

Abstract
Trefoil factor family peptides (TFF1, TFF2, TFF3) are typically co-secreted together with mucins. Tff1 represents a gastric tumor suppressor gene in mice. TFFs are also synthesized in minute amounts in the immune and central nervous systems. In mucous epithelia, they support rapid repair by enhancing cell migration (“restitution”) via their weak chemotactic and anti-apoptotic effects. For a long time, as a paradigm, this was considered as their major biological function. Within recent years, the formation of disulfide-linked heterodimers was documented for TFF1 and TFF3, e.g., with gastrokine-2 and IgG Fc binding protein (FCGBP). Furthermore, lectin activities were recognized as enabling binding to a lipopolysaccharide of Helicobacter pylori (TFF1, TFF3) or to a carbohydrate moiety of the mucin MUC6 (TFF2). Only recently, gastric TFF1 was demonstrated to occur predominantly in monomeric forms with an unusual free thiol group. Thus, a new picture emerged, pointing to diverse molecular functions for TFFs. Monomeric TFF1 might protect the gastric mucosa as a scavenger for extracellular reactive oxygen/nitrogen species. Whereas, the TFF2/MUC6 complex stabilizes the inner layer of the gastric mucus. In contrast, the TFF3–FCGBP heterodimer (and also TFF1–FCGBP) are likely part of the innate immune defense of mucous epithelia, preventing the infiltration of microorganisms.
Keywords: gastric cancer, reactive oxygen species, reactive nitrogen species, inflammation, trefoil factor, lectin, gastrokine, FCGBP, mucin, innate immunity
1. Introduction
Mammalian trefoil factor family (TFF) peptides (TFF1, TFF2, and TFF3) are characterized by a common structural motif, the TFF domain (formerly: P-domain, trefoil domain), which contains six conserved cysteine residues with three intramolecular disulfide bonds (CysI-V, CysII-IV, and CysIII-VI; Figure 1). TFF1 (human: 60 amino acids) and TFF3 (human: 59 amino acids) consist of single TFF domains and additionally contain free 7th cysteine (CysVII) residues located outside the TFF domain. TFF2 (human: 106 amino acids) contains two TFF domains and the N- and the C-terminals are disulfide-linked via two additional cysteine residues outside the TFF domains (reviews: [1,2,3,4,5,6]).
Figure 1.

Disulfide-linked three-leafed structures of the prototypic trefoil factor family peptides TFF1 and TFF2. Cysteine residues (C) are shown in green (numbering in Roman numerals).
In the skin of frogs, such as Xenopus laevis, even more TFF peptides exist [3]. Furthermore, shuffled TFF modules are present in a number of mosaic proteins, e.g., the human zona pellucida proteins ZP1 and ZPB, the sugar-degrading enzymes sucrase–isomaltase, α-glucosidase, and maltase–glucoamylase [3], the frog skin proteins APEG [7] and “βγ-crystallin and trefoil factor” (βγ-CAT) [8], and the frog integumentary mucins FIM-A.1 and FIM-C.1 [9,10,11]. Generally, TFF domains are encoded by single exons belonging to the class 1-1 and thus represent a unique family of cysteine-rich shuffled modules [3].
1.1. Exocrine and Endocrine Secretion of TFF Peptides
Mucous epithelia are the predominant expression sites of TFF peptides. Here, the exocrine secretion of TFFs occurs together with different mucins [3,6]. TFF1 and TFF2 are predominantly synthesized in the stomach, i.e., in surface mucous cells (SMCs) and mucous neck/antral gland cells, respectively [2,6]. Additionally, TFF2 is expressed in duodenal Brunner glands, and in some species, (such as the pig and mouse, but not human) also in the exocrine pancreas [1,12]. In contrast, TFF3 shows a much wider distribution and is found mainly in intestinal goblet cells, but also in the respiratory and urogenitary tracts, the salivary glands, esophageal submucosal glands, conjunctiva, and the inner ear (Table 1) [3,13,14]. Thus, TFFs appear in many body fluids, such as saliva, gastric juice, urine, tears, and breast milk [5].
Table 1.
Expression sites of human TFF peptides.
| TFF Peptides | Exocrine Secretion (Major/Minor) |
Endocrine Secretion |
|---|---|---|
| TFF1 |
Stomach, conjunctiva, lacrimal apparatus, salivary glands, lung, urogenitary tract |
CNS |
| TFF2 |
Stomach, Brunner’s glands, salivary glands |
CNS, lymphoid tissues |
| TFF3 |
Intestine, salivary glands, lung, uterus, vagina, conjunctiva, lacrimal apparatus, inner ear, esophagus, stomach, gallbladder, Vater’s ampulla, urinary tract |
CNS, thyroid, lymphoid tissues, endocrine pancreas |
Major expression sites are shown in bold.
Furthermore, minute amounts of TFF peptides are also secreted in an endocrine manner, e.g., by the central nervous system (CNS), as well as by the immune system (Table 1). In particular, TFF3 is synthesized in neurons (such as oxytocinergic neurons of the hypothalamus), activated microglial cells, and astrocytes of the brain [15,16,17,18]. All three TFF peptides show developmental changes, particularly in the cerebellum, the most pronounced being TFF3 [19]. In contrast, TFF2 (and to some extent also TFF3) is expressed in peritoneal macrophages and lymphoid tissues, such as the spleen (memory T-cells), thymus, lymph nodes, and bone marrow [20,21,22,23,24]. TFF3 (and little TFF1) is also expressed in the endocrine pancreas [25] and TFF3 is also expressed in the thyroid [26]. This would explain why all three TFF peptides are detectable in human serum [27].
1.2. Ectopic Expression of TFF Peptides during Inflammatory Conditions and in Tumors
TFF peptides, together with epidermal growth factor (EGF), are aberrantly expressed in diverse chronic ulcerative conditions, often in glandular structures termed the “ulcer-associated cell lineage” (UACL) [28]. Thus, TFF peptides are part of a chronic inflammatory response during a variety of diseases, such as ulcerative colitis, diverticulitis, cholecystitis, inflammatory bowel disease, gastro-esophageal reflux disease, pancreatitis, and chronic obstructive pulmonary disease [2,3,5,29,30,31,32].
Of special note, TFF1 in particular is ectopically expressed in various chronic inflammatory conditions, even in murine models of encephalitis and asthma, and in the murine spleen after Toxoplasma gondii infection [31,33,34,35,36].
Further pathological expression of TFF peptides occurs in metaplasias [37], as well as in different kinds of tumors [2,3,5,38,39]. Of note, somatic mutations in the TFF1 gene seem to be associated with gastric cancer and there is a strikingly reduced TFF1 expression in the majority of gastric carcinomas [40,41].
1.3. Phenotypes of Tff-Deficient (TffKO) Animals
For a long time, mice deficient in Tff1, Tff2 and Tff3 have been available. The most prominent phenotype is observed in Tff1KO mice.
Tff1KO mice mainly show a gastric phenotype, i.e., they all develop antropyloric adenomas with ~30% progressing to carcinomas [42,43]. Thus, these mice represent a recognized model for spontaneous antral tumorigenesis [44]. Of note, Tff1KO mice also show strongly reduced Tff2 expression, particularly in the gastric corpus [42,45]. Antral carcinogenesis is an age-dependent multi-step process and is accompanied by NF-κB-mediated chronic inflammation [46,47]. Tumor growth can be suppressed with the selective Cox-2 inhibitor celecoxib [43,47,48]. There are indications that long-lived precursor cells in antral units located at positions +4 (Cckbr+) and +5 (Mist+) are involved in tumorigenesis in Tff1KO mice [45]. Of note, in Tff1KO mice, continuous self-renewal is also dysregulated in fundic units, but this does not lead to carcinogenesis [49]. Furthermore, Tff1KO mice show generally increased chemically-induced tumorigenesis [50]. Taken together, Tff1 represents a gastric tumor suppressor gene in mice [43] and TFF1 mutations and dysregulated TFF1 expression seem to be critical to the pathogenesis of most gastric carcinomas in humans [40,41]. Of note, mutations in the interleukin 6 (IL6) signal transducer gp130, which blocked SHP2–Ras–ERK signaling (gp130757F mice), showed reduced Tff1 levels and a phenotype (antropyloric adenomas, but no carcinomas) highly similar to Tff1KO mice [51,52].
In contrast, Tff2KO mice do not show a striking phenotype. Other than moderate morphological changes [53], they have a highly increased Tff3 expression in the antrum [54]. They also have increased susceptibility to Helicobacter-induced gastritis and to Yersinia enterocolitica infection, as well as a delayed recovery after dextran sodium sulfate (DSS)-induced colitis [22,55,56,57]. Furthermore, Tff2KO mice show the dysregulated expression of immune response-related genes and their macrophages are hyperresponsive to interleulin-1β [22,54].
Nine-day-old (P9) Tff2KO rats are highly sensitive to oral infection with Escherichia coli K1, leading to bacteremia probably due to the loss of Tff2 in the developing small intestine [58]. Of note, Tff2 expression is developmentally regulated in the neonatal rat intestine, reaching a peak at P9 and dropping sharply thereafter [59].
Tff3-deficient mice have strongly increased sensitivity in the DSS colitis model and these animals are particularly sensitive to radiation-induced mucosal injury and chemotherapy [60,61]. Of note, mutations in the interleukin 6 (IL6) signal transducer gp130, which abrogated STAT1/3 signaling (gp130ΔSTAT mice), showed reduced Tff3 levels and a phenotype highly similar to Tff3KO mice [51,52]. Furthermore, Tff3KO mice show minor morphological differences in the foliation of the cerebellum, as well as motoric deficits [62].
1.4. TFF Peptides Enhance Cell Migration: Implications for Mucosal Protection and Immune Responses
There is a large body of literature describing the weak motogenic effects of all three TFF peptides for a variety of different epithelial and immune cells in vitro, starting in the year 1994 [3,5,63]. Thus, relatively rapidly, this became a paradigm explaining the biological function of TFF peptides with no clear differences between TFF1, TFF2, and TFF3. For example, TFFs enhance the cell migration of intestinal epithelial cells [63,64], corneal epithelial cells [65,66], bronchial epithelial cells [67], gastric epithelial cells [68], oral keratinocytes [69], pancreatic cells [70], and monocytes [20]. All three TFF peptides show generally weak chemotactic activity [71,72]. There is also a synergistic effect with epidermal growth factor (EGF) [72,73]. In line with their motogenic effects, TFF peptides were also described as pro-angiogenic factors [74].
For all three TFF peptides, an anti-apoptotic effect has also been observed [75,76,77,78]. However, for TFF3, a pro-apoptotic effect was also reported [79]. A combined motogenic and anti-apoptotic activity of TFF peptides would be in line with the observation that, during invasion of the extracellular matrix, cell migration and survival mechanisms are coordinately regulated [80]. This ensures that both effects synergistically support the important process of cell migration/invasion, e.g., during wound healing, immune responses, and angiogenesis.
A special form of rapid wound healing occurs in mucous epithelia after superficial injury, i.e., by the cell migration of neighboring cells. This process is called “restitution” and starts within minutes after damage. Thus, TFFs would be well designed to act as luminal protection peptides, enhancing restitution only after mucosal injury [81,82].
A protective function of TFF peptides for mucous epithelia is in line with numerous in vivo studies using different animal models of disease (after chemical damage or restraint; for compilations, see [3,5]). Additionally, the active delivery of TFF peptides by genetically modified Lactococcus lactis was effective in preventing and healing DSS-induced colitis [83].
TFF3 is also linked to innate immunity, as its synthesis in intestinal goblet cells is selectively induced after the activation of Toll-like receptor (TLR) 2 by commensal bacteria [84]. This probably occurs by an indirect mechanism. Of note, recombinant TFF3 can rescue Tlr2KO mice from increased morbidity and mortality during acute colonic injury, probably due to its anti-apoptotic effect [84].
Generally, the motogenic and anti-apoptotic effects of TFF peptides are rather weak and were observed in a concentration range of about 10−6 to 10−7 M or even higher [13,71]. Such relatively high concentrations are atypical of a classical high-affinity peptide ligand, such as epidermal growth factor (EGF), which activates its specific receptors even at concentrations as low as 10−10 M [72]. Thus, it is not surprising that the repeated attempts to identify high-affinity receptors for TFF peptides have failed [85].
