

Abstract
This investigation tested the efficiency of nitric oxide (NO) in alleviation of Cu-induced adverse impacts on seed germination and photosynthesis in Indian mustard (Brassica juncea L.). Pre-treatment of B. juncea seeds with sodium nitroprusside (SNP; NO donor) significantly improved the seed germination rate and also alleviated Cu-accrued oxidative stress. However, in the absence of NO, Cu caused a higher reduction in seed germination rate. The presence of NO strengthened the antioxidant defense system (glutathione reductase, ascorbate peroxidase, and superoxide dismutase) and thereby sustained the lower lipid peroxidation, reduced H2O2 content, and thiobarbituric acid reactive substances in Cu-exposed seeds. NO pre-treated seeds also retained a higher amylase activity and exhibited an improved seed germination rate. This effect of NO under Cu stress was also seen in plants originated from the NO pre-treated seeds, where the role of NO pre-treatment was reflected in the improved photosynthetic potential of B. juncea. Overall, NO pre-treatment not only improved the germination rate in seeds but also carried its effects in the grown seedlings evidenced as improved photosynthesis and growth. Potential mechanisms involved in the action of NO pre-treatment included NO-mediated significant strengthening of the antioxidant defense system and decreases in Cu-caused oxidative stress parameters.
Keywords: copper stress, nitric oxide pre-treatment, germination rate, photosynthesis
1. Introduction
Studies on heavy metal accumulation and its continuous addition in the environment affecting the agricultural system and human health have been known for a long time especially in less developed countries [1,2]. In this perspective, Cu has emerged as a severe pollutant because of its extensive use in industries and as a pesticide in the agricultural system [3,4]. Based on its available concentration, Cu performs a remarkable array of functions in plants. Generally, Cu is widely accepted as an essential micronutrient element for plants [4]. Proteins containing Cu as a cofactor take part in various biochemical processes, involved in plant growth and development, and protective mechanisms [5]. The optimum levels of Cu in leaves are established as 10 µg g−1 dry mass [6]. However, the acute levels of toxicity found for most of the crop plants are slightly higher (20–30 µg g−1 dry mass) [7]. Indian mustard (Brassica juncea L.) being the hyperaccumulator of metal(oids) can accumulate a high concentration of Cu and translocate much into above-ground parts that can be detrimental to any plant growth. Thus, maintaining Cu-homeostasis is essential for a plant to survive under high Cu, either through vacuolar sequestration, storage, or transport of metal ions from the cytoplasm to outer compartments [8,9,10]. This ability of B. juncea makes it a suitable model plant material to study the impact of high Cu concentration.
Photosynthetic functions are essentially influenced by the presence of a high Cu level. An elevated Cu level in leaves damages thylakoids; dissipates the electron transport in photosystem (PS) II (PSII), alters photophosphorylation and membrane integrity; and eventually decreases photosynthesis and major gas-exchange parameters [11,12,13]. Additionally, high concentrations of Cu induce oxidative stress that causes membrane disruption and lipid peroxidation [14,15]. Moreover, excess Cu also alters the process of mineral uptake [16,17] and also retards the seed germination process and overall health of the developing seedling [4]. Further, excess Cu stimulates the activity of phytochelatin synthase and increases the production of phytochelatins (PCs), cysteine-rich metal-chelating peptides. The production of PCs results in the chelation of Cu by sequestering the PCs–Cu conjugant in vacuoles decreasing free Cu concentration. Since PCs are oligomers of reduced glutathione (GSH), this can deplete the pool of GSH, an important metabolite involved in the maintenance of the cellular redox balance [18,19]. Thus, evaluating the potential influence of Cu on seed germination is a prerequisite for assessing its effects on various plant developmental stages. The studies have well documented the deleterious effects of Cu on seed germination and associated metabolic processes [4,20,21,22,23,24,25]. Copper toxicity is known to alter metabolism during seed germination and also to interfere with the pathway of ubiquitin-proteasome mechanism to oxidatively damaged proteins [20]. Elevated Cu concentrations were reported to inhibit the radicle elongation rate in Alyssum montanum and Thlaspi ochroleucum [21].
Modulation of the signaling of phytohormones has been used as a potential tool to enhance the adaptability of plants under abiotic stress conditions. In particular, nitric oxide (NO), a versatile signaling molecule has been found to play critical roles in plant defense reactions including induction of antioxidant potential under various plant abiotic stresses [26,27]. Adequate evidence substantiates a wide range of roles of NO in plant physiological processes including germination, photosynthesis, and defense mechanism both under normal and environmental stresses [28,29]. In plants, the cell-protective role of NO has been tested with DNA, proteins, lipids, and chlorophyll [30]. Sorghum seeds were reported to exhibit NO-production during germination [31]. Treatment of sodium nitroprusside (SNP; NO-source) found to enhance seed germination and also to break seed dormancy [32,33,34,35]. Moreover, NO-treatment promoted seed germination of wheat under elevated Cu-levels by improving antioxidant capacity [36]. Nitric oxide was also found to play a prominent role in the regulation of S-assimilation [37,38,39]. The reported involvement of NO in S-assimilation has been argued to result in a higher rate of S-assimilation yielding more cysteine and GSH production, and thereby maintaining PCs level optimum for detoxifying high metal ions [40,41].
In the present reported research, the effect of Cu-toxicity and the significance of NO in averting this toxicity in B. juncea were studied. The role of NO pre-treatment to seeds was studied in the alleviation of Cu-induced inhibition of seed germination in terms of measuring germination rate and amylase activity. Further, it was also investigated if this pre-treatment of NO could carry its effects to influence the antioxidant metabolism, photosynthetic functions, and growth at maturity without supplementation of NO at later developmental stages of plant growth. Modulation of the antioxidant system has always been proved as a strategy for improving stress impact tolerance mechanisms in plants via improved photosynthetic potential.
2. Materials and Methods
2.1. Treatments
Healthy uniform seeds of Indian mustard (Brassica juncea L. cv. Pusa Tarak) were surface sterilized with 0.1% hypochlorite (v/v) solution and washed several times with autoclaved double distilled water. The sterilized seeds were randomly placed on two filter papers covered Petri-plates for germination at 28 ± 2 °C containing 0, 0.5, 1, 2, 3, 4, 5, 6, 7, 8, 9, and 10 mM copper sulphate (CuSO4) for 3 days. Fifty seeds were placed on each Petri plate (n = 50). Standard radicle emergence of seeds was scored as the germination percentage. Since, 3.0 mM Cu concentration reduced the germination percentage near to half of the control, this concentration was considered as semi-lethal (LD50).
To understand NO action on seed germination under Cu stress, seeds of B. juncea were pre-treated with 0, 25, 50, 100, 200, and 250 μM SNP (NO donor) for 3 h, and later exposed to semi-lethal 3.0 mM Cu for 3 days. The screening of SNP concentration was done to find out its optimal concentration that can efficiently alleviate Cu-inhibited seed germination. Sodium nitroprusside SNP at 100 μM proved most effective in reducing 3.0 mM Cu and was used for further experimentations. In further experiments, the influence of NO action in presence of Cu was confirmed using its analogue potassium ferrocyanide (FCy) and its scavenger 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (cPTIO). The treatment included control (seeds germinated in deionized water only), 100 μM SNP (optimum concentration), 100 μM (FCy) or 100 μM SNP plus 200 μM cPTIO (SNP + cPTIO) for 3 h, and later exposed to the semi-lethal Cu concentration (3.0 mM).
In another experiment, the influence of NO action was studied on photosynthesis, growth and antioxidant metabolism at 30 days after germination (DAG). Another aim of this experiment was also to ascertain the effect of NO-mediated alleviation of Cu’s impacts on seed germination in developing plants. For this, B. juncea seeds were washed and surface-sterilized with 0.1% hypochlorite (v/v) and were germinated in pots filled with acid-washed autoclaved sand. The plants were raised in modified Hoagland nutrient solution applied on alternate days consisting of 3.0 mM KNO3, 0.5 μM CuSO4, 2.0 mM Ca(NO3)2, NH4H3 PO4, 25 μM H3BO4, 25 μM H3BO4, 2.0 μM MnCl2, 50 μM KCl, 20 μM ZnSO4, 20 μM Na2Fe-EDTA and 0.5 μM (NH4)6Mo7O24. The pots were placed in a naturally illuminated net house with an average temperature of 22 ± 3 °C during day and 14 ± 2 °C at night. In this experiment, seeds were pre-treated (before sowing) for 3 h with deionized water, 100 μM SNP, 100 μM FCy, or 100 μM SNP plus 200 μM cPTIO and were sown in pots in the presence or absence of 3.0 mM Cu in the form of CuSO4. The experimental design is given in Table 1. The treatments were arranged in a randomly blocked design with three replicates (n = 3) for each treatment. Plants were sampled at 30 DAG for recording different parameters.
Table 1.
Experimental design.
| Treatments | Details |
|---|---|
| Control | Seeds pre-treated with water and raised without Cu stress |
| Cu | Seeds pre-treated with water and raised with Cu stress |
| SNP | Seeds pre-treated with SNP and raised without Cu stress |
| SNP + Cu | Seeds pre-treated with SNP and raised with Cu stress |
| FCy + Cu | Seeds pre-treated with FCy and raised with Cu stress |
| SNP + cPTIO + Cu | Seeds pre-treated with SNP+cPTIO and raised with Cu stress |
Cu—Copper; SNP—Sodium nitroprusside; FCY—Potassium ferrocyanide; cPTIO—2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide.
2.2. Determination of H2O2 Content and Lipid Peroxidation
The assay of H2O2 was done following the method of Okuda et al. [42]. Germinated seeds (0.5 g) and fresh leaves (0.5 g) of the two experiments were ground in ice-cold 200 mM perchloric acid. After centrifugation at 1200× g for 10 min, perchloric acid of the supernatant was neutralized with 4 M KOH. The insoluble potassium perchlorate was eliminated by centrifugation at 500× g for 3 min. The reaction was started by the addition of peroxidase and an increase in the absorbance was recorded at A590 for 3 min. Contents of TBARS were measured according to Dhindsa et al. [43] by recording absorbance at 532 nm. Values obtained were corrected for non-specific turbidity by subtracting the absorbance at 600 nm. The TBARS content was calculated using its extinction coefficient of 155 mM−1 cm−1.
