

ABSTRACT
In this study, we present phenotypic and genetic data characterizing the impact of imidacloprid and caging stress on honey bee Apis mellifera physiological responses and regulation of 45 genes using targeted-RNA seq. The term ‘caging stress’ characterizes the effects of depriving honey bees of all hive aspects and conditions. Two cohorts of 1 day old sister bees were subjected to different conditions. One cohort was caged and fed different imidacloprid-tainted sugar solutions and the second was marked and introduced back to its natal hive. Physiological bee parameters and diet behavior were monitored daily for caged bees over several weeks. Bee samples from both cohorts were sampled weekly for RNA sequencing and oxidative stress analyses. Imidacloprid induced significant protein damage and post-ingestive aversion responses in caged bees, leading to lower tainted syrup consumption and higher water intake compared with the controls. No differentially expressed genes were observed among caged bees in regards to imidacloprid treatment. However, significant upregulation in antioxidant genes was recorded in caged bees as compared with hive bees, with overwhelming downregulation in all gene categories in caged bees at week 4. We identified two sets of genes that were constantly regulated in caged bees, including Rsod with unknown function in insects that could potentially characterize caging stress in honey bees.
KEY WORDS: Honey bee, RNA-seq, Cage stress, Imidacloprid, Gene regulation, Oxidative stress
Summary: Honey bees caged for experimental purposes show significant alteration of their gene regulation and oxidative stress compared with their sister-mates operating under natural hive conditions.
INTRODUCTION
The honey bee Apis mellifera is an important eusocial insect pollinator (Giannini et al., 2015; Morandin et al., 2001; Sampson and Cane, 2000). Although the debate on the overall impact of pesticides on honey bee health is ongoing (Stokstad, 2017), honey bee exposure to agricultural pesticides, particularly the neonicotinoids, is one of many factors that contribute to bee population decline (Sanchez-Bayo and Goka, 2014; Tosi et al., 2017; Williamson et al., 2013; Woodcock et al., 2017). Measuring pesticide toxicity for bees under field conditions is difficult because of the complexity of honey bee biology and foraging behaviors (Alburaki et al., 2015, 2017b; Cutler and Scott-Dupree, 2007; Stewart et al., 2014). Therefore, cage experiments are commonly used to test bees (e.g. toxicological and behavioral assessments) under more controlled conditions (Alburaki et al., 2017a; Gregorc et al., 2017). Despite providing more controlled conditions, little is known about the physiological and molecular responses of bees to caging stress.
Honey bees live in highly organized colonies in which each worker performs a very specific function and changes tasks throughout her physiological development (Winston, 1987). Absence of the queen or any disruption in the pheromone communication inside the bee colony is considered fatal for the whole population. When bees are caged for experimental purposes, significant pheromone disruption occurs, altering bee social behavior (Grozinger et al., 2003). For example, absence of the queen mandibular pheromone delays honey bee behavioral maturation (Robinson et al., 1998), and its presence prevents the rearing of new queens (Winston et al., 1991) and inhibits worker ovary development (Hoover et al., 2003). The ability of worker bees to execute their appropriate tasks and communicate with their hive mates is crucial for colony wellbeing.
Gene regulation in honey bees is often studied in relation to pathogen infection (Gregorc et al., 2012), exposure to abiotic stressors (Alburaki et al., 2017b), and other physiological and genetic factors among honey bee castes and developmental phases (Evans and Wheeler, 1999). These efforts have led to a better understanding of honey bee gene regulation and the identification and annotation of an important number of antioxidant and immune genes evolved in honey bee response to various stressors (Corona and Robinson, 2006; Evans et al., 2006).
Imidacloprid, a broadly used neonicotinoid, is acutely toxic to bees and can impair honey bee performance at sublethal doses (Chakrabarti et al., 2015; Williamson et al., 2014). Imidacloprid has a high agonistic affinity with nicotinic acetylcholine receptors (nAChR), particularly in brain tissue (Decourtye et al., 2004). Bees are equipped with complex detoxification mechanisms, including networks of enzymatic antioxidants, to help reduce potential oxidative damage provoked by biotic and abiotic stressors, including pesticides (Corona and Robinson, 2006). Interestingly, caged bees administrated ad libitum sugar syrup containing 5–10 times their median lethal concentration (LD50) of imidacloprid have shown the ability to survive for a relatively long period of time (4–7 weeks) (Alburaki et al., 2017a; Meikle et al., 2016). It is yet unclear whether those bees survive by minimizing their contaminated-syrup intake or have a metabolic capacity allowing them to quickly process the lethal molecules, or by the two scenarios together. It is also conceivable that the honey bee has the ability to sense the presence of those chemicals within the proposed syrup and refrains from feeding on such contaminated nutrients. It has been demonstrated that honey bees fed on neonicotinoids (thiamethoxam), 1 ng per bee for 12 days, significantly decreased their affinity for sucrose stimulation (Aliouane et al., 2009), which can partially explain the rejection of syrup with optimal sugar concentrations for bees. However, acetamiprid which is another neonicotinoid, orally administrated to bees (0.1 μg per bee) increased their responsiveness to water (Aliouane et al., 2009).
This study was conducted on sister honey bees exposed to two different conditions: (1) honey bees were removed from their natal hive and placed in cages in the laboratory and (2) honey bees were labelled and kept in their natal hive. We addressed two major and interrelated hypotheses: (1) compared with an untreated control, sister honey bees caged under similar conditions and administrated ad libitum sugar syrup tainted with lethal and sublethal concentrations of imidacloprid would exhibit higher mortality and upregulation in their detoxification genes; and (2) depriving honey bees of hive conditions by caging them would trigger significant disruption of gene regulation compared with their sister-mates of the natal hive. This study provides new insights into how imidacloprid affects honey bee physiological responses and diet behavior as well as its implications for the regulation of the major bee antioxidant and immune genes. Furthermore, we characterized potential genes involved in the response to cage stress by studying the differentially expressed genes of caged bees and their sister-mates operating under natural hive conditions.
