxydopamine (6-OHDA).25 6-OHDA is a toxin widely used to cause selective dopaminergic lesions in animal models of PD as well as in dopaminergic cells. In fact, it is selectively taken up by dopamine and norepinephrine reuptake transporters. Once in the cells it induces oxidative stress and impairs mitochondrial activity, thus causing cell death. In rat PC12 cells, rutin reduced neurotoxicity induced by 6-OHDA. In addition, it up-regulates the expression of both tyrosine hydroxylase, the rate limiting enzyme of dopamine biosynthesis, and antiapoptotic genes.26,27
A study undertaken in 6-OHDA-induced Parkinson’s disease (PD) rat model, showed that oral administration of rutin protected dopaminergic neurons as demonstrated by histopathological and immunohistochemical findings. Also, an amelioration of neurobehavioral activity of animals treated with rutin was observed. The authors suggested that the oral consumption of rutin might provide protection against PD.28
Finally, it is worth noting that in PC12 cells, Ruta graveolens was found to enhance the NGF-mediated neurite outgrowth, the latter being a parameter widely used to evaluate neuronal differentiation and plasticity.29
Because of its neuroprotective ability, as mentioned before, MAO-B inhibitor selegiline, has also been tested in AD showing a mild beneficial effect both in preclinical experimental settings and in clinical trials.30
3.2.2. Ruta graveolens in Alzheimer’s disease
Furthermore, the use of Ruta graveolens as well as of its component rutin have also been suggested as a treatment for AD. Previous studies have reported Ruta graveolens’ successful application in in vivo murine AD models as acetylcholinesterase (AChE) inhibitor.31, 32, 33 Acetylcholinesterase catalyzes the hydrolysis of acetylcholine following its release at the synapse. In AD cholinergic neurons within the basal forebrain are severely lost. These neurons provide innervation of cerebral cortex as well as of other important brain structures, involved in memory, attention, learning and other higher cognitive functions. Thus, the rationale for using AChE inhibitors, such as donepezil, galantamine, and rivastigmine, is based on counteracting the loss of cholinergic neurotransmission. Nevertheless, these drugs are endowed with adverse and side effects such as nausea, vomiting and diarrhea due to the action in the gastrointestinal tract. Also, they are contraindicated in some cases, including peptic ulcer, bleeding of the gastrointestinal tract, which may be common conditions in the elderly.34
However, in AD a number of pathogenetic alterations leading to neuronal cell death have been described including the accumulation and aggregation of β amyloid, oxidative stress and inflammation. Although the relations among these players is far to be clear, nevertheless evidence have emerged that interfering with these processes may ameliorate or delay the onset, the development and the outcome of the disease.35
Besides enhancing acetylcholine levels, the natural compound has shown to ameliorate the survival rate of affected neurons inhibiting the apoptotic cascade 36; 37.
