

Abstract
Background
Herpes simplex virus (HSV) can cause encephalitis. Its infected cell polypeptide 47 (ICP47), encoded by immediate-early gene US12, promotes immune escape. ICP47 was modified in the clinically approved oncolytic HSV (oHSV) T-Vec. However, transcription regulatory sequence (TRS) and transcription regulatory factor (TRF) of HSV US12 are seldom reported.
Methods
Previously, our laboratory isolated a new HSV strain named HSV-1-LXMW from a male patient with oral herpes in Beijing, China. Firstly, the genetic tree was used to analyze its genetic relationship. The US12 TRS and TRF in HSV-1-LXMW were found by using predictive software. Secondly, the further verification by the multi-sequence comparative analysis shown that the upstream DNA sequence of HSV US12 gene contained the conserved region. Finally, the results of literature search shown that the expression of transcription factors was related to the tissue affinity of HSV-1 and HSV-2, so as to increase the new understanding of the transcriptional regulation of HSV biology and oncolytic virus (OVs) therapy.
Results
Here we reported the transcriptional regulation region sequence of our new HSV-1-LXMW, and its close relationship with HSV-1-CR38 and HSV-1-17. Importantly we identified eight different kinds of novel TRSs and TRFs of HSV US12 for the first time, and found they are conserved among HSV-1 (c-Rel, Elk-1, Pax-4), HSV-2 (Oct-1, CF2-II, E74A, StuAp) or both HSVs (HNF-4). The TRFs c-Rel and Oct-1 are biologically functional respectively in immune escape and viral replication during HSV infection.
Conclusions
Our findings have important implication to HSV biology, infection, immunity and oHSVs.
Keywords: HSV-1, HSV-2, US12, ICP47, Transcriptional regulation sequence (TRS), Transcriptional regulation factor (TRF)
Background
Tumors are heterogeneous and often resistant to chemotherapy and radiotherapy [1, 2], and no single treatment could be widespread applied or has full effectivity for cancer treatment [3–6]. OVs treatment are different from conventional chemotherapy and radiotherapy, and could provide additional treatment strategies [7]. Additionally, OVs are diverse in structure and biology, which spread among tumors with different kinetics and kill tumor cells through multiple mechanisms [8]. The oHSV T-VEC has been approved by FDA for patients with melanoma [9–11]. HSV, a member of the alpha-herpesviruses subfamily, which is an encapsulated DNA virus, offers particular benefits for use as a gene transfer vector, contains at least 120 kb of double-stranded DNA genome, encoding more than 70 genes [7, 12]. Type 1 and 2 HSVs (HSV-1 and HSV-2) are the most common and acute human pathogens. HSV-1 is normally related to oral-facial infections and may cause encephalitis in severe cases, while HSV-2 mostly induces genital infections and could cause mother-to-child transmission [13, 14].
HSV as an OVs has many favourable properties. Engineered oHSVs have been shown to be remarkably safe in clinical trials, and also have some evidence of their effectiveness [12]. Dlsptk, the first recombinant HSV, was generated by deleting the gene UL23 encoding thymidine kinase (TK) [15]. The selectivity and efficacy of dlsptk established a principled proof for the application of HSV-1 genome deletions to carry out the tumor selectivity. However, from the standpoint of clinical application, the UL23 deletion was eventually problematic for it causes dlsptk impervious to first-line anti-herpes pharmaceuticals, resulting in abandance of dlsptk. The second recombinant HSV, G207, is the first oHSV tested in clinical trials and it deletes the genes UL39 (ICP6) and RL1 (ICP34.5). As is well-known, ICP34.5, a key factor of HSV neurovirulence, can preclude the shut-off of protein synthesis in infected host cells. ICP6 is a determinant viral enzyme for HSV DNA synthesis, which is indispensable for virus replication in normal non-dividing cells [16]. Dlsptk and G207 are designed to weaken viral replication and reduce viral virulence in non-cancer cells. The third generation oHSV-1 vector, G47Δ, is based on G207 with additional ICP47 deletion, which surprisingly enhances viral replication and increases immune recognition of infected cells [17]. ICP47 deletion contains the promoter region of US11 and also attenuates γ34.5 growth [18]. Importantly, because of ICP47 can block peptide loading of major histocompatibility complex I (MHC-I) molecules, G47∆ has induced an antitumor immune response for the ICP47 deletion. Therefore, increasing MHC I antigen presentation, stimulating cytotoxic lymphocytes and reducing NK cytolysis of infected cells can enhance anti-tumor immune response. Anti-tumor immune responses may be the key for the treatment of tumor metastasis .
ICP47, encoded by gene US12, is a polymorphous protein and could block RNA splicing in early infection, and then, shuttle viral mRNA from nucleus to cytoplasm in late infection [19]. ICP47 directly binds antigen-dependent transporter (TAP), limiting antigen trafficing, leading to the occurrence of empty MHC-I [20]. The functional domain of ICP47 has been mapped to 35 residues at the N-terminal, forming an extended helix-loop-helix structure in the lipid bilayer [20]. In addition, since ICP47 is too large to be easily transported by TAP, its high affinity binding traps TAP in an inactive conformation [21]. The binding of ICP47 stabilizes the inward conformation, and therefore blocks TAP from transitioning to the outward state in which the nucleotide binding domains (NBDs) form a closed dimer and the translocation pathway points to the endoplasmic reticulum (ER) cavity [22]. By blocking the entry of viral antigens into ER, HSV could avoid the attrack of cytotoxic T lymphocytes (CTLs), which may lead to immune escape of HSV and establish lifelong infection in the host cells. Interestingly, the ICP47 in G47Δ is deleted, and this keeps the cell surface MHC-I-antigen expression and allows to enhance antigen presentation [18]. Furthermore, G47∆ has been proved effective in animal tumor models of various cancers such as brain cancer, prostate cancer, breast cancer, schwannoma and human melanoma [23–25].
Currently, HSV US12 is widely used in OVs modification, gene therapy and vaccine construction [25–27]. However, there are no reports on TRS and TRF of US12 gene in HSV. As an immediate-early protein, its expression is regulated by the tri-partite Oct-1/HCF/VP16 complex [28, 29]. Identification of additional conserved promoter regulatory sequences that might further regulate its expression is certainly an important question. Here we sequenced the transcriptional regulation region of US12 of our new HSV-1 strain LXMW, and for the first time identified novel TRS and TRF of HSV US12. These findings may have important impactions for HSV biology, infection, immunity and OVs.
Methods
Previously, our laboratory isolated a new HSV strain named HSV-1-LXMW from a male patient with oral herpes in Beijing, China [30]. The detailed content of Cells, HSV-1 isolation and identification, and HSV genomic DNA sequencing analysis have been elaborated [30].
Identification of the US12 potential transcriptional regulation region sequences in HSV
The online program NCBI (National Center for Biotechnology Information: https://www.ncbi.nlm.nih.gov/) was used to determine US12 potential transcriptional regulation region sequences.
