

Abstract
Amino acids are ubiquitous vital biomolecules found in all kinds of living organisms including those in the microbial world. They are utilised as nutrients and control many biological functions in microorganisms such as cell division, cell wall formation, cell growth and metabolism, intermicrobial communication (quorum sensing), and microbial-host interactions. Amino acids in the form of enzymes also play a key role in enabling microbes to resist antimicrobial drugs. Antimicrobial resistance (AMR) and microbial biofilms are posing a great threat to the world’s human and animal population and are of prime concern to scientists and medical professionals. Although amino acids play an important role in the development of microbial resistance, they also offer a solution to the very same problem i.e., amino acids have been used to develop antimicrobial peptides as they are highly effective and less prone to microbial resistance. Other important applications of amino acids include their role as anti-biofilm agents, drug excipients, drug solubility enhancers, and drug adjuvants. This review aims to explore the emerging paradigm of amino acids as potential therapeutic moieties.
Keywords: amino acids, antimicrobial resistance, quorum sensing, microbial biofilm, solubility, excipients, adjuvants
1. Introduction
Amino acids are important biomolecules and the very basic constituents or building blocks of proteins [1]. They have at least one amino group, one carboxyl group, and a side chain in their structures [2,3]. There are 20 different amino acids, represented by a general formula H2N-CαH(R)-CO2H (where -R is a side chain with an exception of glycine, in which the side chain -R has been replaced by -H), which are responsible for protein formation in all living organisms [4,5].
Several amino acids (in different proportions) are joined together by amide bonding (also known as peptide bonding) in such a way that the carboxyl group of one amino acid links to the amino group of another amino acid with the loss of a water molecule (per each peptide bond) to form protein polymers [6]. Figure 1a,b illustrates a general chemical structure for 19 of the 20 amino acids and glycine, respectively.
Figure 1.

(a) depicts a general chemical structure of an amino acid; (b) is the structure of amino acid glycine.
Besides the most common 20 proteogenic amino acids, there are hundreds of other non-protein amino acids with varied biological functions, found in all kinds of living systems including humans, plants (abundantly found in legumes and seed), bacteria, and fungi [7,8,9]. Unlike proteogenic amino acids, non-proteogenic amino acids are not naturally encoded with a genetic code and therefore, are also referred to as non-coded amino acids [10]. Non-proteogenic amino acids are secondary metabolites, produced from natural proteogenic amino acids and are usually toxic in nature [11]. Hypoglycine (also written as hypoglycin), canavanine, and mimosine are examples of non-proteogenic amino acids [9].
Non-proteogenic amino acids are able to pose themselves as if they have certain properties such as chemical structure, size, shape, and charge, etc., similar to those exhibited by normal proteogenic amino acids [12]. By mimicking one or more of these properties, non-proteogenic amino acids are mistaken for normal amino acids and during protein biosynthesis, they are incorporated into different proteins [13]. This incorporation of non-proteogenic amino acids results in the formation of unnatural proteins with improper functionalities and hence causes undesired consequences [9]. Azetidine-2-carboxilic acid is a non-proteogenic amino acid (found in sugar beets), due to structural similarities, behaves the same way as alanine and proline [14]. Azetidine-2-carboxilic acid, if consumed during pregnancy, is believed to be the cause of multiple sclerosis in newborns [13].
Amino acids play an important role in microbial metabolism and in facilitating microbial growth, microbial biofilm formation, and its dispersal [15]. Based on the source of amino acids and nature of the side chain, amino acids can be categorised as essential and non-essential amino acids, acidic, basic and neutral amino acids, polar and nonpolar amino acids, hydrophilic and hydrophobic amino acids [16].
Essential amino acids are those amino acids that cannot be synthesised by living organisms and must be supplemented from dietary means [17]. Essential amino acids such as isoleucine, leucine, lysine, methionine, phenylalanine, taurine, threonine, tryptophan, valine, arginine, glycine, and histidine are the common ones found in almost all living organisms in varied proportions [18]. Non-essential amino acids are biosynthesised inside the cells of living organisms and are rarely required to be supplemented or obtained from dietary sources [17]. Non-essential amino acids include alanine, aspartate, asparagine, arginine, cysteine, glutamate, glutamine, glycine, proline, serine, and tyrosine [19].
This review is an effort to highlight different physicochemical properties of amino acids, their microbial biosynthesis, their role in intermicrobial interaction as well as microbe-host interactions, and more importantly, the role played by amino acids in the fight against antimicrobial resistance (AMR) as potential antimicrobial agents, antibiofilm agents, drug adjuvants and drug excipients.
Types of Amino Acids and Chirality
Amino acids can be classed as acidic, basic, or neutral amino acids on the basis of their side chain. An acidic amino acid possesses an acidic functional group in its side chain and a basic amino acid has a basic functional group attached to its side chain, while a neutral amino acid has neither an acidic nor a basic functional group attached to its side chain [4].
Examples of acidic amino acids are aspartic acid and glutamic acid, basic amino acids include lysine, arginine, and histidine, while serine threonine and tyrosine are examples of neutral amino acids [20,21]. Depiction of chemical structures, each from acidic, basic, and neutral amino acids have been given in Figure 2.
Figure 2.

Chemical structures of acidic, basic and neutral amino acids.
The most common 20 proteogenic amino acids (with the exception of glycine) exhibit chirality as the central carbon in their molecules is attached to four different groups, enabling each of these amino acids to exist as two mirror images that are non-superimposable which are called enantiomers [22]. Glycine being the only achiral amino acid as has two hydrogens attached to the central carbon i.e., the side chain -R = H [4].
Both enantiomers (isomers) of an amino acid have similar properties but differ in their interaction with plane-polarised light i.e., one enantiomeric form rotates the light towards the right side and is called dextro- (right handed) rotatory, abbreviated as d-, whilst the other form rotates the light towards the left side and is therefore referred to as laevo- (left handed) rotatory, abbreviated as l- [23]. Figure 3 is a general representation of d- and l- form of a typical amino acid.
Figure 3.

Two enantiomeric forms, l- and d- of an amino acid.
With the exception of glycine, which is an achiral amino acid, all other 19 amino acids have a center of chirality and exist in two enantiomeric forms i.e., l-amino acids and d-amino acids [24].
In chemistry, d-enantiomer of an amino acid is also referred to as its (R)-enantiomer and l-enantiomer as its S-enantiomer, where “R” and “S” both come from Latin words rectus and sinister (right-handed and left-handed respectively). This type of nomenclature follows Cahn–Ingold–Prelog (CIP) convention, that assigns priorities (based on atomic numbers or atomic mass) to different constituents attached to the asymmetric central carbon [25]. According to R, S system, all l-amino acids are (S)-amino acids with the exception of l-cysteine i.e., l-cysteine is not an (S)-cysteine but is an (R)-cysteine instead [26]. Figure 4 depicts the (S) and (R)-enantiomeric structures for the respective l and d-enantiomers an amino acid.
Figure 4.

(S) and (R)-enantiomers of an amino acid, representing its respective l and d-enantiomers.
Chirality of amino acids is of significant importance when it comes to peptide drug designing and synthetic peptide chemistry [27]. It also plays a significant role in determining if a particular amino acid will participate in protein formation or it will serve to regulate other cellular functions. For instance, l-amino acids are proteogenic and will form proteins as well as participate in microbial cell wall formation, whereas d-amino acids are non-proteogenic and regulate different functions such as microbial cell wall formation and dispersal of microbial biofilm [28,29]. The chiral behaviour of amino acids is also responsible for stereoselective catabolism of amino acids i.e., D-amino acids will be catabolised only by d-amino acid oxidase (DAAOs) enzymes and l-amino acids will have their corresponding l-amino acid oxidases (LAAOs) for their catabolism [30].
Chirality of amino acids also stereo-select the antimicrobial properties of LAAOs and DAAOs i.e., LAAOs will exhibit antibacterial activity only in the presence of corresponding substrate (l-amino acids) and DAAOs will exhibit antibacterial activity in the presence of d-amino acids [31,32]. However, the antibacterial mechanism of both DAAOs and LAAOs remains the same i.e., the generation of hydrogen peroxide (H2O2) during the oxidative deamination of stereo-selected amino acids [33].
2. Microbial Biosynthesis of Amino Acids
Microbial biosynthesis of amino acids requires carbon that comes from either the citric acid cycle, glycolytic pathway or pentose phosphate pathway, and nitrogen that comes from ammonia (microbes convert nitrogen gas into ammonia) [34]. Microorganisms are generally capable of producing all the required proteogenic amino acids, however, the mechanism of amino acid biosynthesis is different in different microorganisms [35,36].
Different organic intermediates and precursors undergo biochemical reactions and form different amino acids [37]. Some precursors such as oxaloacetate from the citric acid cycle, α-ketoglutarate from Krebs cycle, and 3-phosphoglycerate from Calvin cycle are converted into amino acids that act as intermediates in the biosynthesis of other amino acids [34,38,39,40]. Table 1 enlists different precursors for the respective intermediates and amino acids.
Table 1.
Precursors involved in microbial biosynthesis of amino acids. Some of the amino acids mentioned in Table 3, do not involve any intermediates and their formation involves their respective precursors only i.e., serine, tryptophan, phenylalanine, valine, etc., while others, such as tyrosine and isoleucine are formed through their respective intermediates. Some amino acids also serve as intermediates in the formation of other amino acids i.e., threonine and phenylalanine serve as intermediates for isoleucine and tyrosine, respectively.
| Precursors | Intermediates | Amino Acids |
|---|---|---|
| α-ketoglutarate | Glutamate | Glutamine |
| Proline | ||
| Arginine | ||
| 3-phosphoglycerate | Serine | Glycine |
| Cysteine | ||
| Oxaloacetate | Aspartate | Asparagine |
| Methionine | ||
| Isoleucine | ||
| Lysine | ||
| Phosphoenolpyruvate + Erythrose 4-phosphate | ---------------------- | Tyrosine |
| Phenylalanine | ||
| Tryptophan | ||
| Pyruvate | ----------------------- | Alanine |
| Valine | ||
| Leucine | ||
| Ribose 5-phosphate | ------------------------ | Histidine |
2.1. Fungal Biosynthesis of Amino Acids
Fungi synthesise all the amino acids required for protein synthesis and other metabolic activities using different enzymes. The mechanism of biosynthesis of certain amino acids (that cannot be synthesised by mammals) has been reported to be similar in fungi and bacteria with the exception of biosynthesis of l-lysine through α-aminoadipate pathway which is only found in fungi. Biosynthesis of lysine is catalysed by enzymes such as homocitrate synthase, homoaconitase, homoisocitrate dehydrogenase, α-aminoadipate aminotransferase, α-aminoadipate reductase, saccharopine reductase, and saccharopine dehydrogenase [41,42,43] as shown in Figure 5.
Figure 5.