MATERIALS AND METHODS
Honey bee colony
One well-established honey bee colony, the ‘mother hive’, headed by a Carniolan queen (Apis mellifera carnica Pollman 1879) was used as the source of all worker samples of this study. A single hive was used as a source of worker bees in this study to minimize variability in hive conditions and genetic make-up of the target organisms. A total of eight capped worker brood frames ready to hatch were removed from this hive and placed in an incubator at 35°C with 70–80% relative humidity. The following day, several thousand 1 day old sister bees were collected into a sterile plastic box for this study.
Cage and hive experiment
Newly hatched worker bees collected in the plastic box were gently mixed and 500 of them were randomly picked and marked with a white dot on their dorsal tergite using a collective marking box (Alburaki et al., 2017a). These workers were placed in a new plastic box to allow the paint to fully dry, then these marked 1 day old bees were introduced back into their natal hive. Another set of ∼150, 1 day old bees were stored at −80°C and were designated as time 0 reference bees (Fig. 1). A third set of 1800, 1 day old bees were randomly divided into 12 groups (150 bees per group); each group was weighed separately and introduced into 12 separate cages. The cages used in this study (dimensions of 11.4×6.3×15.2 cm width:depth:height; Fig. S4) were specifically designed for feeding experiments and are fully described in Gregorc et al. (2018). The 12 bee cages were randomly assigned to four treatment groups (three treatments and one control) in triplicate (Fig. 1). Bees in each cage were provided with distilled water and 1:1 sugar syrup using 30 ml syringes, and 10 g of protein patty (Mann Lake Bee Pro Patties). Imidacloprid was administrated to bees through the sugar syrup at four different dosages (0, 5, 20 and 100 ppb; Fig. 1). Patty, water and syrup consumption and bee mortality were documented daily. Dead bees were collected daily from the cages and stored at −80°C. The protein patties, which were placed into rubber plugs, were weighed using a ±0.01 g sensitive scale and gently placed back in the cages. Both syrup and water consumption were recorded visually from syringes at 0.5 ml sensitivity.
Fig. 1.

Experimental design and procedures. One-day-old sister bees were hatched in an incubator (35°C, 70% relative humidity) and distributed across 12 cages (150 bees per cage). Three imidacloprid treatments (5, 20, 100 ppb) and one control (0 ppb) were established in triplicate. In addition, 150 bees were stored at −80°C at time 0 and 500 others were marked and put back in the hive. Caged bees were provided with sugar syrup, water and a protein patty. Imidacloprid was administrated to bees of the treatment groups via the sugar syrup. Four samplings (25 bees per cage or hive) were carried out (once a week) from both cages and the hive (marked bees).
Worker bee sampling
Worker bees were sampled at four time points: 25 workers were sampled weekly from each cage as well as 75 marked workers from the mother hive. Each cohort of workers was weighed and stored at −80°C for subsequent molecular analyses. In addition, dead bees collected daily from each cage during the experiment were counted and weighed at the end of the experiment. A representative set of dead bees from each treatment (0, 5, 20, 100 ppb imidacloprid) was sent for chemical pesticide residue analysis. Four neonicotinoid molecules were screened (imidacloprid, imidacloprid olfen, thiamethoxam and clothianidin) by liquid chromatography-mass spectrometry (LC-MS) (Barnett et al., 2007; Walorczyk and Gnusowski, 2009). Chemical analyses for pesticide residue detection were processed at the USDA National Scientific Laboratories (Gastonia, NC, USA).
Oxidative stress
Hydrogen peroxide and protein carbonyl content assays were conducted to assess potential honey bee physiological stress and protein damage resulting from exposure to imidacloprid and caging stress. The level of H2O2 was quantified in the hydrogen peroxide assay using the bee hemolymph of samples collected in week 1. Bees were individually crushed in 1.5 ml tubes with 300 μl ultra-sterilized water and centrifuged at 11,000 g for 3 min. In order to eliminate proteins, the supernatant containing the bee hemolymph was filtered through a 10 kDa filter and the assay was conducted using a BioVision kit (Milpitas, CA, USA) as per the manufacturer's instructions. The protein carbonyl content assay was carried out on samples from weeks 1 and 4. Proteins were solubilized from honey bee thorax in a protein extraction buffer consisting of 20 mmol l−1 Tris-HCl pH 8.0, 30 mmol l−1 NaCl and 10% glycerol. The tissues were crushed using a pestle and sonicated using a Bioruptor Pico (Diagenode) sonication device for 10 cycles of 30 s pulse and 30 s rest at 4°C. Homogenates were centrifuged at 5000 g for 10 min at 4°C and the supernatants were collected. Quantification was conducted using a kit from Sigma-Aldrich (St Louis, MO, USA) as per the manufacturer's protocol.
Brain RNA extraction
Bee brains were dissected under a binocular microscope in an RNase free, sterile work environment. Total RNA was extracted from a pool of 15 bee brains per sample following the TRIzol® Reagent protocol (Invitrogen) (Chomczynski, 1993) with some modifications (see Alburaki et al., 2017a). Briefly, dissected brains were added to 1 ml TRIzol with 5 mg of acid-washed glass beads and gently mixed for 2 min. Then, 200 µl of phenol–chloroform was added, and the total mixture was incubated at room temperature for 15 min followed by a centrifugation at 10,000 g for 15 min at 4°C. The integrity of the RNA was determined by using a Nanodrop (260 nm/280 nm absorbance) and RNA concentration was brought to ∼200 ng µl−1 and stored at −80°C.