2.3. Assay of Antioxidant Enzymes
Germinated seeds (0.5 g) and fresh leaves (0.2 g) of the two experiments were homogenized with mortar and pestle using 100 mM potassium phosphate buffer (pH 7.0) containing 1% polyvinylpyrrolidone (PVP) (w/v) and 0.05% Triton X-100 (v/v). Later the centrifugation of homogenized material was done at 15,000× g for 20 min. The supernatant generated after centrifugation was utilized to assay the activity of glutathione reductase (GR) and superoxide dismutase (SOD). The assay of ascorbate peroxidase (APX) required addition of extraction buffer supplemented with 2.0 mM ascorbate.
Activity of SOD was determined as per the protocol of Beyer and Fridovich [44], and Giannopolitis and Ries [45] by monitoring the inhibition of photochemical reduction of nitro blue tetrazolium (NBT). Reaction mixture (5.0 mL) consisted of 50 mM Na2CO3 (pH 10.0), 5.0 mM HEPES (pH 7.6), 0.1 mM EDTA, 0.025% (v/v) Triton X-100, 13 mM methionine, 63 mM NBT and 1.3 mM of riboflavin. The extract of enzyme was illuminated for 15 min (360 μmol m2 s−1), and a set without illumination acted as a control to correct the turbidity of background absorbance. A unit of enzyme was expressed as the amount of the enzyme that inhibited the NBT reduction by 50% at 560 nm. The amount of the enzyme that inhibited the reduction of NBT by 50% at 560 nm was equal to one unit of SOD. Activity of APX and GR was assessed as per the protocol adopted by Nakano and Asada [46] and Foyer and Halliwell [47]. The detailed procedure for determining APX and GR activity was explained in our earlier study [41].
2.4. Assay of Amylase Activity
Seeds were pre-treated with water or with SNP (100 μM). Later, pre-treated seeds were kept for germination for 48 h in Petri plates containing 3 mM CuSO4 solution. Seeds (0.5 g) were then collected and homogenized in a pre-chilled mortar and pestle with 6 mL of 50 mM Tris-HCl (pH 7.5) containing 1% PVP and 15 mM 2-mercaptoethanol. The homogenate was collected and centrifuged at 4 °C at 10,000× g for 30 min. After centrifugation, the resulting supernatant was used for determining enzyme activity. The amylase activity was determined by using the protocol of starch-iodine following Collins et al. [48]. A unit of enzyme was expressed by taking the enzyme quantity to reach 50% of the original color intensity.
2.5. Determination of GSH Content
The GSH content in fresh leaves was determined spectrophotometrically according to the protocol provided by Anderson [49]. Using pre-chilled mortar and pestle, under 4 °C fresh leaf tissue (0.5 g) was crushed in 2.0 mL of 5% sulphosalicylic acid. Centrifugation of homogenized material was done at 10,000× g for 10 min. Then, 0.6 mL of phosphate buffer (100 mM, pH 7.0) and 40 mL of 5,5′-dithiobis-2-nitrobenzoic acid were added to the 0.5 mL of supernatant. After 2 min, the absorbance was recorded at 412 nm. The detailed protocol has been given in our earlier publication [50].
2.6. Histochemical Detection of Reactive Oxygen Species
Histochemical staining of seed and leaf samples was performed using the protocol given by Kumar et al. [51]. Nitro blue tetrazolium (NBT) and 3,3-diaminobenzidine (DAB) were used for the assay accumulation of both superoxide ion (O2−) and H2O2 in seed and leaf samples of different treatments. The samples from each treatment were kept in NBT solution prepared by dissolving 0.1 g of NBT in 50 mL of 50 mM sodium phosphate buffer (pH 7.5) in an amber-colored bottle and were incubated overnight. The stained samples were immersed in absolute ethanol and boiled in water-bath for 10 min for discoloration to get the staining clear. For DAB staining solution, 50 mg DAB was dissolved in 50 mL double-distilled water in an amber-colored bottle with pH 3.8. The samples from each treatment were incubated for 8 h in DAB solution. Subsequently, the stained samples were immersed in absolute ethanol and boiled in a water bath for 10 min for discoloration to get a clear visualization of the stained samples.
2.7. Confocal Laser Microscopy Study for ROS Imaging and Cell Viability Determination
For ROS imaging, root samples 1–2 cm in length were dipped into the 12.5 μM 2′,7′ dichlorofluorescin diacetate (H2DCFDA) solution in a Petri dish for 15 min and washed 3 times properly with double distilled water. Stained samples were kept on a glass slide and studied under a confocal microscope (excitation 400–490 nm, emission ≥ 520 nm). For the cell viability test, properly washed root samples (1–2 cm length) of each treatment were dipped into 25 μM propidium iodide (PI) solution. Stained samples were washed appropriately, placed on a glass slide, and were observed under a confocal microscope.
2.8. Determination of Cu Concentration
Leaf and root samples were dried in the oven for two days at 80 °C. The oven-dried samples were crushed to a fine powder in mortar and pestle. This fine powder was digested with a solution containing concentrated HNO3/HClO4 (3:1, v/v) and was diluted with water. Concentration of Cu was determined by Atomic Absorption Spectrophotometer (GBC, 932 plus; GBC Scientific Instruments, Braeside, Australia).
2.9. Photosynthetic Characteristics
Net photosynthesis (PN), intercellular CO2 (Ci) and stomatal conductance (gS) were measured in second topmost leaves of plants using Infrared gas analyzer (CID-340, Photosynthesis System, Bio-Science, Washington, USA). These parameters were recorded between 11 am to 12 noon when the photosynthetically active radiation (PAR) was above 780 μmol m−2 s−1 and at 380 ± 5 μmol−1 atmospheric CO2 concentrations.
Maximal quantum efficiency of photosystem II (PSII) (Fv/Fm) of full-fledged leaf second from the top was noted with the help of chlorophyll fluorometer (JUNIOR PAM, Heinz Walz, Germany). Proceeding to get the result of maximum fluorescence (Fm) and minimal fluorescence (Fo) intensity, leaf samples were kept in the dark condition for 30 min. Weak measuring pulses (0.1 µmol m−2 s−1) and saturating pulse (>6000 µmol m−2 s−1) were used to measure Fo and Fm, respectively. Difference between Fo and Fm was used to calculate the variable fluorescence (Fv). The maximum quantum yield efficiency of PS II was calculated as a ratio of Fv to Fm.
Activity of Rubisco in leaves was monitored by adopting the procedure of Usuda [52]. Fresh leaf samples (1.0 g) were ground in a pre-chilled mortar and pestle with an extraction buffer containing 0.25 M Tris-HCl (pH 7.8), 0.0025 M EDTA, 0.05 M MgCl2 and 37.5 mg dithiothreitol (DTT). Centrifugation of homogenized material was done at 10,000× g for 10 min at 4 °C. The resulting supernatant brought after centrifugation was used to measure enzyme activity. Reaction mixture (3.0 mL) contained 100 mM Tris-HCl (pH 8.0), 10 mM MgCl2, 0.2 mM NADH, 40 mM NaHCO3, 5.0 mM DTT, 4 mM ATP, 1U of 3-phosphoglycerate kinase, 1U of glyceraldehyde 3-phosphodehydrogenase, and 0.2 mM ribulose 1,5-bisphosphate (RuBP). Bradford [53] method was adopted to estimate protein content.
2.10. Determination of Growth Parameters
The plant samples from each treatment were dried in a hot-air oven at 80 °C. The dried leaf samples were weighed on an electrical balance and the weight was recorded as whole plant dry mass. To measure leaf area, leaf area meter (LA 211, Systronics, New Delhi, India) was used.
2.11. Physiological Measurements of Guard Cells
Upper leaves of 30 days old plants were plugged from each plant with different treatments and were fixed by 2.5% glutaraldehyde, and stomatal images were taken using scanning electron microscopy (JSM-6510 LV, JEOL, Tokyo, Japan). The epidermal peels of the leaf samples were removed from the abaxial side and stomatal images of the sections at 40× were taken using a compound microscope outfitted with NIKON digital camera. The stomatal aperture width was measured with the help of a micrometer scale.
2.12. NO Generation
The level of NO generation was confirmed by estimating nitrite content adopting the protocol given by Zhou et al. [54] with slight modifications. Using pre-chilled mortar and pestle, leaf samples (500 mg) were ground in 3.0 mL of 50 mM ice-cold acetic acid buffer (pH 3.6) containing 4% zinc acetate; and later, was centrifuged at 11,500× g for 15 min at 4 °C. The pellet obtained was washed twice with 1.0 mL of the extraction buffer and then centrifuged again. The resulting supernatants from the two spin were mixed and neutralized by the addition of 100 mg of charcoal. After brief vortex, the filtrate was collected. Each filtrate of 1.0 mL and Greiss reagent (0.1% N-1-naphthyl ethylenediamine dihydrochloride and 1% sulphanilamide in 5% H2PO4 solution) were mixed in the ratio of (1:1) and then for 30 min these were incubated at room temperature. The absorbance was read at 540 nm and NO content was measured from a calibration curve plotted using sodium nitrite as standard.
2.13. Statistical Analysis
Data obtained in the experiments were analyzed statistically using analysis of variance (ANOVA) by SPSS 17.0 for Windows and presented as treatment mean ± SE. Treatment means were compared using the least significant difference (LSD) at p < 0.05. Bars with the same letter are not significantly different by LSD test at p < 0.05.
3. Results
3.1. Effect of Cu Stress on Seed Germination Percentage
The germination percentage of seeds declined with the increasing Cu concentration from 1 to 10 mM (Figure 1A). At 3.0 mM Cu concentration, germination percentage declined nearly half of the control and was marked as a semi-lethal concentration and was selected for further experimentations. Figure 1B offers an overview showing Cu-induced inhibitory effect on seed germination under various Cu concentrations.
Figure 1.

(A) Germination percentage; (B) phenotype of randomly selected germinating seeds of Indian mustard (Brassica juncea L.) exposed to copper (Cu) concentrations (0, 0.5, 1, 2, 3, 4, 5, 6, 7, 8, 9 and 10 mM) for 3 days.
3.2. Effect of NO Pre-treatment on Germination of Cu Stressed Seeds
Seeds pre-treated with 0, 25, 50, 100, 200, and 250 µM SNP concentration for 3 h were later exposed to 3.0 mM Cu showed increased germination percentage up to 100 µM SNP treatment, then dropped to approximately the same as control at 250 µM. Here, 100 µM SNP was proved to be the most effective treatment against 3.0 mM Cu stress-induced inhibition of seed germination (Figure 2A). Figure 2B is a critique of seed germination under varying SNP concentrations. On the other hand, pre-treatment with SNP showed an increase in seed germination percentage under Cu stress. But the pre-treatment of Cu exposed seeds to NO analogue FCy showed no significant change in germination percentage in comparison to Cu exposed seeds. Further, the use of cPTIO along with SNP in Cu exposed seeds reversed the SNP induced inhibition of Cu stress effect on seed germination (Figure 2C).