However, several transmembrane proteins could be characterized to bind TFF2 with low affinity, such as a porcine β-integrin and a porcine 224k protein (CRP-ductin), with similarity to Deleted in Malignant Brain Tumors 1 (DMBT1) [86]. In addition, TFF3 has been reported to bind to the secreted variant DMBT1gp340 in a Ca2+-dependent manner [87], the latter being an agglutinin playing a role in mucosal innate immunity [88]. Later on, TFF2 and TFF3 were described as low-affinity ligands for the chemokine receptors CXCR4 and CXCR7, i.e., TFF2 and TFF3 were active at a concentration of about 5 × 10−7 M [89,90]. Thus, the ligation of TFF peptides to CXCR4 and CXCR7 would explain their chemotactic effects, as CXCR4 and CXCR7 are the high-affinity receptors for the chemokine CXCL12/stromal cell-derived factor (SDF-1), which is a highly potent chemotactic peptide and regulates apoptosis at a concentration below 10−9 M [91]. Thus, to some extent, the TFF2-CXR4 axis in particular seems to play a direct role in gastric repair [92], as well as in suppressing colorectal carcinogesis via the neural innervation of splenic memory T-cells [24]. Furthermore, cell migration was also promoted by TFF2 (2 × 10−7 M) via the activation of the proteinase-activated receptor PAR4 [93], and TFF3 (10−6 M) was claimed to activate PAR2 [94].
Taken together, TFF peptides cannot be considered as high-affinity ligands for specific transmembrane receptors. As their concentrations in mucous gels are rather high, it seems highly unlikely that their protective function is based solely on their action as low-affinity ligands, e.g., for CXCR4 and CXCR7. It is more realistic to expect additional molecular functions for TFF peptides.
2. Molecular Forms of TFF Peptides and Their Interaction Partners: Functional Implications
TFF1-3 are typical secretory peptides. TFF1 and TFF3 are special, as they contain an odd number of cysteine residues, with CysVII located outside the conserved TFF domain. Generally, the existence of unpaired cysteine residues is highly unlikely for secretory proteins, as disulfide formation is enforced in the endoplasmic reticulum (ER) [95]. Thus, TFF1 and TFF3 were expected to occur naturally as homodimers and most of the in vitro wound healing experiments and the in vivo studies using animal models were indeed performed with homodimers. There are reports that the dimeric forms are biologically more active than the monomeric forms [71,96,97,98]. Even the binding of TFF1 to Helicobacter pylori and TFF3 to DMBT1gp340 was reported to depend on dimerization [87,99].
Only later did biochemical studies reveal that TFF1 and TFF3 occur in vivo in different molecular forms and are also capable of forming disulfide-linked heterodimers with at least gastrokine 2 (GKN2) and IgG Fc binding protein (FCGBP). Thus far, the following hetero(di)meric forms have been identified: TFF1-GKN2, TFF1-FCGBP, and TFF3-FCGBP [45,100,101,102,103]. This indicates that the biological function of TFF peptides is obviously more complex.
2.1. TFF Domains Have Different Lectin Activities
All three TFF peptides were shown to bind to carbohydrate moieties, but to quite different extents and probably also with different specificities. Dimeric TFF1 interacts with a core oligosaccharide portion of the H. pylori lipopolysaccharide in a pH-dependent manner and TFF3 binds to a lesser extent [99,104,105]. Binding studies with truncated lipopolysaccharides from H. pylori mutants point to N-acetylglucosamine (GlcNAc) being part of the carbohydrate structure recognized by TFF1 [105]. Dimeric TFF1 also binds weakly to the gastric mucin MUC6, probably to a carbohydrate moiety conserved from frog to human, e.g., the terminal αGlcNAc residue (see also Section 2.3) [103,105]. TFF2 is strongly, but non-covalently, bound to gastric mucus [12,106]; the specific Ca2+- and pH-dependent lectin activity of TFF2 was narrowed down to the unusual O-linked terminal carbohydrate moiety GlcNAcα1→4Galβ1→R of the mucin MUC6 [107,108,109,110]. Thus, the lectin specificities of TFF1 and TFF2 seem to be different, but both could share GlcNAc as a common part of a complex ligand.
Furthermore, the frog integumentary mucins FIM-A.1 and FIM-C.1 from Xenopus laevis are capable of forming high-molecular-mass complexes probably via the lectin activities of their intrinsic multiple TFF modules [111]. In addition, the TFF modules in the sugar-degrading enzymes sucrase–isomaltase, α-glucosidase, and maltase–glucoamylase are also expected to have lectin activities [3]. Finally, the TFF modules in the human zona pellucida proteins ZP1 and ZPB would be perfectly designed to play a key role as lectins during fertilization; this could account for a species specificity.
Generally, the lectin activities of TFF peptides might explain diverse biological effects, such as their weak motogenic, (anti)-apoptotic, and angiogenic effects [108]. Theoretically, TFF peptides could activate a plethora of transmembrane glycoproteins by binding to their carbohydrate moieties, such as the receptors CXCR4 and CXCR7 [89,90], integrins [86], and CRP-ductin/DMBT1gp340 [86,87]. Furthermore, similar to galectins, TFF2 and homodimeric forms of TFF1 and TFF3 in particular could form two- and three-dimensional cross-linked lattices with various transmembrane glycoproteins, leading to supermolecular assembly and signal transduction [112,113].
2.2. Gastric TFF1 Mainly Occurs in a Monomeric Form with an Unusual Free Thiol Group: Possible Intracellular and Extracellular Functions in the Gastric Mucus Barrier and during Inflammation
Human gastric TFF1 occurs in different molecular forms, i.e., monomeric TFF1, homodimeric TFF1, and heterodimeric forms (TFF1-GKN2, TFF1-X/60k, TFF1-FCGBP) [103,114]. Generally, only minor amounts of gastric TFF1 are associated with the gastric mucus, as shown for humans [103,106] and X. laevis (ortholog xP1) [115]. This is even visible at the electron microscopic level, where TFF1/xP1 is localized in dense core regions of secretory granules and is not mixed with mucins [49,116]. However, dimeric TFF1 is capable of weakly binding in vitro to the mucin MUC6 [103].
Gastric TFF1 is also capable of forming a heterodimer with GKN2, as shown for humans and mice [45,100,103,106]. Of note, when gastric specimens are classically extracted (without SDS), TFF1–GKN2 is hardly detectable, particularly in the corpus [45,103]. Part of it seems to be soluble only in 1% SDS [45,103]. Thus, we postulated that TFF1–GKN2 might be a constituent of the inner insoluble layer of the gastric mucus [103]. This aspect is discussed further in Section 2.3.
Minute amounts of gastric TFF1 form a disulfide-linked heterodimer with FCGBP, as shown for humans and mice [45,103]. This is not surprising, as both TFF1 and FCGBP contain an odd number of cysteine residues and are synthesized in SMCs. The concentration of TFF1–FCGBP is higher in the antrum when compared with the corpus, as FCGBP is mainly expressed in the antrum of humans and mice [45,117]. The formation of the TFF1–FCGBP heterodimer is analogous, as described for TFF3–FCGBP in the intestine and saliva [101,102]. The postulated function of TFF1/3-FCGBP heterodimers in the innate immune defense is discussed in more detail in Section 2.4.
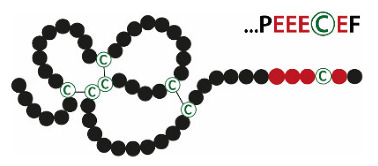
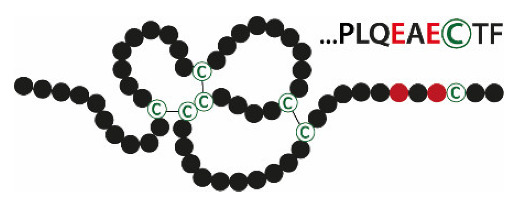
Systematic biochemical investigations revealed that, in gastric extracts, the major form of TFF1 is a secreted monomer with a free thiol group at CysVII. This has been shown for X. laevis (ortholog xP1) [115], mice [45], and humans [103]. Normally, such free thiol groups act as a three-way switch, mediating assembly, retention, or degradation in the ER [118], but they are not secreted [95]. However, the unusual free thiol group in human TFF1 at CysVII is probably masked by four flanking acidic amino acids (see Table 2), and thus TFF1 mainly escapes assembly (dimerization), retention, or degradation and is predominantly secreted as a monomer. This explanation is based upon similar results obtained from Ig light chains, where flanking acid residues also allow the secretion of an unpaired cysteine [119]. Such flanking acid residues can drastically change the pKa of cysteine residues [120,121]. Of note, the unusual acid residues in TFF1/xP1 flanking CysVII are conserved in X. laevis (…PECVII), mice (…QEEECVIIPF), and humans (…PPEEECVIIEF) [122]. It is expected that CysVII is exceptionally reactive (nucleophilic) because of the pronounced steric exposure outside the TFF domain and separation by proline residues [103]. Thus, it is hypothesized that TFF1, via its free thiol at CysVII, has special intracellular as well as extracellular molecular functions.
Table 2.
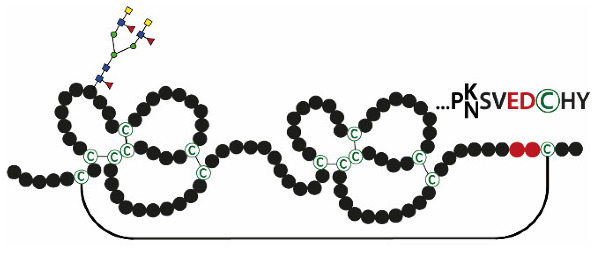
Structures and natural forms of human TFF peptides at their predominant expression sites.
| TFFs | Structures | Expression Sites | Natural Forms (Major/Minor) |
|---|---|---|---|
| TFF1 |
|
Stomach |
TFF1mono TFF1-FCGBP TFF1-X (60 k) TFF1-GKN2 TFF1-TFF1 |
| TFF2 |
|
Stomach |
TFF2/MUC6 TFF2 |
| TFF3 |
|
Intestine, salivary glands |
TFF3-FCGBP TFF3-TFF3 TFF3mono |
Acidic residues flanking the C-terminal cysteines are shown in red. For TFF2, the N-linked carbohydrate moiety is depicted (yellow squares, GalNAc; blue squares, GlcNAc; green circles, Man; red triangles, Fuc). Major natural TFF forms are shown in bold.
Intracellularly, TFF1 is expected to be involved in the folding of cysteine-rich glycoproteins (such as the mucin MUC5AC), as in Tff1KO mice the unfolded protein response (UPR) is activated [123]. For example, the expression of the protein disulfide isomerases (PDI) Pdia3/ERp57, Pdia4/ERp72, Pdia17/Agr2, and Grp78 is significantly elevated in Tff1KO mice [45,123]. This led to the hypothesis that TFF1 plays a role in the ER protein folding machinery [123]. Thus far, the precise molecular function of TFF1 has not been elucidated. For example, it could have an intracellular function for the correct assembly of the mucin MUC5AC. Of note, recently a disulfide-linked TFF1 heterodimer (TFF1-X) with an Mr of about 60k was detected in humans [103], which hypothetically might represent a heterodimer with ERp57. ERp57 is capable of forming intermolecular disulfide bonds, e.g., with the major histocompatibility complex (MHC) class I [124], and is also known for its role in oncogenic transformation [125].