Moreover, rutin has been clearly proven to exert a beneficial effect in a variety of animal models of AD and in cell cultures. In particular, its oral administration reduced Aβ oligomers in brain, decreased inflammation parameters like oxidative stress, gliosis (i.e. microgliosis and astrogliosis) and pro-inflammatory cytokines (i.e. IL-1, IL-6) and finally attenuated memory loss.38
Similar results were observed in the human neuronal cells SH-SY5Y, where rutin was able to inhibit β-amyloid aggregation and also reduced nitric oxide and inflammatory cytokines.39
An extensive investigation has been very recently published on the effects of rutin salts on two different transgenic mouse models of AD. Not only did this study confirm the beneficial effects on inflammation and Aβ amyloid deposition but it clearly showed that learning and memory deficits were ameliorated upon rutin treatment. Interestingly, the authors also found out that the natural compound is able to activate microglial phagocytosis of Aβ amyloid via up-regulation of phagocytosis-related receptors such as TREM2. In addition, the clearance of Aβ amyloid were promoted by the mitochondrial oxidative phosphorylation (OXPHOS) that was selectively enhanced in the microglial cells since neither neurons nor astrocytes showed any alteration of mitochondrial OXPHOS.40 It is worth to note that while other anti-inflammatory drugs, for example those acting on cyclooxygenase (COX) activity, are currently available to curb neuroinflammation and thus have been tested in preclinical and clinical settings (i.e. Celecoxib), nevertheless, none of them has proven to play such a broad effect on microglial functions.35,41
3.2.3. Ruta graveolens in Huntington’s diseases
Rutin proved to be also effective in both rat and Caenorhabditis elegans (C. elegans) models of Huntington’s diseases (HD). HD is an autosomal dominant, genetic, inherited disease that causes uncontrolled, excessive movements (coreic movements) and cognitive and emotional problems. It is caused by mutations in the HTT gene leading, in turn, to a polyglutamine expansion in the huntingtin protein (Htt), which, thus, gains toxic functions. Spiny gabaergic neurons in the striatum are preferentially affected.42
Notably, pretreatment with rutin administered orally was able to counteract the noxious effects of 3-Nitropropionic acid, a succinate dehydrogenase inhibitor that determines, biochemical, histological and behavioral changes in the striatum, thus reproducing the main phenotypic hallmarks of HD.43 Very recently, a more in-depth study was carried out using the nematode C. elegans to investigate the molecular mechanisms underlying the action of rutin. It is worth mentioning that although the C. elegans model of HD is unable to mimic important features of HD, such as the motor manifestations, nevertheless it is a powerful tool to unveil the molecular mechanisms underlying the action of the mutated Htt protein. Chronic administration of rutin was able to delay ageing and increase lifespan. Moreover, it reduced polyQ-mediated neuronal cell death likely via activation of autophagic protein degradation and insulin/IGF1 signaling pathway.44
3.2.4. Ruta graveolens, signaling pathways and neural plasticity
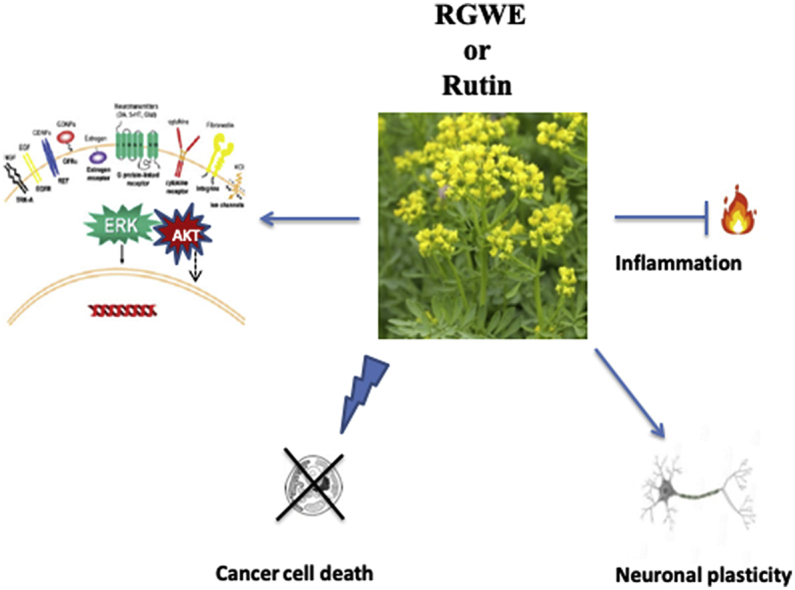
New investigations have shown that other signaling cascades can be influenced by Ruta graveolens. RGWE can affect neural progenitor cells (NPCs) fate through extracellular signal-regulated kinase 1 and 2 (ERK 1/2) and the serine/threonine kinase Akt/Protein kinase B (Akt/PKB) activation, whose signaling cascade are involved in cell proliferation and survival.45,46 Indeed, it is well established that alterations of ERK 1/2 pathway is involved in the pathophysiology of a number of neurodegenerative diseases including PD, AD and HD.47, 48, 49 Surprisingly, ERK also exerts a pivotal role in the action of pro-survival and differentiation factors such as neurotrophins. It has been suggested that the reason underlying such a paradox may be due to a different kinetic of ERK 1/2 activation, which, in turn, depends on the cell type and/or the stimulus administered. In this regard, it is worth noting that RGWE can exert different effects according to the cell type.47 Indeed, in case of neuroepithelial cells RGWE administration increases ERK 1/2 phosphorylation whereas it decreases phosphorylation when given to human endothelial cells.20,46