Phylogenetic analysis of the transcriptional regulation region of the US12 gene
We used MEGA7 (https://www.megasoftware.net/), the application (APP), to analyze phylogenetic relationship.
Prediction of US12 transcription regulatory sequences and factors
We used online program Match (http://gene-regulation.com/pub/programs.html) to predict the gene US12 TRS and TRF according to their instruction.
Alignment of the transcriptional regulation region sequences of US12
ApE (A plasmid Editor: http://biologylabs.utah.edu/jorgensen/wayned/ape/), the application (APP), was used to make the potential transcriptional regulation region sequences alignment of gene US12 according to their manual.
Results
Identification of the US12 transcription regulatory region of HSV-1-LXMW
Using the nucleotide sequence database, we identified transcription regulatory regions of US12 (145851–148,050) as shown in Fig. 1. Please refer to supplementary material for the sequence of the transcription regulatory region. The transcription regulatory regions are 2000 bp upstream and 200 bp downstream of US12 transcription initiation sites. Interestingly, the gene that encodes ICP47 is US10, not US12 in HSV-2 strain HG52 and H1226. We summarized information about the HSV US12 genomic DNA transcription regulatory regions of our new strain and 11 other strains studied in this article (Table 1).
Fig. 1.

Transcription regulatory regions of US12. a. Schematic of the HSV-1 genome showing the regions of US12. The HSV-1 genome consists of long and short unique regions (UL and US) each bounded by terminal (T) and internal (I) repeat regions (RL and RS). b. The DNA sequence of the US12 gene is marked in green and the transcription regulatory regions of US12 is marked in blue
Table 1.
HSV US12 genomic DNA sequencing
| HSV Strain | Gene Bank ID | Tax-ID | Sub-Date | US12 transcription regulatory regions | University, Country |
|---|---|---|---|---|---|
| HSV-1 strain XLMW | 145,851 / 148,050 | Yangtze University, Jingzhou, China | |||
| HSV-1 strain 17 | JN555585 | 10,299 | 2011-08-02 | 145,869 /148,068 | RC University, Glasgow, UK |
| HSV-1 strain H129 | GU734772 | 744,249 | 2010-01-18 | 145,769/ 147,968 | Princeton University, USA |
| HSV-1 Strain CR38 | HM585508 | 10,298 | 2013-10-17 | 145,315 / 147,514 | MRC Virology Unit, UK |
| HSV-1 strain SC16 | KX946970 | 10,309 | 2016-10-30 | 113,629/114,168 | Severo Ochoa, Spain |
| HSV-1 strain KOS | JQ673480 | 10,306 | 2012-03-10 | 145,672 /147,871 | University of Kansas, USA |
| HSV-1 strain Patton isolate | MF959544 | 10,308 | 2017-10-11 | 146,470 / 148,669 | NYU, New York, USA |
| HSV-1 strain E19 | HM585511 | 10,298 | 2013-10-22 | 144,988 / 147,187 | University of Glasgow Centre for Virus Research, UK |
| HSV-1 strain F | GU734771 | 10,304 | 2010-01-18 | 145,795 / 147,994 | Princeton University, USA |
| HSV-2 strain SD90e | KF781518 | 1,177,628 | 2013-10-25 | 147,503 / 149,702 | Harvard Medical School, Boston, US |
| HSV-2 strain HG52 | JN561323 | 10,310 | 2011-08-05 | 147,834 / 150,033 | University of Glasgow, UK |
| HSV-2 strain H1226 | KY922720 | 16,866 | 2017-09-27 | 147,094 /149,293 | Pennslyvania State University, USA |
Phylogenetic analysis of HSV-1-LXMW and other 11 HSV strains
Based on the gene US12 transcription regulatory region sequences in Table 1 of HSV-1-LXMW and other 11 HSV strains, including 8 HSV-1 strains (17, CR38, H129, SC16, KOS, Patton, E19 and F) and 3 HSV-2 strains (SD90e, HG52 and H1226), the phylogenetic analysis about the evolutionary relationship among HSV-1-LXMW and other 11 HSV strains were performed. The result shown high homology among our new strain HSV-1-LXMW and strains HSV-1-CR38, HSV-1-17 and HSV-1-H129. Our data again support that HSV-1-LXMW is a strain of HSV-1 (Fig. 2).
Fig. 2.

Phylogenetic analysis of HSV-1-LXMW together with 11 other HSV strains. Evolutionary analyses were conducted in MEGA7. The evolutionary history was inferred by using the Maximum Likelihood method based on the Tamura-Nei model. The bootstrap consensus tree inferred from 1000 replicates is taken to represent the evolutionary history of the taxa analyzed. Branches corresponding to partitions reproduced in less than 50% bootstrap replicates are collapsed
Identification of the US12 TRS and TRF
Better understanding of US12 transcriptional regulation is crucial for HSV biology and antitumor immune responses of oHSVs. Using Match, the online program, we find four major TRS of HSV-1-LXMW, which bind to c-Rel, HNF-4, ElK-1 and Pax-4, and three of HSV-1-17, which bind to c-Rel, HNF-4 and Pax-4. Interestingly, compared with the TRF of HSV-2, the difference between HSV-1 and HSV-2 is quite large. We find five major different kinds of TRSs of HSV-2-SD90e, which bind to HNF-4, CF2-II, E74A, Oct-1 and StuAp (Table 2).
Table 2.
The US12 TRS and TRF in HSVs
| HSV strain | Matrix identifier | Position strand | Core match | Matrix match | Sequence | Factor name |
|---|---|---|---|---|---|---|
| HSV-1 strain XLMW | V$CREL_01 | 37 (+) | 1.000 | 0.982 | gggtcTTTCC | c-Rel |
| V$HNF4_01 | 1020 (−) | 0.883 | 0.898 | ccctgtcCTTTTtcccacc | HNF-4 | |
| V$ELK1_02 | 1875(+) | 1.000 | 0.984 | ggcgcCGGAAgccc | Elk-1 | |
| V$PAX4_01 | 2029 (−) | 0.888 | 0.833 | gccacgggccgCTTCAcggcc | Pax-4 | |
| HSV-1 strain 17 | V$CREL_01 | 37 (+) | 1.000 | 0.982 | gggtcTTTCC | c-Rel |
| V$HNF4_01 | 1023 (−) | 0.883 | 0.898 | ccctgtcCTTTTtcccacc | HNF-4 | |
| V$PAX4_01 | 2032 (−) | 0.888 | 0.833 | gccacgggccgCTTCAcggcc | Pax-4 | |
| HSV-2 strain SD90e | V$HNF4_01 | 746 (−) | 1.000 | 0.928 | gctcgcaCTTTGccctaat | HNF-4 |
| I$CF2II_01 | 767 (−) | 1.000 | 1.000 | tatATATAc | CF2-II | |
| V$HNF4_01 | 886 (−) | 1.000 | 0.928 | gctcgcaCTTTGccctaat | HNF-4 | |
| I$CF2II_01 | 907 (−) | 1.000 | 1.000 | tatATATAc | CF2-II | |
| I$E74A_01 | 1076(+) | 1.000 | 0.954 | cgaaccCGGAAgggcag | E74A | |
| V$OCT1_Q6 | 1120 (−) | 0.883 | 0.911 | ctcaTTAGCatcgcg | Oct-1 | |
| F$STUAP_01 | 1637 (−) | 1.000 | 1.000 | ggtCGCGAtg | StuAp |
Further analysis of three more HSV-1 strains found that their TRS and TRF binding sites are similar with HSV-1-LXMW, but have minor differences (Fig. 3). Comparing to HSV-1-LXMW, there is no c-Rel binding site for HSV-1-SC16, no Elk-1 binding site for HSV-1-Patton and 17, and no Pax-4 binding site for HSV-1-Patton and E19. There is only one US12 TRS binding to HNF-4 in all the HSV-1 strains analyzed, but there are two HNF-4 binding sites in HSV-2 strains.