Schematic representation of l-lysine biosynthesis via α-aminoadipate pathway. It illustrates that precursors acetyl-CoA and α-ketoglutarate in the presence of homocitrate synthase form homocitrate (a tricarboxylic acid). Homoaconitase enzyme dehydrates homocitrate to form homoaconitate (homoaconitatic acid) that leads to the formation of homoisocitrate in the presence of homoaconitase and water. Homoisocitrate dehydrogenase converts homoisocitrate into α-ketoadipate. NAD is converted into NADH and CO2 is generated during this step. Now, glutamate reacts with α-ketoadipate and forms α-amino adipate which converts into l-lysine. 2-oxoglutatarte (de-amination of glutamate) is also generated in the process. Concept adopted (and modified) from Fazius, F., 2012, Jastrzebowska, K., 2015 and Miyazaki, J., 2001 [41,42,43].
Despite being able to biosynthesise amino acids, fungi have also evolved certain mechanisms such as nitrogen catabolic repression (NCR), transceptor-mediated amino acid sensing, Ssy1-Ptr3-Ssy5 (SPS) and target of rapamycin (TOR) pathway to monitor and use the amino acids present in the environment [44]. SPS is used to sense and uptake exogenous amino acids and TOR serves as a sensory system to monitor intracellular amino acids [45].
2.2. Bacterial Biosynthesis of Amino Acids
Bacteria such as Escherichia coli can biosynthesise all the required 20 amino acids (proteogenic amino acids), including the essential ones, while others such as Lactobacillus plantarum need to acquire them from their external environment [38,46,47,48]. Several enzymes participate to catalyse the formation of each amino acid e.g., bacterial biosynthesis of branched-chain amino acids (leucine, isoleucine, and valine) for instance, is catalysed by at least eight different enzymes i.e., l-threonine dehydratase, acetolactate synthase, keto acid isomeroreductase, dihydroxy acid dehydratase, isopropylmalate synthase, isopropylmalate isomerase, isopropylmalate dehydrogenase, and branched-chain aminotransferase [38,49]. Figure 6 illustrates various steps involved in bacterial biosynthesis of branched amino acids.
Figure 6.

Schematic representation of branched chain amino acids in bacteria. It shows how two pyruvate molecules (product of glycolysis) in the presence of acetohydroxy acid synthase react to form acetolactate. Isomero-reductase enzyme converts acetolactate into an intermediate called dihydroxyisovalerate which, in the presence of dihydroxy acid dehydratase enzyme, dehydrates and produces the final intermediate, ketoisovalerate. Enzyme transaminase B converts ketoisovalerate into l-valine, while enzyme leu BCD converts it into l-leucine. Concept adopted from Amorim Franco, T.M., 2017 [38].
Due to a variety of industrial applications (pharmaceuticals, cosmetics, animal feeds, etc.) of amino acids, microbial biosynthesis of amino acids has been adopted as a means of large-scale production by the process of fermentation [50]. The fermentation process involves the growth of microorganisms on less expensive growth media on the industrial level to produce tons of high-quality amino acids each year e.g., industrial production of l-glutamic acid and l-lysine [51,52,53].
3. Antimicrobial Resistance (AMR) and Role of Amino Acids
Antimicrobial resistance (AMR) is not only taking its toll on the world’s human and animal population, but is also causing serious financial losses to the global economy [54]. According to a report published by the World Health Organisation (WHO) on AMR related worldwide mortalities, at least 700,000 people die each year because of AMR. This number expected to escalate to 10 million per year by the year 2050 [55]. To avert serious pandemics such as the situation due to AMR, the world needs novel therapeutic strategies such as the development of novel antimicrobial drugs as well as repurposing the existing ones, including the drugs that were previously abandoned for different reasons.
Microorganisms have developed different strategies to resist antimicrobial drugs, either by reducing their efficacy or by rendering the drugs fully inactive. Different mechanisms through which microbes develop and spread resistance against antimicrobials include efflux pumps, modification of drugs, alteration of the target sites, decrease in cellular permeability to reduce drug penetration, enzymatic degradation of antimicrobial compounds, and biofilm formation [56].
Microbial cellular envelopes have protein-made designated transport channels called porins [57]. These porins act in the same way as efflux pumps that expel antimicrobial drugs out of the cell, thereby reducing the drug concentration to avoid cytocidal effects [58]. The lipophilic outer membrane in Gram-negative bacteria limits the passage of hydrophilic drugs through porins (protein channels meant for the passage of substances) to a level where the drug concentration is no longer bactericidal [59].
Amino acids (in the form of enzymes) also contribute to antimicrobial resistance in different ways i.e., drug modification, and inactivation via enzymatic degradation e.g., phosphoethanolamine transferase, hydrolases and redox enzymes [60]. Enzymes such as amidases and acyl transferases degrade the antimicrobial drugs (β-lactams and macrolides) while others such as epoxidases, macrolide esterases, etc., modify the antimicrobial drugs (rifamycin, aminoglycoside, etc.) and help the microbes to resist the antimicrobial effect of the drugs [61].
Alongside other strategies (such as developing novel drugs and repurposing the existing ones), amino acids can also be important drug candidates in the fight against antimicrobial resistance, both as antimicrobial as well as antibiofilm agents [62]. Amino acids have been used to produce antimicrobial peptides that are more efficient as antimicrobial and antibiofilm agents and less prone to resistance [63,64]. Amino acids have also been used to increase the efficacy (in vitro) of existing drugs i.e., the efficacy of antimicrobial drug trimethoprim has been enhanced by amino acids-based trimethoprim salt formation [65].
Other applications of amino acids in the fight against AMR, include their use as antibiofilm agents, solubility enhancers for existing drugs and their physical combination with antimicrobial drugs to enhance their efficacy by additive effect or synergism [62,66,67,68]. Different roles and applications of amino acids in microbial control and tackling AMR, have been discussed in the following sections.
4. Role of Amino Acids in Microbial Infections and Quorum Sensing
In any particular microbial-host environment, microbes and their host communicate at the site of infection [69,70]. D-amino acids, in particular, which facilitate communication amongst the microbial communities, play a role in microbial-host interaction as well as affect the host immune system by functioning as host defense peptides [71,72,73].
Trillions (10–100 trillions) of microorganism including archaea, bacteria, fungi, viruses, and protozoans inhabit the human body (both, externally and internally), living in a symbiotic environment collectively known by the term “microbiota” whereas the total number of microbial genes in a human microbiota are referred to as the human microbiome [74,75,76].
The human microbiome, particularly hosted by the human gut (also known as a virtual organ within an organ due to its collective metabolic activity), plays an important role in homeostasis (a balanced, stable physical and chemical environment) and immune system, and any imbalance in the microbiome can have its consequences in the form of pathogenic and other diseases [77,78].
The symbiotic relationship is beneficial for human gut microbiota and the host itself, where the microbes (bacteria in particular) break down the available proteins in the host environment and utilise the resultant amino acids, while their human host is benefited from the amino acids secreted by the microbes [79]. These microbial amino acids can, sometimes, if secreted in excess, can disturb the normal human microbiota (a condition also known as dysbiosis), leading to the production of short-chain fatty acids (SCFA) causing liver problems, obesity and diabetes [80].
Microorganisms exchange cell to cell information and communicate to one another through an intercellular microbial system of communication, known as quorum sensing (QS) [81]. Microbial quorum sensing is activated by certain extracellular chemical signals, called autoinducers [82,83]. Autoinducers that activate the quorum sensing in Gram-positive bacteria are peptides [84]. The quorum sensing peptides (QSP) are composed of different amino acids such as serine, tyrosine, tryptophan, isoleucine, leucine, cysteine, threonine, glycine, glutamic acid, phenyl alanine, proline, and valine [85]. Quorum sensing plays a key role in controlling different microbial properties such as microbial biofilm formation, antimicrobial resistance and microbial infections [86].
The initiation of microbial infection leads to the activation of host immune system which prompts a hypercatabolic response from the host and thus results in an increased consumption of host amino acids and other nutrients such as vitamins, fatty acids, etc. [87]. Microbial infections cause amino acid depletion and a loss of host body proteins, hence it is important to provide the infected host with extra dietary proteins and amino acids (20 to 25% extra as compared to normal intake) during any microbial infection-mediated hypercatabolic phase and the recovery phase, to stop any further depletion of proteins [88].
Amino acids regulate different biological events in microbes such as spore germination, microbial growth, and remodeling of the microbial cell wall [28]. Microbial infections are a direct consequence of microbial growth as microbes utilise amino acids as a source of nutrients and energy for their survival [89,90].
Microorganisms such as bacteria and fungi, when faced with adverse environmental conditions (either living freely or on/inside a host) such as lack of essential nutrients, lack of moisture or even when under the influence of toxic chemicals and high temperatures, produce certain types of highly infectious cells, called spores which can survive the aforementioned extreme conditions [91,92]. Spores germinate into active cells after sensing certain biomolecules, known as germinants (which are amino acids and sugar molecules), and cause pathogenic illnesses [93]. Amino Acids also play an important role in the development of microbial appendages (such as pili in bacteria and hyphae in fungi), that are used for nutrients uptake, locomotion and more importantly for adhesion to the host surface, thereby help in developing and spreading microbial diseases [28,94,95,96].
Despite their role in the development and spread of microbial infections, certain peptides and amino acids, d-amino acids in particular, have been reported to exhibit broad-spectrum antimicrobial, anti-quorum sensing as well as antibiofilm activity. They are less likely to be resisted by microbes, hence offer an advantageous alternative to traditional antimicrobial drugs [97,98,99,100]. The aforementioned different roles of amino acids have been discussed in detail, in the forthcoming sections.
5. Role of Amino Acids in Bacterial Cell Wall Formation and Controlling Planktonic Bacteria
Amino acids perform different functions in planktonic bacteria, ranging from exhibiting protective roles to being used as nutritional substrates. Bacterial peptidoglycan (PG), an important functionary in the biosynthesis of the bacterial cell wall, has amino acid constituents in it [101]. PG is a polymer that gives strength to the bacterial cell wall and has sugar molecules alongside its amino acid residues [102]. PG relies on penicillin-binding proteins (PBP) for its synthesis, while its strength and elasticity are dependent on amino acids that regulate the function of PBP [103].
The synthesis of peptidoglycan takes place in the cytoplasm in three stages i.e., i) synthesis of nucleotide precursors namely UDP-N-acetylglucosamine and UDP-N-acetylmuramyl pentapeptide, ii) assembly of precursors into monomer subunits and iii) polymerisation of monomers in the presence of glycosyltransferases [104]. The precursor nucleotide UDP-N-acetylglucosamine is synthesised from fructose-6-phosphate in the presence of enzyme glucoamylase (Glm), the precursor UDP-N-acetylglucosamine is converted into nucleotide UDP-N-acetylmuramyl pentapeptide by MurA, MurB, MurC, MurD, MurE and MurF enzymes [105,106]. A typical pentapeptide has -(l-alanine-d-glutamine-l-lysine-d-alanine-d-alanine)- sequence of amino acids [107].
The precursor nucleotides are then bonded to undecaprenyl phosphate and form the monomeric lipid-anchored disaccharide pentapeptide units (also known as lipid II) which are cross-linked and polymerised by enzymes glycosyltransferases and penicillin-binding proteins (PBPs) to form peptidoglycan layers [104,108,109]. Figure 7 is a depiction of disaccharide pentapeptide monomer of peptidoglycan.
Figure 7.

Disaccharide pentapeptide monomeric unit of peptidoglycan.