Targeted RNA-seq
In total, 45 honey bee genes involved in various functions (detoxification, immune defense and chemosensory roles, physiological development and nervous system regulation) were studied (Table S1). Approximately 200 bp of each exon was processed using the online primer picking program Batch Primer 3 (https://probes.pw.usda.gov/batchprimer3/) under the default setting for general primers to select forward and reverse primers that produce amplicons of 60–70 bp in length (You et al., 2008). The resulting primers were then tested in a single multiplex mixture using bee genomic DNA as the template. The genomic DNA was extracted using a MagJET system according to the manufacturer's directions (ThermoFisher). All amplifications from genomic DNA or cDNA were accomplished by Floodlight Genomics (Floodlight Genomics, Knoxville, TN, USA) through their no-cost Educational Outreach Program using an optimized Hi-plex approach (Nguyen-Dumont et al., 2013). Briefly, primers were mixed into a single multiplex mixture and amplified according to the parameters previously outlined for the Hi-plex approach. The resulting amplicons have a sample-specific barcode sequence incorporated during the PCR and all amplicons were pooled and a dual-index Illumina library constructed and quantified using a KAPA PCR-free kit according to the manufacturer's directions (Roche Sequencing and Life Sciences). Sequencing was accomplished on an Illumina HiSeq device running a 2×150 configuration (Novogene). The resulting sequences were mapped to the original target sequences using CLC Genomics Workbench version 9.5.2 (Qiagen) at default settings and the sequence coverage assessed manually to determine targets with no amplification.
cDNA synthesis and determination of relative gene expression
Total RNA was converted to cDNA using the Promega GoScript reverse transcription system (Madison, WI, USA) according to the manufacturer's directions. The resulting cDNA was used as a template for multiplex amplifications as described above. Each amplification was accomplished twice and an RNA template (no cDNA) reaction was included as a third reaction for each sample. The resulting amplicons were sequenced and mapped as described above. To normalize the data and determine the relative gene expression, the total number of reads mapping to each target was divided by the average sequence coverage of four housekeeping genes (Actin, CaMKII, GAPDH and E2F) (Scharlaken et al., 2008). The sequences of the gene-specific primers have been published in Scharlaken et al. (2008) and Alburaki et al. (2017a).
Statistical analysis
Statistical analyses and figure generation were conducted within the R environment (http://www.R-project.org/). Analysis of variance (ANOVA) was performed to study the difference between variables regarding the treatment at a 95% confidence level. Sequences were checked for their quality using FastQC software and normalized using four housekeeping genes as mentioned above. A few genes that were very weakly amplified were discarded from the dataset. Bioconductor package EdgeR (Robinson et al., 2010) version 3.6 was used in R environment version 3.4.3 for differential expression analyses of read counts arising from our RNA-seq data (Robinson and Oshlack, 2010). Simple list-based data objects (DGEList) were created using the function readDGE, and genes with very low counts were filtered not by direct count but with counts per million (cpm). RNA composition was normalized using the function CalcNormFactors by finding a set of scaling factors that minimize the log-fold change (logFC) between samples for most genes, and these scale factors were computed using a trimmed mean of M-values between each pair of samples (Robinson and Oshlack, 2010). Heatmaps were carried out using the library ComplexHeatmap of the package bioclite Limma. Other plots such as multi-dimensional scaling (MDS), hierarchical clustering and differential gene expression (DEG) were generated using their appropriate and respected functions in EdgeR. DEG data were calculated using EdgeR's Fisher's exact test after sample normalization. False discovery rates (FDRs) in DGEExact object were calculated at three different P-cutoffs (0.05, 0.01, 0.001) and their values were retained for each gene found to be upregulated or downregulated at 5%.
RESULTS
Nutrient consumption
Average syrup intake per week showed no differences among the groups until week 4, when bees in the 100 ppb imidacloprid treatment significantly reduced their syrup consumption (P<0.05, Fig. 2). Similar results were obtained when accounting for overall syrup consumption (P<0.01, Fig. 2). Interestingly, all groups treated with imidacloprid consumed a significantly (P<0.01) higher quantity of water than the control treatment group (0 ppb, Fig. 2), while protein patty consumption did not differ among treatments (data not shown). The correlation coefficients of syrup consumption and imidacloprid concentration indicated significant negative correlations in all treatment categories, particularly for 100 ppb (R=−0.15, P<0.001), with the exception of the 5 ppb treatment, in which no correlation was found (Fig. 2).
Fig. 2.

Syrup consumption of treatment groups during the 24 day experiment. Left, scatterplots showing daily syrup consumption and Pearson correlation coefficient (R) are provided for each treatment category. Right, weekly and overall syrup intake and water intake calculated per treatment. ANOVA was conducted at a 95% confidence level and error bars represent the s.e.m. (*P<0.05; **P<0.01).
Bee weight and mortality
The 1 day old sister bees used in cage and hive experiments had similar body mass at time 0, at week 3 and for overall mass (Fig. 3). However, significant differences between the mass of caged and hive bees were recorded at weeks 1, 2 and 4, with no differences among caged bees (Fig. 3). In the cage experiment, there were no significant differences in the total number of dead bees collected throughout the experiment (P=0.7) amongst the treatments. However, dead bees of the control treatment were significantly (P<0.05) greater in mass than those in all other treatment groups (Fig. 3). LC-MS pesticide residue analysis of the dead bees showed trace levels of imidacloprid and imidacloprid olfen only in bees of the 100 ppb treatment (Fig. 3).
Fig. 3.

Average mass of bees at time 0 and sampled weekly from cages and hive for the different treatment groups. Average mass and total number of dead bees collected daily from cages are also shown. LC-MS screening for four neonicotinoid molecules conducted on the total number of dead bees collected for each treatment is displayed in the table with values of the limit of detection LOD. ANOVA was conducted at a 95% confidence level (*P<0.05; **P<0.01; ***P<0.001).
Honey bee oxidative stress
At week 1, caged bees of the control treatment (0 ppb) showed a significantly higher level of hydrogen peroxide (P<0.001) than all other groups including the hive bees (Fig. 4). Protein carbonyl content was significantly higher in caged bees given the highest concentration of imidacloprid (100 ppb; P<0.05) at week 1. However, the same assay applied on samples at week 4 showed higher carbonyl content in bees of the 5 ppb group (Fig. 4). Similar results were obtained for whole-bee body protein conducted on samples at week 4 (Fig. 4).
Fig. 4.