Fig. 3.

The US12 TRSs and TRFs in HSVs. TRFs are represented in different colors, and the number represents the specific location of their binding TRFs
The TRS and TRF are conserved
Conserved sequences refer to highly similar or identical nucleic acid sequences (RNA or DNA sequences), protein sequences, and their structures. Conserved sequences generally have functional value. Here, we found that the US12 TRSs and TRF binding sites are conserved among the 9 HSV-1 and 3 HSV-2 strains (Fig. 4), indicating these conserved TRSs and TRFs are likely to be biologically functional.
Fig. 4.
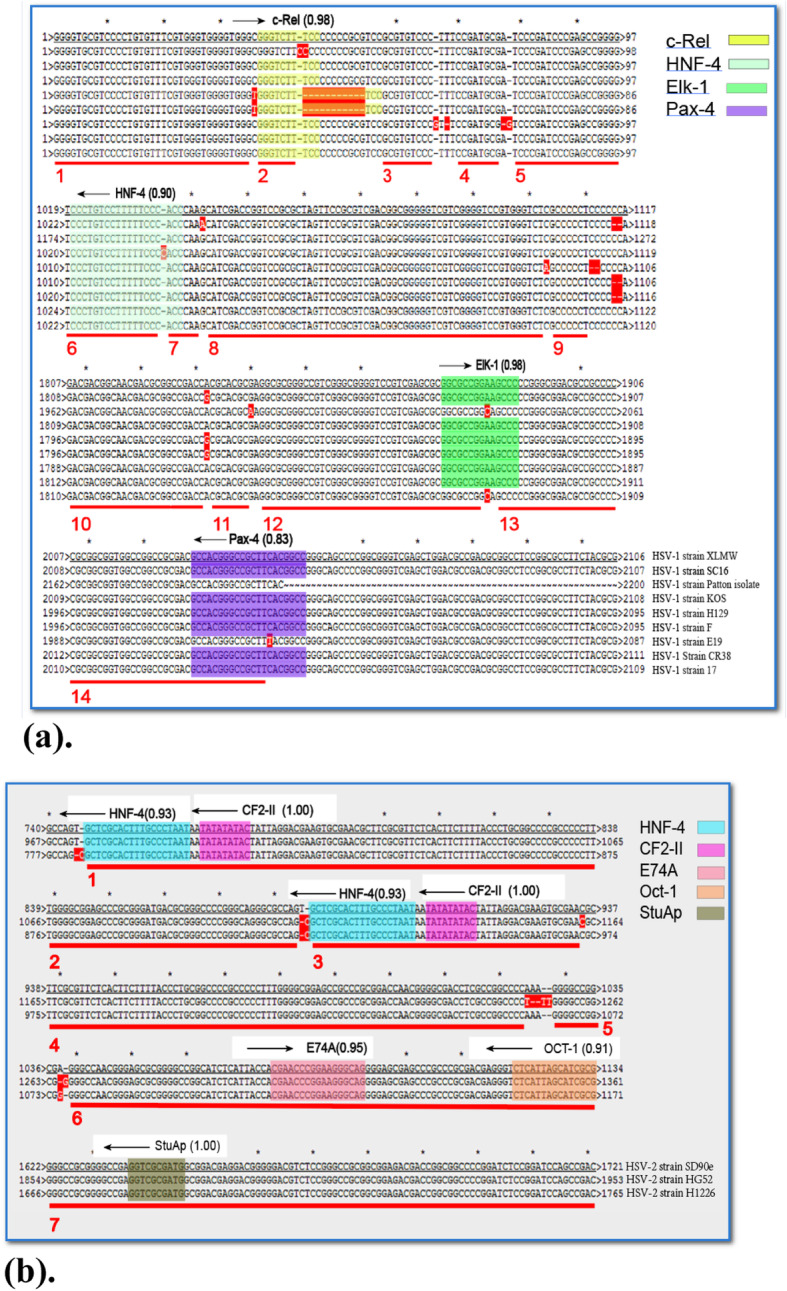
a. The US12 TRSs and TRFs are conserved among HSV-1 strains. Red lines indicate the conserved regions 1–14. The TRSs and TRFs are shown in colored boxes. b The US12 TRSs and TRFs are identical among HSV-2 strains. Red lines indicate the conserved regions 1–17. The TRSs and TRFs are shown in colored boxes
Our multi-sequence alignment results indicated that in the US12 transcriptional regulation regions there are less mutations among HSV-1 strains than mutations between HSV-1 and HSV-2. Between HSV-1-LXMW and HSV-1-172,184 base pairs are matched and only 5 base pairs are mismatched. However, there are only 468 base pairs matched between the HSV-1- LXMW and HSV-2-SD90e. Therefore, we decided to compare HSV type 1 and type 2 separately in ApE Program (Aligment parameters: Blocks: 10, mismatch penalty: 0, gap penalty: 0, gap Ext penalty: 0, everything else is at default). Results shown that our new strain HSV-1-LXMW was highly similar to HSV-1-17 and HSV-1-CR38 (Fig. 4a).
The alignment of the transcription regulatory region (nucleotides 1–2200) of HSV-1-XLMW strain to other 8 HSV-1 strains SC16, Patton, KOS, H129, F, E19, CR38 and 17, respectively, shown 2177, 2023, 2182, 2165, 2167, 2150, 2184 and 2184 matched base pairs, 11, 6, 6, 11, 11, 10, 3 and 5 mismatched base pairs, and 24, 324, 26, 48, 44, 80, 26 and 21 base pair gaps (Fig. 4a). The alignment of the transcription regulatory region (nucleotides 1–2200) of HSV-2- SD90e strain to other 2 HSV-2 strains HG52 and H1226, respectively, shown 1956 and 2128 matched base pairs, 3 and 2 mismatched base pairs, and 480 and 138 base pair gaps (Fig. 4b).