Extracellularly, the free thiol group of TFF1 has been proposed to act as a scavenger for reactive oxygen/nitrogen species (ROS/RNS) [45,103,115]. The formation of sulfur-centered radicals is a characteristic feature after the reaction of thiols with ROS [126,127]. As a consequence, sulfur-containing amino acids, such as cysteine, have a protective role against free radicals and heavy metals [128]. Protection from ROS/RNS is particularly important for the stomach, as oxidative stress plays a major role in stomach disorders [129]. Thus, TFF1 would be particularly well suited to protect the gastric surface mucous cells and probably also the adjacent population of highly proliferating precursor cells in the isthmus (review: [130]). The firmly adherent inner layer of the gastric mucus–bicarbonate barrier maintains a pH gradient with a nearly neutral pH at the mucosal surface [131]. This neutral pH allows the formation of thiolate anions and they are more apt to react chemically as a nucleophile with ROS/RNS [120,121,127]. Such a scavenger function for TFF1 is physiologically most important in the stomach, as here the extracellular level of ROS (e.g., H2O2 and O2-) is high due to dual oxidases (DUOX) and NADPH oxidases (NOX), respectively. DUOX plays a key role in innate mucosal immunity, preventing gastric colonization with bacteria [132,133]. Additionally, NOX enzymes are expressed in the gastric mucosa, the different forms being species-dependent [134]. Furthermore, the level of RNS can also be particularly high in the gastric lumen. After the reduction in exogenous nitrate (NO3-) from food to nitrite (NO2-) by saliva and microbiota, HO-NO forms in the gastric juice and disproportionates into NO [135], which is a highly reactive RNS-inducing S-nitrosylation of reactive cysteines [136,137]. In addition, NO can also react with O2− forming peroxynitrite (ONOO−), which is the prototype of a toxic RNS [138]. Critical features for S-nitrosylation are acidic residues flanking the reactive cysteine [139], as well as copper ions [136]. Of special note, copper ions are known to bind to the glutamic acid residues flanking the CysVII of human TFF1 [140]. Taken together, TFF1 would be particularly well suited to protect the stomach from RNS.
The reactive CysVII of monomeric TFF1 could not only have intracellular and extracellular protective functions as a gastric tumor suppressor. TFF1 is also expected to play a protective role during inflammatory processes, as there is a fundamental connection to oxidative stress (“oxidative burst”) [137]. This assumption is supported by the observation that TFF1 is ectopically expressed during various chronic inflammatory conditions in humans [29,141], as well as in animal models of pancreatitis [31], asthma [34], and encephalitis [35], and in the murine spleen after T. gondii infection [36].
2.3. TFF2 is a MUC6-Binding Lectin: Function for the Stabilization of the Inner Insoluble Layer of the Gastric Mucus–Bicarbonate Barrier and More
In contrast to TFF1, gastric TFF2 is predominantly associated with mucins, as shown for humans [106], pigs [12], and X. laevis (ortholog xP4) [115]. Here, it binds to the mucin MUC6, but not MUC5AC [110]. Binding occurs as a lectin to the terminal oligosaccharide GlcNAcα1→4Galβ1→R in a Ca2+- and pH-dependent manner [107,108,109,110]. This explains why TFF2 and MUC6 are co-localized in the gastric mucus barrier [142]. The unusual αGlcNAc residue at the non-reducing terminal of the O-linked glycan of MUC6 is conserved from frog to human and is recognized by both the monoclonal antibody HIK1083, as well as the lectin GSA-II from Griffonia simplicifolia [143,144]. MUC6 is expressed early in vertebrate evolution, but was lost in teleost fishes [145]. This explains why even porcine TFF2 binds to X. laevis gastric mucin [115]. Of special note, α1,4GlcNAc-capped mucin-type O-glycans function as natural antibiotics against H. pylori infection by inhibiting the cholesterol α-glucosyltransferase of H. pylori [146,147]. That is probably one reason why H. pylori co-localizes with MUC5AC in the human stomach [148]. A key enzyme for the synthesis of the terminal αGlcNAc residue is α1,4-N-acetylglucosaminyltransferase (α4GnT); all mice lacking this enzyme (A4gntKO) spontaneously develop antral adenocarcinomas through an inflammation-associated pathway [149]. Thus, αGlcNAc seems to serve as a gastric tumor suppressor.
A characteristic feature of human TFF2 is the N-glycosylation site (Table 2), which is lacking in the porcine and murine homologs. As a hallmark, human gastric TFF2 mainly contains the unusual N-linked monofucosylated N,N’-diacetyllactosediamine (LacdiNAc) oligosaccharide [150]. There is a dramatic diurnal variation of the TFF2 concentration in the gastric juice, with a maximum between 05:00 and 07:00, and N-glycosylation also varied diurnally, with a maximum between 17:00 and 23:00 [151]. The LacdiNAc moiety is also present in the gastric mucin MUC5AC and is recognized by the H. pylori adhesin LabA [152]. This is another reason why H. pylori mainly adheres to MUC5AC in the gastric mucus barrier.
TFF2 binding to MUC6 differs characteristically between human (mainly N-glycosylated TFF2) and pig (non-glycosylated TFF2). In porcine gastric mucus, the binding of TFF2 is much stronger than in human mucus and is even resistant to boiling in SDS [12,109,110]. This remarkable discrepancy cannot be explained by the differences in N-glycosylation, as murine Tff2, which is not N-glycosylated, behaves like human TFF2. Thus, it is assumed that it is rather the amino acid sequence which determines the strength of the TFF2 binding to MUC6. This view is also in agreement with recent results of the two X. laevis orthologs of TFF2, i.e., xP4.1 (N-glycosylated) and xP4.2 (non-glycosylated), where glycosylation has no influence on lectin binding to gastric mucin [115].
Taken together, the highly specific lectin binding of TFF2 to a conserved O-linked oligosaccharide in MUC6 has implications for understanding the structure of the two-layered gastric mucus–bicarbonate barrier, which is composed of a loosely adherent (outer) layer and a firmly adherent, water-insoluble (inner) layer (Figure 2) [108,153].
Figure 2.

Model of the gastric mucus–bicarbonate barrier in the fundus/corpus and the antrum, respectively. The gastric mucus consists of an outer and an inner layer and originates from different cell populations, i.e., surface mucous cells (SMCs) and glandular cells (fundic mucous neck cells (MNCs) and antral gland cells (AGCs); shown in red). The SMCs in the fundus and corpus differ somewhat and they originate from different stem cells (review: [130]). Both SMC types synthesize TFF1, the mucin MUC5AC, and the gastrokines GKN1, and GKN2, whereas the fundic SMCs additionally secrete gastric lipase (LIPF), and the antral SMCs, lysozyme (LYZ), TFF3, and the major portion of Ig Fc binding protein (FCGBP) [117]. The MNCs and AGCs secrete TFF2 and the mucin MUC6 and the TFF2/MUC6 complex are expected to form the rather insoluble matrix of the inner layer. Further components of the inner layer are MUC5AC and probably TFF1–GKN2 and GKN2. The very hydrophobic LIPF is expected to be a component of the inner fundic mucus layer [117]. SMCs also secrete bicarbonate (HCO3−; shown in blue) in exchange with Cl− creating a pH gradient along the inner layer. The dual oxidase (DUOX) in SMCs and the NOX enzymes generate H2O2 and O2−, respectively, at the apical surface, restricting microbial colonization (innate immune defense). The formation of NO in the gastric juice by the disproportionation of HO-NO is also shown.
The outer layer consists mainly of the mucin MUC5AC and only little MUC6 [110,153,154], and mixes with the gastric juice, which also contains TFF2 and mucins (mainly MUC5AC and only little MUC6) [151,155]. The adherent inner layer is composed of alternating layers of the mucins MUC5AC and MUC6 [154]. Here, the tight association of TFF2 and MUC6 is even visible by immunohistochemistry [142]. There are multiple indications that TFF2 stabilizes the water-insoluble inner layer physically [110]. For example, TFF2 affects the viscoelastic properties of mucous gels [156] and the X. laevis ortholog of TFF2 (xP4) prevents the shrinking of secretory granules of esophageal goblet cells during the processing of the samples for electron microscopy, due to its interaction with mucin [116]. Furthermore, Tff2KO mice show accelerated progression of Helicobaccter-induced gastritis, as the inner mucus barrier is probably no longer very tight [55]. Thus, TFF2 and MUC6 are expected to form a mesh-like insoluble matrix (TFF2/MUC6 complex), where TFF2 may crosslink the MUC6 subunits in a Ca2+-dependent manner [109,110]. This matrix also creates an unstirred environment where neutralization by the secreted bicarbonate anions (HCO3−) occurs [131]. This nearly neutral pH in the inner layer also favors the lectin binding of TFF2 to MUC6, whereas an acidic pH in the outer layer favors the dissociation of TFF2 into the gastric juice [109]. Additionally, a large portion of TFF1–GKN2 and probably GKN2 are expected to be constituents of the inner layer, as these molecules are hardly soluble and Gkn2KO mice have increased susceptibility to H. pylori-dependent immunopathology [103,157]. This model is depicted in Figure 2.
In contrast to human, pig TFF2 is also expressed in the pancreas. Here, TFF2 is clearly not associated with mucins and rather occurs in an unusual low-molecular homodimeric form, which is resistant to boiling in SDS [12]. This is probably due to the lack of MUC6 or of α4GnT in the porcine pancreas. Thus, porcine pancreatic TFF2 probably forms a complex with MUC6 only after having reached the pancreatic ducts and then the duodenum via Vater’s ampulla.
Finally, it will be a challenging goal for the future to identify the binding partner and to elucidate the molecular function of TFF2 in the lymphoid system, as Tff2 deficiency influences the immune system [54]. It could well be that it also forms a matrix with MUC6 there.
2.4. TFF3 Mainly Forms a Disulfide-linked Heteromer with FCGBP: Postulated Function in the Innate Immune Defense
Human TFF1 and TFF3 are highly similar, as both consist of a single TFF domain and additionally contain a free CysVII residue (Table 2). However, as a hallmark, TFF1 and TFF3 differ, as TFF1 (in the stomach) is mainly not associated with the high-molecular-mass fraction after size exclusion chromatography [103,106], whereas TFF3 (in the intestine and saliva) is [101,102]. Here, TFF3 forms a disulfide-linked heterodimer with FCGBP, whose mature form consists of nearly 5400 amino acid residues, 435 being cysteine residues [101,158,159]. TFF3–FCGBP can easily form, as both TFF3 and FCGBP are secreted from the same cells, i.e., intestinal goblet cells and the mucous acini of salivary glands, respectively [159,160]. Furthermore, both TFF3 and FCGBP contain an odd number of cysteine residues. Based on quantitative measurements, it was suggested previously that probably only a single TFF3 molecule is linked to FCGBP, despite its enormous number of cysteine residues [101].
As shown for humans and mice, gastric TFF1 is also capable of forming a heteromer with FCGBP, but only to a very small extent [45,103]. Only little TFF1–FCGBP is generated, although both TFF1 and FCGBP are secreted from gastric SMCs. The reason might be the different reactivities of CysVII in TFF1 versus TFF3, as the flanking acidic residues are more numerous in TFF1 (see Table 2), which might prevent its heterodimerization with FCGBP. However, it is expected that the molecular function of TFF1–FCGBP (in the stomach) and TFF3–FCGBP (in the intestine, saliva, and many other mucous epithelia) is similar.
Currently, the precise molecular function of FCGBP and its heteromers with TFF3 (TFF3–FCGBP) and TFF1 (TFF1–FCGBP) is not known. The repetitive glycoprotein FCGBP is ubiquitous in vertebrates and cephalochordates and it is secreted by most mucous epithelia from a variety of mucin-producing cells (similar as TFF3), and thus appears in the corresponding body fluids [145,159]. There are indications that FCGBP has a role in the innate immunity of mucous epithelia, where it is thought to regulate pathogen attachment and disease progression [161]. For example, FCGBP is an early response gene after microbial infection from fishes to humans [161,162] and it has also been hypothesized to act as a viral trap for HIV–antibody complexes [163]. Its expression is strongly induced by the TH2 cytokine interleukin-13 [164]. The multiple modular cysteine-rich domains of FCGBP might support the clearing of microorganisms or prevent bacterial infiltration through the mucus layer and even a number of bacterial proteins bear N-terminal domains homologous to the N-terminal domain of FCGBP [145]. The latter shows similarity to an enzyme from frog skin, which catalyzes the isomerization of L-amino acid residues into their corresponding D-enantiomers [165]. The heterodimerization of FCGBP with TFF3 (and also with TFF1) could synergistically enhance or modulate the binding of microorganisms due to the lectin activities of TFF1 and TFF3 [99]. Furthermore, TFF3–FCGBP could form a network by interacting with DMBTgp340, which is an agglutinin that also plays a general role in mucosal innate immunity [87,88]. For example, DMBT-1 aggregates Streptococcus mutans and S. sanguis [166]. The interaction could be lectin-based, similar to that of DMBT1 and galectin 3 [167]. Of note, FCGBP was also reported to be able to bind covalently to the mucin MUC2, probably as a component of the inner colonic mucus layer [168]. TFF3 may also play a role here. This might help to explain the phenotype of Tff3KO mice in the DSS colitis model [60,169].