Quantification of hydrogen peroxide and protein carbonyl content in caged and hive bees. The hydrogen peroxide assay was conducted on bees sampled at week 1 and the protein carbonyl assay was conducted on bees at week 1 and 4. Each boxplot represents an average of six biological replicates. Bovine serum albumin (BSA) data were not included in the statistical analysis. Error bars are quartiles and outliers; ANOVA levels of significance are *P<0.05 and ***P<0.001.
DEGs
In order to test the effect of both factors (imidacloprid and caging stress) on honey bee gene regulation, DEG analyses were carried out among caged bees only (imidacloprid versus control) and between caged and hive bees (caged versus hive). To gain an initial overview of the data, heatmaps for the whole dataset, including all samples, were generated. Major variation in color pattern was visually identifiable between the hive bee group and the rest of the samples (Fig. S1). We refined our analysis by running it on a weekly basis (Fig. 5). In the caged bees, imidacloprid showed no DEG at any time in the four studied weeks (Fig. S2). The only variation in gene regulation was found between caged and hive bees for both per-week (Fig. 5) and overall week data (Fig. S3). The biological coefficient of variance constantly distinguished two major groups of variables across the weeks with DEGs (in-hive bees versus caged bees, Fig. 5A–C). Heatmaps carried out weekly produced similar findings; genes with potentially different regulation are marked with asterisks in Fig. 5A–C. Based on their function (Table S1), antioxidant genes were the major DEGs in caged bees compared to hive bees in all weeks. At week 1, caged bees upregulated seven antioxidant genes and downregulated nine with no upregulation in the developmental and hormonal genes (Fig. 5A). Relatively similar regulation was observed in week 2 (Fig. 5B), with an increasing downregulation in all DEG categories up to week 4 (Fig. 5D). At week 4, upregulation of both immune defense and hormone genes depleted completely and a significant downregulation in all DEGs took place in caged bees. Gathering together the weekly DEG data in the caged bees, we identified constant upregulation of two antioxidant genes (Rsod and Trx-1) and downregulation of six others (Trx-2, Nmdar1, Vg, CSP3, AChE-2 and MsrA; Table S2; Fig. 6) Rsod is a poorly known gene that encodes an uncharacterized protein of 1123 amino acids, while Trx-1 is a gene encoding a major honey bee antioxidant of 136 amino acids and belongs to the thioredoxin family.
Fig. 5.
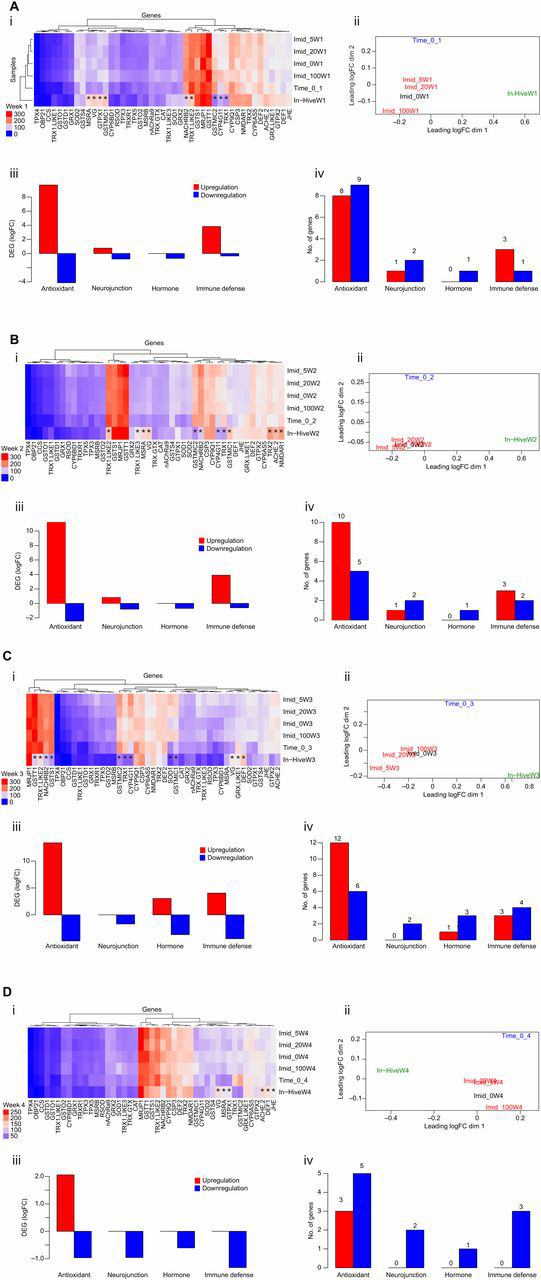
Honey bee gene regulation. (A–D) Data for weeks 1–4, respectively. (i) Heatmap of all samples and target genes (45) using moderated log counts per million. Asterisks represent genes differentially expressed in hive bees. (ii) Multi-dimensional scaling (MDS) or biological coefficient of variance of the samples based on the log fold-change (logFC) between each pair of RNA samples. (iii,iv) Data sorted by gene function for differential gene expression studies (DEG; iii) and the number of genes regulated (iv) in caged bees compared with hive bees in week 1.
Fig. 6.

Illustration of the DEG between caged and hive bees, emphasizing the cage stress factor. The gene regulation for caged bees and the top five significantly regulated genes (FDR<0.05) are displayed per week. Major potential functions for genes are described.