Figure 2.
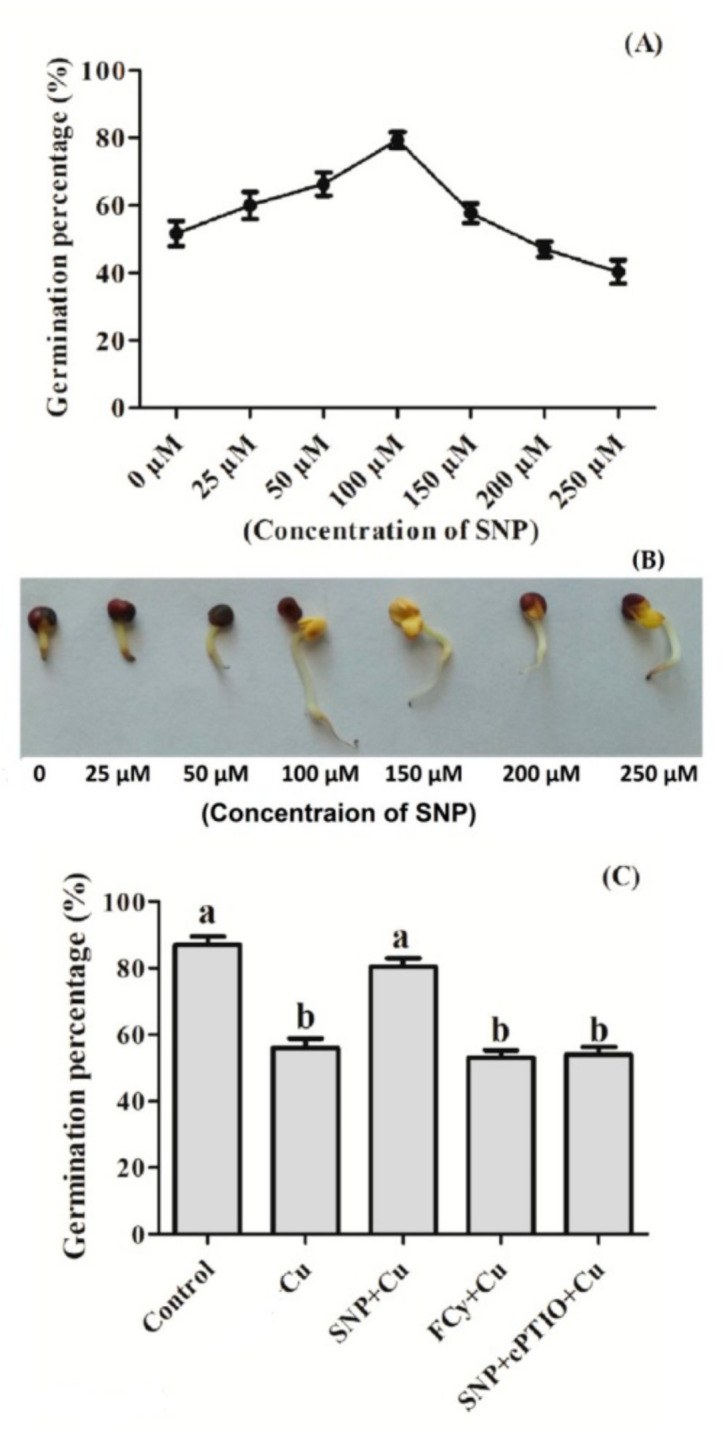
(A) Germination percentage under varying concentration of SNP under Cu stress; (B) phenotype of randomly selected germinating seeds of Indian mustard (Brassica juncea L.) pre-treated with 0, 25, 50, 100, 150, 200, and 250 µM SNP and exposed to 3.0 mM Cu; and (C) germination percentage in B. juncea seeds pre-treated with water, SNP, FCy (SNP analogue) and SNP + cPTIO (cPTIO as a specific NO-scavenger), later exposed to 3.0 mM Cu for 3 days. Same letter above bars show that data did not differ significantly by LSD test at p < 0.05.
3.3. Effect of NO on H2O2 and TBARS Contents in Germinating Seeds under Cu Stress
Copper stress markedly increased the content of H2O2 and TBARS where the values increased by 152.7 % and 106.7% compared to that of control seeds. On the other hand, application of SNP (prior germination) substantially reduced H2O2 by 33.1% and TBARS by 42.6% over the untreated control seeds. In addition, pre-germination application of FCy or SNP in combination with cPTIO in Cu-stressed seeds showed very similar result with stressed seeds (water pre-treated) over the control (Figure 3).
Figure 3.

H2O2 (A) and TBARS (B) content in germinating seeds of Indian mustard (Brassica juncea L.). Seeds were germinated with/without Cu stress and pre-treated with water, SNP, FCy (SNP analogue) and SNP + cPTIO (cPTIO as a specific NO-scavenger), later exposed to 3.0 mM Cu stress for 3 days. Same letters above bars show that data did not differ significantly by LSD test at p < 0.05. FW—fresh weight.
Further SNP pre-treatment under Cu stress reduced H2O2 content by about 47.3% and TBARS 31% in comparison to the Cu-stressed seeds. However, FCy or SNP in combination with cPTIO pre-treatment reduced H2O2 content by 7.6% and 6.8%, respectively, as compared to Cu-stressed seeds. In the case of TBARS content, FCy or SNP in combination with cPTIO reduced it equally by 2.5% and 2.6% respectively in comparison to stressed seeds (Figure 3A,B). These results of FCy or SNP in combination with cPTIO were comparable to Cu-stressed seeds.
3.4. Effect of NO Application on Activities of Antioxidant Enzymes under Cu Stress during Seed Germination
The performances of enzymatic antioxidants, such as APX, SOD, and GR, in mustard seeds are depicted in Figure 4. Results showed that activity of APX, GR and SOD were notably decreased in Cu-stressed seeds. In the presence of Cu, SNP did not influence activity of APX but significantly increased GR activity over the unstressed control seeds. On the other hand, SOD activity was lower in SNP treated seeds than those of control. However, the influence of FCy or SNP in combination with cPTIO showed results similar to stressed seeds.
Figure 4.

Activity of APX (A), SOD (B), and GR (C) in germinating seeds of Indian mustard (Brassica juncea L.). Seeds were germinated with/without Cu stress and pre-treated with water, SNP, FCy (SNP analogue) and SNP + cPTIO (cPTIO as a specific NO-scavenger), later exposed to 3.0 mM Cu stress for 3 days. Same letters above bars show that data did not differ significantly by LSD test at p < 0.05.
Application of SNP with Cu boosted the activity of antioxidant enzymes to a higher level than Cu-stressed seeds, increasing the activity of APX by about 99.5%, SOD by about 59.6%, and GR by 210.3% in contrast to the stressed seeds. However, in the presence of Cu, SNP in combination with cPTIO showed no difference than the Cu-stressed seeds in the activities of the above enzymes. Similarly, FCy also showed significantly similar activity of APX, SOD, and GR as was under Cu treatment alone (Figure 4).
3.5. Effect of NO Application on ROS Accumulation in Germinating Seeds
Under Cu stressed conditions, seeds pre-treated with water, FCy or SNP in combination with cPTIO showed an increased O2− accumulation as compared to SNP pre-treated seeds, which was observed by histochemical staining with NBT, showed dark blue stained area on radicle (a marker for O2− ion accumulation). While H2O2 accumulation revealed using DAB staining showed more reddish-brown precipitate on water pre-treated seeds, FCy or SNP + cPTIO pre-treated seeds as compared to SNP treatment under Cu stress (Figure 5A,B).
Figure 5.

Accumulation of superoxide ion (O2−) using NBT staining, and (A) H2O2 using DAB staining (B) in germinating seeds of Indian mustard (Brassica juncea L.) pre-treated with water, SNP, FCy (SNP analogue) or SNP + cPTIO (cPTIO as a specific NO-scavenger), exposed to 3.0 mM Cu for 3 days.
3.6. Effect of NO Application on the Activity of Amylase
From Figure 6, it can be observed that the activity of amylase increased substantially by the application of SNP to Cu-treated seeds compared to the seeds grown under Cu-stress. However, the optimum increase was noted at 12 and 24 h duration of SNP treatment, and then showed a reduction from 36 to 48 h duration.
Figure 6.

Activity of amylase in germinating seeds of Indian mustard (Brassica juncea L.) pre-treated with water or 100 µM SNP for 3 h (shown as from 0″ to 0/3 h of the treatment times) prior to exposing to 3.0 mM Cu for further 48 h (shown as 3/0, 12, 24, 36 and 48 h respectively). Same letters above bars show that data did not differ significantly by LSD test at p < 0.05. DW—dry weight.
3.7. Effect of NO Application on Cu Uptake and Levels of H2O2 and TBARS
To establish the role of NO in the Cu accumulation in Cu-exposed B. juncea plants, Cu content was measured in both roots and leaves (Figure 7A,B). The Cu accumulation was found much higher in roots (470 µg g−1) than in leaves (28.02 µg g−1). Pre-germination application of SNP (NO donor) resulted in a decrease in Cu content level in both roots and leaves of Cu treated plants. However, the NO-mediated diminished accumulation of Cu was reversed by NO-scavenger, cPTIO when applied in combination with SNP. Similarly, FCy treatment showed a little or no significant reversal in Cu accumulation by plants.
Figure 7.

Contents of Cu in root (A) and leaf (B), and contents of both H2O2 (C) and TBARS (D) in leaves of Indian mustard (Brassica juncea L.) at 30 DAG. Plants were grown with/without Cu stress and treated during pre-germination for 3 h with water, SNP, FCy (SNP analogue) and SNP + cPTIO (cPTIO as a specific NO-scavenger). Same letters above bars show that data did not differ significantly by LSD test at p < 0.05. FW, fresh weight; DW, dry weight.