Moreover, on the whole, these studies reflect the findings observed when using other phenolic compounds, like resveratrol, amentoflavone and curcumin, which exert their actions through the same pathways.50, 51, 52 Indeed, it is well documented that Akt/PKB acts as an upstream signaling molecule of the mTOR pathway, whose activation is involved in various aspects of NPCs and/or neural stem cells development including cell proliferation, differentiation and fate.53, 54, 55
Furthermore, one of the latest studies involving the use of RGWE, has drawn the attention on the capability of the water extract to induce plasticity and promote cell cycle re-entry of fully differentiated A1 mes c-myc neural cells (A1). Notably, upon RGWE stimulation differentiated A1 cells, increase neurites length and express plasticity genes.56 A1 mes c-myc cells is a neural mesencephalic cell line that has been extensively characterized also via advanced proteomic techniques thus allowing to unveil molecules associated to neural cell proliferation, differentiation and maturation.
57, 58, 59. It can be cultured as undifferentiated neural progenitor while after serum withdrawal and cAMP stimulation undergoes cell cycle exit and neuronal differentiation.60 These cells have been used in a variety of experimental settings to understand how drugs differently affect the vulnerability and/or the responsiveness of neural cells according to their differentiation properties.61,62
Differently from temozolomide, a well-known chemotherapeutic drug, regarded as the golden standard in the treatment of glioblastoma, RGWE has been shown to distinguish between glioblastoma and differentiated neuronal cells (Fig. 1). As a matter of fact, RGWE was able to halt the proliferation of glioblastoma cells without harming neuronal differentiated cells. Moreover, RGWE also caused cell death of proliferating, undifferentiated neural cells while promoting plasticity of the differentiated counterpart (Fig. 2). On the contrary, temozolomide was toxic, although to different extent, to both undifferentiated and differentiated neural cells46,56 (Fig. 3). Interestingly, in these experimental settings, rutin has proven to be ineffective against glioblastoma cells or to modify neuronal plasticiy, thus suggesting that either it is not involved in the cell death and plasticity mechanisms or it is needed in combination with other compounds, currently unknown, present in the RGWE.
Fig. 1.

RGWE is able to halt the proliferation of glioblastoma cells without harming neuronal differentiated cells.
Fig. 2.

RGWE causes cell death of proliferating, undifferentiated neural cells while promoting plasticity of the differentiated counterpart.
Fig. 3.

Temozolomide is toxic to both undifferentiated and differentiated neural cells.
4. Conclusions
Past and recent experimental evidence show that Ruta graveolens and/or its main component rutin are endowed with a number of interesting properties that, if properly handled, could be exploited to develop potential drugs, targeting some devastating diseases and pathological conditions such as neurodegeneration and glioblastomas. In particular, it is worth mentioning the recent findings showing the ability of RGWE to distinguish between neuroepithelial cells and normal cells, by halting or killing the former and sparing the latter. Moreover, interestingly when administered on neural differentiated cells RGWE is able to promote cell cycle re-entry and morphological and molecular differentiation. Such an effect on cell plasticity could be exploited in neurodegeneration to both prevent cells from dying and/or to slow down the rate of cell death.
Current results, despite being promising, leave a number of questions open. Future studies will address a number of issues including the effects of Ruta graveolens on different types of brain cells (i.e. neurons, astrocytes, oligodendrocytes, and microglia), the molecular mechanisms underlying its action, the features of ruta-induced neuron differentiation and more importantly the ability to protect the brain in cellular and/or animal models of disease.