The basic two-dimensional structure of peptidoglycan mesh remains the same in both Gram-positive and Gram-negative bacteria i.e., glycan strands are oriented in circumferential order and crossly linked by peptide side chains [110]. Figure 8 is a general depiction of pentaglycine cross-linkages in Gram-negative and Gram-positive bacterial peptidoglycan.
Figure 8.

Pentaglycine cross-linkages in Gram-negative and Gram-positive bacterial peptidoglycan. Disaccharide pentapeptide monomer in Gram-negative bacterial peptidoglycan possesses meso-diaminopimelic acid (mDAP), whereas Gram-positive bacterial peptidoglycan has l-lysine instead. Basic concept (with adaptations) has been drawn from Irazoki, Hernandez, and Cava, 2019 [111]. Figure 8 has been drawn using Microsoft Word.
Gram-positive bacteria possess multilayers (10 or more) of peptidoglycan and is about 50% of the cell wall weight, whereas the peptidoglycan in Gram-negative bacteria is 1–2 layered, constituting 10–20% of the cell envelop weight [112,113].
The thin layered peptidoglycan of Gram-negative bacteria is sandwiched between the outer membrane (OM) and inner membrane (IM), compared to the multi-layered thick peptidoglycan of Gram-positive bacteria which is located exterior to the cytoplasmic membrane and is exposed to the extracellular environment [114]. Figure 9 is a depiction of Gram-negative and Gram-positive bacterial cell wall types, showing the location of peptidoglycan layers.
Figure 9.

Gram-negative and Gram-positive bacterial cell walls, showing the location of peptidoglycan (PG) alongside other components in the respective cell walls. Basic concept (with some changes) has been drawn from MartÃ-nez-Carmona, Gun’ko and Vallet-RegÃ, 2018 [115]. Figure 9 has been drawn using ChemDraw software.
Peptidoglycan of the bacterial cell wall serves as a target for many antibiotics [113]. It also serves as a target for antimicrobial amino acids. The incorporation of exogenous d-amino acids into bacterial PG during its biosynthesis disrupts the natural sequence of amino acids in PG and hence produce bactericidal effect [104]. Exogenous amino acids such as d-methionine, d-tryptophan, and d-phenylalanine, when incorporated into bacterial PG, replace l-alanine at position 1 and d-alanine at position 4 and 5 at a terminal position, results in bacterial killing in planktonic or suspended form [116]. Amino acid glycine when provided exogenously, replaces l-alanine at position 1 and d-alanine at positions 4 and 5 in PG, disrupting the natural sequence in it and results in bacterial growth inhibition [117].
Amino acids have been used (in vitro) in combination with antimicrobial drugs and have been shown to have successfully enhanced the efficacy of the drugs against planktonic bacteria [48]. For example, aspartic acid and glutamic acid have been used to improve the efficacy of antibiotic trimethoprim via amino acids-based trimethoprim salts formation [65]. The resultant salts have better solubilities, increased absorption, and hence more effectiveness even at lower concentrations [68].
Different free amino acids and peptides have been reported to exhibit antibacterial activity i.e., nisin, a polycyclic peptide consisted of different amino acids such as alanine, valine, serine lysine, etc., have been shown to have antimicrobial activity against both Gram-positive and Gram-negative bacteria [101,118,119].
Apart from the aforementioned antimicrobial roles of amino acids in planktonic bacterial control, d-forms of different amino acids have been reported to have antibiofilm activity as well, as discussed in the following section.
6. d-Amino Acids as Antibiofilm Agents, Adjuvants, and Potential Drugs Excipients
Free-living microbial cells attach to the host surface, initiate colonisation and secrete an extracellular polymeric substance called biofilm, composed of polysaccharides, proteins, and DNA (extracellular DNA) [72,120,121]. Biofilm makes it difficult to treat microbial infections as it protects the attached cells residing inside it and thereby offer resistance against antimicrobial drugs [122]. Once a biofilm reaches its maturity, it is dispersed and the microbial cells are released into the surrounding environment, ready to recolonise the host surface and repeat the process of biofilm formation [123], as illustrated in Figure 10.
Figure 10.

The different stages of microbial biofilm formation are as follows 1) attachment of cells to the surface (host or inanimate surfaces), 2) colonisation and biofilm formation, 3) biofilm maturation and 4) biofilm dispersal. Basic concept has been adopted from Di Luca, Maccari and Nifos, 2014 [124]. Figure 10 has been drawn using Paint 3D app.
Prevention and dispersal of microbial biofilm are of utmost importance in treating microbial infections and controlling antimicrobial resistance. Certain enzymes, called racemases facilitate the conversion of l-amino acids into their corresponding d-enantiomers through the process of racemisation [24,125]. These d-enantiomers or d-amino acids have been reported to have the ability to inhibit and disperse microbial biofilms [66]. A mixture of four amino acids, d-leucine, d-tryptophan, d-methionine, and d-tyrosine was able to disperse the microbial biofilm in Bacillus subtilis [63]. d-amino acids work via the mechanism of incorporation into the PG of the bacterial cell wall and disrupt the natural sequence of amino acids by replacing d-alanine in PG [103]. This mechanism leads to the release of amyloid fibers from bacterial PG and the dispersal of biofilm [63]. Amyloid fibers are protein fibers that are embedded in bacterial PG and biofilm at opposite ends [126].
d-amino acids were also found to have prevented biofilm formation in Staphylococcus aureus and Streptococcus mutans [127,128]. Microbial cells are connected to the biofilm via cellulose fibres, embedded into microbial peptidoglycan [129]. The incorporation of d-amino acids into peptidoglycan (during its biosynthesis) disrupts the sequence of existing amino acids and the microfibres-microbial cell linkage breaks up, hence dispersing the biofilm and releasing the sessile microbial cells [63,103].
Amino acids may have potential therapeutic applications as antibiofilm agents and drug excipients as the available literature mentions numerous research studies on the role of amino acids as potential antibiofilm agents and drug excipients. One such study found that the fungicidal and antibiofilm activity of antifungal drug amphotericin B was enhanced when combined (physically) with amino acid lysine, through a possible lysine-mediated generation of endogenous reactive oxygen species (ROS) [130]. The antibacterial and antibiofilm activity of known drugs such as clindamycin, cefazolin, oxacillin, rifampin, and vancomycin (for Gram-positive) and amikacin, colistin, ciprofloxacin, imipenem, and ceftazidime (for Gram-negative) has been reported to have enhanced when combined (physically) with d-amino acids [51].
Amino acids have also been used as adjuvants to enhance the efficacy of inactivated pdm H1N1 vaccine used in the treatment of viral influenza by promoting the cellular uptake of pdm H1N1 and activation of macrophages in the host environment [67,131]. Similarly, the known biocide, hydroxymethyl phosphonium sulphate (THPS) exhibited an enhanced biocidal activity when physically mixed with a mixture of d-tyrosine, d-leucin, d-tryptophan, and d-methionine, compared to the biocidal activity of the THPS alone [132]. One of the possible mechanisms of action for such amino acids-drugs combination is that d-amino acids disperse the microbial biofilm to release and expose the sessile cells (which are now in the planktonic state), thereby making it easier for the drug to reach and eliminate the microbial cells more effectively [133].
A second probable mechanism is the incorporation of D-amino acids into the microbial peptidoglycan that disrupts the sequence of existing amino acids and replace the terminal d-alanine (at fifth position) in the peptidoglycan, enhancing its sensitivity and susceptibility towards antimicrobial drugs [134]. Amino acids have also been used as formulation protectants in freeze-drying [135]. The development of vaccine formulation traditionally involves the use of sugars as cryoprotectants in freeze-drying [136]. However, recent work has reported the use of amino acids as cryoprotectants in freeze-drying, offering similar advantages with improved immune response [137].
Freeze-drying or the lyophilisation technique is employed in the formulation and development of vaccines, mainly to stabilise the vaccines against hydrolysis and physical degradation during storage [138]. However, this technique itself can damage the vaccine during lyophilisation/freeze-drying [139]. To avoid any degradation or damage to the vaccine, amino acids have been used as cryoprotectants. Amino acids being charged molecules, form a protective layer on the surface of liposome by their electrostatic interaction with phosphate head groups of the liposomal lipid molecules, thereby protect the formulation against ice-crystal damage [137].
Despite a variety of pharmaceutical and therapeutic applications, the concentration of amino acids should be carefully optimised in pharmaceutical formulations to avoid any undesired effects. For instance, an overconsumption of amino acids can lead to different undesired consequences such as depressed growth due to antagonism and even toxicity due to high plasma concentrations of the amino acids taken in excess [140].
7. Amino Acids as Solubility Enhancing Agents
Solubility is a phenomenon exhibited by any solid, liquid, or gaseous chemical substance with reference to its maximum amount dissolved in a given amount of solvent (solid, liquid, or gas phase) at a specific temperature to form a solution of uniform composition [141,142]. The solubility of chemical substances plays an important role in different scientific fields such as chemistry, biological science, food science, and pharmaceutical science [143].
As the discovery of new drugs continues, about 90% of these potential new drugs have been reported to have poor water-solubility [144]. Poorly water-soluble drugs have problematic absorption and low-bioavailability [142]. Different strategies such as nanotechnology, co-crystals, adsorption enhancers, and salt formation have been adopted to overcome the issue of poor solubility of drugs in aqueous media [145,146].
Amino acids have been reported to have enhanced the solubility of drugs through salt formation technique i.e., solubility of Indomethacin (a nonsteroidal anti-inflammatory drug) was enhanced through salt formation by using basic amino acids l-arginine and l-lysine as counterions [147,148]. ElShaer et al., [65] used acidic amino acids, aspartic acid, and glutamic acid as counterions to form amino acid-trimethoprim salts, enhancing water solubility of trimethoprim by 280 folds.
Similarly, the water solubility of insulin (an antidiabetic drug) and ciprofloxacin was improved by salt formation with cationic and anionic amino acids, respectively [68,149,150]. The increase in solubility of antimicrobial drugs results in improved bioavailability, better absorption and enhanced efficacy of the antimicrobial drugs i.e., the counterion-based salts of antimicrobial drugs are more effective at lower concentrations compared to their free forms [65,144].
Drugs solubility improvement by salt formation with acidic or basic salting agents as counterions, is regulated by parameters such as the acid-base dissociation constant (pKa), pH, and isoelectronic point. The acidity or basicity of a substance is expressed by its pKa value, the acid-base dissociation constant of that substance [151]. The pKa scale provides information regarding the ionizability of a chemical substance and also holds a significant pharmaceutical and chemical importance at an industrial level [152]. The pKa value and acid strength are inversely related i.e., strong acids have low pKa values and vice versa [153]. Moreover, salts that are formed from acidic drugs require counterions with a pKa < pKa of the drugs and salts that are formed from basic drugs require counterions with a pKa > pKa of the drugs [154].
Amino acids generally exhibit zwitterionic character in aqueous media, therefore, their acidic or basic characters can also be determined by measuring their isoelectronic point (pI). The isoelectronic point, pI of a substance is its pH at which it carries no charge at all i.e., the net charge on substance is equal to zero [155]. Zwitterions represent those compounds that possess both acidic and basic properties, with positive and negative charged species within the same molecule (dipolar) as shown in Figure 11, but the molecule as whole stays neutral [156,157].