Compared to control, plants receiving Cu showed greater content of H2O2 and TBARS (Figure 7C,D). The lone application of SNP exhibited decrease in H2O2 and TBARS content by about 35% and 45% without Cu stress. However, under Cu stress, this decrease in H2O2 and TBARS content was about 40% and 47% in comparison to Cu-treated plants. The protective effect of NO application was reversed by addition of cPTIO in Cu-fed plants. Further, FCy effect was significantly similar to the effect generated on application of cPTIO to SNP- and Cu-treated plants.
3.8. Effect of NO Treatment on ROS Accumulation by Leaves
Both H2O2 and O2− markedly accumulated in Cu-stressed plant (Figure 8A–F,G–L, represent H2O2 and O2− accumulation respectively). Accumulation of H2O2 and O2− was shown by histochemical staining with DAB and NBT, respectively. The leaves from the Cu-treated plants, with FCy and cPTIO in combination with NO applied plants showed discrete and deepest blue staining, which is a marker of O2− accumulation. Addition of SNP to non-stressed seeds seemed closer to control in terms of staining markers. However, application of NO noticeably diminished ROS accumulation in the leaves of Cu-stressed B. juncea leaves. A similar result was observed in DAB staining; but in this case, brownish patches were observed which was a marker of H2O2 accumulation in leaves, showing NO induced reduction in H2O2.
Figure 8.

Accumulation of superoxide ion (O2−) and H2O2 in leaves of Indian mustard (Brassica juncea L.) stained with NBT (A–F) and DAB (G–L) respectively at 30 DAG. Plants were grown with/without Cu stress and treated during pre-germination for 3 h with water, SNP, FCy (SNP analogue) and SNP + cPTIO (cPTIO as a specific NO-scavenger). Control: A and G; B and H: Cu; C and I: SNP; D and J: SNP + Cu; E and K: FCy + Cu; F and L: SNP + cPTIO + Cu.
3.9. Confocal Laser Scanning Microscopy
Copper induced H2O2 generation was visualized in roots by staining with H2DCFDA an indicator for ROS predominantly H2O2 in cells (Figure 9A–F). This reagent passively diffused into cells and its acetate groups cleaved by esterases. Upon oxidation, by H2O2 the non-fluorescent probe H2DCFDA gets converted to the highly fluorescent 2′,7-dichlorofluorescein (DCF). In this study, root cells of Cu stressed and FCy or SNP in combination with cPTIO-treated plants in the presence of Cu yielded higher intensity of green fluorescence while SNP reduced the effect of Cu stress and showed the less intensity of green fluorescence like that produced in control plants (Figure 9A–F).
Figure 9.

Confocal microscopic images of H2O2 formation in roots using H2DCFDA staining (A–F) and cell viability test (G–L) by propidium iodide staining, was performed on 30 days old roots of Indian mustard (Brassica juncea L.). Plants were grown with/without Cu stress and treated during pre-germination for 3 h with water, SNP, FCy (SNP analogue) and SNP + cPTIO (cPTIO as a specific NO-scavenger). Control: A and G; B and H: Cu; C and I: SNP; D and J: SNP+Cu; E and K: FCy + Cu; F and L: SNP+cPTIO+Cu.
Propidium iodide staining (an indicator of cell death) used to visualize cell viability by identifying the nucleic acid staining. Propidium iodide is membrane impairment and generally excluded from viable cells and in dead cells reaches the nucleus through distorted areas of dead cell membranes. In our study root cells of Cu stressed, FCy and of SNP in combination with cPTIO were less viable. However, Cu induced cell death was reduced by SNP application, showing similar response as was shown in control plants (Figure 9G–L).
3.10. Effect of NO on the Activity of Antioxidant Enzymes
In Figure 10, it is apparent that in the leaves of B. juncea, SOD activity increased with Cu treatment in contrast to the control. Sodium nitroprusside enhanced SOD activity to a higher level than in the presence of Cu alone. However, pre-treatment of SNP in stressed plant maximally elevated SOD activity, which was by 99.6% when compared to the control. Potassium ferrocyanide (FCy) and cPTIO in combination with SNP did not alter the SOD enzyme activity in the stressed plant and showed a response comparable to the control.
Figure 10.

APX (A), SOD (B) and GR (C), activity in leaves of Indian mustard (Brassica juncea L.) at 30 DAG. Plants were grown with/without Cu stress and treated during pre-germination for 3 h with water, SNP, FCy (SNP analogue) and SNP + cPTIO (cPTIO as a specific NO-scavenger). Same letters above bars show that data did not differ significantly by LSD test at p < 0.05.
Copper treatment increased the activity of GR and APX by 60.1% and 56.3%, respectively, as compared to control. Maximum increase in GR and APX activity was noted with the supplementation of NO (Cu + SNP), which increased the activity of both by about 40% and 60%, respectively, in leaves in comparison to the Cu-treated plant. The addition of NO-scavenger cPTIO nullified the influence of NO. Apart from that FCy also showed null effect on antioxidant enzymes activities in the Cu exposed plant.
3.11. Impact of NO on Photosynthetic Performance
Compared to the control plants getting NO in the form of SNP exhibited higher values for photosynthetic characteristics, such as PN, Ci, gs, maximal PSII photochemical efficiency, and Rubisco activity. Supplementation of NO in non-Cu fed plants improved PN value by 57%, Ci by 49.3%, gs by 36.2%, maximal PSII photochemical efficiency by 33.3% and Rubisco enzyme activity by 35% when compared to the control. However, Cu treatment reduced the values of above parameters by 38%, 21.7%, 29.5%, 45.1% and 40.7%, respectively in comparison to control. Further, in Cu-treated plants, NO alleviated Cu induced reduction and showed increased values for PN, Ci, gs, and PSII photochemical efficiency and Rubisco enzyme activity by 114%, 74.8%, 84.4%, 114.7% and 89%, respectively, in comparison to the Cu treated plants. However, in case of supplementation of FCy the results were same for all photosynthetic parameters as shown under Cu alone condition. The treatment of cPTIO in combination with NO nullified the effect of NO, resulted in reduced photosynthetic parameters in Cu exposed plants (Table 2).
Table 2.
Net photosynthesis, intercellular CO2 concentration, Maximal PSII photochemical efficiency, stomatal conductance, and Rubisco activity in Indian mustard (Brassica juncea L.) at 30 DAG. Seeds were treated at pre-germination time with water, 100 µM SNP or SNP with cPTIO and FCy in presence or absence of 3.0 mM Cu.
| Treatments | Net Photosynthesis (µmol CO2 m−2 s−1) | Internal CO2 Concentration (µmol CO2 mol−1) | Maximal PSII Photochemical Efficiency | Stomatal Conductance (mmol CO2 m−2 s−1) | Rubisco Activity (µmol CO2 mg1 protein min−1) |
|---|---|---|---|---|---|
| Control | 16.67 ± 0.89 c | 203 ± 5.57 c | 0.6 ± 0.09 c | 288 ± 9.54 b | 0.82 ± 0.06 b |
| Cu | 10.27 ± 0.92 e | 159 ± 5.51 d | 0.34 ± 0.06 d | 203 ± 8.54 c | 0.49 ± 0.05 c |
| SNP | 26.16 ± 0.72 a | 303 ± 4.04 a | 0.8 ± 0.08 a | 392.67 ± 8.19 a | 1.06 ± 0.07 a |
| SNP + Cu | 22 ± 0.58 b | 278 ± 6.08 b | 0.7 ± 0.06 b | 374.33 ± 8.68 a | 0.92 ± 0.09 ab |
| FCy + Cu | 12.96 ± 0.80 d | 168 ± 5.29 d | 0.38 ± 0.07 d | 221 ± 6.66 c | 0.52 ± 0.04 c |
| SNP + cPTIO + Cu | 11.97 ± 58 de | 162 ± 4.51 d | 0.35 ± 0.05 d | 208 ± 5.51 c | 0.51 ± 0.04 c |
The data followed by same superscripted letter are not significantly different by LSD test at p < 0.05.
3.12. Effect of NO on Physiology of Guard Cells
Changes in stomatal structure in response to SNP, FCy and cPTIO were studied through electron microscopy. Stomatal analysis depicted a noteworthy change in the stomatal functioning in Cu exposed plants. Stomatal opening (length and width) was 7.96 and 0.92 μm on 30th DAG in control plants (Figure 11). The stomatal opening was about 9.42 μm in pore length and stomata were found closed in leaf samples of Cu-exposed plants. However, the supplementation of SNP in the presence of Cu showed an increase in the stomatal aperture by about 2.29 μm in diameter. Conversely, treatment of SNP applied plants resulted in the maximal stomatal opening of 2.53 μm and 12.54 μm in pore length and pore width respectively. Further, the application of SNP treated Cu-stressed plants with its scavenger cPTIO showed decrease in the stomatal aperture by about 0.81 μm in diameter. However, when FCy was applied to Cu-treated plants showed the same result as plants grown under Cu treatment alone (Figure 11A–F).
Figure 11.

Stomatal response of Indian mustard (Brassica juncea L.) leaves at 30 DAG under control (A), Cu (B), SNP (C), SNP + Cu (D), FCy+ Cu (E) and SNP + Cu +cPTIO (F) at 4000× using scanning microscope.
The assertions of electron microscopy were further expanded by observations under compound microscopy. Stomata in leaf samples of control were normal with specialized guard cells, SNP applied in Cu treated plants showed an increase in stomatal aperture as compared to closed with distorted guard cells in stressed plants (Figure 12A–F).
Figure 12.

Stomatal response of Indian mustard (Brassica juncea L.) leaves at 30 DAG under control (A), Cu (B), SNP (C), SNP+Cu (D), FCy+Cu (E) and SNP+Cu+cPTIO (F) at 40× using compound microscope.
3.13. Effect of NO on NO Generation, GSH Content, Plant Dry Mass and Leaf Area
Figure 13 revealed that plants grown in Cu, exhibited increased NO generation by 4.3 times compared to control plants, but pre-germination supplementation of SNP decreased NO generation by 1.9 times compared to Cu-exposed plants. However, minimum decrease i.e., not much significant change in NO generation was noted by about 1.2 times with exogenously applied FCy. Furthermore, application of cPTIO in combination with NO resulted in the reversal of the outcome of NO generation in Cu-exposed plants.
Figure 13.

Effect of NO on NO generation (A), GSH (B), plant dry mass (C) and leaf area (D) of Indian mustard (Brassica juncea L.) treated during pre-germination with water, 100 µM SNP or SNP with cPTIO and FCy in presence or absence of 3 mM Cu at 30 DAG. Same letter above bars show that data did not differ significantly by LSD test at p < 0.05. FW—fresh weight; DW—dry weight.