Declaration of competing interest
None.
Acknowledgements
"VALERE: VAnviteLli pER la RicErca" (LCD).
Footnotes
Peer review under responsibility of The Center for Food and Biomolecules, National Taiwan University.
Contributor Information
Luca Colucci-D’Amato, Email: luca.colucci@unicampania.it.
Gloria Cimaglia, Email: cimagliag@cardiff.ac.uk, Gloria.cimaglia@ki.se.
References
- 1.Kalra E.K. Nutraceutical-definition and introduction. AAPS PharmSci. 2003;5(3):27–28. doi: 10.1208/ps050325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Nasri H., Baradaran A., Shirzad H., Rafieian-Kopaei M. New concepts in nutraceuticals as alternative for pharmaceuticals. Int J Prev Med. 2014;5(12):1487–1499. https://www.ncbi.nlm.nih.gov/pubmed/25709784 [PMC free article] [PubMed] [Google Scholar]
- 3.Stafford G.I., Pedersen M.E., Staden J Van, Jäger A.K. Review on plants with CNS-effects used in traditional South African medicine against mental diseases. J Ethnopharmacol. 2008;119:513–537. doi: 10.1016/j.jep.2008.08.010. [DOI] [PubMed] [Google Scholar]
- 4.Kannan R., Babu U.V. Identity and pharmacognosy of Ruta graveolens linn. Ancient Sci Life. 2012;32(1):16–19. doi: 10.4103/0257-7941.113792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Colucci-D’Amato L., di Porzio U. Neurogenesis in adult CNS: from denial to opportunities and challenges for therapy. Bioessays. 2008;30(2):135–145. doi: 10.1002/bies.20703. [DOI] [PubMed] [Google Scholar]
- 6.Parray S., Bhat J., Ahmad G., Jahan N. Ruta graveolens: from traditional system of medicine to modern pharmacology: an overview. Am J PharmTech Res. 2012;2:239–252. [Google Scholar]
- 7.Kuzovkina I., Al’terman I., Schneider B. Specific accumulation and revised structures of acridone alkaloid glucosides in the tips of transformed roots of Ruta graveolens. Phytochemistry. 2004;65(8):1095–1100. doi: 10.1016/j.phytochem.2004.03.003. [DOI] [PubMed] [Google Scholar]
- 8.Pollio A., de Natale A., Appetitil E., Aliotta G. Touwaide A Continuity and change in the Mediterranean medical tradition: ruta sp. (Rutaceae) in Hippocratic medicine and present practices. J Ethnopharmacol. 2008;116:469–482. doi: 10.1016/j.jep.2007.12.013. [DOI] [PubMed] [Google Scholar]
- 9.Malik S., Moraes D.F.C., do Amaral F.M.M., Ribeiro M.N.S. Ruta graveolens: phytochemistry, pharmacology, and biotechnology. In: Jha S., editor. Transgenesis and Secondary Metabolism. Springer International Publishing; Cham: 2016. pp. 1–28. [DOI] [Google Scholar]
- 10.Gutiérrez-pajares J.L., Zúñiga L., Pino J. Ruta graveolens aqueous extract retards mouse preimplantation embryo development. Reprod Toxicol. 2003;17:667–672. doi: 10.1016/j.reprotox.2003.07.002. [DOI] [PubMed] [Google Scholar]
- 11.Denham A., Tobyn G., Denham A., Whitelegg M. Singing Dragon; London, UK: 2016. The Western Herbal Tradition: 2000 Years of Medicinal Plant Knowledge.https://www.elsevier.com/books/the-western-herbal-tradition/tobyn/978-0-443-10344-5 Reprint of Ist ed. 2011. [Google Scholar]