A sequence with five or more conserved base pairs is defined as a conservative region. From the alignment analysis, we found 14 conserved regions of 9 HSV-1 strains and 7 conserved regions of 3 HSV-2 strains (Fig. 4). The conserved regions of HSV-1-LXMW US12 started respectively at 1, 37, 58, 70, 79, 1019, 1036, 1043, 1104, 1807, 1833, 1842, 1886, and 2007 base pairs. Interestingly, we found that most of the US12 transcriptional regulation regions are overlapped with the TRSs identified above.
There are four TRFs in HSV-1-LXMW, KOS, H129, F and CR38 strains and three TRFs in HSV-1-SC16, E19 and 17 strains and two transcription factors in HSV-1-patton strain. Additionally, the TRFs binding sites of the HSV-1 strains are basically conserved (Fig. 4a). Interestingly, TRSs in the US12 transcriptional regulatory region are identical in HSV-2 strains SD90e, HG52 and H1226 (Fig. 4b). Importantly, binding sites of c-Rel, HNF-4, Elk-1, Pax-4, CF2-II, E74A, Oct-1 or StuAp in US12 transcriptional regulation regions are also conserved among HSV-1/2. HNF-4 is conserved in both HSV-1 and HSV-2 strains. Our findings support that the conserved TRSs and TRFs binding sites are closely linked with the gene US12 functions. However, the TRSs and TRFs between HSV-1 and HSV-2 strains are quite different. Many bases are found unpaired in the sequence alignment, and that indicates different biological functions of these TRSs and TRFs in HSV-1 or HSV-2. Whether their functions exist in vivo or are related to the immune escape of HSV is not clear, thus, to validate these, further functional studies are needed.
The TRFs c-Rel and Oct-1 are functional during HSV infection
To understand whether the identified conserved TRSs and TRFs are functional or are involved in the immune escape of HSV, we did a literature search of each of these TRSs and TRFs related to HSV-1 or HSV-2. We found that the TRFs of c-Rel and Oct-1 have been reported to be expressed and functional respectively in HSV-1 and HSV-1/2 infected cells (Table 3). These data are consistent with the identification of TRF c-Rel binding site in HSV-1 and Oct-1 binding site in HSV-2 (Fig. 3), supporting that these TRSs and TRFs are biologically functional.
Table 3.
The TRFs c-Rel and Oct-1 are functional during HSV infection
| Tissue type | HSV strain | Oct-1 | c-Rel | Function | Ref. |
|---|---|---|---|---|---|
| Kidney: Vero cells | HSV-1 strain 17 | – | c-Rel | As a novel cause of HSE disease susceptibility. | [31] |
| Hematological: Jurkat cells | HSV-1 | – | p65/c-Rel | the p65/c-Rel heterodimer is responsible for the NF-kB-dependent induction of HIV-1 LTR in HSV- 1-infected cells. | [32] |
| Embryonic: WT and dOct MEF cells | HSV-1 strain F | Oct-1 | – | Oct-1 is required for the formation of HSV replication factories and late gene expression. | [33] |
| Digestive: Hep2 cells | HSV-1 strain KOS | Oct-1 | – | Oct-1 directly recognizes TAATGARAT elements in promoters of IE genes. | [34] |
| Urinary: COS-7 cells | HSV-1 strain KOS | Oct-1 | – | Distinct conformations of Oct-1 on the BHV IE1 sites and on the HSV IE110 sites. | [35] |
| Genital: HeLa cells | HSV-1 strain F | Oct-1 | – | late in infection Oct-1 is posttranslationally modified and exhibits a reduced capacity to bind to its cognate sites. | [36] |
| Genital: HeLa cells | HSV-1 strain KOS | Oct-1 | – | Ser375 is important for the interaction of VP16 with Oct-1, and that the interaction is required to enable both proteins to bind to IE promoters. | [28] |
| Genital: HFF | HSV-1 strain KOS | Oct-1 | – | forms a transactivation complex with the cellular proteins HCF-1 and HSV-1 VP16 tegument protein. | [29] |
| Genital: HeLa cells | HSV-2 strain 333 | Oct-1 | – | the HSV-2 protein forms a transcriptional complex with the cellular Oct-1 protein and target TAATGARAT elements from immediate-early promoters. | [37] |
HSE Herpes simplex encephalitis, HIV human immunodeficiency virus, LTR long terminal repeat
It is well known that HSV-1 causes buccal ulcers and encephalitis. Interestingly, it’s reported that c-Rel is a novel cause of herpes simplex encephalitis susceptibility [38]. Studies have also shown that c-Rel is involved in immune evasion via interacting with viral nuclear protein UL24 and endogenous NF-κB subunits p65 and p50, and inhibiting cGAS-STING mediated NF-κB promoter activity in HSV-1 infected cells. We would hypothesize that our newly identified HSV-1 specific c-Rel may bind to its US12 TRSs, and activate US12 (ICP47) expression in HSV-1 infected cells. In turn, ICP47 blocked HSV-1 antigen presentation, and promoted HSV-1 infection spread and herpes simplex encephalitis. Oct-1 activates IE-gene transcription through forming a transactivation complex with the cellular proteins HCF-1 and VP16 tegument protein in HSV-1 infected tissues [28, 29].
However, there was no report of US12 transcriptional regulation by c-Rel or Oct-1, no report on c-Rel expression in HSV-2 infected tissues, and no report about expression and function of the other 6 identified TRFs of HNF-4, Elk-1, Pax-4, CF2-II, E74A and StuAp, and no report of any of the TRSs identified above in HSV.
The HSV-1/2 tissue tropism and TRFs expression in different tissues
Tissue tropism is the cells and tissues of a host that support the growth of a particular virus or bacterium. Some bacteria and viruses have a wide range of tissue tropism and can infect many types of cells and tissues, while other viruses may infect mainly individual tissues. Here we summarized the HSV-1/2 tissue tropism and the TRFs expression in different tissues (Table 4). According to the results, HSV-1 specific c-Rel, Elk-1, and Pax-4 are highly expressed in tissues above the abdomen, including oral cavity, tongue and head, and Oct-1, HNF-4 and CF2-II are highly expressed in tissues within the genital system. c-Rel belongs to the nuclear factor κB (NF-κB) family, and plays a crucial role in mammalian B and T cell function [39]. Elk-1 is involved in ERK-induced cellular proliferation, and its transcriptional activity is regulated by ubiquitination at lysine 35 (K35) [40]. Pax proteins are crucial in stem cell biology and organ development. Pax-4 is known to be a major regulator of pancreatic cell development and differentiation, and its transactivation domain was localized within its C-terminal region [41]. OCT-1 (Pou2f1) is well known as a widely expressed TRFs in most cells and tissues. Recently, a series of studies have reported that OCT-1 plays a critical role in CD4+ T cell function through mediating long-range chromosomal interactions and regulating gene expression during differentiation [42]. Hepatocyte nuclear factor 4 (HNF-4) is enriched in liver extracts and belongs to the steroid hormone receptor superfamily [43]. C(2)-H(2)-type zinc-finger transcription factor II (CF2-II) may potentially regulates diverse sets of target genes during cell development and the CF2-II recognition properties depends largely on the COOH-terminal DNA binding domain [44]. E74A belongs to Ets transcription protein, which is involved in multifarious important biological processes. Study has demonstrated that ecdysone inducible TRFs E74A could directly regulate the EO gene expression in silkworm [45]. StuAp is a member of fungal TRFs family that regulates cell cycle progression or development. Further, StuAp belongs to a sub-family possessing the conserved APSES domain. Study has shown that StuAp acts as a transcriptional repressor in A.nidulans, but as a weak activator in budding yeast [46].