B. juncea treated with 3.0 mM Cu increased GSH content while there was a reduction in plant dry mass and leaf area as compared to control. Plants with pre-germination treatment of SNP increased GSH content, and improved growth characteristics, plant dry mass and leaf area. Application of 100 µM SNP to Cu grown plants increased GSH content, plant dry mass and leaf area by 21%, 128%, and 100%, respectively as compared to Cu-treated plants, while the addition of cPTIO reversed the influence of exogenously applied NO and FCy application showed no significant change to Cu-exposed plants (Figure 13).
4. Discussion
Copper is an essential micronutrient required in a very low concentration by the plants for their proper growth and development and is a vital component of several biomolecules [6]. In excess of its permissible concentration, it has a high detrimental influence on seed germination and growth of plants [4,55]. In plants, NO acts as an important signaling molecule and has achieved an evident concern due to the fact of its crucial mitigating role in abiotic and biotic stresses [41,56,57,58].
In the current study, SNP as an efficient NO donor was used because it provides an absolute form of NO and supplies it for an extensively longer duration as compared to other NO donors [59,60]. However, SNP can also release cyanide (CN) and/or can form CN-Fe complex. It has been proposed that these complexes of CN may have an overlapping or distinct role with the role of NO alone on biological tissues [60]. Earlier, NO has been reported to play a significant role in the seed germination process in different plant species including warm-season grasses [35], Arabidopsis [34], and lettuce [32]. NO-mediated attenuation of the inhibition of germination of rice seed and the amelioration of the inhibition of seedling growth have also been found under cadmium (Cd) regimes [61]. These authors have also noticed a cPTIO-mediated reversal of the protective effect of exogenous supplementation of SNP. Besides, the incapability of potassium ferricyanide in the release NO like FCy and its antagonistic effects in contrast to SNP was also observed. In Paulownia elongata, SNP application was found involved in the breaking of seed dormancy and induction of seed germination [62]. Pre-soaking treatment also showed an advantageous effect on the attenuation of NaCl-induced inhibition of seed germination and growth of pakchoi radicles and plumules [63]. In agreement with the data published on other plants, our results proved the protective and stimulatory role of SNP pre-treatment on B. juncea seed germination. Conversely, the NO-scavenger cPTIO reversed the NO-induced seed germination under Cu stress, while its analogue FCy showed no change in germination percentage in comparison to Cu treated seeds. Results also confirmed that NO derived from SNP but not any other compound FCy was responsible for germination under Cu stress in mustard seeds. Additionally, the use of its analogue (FCy) did not work in mitigation but also inhibition of NO generation using cPTIO was proved ineffective. Sodium nitroprusside pre-treated B. juncea seeds were able to induce a rapid rise in the activity of amylase enzyme as compared to the stressed seeds with a steady rise to a maximum till 24 h (Figure 6). According to Zhang et al. [64], the amylase activity induced by NO was independent of gibberellic acid, showing that NO was involved in seed germination under various conditions. Similarly, Patel et al. [65] showed the involvement of NO in inducing germination of primed maize seeds.
Under Cu stress, B. juncea seeds not only showed inhibition in germination but also exhibited enhanced levels of TBARS and H2O2 content, even higher content was observed at the vegetative phase. These outcomes are in harmony with the earlier work of Hu et al. [36] and Fatma et al. [39], showing Cu and salt stress increased the accumulation of H2O2 and TBARS contents in wheat seeds and B. juncea leaves respectively. Per et al. [41] emphasized that NO tends to decrease the levels of TBARS and H2O2 content safeguarding the cell membrane to reduce the cell membrane damage through lipid peroxidation. An appropriate concentration of NO is required to induce an antioxidant system to prevent ROS induced oxidative damage that alleviates plant fitness loss [66]. In confronting heavy metal stress, NO was found to regulate the antioxidant system, including both the enzymatic and the non-enzymatic antioxidant system to alleviate ROS production [67]. This study further confirmed that the NO application reduced the production of ROS, H2O2, and O2− using DAB and NBT staining methods in both Cu fed plants and germinating seeds. This may be because NO detoxifying ROS by forming new compound peroxynitrite when reacts directly with the O2−. This newly formed compound itself acts as a potential signaling molecule in stress response, and also functions in the regulation of protein activity [68]. It has also been reported that NO improves Cu-tolerance by regulating H2O2 and O2− [69]. Furthermore, a plethora of literature depicts the involvement of NO in the inhibition of ROS accumulation of O2− and H2O2 and their subsequent localization using NBT and DAB staining [70,71,72]. Nitric oxide application also exhibited an inhibitory effect on Cu uptake in both shoots and roots in Cu fed plants (Figure 7A,B). Our results are following the outcomes of Wang et al. [73] in tomato, Mostofa et al. [74] in rice and Zhang et al. [75] in tomato seedlings.
The present study also indicated the participation of NO in enhancing the activities of antioxidant enzymes including APX, GR, and SOD in pre-treated germinating seeds and leaves of B. juncea grown plants under Cu stressed condition. In compliance, various earlier published reports have proved the expression of antioxidant enzymes by the supplementation of NO in Oryza sativa under nickel toxicity [76], Lycopersicon esculentum under salinity stress [58], and in cotton under NaCl stress [77]. Cu-pretreatment exhibited several times increase in Cu-concentrations both in roots and leaves of plants. However, the accumulation in leaves was lesser compared to roots (Figure 7). Limiting heavy metal in roots has been thought to be a mechanism of plant heavy metal stress tolerance. In earlier studies, as a primary strategy of a plant to counter potential metal-toxicity, roots have been found as efficient barriers in the translocation of most of the metals to the aboveground plant parts [78,79,80,81,82]. Herein, NO-addition increased the accumulation of Cu in roots and thereby decreased its contents in shoots. This result indicated the NO-mediated inhibition of excess Cu transport toward shoots. Involvement of NO-mediated S-nitrosylation (a regulator of protein activation) and the actions of PCs and some other -SH containing thiols were argued to be involved in the detoxification of heavy metals [83]. Therefore, NO-induced low Cu concentration in B. juncea leaves contributed to the increase of Cu tolerance. It was also observed that cPTIO treatment for 48 h could induce Cu accumulation in Cu-stressed periwinkle seedlings [84]. From these results, it can be concluded that NO might play an important role in reducing the Cu accumulation in mustard plants. In this study, SNP application was found to significantly upregulated GR activity and thereby increased the reduced pool of GSH, a major constituent of metal-chelating PCs (Figure 10 and Figure 13). Mostafa et al. [74] also confirmed that NO and GSH co-treatment was more effective in reducing Cu accumulation than lone treatment of SNP.
It has also been concluded by various studies that NO could confront much of the oxidative damage by regulating the activity of antioxidant enzymes in plants under heavy metal stress [85,86,87]. Our results are in agreement with the above works where APX, SOD, and GR activity were induced under SNP pre-treatment. Nitric oxide proved to be more promising and impactful in increasing photosynthetic characteristics, PN, Ci, gs, Rubisco activity, and PSII photochemical efficiency in the presence and absence of Cu stress (Table 1). The mechanism underlying the positive influence of NO on photosynthesis under Cu-toxicity may be correlated with the increased activity of antioxidant enzymes, the protection of chlorophyll, and decreased ROS accumulation. Earlier, it has been suggested that the NO can control the photosynthetic efficiency rate by controlling the size of the stomatal aperture, hence influencing the stomatal conductance [39,88]. SNP application also has some positive role in improving the activity of the Rubisco enzyme and ultimately enhancing photosynthetic activity. The study is supported by numerous findings such as protection of chlorophyll damage and enhancement in photosynthetic pigments in NO-treated Lolium perenne under Cu stress [79], Helianthus annuus exposed to Cd stress [89], and Triticum aestivum [90] and B. juncea [39] under salt stress. Treatment of cucumber seedlings, with SNP, enhanced the chlorophyll content, rate of photosynthesis, and transpiration rate and stomatal conductance [91]. These results were mainly due to the enhancement of antioxidant machinery, prevention, and recovery of chlorophyll damage and increased GSH content. In our study, the effect of NO prominently increased the content of GSH in the absence and presence of Cu stress. The study of Per et al. [41] also showed that NO accelerated GSH production in mustard plants treated with Cd.
The application of SNP eased the effect of Cu stress on plant dry weight and leaf area (Figure 13). Foliar spray of NO on several crops protected against Cu toxicity in Lycopersicon esculentum [75], B. juncea [92], Nicotiana tabaccum [93] and also alleviated the effect of Ni stress in rice [76]. Growth improvement can also be associated with the escalation in antioxidant machinery after NO supplementation. The study of Bai et al. [94] in L. perenne revealed that the application of NO mitigated Pb stress, and alleviated negative effects on leaf growth by increasing the mineral nutrient and by differing the oxidative stress parameters. Maximum NO accumulation was observed in the Cu stressed plants (Figure 13). It is because excess Cu also induces nitrosative stress by excessive production of NO, which subsequently reacts with ROS such as O2, O2− and H2O2 and results in other reactive nitrogen species in Cu stressed plants [95]. Thus, excessive accumulation of NO, along with H2O2 might be a potential mechanism for enhanced lipid peroxidation. Exogenously-sourced NO has been established to prevent the increase of oxidative damage and endogenous NO content in plants and increase tolerance of plants to Cu stress by up-regulation of the antioxidant system (APX, GR, and SOD) thereby ROS quenching which retained the cellular osmotic adjustment and protected photosynthetic machinery from Cu stress by acting as oxygen radical scrapper. SNP application resulted in the reduction of NO generation in plants grown with or without Cu stress. However, a minimum reduction of NO generation was found in Cu stressed plant and exogenously applied FCy as NO analogue which did not release NO on its breakdown in Cu stressed plant. Further, the application of cPTIO in combination with NO resulted in the reversal of the effect imposed by SNP on plants exposed to Cu. Wang et al. [96] also found that NO content increased in tomato plants in presence of Cu toxicity. We have earlier shown the mitigating effect of exogenous NO application under various abiotic stresses [39,41,50,88], where SNP was sprayed on the vegetative phase of the plants. In the following experiment, seeds were pretreated showing similar responses as procured in the above experiments, and the dose of SNP used was reduced to a much smaller level. Suggesting the use of pre-treatment of seeds could be a better strategy in improving the stress tolerance mechanism in plants with better germination percentage.