- 12.Forsatkar M.N., Hedayatirad M., Luchiari A.C. “ Not tonight zebrafish ”: the effects of Ruta graveolens on reproduction. Pharm Biol. 2018;56(1):60–66. doi: 10.1080/13880209.2017.1421234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Yamada M., Yasuhara H. Clinical pharmacology of MAO Inhibitors : safety and future. Neurotoxicology. 2004;25:215–221. doi: 10.1016/S0161-813X(03)00097-4. [DOI] [PubMed] [Google Scholar]
- 14.Adams M., Gmunder F., Hamburger M. Plants traditionally used in age related brain disorders — a survey of ethnobotanical literature. J Ethnopharmacol. 2007;113:363–381. doi: 10.1016/j.jep.2007.07.016. [DOI] [PubMed] [Google Scholar]
- 15.Stafford G.I., Pedersen P.D., Jäger A.K., Staden J Van. Monoamine oxidase inhibition by southern African traditional medicinal plants. South Afr J Bot. 2007;73:384–390. doi: 10.1016/j.sajb.2007.03.001. [DOI] [Google Scholar]
- 16.Russo P., Frustaci A., Del Bufalo A., Fini M., Cesario A. From traditional European medicine to discovery of new drug candidates for the treatment of dementia and Alzheimer’s disease: acetylcholinesterase inhibitors. Curr Med Chem. 2013;20(8):976–983. doi: 10.2174/0929867311320080002. [DOI] [PubMed] [Google Scholar]
- 17.Wai Kan Yeung A., Georgieva M.G., Atanasov A.G., Tzvetkov N.T. Monoamine oxidases (MAOs) as privileged molecular targets in neuroscience: research literature analysis. Front Mol Neurosci. 2019 doi: 10.3389/fnmol.2019.00143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hauser R.A., Li R., Perez A., Ren X. Longer duration of MAO-b inhibitor exposure is associated with less clinical decline in Parkinson’s disease: an analysis of NET-PD LS1. J Parkinsons Dis. 2017;7(1):117–127. doi: 10.3233/JPD-160965. [DOI] [PubMed] [Google Scholar]
- 19.Naoi M. Maruyama W Monoamine oxidase inhibitors as neuroprotective agents in age-dependent neurodegenerative disorders. Curr Pharmaceut Des. 2010;16(25):2799–2817. doi: 10.2174/138161210793176527. [DOI] [PubMed] [Google Scholar]
- 20.Gentile M.T., Russo R., Pastorino O. Ruta graveolens water extract inhibits cell-cell network formation in human umbilical endothelial cells via MEK-ERK1/2 pathway. Exp Cell Res. 2018 doi: 10.1016/j.yexcr.2018.01.025. July 2017. [DOI] [PubMed] [Google Scholar]
- 21.Mazumder M.K., Borah A., Choudhury S. Inhibitory potential of plant secondary metabolites on anti-parkinsonian drug targets: relevance to pathophysiology, and motor and non-motor behavioural abnormalities. Med Hypotheses. 2020;137:109544. doi: 10.1016/j.mehy.2019.109544. Jan 2. [DOI] [PubMed] [Google Scholar]
- 22.Grzybowski A., Pietrzak K. Albert Szent-Györgyi (1893-1986): the scientist who discovered vitamin C. Clin Dermatol. 2013;31(3):327–331. doi: 10.1016/J.CLINDERMATOL.2012.08.00. [DOI] [PubMed] [Google Scholar]
- 23.Kalra E.K., Russo P., Frustaci A. Rutin abrogates manganese—induced striatal and hippocampal toxicity via inhibition of iron depletion, oxidative stress, inflammation and suppressing the NF-κB signaling pathway. Int J Prev Med. 2013;5(8):92–99. doi: 10.1208/ps050325. [DOI] [PubMed] [Google Scholar]
- 24.Nkpaa K.W., Onyeso G.I. Rutin attenuates neurobehavioral deficits, oxidative stress, neuro-inflammation and apoptosis in fluoride treated rats. Neurosci Lett. 2018;682:92–99. doi: 10.1016/j.neulet.2018.06.023. [DOI] [PubMed] [Google Scholar]