Figure 11.
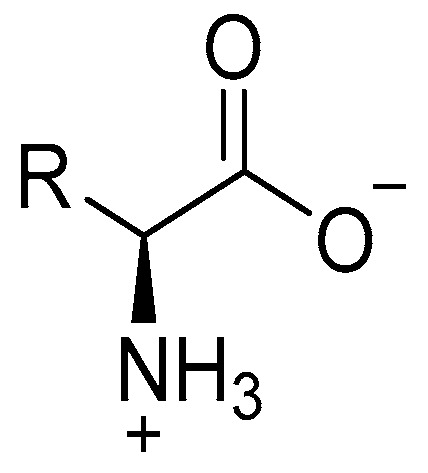
Zwitterionic structure of amino acids.
An amino acid with a pI less than seven (pI < 7) will exhibit acidic character and an amino acid with a pI greater than seven (pI > 7) will exhibit a basic character [158]. The pKa and pI values for different amino acids are listed in Table 2.
Table 2.
Enlists all 20 proteogenic amino acids. It highlights acidic, basic and neutral amino acids with their pKa and pI values. Data collected from Haynes, William M., 2014 [159].
| Name | Acidic | Basic | Neutral | pKa1 (α-Carboxyl Group) | PKa2 (α-Amino Group) | pKa3 (Side Chain) | pI (Isoelectronic Point) |
|---|---|---|---|---|---|---|---|
| Alanine | √ | 2.34 | 9.69 | 6.00 | |||
| Arginine | √ | 2.17 | 9.04 | 12.48 | 10.76 | ||
| Asparagine | √ | 2.02 | 8.80 | 5.41 | |||
| Aspartic acid | √ | 1.88 | 9.60 | 3.65 | 2.77 | ||
| Cysteine | √ | 1.96 | 10.28 | 8.18 | 5.07 | ||
| Glutamic acid | √ | 2.19 | 9.67 | 4.25 | 3.22 | ||
| Glutamine | √ | 2.17 | 9.13 | 5.65 | |||
| Glycine | √ | 2.34 | 9.60 | 5.97 | |||
| Histidine | √ | 1.82 | 9.17 | 6.00 | 7.95 | ||
| Isoleucine | √ | 2.36 | 9.60 | 6.02 | |||
| Leucine | √ | 2.36 | 9.60 | 5.98 | |||
| Lysine | √ | 2.18 | 8.95 | 10.53 | 9.74 | ||
| Methionine | √ | 2.28 | 9.21 | 5.74 | |||
| Phenylalanine | √ | 1.83 | 9.13 | 5.48 | |||
| Proline | √ | 1.99 | 10.60 | 6.30 | |||
| Serine | √ | 2.21 | 9.15 | 5.68 | |||
| Threonine | √ | 2.09 | 9.10 | 5.60 | |||
| Tryptophan | √ | 2.83 | 9.39 | 5.89 | |||
| Tyrosine | √ | 2.20 | 9.11 | 10.07 | 5.66 | ||
| Valine | √ | 2.32 | 9.62 | 5.96 |
Table 2 shows that the pKa values for α-carboxyl group fall in the range of 1.82–2.21 (acidic), while for α-amino group, the pKa values range from 8.95 to 9.67 (basic). The side chain pKa values of 3.65 for aspartic acid and 4.25 for glutamic acid are representative of their acidic behavior. Similarly, arginine and lysine side chains have higher pKa values i.e., 12.48 and 10.53 respectively, are more basic compared to Histidine with a side chain having a pKa value of 6.
Acidic amino acids i.e., aspartic acid and glutamic acid, have low (2.77 and 3.22) isoelectronic points (pI), compared to basic amino acids such as arginine with a pI value of 10.76, lysine 9.74 and histidine with a value of 7.95. Neutral amino acids have their pI ranging from 5.6 to 5.68.
The pKa and pI parameters are important in amino acid separation, drug solubility determination, predicting permeability of drugs across the microbial cell envelopes and therefore helpful in designing new antimicrobial drugs [64,160,161].
The side chain of an amino acid also determines if a particular amino acid is hydrophilic or hydrophobic, and whether an amino acid will exist as a charged or neutral molecule [162]. Hydrophilic amino acids are water-soluble and tend to exist as positively or negatively charged species due the presence of ionized or polar side chain in their molecules, while the hydrophobic amino acids do not have such side chains and are therefore either insoluble or barely soluble in water [163]. Hydrophobic and hydrophilic amino acids have been listed in Table 3.
Table 3.
List of different hydrophilic and hydrophobic amino acids with corresponding logP values. The lipophilicity of these amino acids, expressed as their logP, ranges from −4.3 to −1.38, where asparagine with logP value of −4.3 being the least lipophilic and phenylalanine with a logP of −1.38 being the most lipophilic or hydrophobic amino acid. It also shows that hydrophilic amino acids are either charged, uncharged or polar molecules, while hydrophobic amino acids are mostly uncharged and nonpolar.
| Hydrophilic Amino Acids | Charged (+/−/Uncharged) | logP | Hydrophobic Amino Acids | Charged (+/−/Uncharged) | logP |
|---|---|---|---|---|---|
| Asparagine | Uncharged | −4.3 | Arginine | + | −4.2 |
| Aspartic acid | − | −3.89 | Cysteine | Uncharged | −2.49 |
| Glutamine | Uncharged | −3.64 | Isoleucine | Uncharged | −1.7 |
| Glutamic acid | − | −3.69 | Glycine | Uncharged | −3.4 |
| Histidine | + | −3.32 | Phenylalanine | Uncharged | −1.38 |
| Lysine | + | −3.05 | Serine | Uncharged | −3.07 |
| Threonine | Uncharged | −3.5 | Leucine | Uncharged | −1.6 |
| Tyrosine | Uncharged | −2.4 | Valine | Uncharged | −2.26 |
The addition of acidic counterions to a poorly soluble basic drug solution usually lowers the pH of the drug solution with salt formation and thereby improves the solubility of the drug, whereas the addition of basic counterions to poorly soluble acidic drugs increase the solution pH and improve drug solubility [65,152,164]. Drugs usually have better solubilities at a pH that is either above or below their pI i.e., at a pH = pI the total charge on the individual solute particles (both counterion and the drug molecules) = zero, a point at which the drug precipitates out in its free from [165,166].
Selection of a suitable amino acid as a counterion for a poorly soluble drug involves a thorough assessment of certain factors such as screening different amino acids for concentration optimisation and selection of oppositely charged amino acids [68,148]. Concentration of amino acid as a counterion affects drug solubility as pH of the drug solution varies with varying amino acid concentration. For instance, amino acids arginine and lysin solutions having concentrations > 100 µg/mL, were able to change the pH of insulin in Hank’s balanced salt solution (HBSS) by almost 2 units i.e., from 7.4 to 9.2, improving insulin’s solubility [149].
Similarly, the overall charge of an amino acid has to be assessed to determine if it can be a suitable counterion for a particular acidic or basic ionisable drug. Amino acids that have overall positive or negative charges are all of hydrophilic nature, with an exception of hydrophobic arginine [162]. Amino acid-based salt formation of a drug requires the selection of appositely charged amino acids to balance the total charge on the resultant salt [167]. These appositely charged counterions interact with acidic or basic ionised drugs in their solutions via strong electrostatic forces of attraction to form stable and neutral salts [154]. The resultant ionic salts have better solubility, improved absorption, and hence enhanced efficacy [65,168].
Amino acids as salt formers also impact the membrane permeability of the resultant salts and thereby affect their overall efficacy. Salt formation often increases the stability and alters lipophilicity of the drug and hence eases the drug permeation across the lipophilic cellular barriers [169]. However, amino acids can be used to improve membrane permeability of drugs by increasing their hydrophilicity via salt formation. For example, one such study has reported aspartic acid to have increased the membrane permeability of antimicrobial drug ciprofloxacin, by enhancing its water solubility through hydrophilic salt formation [68].
Apart from their aforementioned applications, d-forms of amino acids can also be used as sweeteners for taste masking. Since l-amino acids are usually bitter or less sweet compared to their respective d-forms, their racemisation into d-amino acids results in the formation of sweet amino acids or in converting less sweet amino acids into even sweeter ones [170].
8. Expert Opinion
Amino acids are naturally occurring substances which play a key role in all living organisms. There are numerous phenotypic studies evaluating the effect of amino acids on growth, metabolism, and control of microorganisms. However, there is a lack of data on the exact mechanism of action of amino acids in various scenarios such as biofilm disruption, synergy with antibiotics, and microbial regulation.
Amino acids present an interesting biological paradox with huge potential in the process of drug development. On one hand, they are required for growth and metabolism in microorganisms, and on the other hand, careful optimisation of composition and concentration can produce antibiofilm and antibacterial effects. The above attribute allows them to be used as drug adjuvants/excipients which ultimately improves the efficacy of drugs. This not only results in better clinical outcomes but also provides a tool to fight antimicrobial resistance which has become a global threat. Improvement in efficacy is achieved by increasing the solubility of poorly soluble drugs and ultimately increasing their bioavailability. As amino acids, when used as excipients/adjuvants, are non-toxic to humans, the development of resulting product/formulation would incur lower costs when compared to developing a new active pharmaceutical ingredient/antibiotic. In addition, understanding the mechanism of amino acid-mediated processes such as disruption of microbial biofilms and their role in enhancing antibiotic activity will enable the development of simple, practical, and cost-effective strategies to reduce AMR. By disrupting biofilms, amino acids will allow increased penetration of antibiotics into the deeper layers of the biofilm, thereby lowering the concentration of the antibiotic required to treat biofilm-related infections. This will result in broadening the use of existing antibiotics to which microorganisms have become resistant thus aiding to repurpose existing antibiotics for a wide range of microbial infections. Impact of such combinations in vivo will have to be established to determine the applicability and safety of amino acid-drug combinations. One of the challenges in this field revealed by reviewing existing literature on the dispersal activity of d-amino acids on biofilms of Bacillus subtilis, Staphylococcus aureus, Streptococcus mutans, and Pseudomonas aeruginosa is that microorganisms vary in their response to a given amino acid. Therefore, universal mechanism by which amino acids disrupt/disperse biofilms is implausible.
Amino acids have generated considerable interest as biofilm disrupting agents and this strategy has applications in treating biofilm-related infections such as those caused in cystic fibrosis patients and medical device-related infections. The impact of amino acids on biofilms can also be exploited in the food industry and the environment where biofilms are an issue causing spoilage and leading to an increased threat of infection. Thus, amino acids are versatile biomolecules that have a wide range of applications in the pharmaceutical, food, and the industrial environment.