Other than playing a role in the innate immune defense of mucous epithelia, FCGBP could also fulfill a structural matrix function, as FCGBP seems to be expressed in the aorta, where it belongs to the most downregulated genes in patients with ruptured abdominal aortic aneurisms [170].
3. Conclusions and Clinical Perspectives
Taken together, within recent years, a body of evidence has been accumulated that TFF peptides fulfill diverse roles in the protection of mucous epithelia, other than acting solely as motogens (old paradigm). Their relatively high abundance in mucous epithelia does not fit with a predominant role as typical high-affinity ligands for receptors in mucous epithelia. However, even as low-affinity ligands, they can synergistically support mucosal repair, e.g., by restitution. In this case, the activation of certain glycosylated receptors, such as CXCR4 and CXCR7, might occur via the lectin activities of TFF peptides and would depend on the glycosylation of these receptors; the latter could vary in different cell types and would thus allow some specificity and diversity.
Currently, TFF peptides seem to have rather structural roles as part of the insoluble gastric inner mucus–bicarbonate barrier layer (TFF2/MUC6 complex) or as part of a postulated mucosal clearance/barrier system for different microbes (TFF3–FCGBP, TFF1–FCGBP). The latter might also be able to trap viruses with the help of immunoglobulins on mucosal surfaces. Both of these functions would indicate important roles of TFF2 and TFF3/TFF1 for the mucosal innate immune defense against pathogenic bacteria and viruses [171]. This is in line with the observation that TFF3 expression is regulated via TLR2 by commensal bacteria [84]. In contrast, the gastric tumor suppressor TFF1, with its free thiol group, could extracellularly protect the stomach from ROS and RNS. Furthermore, an intracellular function of TFF1 is still within the limits of expectation (folding of glycoproteins, such as MUC5AC, in the ER or even regulating gene expression).
The function of TFF peptides in sites other than mucous epithelia is currently less well understood. Particularly, TFF1 might play a protective role during various inflammatory processes, even in the brain. TFF3 could also have a function during immune reactions in the CNS (expression in activated microglial cells). Furthermore, all three TFF peptides (especially TFF3) could also act as neural chemotactic factors, as they show developmental changes, particularly in the cerebellum [19]. In contrast, TFF2 seems to play a special role in the immune system, maybe as part of a matrix.
Based on these diverse molecular functions of TFF peptides, interesting clinical applications might arise in the future [172]. For example, TFF1, as well as TFF3, have been used to prevent oral mucositis after different regimes of chemotherapy [173,174]. Furthermore, the pharmacological inhibition of the TFF3 homodimer by monomerization via a synthetic drug has been reported to reduce the growth of colorectal carcinoma in vivo [175]. There are many more applications possible, as microbiota probably play a pivotal role in the development and severity of chemotherapy-induced mucositis [176]. For example, TFF1–FCGBP, as well as TFF3–FCGBP, could be components of new anti-microbial formulations, e.g., used as artificial saliva. It could be also tested whether synthetic peptides mimicking the C-terminal of TFF1, including the activated CysVII, can prevent Tff1KO mice from developing antral carcinomas.
In a further attempt, the region in TFF modules responsible for lectin activities could be defined in detail. In combination with theoretical modeling methods, a mutagenesis program could create variants with new/novel affinities for different carbohydrate moieties. Such a program should be greatly facilitated by the fact that biologically active TFF peptides can be generated, not only by recombinant technologies, but also pure chemical synthesis, where even the correct formation of disulfide bridges is possible [103,105].
Taken together, TFF peptides appear to share characteristic features with the multifunctional family of galectins [177]. For the future, many interesting medical applications concerning TFF modules can be expected, particularly for treating carcinomas and mucosal infections, as well as for supporting chemotherapy.
Acknowledgments
I thank Daniela Lorenz (Otto-von-Guericke University, Magdeburg) for her secretarial help, and Jonathan A. Lindquist (Otto-von-Guericke University, Magdeburg) for his valuable comments on the manuscript.
Abbreviations
| DUOX | Dual oxidase |
| ER | Endoplasmic reticulum |
| FCGBP | IgG Fc binding protein |
| GKN | Gastrokine |
| NOX | NADPH oxidase |
| SDS | Sodium dodecyl sulfate |
| SMC | Surface mucous cell |
| TFF | Trefoil factor family |
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflict of interest.
References
- 1.Thim L. Trefoil peptides: From structure to function. Cell. Mol. Life Sci. 1997;53:888–903. doi: 10.1007/s000180050108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ribieras S., Tomasetto C., Rio M.-C. The pS2/TFF1 trefoil factor, from basic research to clinical applications. Biochim. Biophys. Acta. 1998;1378:F61–F77. doi: 10.1016/S0304-419X(98)00016-X. [DOI] [PubMed] [Google Scholar]
- 3.Hoffmann W., Jagla W. Cell type specific expression of secretory TFF peptides: Colocalization with mucins and synthesis in the brain. Int. Rev. Cytol. 2002;213:147–188. doi: 10.1016/s0074-7696(02)13014-2. [DOI] [PubMed] [Google Scholar]
- 4.Thim L., May F.E. Structure of mammalian trefoil factors and functional insights. Cell. Mol. Life Sci. 2005;62:2956–2973. doi: 10.1007/s00018-005-5484-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kjellev S. The trefoil factor family—Small peptides with multiple functionalities. Cell. Mol. Life Sci. 2009;66:1350–1369. doi: 10.1007/s00018-008-8646-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hoffmann W. TFF peptides. In: Kastin A., editor. Handbook of Biologically Active Peptides. 2nd ed. Elsevier; Amsterdam, The Netherlands: 2013. pp. 1338–1345. [Google Scholar]
- 7.Hauser F., Roeben C., Hoffmann W. xP2, a new member of the P-domain peptide family of potential growth factors, is synthesized in Xenopus laevis skin. J. Biol. Chem. 1992;267:14451–14455. [PubMed] [Google Scholar]
- 8.Liu S.-B., He Y.-Y., Zhang Y., Lee W.-H., Qian J.-Q., Lai R., Jin Y. A novel non-lens βγ-crystallin and trefoil factor complex from amphibian skin and its functional implications. PLoS ONE. 2008;3:e1770. doi: 10.1371/journal.pone.0001770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hauser F., Gertzen E.M., Hoffmann W. Expression of spasmolysin (FIM-A.1): An integumentary mucin from Xenopus laevis. Exp. Cell Res. 1990;189:157–162. doi: 10.1016/0014-4827(90)90230-8. [DOI] [PubMed] [Google Scholar]
- 10.Hauser F., Hoffmann W. P-domains as shuffled cysteine-rich modules in integumentary mucin C.1 (FIM-C.1) from Xenopus laevis. Polydispersity and genetic polymorphism. J. Biol. Chem. 1992;267:24620–24624. [PubMed] [Google Scholar]
- 11.Hoffmann W., Hauser F. Biosynthesis of frog skin mucins: Cysteine-rich shuffled modules, polydispersities and genetic polymorphism. Comp. Biochem. Physiol. B. 1993;105:465–472. doi: 10.1016/0305-0491(93)90075-G. [DOI] [PubMed] [Google Scholar]
- 12.Stürmer R., Müller S., Hanisch F.G., Hoffmann W. Porcine gastric TFF2 is a mucus constituent and differs from pancreatic TFF2. Cell. Physiol. Biochem. 2014;33:895–904. doi: 10.1159/000358662. [DOI] [PubMed] [Google Scholar]
- 13.Hoffmann W., Jagla W., Wiede A. Molecular medicine of TFF-peptides: From gut to brain. Histol. Histopathol. 2001;16:319–334. doi: 10.14670/HH-16.319. [DOI] [PubMed] [Google Scholar]
- 14.Madsen J., Nielsen O., Tornoe I., Thim L., Holmskov U. Tissue localization of human trefoil factors 1, 2, and 3. J. Histochem. Cytochem. 2007;55:505–513. doi: 10.1369/jhc.6A7100.2007. [DOI] [PubMed] [Google Scholar]
- 15.Jagla W., Wiede A., Dietzmann K., Rutkowski K., Hoffmann W. Co-localization of TFF3 peptide and oxytocin in the human hypothalamus. FASEB J. 2000;14:1126–1131. doi: 10.1096/fasebj.14.9.1126. [DOI] [PubMed] [Google Scholar]
- 16.Fu T., Stellmacher A., Znalesniak E.B., Dieterich D.C., Kalbacher H., Hoffmann W. Tff3 is expressed in neurons and microglial cells. Cell. Physiol. Biochem. 2014;34:1912–1919. doi: 10.1159/000366389. [DOI] [PubMed] [Google Scholar]
- 17.Bernstein H.G., Dobrowolny H., Trubner K., Steiner J., Bogerts B., Hoffmann W. Differential regional and cellular distribution of TFF3 peptide in the human brain. Amino Acids. 2015;47:1053–1063. doi: 10.1007/s00726-015-1938-9. [DOI] [PubMed] [Google Scholar]
- 18.Arnold P., Rickert U., Helmers A.K., Spreu J., Schneppenheim J., Lucius R. Trefoil factor 3 shows anti-inflammatory effects on activated microglia. Cell Tissue Res. 2016;365:3–11. doi: 10.1007/s00441-016-2370-5. [DOI] [PubMed] [Google Scholar]
- 19.Hinz M., Schwegler H., Chwieralski C.E., Laube G., Linke R., Pohle W., Hoffmann W. Trefoil factor family (TFF) expression in the mouse brain and pituitary: Changes in the developing cerebellum. Peptides. 2004;25:827–832. doi: 10.1016/j.peptides.2004.01.020. [DOI] [PubMed] [Google Scholar]
- 20.Cook G.A., Familari M., Thim L., Giraud A.S. The trefoil peptides TFF2 and TFF3 are expressed in rat lymphoid tissues and participate in the immune response. FEBS Lett. 1999;456:155–159. doi: 10.1016/S0014-5793(99)00940-0. [DOI] [PubMed] [Google Scholar]
- 21.Baus-Lončar M., Kayademir T., Takaishi S., Wang T. Trefoil factor family 2 deficiency and immune response. Cell Mol. Life Sci. 2005;62:2947–2955. doi: 10.1007/s00018-005-5483-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kurt-Jones E.A., Cao L., Sandor F., Rogers A.B., Whary M.T., Nambiar P.R., Cerny A., Bowen G., Yan J., Takaishi S., et al. Trefoil family factor 2 is expressed in murine gastric and immune cells and controls both gastrointestinal inflammation and systemic immune responses. Infect. Immun. 2007;75:471–480. doi: 10.1128/IAI.02039-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kont V., Laan M., Kisand K., Merits A., Scott H.S., Peterson P. Modulation of Aire regulates the expression of tissue-restricted antigens. Mol. Immunol. 2008;45:25–33. doi: 10.1016/j.molimm.2007.05.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Dubeykovskaya Z., Si Y., Chen X., Worthley D.L., Renz B.W., Urbanska A.M., Hayakawa Y., Xu T., Westphalen C.B., Dubeykovskiy A., et al. Neural innervation stimulates splenic TFF2 to arrest myeloid cell expansion and cancer. Nat. Commun. 2016;7:10517. doi: 10.1038/ncomms10517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Jackerott M., Lee Y.C., Møllgård K., Kofod H., Jensen J., Rohleder S., Neubauer N., Gaarn L.W., Lykke J., Dodge R., et al. Trefoil factors are expressed in human and rat endocrine pancreas: Differential regulation by growth hormone. Endocrinology. 2006;147:5752–5759. doi: 10.1210/en.2006-0601. [DOI] [PubMed] [Google Scholar]