- 25.Yu X.L., Li Y.N., Zhang H. Rutin inhibits amylin-induced neurocytotoxicity and oxidative stress. Food Funct. 2015;6(10):3296–3306. doi: 10.1039/c5fo00500k. [DOI] [PubMed] [Google Scholar]
- 26.Magalingam K.B., Radhakrishnan A., Haleagrahara N. Rutin, a bioflavonoid antioxidant protects rat pheochromocytoma (PC-12) cells against 6-hydroxydopamine (6-OHDA)-induced neurotoxicity. Int J Mol Med. 2013;32(1):235–240. doi: 10.3892/ijmm.2013.1375. [DOI] [PubMed] [Google Scholar]
- 27.Magalingam K.B., Radhakrishnan A., Ramdas P., Haleagrahara N. Quercetin glycosides induced neuroprotection by changes in the gene expression in a cellular model of Parkinson’s disease. J Mol Neurosci. 2015;55(3):609-617. doi: 10.1007/s12031-014-0400-x. [DOI] [PubMed] [Google Scholar]
- 28.Khan M.M., Raza S.S., Javed H. Rutin protects dopaminergic neurons from oxidative stress in an animal model of Parkinson’s disease. Neurotox Res. 2012;22(1):1–15. doi: 10.1007/s12640-011-9295-2. [DOI] [PubMed] [Google Scholar]
- 29.Li P., Matsunaga K., Ohizumi Y. Enhancement of the nerve growth factor-mediated neurite outgrowth from PC12D cells by Chinese and Paraguayan medicinal plants. Biol Pharm Bull. 1999;22(7):752–755. doi: 10.1248/bpb.22.752. [DOI] [PubMed] [Google Scholar]
- 30.Tábi T., Vécsei L., Youdim M.B., Riederer P., Szök É. Selegiline: a molecule with innovative potential. J Neural Transm. 2019 doi: 10.1007/s00702-019-02082-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Adsersen A., Gauguin B., Gudiksen L., Jäger A.K. Screening of plants used in Danish folk medicine to treat memory dysfunction for acetylcholinesterase inhibitory activity. J Ethnopharmacol. 2006;104(3):418–422. doi: 10.1016/j.jep.2005.09.032. [DOI] [PubMed] [Google Scholar]
- 32.Wszelaki N., Kuciun A., Kiss A.K. Screening of traditional European herbal medicines for acetylcholinesterase and butyrylcholinesterase inhibitory activity. Acta Pharm. 2010;60(1):119–128. doi: 10.2478/v10007-010-0006-y. [DOI] [PubMed] [Google Scholar]
- 33.Russo P., Frustaci A., Del Bufalo A., Fini M., Cesario A. From traditional European medicine to discovery of new drug candidates for the treatment of dementia and Alzheimer’s disease: acetylcholinesterase inhibitors. Curr Med Chem. 2013;20(8):976–983. doi: 10.2174/0929867311320080002. [DOI] [PubMed] [Google Scholar]
- 34.Atri A. Current and future treatments in alzheimer’s disease. Semin Neurol. 2019;39(2):227–240. doi: 10.1055/s-0039-1678581. [DOI] [PubMed] [Google Scholar]
- 35.Wyss-Coray T., Rogers J. Inflammation in Alzheimer disease-a brief review of the basic science and clinical literature. Cold Spring Harb Perspect Med. 2012;2(1) doi: 10.1101/cshperspect.a006346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Salem A.M., Sabry G.M., Ahmed H.H. Amelioration of neuroinflammation and apoptosis characterizing Alzheimer’s disease by natural products. Int J Pharm Pharmaceut Sci. 2013;5(SUPPL. 2):87–94. [Google Scholar]
- 37.Shojaii A., Ghods R., Fard M.A. Medicinal herbs in Iranian traditional medicine for learning and memory. Iran J Med Sci. 2016;41(3 suppl l) https://www.ncbi.nlm.nih.gov/pubmed/27840509 S43-S43. [PMC free article] [PubMed] [Google Scholar]