DISCUSSION
Honey bees are social insects, living together in highly organized colonies (Winston, 1987). Worker bees carry out different tasks throughout their developmental cycle from nurses to foragers, a behavioral maturation that is mainly governed by gene regulation (Grozinger et al., 2003; Robinson, 2002). Cage experiments offer a more controllable environment and are widely used in honey bee behavioral and toxicological studies. Nevertheless, little is known about the physiological and gene regulation changes in bees when caged and deprived of hive conditions. In this study, we found that imidacloprid induced a slight effect on honey bee diet behavior; however, an overwhelming difference in gene regulation was observed in bees deprived of hive conditions. Cohorts of caged sister bees consumed the same amount of syrup during the first 3 weeks of the study, regardless of the imidacloprid dose (0, 5, 20, 100 ppb) in the syrup. There were negative correlations between syrup ingestion and time for all concentrations except 5 ppb. This correlation was more pronounced at the highest dose (100 ppb, r=−0.15, P<0.001; Fig. 2), and during week 4, bees consumed significantly less syrup in the 100 ppb imidacloprid group. Consequently, overall consumption was less for syrup at the 100 ppb dose (P<0.05). It is not clear whether bees sensed the presence of imidacloprid in the tainted syrup and avoided it (Meikle et al., 2016) or whether a post-ingestive aversion response previously described in other invertebrates occurred (Behmer et al., 2005). Bees exposed to imidacloprid showed similar survival to the control group (P=0.7; Fig. 4). Despite surviving the ingestion of imidacloprid, symptoms of insecticide toxicity such as increased grooming and ventilation, disorientation, slower activity and difficulty in flying (Williamson et al., 2014) were clearly observed in the treated bees. Although only traces of imidacloprid and its metabolite molecule (imidacloprid olfen) were detected in bees fed 100 ppb, imidacloprid did induce significant protein damage in honey bees at weeks 1 and 4 (Figs 3 and 4). Surprisingly, and presumably as a mechanism to minimize the effects of exposure to pesticides, treated bees consumed more water than the control group (Fig. 2). Dead bees in the control group weighed more than those exposed to imidacloprid (P<0.05; Fig. 4), but there were no differences in the mass of living bees (Fig. 3). Pairing these results together, it appears that the evaporation of water content from the bodies of dead bees explains this contradictory finding.
While caged bees showed no weekly differences in mass, hive bees exhibited completely different patterns (Fig. 3). The mass patterns of hive bees clearly reflect their physiological development from nurse to forager bees. Hive bees were significantly heavier than caged bees at week 1 as well as week 2 (P<0.05), until becoming equal in mass at week 3. When they switched to foraging behavior at week 4, hive bees lost significant mass (P<0.001) compared with the relatively inactive caged bees (Fig. 3). Overall, our data indicate that caging bees induces a profound alteration in their physiological development compared with their sister-mates operating under hive conditions.
The DEG analysis showed a clear divergence in gene regulation of both bee sets (caged versus hive). Based on the genes with the five highest FDR values found in our DEG study, gene upregulation is the predominant response during the first 2 weeks in caged bees followed by overwhelming downregulation at weeks 3 and 4 (Fig. 5C,D), which points to potential depletion in caged bees compared with that of bees in the hive. Upregulation of the gene group involved in the antioxidative process was recorded in caged bees as compared with hive bees at all times and particularly during the first 3 weeks (Fig. 5A–D). Some of those genes belong to the thioredoxin family, such as Trx-1, Trx1-like2 and Trxr-1, as well as the glutaredoxin and glutathione families, all known to be involved with Apis cerana’s response to oxidative stress (Meng et al., 2014). Fewer genes coding for other functions were found to be differentially expressed in caged bees, such as Mrjp1, Fed1 and Vg (Table S2). Vitellogenin, a glycolipoprotein, which is often more abundant in nurse bees (Amdam et al., 2005; Ihle et al., 2009; Nelson et al., 2007), was among the six genes that were constantly downregulated (Vg, CSP3, Trx2, Nmdar1, AChE-2, MsrA) in caged bees throughout the 4 week experiment (Fig. 6). The honey bee genome contains 21 genes encoding odorant-binding proteins (Obps; The Honeybee Genome Sequencing Consortium, 2006) which, beside chemosensory proteins (CSPs), mediate both bee perception and release of chemical stimuli and probably have other functions (Iovinella et al., 2011). Obp21 was only upregulated in caged bees at week 3, while CSP3 was constantly downregulated, which suggest different functions of these genes. The constant downregulation of AChE suggests an absence of toxicity stress in caged bees as this gene was described in previous studies to be well linked to neonicotinoid toxicity (Alburaki et al., 2015; Badiou-Bénéteau et al., 2012). In this study, we identified two genes that were constantly upregulated in caged bees; Rsod, an uncharacterized gene, and thioredoxin (Trx1) (Fig. 6; Table S2). Rsod was described in Drosophila as an atypical member of the Cu/ZNSOD family with a highly unusual number of introns (18) and a duplicated SOD domain. Homologous genes were described in Apis, protozoa and fish but not found in mammals (Corona and Robinson, 2006). Nonetheless, the function of this gene (Rsod) remains unknown in insects. The thioredoxin genes (Trx) encode small and highly conserved oxidoreducatse proteins and are required to maintain redox homeostasis of the cell (Holmgren et al., 2005). Thus, we assume that the caging stress challenge may have triggered the bee biological system to mitigate this stress and prevent further cellular damage. This assumption is supported by the significant upregulation of the antioxidant genes and higher protein carbonyl content in caged bees compared with hive bees.
In conclusion, our study provides new insights into honey bee gene regulation when bees are exposed to cage stress compared with a typical hive environment. We showed that major honey bee antioxidants were constantly upregulated when bees are caged, including genes with uncharacterized function such as Rsod and Trx1. Furthermore, to the best of our knowledge, this is the first time that eight genes have been reported to potentially characterize the honey bee caging stress while conducting cage experiments. Our results add a significant contribution to the body of knowledge related to the effect of stressors on honey bee gene regulation.
Supplementary Material
Acknowledgements
We are grateful to Dr William Meikle and Milagra Weiss (USDA-ARS Carl Hayden Bee Research Center, Tucson, AZ, USA) for providing material support for this study.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Author contributions
Conceptualization: M.A.; Methodology: M.A., S.K.; Software: M.A., K.L.; Validation: M.A., S.K., S.D.S.; Formal analysis: K.L.; Investigation: J.A.; Resources: S.K., K.L., J.A.; Data curation: S.K., K.L., J.A., S.D.S.; Writing - original draft: M.A.; Writing - review & editing: M.A., S.K., K.L., J.A., S.D.S.; Supervision: S.K., J.A., S.D.S.; Project administration: S.D.S.; Funding acquisition: S.K., K.L., J.A., S.D.S.