Table 4.
The HSV-1/2 tissue tropism and the TRFs expression in different tissues
H high-expression, M middle expression, L little expression, N no-expression; The result from: http://biogps.org. Grading was based on fold increases compared to median fluorescence intensity on Affymetrix microarray chips at 0–2.5 (L), > 2.5- < 5 (M), > 5 (H)
| System | Cell/ tissue | HSV-1 | HSV-2 | c-Rel | HNF-4 | Elk-1 | Pax-4 | CF2-II | OCT-1 |
|---|---|---|---|---|---|---|---|---|---|
| Blood system | CD34+ stem cell | + | _ | H | H | H | H | H | H |
| 721 B lymphoblasts | + | _ | H | H | H | H | M | H | |
| CD19 + B cell | + | _ | H | H | H | H | M | H | |
| Leukemia lymphoblastic | + | _ | M | M | M | M | L | L | |
| Bonemarrow | + | _ | H | M | H | H | M | H | |
| Pituitariy | + | _ | H | H | H | H | H | M | |
| Head | Prefrontal Cortex | + | _ | H | H | H | H | H | H |
| Pineal | + | _ | H | H | H | H | H | M | |
| Tongue | + | _ | H | M | H | H | L | M | |
| Tonsil | + | _ | H | M | M | H | L | L | |
| Retina | + | _ | H | H | H | H | H | M | |
| Trigeminal ganglion | + | _ | M | H | H | H | L | L | |
| Cerebellum | + | _ | H | H | M | H | H | M | |
| Viscera | Heart | + | _ | H | H | H | H | H | M |
| Lung | + | _ | H | H | H | H | H | M | |
| Liver | + | _ | H | H | H | H | H | M | |
| Kidney | + | _ | M | M | M | M | H | L | |
| Smooth Muscles | + | _ | H | H | H | H | H | M | |
| Adipocyte | + | _ | H | M | M | H | L | L | |
| Secretory system | Adrenalgland | + | _ | M | M | H | H | L | L |
| Pancreaticlstet | + | _ | H | H | H | H | H | M | |
| Genital system | Placenta | + | + | H | H | H | M | H | M |
| Fetalthyroid | + | + | H | M | M | M | H | M | |
| Uterus | + | + | M | M | M | M | M | L | |
| Testis | + | + | M | M | M | M | H | L | |
| Ovary | + | + | M | M | L | L | L | M |
The US12 transcription and ICP47 function during the HSV infection process
To better understand the significance of the newly identified US12 transcriptional regulation, we summarize the US12 transcription and ICP47 function during the HSV infection process (Fig. 5). TAP plays a crucial role in MHC I antigen presentation and has become an important target for viral immune escape strategies. In the long-term process of virus-host co-evolution, herpes viruses independently obtained an efficient way to block TAP-mediated peptide transport via the viral immune evasion protein ICP47, which blocks the binding of peptide to TAP by capturing TAP in the endogenous conformation [27]. Interestingly, in our study, we found that two crucial TRFs, c-Rel and Oct-1, play a variety of roles in the growth, proliferation, and survival of mature T cells, which might associate with the viral immune evasion via HSV ICP47. Studies have shown that HSV-1 have evolved complex mechanisms to disrupt the antiviral response via affecting the NF-κB. For example, in HSV-1, ICP0 interacts with p65 and p50 and then degrades p50 through regulating E3 ubiquitin ligase activity [47]. Protein kinase US3 was shown to inhibit NF-κB activity via making p65 hyperphosphorylation at serine 75 and blocking its nuclear translocation [48]. Besides, ICP27 blocks the phosphorylation of IκB to inhibit NF-κB activation. Furthermore, our data also shown that c-Rel is conserved in HSV-1, which inhibits NF-κB promoter activity. Importantly, Oct-1 plays a key role in CD4 T cells, mediating long-range chromosomal interactions and differentiation through regulating gene expression, and has a critical protection effect on viruses and pathogens [42] and further, Oct-1 also is conserved in HSV-2.
Fig. 5.

ICP47 function during HSV infection. a Intracellular antigenic peptides, mainly generated by the proteasome, are transported into the endoplasmic reticulum by the TAP and then loaded onto the nascent MHC I molecules, which are exported to the cell surface and present peptides to the immune cells. Cytotoxic T lymphocytes recognize and kill the infected cells by granzyme and perforin. b ICP47 can preclude peptide binding and traps TAP in an inward-facing conformation. Binding of ICP47 stabilizes the inward-facing conformation, and thus prevents TAP from transitioning to an outward-facing state, resulting in the emergence of empty carrier MHC I molecules. Therefore, CD8+ T cells could not recognize them, and HSVs could avoid the immune responses. We hypothesize that c-Rel bind to its US12 TRS, and enhance US12 (ICP47) expression, leading to HSV-1 immune evasion and HSV-1 encephalitis
Discussion
HSV ICP47 can bind TAP and block antigen presentation. Transcriptional regulation of US12 is important for ICP47 functioning. However, TRSs and TRFs of HSV US12 are seldom reported. In this study, we reported the transcriptional regulation region sequence of our newly isolated strain HSV-1-LXMW in China, and found it is closely related to HSV-1-CR38 and HSV-1-17 in UK. We identified eight different kinds of novel TRSs and TRFs of HSV US12 for the first time. These identified TRSs and TRFs are conserved among HSV-1 (c-Rel, Elk-1, Pax-4), HSV-2 (Oct-1, CF2-II, E74A, StuAp) or both of them (HNF-4). Two of the TRFs c-Rel and Oct-1 are biologically functional in vitro respectively in immune escape and viral replication during HSV infection. We further hypothesize a novel mechanism of HSV-1 encephalitis by c-Rel activated ICP47-mediated immune escape. These findings may have important implication to our understanding of HSV biology, infection, immunity and OVs.
oHSV-1 has become one of the most promising OVs at present [49]. In 2015, talimogene laherparepvec (T-VEC), a kind of oHSV, was approved by FDA for the treatment of advanced melanoma [50–52]. In T-VEC, ICP47 was deleted to prevent limitation of viral antigen presentation, and increase the US11 gene expression, and virus replication in cancer cells without reducing tumor selectivity [53]. Considering that the immune escape function of ICP47, the construction of gene therapy vectors precede a new perspective. For instance, Adeno-associated virus gene therapy of Duchenne muscular dystrophy was achieved by expression ICP47 [54]. Additionally, study has also reported another recombinant adenovirus vector expressing ICP47 protein to reduce the stimulation of dendritic cells [55].