- 38.Xu P.X., Wang S.W., Yu X.L. Rutin improves spatial memory in Alzheimer’s disease transgenic mice by reducing Aβ oligomer level and attenuating oxidative stress and neuroinflammation. Behav Brain Res. 2014;264:173-180. doi: 10.1016/j.bbr.2014.02.002. [DOI] [PubMed] [Google Scholar]
- 39.Wang S.W., Wang Y.J., Su Y.J. Rutin inhibits β-amyloid aggregation and cytotoxicity, attenuates oxidative stress, and decreases the production of nitric oxide and proinflammatory cytokines. Neurotoxicology. 2012;33(3):482-490. doi: 10.1016/j.neuro.2012.03.003. [DOI] [PubMed] [Google Scholar]
- 40.Pan R.Y., Ma J., Kong X.X. Sodium rutin ameliorates Alzheimer’s disease-like pathology by enhancing microglial amyloid-β clearance. Sci Adv. 2019;5(2) doi: 10.1126/sciadv.aau6328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Villa V., Thellung S., Corsaro A. Celecoxib inhibits prion protein 90-231-mediated pro-inflammatory responses in microglial cells. Mol Neurobiol. 2016;53(1):57–72. doi: 10.1007/s12035-014-8982-4. [DOI] [PubMed] [Google Scholar]
- 42.McColgan P., Tabrizi S.J. Huntington’s disease: a clinical review. Eur J Neurol. 2018;25(1):24–34. doi: 10.1111/ene.13413. [DOI] [PubMed] [Google Scholar]
- 43.Suganya S.N., Sumathi T. Effect of rutin against a mitochondrial toxin, 3-nitropropionicacid induced biochemical, behavioral and histological alterations-a pilot study on Huntington’s disease model in rats. Metab Brain Dis. 2017;32(2):471–481. doi: 10.1007/s11011-016-9929-4. [DOI] [PubMed] [Google Scholar]
- 44.Cordeiro L.M., Machado M.L., da Silva A.F. Rutin protects Huntington’s disease through the insulin/IGF1 (IIS) signaling pathway and autophagy activity: study in Caenorhabditis elegans model [published online ahead of print, 2020 Apr 8] Food Chem Toxicol. 2020;141:111323. doi: 10.1016/j.fct.2020.111323. [DOI] [PubMed] [Google Scholar]
- 45.Colucci-D’Amato L., Gentile M.T. Neural stem Cell : tools to unravel pathogenetic mechanisms and to test novel drugs for CNS diseases. In: Bonfanti L., editor. Neural Stem Cells. New Perspectives IntechOpen; 2013. [DOI] [Google Scholar]
- 46.Gentile M.T., Ciniglia C., Reccia M.G., Volpicelli F. Ruta graveolens L . Induces death of glioblastoma cells and neural progenitors , but not of neurons , via ERK 1/2 and AKT activation. PloS One. 2015:1–18. doi: 10.1371/journal.pone.0118864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Colucci-D’Amato L., di Porzio U. Neurogenesis in adult CNS: from denial to opportunities and challenges for therapy. Bioessays. 2008;30(2):135–145. doi: 10.1002/bies.20703. [DOI] [PubMed] [Google Scholar]
- 48.Nizzari M., Barbieri F., Gentile M.T. Amyloid-β protein precursor regulates phosphorylation and cellular compartmentalization of microtubule associated protein tau. J Alzheimers Dis. 2012;29(1):211-227. doi: 10.3233/JAD-2011-101590. [DOI] [PubMed] [Google Scholar]
- 49.Fusco F.R., Anzilotti S., Giampà C. Changes in the expression of extracellular regulated kinase (ERK 1/2) in the R6/2 mouse model of Huntington’s disease after phosphodiesterase IV inhibition. Neurobiol Dis. 2012;46(1):225–233. doi: 10.1016/j.nbd.2012.01.011. [DOI] [PubMed] [Google Scholar]