Funding
This study was supported by the University of Tennessee, the University of Southern Mississippi and a USDA-ARS (U.S. Department of Agriculture–Agricultural Research Service) contract.
Supplementary information
Supplementary information available online at http://jeb.biologists.org/lookup/doi/10.1242/jeb.207761.supplemental
References
- Alburaki M., Boutin S., Mercier P.-L., Loublier Y., Chagnon M. and Derome N. (2015). Neonicotinoid-coated zea mays seeds indirectly affect honeybee performance and pathogen susceptibility in field trials. PLoS ONE 10, e0125790 10.1371/journal.pone.0125790 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alburaki M., Steckel S. J., Chen D., McDermott E., Weiss M., Skinner J. A., Kelly H., Lorenz G., Tarpy D. R., Meikle W. G. et al. (2017a). Landscape and pesticide effects on honey bees: forager survival and expression of acetylcholinesterase and brain oxidative genes. Apidologie 48, 556-571. 10.1007/s13592-017-0497-3 [DOI] [Google Scholar]
- Alburaki M., Steckel S. J., Williams M. T., Skinner J. A., Tarpy D. R., Meikle W. G., Adamczyk J. and Stewart S. D. (2017b). Agricultural landscape and pesticide effects on honey bee (Hymenoptera: Apidae) biological traits. J. Econ. Entomol. 110, 835-847. 10.1093/jee/tox111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aliouane Y., El Hassani A. K., Gary V., Armengaud C., Lambin M. and Gauthier M. (2009). Subchronic exposure of honeybees to sublethal doses of pesticides: effects on behavior. Environ. Toxicol. Chem. 28, 113-122. 10.1897/08-110.1 [DOI] [PubMed] [Google Scholar]
- Alptekin S., Bass C., Nicholls C., Paine M. J. I., Clark S. J., Field L. and Moores G. D. (2016). Induced thiacloprid insensitivity in honeybees (Apis mellifera L.) is associated with up-regulation of detoxification genes. Insect Mol. Biol. 25, 171-180. 10.1111/imb.12211 [DOI] [PubMed] [Google Scholar]
- Amdam G. V., Norberg K., Omholt S. W., Kryger P., Lourenço A. P., Bitondi M. M. G. and Simões Z. L. P. (2005). Higher vitellogenin concentrations in honey bee workers may be an adaptation to life in temperate climates. Insectes Soc. 52, 316-319. 10.1007/s00040-005-0812-2 [DOI] [Google Scholar]
- Amdam G. V., Norberg K., Page R. E., Erber J. and Scheiner R. (2006). Downregulation of vitellogenin gene activity increases the gustatory responsiveness of honey bee workers (Apis mellifera). Behav. Brain Res. 169, 201-205. 10.1016/j.bbr.2006.01.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aronstein K. and Saldivar E. (2005). Characterization of a honey bee Toll related receptor gene Am18w and its potential involvement in antimicrobial immune defense. Apidologie 36, 3-14. 10.1051/apido:2004062 [DOI] [Google Scholar]
- Badiou-Bénéteau A., Carvalho S. M., Brunet J.-L., Carvalho G. A., Buleté A., Giroud . and Belzunces L. P. (2012). Development of biomarkers of exposure to xenobiotics in the honey bee Apis mellifera: application to the systemic insecticide thiamethoxam. Ecotoxicol. Environ. Saf. 82, 22-31. 10.1016/j.ecoenv.2012.05.005 [DOI] [PubMed] [Google Scholar]
- Barnett A. E., Charlton J. A. and Fletcher R. M. (2007). Incidents of bee poisoning with pesticides in the United Kingdom, 1994-2003. Pest Manag. Sci. 63, 1051-1057. 10.1002/ps.1444 [DOI] [PubMed] [Google Scholar]
- Behmer S. T., Lloyd C. M., Raubenheimer D., Stewart-Clark J., Knight J., Leighton R. S., Harper F. A. and Smith J. A. C. (2005). Metal hyperaccumulation in plants: mechanisms of defence against insect herbivores. Funct. Ecol. 19, 55-66. 10.1111/j.0269-8463.2005.00943.x [DOI] [Google Scholar]
- Bordier C., Dechatre H., Suchail S., Peruzzi M., Soubeyrand S., Pioz M., Pélissier M., Crauser D., Le Conte Y. and Alaux C. (2017). Colony adaptive response to simulated heat waves and consequences at the individual level in honeybees (Apis mellifera). Sci. Rep. 7, 3760 10.1038/s41598-017-03944-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Briand L., Swasdipan N., Nespoulous C., Bézirard V., Blon F., Huet J.-C., Ebert P. and Pernollet J.-C. (2002). Characterization of a chemosensory protein (ASP3c) from honeybee (Apis mellifera L.) as a brood pheromone carrier. Eur. J. Biochem. 269, 4586-4596. 10.1046/j.1432-1033.2002.03156.x [DOI] [PubMed] [Google Scholar]
- Buttstedt A., Moritz R. F. A. and Erler S. (2014). Origin and function of the major royal jelly proteins of the honeybee (Apis mellifera) as members of the yellow gene family. Biol. Rev. Camb. Philos. Soc. 89, 255-269. 10.1111/brv.12052 [DOI] [PubMed] [Google Scholar]
- Chakrabarti P., Rana S., Sarkar S., Smith B. and Basu P. (2015). Pesticide-induced oxidative stress in laboratory and field populations of native honey bees along intensive agricultural landscapes in two Eastern Indian states. Apidologie 46, 107-129. 10.1007/s13592-014-0308-z [DOI] [Google Scholar]
- Chomczynski P. (1993). A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. BioTechniques 15, 532-534. [PubMed] [Google Scholar]
- Corona M. and Robinson G. E. (2006). Genes of the antioxidant system of the honey bee: annotation and phylogeny. Insect Mol. Biol. 15, 687-701. 10.1111/j.1365-2583.2006.00695.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cutler G. C. and Scott-Dupree C. D. (2007). Exposure to clothianidin seed-treated canola has no long-term impact on honey bees. J. Econ. Entomol. 100, 765-772. 10.1603/0022-0493(2007)100[765:ETCSCH]2.0.CO;2 [DOI] [PubMed] [Google Scholar]