Future functional studies of these novel TRSs and TRFs, and their roles in HSV replication, infection, immunity, tissue tropism, encephalitis and OVs are warranted.
Conclusions
We identified eight different kinds of novel TRFs and TRFs of HSV US12 for the first time, and found they are conserved among HSV-1 (c-Rel, Elk-1, Pax-4), HSV-2 (Oct-1, CF2-II, E74A, StuAp) or both HSVs (HNF-4). The c-Rel and Oct-1 are biologically functional respectively in immune escape and viral replication during HSV infection. We further hypothesized a novel mechanism of HSV-1 encephalitis caused by c-Rel activated ICP47-mediated immune escape.
Supplementary information
Additional file 1. Using the nucleotide sequence database, we identified transcription regulatory regions of US12 (145851–148,050). The transcription regulatory regions are 2000 bp upstream and 200 bp downstream of US12 transcription initiation sites.
Acknowledgements
None.
Abbreviations
- HSV
Herpes simplex virus
- ICP47
Infected cell polypeptide 47
- oHSV
oncolytic HSV
- TRSs
Transcription regulatory sequences
- TRFs
Transcription regulatory factors
- OVs
Oncolytic viruses
- TK
Thymidine kinase
- MHC-I
Major histocompatibility complex I
- TAP
Transporter associated with antigen processing
- NBDs
Nucleotide binding domains
- ER
Endoplasmic reticulum
- CTLs
Cytotoxic T lymphocytes
- NF-κB
Nuclear factor κB
- HNF-4
Hepatocyte nuclear factor 4
- CF2-II
C(2)-H(2)-type zinc-finger transcription factor II
- T-VEC
Talimogene laherparepvec
Authors’ contributions
J.-T.C. and Y.-Y.W. designed the outline of the article and wrote it. Z.M. and H.-W.X designed the outline of the article, revised the initial draft and expanded the manuscript. L.-Z.Z. and B.-R.L. revised the manuscript. All made intellectual contributions. All authors approved the final manuscript.
Funding
This work was mainly supported by National Natural Science Foundation of China (81872412 to XHW, 81602303 to XY, 31700736 to WXW). We thank the Natural Science Foundation of Hubei Province (2019CFB591 to Z.M.), and Guangzhou Key Medical Discipline Construction Project (CSZ).
Availability of data and materials
All data from the current study are available from the corresponding author on request.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors have declared that no competing interest exists. Consent for publication.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Jun-Ting Cheng, Ying-Ying Wang and Lin-Zhong Zhu contributed equally to this work.
Contributor Information
Zhaowu Ma, Email: zhaowu823@126.com.
Bing-Rong Liu, Email: bingrongliu@qq.com.
Hong-Wu Xin, Email: hongwu_xin@126.com.
Supplementary information
Supplementary information accompanies this paper at 10.1186/s12985-020-01365-3.
References
- 1.Ellison LM, Man Y, Stojadinovic A, Xin H, Avital I. Cytoreductive surgery and hyperthermic intraperitoneal chemotherapy in treatment of gastric cancer with peritoneal carcinomatosis. Chin J Cancer Res. 2017;29:86–92. doi: 10.21147/j.issn.1000-9604.2017.01.10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Liu Y, Yu C, Wu Y, Sun X, Su Q, You C, Xin H. CD44(+) fibroblasts increases breast cancer cell survival and drug resistance via IGF2BP3-CD44-IGF2 signalling. J Cell Mol Med. 2017;21:1979–1988. doi: 10.1111/jcmm.13118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Xin HW, Ambe CM, Ray S, Kim BK, Koizumi T, Wiegand GW, Hari D, Mullinax JE, Jaiswal KR, Garfield SH, et al. Wnt and the cancer niche: paracrine interactions with gastrointestinal cancer cells undergoing asymmetric cell division. J Cancer. 2013;4:447–457. doi: 10.7150/jca.6896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Xin HW, Ambe CM, Hari DM, Wiegand GW, Miller TC, Chen JQ, Anderson AJ, Ray S, Mullinax JE, Koizumi T, et al. Label-retaining liver cancer cells are relatively resistant to sorafenib. Gut. 2013;62:1777–1786. doi: 10.1136/gutjnl-2012-303261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Xin HW, Ambe CM, Miller TC, Chen JQ, Wiegand GW, Anderson AJ, Ray S, Mullinax JE, Hari DM, Koizumi T, et al. Liver label retaining Cancer cells are relatively resistant to the reported anti-Cancer stem cell drug metformin. J Cancer. 2016;7:1142–1151. doi: 10.7150/jca.10047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hari D, Xin HW, Jaiswal K, Wiegand G, Kim BK, Ambe C, Burka D, Koizumi T, Ray S, Garfield S, et al. Isolation of live label-retaining cells and cells undergoing asymmetric cell division via nonrandom chromosomal cosegregation from human cancers. Stem Cells Dev. 2011;20:1649–1658. doi: 10.1089/scd.2010.0455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Liu XQ, Xin HY, Lyu YN, Ma ZW, Peng XC, Xiang Y, Wang YY, Wu ZJ, Cheng JT, Ji JF, et al. Oncolytic herpes simplex virus tumor targeting and neutralization escape by engineering viral envelope glycoproteins. Drug Deliv. 2018;25:1950–1962. doi: 10.1080/10717544.2018.1534895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Russell SJ, Barber GN. Oncolytic viruses as antigen-agnostic Cancer vaccines. Cancer Cell. 2018;33:599–605. doi: 10.1016/j.ccell.2018.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lawler SE, Speranza MC, Cho CF, Chiocca EA. Oncolytic viruses in Cancer treatment: a review. JAMA Oncol. 2017;3:841–849. doi: 10.1001/jamaoncol.2016.2064. [DOI] [PubMed] [Google Scholar]
- 10.Puzanov I, Milhem MM, Minor D, Hamid O, Li A, Chen L, Chastain M, Gorski KS, Anderson A, Chou J, et al. Talimogene Laherparepvec in combination with Ipilimumab in previously untreated, Unresectable stage IIIB-IV melanoma. J Clin Oncol. 2016;34:2619–2626. doi: 10.1200/JCO.2016.67.1529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Poh A. First Oncolytic viral therapy for melanoma. Cancer Discov. 2016;6:6. doi: 10.1158/2159-8290.CD-NB2015-158. [DOI] [PubMed] [Google Scholar]