- 50.Zeng W., Zhang W., Lu F., Gao L., Gao G. Resveratrol attenuates MPP + -induced mitochondrial dysfunction and cell apoptosis via AKT/GSK-3β pathway in SN4741 cells. Neurosci Lett. 2017;637:50–56. doi: 10.1016/j.neulet.2016.11.054. [DOI] [PubMed] [Google Scholar]
- 51.Cui Q., Xin Li, Zhu H. Curcumin ameliorates dopaminergic neuronal oxidative damage via activation of the Akt/Nrf2 pathway. Mol Med Rep. 2016:1381–1388. doi: 10.3892/mmr.2015.4657. [DOI] [PubMed] [Google Scholar]
- 52.Cao Q., Qin L., Huang F. Amento flavone protects dopaminergic neurons in MPTP-induced Parkinson ’ s disease model mice through PI3K/Akt and ERK signaling pathways. Toxicol Appl Pharmacol. 2017;319:80–90. doi: 10.1016/j.taap.2017.01.019. [DOI] [PubMed] [Google Scholar]
- 53.Mahoney C., Feliciano D.M., Bordey A., Hartman N.W. Switching on mTORC1 induces neurogenesis but not proliferation in neural stem cells of young mice. Neurosci Lett. 2016;614:112–118. doi: 10.1016/j.neulet.2015.12.042. [DOI] [PubMed] [Google Scholar]
- 54.Lee D.Y. Roles of mTOR signaling in brain development. Exp Neurobiol. 2015;24(3):177–185. doi: 10.5607/en.2015.24.3.177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Guérout N., Li X., Barnabé-heider F. Cell fate control in the developing central nervous system. Exp Cell Res. 2014;321(1):77–83. doi: 10.1016/j.yexcr.2013.10.003. [DOI] [PubMed] [Google Scholar]
- 56.Cimaglia G., Gentile M.T., Pastorino O., Colucci-D’Amato L. Society for Neuroscience; Chicago: 2019. Ruta Graveolens Stimulates Neural Plasticity of Human and Mouse Embryonic Mesencephalic Neuronal Cell Progenitors. 640.11/A40. [Google Scholar]
- 57.Chambery A., Colucci-D’Amato L., Vissers J.P. J Proteome Res. 2009 Jan;8(1):227–238. doi: 10.1021/pr800454n. [DOI] [PubMed] [Google Scholar]
- 58.Colucci-D’Amato L., Farina A., Vissers J.P., Chambery A. Quantitative neuroproteomics: classical and novel tools for studying neural differentiation and function. Stem Cell Rev Rep. 2011;7(1):77-93. doi: 10.1007/s12015-010-9136-3. [DOI] [PubMed] [Google Scholar]
- 59.Severino V., Farina A., Colucci-D’Amato L. Secretome profiling of differentiated neural mes-c-myc A1 cell line endowed with stem cell properties. Biochim Biophys Acta. 2013;1834(11):2385–2395. doi: 10.1016/j.bbapap.2012.12.005. [DOI] [PubMed] [Google Scholar]
- 60.Colucci-D’Amato G.L., Tino A., Pernas-Alonso R. Neu ronal and glial properties coexist in a novel mouse CNS immortalized cell line. Exp Cell Res. 1999;252(2):383–391. doi: 10.1006/excr.1999.4636. 1 November. [DOI] [PubMed] [Google Scholar]
- 61.Di Lieto A., Leo D., Volpicelli F. FLUOXETINE modifies the expression of serotonergic markers in a differentiation-dependent fashion in the mesencephalic neural cell line A1 mes c-myc. Brain Res. 2007;1143:1–10. doi: 10.1016/j.brainres.2007.01.076. [DOI] [PubMed] [Google Scholar]
- 62.Gentile M.T., Nawa Y., Lunardi G. Tryptophan hydroxylase 2 (TPH2) in a neuronal cell line: modulation by cell differentiation and NRSF/rest activity. J Neurochem. 2012;123(6):963–970. doi: 10.1111/jnc.12004. [DOI] [PubMed] [Google Scholar]