5. Conclusions
This study demonstrated that NO pretreatment was able to improve seed germination and reduced inhibitory effect of Cu by modulating antioxidant system, ROS accumulation, lipid peroxidation, and amylase activity. It also confirmed that pretreatment of NO significantly mitigated Cu toxicity also during the vegetative phase through improving the antioxidant system, photosynthetic efficiency, and reducing Cu induced accumulation of ROS accompanied by a reduction in lipid peroxidation of the mustard plant. Nitric oxide-pretreatment could be argued as a potential way of application that has shown here exhibiting both pre and post-germination-effects on B. juncea plants. From our results, it is concluded that no other compounds except NO derived from SNP, was able to promote seed germination and photosynthetic efficiency under Cu stress in Indian mustard. Therefore, pre-treatment of seeds with NO could be employed as a key biochemical approach for alleviating Cu-toxicity during seed germination, as it not only resulted in improving germination rate, but the applied NO also led to an improved vegetative phase of the plants.
Acknowledgments
The Authors are also grateful to the University Sophisticated Instruments Facility (USIF) of the Aligarh Muslim University for providing the necessary instrument facilities.
Author Contributions
Conceptualization, A.M., N.A.K. and N.A.A.; investigation and data curation, B.A.R. and I.R.M.; writing—original draft preparation, B.A.R. and I.R.M.; editing and content improvement, A.M., N.A.K. and N.A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Department of Science and Technology SERB (Project code: SB/YS/LS-108/2014), New Delhi India awarded to A.M. for advancing the laboratory facilities.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 1.Mostofa M.G., Hossain M.A., Fujita M., Tran L.-S.P. Physiological and biochemical mechanisms associated with trehalose-induced copper-stress tolerance in rice. Sci. Rep. 2015;5:11433. doi: 10.1038/srep11433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kumar V., Sharma A., Kaur P., Sidhu G.P.S., Bali A.S., Bhardwaj R., Thukral A.R., Cerda A. Pollution assessment of heavy metals in soils of India and ecological risk assessment: A state-of-the-art. Chemosphere. 2019;216:449–462. doi: 10.1016/j.chemosphere.2018.10.066. [DOI] [PubMed] [Google Scholar]
- 3.Gallagher D.L., Johnston K.M., Dietrich A.M. Fate and transport of copper-based crop protectants in plasticulture runoff and the impact of sedimentation as a best management practice. Water Res. 2001;35:2984–2994. doi: 10.1016/S0043-1354(00)00594-7. [DOI] [PubMed] [Google Scholar]
- 4.Adrees M., Ali S., Rizwan M., Ibrahim M., Abbas F., Farid M., Zia-Ur-Rehman M., Irshad M.K., Bharwana S.A. The effect of excess copper on growth and physiology of important food crops: A review. Environ. Sci. Pollut. Res. 2015;22:8148–8162. doi: 10.1007/s11356-015-4496-5. [DOI] [PubMed] [Google Scholar]
- 5.Ravet K., Pilon M. Copper and iron homeostasis in plants: The challenges of oxidative stress. Antiox. Redox Sign. 2013;19:919–932. doi: 10.1089/ars.2012.5084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Yruela I. Copper in plants: Acquisition, transport and interactions. Funct. Plant Biol. 2009;36:409–430. doi: 10.1071/FP08288. [DOI] [PubMed] [Google Scholar]
- 7.Alaoui-Sossé B., Genet P., Vinit-Dunand F., Toussaint M.L., Epron D., Badot P.M. Effect of copper on growth in cucumber plants (Cucumis sativus) and its relationships with carbohydrate accumulation and changes in ion contents. Plant Sci. 2004;166:1213–1218. doi: 10.1016/j.plantsci.2003.12.032. [DOI] [Google Scholar]
- 8.Purakayastha T.J., Viswanath T., Bhadraray S., Chhonkar P.K., Adhikari P.P., Suribabu K. Phytoextraction of zinc, copper, nickel and lead from a contaminated soil by different species of Brassica. Int. J. Phytorem. 2008;10:61–72. doi: 10.1080/15226510701827077. [DOI] [PubMed] [Google Scholar]
- 9.Mourato M.P., Moreira I.N., Leitão I., Pinto F.R., Sales J.R., Martins L.L. Effect of heavy metals in plants of the genus Brassica. Int. J. Mol. Sci. 2015;16:17975–17998. doi: 10.3390/ijms160817975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Rahman M.M., Azirun S.M., Boyce A.N. Enhanced accumulation of copper and lead in amaranth (Amaranthus paniculatus), Indian mustard (Brassica juncea) and sunflower (Helianthus annuus) PLoS ONE. 2013;8:e62941. doi: 10.1371/journal.pone.0062941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Küpper H., Götz B., Mijovilovich A., Küpper F.C., Meyer-Klaucke W. Complexation and toxicity of copper in higher plants. I. Characterization of copper accumulation, speciation, and toxicity in Crassula helmsii as a new copper accumulator. Plant Physiol. 2009;151:702–714. doi: 10.1104/pp.109.139717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Maksymiec W., Russa R., Urbanik-Sypniewska T., Baszyński T. Effect of excess Cu on the photosynthetic apparatus of runner bean leaves treated at two different growth stages. Physiol. Plant. 1994;91:715–725. doi: 10.1111/j.1399-3054.1994.tb03010.x. [DOI] [Google Scholar]
- 13.Emamverdian A., Ding Y., Mokhberdoran F., Xie Y. Heavy metal stress and some mechanisms of plant defense response. Sci. World J. 2015;2015 doi: 10.1155/2015/756120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhou Y.-L., Huo S.-T., Wang L.-T., Meng J.-F., Zhang Z.-W., Xi Z.-M. Exogenous 24-Epibrassinolide alleviates oxidative damage from copper stress in grape (Vitis vinifera L.) cuttings. Plant Physiol. Biochem. 2018;130:555–565. doi: 10.1016/j.plaphy.2018.07.029. [DOI] [PubMed] [Google Scholar]
- 15.Contreras L., Mella D., Moenne A., Correa J.A. Differential responses to copper-induced oxidative stress in the marine macro algae Lessonia nigrescens and Scytosiphon lomentaria (Phaeophyceae) Aquat. Toxic. 2009;94:94–102. doi: 10.1016/j.aquatox.2009.06.004. [DOI] [PubMed] [Google Scholar]
- 16.Ke W., Xiong Z.-T., Chen S., Chen J. Effects of copper and mineral nutrition on growth, copper accumulation and mineral element uptake in two Rumex japonicus populations from a copper mine and an uncontaminated field sites. Environ. Exp. Bot. 2007;59:59–67. doi: 10.1016/j.envexpbot.2005.10.007. [DOI] [Google Scholar]
- 17.Puig S., Andrés-Colás N., García-Molina A., Penarrubia L. Copper and iron homeostasis in Arabidopsis: Responses to metal deficiencies, interactions and biotechnological applications. Plant Cell Environ. 2007;30:271–290. doi: 10.1111/j.1365-3040.2007.01642.x. [DOI] [PubMed] [Google Scholar]
- 18.Gonzalez-Mendoza D., Moreno A.Q., Zapata-Perez O. Coordinated responses of phytochelatin synthase and metallothionein genes in black mangrove, Avicennia germinans, exposed to cadmium and copper. Aquat. Toxicol. 2007;83:306–314. doi: 10.1016/j.aquatox.2007.05.005. [DOI] [PubMed] [Google Scholar]
- 19.Navarrete A., González A., Gómez M., Contreras R.A., Díaz P., Lobos GMoenne A. Copper excess detoxification is mediated by a coordinated and complementary induction of glutathione, phytochelatins and metallothioneins in the green seaweed Ulva compressa. Plant Physiol. Biochem. 2019;135:423–431. doi: 10.1016/j.plaphy.2018.11.019. [DOI] [PubMed] [Google Scholar]
- 20.Karmous I., Trevisan R., El Ferjani E., Chaoui A., Sheehan D. Redox biology response in germinating Phaseolus vulgaris seeds exposed to copper: Evidence for differential redox buffering in seedlings and cotyledon. PLoS ONE. 2017;12:e0184396. doi: 10.1371/journal.pone.0184396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ouzounidou G. Effect of copper on germination and seedling growth of Minuartia, Silene, Alyssum and Thlaspi. Biol. Plant. 1995;37:411. doi: 10.1007/BF02913990. [DOI] [Google Scholar]
- 22.Bes C.M., Jaunatre R., Mench M. Seed bank of Cu-contaminated top soils at a wood preservation site: Impacts of copper and compost on seed germination. Environ. Monit. Assess. 2013;185:2039–2053. doi: 10.1007/s10661-012-2686-x. [DOI] [PubMed] [Google Scholar]
- 23.Ashagre H., Almaw D., Feyisa T. Effect of copper and zinc on seed germination, phytotoxicity, tolerance and seedling vigor of tomato (Lycopersicon esculentum L. cultivar Roma VF) Int. J. Agric. Sci. Res. 2013;2:312–317. [Google Scholar]
- 24.Adhikari T., Kundu S., Biswas A.K., Tarafdar J.C., Rao A.S. Effect of copper oxide nanoparticle on seed germination of selected crops. J. Agric. Sci. Technol. 2012;2:815. [Google Scholar]
- 25.Muccifora S., Bellani L.M. Effects of copper on germination and reserve mobilization in Vicia sativa L. seeds. Environ. Pollut. 2013;179:68–74. doi: 10.1016/j.envpol.2013.03.061. [DOI] [PubMed] [Google Scholar]
- 26.Gould K., Lamotte O., Klinguer A., Pugin A., Wendehenne D. Nitric oxide production in tobacco leaf cells: A generalized stress response? Plant Cell Environ. 2003;26:1851–1862. doi: 10.1046/j.1365-3040.2003.01101.x. [DOI] [Google Scholar]