- Decourtye A., Armengaud C., Renou M., Devillers J., Cluzeau S., Gauthier M. and Pham-Delègue M.-H. (2004). Imidacloprid impairs memory and brain metabolism in the honeybee (Apis mellifera L.). Pestic. Biochem. Physiol. 78, 83-92. 10.1016/j.pestbp.2003.10.001 [DOI] [Google Scholar]
- Evans J. D. and Wheeler D. E. (1999). Differential gene expression between developing queens and workers in the honey bee, Apis mellifera. Proc. Natl Acad. Sci. USA 96, 5575-5580. 10.1073/pnas.96.10.5575 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans J. D., Aronstein K., Chen Y. P., Hetru C., Imler J.-L., Jiang H., Kanost M., Thompson G. J., Zou Z. and Hultmark D. (2006). Immune pathways and defence mechanisms in honey bees Apis mellifera. Insect Mol. Biol. 15, 645-656. 10.1111/j.1365-2583.2006.00682.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giannini T. C., Cordeiro G. D., Freitas B. M., Saraiva A. M. and Imperatriz-Fonseca V. L. (2015). The dependence of crops for pollinators and the economic value of pollination in Brazil. J. Econ. Entomol. 108, 849-857. 10.1093/jee/tov093 [DOI] [PubMed] [Google Scholar]
- Gong Y. H. and Diao Q. Y. (2017). Current knowledge of detoxification mechanisms of xenobiotic in honey bees. Ecotoxicology 26, 1-12. 10.1007/s10646-016-1742-7 [DOI] [PubMed] [Google Scholar]
- Gregorc A., Evans J. D., Scharf M. and Ellis J. D. (2012). Gene expression in honey bee (Apis mellifera) larvae exposed to pesticides and Varroa mites (Varroa destructor). J. Insect Physiol. 58, 1042-1049. 10.1016/j.jinsphys.2012.03.015 [DOI] [PubMed] [Google Scholar]
- Gregorc A., Alburaki M., Werle C., Knight P. R. and Adamczyk J. (2017). Brood removal or queen caging combined with oxalic acid treatment to control varroa mites (Varroa destructor) in honey bee colonies (Apis mellifera). Apidologie 48, 821-832. 10.1007/s13592-017-0526-2 [DOI] [Google Scholar]
- Gregorc A., Alburaki M., Sampson B., Knight P. R. and Adamczyk J. (2018). Toxicity of selected acaricides to honey bees (Apis mellifera) and Varroa (Varroa destructor Anderson and Trueman) and their use in controlling varroa within honey bee colonies. Insects 9, 55 10.3390/insects9020055 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grozinger C. M., Sharabash N. M., Whitfield C. W. and Robinson G. E. (2003). Pheromone-mediated gene expression in the honey bee brain. Proc. Natl. Acad. Sci. USA 100, 14519-14525. 10.1073/pnas.2335884100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmgren A., Johansson C., Berndt C., Lönn M. E., Hudemann C. and Lillig C. H. (2005). Thiol redox control via thioredoxin and glutaredoxin systems. Biochem. Soc. Trans. 33, 1375-1377. 10.1042/BST0331375 [DOI] [PubMed] [Google Scholar]
- Hoover S. E. R., Keeling C. I., Winston M. L. and Slessor K. N. (2003). The effect of queen pheromones on worker honey bee ovary development. Naturwissenschaften 90, 477-480. 10.1007/s00114-003-0462-z [DOI] [PubMed] [Google Scholar]
- Ihle K. E., Page R. E., Fondrk M. K. and Amdam G. V. (2009). Vitellogenin modulates foraging behavior in selected strains of honey bees (Apis mellifera). Integr. Comp. Biol. 49, E247-E247. [Google Scholar]
- Iovinella I., Dani F. R., Niccolini A., Sagona S., Michelucci E., Gazzano A., Turillazzi S., Felicioli A. and Pelosi P. (2011). Differential expression of odorant-binding proteins in the mandibular glands of the honey bee according to caste and age. J. Proteome Res. 10, 3439-3449. 10.1021/pr2000754 [DOI] [PubMed] [Google Scholar]
- Li C. C., Xu B. H., Wang Y. X., Yang Z. B. and Yang W. R. (2014). Protein content in larval diet affects adult longevity and antioxidant gene expression in honey bee workers. Entomol. Exp. Appl. 151, 19-26. 10.1111/eea.12167 [DOI] [Google Scholar]
- Liu S. C., Liu F., Jia H. H., Yan Y., Wang H. F., Guo X. Q. and Xu B. H. (2016). A glutathione S-transferase gene associated with antioxidant properties isolated from Apis cerana cerana. Sci. Nat. 103, 43 10.1007/s00114-016-1362-3 [DOI] [PubMed] [Google Scholar]
- Mao W. F., Schuler M. A. and Berenbaum M. R. (2011). CYP9Q-mediated detoxification of acaricides in the honey bee (Apis mellifera). Proc. Natl. Acad. Sci. USA 108, 12657-12662. 10.1073/pnas.1109535108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meikle W. G., Adamczyk J. J., Weiss M., Gregorc A., Johnson D. R., Stewart S. D., Zawislak J., Carroll M. J. and Lorenz G. M. (2016). Sublethal effects of imidacloprid on honey bee colony growth and activity at three sites in the U.S. PLoS ONE 11, e0168603 10.1371/journal.pone.0168603 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meng F., Zhang Y. Y., Liu F., Guo X. Q. and Xu B. H. (2014). Characterization and mutational analysis of omega-class GST (GSTO1) from Apis cerana cerana, a gene involved in response to oxidative stress. PLoS ONE 9, e93100 10.1371/journal.pone.0093100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morandin L. A., Laverty T. M. and Kevan P. G. (2001). Effect of bumble bee (Hymenoptera: Apidae) pollination intensity on the quality of greenhouse tomatoes. J. Econ. Entomol. 94, 172-179. 10.1603/0022-0493-94.1.172 [DOI] [PubMed] [Google Scholar]