- 26.Takano T., Miyauchi A., Yoshida H., Kuma K., Amino N. High-throughput differential screening of mRNAs by serial analysis of gene expression: Decreased expression of trefoil factor 3 mRNA in thyroid follicular carcinomas. Br. J. Cancer. 2004;90:1600–1605. doi: 10.1038/sj.bjc.6601702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Vestergaard E.M., Brynskov J., Ejskjaer K., Clausen J.T., Thim L., Nexø E., Poulsen S.S. Immunoassays of human trefoil factors 1 and 2: Measured on serum from patients with inflammatory bowel disease. Scand. J. Clin. Lab Investig. 2004;64:146–156. doi: 10.1080/00365510410001176. [DOI] [PubMed] [Google Scholar]
- 28.Wright N.A. Aspects of the biology of regeneration and repair in the human gastrointestinal tract. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1998;353:925–933. doi: 10.1098/rstb.1998.0257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Rio M.-C., Chenard M.P., Wolf C., Marcellin L., Tomasetto C., Lathe R., Bellocq J.P., Chambon P. Induction of pS2 and hSP genes as markers of mucosal ulceration of the digestive tract. Gastroenterology. 1991;100:375–379. doi: 10.1016/0016-5085(91)90205-Y. [DOI] [PubMed] [Google Scholar]
- 30.Poulsom R., Wright N.A. Trefoil peptides: A newly recognized family of epithelial mucin-associated molecules. Am. J. Physiol. 1993;265:G205–G213. doi: 10.1152/ajpgi.1993.265.2.G205. [DOI] [PubMed] [Google Scholar]
- 31.Ebert M.P.A., Hoffmann J., Haeckel C., Rutkowski K., Schmid R.M., Wagner M., Adler G., Schulz H.U., Roessner A., Hoffmann W., et al. Induction of TFF1 gene expression in pancreas overexpressing transforming growth factor α. Gut. 1999;45:105–111. doi: 10.1136/gut.45.1.105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Viby N.E., Nexø E., Kissow H., Andreassen H., Clementsen P., Thim L., Poulsen S.S. Trefoil factors (TFFs) are increased in bronchioalveolar lavage fluid from patients with chronic obstructive lung disease (COPD) Peptides. 2015;63:90–95. doi: 10.1016/j.peptides.2014.09.026. [DOI] [PubMed] [Google Scholar]
- 33.Longman R.J., Poulsom R., Corfield A.P., Warren B.F., Wright N.A., Thomas M.G. Alterations in the composition of the supramucosal defense barrier in relation to disease severity of ulcerative colitis. J. Histochem. Cytochem. 2006;54:1335–1348. doi: 10.1369/jhc.5A6904.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kouznetsova I., Chwieralski C.E., Bälder R., Hinz M., Braun A., Krug N., Hoffmann W. Induced trefoil factor family 1 expression by trans-differentiating Clara cells in a murine asthma model. Am. J. Respir. Cell Mol. Biol. 2007;36:286–295. doi: 10.1165/rcmb.2006-0008OC. [DOI] [PubMed] [Google Scholar]
- 35.Znalesniak E.B., Fu T., Guttek K., Händel U., Reinhold D., Hoffmann W. Increased Cerebral Tff1 Expression in Two Murine Models of Neuroinflammation. Cell. Physiol. Biochem. 2016;39:2287–2296. doi: 10.1159/000447921. [DOI] [PubMed] [Google Scholar]
- 36.Znalesniak E.B., Fu T., Salm F., Händel U., Hoffmann W. Transcriptional Responses in the Murine Spleen after Toxoplasma gondii Infection: Inflammasome and Mucus-Associated Genes. Int. J. Mol. Sci. 2017;18:1245. doi: 10.3390/ijms18061245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Goldenring J.R., Nam K.T., Wang T.C., Mills J.C., Wright N.A. Spasmolytic polypeptide-expressing metaplasia and intestinal metaplasia: Time for reevaluation of metaplasias and the origins of gastric cancer. Gastroenterology. 2010;138:2207–2210. doi: 10.1053/j.gastro.2010.04.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Katoh M. Trefoil factors and human gastric cancer. Int. J. Mol. Med. 2003;12:3–9. doi: 10.3892/ijmm.12.1.3. [DOI] [PubMed] [Google Scholar]
- 39.Regalo G., Wright N.A., Machado J.C. Trefoil factors: From ulceration to neoplasia. Cell. Mol. Life Sci. 2005;62:2910–2915. doi: 10.1007/s00018-005-5478-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Park W.S., Oh R.R., Park J.Y., Lee J.H., Shin M.S., Kim H.S., Lee H.K., Kim Y.S., Kim S.Y., Lee S.H. Somatic mutations of the trefoil factor family 1 gene in gastric cancer. Gastroenterology. 2000;119:691–698. doi: 10.1053/gast.2000.16483. [DOI] [PubMed] [Google Scholar]
- 41.Beckler A.D., Roche J.K., Harper J.C., Petroni G., Frierson H.F., Jr., Moskaluk C.A., El-Rifai W., Powell S.M. Decreased abundance of trefoil factor 1 transcript in the majority of gastric carcinomas. Cancer. 2003;98:2184–2191. doi: 10.1002/cncr.11789. [DOI] [PubMed] [Google Scholar]
- 42.Lefebvre O., Chenard M.P., Masson R., Linares J., Dierich A., LeMeur M., Wendling C., Tomasetto C., Chambon P., Rio M.-C. Gastric mucosa abnormalities and tumorigenesis in mice lacking the pS2 trefoil protein. Science. 1996;274:259–262. doi: 10.1126/science.274.5285.259. [DOI] [PubMed] [Google Scholar]
- 43.Tomasetto C., Rio M.-C. Pleiotropic effects of Trefoil Factor 1 deficiency. Cell. Mol. Life Sci. 2005;62:2916–2920. doi: 10.1007/s00018-005-5479-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Hayakawa Y., Fox J.G., Gonda T., Worthley D.L., Muthupalani S., Wang T.C. Mouse models of gastric cancer. Cancers. 2013;5:92–130. doi: 10.3390/cancers5010092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Znalesniak E.B., Salm F., Hoffmann W. Molecular Alterations in the Stomach of Tff1-Deficient Mice: Early Steps in Antral Carcinogenesis. Int. J. Mol. Sci. 2020;21:644. doi: 10.3390/ijms21020644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Karam S.M., Tomasetto C., Rio M.-C. Amplification and invasiveness of epithelial progenitors during gastric carcinogenesis in trefoil factor 1 knockout mice. Cell Prolif. 2008;41:923–935. doi: 10.1111/j.1365-2184.2008.00562.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Soutto M., Belkhiri A., Piazuelo M.B., Schneider B.G., Peng D., Jiang A., Washington M.K., Kokoye Y., Crowe S.E., Zaika A., et al. Loss of TFF1 is associated with activation of NF-κB-mediated inflammation and gastric neoplasia in mice and humans. J. Clin. Investig. 2011;121:1753–1767. doi: 10.1172/JCI43922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Saukkonen K., Tomasetto C., Narko K., Rio M.-C., Ristimaki A. Cyclooxygenase-2 expression and effect of celecoxib in gastric adenomas of trefoil factor 1-deficient mice. Cancer Res. 2003;63:3032–3036. [PubMed] [Google Scholar]
- 49.Karam S.M., Tomasetto C., Rio M.-C. Trefoil factor 1 is required for the commitment programme of mouse oxyntic epithelial progenitors. Gut. 2004;53:1408–1415. doi: 10.1136/gut.2003.031963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Buache E., Etique N., Alpy F., Stoll I., Muckensturm M., Reina-San-Martin B., Chenard M.P., Tomasetto C., Rio M.C. Deficiency in trefoil factor 1 (TFF1) increases tumorigenicity of human breast cancer cells and mammary tumor development in TFF1-knockout mice. Oncogene. 2011;30:3261–3273. doi: 10.1038/onc.2011.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Tebbutt N.C., Giraud A.S., Inglese M., Jenkins B., Waring P., Clay F.J., Malki S., Alderman B.M., Grail D., Hollande F. Reciprocal regulation of gastrointestinal homeostasis by SHP2 and STAT-mediated trefoil gene activation in gp130 mutant mice. Nat. Med. 2002;8:1089–1097. doi: 10.1038/nm763. [DOI] [PubMed] [Google Scholar]
- 52.Wang T.C., Goldenring J.R. Inflammation intersection: gp130 balances gut irritation and stomach cancer. Nat. Med. 2002;8:1080–1082. doi: 10.1038/nm1002-1080. [DOI] [PubMed] [Google Scholar]
- 53.Farrell J.J., Taupin D., Koh T.J., Chen D., Zhao C.-M., Podolsky D.K., Wang T.C. TFF2/SP-deficient mice show decreased gastric proliferation, increased acid secretion, and increased susceptibility to NSAID injury. J. Clin. Investig. 2002;109:193–204. doi: 10.1172/JCI0212529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Baus-Loncar M., Schmid J., Lalani E.-N., Rosewell I., Goodlad R.A., Stamp G.W.H., Blin N., Kayademir T. Trefoil factor 2 (Tff2) deficiency in murine digestive tract influences the immune system. Cell. Physiol. Biochem. 2005;16:31–42. doi: 10.1159/000087729. [DOI] [PubMed] [Google Scholar]
- 55.Fox J.G., Rogers A.B., Whary M.T., Ge Z., Ohtani M., Jones E.K., Wang T.C. Accelerated progression of gastritis to dysplasia in the pyloric antrum of TFF2 -/- C57BL6 x Sv129 Helicobacter pylori-infected mice. Am. J. Pathol. 2007;171:1520–1528. doi: 10.2353/ajpath.2007.070249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Shah A.A., Mihalj M., Ratkay I., Lubka-Pathak M., Balogh P., Klingel K., Bohn E., Blin N., Baus-Lončar M. Increased susceptibility to Yersinia enterocolitica Infection of Tff2 deficient mice. Cell. Physiol. Biochem. 2012;30:853–862. doi: 10.1159/000341463. [DOI] [PubMed] [Google Scholar]
- 57.Judd L.M., Chalinor H.V., Walduck A., Pavlic D.I., Dabritz J., Dubeykovskaya Z., Wang T.C., Menheniott T.R., Giraud A.S. TFF2 deficiency exacerbates weight loss and alters immune cell and cytokine profiles in DSS colitis, and this cannot be rescued by wild-type bone marrow. Am. J. Physiol. Gastrointest. Liver Physiol. 2015;308:G12–G24. doi: 10.1152/ajpgi.00172.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.McCarthy A.J., Birchenough G.M.H., Taylor P.W. Loss of Trefoil Factor 2 Sensitizes Rat Pups to Systemic Infection with the Neonatal Pathogen Escherichia coli K1. Infect. Immun. 2019;87:e00878-18. doi: 10.1128/IAI.00878-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Birchenough G.M., Johansson M.E., Stabler R.A., Dalgakiran F., Hansson G.C., Wren B.W., Luzio J.P., Taylor P.W. Altered innate defenses in the neonatal gastrointestinal tract in response to colonization by neuropathogenic Escherichia coli. Infect. Immun. 2013;81:3264–3275. doi: 10.1128/IAI.00268-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Mashimo H., Wu D.C., Podolsky D.K., Fishman M.C. Impaired defense of intestinal mucosa in mice lacking intestinal trefoil factor. Science. 1996;274:262–265. doi: 10.1126/science.274.5285.262. [DOI] [PubMed] [Google Scholar]
- 61.Beck P.L., Wong J.F., Li Y., Swaminathan S., Xavier R.J., Devaney K.L., Podolsky D.K. Chemotherapy- and radiotherapy-induced intestinal damage is regulated by intestinal trefoil factor. Gastroenterology. 2004;126:796–808. doi: 10.1053/j.gastro.2003.12.004. [DOI] [PubMed] [Google Scholar]
- 62.Blaschke K. M.D. Thesis. Otto-von-Guericke University; Magdeburg, Germany: 2010. Vergleichende neurobiologische Untersuchungen von TFF3-defizienten Mäusen und Wildtyptieren. [Google Scholar]