- 27.Del Castello F., Nejamkin A., Cassia R., Correa-Aragunde N., Fernández B., Foresi N., Lombardo C., Ramirez L., Lamattina L. The era of nitric oxide in plant biology: Twenty years tying up loose ends. Nitric Oxide. 2019;85:17–27. doi: 10.1016/j.niox.2019.01.013. [DOI] [PubMed] [Google Scholar]
- 28.Corpas F.J., Palma J.M. Assessing nitric oxide (NO) in higher plants: An outline. Nitrogen. 2018;1:12–20. doi: 10.3390/nitrogen1010003. [DOI] [Google Scholar]
- 29.Santisree P., Adimulam S.S., Sharma K., Bhatnagar-Mathur P., Sharma K.K. Insights into the Nitric Oxide Mediated Stress Tolerance in Plants. In: Khan M.I.R., Reddy P.S., Ferrante A., Khan N.A., editors. Plant Signaling Molecules. Elsevier; Amsterdam, The Netherlands: 2019. pp. 385–406. [DOI] [Google Scholar]
- 30.Lamattina L., García-Mata C., Graziano M., Pagnussat G. Nitric oxide: The versatility of an extensive signal molecule. Ann. Rev. Plant Biol. 2003;54:109–136. doi: 10.1146/annurev.arplant.54.031902.134752. [DOI] [PubMed] [Google Scholar]
- 31.Simontacchi M., Jasid S., Puntarulo S. Nitric oxide generation during early germination of sorghum seeds. Plant Sci. 2004;167:839–847. doi: 10.1016/j.plantsci.2004.05.028. [DOI] [Google Scholar]
- 32.Beligni M.V., Lamattina L. Nitric oxide stimulates seed germination and de-etiolation, and inhibits hypocotyl elongation, three light-inducible responses in plants. Planta. 2000;210:215–221. doi: 10.1007/PL00008128. [DOI] [PubMed] [Google Scholar]
- 33.Zhang H., Shen W.-B., Xu L.-L. Effects of nitric oxide on the germination of wheat seeds and its reactive oxygen species metabolisms under osmotic stress. Acta Bot. Sin. 2003;45:901–905. [Google Scholar]
- 34.Bethke P.C., Gubler F., Jacobsen J.V., Jones R.L. Dormancy of Arabidopsis seeds and barley grains can be broken by nitric oxide. Planta. 2004;219:847–855. doi: 10.1007/s00425-004-1282-x. [DOI] [PubMed] [Google Scholar]
- 35.Sarath G., Bethke P.C., Jones R., Baird L.M., Hou G., Mitchell R.B. Nitric oxide accelerates seed germination in warm-season grasses. Planta. 2006;223:1154–1164. doi: 10.1007/s00425-005-0162-3. [DOI] [PubMed] [Google Scholar]
- 36.Hu K.-D., Hu L.-Y., Li Y.-H., Zhang F.-Q., Zhang H. Protective roles of nitric oxide on germination and antioxidant metabolism in wheat seeds under copper stress. Plant Growth Regul. 2007;53:173–183. doi: 10.1007/s10725-007-9216-9. [DOI] [Google Scholar]
- 37.Fatma M., Masood A., Per T.S., Rasheed F., Khan N.A. Interplay between nitric oxide and sulfur assimilation in salt tolerance in plants. Crop J. 2016;4:153–161. doi: 10.1016/j.cj.2016.01.009. [DOI] [Google Scholar]
- 38.Jahan B., Al Ajmi M.F., Rehman M.T., Khan N.A. Treatment of nitric oxide supplemented with nitrogen and sulfur regulates photosynthetic performance and stomatal behavior in mustard under salt stress. Physiol. Plant. 2020;168:490–510. doi: 10.1111/ppl.13056. [DOI] [PubMed] [Google Scholar]
- 39.Fatma M., Masood A., Per T.S., Khan N.A. Nitric oxide alleviates salt stress inhibited photosynthetic performance by interacting with sulfur assimilation in mustard. Front. Plant Sci. 2016;7:521. doi: 10.3389/fpls.2016.00521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wang J., Yu S.X., Zhang M., Cui X.M. Exogenous nitric oxide-mediated GSH-PC synthesis pathway in tomato under copper stress. Russ. J. Plant Physiol. 2015;62:349–359. doi: 10.1134/S1021443715030188. [DOI] [Google Scholar]
- 41.Per T.S., Masood A., Khan N.A. Nitric oxide improves S-assimilation and GSH production to prevent inhibitory effects of cadmium stress on photosynthesis in mustard (Brassica juncea L.) Nitric Oxide. 2017;68:111–124. doi: 10.1016/j.niox.2016.12.012. [DOI] [PubMed] [Google Scholar]
- 42.Okuda T., Matsuda Y., Yamanaka A., Sagisaka S. Abrupt increase in the level of hydrogen peroxide in leaves of winter wheat is caused by cold treatment. Plant Physiol. 1991;9:1265–1267. doi: 10.1104/pp.97.3.1265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Dhindsa R.S., Plumb-Dhindsa P., Thorpe T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981;32:93–101. doi: 10.1093/jxb/32.1.93. [DOI] [Google Scholar]
- 44.Beyer W.F., Jr., Fridovich I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987;161:559–566. doi: 10.1016/0003-2697(87)90489-1. [DOI] [PubMed] [Google Scholar]
- 45.Giannopolitis C.N., Ries S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977;59:309–314. doi: 10.1104/pp.59.2.309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Nakano Y., Asada K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981;22:867–880. doi: 10.1093/oxfordjournals.pcp.a076232. [DOI] [Google Scholar]
- 47.Foyer C.H., Halliwell B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta. 1976;133:21–25. doi: 10.1007/BF00386001. [DOI] [PubMed] [Google Scholar]
- 48.Collins G., Jenner C., Paleg L. The metabolism of soluble nucleotides in wheat aleurone layers treated with gibberellic acid. Plant Physiol. 1972;49:404–410. doi: 10.1104/pp.49.3.404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Anderson M.E. Determination of glutathione and glutathione disulfide in biological samples. Methods Enzymol. 1985;113:548–555. doi: 10.1016/s0076-6879(85)13073-9. [DOI] [PubMed] [Google Scholar]
- 50.Fatma M., Khan N.A. Nitric oxide protects photosynthetic capacity inhibition by salinity in Indian mustard. J. Funct. Environ. Bot. 2014;4:106–116. doi: 10.5958/2231-1750.2014.00009.2. [DOI] [Google Scholar]
- 51.Kumar D., Yusuf M.A., Singh P., Sardar M., Sarin N.B. Histochemical detection of superoxide and H2O2 accumulation in Brassica juncea seedlings. Biol. Protoc. 2014;4:e1108. doi: 10.21769/BioProtoc.1108. [DOI] [Google Scholar]
- 52.Usuda H. The activation state of ribulose 1, 5-bisphosphate carboxylase in maize leaves in dark and light. Plant Cell Physiol. 1985;26:1455–1463. [Google Scholar]
- 53.Bradford M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 54.Zhou B., Guo Z., Xing J., Huang B. Nitric oxide is involved in abscisic acid-induced antioxidant activities in Stylosanthes guianensis. J. Exp. Bot. 2005;56:3223–3228. doi: 10.1093/jxb/eri319. [DOI] [PubMed] [Google Scholar]
- 55.Zhang H., Hu L.Y., Hu K.D., He Y.D., Wang S.H., Luo J.P. Hydrogen sulfide promotes wheat seed germination and alleviates oxidative damage against copper stress. J. Integr. Plant Biol. 2008;50:1518–1529. doi: 10.1111/j.1744-7909.2008.00769.x. [DOI] [PubMed] [Google Scholar]
- 56.Delledonne M., Xia Y., Dixon R.A., Lamb C. Nitric oxide functions as a signal in plant disease resistance. Nature. 1998;394:585. doi: 10.1038/29087. [DOI] [PubMed] [Google Scholar]
- 57.Wimalasekera R., Tebartz F., Scherer G.F. Polyamines, polyamine oxidases and nitric oxide in development, abiotic and biotic stresses. Plant Sci. 2011;181:593–603. doi: 10.1016/j.plantsci.2011.04.002. [DOI] [PubMed] [Google Scholar]
- 58.Manai J., Kalai T., Gouia H., Corpas F. Exogenous nitric oxide (NO) ameliorates salinity-induced oxidative stress in tomato (Solanum lycopersicum) plants. J. Soil. Sci. 2014;14:433–446. doi: 10.4067/S0718-95162014005000034. [DOI] [Google Scholar]
- 59.Mur L.A., Mandon J., Persijn S., Cristescu S.M., Moshkov I.E., Novikova G.V., Hall M.A., Harren F.J.M., Hebelstrup K.H., Gupta K.J. Nitric oxide in plants: An assessment of the current state of knowledge. AoB Plants. 2013;5 doi: 10.1093/aobpla/pls052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Bethke P.C., Libourel I.G., Reinöhl V., Jones R.L. Sodium nitroprusside, cyanide, nitrite, and nitrate break Arabidopsis seed dormancy in a nitric oxide-dependent manner. Planta. 2006;223:805–812. doi: 10.1007/s00425-005-0116-9. [DOI] [PubMed] [Google Scholar]
- 61.He J., Ren Y., Chen X., Chen H. Protective roles of nitric oxide on seed germination and seedling growth of rice (Oryza sativa L.) under cadmium stress. Ecotoxicol. Environ. Saf. 2014;108:114–119. doi: 10.1016/j.ecoenv.2014.05.021. [DOI] [PubMed] [Google Scholar]
- 62.Liu J., Xue T., Shen Y. Effect of Nitric Oxide on Seed Germination and Dormancy in Empress Trees. HortTechnology. 2019;29:271–275. doi: 10.21273/HORTTECH04250-18. [DOI] [Google Scholar]
- 63.Ren Y., Wang W., He J., Zhang L., Wei Y., Yang M. Nitric oxide alleviates salt stress in seed germination and early seedling growth of pakchoi (Brassica chinensis L.) by enhancing physiological and biochemical parameters. Ecotoxicol. Environ. Saf. 2020;187:109785. doi: 10.1016/j.ecoenv.2019.109785. [DOI] [PubMed] [Google Scholar]
- 64.Zhang H., Shen W.-B., Zhang W., Xu L.-L. A rapid response of β-amylase to nitric oxide but not gibberellin in wheat seeds during the early stage of germination. Planta. 2005;220:708–716. doi: 10.1007/s00425-004-1390-7. [DOI] [PubMed] [Google Scholar]