- Mussig L., Richlitzki A., Rossler R., Eisenhardt D., Menzel R. and Leboulle G. (2010). Acute disruption of the NMDA receptor subunit NR1 in the honeybee brain selectively impairs memory formation. J. Neurosci. 30, 7817-7825. 10.1523/JNEUROSCI.5543-09.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson C. M., Ihle K. E., Fondrk M. K., Page R. E. and Amdam G. V. (2007). The gene vitellogenin has multiple coordinating effects on social organization. PLoS Biol. 5, 673-677. 10.1371/journal.pbio.0050062 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nguyen-Dumont T., Pope B. J., Hammet F., Southey M. C. and Park D. J. (2013). A high-plex PCR approach for massively parallel sequencing. BioTechniques 55, 69-74. 10.2144/000114052 [DOI] [PubMed] [Google Scholar]
- Richard F.-J., Holt H. L. and Grozinger C. M. (2012). Effects of immunostimulation on social behavior, chemical communication and genome-wide gene expression in honey bee workers (Apis mellifera). BMC Genomics 13, 558 10.1186/1471-2164-13-558 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson G. E. (2002). Genomics and integrative analyses of division of labor in honeybee colonies. Am. Nat. 160 Suppl. 6, S160-S172. 10.1086/342901 [DOI] [PubMed] [Google Scholar]
- Robinson M. D. and Oshlack A. (2010). A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 11, R25 10.1186/gb-2010-11-3-r25 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson G. E., Winston M. L., Huang Z.-Y. and Pankiw T. (1998). Queen mandibular gland pheromone influences worker honey bee (Apis mellifera L.) foraging ontogeny and juvenile hormone titers. J. Insect Physiol. 44, 685-692. 10.1016/S0022-1910(98)00040-7 [DOI] [PubMed] [Google Scholar]
- Robinson M. D., McCarthy D. J. and Smyth G. K. (2010). edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139-140. 10.1093/bioinformatics/btp616 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sampson B. J. and Cane J. H. (2000). Pollination efficiencies of three bee (Hymenoptera: Apoidea) species visiting rabbiteye blueberry. J. Econ. Entomol. 93, 1726-1731. 10.1603/0022-0493-93.6.1726 [DOI] [PubMed] [Google Scholar]
- Sanchez-Bayo F. and Goka K. (2014). Pesticide residues and bees--a risk assessment. PLoS One 9, e94482 10.1371/journal.pone.0094482 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scharlaken B., de Graaf D. C., Goossens K., Brunain M., Peelman L. J. and Jacobs F. J. (2008). Reference gene selection for insect expression studies using quantitative real-time PCR: The head of the honeybee, Apis mellifera, after a bacterial challenge. J. Insect Sci. 8, 33 10.1673/031.008.3301 [DOI] [Google Scholar]
- Stewart S. D., Lorenz G. M., Catchot A. L., Gore J., Cook D., Skinner J., Mueller T. C., Johnson D. R., Zawislak J. and Barber J. (2014). Potential exposure of pollinators to neonicotinoid insecticides from the use of insecticide seed treatments in the mid-southern United States. Environ. Sci. Technol. 48, 9762-9769. 10.1021/es501657w [DOI] [PubMed] [Google Scholar]
- Stokstad E. (2017). European bee study fuels debate over pesticide ban. Science 356, 1321-1321. 10.1126/science.356.6345.1321 [DOI] [PubMed] [Google Scholar]
- The Honeybee Genome Sequencing Consortium. (2006). Insights into social insects from the genome of the honeybee Apis mellifera. Nature 443, 931-949. 10.1038/nature05260 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tosi S., Burgio G. and Nieh J. C. (2017). A common neonicotinoid pesticide, thiamethoxam, impairs honey bee flight ability. Sci. Rep. 7, 1201 10.1038/s41598-017-01361-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walorczyk S. and Gnusowski B. (2009). Development and validation of a multi-residue method for the determination of pesticides in honeybees using acetonitrile-based extraction and gas chromatography-tandem quadrupole mass spectrometry. J. Chromatogr. A 1216, 6522-6531. 10.1016/j.chroma.2009.07.045 [DOI] [PubMed] [Google Scholar]
- Williamson S. M., Moffat C., Gomersall M. A. E., Saranzewa N., Connolly C. N. and Wright G. A. (2013). Exposure to acetylcholinesterase inhibitors alters the physiology and motor function of honeybees. Front. Physiol. 4, 13 10.3389/fphys.2013.00013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williamson S. M., Willis S. J. and Wright G. A. (2014). Exposure to neonicotinoids influences the motor function of adult worker honeybees. Ecotoxicology 23, 1409-1418. 10.1007/s10646-014-1283-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winston M. L. (1987). The Biology of the Honey Bee. Massachusetts: Harvard University Press, Cambridge. [Google Scholar]
- Winston M. L., Higo H. A., Colley S. J., Pankiw T. and Slessor K. N. (1991). The role of queen mandibular pheromone and colony congestion in honey bee (Apis mellifera L.) reproductive swarming (Hymenoptera: Apidae). J. Insect Behav. 4, 649-660. 10.1007/BF01048076 [DOI] [Google Scholar]
- Woodcock B. A., Bullock J. M., Shore R. F., Heard M. S., Pereira M. G., Redhead J., Ridding L., Dean H., Sleep D., Henrys P. et al. (2017). Country-specific effects of neonicotinoid pesticides on honey bees and wild bees. Science 356, 1393-1395. 10.1126/science.aaa1190 [DOI] [PubMed] [Google Scholar]
- You F. M., Huo N., Gu Y. Q., Luo M.-C., Ma Y., Hane D., Lazo G. R., Dvorak J. and Anderson O. D. (2008). BatchPrimer3: a high throughput web application for PCR and sequencing primer design. BMC Bioinformatics 9, 253 10.1186/1471-2105-9-253 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.