- 63.Dignass A., Lynch-Devaney K., Kindon H., Thim L., Podolsky D.K. Trefoil peptides promote epithelial migration through a transforming growth factor β-independent pathway. J. Clin. Investig. 1994;94:376–383. doi: 10.1172/JCI117332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Playford R.J., Marchbank T., Chinery R., Evison R., Pignatelli M., Boulton R.A., Thim L., Hanby A.M. Human spasmolytic polypeptide is a cytoprotective agent that stimulates cell migration. Gastroenterology. 1995;108:108–116. doi: 10.1016/0016-5085(95)90014-4. [DOI] [PubMed] [Google Scholar]
- 65.Göke M.N., Cook J.R., Kunert K.S., Fini M.E., Gipson I.K., Podolsky D.K. Trefoil peptides promote restitution of wounded corneal epithelial cells. Exp. Cell. Res. 2001;264:337–344. doi: 10.1006/excr.2000.5116. [DOI] [PubMed] [Google Scholar]
- 66.Schulze U., Hampel U., Sel S., Contreras-Ruiz L., Schicht M., Dieckow J., Diebold Y., Paulsen F. Trefoil factor family peptide 3 (TFF3) is upregulated under experimental conditions similar to dry eye disease and supports corneal wound healing effects in vitro. Investig. Ophthalmol. Vis. Sci. 2014;55:3037–3042. doi: 10.1167/iovs.13-13423. [DOI] [PubMed] [Google Scholar]
- 67.Graness A., Chwieralski C.E., Reinhold D., Thim L., Hoffmann W. Protein kinase C and ERK activation are required for TFF-peptide-stimulated bronchial epithelial cell migration and tumor necrosis factor-α-induced interleukin-6 (IL-6) and IL-8 secretion. J. Biol. Chem. 2002;277:18440–18446. doi: 10.1074/jbc.M200468200. [DOI] [PubMed] [Google Scholar]
- 68.Fu T., Kalbacher H., Hoffmann W. TFF1 is differentially expressed in stationary and migratory rat gastric epithelial cells (RGM-1) after in vitro wounding: Influence of TFF1 RNA interference on cell migration. Cell. Physiol. Biochem. 2013;32:997–1010. doi: 10.1159/000354501. [DOI] [PubMed] [Google Scholar]
- 69.Storesund T., Hayashi K., Kolltveit K.M., Bryne M., Schenck K. Salivary trefoil factor 3 enhances migration of oral keratinocytes. Eur. J. Oral Sci. 2008;116:135–140. doi: 10.1111/j.1600-0722.2007.00516.x. [DOI] [PubMed] [Google Scholar]
- 70.Guppy N.J., El-Bahrawy M.E., Kocher H.M., Fritsch K., Qureshi Y.A., Poulsom R., Jeffery R.E., Wright N.A., Otto W.R., Alison M.R. Trefoil factor family peptides in normal and diseased human pancreas. Pancreas. 2012;41:888–896. doi: 10.1097/MPA.0b013e31823c9ec5. [DOI] [PubMed] [Google Scholar]
- 71.Prest S.J., May F.E., Westley B.R. The estrogen-regulated protein, TFF1, stimulates migration of human breast cancer cells. FASEB J. 2002;16:592–594. doi: 10.1096/fj.01-0498fje. [DOI] [PubMed] [Google Scholar]
- 72.Chwieralski C.E., Schnurra I., Thim L., Hoffmann W. Epidermal growth factor and trefoil factor family 2 synergistically trigger chemotaxis on BEAS-2B cells via different signaling cascades. Am. J. Cell Mol. Biol. 2004;31:528–537. doi: 10.1165/rcmb.2003-0433OC. [DOI] [PubMed] [Google Scholar]
- 73.Chinery R., Playford R.J. Combined intestinal trefoil factor and epidermal growth factor is prophylactic against Indomethacin-Induced gastric damage in the rat. Clin. Sci. 1995;88:401–403. doi: 10.1042/cs0880401. [DOI] [PubMed] [Google Scholar]
- 74.Rodrigues S., Van Aken E., Van Bocxlaer S., Attoub S., Nguyen Q.D., Bruyneel E., Westley B.R., May F.E., Thim L., Mareel M., et al. Trefoil peptides as proangiogenic factors in vivo and in vitro: Implication of cyclooxygenase-2 and EGF receptor signaling. FASEB J. 2003;17:7–16. doi: 10.1096/fj.02-0201com. [DOI] [PubMed] [Google Scholar]
- 75.Lalani E.N., Williams R., Jayaram Y., Gilbert C., Chaudhary K.S., Siu L.S., Koumarianou A., Playford R., Stamp G.W. Trefoil factor-2, human spasmolytic polypeptide, promotes branching morphogenesis in MCF-7 cells. Lab. Investig. 1999;79:537–546. [PubMed] [Google Scholar]
- 76.Taupin D.R., Kinoshita K., Podolsky D.K. Intestinal trefoil factor confers colonic epithelial resistance to apoptosis. Proc. Natl. Acad. Sci. USA. 2000;97:799–804. doi: 10.1073/pnas.97.2.799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Bossenmeyer-Pourié C., Kannan R., Ribieras S., Wendling C., Stoll I., Thim L., Tomasetto C., Rio M.-C. The trefoil factor 1 participates in gastrointestinal cell differentiation by delaying G1-S phase transition and reducing apoptosis. J. Cell Biol. 2002;157:761–770. doi: 10.1083/jcb200108056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Siu L.S., Romanska H., Abel P.D., Baus-Lončar M., Kayademir T., Stamp G.W., Lalani E.-N. TFF2 (trefoil family factor2) inhibits apoptosis in breast and colorectal cancer cell lines. Peptides. 2004;25:855–863. doi: 10.1016/j.peptides.2003.11.023. [DOI] [PubMed] [Google Scholar]
- 79.Rösler S., Haase T., Claassen H., Schulze U., Schicht M., Riemann D., Brandt J., Wohlrab D., Müller-Hilke B., Goldring M.B., et al. Trefoil factor 3 is induced during degenerative and inflammatory joint disease, activates matrix metalloproteinases, and enhances apoptosis of articular cartilage chondrocytes. Arthritis Rheumatol. 2010;62:815–825. doi: 10.1002/art.27295. [DOI] [PubMed] [Google Scholar]
- 80.Cho S.Y., Klemke R.L. Extracellular-regulated kinase activation and CAS/Crk coupling regulate cell migration and suppress apoptosis during invasion of the extracellular matrix. J. Cell Biol. 2000;149:223–236. doi: 10.1083/jcb.149.1.223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Taupin D., Podolsky D.K. Trefoil factors: Initiators of mucosal healing. Nat. Rev. Mol. Cell. Biol. 2003;4:721–732. doi: 10.1038/nrm1203. [DOI] [PubMed] [Google Scholar]
- 82.Hoffmann W. Trefoil factors TFF (trefoil factor family) peptide-triggered signals promoting mucosal restitution. Cell. Mol. Life Sci. 2005;62:2932–2938. doi: 10.1007/s00018-005-5481-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Vandenbroucke K., Hans W., Van Huysse J., Neirynck S., Demetter P., Remaut E., Rottiers P., Steidler L. Active delivery of trefoil factors by genetically modified Lactococcus lactis prevents and heals acute colitis in mice. Gastroenterology. 2004;127:502–513. doi: 10.1053/j.gastro.2004.05.020. [DOI] [PubMed] [Google Scholar]
- 84.Podolsky D.K., Gerken G., Eyking A., Cario E. Colitis-associated variant of TLR2 causes impaired mucosal repair because of TFF3 deficiency. Gastroenterology. 2009;137:209–220. doi: 10.1053/j.gastro.2009.03.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Otto W.R., Thim L. Trefoil factor family-interacting proteins. Cell. Mol. Life Sci. 2005;62:2939–2946. doi: 10.1007/s00018-005-5482-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Thim L., Mørtz E. Isolation and characterization of putative trefoil peptide receptors. Regul. Pept. 2000;90:61–68. doi: 10.1016/S0167-0115(00)00110-5. [DOI] [PubMed] [Google Scholar]
- 87.Madsen J., Sorensen G.L., Nielsen O., Tornøe I., Thim L., Fenger C., Mollenhauer J., Holmskov U. A variant form of the human deleted in malignant brain tumor 1 (DMBT1) gene shows increased expression in inflammatory bowel diseases and interacts with dimeric trefoil factor 3 (TFF3) PLoS ONE. 2013;8:e64441. doi: 10.1371/journal.pone.0064441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Leito J.T., Ligtenberg A.J., van Houdt M., van den Berg T.K., Wouters D. The bacteria binding glycoprotein salivary agglutinin (SAG/gp340) activates complement via the lectin pathway. Mol. Immunol. 2011;49:185–190. doi: 10.1016/j.molimm.2011.08.010. [DOI] [PubMed] [Google Scholar]
- 89.Dubeykovskaya Z., Dubeykovskiy A., Solal-Cohen J., Wang T.C. Secreted trefoil factor 2 activates the CXCR4 receptor in epithelial and lymphocytic cancer cell lines. J. Biol. Chem. 2009;284:3650–3662. doi: 10.1074/jbc.M804935200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Dieckow J., Brandt W., Hattermann K., Schob S., Schulze U., Mentlein R., Ackermann P., Sel S., Paulsen F.P. CXCR4 and CXCR7 mediate TFF3-Induced cell migration independently from the ERK1/2 signaling pathway. Investig. Ophthalmol. Sci. 2016;57:56–65. doi: 10.1167/iovs.15-18129. [DOI] [PubMed] [Google Scholar]
- 91.Hoffmann W. Trefoil factor family (TFF) peptides and chemokine receptors: A promising relationship. J. Med. Chem. 2009;52:6505–6510. doi: 10.1021/jm9008136. [DOI] [PubMed] [Google Scholar]
- 92.Engevik K.A., Hanyu H., Matthis A.L., Zhang T., Frey M.R., Oshima Y., Aihara E., Montrose M.H. Trefoil factor 2 activation of CXCR4 requires calcium mobilization to drive epithelial repair in gastric organoids. J. Physiol. 2019;597:2673–2690. doi: 10.1113/JP277259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Zhang Y., Yu G., Wang Y., Xiang Y., Gao Q., Jiang P., Zhang J., Lee W., Zhang Y. Activation of protease-activated receptor (PAR) 1 by frog trefoil factor (TFF) 2 and PAR4 by human TFF2. Cell. Mol. Live Sci. 2011;68:3771–3780. doi: 10.1007/s00018-011-0678-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Barrera Roa G.J., Sanchez Tortolero G.S. Trefoil factor 3 (TFF3) from human breast milk activates PAR-2 receptors, of the intestinal epithelial cells HT-29, regulating cytokines and defensins. Bratisl. Med. J. 2016;117:332–339. doi: 10.4149/BLL_2016_066. [DOI] [PubMed] [Google Scholar]
- 95.Riemer J., Bulleid N., Herrmann J.M. Disulfide formation in the ER and mitochondria: Two solutions to a common process. Science. 2009;324:1284–1287. doi: 10.1126/science.1170653. [DOI] [PubMed] [Google Scholar]
- 96.Marchbank T., Westley B.R., May F.E.B., Calnan D.P., Playford R.J. Dimerization of human pS2 (TFF1) plays a key role in its protective/healing effects. J. Pathol. 1998;185:153–158. doi: 10.1002/(SICI)1096-9896(199806)185:2<153::AID-PATH87>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- 97.Calnan D.P., Westley B.R., May F.E.B., Floyd D.N., Marchbank T., Playford R.J. The trefoil peptide TFF1 inhibits the growth of the human gastric adenocarcinoma cell line AGS. J. Pathol. 1999;188:312–317. doi: 10.1002/(SICI)1096-9896(199907)188:3<312::AID-PATH360>3.0.CO;2-P. [DOI] [PubMed] [Google Scholar]
- 98.Kinoshita K., Taupin D.R., Itoh H., Podolsky D.K. Distinct pathways of cell migration and antiapoptotic response to epithelial injury: Structure-function analysis of human intestinal trefoil factor. Mol. Cell. Biol. 2000;20:4680–4690. doi: 10.1128/MCB.20.13.4680-4690.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Reeves E.P., Ali T., Leonard P., Hearty S., O’Kennedy R., May F.E.B., Westley B.R., Josenhans C., Rust M., Suerbaum S., et al. Helicobacter pylori lipopolysaccharide interacts with TFF1 in a pH-dependent manner. Gastroenterology. 2008;135:2043–2054. doi: 10.1053/j.gastro.2008.08.049. [DOI] [PubMed] [Google Scholar]
- 100.Westley B.R., Griffin S.M., May F.E.B. Interaction between TFF1, a gastric tumor suppressor trefoil protein, and TFIZ1, a brichos domain-containing protein with homology to SP-C. Biochemistry. 2005;44:7967–7975. doi: 10.1021/bi047287n. [DOI] [PubMed] [Google Scholar]
- 101.Albert T.K., Laubinger W., Müller S., Hanisch F.-G., Kalinski T., Meyer F., Hoffmann W. Human intestinal TFF3 forms disulfide-linked heteromers with the mucus-associated FCGBP protein and is released by hydrogen sulfide. J. Proteome Res. 2010;9:3108–3117. doi: 10.1021/pr100020c. [DOI] [PubMed] [Google Scholar]