Author Contributions
Conceptualization, A.R.M. & A.R.; methodology, A.R. & M.I.; software ChemDraw, M.I.; writing—original draft preparation, M.I.; writing—review and editing, A.R.M., A.R. & N.K.; supervision, A.R. & N.K.; project administration, A.R. & N.K.; funding acquisition, A.R. & N.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Faculty of Science and Engineering, University of Wolverhampton (UoW).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- 1.Moini J. Epidemiology of Diet and Diabetes Mellitus. In: Moini J., editor. Epidemiology of Diabetes. 1st ed. Elsevier; Amsterdam, The Netherlands: 2019. pp. 57–73. [Google Scholar]
- 2.Bischoff R., Schlüter H. Amino acids: Chemistry, functionality and selected non-enzymatic post-translational modifications. J. Proteom. 2012;75:2275–2296. doi: 10.1016/j.jprot.2012.01.041. [DOI] [PubMed] [Google Scholar]
- 3.Sparkman O.D., Penton Z.E., Kitson F.G. Amino Acids. In: Sparkman O.D., Penton Z.E., Kitson F.G., editors. Gas Chromatography and Mass Spectrometry. Academic Press; Amsterdam, The Netherlands: 2011. pp. 265–271. [Google Scholar]
- 4.Ouellette R.J., Rawn J.D. Amino acids, peptides, and proteins. In: Ouellette R.J., Rawn J.D., editors. Organic Chemistry Study Guide. Elsevier; Boston, MA, USA: 2015. pp. 569–586. [Google Scholar]
- 5.Makishima A. Fundamental knowledges and techniques in biochemistry. In: Makishima A., editor. Biochemistry for Materials. Elsevier Science; Amsterdam, The Netherlands: 2019. pp. 35–51. [Google Scholar]
- 6.Parker J. Peptide Bond. In: Brenner S., Miller J.H., editors. Encyclopedia of Genetics. 1st ed. Academic Press; New York, NY, USA: 2001. pp. 1429–1430. [Google Scholar]
- 7.Singh P., Manda S.L.K., Samanta K., Panda G. α-Amino acids with electrically charged and polar uncharged side chains as chiral synthon: Application to the synthesis of bioactive alkaloids (1996–Dec 2013) Tetrahedron. 2017;73:1911–2008. doi: 10.1016/j.tet.2017.02.029. [DOI] [Google Scholar]
- 8.Pizzarello S. Non-Protein amino acids. In: Gargaud M., Amils R., Quintanilla J.C., Cleaves H.J., Irvine W.M., Pinti D.L., Viso M., editors. Encyclopedia of Astrobiology. 1st ed. Springer; Berlin/Heidelberg, Germany: 2011. pp. 1128–1132. [Google Scholar]
- 9.Yamane H., Konno K., Sabelis M., Takabayashi J., Sassa T., Oikawa H. Chemical defence and toxins of plants. In: Liu H., Mander L., editors. Comprehensive Natural Products II. 1st ed. Elsevier; Oxford, UK: 2010. pp. 339–385. [Google Scholar]
- 10.Ribas de Pouplana L. Not an inside job: Non-coded amino acids compromise the genetic code. EMBO J. 2014;33:1619–1620. doi: 10.15252/embj.201489108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Huang T., Jander G., de Vos M. Non-protein amino acids in plant defense against insect herbivores: Representative cases and opportunities for further functional analysis. Phytochemistry. 2011;72:1531–1537. doi: 10.1016/j.phytochem.2011.03.019. [DOI] [PubMed] [Google Scholar]
- 12.Rodgers K.J., Samardzic K., Main B.J. Toxic nonprotein amino acids. In: Carlini C.R., Ligabue-Braun R., editors. Plant Toxins. Springer; Dordrecht, The Netherlands: 2017. pp. 263–285. [Google Scholar]
- 13.Song Y., Zhou H., Vo M.N., Shi Y., Nawaz M.H., Vargas-Rodriguez O., Diedrich J.K., Yates J.R., Kishi S., Musier-Forsyth K., et al. Double mimicry evades tRNA synthetase editing by toxic vegetable-sourced non-proteinogenic amino acid. Nat. Commun. 2017;8:2281. doi: 10.1038/s41467-017-02201-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Rubenstein E., Zhou H., Krasinska K.M., Chien A., Becker C.H. Azetidine-2-carboxylic acid in garden beets (Beta vulgaris) Phytochemistry. 2006;67:898–903. doi: 10.1016/j.phytochem.2006.01.028. [DOI] [PubMed] [Google Scholar]
- 15.Wu G. Amino acids: Metabolism, functions, and nutrition. Amino Acids. 2009;37:1–17. doi: 10.1007/s00726-009-0269-0. [DOI] [PubMed] [Google Scholar]
- 16.Kevin A., Indira R., Taralyn T. Structure & Function—Amino Acids—Biology LibreTexts. [(accessed on 12 November 2019)]; Available online: https://bio.libretexts.org/Bookshelves/Biochemistry/Book:_Biochemistry_Free_For_All_(Ahern,_Rajagopal,_and_Tan)/2:_Structure_and_Function/2.2:_Structure_and_Function_-_Amino_Acids.
- 17.Litwack G. Human Biochemistry. Academic Press; Boston, MA, USA: 2018. Proteins; pp. 63–94. [Google Scholar]
- 18.Engelking L.R. Amino acid modifications. In: Engelking L.R., editor. Textbook of Veterinary Physiological Chemistry. 3rd ed. Academic Press; Boston, MA, USA: 2015. pp. 12–17. [Google Scholar]
- 19.Bo-Hyun C., Coloff J.L. The diverse functions of non-essential amino acids in cancer. Cancers. 2019;11:675. doi: 10.3390/cancers11050675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Naef R. A generally applicable computer algorithm based on the group additivity method for the calculation of seven molecular descriptors: Heat of combustion, LogP O/W, LogS, refractivity, polarizability, toxicity and LogBB of organic compounds; scope and limits of applicability. Molecules. 2015;20:18279–18351. doi: 10.3390/molecules201018279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.National Center for Biotechnology Information PubChem Database Glutamic Acid | C5H9NO4—PubChem. [(accessed on 27 January 2020)]; Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Glutamic-acid.
- 22.Burton A.S., Berger E.L. Berger Insights into Abiotically-Generated Amino Acid Enantiomeric Excesses Found in Meteorites. [(accessed on 12 November 2019)]; doi: 10.3390/life8020014. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6027462/ [DOI] [PMC free article] [PubMed]
- 23.Fujii N., Kaji Y., Fujii N. d-Amino acids in aged proteins: Analysis and biological relevance. J. Chromatogr. B. 2011;879:3141–3147. doi: 10.1016/j.jchromb.2011.05.051. [DOI] [PubMed] [Google Scholar]
- 24.Zhang G., Sun H.J. Racemization in reverse: Evidence that D-Amino acid toxicity on earth is controlled by bacteria with racemases. PLoS ONE. 2014;9:e92101. doi: 10.1371/journal.pone.0092101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Nguyen L.A., He H., Pham-Huy C. Chiral drugs: An overview. Int. J. Biomed. Sci. 2006;2:85–100. [PMC free article] [PubMed] [Google Scholar]
- 26.Jakubowski H. Amino Acid Stereochemistry—Biology LibreTexts. [(accessed on 23 April 2020)]; Available online: https://bio.libretexts.org/Bookshelves/Biochemistry/Book:_Biochemistry_Online_(Jakubowski)/02:_PROTEIN_STRUCTURE/2A:_Amino_Acids/A02._Amino_Acid_Stereochemistry.
- 27.Patzold R., Bruckner H. Chiral separation of amino acids by gas chromatography. In: MolnÃir-Perl I., editor. Journal of Chromatography Library. Volume 70. Elsevier; Amsterdam, The Netherlands: 2005. pp. 98–118. [Google Scholar]
- 28.Cava F., Lam H., de Pedro M.A., Waldor M.K. Emerging knowledge of regulatory roles of D-Amino acids in Bacteria. Cell. Mol. Life Sci. 2010;68:817–831. doi: 10.1007/s00018-010-0571-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kiriyama Y., Nochi H. D-Amino acids in the nervous and endocrine systems. Scientifica. 2016;216:6494621. doi: 10.1155/2016/6494621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Pawelek P.D., Cheah J., Coulombe R., Macheroux P., Ghisla S., Vrielink A. The structure of L-amino acid oxidase reveals the substrate trajectory into an enantiomerically conserved active site. EMBO J. 2000;19:4204–4215. doi: 10.1093/emboj/19.16.4204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Marcone G.L., Binda E., Rosini E., Abbondi M., Pollegioni L. Antibacterial Properties of D-Amino acid oxidase: Impact on the food industry. Front. Microbiol. 2019;10:2786. doi: 10.3389/fmicb.2019.02786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kitani Y., Kikuchi N., Zhang G., Ishizaki S., Shimakura K., Shiomi K., Nagashima Y. Antibacterial action of L-amino acid oxidase from the skin mucus of rockfish Sebastes schlegelii. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008;149:394–400. doi: 10.1016/j.cbpb.2007.10.013. [DOI] [PubMed] [Google Scholar]
- 33.Zhang H., Yang Q., Sun M., Teng M., Niu L. Hydrogen peroxide produced by two amino acid oxidases mediates antibacterial actions. J. Microbiol. 2004;42:336–339. [PubMed] [Google Scholar]
- 34.Berg J.M., Tymoczko J., Stryer L., editors. The Biosynthesis of Amino Acids. Volume 2019 W. H. Freeman; New York, NY, USA: 2002. [Google Scholar]
- 35.Nakayama K., Araki K., Kase H. Microbial production of essential amino acid with Corynebacterium glutamicum mutants. Adv. Exp. Med. Biol. 1978;105:649–661. doi: 10.1007/978-1-4684-3366-1_31. [DOI] [PubMed] [Google Scholar]
- 36.Hernández-Montes G., Díaz-Mejía J.J., Pérez-Rueda E., Segovia L. The hidden universal distribution of amino acid biosynthetic networks: A genomic perspective on their origins and evolution. Genome Biol. 2008;9:R95. doi: 10.1186/gb-2008-9-6-r95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Akashi H., Gojobori T. Metabolic efficiency and amino acid composition in the proteomes of Escherichia coli and Bacillus subtilis. Proc. Natl. Acad. Sci. USA. 2002;99:3695. doi: 10.1073/pnas.062526999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Amorim Franco T.M., Blanchard J.S. Bacterial branched-chain amino acid biosynthesis: Structures, mechanisms, and drugability. Biochemistry. 2017;56:5849–5865. doi: 10.1021/acs.biochem.7b00849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Wu N., Yang M., Gaur U., Xu H., Yao Y., Li D. Alpha-Ketoglutarate: Physiological functions and applications. Biomol. Ther. 2016;24:1–8. doi: 10.4062/biomolther.2015.078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Pelley J.W. Citric acid cycle, electron transport chain, and oxidative phosphorylation. In: Pelley J.W., editor. Elsevier’s Integrated Biochemistry. Mosby; Philadelphia, PA, USA: 2007. pp. 55–63. [Google Scholar]
- 41.Jastrzebowska K., Gabriel I. Inhibitors of amino acids biosynthesis as antifungal agents. Amino Acids. 2015;47:227–249. doi: 10.1007/s00726-014-1873-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Fazius F., Shelest E., Gebhardt P., Brock M. The fungal α-aminoadipate pathway for lysine biosynthesis requires two enzymes of the aconitase family for the isomerization of homocitrate to homoisocitrate. Mol. Microbiol. 2012;86:1508–1530. doi: 10.1111/mmi.12076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Miyazaki J., Kobashi N., Nishiyama M., Yamane H. Functional and evolutionary relationship between arginine biosynthesis and prokaryotic lysine biosynthesis through. α-Aminoadipate. J. Bacteriol. 2001;183:5067. doi: 10.1128/JB.183.17.5067-5073.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Garbe E.V. Role of amino acid metabolism in the virulence of human pathogenic fungi. Curr. Clin. Microbiol. Rep. 2019;6:108–119. doi: 10.1007/s40588-019-00124-5. [DOI] [Google Scholar]
- 45.Zhang W., Du G., Zhou J., Chen J. Regulation of Amino Acid, Nucleotide, and Phosphate Metabolism in Saccharomyces Cerevisiae. [(accessed on 25 December 2019)]; Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3296254/?report=reader.