- 65.Patel P., Kadur Narayanaswamy G., Kataria S., Baghel L. Involvement of nitric oxide in enhanced germination and seedling growth of magneto primed maize seeds. Plant. Signal. Behav. 2017;12:e1293217. doi: 10.1080/15592324.2017.1293217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Terrón-Camero L.C., Peláez-Vico M.Á., Del-Val C., Sandalio L.M., Romero-Puertas M.C. Role of nitric oxide in plant responses to heavy metal stress: Exogenous application versus endogenous production. J. Exp. Bot. 2019;70:4477–4488. doi: 10.1093/jxb/erz184. [DOI] [PubMed] [Google Scholar]
- 67.Wei L., Zhang J., Wang C., Liao W. Recent progress in the knowledge on the alleviating effect of nitric oxide on heavy metal stress in plants. Plant Physiol. Biochem. 2020;147:161–171. doi: 10.1016/j.plaphy.2019.12.021. [DOI] [PubMed] [Google Scholar]
- 68.Baudhouin E. The language of nitric oxide signaling. Plant Biol. 2011;13:233–242. doi: 10.1111/j.1438-8677.2010.00403.x. [DOI] [PubMed] [Google Scholar]
- 69.Pető A., Lehotai N., Feigl G., Tugyi N., Ördög A., Gémes K., Kolbert Z. Nitric oxide contributes to copper tolerance by influencing ROS metabolism in Arabidopsis. Plant Cell Rep. 2013;32:1913–1923. doi: 10.1007/s00299-013-1503-5. [DOI] [PubMed] [Google Scholar]
- 70.Wang H., Hou J.L.Y., Zhang Y., Huang J., Liang W. Nitric oxide-mediated cytosolic glucose-6-phosphate dehydrogenase is involved in aluminum toxicity of soybean under high aluminum concentration. Plant Soil. 2017;416:39–52. doi: 10.1007/s11104-017-3197-x. [DOI] [Google Scholar]
- 71.Singh A.P., Dixit G., Kumar A., Mishra S., Kumar N., Dixit S., Dhankher O.P. A protective role for nitric oxide and salicylic acid for arsenite phytotoxicity in rice (Oryza sativa L.) Plant Physiol. Biochem. 2017;115:163–173. doi: 10.1016/j.plaphy.2017.02.019. [DOI] [PubMed] [Google Scholar]
- 72.Gong B., Nie W., Yan Y., Gao Z., Shi Q. Unraveling cadmium toxicity and nitric oxide induced tolerance in Cucumis sativus: Insight into regulatory mechanisms using proteomics. J. Hazard. Mater. 2017;336:202–213. doi: 10.1016/j.jhazmat.2017.04.058. [DOI] [PubMed] [Google Scholar]
- 73.Wang Y.J., Hu M.M., Cui X.M., Lou Y.H., Zhuge Y.P. Exogenous NO mediated the detoxification pathway of tomato seedlings under different stress of Cu and Cd. J. Appl. Ecol. 2018;29:4199. doi: 10.13287/j.1001-9332.201812.031. [DOI] [PubMed] [Google Scholar]
- 74.Mostofa M.G., Seraj Z.I., Fujita M. Exogenous sodium nitroprusside and glutathione alleviate copper toxicity by reducing copper uptake and oxidative damage in rice (Oryza sativa L.) seedlings. Protoplasma. 2014;251:1373–1386. doi: 10.1007/s00709-014-0639-7. [DOI] [PubMed] [Google Scholar]
- 75.Zhang Y., Han X., Chen X., Jin H., Cui X. Exogenous nitric oxide on antioxidative system and ATPase activities from tomato seedlings under copper stress. Sci. Hortic. 2009;123:217–223. doi: 10.1016/j.scienta.2009.08.015. [DOI] [Google Scholar]
- 76.Rizwan M., Mostofa M.G., Ahmad M.Z., Imtiaz M., Mehmood S., Adeel M., Dai Z., Li Z., Aziz O., Zhang Y., et al. Nitric oxide induces rice tolerance to excessive nickel by regulating nickel uptake, reactive oxygen species detoxification and defense-related gene expression. Chemosphere. 2018;191:23–35. doi: 10.1016/j.chemosphere.2017.09.068. [DOI] [PubMed] [Google Scholar]
- 77.Dong Y., Jinc S., Liu S., Xu L., Kong J. Effects of exogenous nitric oxide on growth of cotton seedlings under NaCl stress. J. Soil Sci. Plant Nutr. 2014;14:1–13. doi: 10.4067/S0718-95162014005000001. [DOI] [Google Scholar]
- 78.Dong Y.X., Wang X.F., Cui X.M. Exogenous nitric oxide involved in subcellular distribution and chemical forms of Cu2+ under copper stress in tomato seedlings. J. Integr. Agric. 2013;12:1783–1790. doi: 10.1016/S2095-3119(13)60367-6. [DOI] [Google Scholar]
- 79.Dong Y., Xu L., Wang Q., Fan Z., Kong J., Bai X. Effects of exogenous nitric oxide on photosynthesis, antioxidative ability, and mineral element contents of perennial ryegrass under copper stress. J. Plant Interact. 2014;9:402–411. doi: 10.1080/17429145.2013.845917. [DOI] [Google Scholar]
- 80.Hattab S., Chouba L., Benkheder M., Mahouachi T., Boussetta H. Cadmium- and copper-induced DNA damage in Pisum sativum roots and leaves as determined by the Comet assay. Plant Biosyst. 2009;143:6–11. doi: 10.1080/11263500903187035. [DOI] [Google Scholar]
- 81.Xiong J., An L., Lu H., Yhu C. Exogenous nitric oxide enhances cadmium tolerance of rice by increasing pectin and hemicellulose contents in root cell wall. Planta. 2009;230:755–765. doi: 10.1007/s00425-009-0984-5. [DOI] [PubMed] [Google Scholar]
- 82.Wang Q.H., Liang X., Dong Y.J., Xu L.L., Zhang X.W., Hou J., Fan Z.Y. Effects of exogenous nitric oxide on cadmium toxicity, element contents and antioxidative system in perennial ryegrass. Plant Growth Regul. 2013;69:11–20. doi: 10.1007/s10725-012-9742-y. [DOI] [Google Scholar]
- 83.Fecht-Christoffers M.M., Braun H.P., Lemaitre-Guillier C., van Dorsselaer A., Horst W.J. Effect of manganese toxicity on the proteome of the leaf apoplast in cowpea. Plant Physiol. 2003;133:1935–1946. doi: 10.1104/pp.103.029215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Liu S., Yang R., Pan Y., Ren B., Chen Q., Li X., Ma M. Beneficial behavior of nitric oxide in copper-treated medicinal plants. J. Hazard. Mater. 2016;314:140–154. doi: 10.1016/j.jhazmat.2016.04.042. [DOI] [PubMed] [Google Scholar]
- 85.Kaur G., Singh H.P., Batish D.R., Mahajan P., Kohli R.K., Rishi V. Exogenous nitric oxide (NO) interferes with lead (Pb)-induced toxicity by detoxifying reactive oxygen species in hydroponically grown wheat (Triticumaestivum) roots. PLoS ONE. 2015;10:e0138713. doi: 10.1371/journal.pone.0138713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Tripathi D.K., Singh S., Singh S., Srivastava P.K., Singh V.P., Singh S., Prasad S.M., Singh P.K., Dubey N.K., Pandey A.C., et al. Nitric oxide alleviates silver nanoparticles (AgNps)-induced phytotoxicity in Pisumsativum seedlings. Plant Physiol. Biochem. 2017;110:167–177. doi: 10.1016/j.plaphy.2016.06.015. [DOI] [PubMed] [Google Scholar]
- 87.Ahmad P., Ahanger M.A., Alyemeni M.N., Wijaya L., Alam P. Exogenous application of nitric oxide modulates osmolyte metabolism, antioxidants, enzymes of ascorbate-glutathione cycle and promotes growth under cadmium stress in tomato. Protoplasma. 2018;255:79–93. doi: 10.1007/s00709-017-1132-x. [DOI] [PubMed] [Google Scholar]
- 88.Sehar Z., Masood A., Khan N.A. Nitric oxide reverses glucose-mediated photosynthetic repression in wheat (Triticum aestivum L.) under salt stress. Environ. Exp. Bot. 2019;161:277–289. doi: 10.1016/j.envexpbot.2019.01.010. [DOI] [Google Scholar]
- 89.Laspina N., Groppa M., Tomaro M., Benavides M. Nitric oxide protects sunflower leaves against Cd-induced oxidative stress. Plant Sci. 2005;169:323–330. doi: 10.1016/j.plantsci.2005.02.007. [DOI] [Google Scholar]
- 90.Ruan H.-H., Shen W.-B., Xu L.-L. Nitric oxide involved in the abscisic acid induced proline accumulation in wheat seedling leaves under salt stress. Acta Bot. Sin. 2004;46:1307–1315. [Google Scholar]
- 91.Fan H., Guo S., Jiao Y., Zhang R., Li J. Effects of exogenous nitric oxide on growth, active oxygen species metabolism, and photosynthetic characteristics in cucumber seedlings under NaCl stress. Front. Agric. China. 2007;1:308–314. doi: 10.1007/s11703-007-0052-5. [DOI] [Google Scholar]
- 92.Feigl G., Kumar D., Lehotai N., Tugyi N., Molnár Á., Ördög A., Szepesi A., Gemes K., Laskay G., Erdei L., et al. Physiological and morphological responses of the root system of Indian mustard (Brassica juncea L. Czern.) and rapeseed (Brassica napus L.) to copper stress. Ecotoxicol. Environ. Saf. 2013;94:179–189. doi: 10.1016/j.ecoenv.2013.04.029. [DOI] [PubMed] [Google Scholar]
- 93.Khairy A.I.H., Oh M.J., Lee S.M., Roh K.S. Nitric oxide overcomes Cd and Cu toxicity in in vitro-grown tobacco plants through increasing contents and activities of rubisco and rubisco activase. Biochim. Open. 2016;2:41–51. doi: 10.1016/j.biopen.2016.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Bai X., Dong Y., Wang Q., Xu L., Kong J., Liu S. Effects of lead and nitric oxide on photosynthesis, antioxidative ability, and mineral element content of perennial ryegrass. Biol. Plant. 2015;59:163–170. doi: 10.1007/s10535-014-0476-8. [DOI] [Google Scholar]
- 95.Tewari R.K., Hahn E.J., Paek K.Y. Modulation of copper toxicity-induced oxidative damage by nitric oxide supply in the adventitious roots of Panax ginseng. Plant Cell Rep. 2008;27:171–181. doi: 10.1007/s00299-007-0423-7. [DOI] [PubMed] [Google Scholar]
- 96.Wang L., Yang L., Yang F., Li X., Song Y., Wang X., Hu X. Involvements of H2O2 and metallothionein in NO-mediated tomato tolerance to copper toxicity. J. Plant Physiol. 2010;167:1298–1306. doi: 10.1016/j.jplph.2010.04.007. [DOI] [PubMed] [Google Scholar]