- 102.Houben T., Harder S., Schlüter H., Kalbacher H., Hoffmann W. Different Forms of TFF3 in the Human Saliva: Heterodimerization with IgG Fc Binding Protein (FCGBP) Int. J. Mol. Sci. 2019;20:5000. doi: 10.3390/ijms20205000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Heuer J., Heuer F., Stürmer R., Harder S., Schlüter H., Braga Emidio N., Muttenthaler M., Jechorek D., Meyer F., Hoffmann W. The Tumor Suppressor TFF1 Occurs in Different Forms and Interacts with Multiple Partners in the Human Gastric Mucus Barrier: Indications for Diverse Protective Functions. Int. J. Mol. Sci. 2020;21:2508. doi: 10.3390/ijms21072508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Dolan B., Naughton J., Tegtmeyer N., May F.E.B., Clyne M. The interaction of Helicobacter pylori with the adherent mucus gel layer secreted by polarized HT29-MTX-E12 cells. PLoS ONE. 2012;7:e47300. doi: 10.1371/journal.pone.0047300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Braga Emidio N., Baik H., Lee D., Stürmer R., Heuer J., Elliott A.G., Blaskovich M.A., Haupenthal K., Tegtmeyer N., Hoffmann W., et al. Chemical synthesis of human trefoil factor 1 (TFF1) and its homodimer provides novel insights into their mechanisms of action. Chem. Commun. 2020;56:6420–6423. doi: 10.1039/D0CC02321C. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Kouznetsova I., Laubinger W., Kalbacher H., Kalinski T., Meyer F., Roessner A., Hoffmann W. Biosynthesis of gastrokine-2 in the human gastric mucosa: Restricted spatial expression along the antral gland axis and differential interaction with TFF1, TFF2 and mucins. Cell. Physiol. Biochem. 2007;20:899–908. doi: 10.1159/000110450. [DOI] [PubMed] [Google Scholar]
- 107.Hanisch F.-G., Bonar D., Schloerer N., Schroten H. Human trefoil factor 2 is a lectin that binds α-GlcNAc-capped mucin glycans with antibiotic activity against Helicobacter pylori. J. Biol. Chem. 2014;289:27363–27375. doi: 10.1074/jbc.M114.597757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Hoffmann W. TFF2, a MUC6-binding lectin stabilizing the gastric mucus barrier and more. Int. J. Oncol. 2015;47:806–816. doi: 10.3892/ijo.2015.3090. [DOI] [PubMed] [Google Scholar]
- 109.Stürmer R., Harder S., Schlüter H., Hoffmann W. Commercial porcine gastric mucin preparations, also used as artificial saliva, are a rich source for the lectin TFF2: In vitro binding studies. ChemBioChem. 2018;19:2598–2608. doi: 10.1002/cbic.201800622. [DOI] [PubMed] [Google Scholar]
- 110.Heuer F., Stürmer R., Heuer J., Kalinski T., Lemke A., Meyer F., Hoffmann W. Different Forms of TFF2, A Lectin of the Human Gastric Mucus Barrier: In Vitro Binding Studies. Int. J. Mol. Sci. 2019;20:5871. doi: 10.3390/ijms20235871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Stürmer R., Reising J., Hoffmann W. Trefoil Factor Family (TFF) Modules Are Characteristic Constituents of Separate Mucin Complexes in the Xenopus laevis Integumentary Mucus: In Vitro Binding Studies with FIM-A. 1. Int. J. Mol. Sci. 2020;21:2400. doi: 10.3390/ijms21072400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Sacchettini J.C., Baum L.G., Brewer C.F. Multivalent protein-carbohydrate interactions. A new paradigm for supermolecular assembly and signal transduction. Biochemistry. 2001;40:3009–3015. doi: 10.1021/bi002544j. [DOI] [PubMed] [Google Scholar]
- 113.Rabinovich G.A., Toscano M.A., Jackson S.S., Vasta G.R. Functions of cell surface galectin-glycoprotein lattices. Curr. Opin. Struct. Biol. 2007;17:513–520. doi: 10.1016/j.sbi.2007.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Newton J., Allen A., Westley B.R., May F.E.B. The human trefoil peptide, TFF1, is present in different molecular forms that are intimately associated with mucus in normal stomach. Gut. 2000;46:312–320. doi: 10.1136/gut.46.3.312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Stürmer R., Reising J., Hoffmann W. The TFF Peptides xP1 and xP4 Appear in Distinctive Forms in the Xenopus laevis Gastric Mucosa: Indications for Different Protective Functions. Int. J. Mol. Sci. 2019;20:6052. doi: 10.3390/ijms20236052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Schwarz H., Hoffmann W. Subcellular Localization of the TFF Peptides xP1 and xP4 in the Xenopus laevis Gastric/Esophageal Mucosa: Different Secretion Modes Reflecting Diverse Protective Functions. Int. J. Mol. Sci. 2020;21:761. doi: 10.3390/ijms21030761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Kouznetsova I., Kalinski T., Meyer F., Hoffmann W. Self-renewal of the human gastric epithelium: New insights from expression profiling using laser microdissection. Mol. BioSyst. 2011;7:1105–1112. doi: 10.1039/c0mb00233j. [DOI] [PubMed] [Google Scholar]
- 118.Fra A.M., Fagioli C., Finazzi D., Sitia R., Alberini C.M. Quality control of ER synthesized proteins: An exposed thiol group as a three-way switch mediating assembly, retention and degradation. EMBO J. 1993;12:4755–4761. doi: 10.1002/j.1460-2075.1993.tb06164.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Reddy P., Sparvoli A., Fagioli C., Fassina G., Sitia R. Formation of reversible disulfide bonds with the protein matrix of the endoplasmic reticulum correlates with the retention of unassembled Ig light chains. EMBO J. 1996;15:2077–2085. doi: 10.1002/j.1460-2075.1996.tb00561.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Gilbert H.F. Molecular and cellular aspects of thiol-disulfide exchange. Adv. Enzymol. Relat. Areas Mol. Biol. 1990;63:69–172. doi: 10.1002/9780470123096.ch2. [DOI] [PubMed] [Google Scholar]
- 121.Poole L.B. The basics of thiols and cysteines in redox biology and chemistry. Free Radic. Biol. Med. 2015;80:148–157. doi: 10.1016/j.freeradbiomed.2014.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Hoffmann W., Hauser F. The P-domain or trefoil motif: A role in renewal and pathology of mucous epithelia? Trends Biochem. Sci. 1993;18:239–243. doi: 10.1016/0968-0004(93)90170-R. [DOI] [PubMed] [Google Scholar]
- 123.Torres L.-F., Karam S.M., Wendling C., Chenard M.-P., Kershenobich D., Tomasetto C., Rio M.-C. Trefoil factor 1 (TFF1/pS2) deficiency activates the unfolded protein response. Mol. Med. 2002;8:273–282. doi: 10.1007/BF03402153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Lindquist J.A., Hämmerling G.J., Trowsdale J. ER60/ERp57 forms disulfide-bonded intermediates with MHC class I heavy chain. FASEB J. 2001;15:1448–1450. doi: 10.1096/fj.00-0720fje. [DOI] [PubMed] [Google Scholar]
- 125.Shishkin S., Eremina L., Kovalev L., Kovaleva M. AGR2, ERp57/GRP58, and some other human protein disulfide isomerases. Biochemistry. 2013;78:1415–1430. doi: 10.1134/S000629791313004X. [DOI] [PubMed] [Google Scholar]
- 126.Giles G.I., Jacob C. Reactive sulfur species: An emerging concept in oxidative stress. Biol. Chem. 2002;383:375–388. doi: 10.1515/BC.2002.042. [DOI] [PubMed] [Google Scholar]
- 127.Ying J., Clavreul N., Sethuraman M., Adachi T., Cohen R.A. Thiol oxidation in signaling and response to stress: Detection and quantification of physiological and pathophysiological thiol modifications. Free Radic. Biol. Med. 2007;43:1099–1108. doi: 10.1016/j.freeradbiomed.2007.07.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Colovic M.B., Vasic V.M., Djuric D.M., Krstic D.Z. Sulphur-containing amino acids: Protective role against free radicals and heavy metals. Curr. Med. Chem. 2018;25:324–335. doi: 10.2174/0929867324666170609075434. [DOI] [PubMed] [Google Scholar]
- 129.Suzuki H., Nishizawa T., Tsugawa H., Mogami S., Hibi T. Roles of oxidative stress in stomach disorders. J. Clin. Biochem. Nutr. 2012;50:35–39. doi: 10.3164/jcbn.11-115SR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Hoffmann W. Current status on stem cells and cancers of the gastric epithelium. Int. J. Mol. Sci. 2015;16:19153–19169. doi: 10.3390/ijms160819153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Phillipson M., Atuma C., Henriksnas J., Holm L. The importance of mucus layers and bicarbonate transport in preservation of gastric juxtamucosal pH. Am. J. Physiol. Gastrointest. Liver Physiol. 2002;282:G211–G219. doi: 10.1152/ajpgi.00223.2001. [DOI] [PubMed] [Google Scholar]
- 132.Bae Y.S., Choi M.K., Lee W.J. Dual oxidase in mucosal immunity and host-microbe homeostasis. Trends Immunol. 2010;31:278–287. doi: 10.1016/j.it.2010.05.003. [DOI] [PubMed] [Google Scholar]
- 133.Grasberger H., El-Zaatari M., Dang D.T., Merchant J.L. Dual oxidases control release of hydrogen peroxide by the gastric epithelium to prevent Helicobacter felis infection and inflammation in mice. Gastroenterology. 2013;145:1045–1054. doi: 10.1053/j.gastro.2013.07.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Bedard K., Krause K.-H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007;87:245–313. doi: 10.1152/physrev.00044.2005. [DOI] [PubMed] [Google Scholar]
- 135.Björne H., Petersson J., Phillipson M., Weitzberg E., Holm L., Lundberg J.O. Nitrite in saliva increases gastric mucosal blood flow and mucus thicknes. J. Clin. Investig. 2004;113:106–114. doi: 10.1172/JCI19019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Foster M.W., McMahon T.J., Stamler J.S. S-nitrosylation in health and disease. Trends Mol. Med. 2003;9:160–168. doi: 10.1016/S1471-4914(03)00028-5. [DOI] [PubMed] [Google Scholar]
- 137.Lugrin J., Rosenblatt-Velin N., Parapanov R., Liaudet L. The role of oxidative stress during inflammatory processes. Biol. Chem. 2014;395:203–230. doi: 10.1515/hsz-2013-0241. [DOI] [PubMed] [Google Scholar]
- 138.Pacher P., Beckman J.S., Liaudet L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007;87:315–424. doi: 10.1152/physrev.00029.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Gu Z., Kaul M., Yan B., Kridel S.J., Cui J., Strongin A., Smith J.W., Liddington R.C., Lipton S.A. S-nitrosylation of matrix metalloproteinases: Signaling pathway to neuronal cell death. Science. 2002;297:1186–1190. doi: 10.1126/science.1073634. [DOI] [PubMed] [Google Scholar]
- 140.Tosco A., Monti M.C., Fontanella B., Montefusco S., D’Andrea L., Ziaco B., Baldantoni D., Rio M.-C., Marzullo L. Copper binds the carboxy-terminus of trefoil protein 1 (TFF1), favoring its homodimerization and motogenic activity. Cell. Mol. Life Sci. 2010;67:1943–1955. doi: 10.1007/s00018-010-0309-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Wright N.A., Poulsom R., Stamp G., Van Noorden S., Sarraf C., Elia G., Ahnen D., Jeffery R., Longcroft J., Pike C. Trefoil peptide gene expression in gastrointestinal epithelial cells in inflammatory bowel disease. Gastroenterology. 1993;104:12–20. doi: 10.1016/0016-5085(93)90830-6. [DOI] [PubMed] [Google Scholar]