- 46.Price M.N., Zane G.M., Kuehl J.V., Melnyk R.A., Wall J.D., Deutschbauer A.M., Arkin A.P. Filling gaps in bacterial amino acid biosynthesis pathways with high-throughput genetics. PLoS Genet. 2018;14:1007147. doi: 10.1371/journal.pgen.1007147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Yamamoto K., Tsuchisaka A., Yukawa H. Branched-Chain amino acids. Adv. Biochem. Eng. Biotechnol. 2017;159:103–128. doi: 10.1007/10_2016_28. [DOI] [PubMed] [Google Scholar]
- 48.Bringel F., Hubert J. Extent of genetic lesions of the arginine and pyrimidine biosynthetic pathways in Lactobacillus plantarum, L. paraplantarum, L. pentosus, and L. casei: Prevalence of CO(2)-dependent auxotrophs and characterization of deficient arg genes in L. plantarum. Appl. Environ. Microbiol. 2003;69:2674–2683. doi: 10.1128/AEM.69.5.2674-2683.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Radmacher E., Vaitsikova A., Burger U., Krumbach K., Sahm H., Eggeling L. Linking Central metabolism with increased pathway flux: L-Valine accumulation by Corynebacterium glutamicum. Appl. Environ. Microbiol. 2002;68:2246. doi: 10.1128/AEM.68.5.2246-2250.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.D’Este M., Alvarado-Morales M., Angelidaki I. Amino acids production focusing on fermentation technologies—A review. Biotechnol. Adv. 2018;36:14–25. doi: 10.1016/j.biotechadv.2017.09.001. [DOI] [PubMed] [Google Scholar]
- 51.Sanchez S., Rodriguez-Sanoja R., Ramos A., Demain A.L. Our microbes not only produce antibiotics, they also overproduce amino acids. J. Antibiot. 2017;71:26–36. doi: 10.1038/ja.2017.142. [DOI] [PubMed] [Google Scholar]
- 52.Ikeda M. Amino acid production processes. Adv. Biochem. Eng. Biotechnol. 2003;79:1–35. doi: 10.1007/3-540-45989-8_1. [DOI] [PubMed] [Google Scholar]
- 53.Hermann T. Industrial production of amino acids by coryneform bacteria. J. Biotechnol. 2003;104:155–172. doi: 10.1016/S0168-1656(03)00149-4. [DOI] [PubMed] [Google Scholar]
- 54.Sugden R., Kelly R., Davies S. Combatting antimicrobial resistance globally. Nat. Microbiol. 2016;1:1–2. doi: 10.1038/nmicrobiol.2016.187. [DOI] [PubMed] [Google Scholar]
- 55.World Health Organisation New Report Calls for Urgent Action to Avert Antimicrobial Resistance Crisis. [(accessed on 20 May 2020)]; Available online: https://www.who.int/news-room/detail/29–04–2019-new-report-calls-for-urgent-action-to-avert-antimicrobial-resistance-crisis.
- 56.Munita J.M., Arias C.A. Mechanisms of antibiotic resistance. Microbiol. Spectr. 2016;4 doi: 10.1128/microbiolspec.VMBF-0016-2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kostyanev T., Can F. The global crisis of antimicrobial resistance. In: Pulcini C., Ergönül Ö., Can F., Beović B., editors. Antimicrobial Stewardship. Academic Press; Cambridge, MA, USA: 2017. pp. 3–12. [Google Scholar]
- 58.Hasdemir U. The role of cell wall organization and active efflux pump systems in multidrug resistance of bacteria. Mikrobiyol. Bul. 2007;41:309–327. [PubMed] [Google Scholar]
- 59.Nguyen Van J.C., Gutmann L. Resistance to antibiotics caused by decrease of the permeability in gram-negative bacteria. Presse Med. 1994;23:522, 527–531. [PubMed] [Google Scholar]
- 60.Egorov A.M., Ulyashova M.M., Rubtsova M.Y. Bacterial enzymes and antibiotic resistance. Acta Nat. 2018;10:33. doi: 10.32607/20758251-2018-10-4-33-48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Wright G.D. Bacterial resistance to antibiotics: Enzymatic degradation and modification. Adv. Drug Deliv. Rev. 2005;57:1451–1470. doi: 10.1016/j.addr.2005.04.002. [DOI] [PubMed] [Google Scholar]
- 62.Sanchez C.J., Jr., Akers K.S., Romano D.R., Woodbury R.L., Hardy S.K., Murray C.K., Wenke J.C. D-amino acids enhance the activity of antimicrobials against biofilms of clinical wound isolates of Staphylococcus aureus and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2014;58:4353–4361. doi: 10.1128/AAC.02468-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Kolodkin-Gal I., Romero D., Cao S., Clardy J., Kolter R., Losick R. D-amino acids trigger biofilm disassembly. Science. 2010;328:627–629. doi: 10.1126/science.1188628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Pfalzgraff A., Brandenburg K., Weindl G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018;9:281. doi: 10.3389/fphar.2018.00281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Elshaer A., Hanson P., Worthington T., Lambert P., Mohammed A.R. Preparation and characterization of amino acids-based trimethoprim salts. Pharmaceutics. 2012;4:179–196. doi: 10.3390/pharmaceutics4010179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Ampornaramveth R.S., Akeatichod N., Lertnukkhid J., Songsang N. Application of D-Amino acids as biofilm dispersing agent in dental unit waterlines. Int. J. Dent. 2018;2018:9413925. doi: 10.1155/2018/9413925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Lim J.W., Na W., Kim H.O., Yeom M., Park G., Kang A., Chun H., Park C., Oh S., Le V.P., et al. Cationic Poly(Amino Acid) vaccine adjuvant for promoting both cell-mediated and humoral immunity against influenza virus. Adv. Healthc. Mater. 2019;8:1800953. doi: 10.1002/adhm.201800953. [DOI] [PubMed] [Google Scholar]
- 68.ElShaer A., Ouyang D., Hanson P., Mohammed A.R. Preparation and evaluation of amino acid based salt forms of model zwitterionic drug ciprofloxacin. J. Pharm. Drug. Deliv. Res. 2013;2:1–10. [Google Scholar]
- 69.Kaiko G.E., Stappenbeck T.S. Host-microbe interactions shaping the gastrointestinal environment. Trends Immunol. 2014;35:538–548. doi: 10.1016/j.it.2014.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Bahia D., Satoskar A.R., Dussurget O. Editorial: Cell signaling in host-pathogen interactions: The host point of view. Front. Immunol. 2018;9:221. doi: 10.3389/fimmu.2018.00221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ren W., Rajendran R., Zhao Y., Tan B., Wu G., Bazer F.W., Zhu G., Peng Y., Huang X., Deng J., et al. Amino acids as mediators of metabolic cross talk between host and pathogen. Front. Immunol. 2018;9:319. doi: 10.3389/fimmu.2018.00319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Aliashkevich A., Alvarez L., Cava F. New insights into the mechanisms and biological roles of D-Amino acids in complex Eco-Systems. Front. Microbiol. 2018;9:683. doi: 10.3389/fmicb.2018.00683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Li H., Anuwongcharoen N., Malik A.A., Prachayasittikul V., Wikberg J.E., Nantasenamat C. Roles of d-Amino acids on the bioactivity of host defense peptides. Int. J. Mol. Sci. 2016;17:1023. doi: 10.3390/ijms17071023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Wang B., Yao M., Lv L., Ling Z., Li L. The human microbiota in health and disease. Engineering. 2017;3:71–82. doi: 10.1016/J.ENG.2017.01.008. [DOI] [Google Scholar]
- 75.Ursell L.K., Metcalf J.L., Parfrey L.W., Knight R. Defining the human microbiome. Nutr. Rev. 2012;70:38–44. doi: 10.1111/j.1753-4887.2012.00493.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Turnbaugh P.J., Ley R.E., Hamady M., Fraser-Liggett C.M., Knight R., Gordon J.I. The human microbiome project. Nature. 2007;449:804–810. doi: 10.1038/nature06244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Thursby E., Juge N. Introduction to the human gut microbiota. Biochem. J. 2017;474:1823–1836. doi: 10.1042/BCJ20160510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.O’Hara A.M., Shanahan F. The gut flora as a forgotten organ. EMBO Rep. 2006;7:688–693. doi: 10.1038/sj.embor.7400731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Neis E.P., Dejong C.H., Rensen S.S. The role of microbial amino acid metabolism in host metabolism. Nutrients. 2015;7:2930–2946. doi: 10.3390/nu7042930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Baothman O.A., Zamzami M.A., Taher I., Abubaker J., Abu-Farha M. The role of Gut Microbiota in the development of obesity and Diabetes. Lipids Health Dis. 2016;15:108. doi: 10.1186/s12944-016-0278-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Bauer M., Knebel J., Lechner M., Pickl P., Frey E. Ecological feedback in quorum-sensing microbial populations can induce heterogeneous production of autoinducers. eLife. 2017;6:25773. doi: 10.7554/eLife.25773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Fatimah N., Pande G.S.J., Natrah F.M.I., Meritha W.W., Widanarni W., Sucipto A., Ekasari J. The role of microbial quorum sensing on the characteristics and functionality of bioflocs in aquaculture systems. Aquaculture. 2019;504:420–426. doi: 10.1016/j.aquaculture.2019.02.022. [DOI] [Google Scholar]
- 83.Sturme M.H., Kleerebezem M., Nakayama J., Akkermans A.D., Vaugha E.E., de Vos W.M. Cell to cell communication by autoinducing peptides in gram-positive bacteria. Antonie Van Leeuwenhoek. 2002;81:233–243. doi: 10.1023/A:1020522919555. [DOI] [PubMed] [Google Scholar]
- 84.Reading N.C., Sperandio V. Quorum sensing: The many languages of bacteria. FEMS Microbiol. Lett. 2006;254:1–11. doi: 10.1111/j.1574-6968.2005.00001.x. [DOI] [PubMed] [Google Scholar]
- 85.Rajput A., Gupta A.K., Kumar M. Prediction and analysis of quorum sensing peptides based on sequence features. PLoS ONE. 2015;10:e0120066. doi: 10.1371/journal.pone.0120066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Jiang Q., Chen J., Yang C., Yin Y., Yao K. Quorum sensing: A prospective therapeutic target for bacterial diseases. BioMed Res. Int. 2019;2019:2015978. doi: 10.1155/2019/2015978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Schaible U.E., Kaufmann S.H.E. Malnutrition and infection: Complex mechanisms and global impacts (research in translation) PLoS Med. 2007;4:115. doi: 10.1371/journal.pmed.0040115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Kurpad A.V. The requirements of protein & amino acid during acute & chronic infections. Indian J. Med. Res. 2006;124:129–148. [PubMed] [Google Scholar]