- 12.Agarwalla PK, Aghi MK. Oncolytic herpes simplex virus engineering and preparation. Methods Mol Biol. 2012;797:1–19. doi: 10.1007/978-1-61779-340-0_1. [DOI] [PubMed] [Google Scholar]
- 13.Dai X, Zhou ZH. Structure of the herpes simplex virus 1 capsid with associated tegument protein complexes. Science. 2018;360. [DOI] [PMC free article] [PubMed]
- 14.Johnston C, Koelle DM, Wald A. HSV-2: in pursuit of a vaccine. J Clin Invest. 2011;121:4600–4609. doi: 10.1172/JCI57148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Duan J, Liuzzi M, Paris W, Liard F, Browne A, Dansereau N, Simoneau B, Faucher AM, Cordingley MG. Oral bioavailability and in vivo efficacy of the helicase-primase inhibitor BILS 45 BS against acyclovir-resistant herpes simplex virus type 1. Antimicrob Agents Chemother. 2003;47:1798–1804. doi: 10.1128/AAC.47.6.1798-1804.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bernstock JD, Vicario N, Li R, Nan L, Totsch SK, Schlappi C, Gessler F, Han X, Parenti R, Beierle EA, et al. Safety and efficacy of oncolytic HSV-1 G207 inoculated into the cerebellum of mice. Cancer Gene Ther. 2019. [DOI] [PMC free article] [PubMed]
- 17.Russell SJ, Peng KW, Bell JC. Oncolytic virotherapy. Nat Biotechnol. 2012;30:658–670. doi: 10.1038/nbt.2287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Todo T, Martuza RL, Rabkin SD, Johnson PA. Oncolytic herpes simplex virus vector with enhanced MHC class I presentation and tumor cell killing. Proc Natl Acad Sci. 2001;98:6396–6401. doi: 10.1073/pnas.101136398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Whitley RJ, Roizman B. Herpes simplex virus infections. Lancet. 2001;357:1513–1518. doi: 10.1016/S0140-6736(00)04638-9. [DOI] [PubMed] [Google Scholar]
- 20.Praest P, Luteijn RD, Brak-Boer IGJ, Lanfermeijer J, Hoelen H, Ijgosse L, Costa AI, Gorham RD, Lebbink RJ, Wiertz EJHJ. The influence of TAP1 and TAP2 gene polymorphisms on TAP function and its inhibition by viral immune evasion proteins. Mol Immunol. 2018;101:55–64. doi: 10.1016/j.molimm.2018.05.025. [DOI] [PubMed] [Google Scholar]
- 21.Oldham ML, Hite RK, Steffen AM, Damko E, Li Z, Walz T, Chen J. A mechanism of viral immune evasion revealed by cryo-EM analysis of the TAP transporter. Nature. 2016;529:537–540. doi: 10.1038/nature16506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Matschulla T, Berry R, Gerke C, Döring M, Busch J, Paijo J, Kalinke U, Momburg F, Hengel H, Halenius A. A highly conserved sequence of the viral TAP inhibitor ICP47 is required for freezing of the peptide transport cycle. Sci Rep. 2017;7. [DOI] [PMC free article] [PubMed]
- 23.Liu R, Varghese S, Rabkin SD. Oncolytic herpes simplex virus vector therapy of breast cancer in C3(1)/SV40 T-antigen transgenic mice. Cancer Res. 2005;65:1532–1540. doi: 10.1158/0008-5472.CAN-04-3353. [DOI] [PubMed] [Google Scholar]
- 24.Messerli SM, Prabhakar S, Tang Y, Mahmood U, Giovannini M, Weissleder R, Bronson R, Martuza R, Rabkin S, Breakefield XO. Treatment of schwannomas with an oncolytic recombinant herpes simplex virus in murine models of neurofibromatosis type 2. Hum Gene Ther. 2006;17:20–30. doi: 10.1089/hum.2006.17.20. [DOI] [PubMed] [Google Scholar]
- 25.Fukuhara H, Martuza RL, Rabkin SD, Ito Y, Todo T. Oncolytic herpes simplex virus vector g47delta in combination with androgen ablation for the treatment of human prostate adenocarcinoma. Clin Cancer Res. 2005;11:7886–7890. doi: 10.1158/1078-0432.CCR-05-1090. [DOI] [PubMed] [Google Scholar]
- 26.Raafat N, Sadowski-Cron C, Mengus C, Heberer M, Spagnoli GC, Zajac P. Preventing vaccinia virus class-I epitopes presentation by HSV-ICP47 enhances the immunogenicity of a TAP-independent cancer vaccine epitope. Int J Cancer. 2012;131:E659–E669. doi: 10.1002/ijc.27362. [DOI] [PubMed] [Google Scholar]
- 27.Scaria A, Sullivan JA, St. George JA, Kaplan JM, Lukason MJ, Morris JE, Plog M, Nicolette C, Gregory RJ, Wadsworth SC. Adenoviral vector expressing ICP47 inhibits adenovirus-specific cytotoxic T lymphocytes in nonhuman primates. Mol Ther. 2000;2:505–514. doi: 10.1006/mthe.2000.0197. [DOI] [PubMed] [Google Scholar]
- 28.S O, FJ H, JR D, A H, S M, WS L, SJ T: Phosphorylation of the VP16 transcriptional activator protein during herpes simplex virus infection and mutational analysis of putative phosphorylation sites. Virology 2006, 345:468–481. [DOI] [PMC free article] [PubMed]
- 29.Suk H, Knipe DM. Proteomic analysis of the herpes simplex virus 1 virion protein 16 transactivator protein in infected cells. Proteomics. 2015;15:1957–1967. doi: 10.1002/pmic.201500020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wang YY, Lyu YN, Xin HY, Cheng JT, Liu XQ, Wang XW, Peng XC, Xiang Y, Xin VW, Lu CB, et al. Identification of putative UL54 (ICP27) transcription regulatory sequences binding to Oct-1, v-Myb, Pax-6 and hairy in herpes simplex viruses. J Cancer. 2019;10:430–440. doi: 10.7150/jca.29787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mancini M, Caignard G, Charbonneau B, Dumaine A, Wu N, Leiva-Torres GA, Gerondakis S, Pearson A, Qureshi ST, Sladek R, Vidal SM. Rel-dependent immune and central nervous system mechanisms control viral replication and inflammation during mouse herpes simplex encephalitis. J Immunol. 2019;202:1479–1493. doi: 10.4049/jimmunol.1800063. [DOI] [PubMed] [Google Scholar]
- 32.SL S, J H, PM P: Differential regulation of the HIV-1 LTR by specific NF-kappa B subunits in HSV-1-infected cells. Virology 1996, 224:214–223. [DOI] [PubMed]