- 142.Ota H., Hayama M., Momose M., El-Zimaity H.M., Matsuda K., Sano K., Maruta F., Okumura N., Katsuyama T. Co-localization of TFF2 with gland mucous cell mucin in gastric mucous cells and in extracellular mucous gel adherent to normal and damaged gastric mucosa. Histochem. Cell Biol. 2006;126:617–625. doi: 10.1007/s00418-006-0197-y. [DOI] [PubMed] [Google Scholar]
- 143.Oinuma T., Ide S., Kawano J., Suganuma T. Purification and immunohistochemistry of Griffonia simplicifolia agglutinin-II-binding mucus glycoprotein in rat stomach. Glycobiology. 1994;4:469–475. doi: 10.1093/glycob/4.4.469. [DOI] [PubMed] [Google Scholar]
- 144.Ishihara K., Kurihara M., Goso Y., Urata T., Ota H., Katsuyama T., Hotta K. Peripheral α -linked N-acetylglucosamine on the carbohydrate moiety of mucin derived from mammalian gastric gland mucous cells: Epitope recognized by a newly characterized monoclonal antibody. Biochem. J. 1996;318:409–416. doi: 10.1042/bj3180409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Lang T., Klasson S., Larsson E., Johansson M.E., Hansson G.C., Samuelsson T. Searching the Evolutionary Origin of Epithelial Mucus Protein Components-Mucins and FCGBP. Mol. Biol. Evol. 2016;33:1921–1936. doi: 10.1093/molbev/msw066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Kawakubo M., Ito Y., Okimura Y., Kobayashi M., Sakura K., Kasama S., Fukuda M.N., Fukuda M., Katsuyama T., Nakayama J. Natural antibiotic function of a human gastric mucin against Helicobacter pylori infection. Science. 2004;305:1003–1006. doi: 10.1126/science.1099250. [DOI] [PubMed] [Google Scholar]
- 147.Lee H., Wang P., Hoshino H., Ito Y., Kobayashi M., Nakayama J., Seeberger P.H., Fukuda M. α1,4GlcNAc-capped mucin-type O-glycan inhibits cholesterol α-glucosyltransferase from Helicobacter pylori and suppresses H. pylori growth. Glycobiology. 2008;18:549–558. doi: 10.1093/glycob/cwn037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Van den Brink G.R., Tytgat K.M.A.J., Van der Hulst R.W.M., Van der Loos C.M., Einerhand A.W., Büller H.A., Dekker J. H pylori colocalises with MUC5AC in the human stomach. Gut. 2000;46:601–607. doi: 10.1136/gut.46.5.601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Karasawa F., Shiota A., Goso Y., Kobayashi M., Sato Y., Masumoto J., Fujiwara M., Yokosawa S., Muraki T., Miyagawa S. Essential role of gastric gland mucin in preventing gastric cancer in mice. J. Clin. Investig. 2012;122:923–934. doi: 10.1172/JCI59087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Hanisch F.-G., Ragge H., Kalinski T., Meyer F., Kalbacher H., Hoffmann W. Human gastric TFF2 peptide contains an N-linked fucosylated N,N’-diacetyllactosediamine (LacdiNAc) oligosaccharide. Glycobiology. 2013;23:2–11. doi: 10.1093/glycob/cws131. [DOI] [PubMed] [Google Scholar]
- 151.Semple J.I., Newton J.L., Westley B.R., May F.E.B. Dramatic diurnal variation in the concentration of the human trefoil peptide TFF2 in gastric juice. Gut. 2001;48:648–655. doi: 10.1136/gut.48.5.648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Rossez Y., Gosset P., Boneca I.G., Magalhães A., Ecobichon C., Reis C.A., Cieniewski-Bernard C., Joncquel Chevalier Curt M., Léonard R., Maes E. The lacdiNAc-specific adhesin LabA mediates adhesion of Helicobacter pylori to human gastric mucosa. J. Infect. Dis. 2014;210:1286–1295. doi: 10.1093/infdis/jiu239. [DOI] [PubMed] [Google Scholar]
- 153.Phillipson M., Johansson M.E., Henriksnas J., Petersson J., Gendler S.J., Sandler S., Persson A.E., Hansson G.C., Holm L. The gastric mucus layers: Constituents and regulation of accumulation. Am. J. Physiol. Gastrointest. Liver Physiol. 2008;295:G806–G812. doi: 10.1152/ajpgi.90252.2008. [DOI] [PubMed] [Google Scholar]
- 154.Ho S.B., Takamura K., Anway R., Shekels L.L., Toribara N.W., Ota H. The adherent gastric mucous layer is composed of alternating layers of MUC5AC and MUC6 mucin proteins. Dig. Dis. Sci. 2004;49:1598–1606. doi: 10.1023/B:DDAS.0000043371.12671.98. [DOI] [PubMed] [Google Scholar]
- 155.Paulo J.A., Lee L.S., Wu B., Repas K., Banks P.A., Conwell D.L., Steen H. Proteomic analysis of endoscopically (ePFT) collected gastroduodenal fluid using in-gel tryptic digestion followed by LC-MS/MS. Proteomics Clin. Appl. 2010;4:715–725. doi: 10.1002/prca.201000018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Thim L., Madsen F., Poulsen S.S. Effect of trefoil factors on the viscoelastic properties of mucus gels. Eur. J. Clin. Investig. 2002;32:519–527. doi: 10.1046/j.1365-2362.2002.01014.x. [DOI] [PubMed] [Google Scholar]
- 157.Menheniott T.R., O’Connor L., Chionh Y.T., Däbritz J., Scurr M., Rollo B.N., Ng G.Z., Jacobs S., Catubig A., Kurklu B. Loss of gastrokine-2 drives premalignant gastric inflammation and tumor progression. J. Cin. Investig. 2016;126:1383–1400. doi: 10.1172/JCI82655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Harada N., Iijima S., Kobayashi K., Yoshida T., Brown W.R., Hibi T., Oshima A., Morikawa M. Human IgGFc binding protein (FcγBP) in colonic epithelial cells exhibits mucin-like structure. J. Biol. Chem. 1997;272:15232–15241. doi: 10.1074/jbc.272.24.15232. [DOI] [PubMed] [Google Scholar]
- 159.Kobayashi K., Ogata H., Morikawa M., Iijima S., Harada N., Yoshida T., Brown W., Inoue N., Hamada Y., Ishii H. Distribution and partial characterisation of IgG Fc binding protein in various mucin producing cells and body fluids. Gut. 2002;51:169–176. doi: 10.1136/gut.51.2.169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Kouznetsova I., Gerlach K.L., Zahl C., Hoffmann W. Expression analysis of human salivary glands by laser microdissection: Differences between submandibular and labial glands. Cell. Physiol. Biochem. 2010;26:375–382. doi: 10.1159/000320561. [DOI] [PubMed] [Google Scholar]
- 161.Li C., Wang R., Su B., Luo Y., Terhune J., Beck B., Peatman E. Evasion of mucosal defenses during Aeromonas hydrophila infection of channel catfish (Ictalurus punctatus) skin. Dev. Comp. Immunol. 2013;39:447–455. doi: 10.1016/j.dci.2012.11.009. [DOI] [PubMed] [Google Scholar]
- 162.Wang Y., Liu Y., Liu H., Zhang Q., Song H., Tang J., Fu J., Wang X. FcGBP was upregulated by HPV infection and correlated to longer survival time of HNSCC patients. Oncotarget. 2017;8:86503–86514. doi: 10.18632/oncotarget.21220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Schwartz J.L. Fcgbp—A Potential Viral Trap in RV144. Open AIDS J. 2014;8:21–24. doi: 10.2174/1874613601408010021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Zhen G., Park S.W., Nguyenvu L.T., Rodriguez M.W., Barbeau R., Paquet A.C., Erle D.J. IL-13 and epidermal growth factor receptor have critical but distinct roles in epithelial cell mucin production. Am. J. Respir. Cell. Mol. Biol. 2007;36:244–253. doi: 10.1165/rcmb.2006-0180OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Jilek A., Mollay C., Tippelt C., Grassi J., Mignogna G., Müllegger J., Sander V., Fehrer C., Barra D., Kreil G. Biosynthesis of a D-amino acid in peptide linkage by an enzyme from frog skin secretions. Proc. Natl. Acad. Sci. USA. 2005;102:4235–4239. doi: 10.1073/pnas.0500789102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Nieuw Amerongen A.V., Veerman E.C. Saliva—The defender of the oral cavity. Oral Dis. 2002;8:12–22. doi: 10.1034/j.1601-0825.2002.1o816.x. [DOI] [PubMed] [Google Scholar]
- 167.Rossez Y., Coddeville B., Elass E., Quinchon J.F., Vidal O., Corfield A.P., Gosset P., Lacroix J.M., Michalski J.C., Robbe-Masselot C. Interaction between DMBT1 and galectin 3 is modulated by the structure of the oligosaccharides carried by DMBT1. Biochimie. 2011;93:593–603. doi: 10.1016/j.biochi.2010.12.002. [DOI] [PubMed] [Google Scholar]
- 168.Johansson M.E., Thomsson K.A., Hansson G.C. Proteomic analyses of the two mucus layers of the colon barrier reveal that their main component, the Muc2 mucin, is strongly bound to the Fcgbp protein. J. Proteome Res. 2009;8:3549–3557. doi: 10.1021/pr9002504. [DOI] [PubMed] [Google Scholar]
- 169.Johansson M.E., Gustafsson J.K., Sjoberg K.E., Petersson J., Holm L., Sjovall H., Hansson G.C. Bacteria penetrate the inner mucus layer before inflammation in the dextran sulfate colitis model. PLoS ONE. 2010;5:e12238. doi: 10.1371/journal.pone.0012238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Gäbel G., Northoff B.H., Weinzierl I., Ludwig S., Hinterseher I., Wilfert W., Teupser D., Doderer S.A., Bergert H., Schönleben F., et al. Molecular Fingerprint for Terminal Abdominal Aortic Aneurysm Disease. J. Am. Heart Assoc. 2017;6:e006798. doi: 10.1161/JAHA.117.006798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Lieleg O., Lieleg C., Bloom J., Buck C.B., Ribbeck K. Mucin biopolymers as broad-spectrum antiviral agents. Biomacromolecules. 2012;13:1724–1732. doi: 10.1021/bm3001292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Braga Emidio N., Hoffmann W., Brierley S.M., Muttenthaler M. Trefoil Factor Family: Unresolved Questions and Clinical Perspectives. Trends Biochem. Sci. 2019;44:387–390. doi: 10.1016/j.tibs.2019.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Limaye S.A., Haddad R.I., Cilli F., Sonis S.T., Colevas A.D., Brennan M.T., Hu K.S., Murphy B.A. Phase 1b, multicenter, single blinded, placebo-controlled, sequential dose escalation study to assess the safety and tolerability of topically applied AG013 in subjects with locally advanced head and neck cancer receiving induction chemotherapy. Cancer. 2013;119:4268–4276. doi: 10.1002/cncr.28365. [DOI] [PubMed] [Google Scholar]
- 174.Peterson D.E., Barker N.P., Akhmadullina L.I., Rodionova I., Sherman N.Z., Davidenko I.S., Rakovskaya G.N., Gotovkin E.A., Shinkarev S.A., Kopp M.V., et al. Phase II, randomized, double-blind, placebo-controlled study of recombinant human intestinal trefoil factor oral spray for prevention of oral mucositis in patients with colorectal cancer who are receiving fluorouracil-based chemotherapy. J. Clin. Oncol. 2008;27:4333–4338. doi: 10.1200/JCO.2008.21.2381. [DOI] [PubMed] [Google Scholar]
- 175.Chen R.-M., Chiou Y.-S., Chong Q.-Y., Poh H.-M., Tan T.-Z., Zhang M.-Y., Ma L., Zhu T., Pandey V., Kumar A.P., et al. Pharmacological Inhibition of TFF3 Enhances Sensitivity of CMS4 Colorectal Carcinoma to 5-Fluorouracil through Inhibition of p44/42 MAPK. Int. J. Mol. Sci. 2019;20:6215. doi: 10.3390/ijms20246215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Van Vliet M., Harmsen H.J., de Bont E.S., Tissing W.J. The role of intestinal microbiota in the development and severity of chemotherapy-induced mucositis. PLoS Pathog. 2010;6:e1000879. doi: 10.1371/journal.ppat.1000879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Caballero G.G., Kaltner H., Kutzner T.J., Ludwig A.-K., Manning J.C., Schmidt S., Sinowatz F., Gabius H.-J. How galectins have become multifunctional proteins. Histol. Histopathol. 2020;35:509–539. doi: 10.14670/HH-18-199. [DOI] [PubMed] [Google Scholar]