- 89.Shrestha R., Sorg J.A. Hierarchical recognition of amino acid co-germinants during Clostridioides difficile spore germination. Anaerobe. 2018;49:41–47. doi: 10.1016/j.anaerobe.2017.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Pollegioni L., Piubelli L., Sacchi S., Pilone M.S., Molla G. Physiological functions of D-amino acid oxidases: From yeast to humans. Cell. Mol. Life Sci. 2007;64:1373–1394. doi: 10.1007/s00018-007-6558-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Eissa M.E., Abd El Naby M., Beshir M.M. Bacterial vs. fungal spore resistance to peroxygen biocide on inanimate surfaces. Bull. Fac. Pharm. Cairo Univ. 2014;52:219–224. doi: 10.1016/j.bfopcu.2014.06.003. [DOI] [Google Scholar]
- 92.Swick M.C., Koehler T.M., Driks A. Surviving between hosts: Sporulation and transmission. Microbiol. Spectr. 2016;4 doi: 10.1128/microbiolspec.VMBF-0029-2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Zhu D., Sorg J.A., Sun X. Biology: Sporulation, germination, and corresponding therapies for infection. Front. Cell. Infect. Microbiol. 2018;8:29. doi: 10.3389/fcimb.2018.00029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Vylkova S., Carman A.J., Danhof H.A., Collette J.R., Zhou H., Lorenz M.C. The fungal pathogen Candida albicans autoinduces hyphal morphogenesis by raising extracellular pH. mBio. 2011;2:e00055-11. doi: 10.1128/mBio.00055-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Soriani M., Telford J.L. Relevance of pili in pathogenic streptococci pathogenesis and vaccine development. Future Microbiol. 2010;5:735–747. doi: 10.2217/fmb.10.37. [DOI] [PubMed] [Google Scholar]
- 96.Mandlik A., Swierczynski A., Das A., Ton-That H. Pili in Gram-positive bacteria: Assembly, involvement in colonization and biofilm development. Trends Microbiol. 2008;16:33–40. doi: 10.1016/j.tim.2007.10.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Padhi A., Sengupta M., Sengupta S., Roehm K.H., Sonawane A. Antimicrobial peptides and proteins in mycobacterial therapy: Current status and future prospects. Tuberculosis. 2014;94:363–373. doi: 10.1016/j.tube.2014.03.011. [DOI] [PubMed] [Google Scholar]
- 98.Carvajal-Rondanelli P., Arostica M., Alvarez C.A., Ojeda C., Albericio F., Aguilar L.F., Marshall S.H., Guzman F. Understanding the antimicrobial properties/activity of an 11-residue Lys homopeptide by alanine and proline scan. Amino Acids. 2018;50:557–568. doi: 10.1007/s00726-018-2542-6. [DOI] [PubMed] [Google Scholar]
- 99.She P., Chen L., Liu H., Zou Y., Luo Z., Koronfel A., Wu Y. The effects of D-Tyrosine combined with amikacin on the biofilms of Pseudomonas aeruginosa. Microb. Pathog. 2015;86:38–44. doi: 10.1016/j.micpath.2015.07.009. [DOI] [PubMed] [Google Scholar]
- 100.Yehuda A., Slamti L., Malach E., Lereclus D., Hayouka Z. Elucidating the hot spot residues of quorum sensing peptidic autoinducer PapR by multiple amino acid replacements. Front. Microbiol. 2019;10:1246. doi: 10.3389/fmicb.2019.01246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Santos J.C.P., Sousa R.C.S., Otoni C.G., Moraes A.R.F., Souza V.G.L., Medeiros E.A.A., Espitia P.J.P., Pires A.C.S., Coimbra J.S.R., Soares N.F.F. Nisin and other antimicrobial peptides: Production, mechanisms of action, and application in active food packaging. Innov. Food Sci. Emerg. Technol. 2018;48:179–194. doi: 10.1016/j.ifset.2018.06.008. [DOI] [Google Scholar]
- 102.Foster S.J., Popham D.L. Bacillus Subtilis and Its Closest Relatives. American Society of Microbiology; Washington, DC, USA: 2002. Structure and synthesis of cell wall, spore cortex, teichoic acids, S-Layers, and Capsules. [Google Scholar]
- 103.Lam H., Oh D., Cava F., Takacs C.N., Clardy J., de Pedro M.A., Waldor M.K. D-amino acids govern stationary phase cell wall remodeling in bacteria. Science. 2009;325:1552. doi: 10.1126/science.1178123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Typas A., Banzhaf M., Gross C.A., Vollmer W. From the regulation of peptidoglycan synthesis to bacterial growth and morphology. Nat. Rev. Microbiol. 2011;10:123–136. doi: 10.1038/nrmicro2677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Rani C., Khan I.A. UDP-GlcNAc pathway: Potential target for inhibitor discovery against M. tuberculosis. Eur. J. Pharm. Sci. 2016;83:62–70. doi: 10.1016/j.ejps.2015.12.013. [DOI] [PubMed] [Google Scholar]
- 106.Lovering A.L., Safadi S.S., Strynadka N.C. Structural perspective of peptidoglycan biosynthesis and assembly. Annu. Rev. Biochem. 2012;81:451–478. doi: 10.1146/annurev-biochem-061809-112742. [DOI] [PubMed] [Google Scholar]
- 107.Kaiser G. The Peptidoglycan Cell Wall—Biology LibreTexts. [(accessed on 19 April 2020)]; Available online: https://bio.libretexts.org/Bookshelves/Microbiology/Book:_Microbiology_(Kaiser)/Unit_1:_Introduction_to_Microbiology_and_Prokaryotic_Cell_Anatomy/2:_The_Prokaryotic_Cell_-_Bacteria/2.3:_The_Peptidoglycan_Cell_Wall.
- 108.Liu Y., Breukink E. The membrane steps of bacterial cell wall synthesis as antibiotic targets. Antibiotics. 2016;5:28. doi: 10.3390/antibiotics5030028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Rohs P.D.A., Buss J., Sim S.I., Squyres G.R., Srisuknimit V., Smith M., Cho H., Sjodt M., Kruse A.C., Garner E.C., et al. A central role for PBP2 in the activation of peptidoglycan polymerization by the bacterial cell elongation machinery. PLoS Genet. 2018;14:1007726. doi: 10.1371/journal.pgen.1007726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Popham D.L. Visualizing the production and arrangement of peptidoglycan in Gram-positive cells. Mol. Microbiol. 2013;88:645–649. doi: 10.1111/mmi.12212. [DOI] [PubMed] [Google Scholar]
- 111.Irazoki O., Hernandez S.B., Cava F. Peptidoglycan muropeptides: Release, perception, and functions as signaling molecules. Front. Microbiol. 2019;10:500. doi: 10.3389/fmicb.2019.00500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Que Y., Moreillon P. Staphylococcus aureus (Including Staphylococcal Toxic Shock Syndrome) In: Bennett J.E., Dolin R., Blaser M.J., editors. Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases. 8th ed. Content Repository Only; Philadelphia, PA, USA: 2015. pp. 2237–2271. [Google Scholar]
- 113.Allison D.G., Lambert P.A. Modes of action of antibacterial agents. In: Tang Y.-W., Sussman M., Liu D., Poxton I., Schwartzman J., editors. Molecular Medical Microbiology. 2nd ed. Academic Press; Boston, MA, USA: 2015. p. 583. [Google Scholar]
- 114.Silhavy T.J., Kahne D., Walker S. The bacterial cell envelope. Cold Spring Harbor Perspect. Biol. 2010;2:a000414. doi: 10.1101/cshperspect.a000414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.MartÃnez-Carmona M., Gun’ko Y.K., Vallet-Regà M. Mesoporous silica materials as drug delivery: “The nightmare” of bacterial infection. Pharmaceutics. 2018;10:279. doi: 10.3390/pharmaceutics10040279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Caparros M., Pisabarro A.G., de Pedro M.A. Effect of D-amino acids on structure and synthesis of peptidoglycan in Escherichia coli. J. Bacteriol. 1992;174:5549–5559. doi: 10.1128/JB.174.17.5549-5559.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Hammes W., Schleifer K.H., Kandler O. Mode of action of glycine on the biosynthesis of peptidoglycan. J. Bacteriol. 1973;116:1029–1053. doi: 10.1128/JB.116.2.1029-1053.1973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Li Q., Montalban-Lopez M., Kuipers O.P. Increasing the antimicrobial activity of nisin-based lantibiotics against gram-negative pathogens. Appl. Environ. Microbiol. 2018;84:e00052-18. doi: 10.1128/AEM.00052-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Williams G.C., Delves-Broughton J. NISIN. In: Caballero B., editor. Encyclopedia of Food Sciences and Nutrition. 2nd ed. Academic Press; Oxford, UK: 2003. pp. 4128–4135. [Google Scholar]
- 120.Esteban J., García-Coca M. Biofilms. Front. Microbiol. 2017;8:2651. doi: 10.3389/fmicb.2017.02651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Høiby N., Bjarnsholt T., Givskov M., Molin S., Ciofu O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents. 2010;35:322–332. doi: 10.1016/j.ijantimicag.2009.12.011. [DOI] [PubMed] [Google Scholar]
- 122.Aparna M.S., Yadav S. Biofilms: Microbes and disease. Braz. J. Infect. Dis. 2008;12:526–530. doi: 10.1590/S1413-86702008000600016. [DOI] [PubMed] [Google Scholar]
- 123.Crouzet M., Le Senechal C., BrÃzel V.S., Costaglioli P., Barthe C., Bonneu M., Garbay B., Vilain S. Exploring early steps in biofilm formation: Set-up of an experimental system for molecular studies. BMC Microbiol. 2014;14:253. doi: 10.1186/s12866-014-0253-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Di Luca M., Maccari G., Nifosì R. Treatment of microbial biofilms in the post-antibiotic era: Prophylactic and therapeutic use of antimicrobial peptides and their design by bioinformatics tools. Pathog. Dis. 2014;70:257–270. doi: 10.1111/2049-632X.12151. [DOI] [PubMed] [Google Scholar]
- 125.Tanner M.E. Understanding nature’s strategies for enzyme-catalyzed racemization and epimerization. Acc. Chem. Res. 2002;35:237–246. doi: 10.1021/ar000056y. [DOI] [PubMed] [Google Scholar]