- 33.Nogueira ML, Wang VE, Tantin D, Sharp PA, Kristie TM. Herpes simplex virus infections are arrested in Oct-1-deficient cells. Proc Natl Acad Sci U S A. 2004;101:1473–1478. doi: 10.1073/pnas.0307300101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Akhova O, Bainbridge M, Misra V. The neuronal host cell factor-binding protein Zhangfei inhibits herpes simplex virus replication. J Virol. 2005;79:14708–14718. doi: 10.1128/JVI.79.23.14708-14718.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.V M, S W, P Y, S H, P OH: Conformational alteration of Oct-1 upon DNA binding dictates selectivity in differential interactions with related transcriptional coactivators. Mol Cell Biol 1996, 16:4404–4413. [DOI] [PMC free article] [PubMed]
- 36.Advani SJ, Durand LO, Weichselbaum RR, Roizman B. Oct-1 is posttranslationally modified and exhibits reduced capacity to bind cognate sites at late times after infection with herpes simplex virus 1. J Virol. 2003;77:11927–11932. doi: 10.1128/JVI.77.22.11927-11932.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.RF G, P OH: Sequence, function, and regulation of the Vmw65 gene of herpes simplex virus type 2. J Virol 1991, 65:6705–6713. [DOI] [PMC free article] [PubMed]
- 38.Mancini M, Caignard G, Charbonneau B, Dumaine A, Wu N: Rel-Dependent Immune and Central Nervous System Mechanisms Control Viral Replication and Inflammation during Mouse Herpes Simplex Encephalitis 2019, 202:1479–1493. [DOI] [PubMed]
- 39.Gilmore TD, Gerondakis S. The c-Rel transcription factor in development and disease. Genes Cancer. 2011;2:695–711. doi: 10.1177/1947601911421925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ducker C, Chow LKY, Saxton J, Handwerger J, McGregor A, Strahl T, Layfield R, Shaw PE. De-ubiquitination of ELK-1 by USP17 potentiates mitogenic gene expression and cell proliferation. Nucleic Acids Res. 2019;47:4495–4508. doi: 10.1093/nar/gkz166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sakata N, Yoshimatsu G, Kodama S. Development and characteristics of pancreatic epsilon cells. Int J Mol Sci. 2019;20. [DOI] [PMC free article] [PubMed]
- 42.Hwang SS, Kim LK, Lee GR, Flavell RA. Role of OCT-1 and partner proteins in T cell differentiation. Biochim Biophys Acta. 2016;1859:825–831. doi: 10.1016/j.bbagrm.2016.04.006. [DOI] [PubMed] [Google Scholar]
- 43.Tsukamoto D, Ito M, Takamatsu N. HNF-4 participates in the hibernation-associated transcriptional regulation of the chipmunk hibernation-related protein gene. Sci Rep. 2017;7:44279. doi: 10.1038/srep44279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gogos JA, Hsu T, Bolton J, Kafatos FC. Sequence discrimination by alternatively spliced isoforms of a DNA binding zinc finger domain. Science. 1992;257:1951–1955. doi: 10.1126/science.1290524. [DOI] [PubMed] [Google Scholar]
- 45.Sun W, Wang CF, Zhang Z. Transcription factor E74A affects the ecdysone titer by regulating the expression of the EO gene in the silkworm, Bomby mori. Biochim Biophys Acta Gen Subj. 2017;1861:551–558. doi: 10.1016/j.bbagen.2016.11.017. [DOI] [PubMed] [Google Scholar]
- 46.Zhao Y, Su H, Zhou J, Feng H, Zhang KQ, Yang J. The APSES family proteins in fungi: characterizations, evolution and functions. Fungal Genet Biol. 2015;81:271–280. doi: 10.1016/j.fgb.2014.12.003. [DOI] [PubMed] [Google Scholar]
- 47.Zhang J, Wang K, Wang S, Zheng C. Herpes simplex virus 1 E3 ubiquitin ligase ICP0 protein inhibits tumor necrosis factor alpha-induced NF-kappaB activation by interacting with p65/RelA and p50/NF-kappaB1. J Virol. 2013;87:12935–12948. doi: 10.1128/JVI.01952-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wang K, Ni L, Wang S, Zheng C. Herpes simplex virus 1 protein kinase US3 hyperphosphorylates p65/RelA and dampens NF-kappaB activation. J Virol. 2014;88:7941–7951. doi: 10.1128/JVI.03394-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wang D, Wang XW, Peng XC, Xiang Y, Song SB, Wang YY, Chen L, Xin VW, Lyu YN, Ji J, et al. CRISPR/Cas9 genome editing technology significantly accelerated herpes simplex virus research. Cancer Gene Ther. 2018;25:93–105. doi: 10.1038/s41417-018-0016-3. [DOI] [PubMed] [Google Scholar]
- 50.Killock D. Skin cancer: T-VEC oncolytic viral therapy shows promise in melanoma. Nat Rev Clin Oncol. 2015;12:438. doi: 10.1038/nrclinonc.2015.106. [DOI] [PubMed] [Google Scholar]
- 51.Johnson DB, Puzanov I, Kelley MC. Talimogene laherparepvec (T-VEC) for the treatment of advanced melanoma. Immunotherapy. 2015;7:611–619. doi: 10.2217/imt.15.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Andtbacka R, Collichio F, Amatruda T, Senzer N, Chesney J, Delman K, Spitler L, Puzanov I, Doleman S, Ye Y, et al. OPTiM: a multicenter, randomized phase 3 trial of Talimogene Laherparepvec versus subcutaneous GM-CSF for the treatment of Unresected stage IIIB/C and IV melanoma. J der Deutschen Dermatologischen Gesellschaft. 2013;11:8–8. [Google Scholar]
- 53.Grigg C, Blake Z, Gartrell R, Sacher A, Taback B, Saenger Y. Talimogene laherparepvec (T-Vec) for the treatment of melanoma and other cancers. Semin Oncol. 2016;43:638–646. doi: 10.1053/j.seminoncol.2016.10.005. [DOI] [PubMed] [Google Scholar]
- 54.Shao W, Chen X, Samulski RJ, Hirsch ML, Li C. Inhibition of antigen presentation during AAV gene therapy using virus peptides. Hum Mol Genet. 2018;27:601–613. doi: 10.1093/hmg/ddx427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Wang P, Kan Q, Yu Z, Li L, Zhang Z, Pan X, Feng T. Recombinant adenovirus expressing ICP47 gene suppresses the ability of dendritic cells by restricting specific T cell responses. Cell Immunol. 2013;282:129–135. doi: 10.1016/j.cellimm.2013.05.004. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1. Using the nucleotide sequence database, we identified transcription regulatory regions of US12 (145851–148,050). The transcription regulatory regions are 2000 bp upstream and 200 bp downstream of US12 transcription initiation sites.
Data Availability Statement
All data from the current study are available from the corresponding author on request.