- 126.Chen D., Cao Y., Yu L., Tao Y., Zhou Y., Zhi Q., Lin H. Characteristics and influencing factors of amyloid fibers in S. mutans biofilm. AMB Express. 2019;9:31. doi: 10.1186/s13568-019-0753-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Hochbaum A.I., Kolodkin-Gal I., Foulston L., Kolter R., Aizenberg J., Losick R. Inhibitory effects of d-Amino acids on staphylococcus aureus biofilm development. J. Bacteriol. 2011;193:5616. doi: 10.1128/JB.05534-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Tong Z., Zhang L., Ling J., Jian Y., Huang L., Deng D., Tandheelkunde P. An in vitro study on the effect of free amino acids alone or in combination with nisin on biofilms as well as on planktonic bacteria of Streptococcus mutans. PLoS ONE. 2014;9:e99513. doi: 10.1371/journal.pone.0099513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Spiers A.J., Bohannon J., Gehrig S.M., Rainey P.B. Biofilm formation at the air-liquid interface by the Pseudomonas fluorescens SBW25 wrinkly spreader requires an acetylated form of cellulose. Mol. Microbiol. 2003;50:15–27. doi: 10.1046/j.1365-2958.2003.03670.x. [DOI] [PubMed] [Google Scholar]
- 130.Zhao L., Jiang J., Zhu Z., Liao Z., Yao X., Yang Y., Cao Y., Jiang Y. Lysine enhances the effect of amphotericin B against Candida albicans in vitro. Acta Biochim. Biophys. Sin. 2016;48:182–193. doi: 10.1093/abbs/gmv125. [DOI] [PubMed] [Google Scholar]
- 131.Baldo V., Bertoncello C., Cocchio S., Fonzo M., Pillon P., Buja A., Baldovin T. The new pandemic influenza A/(H1N1)pdm09 virus: Is it really “new”? J. Prev. Med. Hyg. 2016;57:19–22. [PMC free article] [PubMed] [Google Scholar]
- 132.Li Y., Jia R., Al-Mahamedh H., Xu D., Gu T. Enhanced biocide mitigation of field biofilm consortia by a mixture of D-Amino Acids. Front. Microbiol. 2016;7:896. doi: 10.3389/fmicb.2016.00896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Xu D., Li Y., Gu T. D-Methionine as a biofilm dispersal signaling molecule enhanced tetrakis hydroxymethyl phosphonium sulfate mitigation of Desulfovibrio vulgaris biofilm and biocorrosion pitting. Mater. Corros. 2014;65:837–845. doi: 10.1002/maco.201206894. [DOI] [Google Scholar]
- 134.De Jonge B.L.M., Gage D., Xu N. The carboxyl terminus of peptidoglycan stem peptides is a determinant for methicillin resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 2002;46:3151–3155. doi: 10.1128/AAC.46.10.3151-3155.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Österberg T., Wadsten T. Physical state of l-histidine after freeze-drying and long-term storage. Eur. J. Pharm. Sci. 1999;8:301–308. doi: 10.1016/S0928-0987(99)00028-7. [DOI] [PubMed] [Google Scholar]
- 136.Hansen L.J.J., Daoussi R., Vervaet C., Remon J., De Beer T.R.M. Freeze-drying of live virus vaccines: A review. Vaccine. 2015;33:5507–5519. doi: 10.1016/j.vaccine.2015.08.085. [DOI] [PubMed] [Google Scholar]
- 137.Mohammed A.R., Coombes A.G.A., Perrie Y. Amino acids as cryoprotectants for liposomal delivery systems. Eur. J. Pharm. Sci. 2007;30:406–413. doi: 10.1016/j.ejps.2007.01.001. [DOI] [PubMed] [Google Scholar]
- 138.Van Winden E.C.A. Methods in Enzymology. Volume 367. Academic Press; Cambridge, MA, USA: 2003. Freeze-Drying of liposomes: Theory and practice; pp. 99–110. [DOI] [PubMed] [Google Scholar]
- 139.Khan I., Elhissi A., Shah M., Alhnan M.A., Ahmed W. Liposome-based carrier systems and devices used for pulmonary drug delivery. In: Davim J.P., editor. Biomaterials and Medical Tribology. Woodhead Publishing; Sawston, UK: 2013. pp. 395–443. [Google Scholar]
- 140.Munro H.M. Nutritional consequences of excess amino acid intake. Adv. Exp. Med. Biol. 1978;105:119–129. doi: 10.1007/978-1-4684-3366-1_8. [DOI] [PubMed] [Google Scholar]
- 141.Mittal B. Pharmacokinetics and preformulation. In: Mittal B., editor. How to Develop Robust Solid Oral Dosage Forms from Conception to Post-Approval. Academic Press; Cambridge, MA, USA: 2017. pp. 17–37. [Google Scholar]
- 142.Savjani K.T., Gajjar A.K., Savjani J.K. Drug solubility: Importance and enhancement techniques. ISRN Pharm. 2012;2012:195727. doi: 10.5402/2012/195727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Gong Y., Grant D.J.W., Brittain H.G. Principles of solubility. In: Augustijns P., Brewster M.E., editors. Solvent Systems and Their Selection in Pharmaceutics and Biopharmaceutics. Springer; New York, NY, USA: 2007. pp. 1–27. [Google Scholar]
- 144.Kalepu S., Nekkanti V. Insoluble drug delivery strategies: Review of recent advances and business prospects. Acta Pharm. Sin. B. 2015;5:442–453. doi: 10.1016/j.apsb.2015.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Zhao J., Yang J., Xie Y. Improvement strategies for the oral bioavailability of poorly water-soluble flavonoids: An overview. Int. J. Pharm. 2019;570:118642. doi: 10.1016/j.ijpharm.2019.118642. [DOI] [PubMed] [Google Scholar]
- 146.Serajuddin A.T. Salt formation to improve drug solubility. Adv. Drug Deliv. Rev. 2007;59:603–616. doi: 10.1016/j.addr.2007.05.010. [DOI] [PubMed] [Google Scholar]
- 147.Lucas S. The Pharmacology of Indomethacin. Headache. 2016;56:436–446. doi: 10.1111/head.12769. [DOI] [PubMed] [Google Scholar]
- 148.ElShaer A., Khan S., Perumal D., Hanson P., Mohammed A.R. Use of amino acids as counterions improves the solubility of the BCS II model drug, indomethacin. Curr. Drug Deliv. 2011;8:363–372. doi: 10.2174/156720111795767924. [DOI] [PubMed] [Google Scholar]
- 149.Iyire A., Alaayedi M., Mohammed A.R. Pre-formulation and systematic evaluation of amino acid assisted permeability of insulin across in vitro buccal cell layers. Sci. Rep. 2016;6:32498. doi: 10.1038/srep32498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Kelwade J., Parekh H., Dukle V., Sethi B.K. How many oral antidiabetic drugs before insulin? Indian J. Endocrinol. Metab. 2017;21:249–250. doi: 10.4103/2230-8210.195994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Reijenga J., van Hoof A., van Loon A., Teunissen B. Development of methods for the determination of pKa values. Anal. Chem. Insights. 2013;8:53–71. doi: 10.4137/ACI.S12304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Manallack D.T. The pK(a) distribution of drugs: Application to drug discovery. Perspect. Med. Chem. 2007;1:25–38. [PMC free article] [PubMed] [Google Scholar]
- 153.Ouellette R.J., Rawn J.D. Organic Chemistry. Academic Press; Boston, MA, USA: 2018. Amino acids, peptides, and proteins; pp. 929–971. [Google Scholar]
- 154.Bhattachar S.N., Deschenes L.A., Wesley J.A. Solubility: It’s not just for physical chemists. Drug Discov. Today. 2006;11:1012–1018. doi: 10.1016/j.drudis.2006.09.002. [DOI] [PubMed] [Google Scholar]
- 155.Kallay N., Kovačević D., Žalac S. Thermodynamics of the solid/liquid interface - its application to adsorption and colloid stability. Interface Sci. Technol. 2006;11:133–170. [Google Scholar]
- 156.Barret R. Importance and Evaluation of the pKa. In: Barret R., editor. Therapeutical Chemistry. Elsevier; Amsterdam, The Netherlands: 2018. pp. 21–51. [Google Scholar]
- 157.Hadjesfandiari N., Parambath A. Stealth coatings for nanoparticles: Polyethylene glycol alternatives. In: Parambath A., editor. Engineering of Biomaterials for Drug Delivery Systems. Woodhead Publishing; Cambridge, UK: 2018. pp. 345–361. [Google Scholar]
- 158.Moldoveanu S.C., David V. Properties of analytes and matrices determining HPLC selection. In: Moldoveanu S.C., David V., editors. Selection of the HPLC Method in Chemical Analysis. Elsevier; Boston, MA, USA: 2017. pp. 189–230. [Google Scholar]
- 159.Haynes W.M., Lide D.R., Bruno T.J. Properties of amino acids. In: Haynes W.M., Lide D.R., Bruno T.J., editors. CRC Handbook of Chemistry and Physics. 97th ed. CRC Press; Boca Raton, FL, USA: 2014. [Google Scholar]
- 160.Kundukad B., Udayakumar G., Grela E., Kaur D., Rice S.A., Kjelleberg S., Doyle P.S. Weak acids as an alternative anti-microbial therapy. Biofilm. 2020;2:100019. doi: 10.1016/j.bioflm.2020.100019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Doran P.M. Bioprocess Engineering Principles. 2nd ed. Academic Press; London, UK: 2013. Unit operations; pp. 445–595. [Google Scholar]
- 162.Aftabuddin M., Kundu S. Hydrophobic, hydrophilic, and charged amino acid networks within protein. Biophys. J. 2007;93:225–231. doi: 10.1529/biophysj.106.098004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Lodish H., Berk A., Zipursky S.L., Matsudaira P., Baltimore D., Darnell J. Hierarchical Structure of Proteins—Molecular Cell Biology—NCBI Bookshelf. [(accessed on 17 November 2019)]; Available online: https://www.ncbi.nlm.nih.gov/books/NBK21581/
- 164.Manallack D.T., Prankerd R.J., Yuriev E., Oprea T.I., Chalmers D.K. The significance of acid/base properties in drug discovery. Chem. Soc. Rev. 2013;42:485–496. doi: 10.1039/C2CS35348B. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Nadendla K., Friedman S.H. Light control of protein solubility through isoelectric point modulation. J. Am. Chem. Soc. 2017;139:17861–17869. doi: 10.1021/jacs.7b08465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Fischel-Ghodsian F., Brown L., Mathiowitz E., Brandenburg D., Langer R. Enzymatically controlled drug delivery. Proc. Natl. Acad. Sci. USA. 1988;85:2403–2406. doi: 10.1073/pnas.85.7.2403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Acharya P.C., Marwein S., Mishra B., Ghosh R., Vora A., Tekade R.K. Role of salt selection in drug discovery and development. In: Tekade R.K., editor. Dosage Form Design Considerations. Academic Press; Cambridge, MA, USA: 2018. pp. 435–472. [Google Scholar]
- 168.Neau S.H., Loka N.C. Pharmaceutical Salts. In: Liu R., editor. Water-Insoluble Drug Formulation. CRC Press; Boca Raton, FL, USA: 2018. pp. 451–469. [Google Scholar]
- 169.Gupta D., Bhatia D., Dave V., Sutariya V., Varghese Gupta S. Salts of therapeutic agents: Chemical, physicochemical, and biological considerations. Molecules. 2018;23:1719. doi: 10.3390/molecules23071719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Barrett G. Chemistry and Biochemistry of the Amino Acids. Springer Science & Business Media; Berlin, Germany: 2012. [Google Scholar]