

Abstract
Molecular and morphological phylogenies concur in indicating that the African lineages formerly referred to Crocodylus niloticus are the sister taxon the four Neotropical crocodiles (Crocodylus intermedius, C. moreleti, C. acutus and C. rhombifer), implying a transoceanic dispersal from Africa to America. So far the fossil record did not contribute to identify a possible African forerunner of the Neotropical species but, curiously, the oldest remains referred to the African C. niloticus are Quaternary in age, whereas the oldest American fossils of Crocodylus are older, being dated to the early Pliocene, suggesting that another species could be involved. We re-described, also thanks to CT imaging, the only well-preserved topotipic skull of Crocodylus checchiai Maccagno, 1947 from the late Miocene (Messinian) African site of As Sahabi in Libya. As previously suggested on the basis of late Miocene material from Tanzania, C. checchiai is a valid, diagnosable species. According to our phylogenetic analyses, C. checchiai is related to the Neotropical taxa and could be even located at the base of their radiation, therefore representing the missing link between the African and the American lineages.
Subject terms: Palaeontology, Zoology, Biogeography
Introduction
Extant crocodylians are represented by 25 species grouped into 9 genera1. The most speciose and widespread genus is Crocodylus Laurenti, 1768 that hosts 12 species inhabiting a longitudinally very broad intertropical belt ranging from Australia to South Asia, to Africa and then America. The origin of Crocodylus was placed in the late Miocene (about 13.6–8.3 Ma) of Asia by Oaks2, on the basis of a time-calibrated species tree stemming from a DNA analysis, and recently reconstructed as Asian by Nicolai and Mazke3 on the basis of historical biogeography models. Both the molecular2,4,5 and the morphological/palaeontological6–9 approaches, as well as the shared presence of parasites10 concur in indicating that the origin of the American clade of Crocodylus should be sought in Africa. The four American species Crocodylus intermedius, C. moreleti, C. acutus and C. rhombiferare grouped together with the African Crocodylus niloticus Laurenti, 1768, which is placed at the base of the American branch. According to Meredith et al.5, the most parsimonious explanation that takes into consideration both the phylogenetic results and the fossil record of Crocodylus supports a relatively recent trans-Atlantic crossing of “C. niloticus” from Africa to America. The reason of the inverted commas placed by these authors around the name of Nile crocodile is related to the fact that the eastern and western clades of this species do not cluster together, but are arranged in paraphyly at the base of the American clade, a result shared by different authors applying different molecular techniques2,4,5,11,12. Hekkala et al.5 eventually proposed the resurrection of the name Crocodylus suchus Geoffroy Saint-Hilaire, 1807 for the western populations, keeping the name C. niloticus for the (predominantly) eastern populations that is directly at the base of the American clade. Probably because of the dominance of the molecular approach involved in the analysis of the phylogenetic relationships and biogeographic history of extant taxa, the African fossil record has been only marginally involved in the discussion concerning the origin of the American clade6,7,8,9, even if Hect13 already underlined in 1987 that an African extinct species shared with the American clade a synapomorphic trait.
Crocodylus checchiai Maccagno, 1947
This species (Fig. 1) was originally described in Italian by Maccagno14 on the basis of an adult, well-preserved skull and associated lower jaw collected in 1938 by Petrocchi in the area of As Sahabi in Libya and then hosted in the collections of the Istituto di Paleontologia dell’Università di Roma (now apparently lost). As Sahabi is a celebrated latest Miocene vertebrate locality in northern Libya, located about 130 km south of Ajdabiya along the road going south into the Libyan Sahara to Gialo and Kufra (Fig. 2). The discovery of the paleontological site and the recovery of abundant fossils at As Sahabi in the 1930′s is due to the invaluable efforts of C. Petrocchi16,17.
Figure 1.

Life appearance of Crocodylus checchiai from As Sahabi (Libya). The reconstruction is based on the information from literature13–15 updated with the complete cranium sn813/lj. Artwork by D. A. Iurino.
Figure 2.

Geographic location of the As Sahabi paleontological locality. Artwork by D. A. Iurino.
Maccagno provided a thorough description of the skull and a lengthy series of comparisons with some of the extinct and extant crocodylian taxa known at that time, comparisons that are now of little value in the context of the currently known phylogenetic relationships. However, among the several characters listed as diagnostic for C. checchiai at least two are still considered of taxonomic and phylogenetic relevance: the marked median gibbosity located in the posterior sector of the nasals (involving also the maxillae and the lacrimals), as well as the anterior tip of the nasals entering the narial opening.
A few years later, in a second paper on the crocodiles from As Sahabi, Maccagno15 reported the presence of four more skulls. The three skulls (one with lower jaw) stored in the Museum of Natural History of Tripoli were destroyed or lost during the II World War, but she published their photograph in lateral view15. The fourth skull, collected by Petrocchi in 1939 and brought to Rome was described by Maccagno15 and referred to the new variety, Crocodylus checchiai var. depressa characterized by slightly broader posterior region of the skull. Few years ago, fragmentary new material from the same Libyan area of As Sahabi (30P24A belonging to the University of Garyounis Earth Science Museum, but currently on loan at the Department of Earth Sciences of the University of Florence, Italy) was referred to C. checchiai by Delfino18 without any phylogenetic consideration. Later Brochu and Storrs9 referred two skulls (KNM-LT 23108 and KNM-LT 26618) from the late Miocene–early Pliocene Nawata Formation at Lothagam (Tanzania) to the same taxon. On the basis of the Tanzanian remains, they included for the first time C. checchiai in a phylogenetic context and concluded that this species cannot be excluded from Crocodylus, but retrieved a broad politomy that did not support any precise relationship.
In order to improve the knowledge about C. checchiai and its possible relationships with the Neotropical crocodile species, we tried to locate the original material described by Maccagno and succeeded in finding in the collection of the MUST (Museo Universitario di Scienze della Terra) of Sapienza University of Rome, the skull sn813/lj on which Maccagno15 based the taxon C. checchiai var. depressa. We here redescribe the specimen also on the basis of tomographic imaging that allow us to study the inner structures, taking into consideration the characters currently considered taxonomically diagnostic and phylogenetically relevant. The phylogenetic relationships were explored thanks to the character matrix by Brochu and Storrs9 that includes several African crocodylines, as well as on the basis of that of Scheyer et al.19 that includes the geological oldest American Crocodylus, e.g. C. falconensis.
Systematic palaeontology
Crocodylia Gmelin, 1789, sensu Benton and Clark, 1988.
Crocodylidae Cuvier, 1807, sensu Brochu, 2003.
Crocodylus Laurenti, 1769.
Crocodylus checchiai Maccagno, 1947.
Institutional abbreviations
KNM, National Museums of Kenya, Nairobi, Kenya; MPURLS, Museo di Paleontologia dell’Università di Roma La Sapienza, Rome, Italy; MUST, Museo Universitario di Scienze della Terra, Sapienza University of Rome, Italy; sn, senza numero d’inventario (temporary inventory number).
Referred material
Cranium sn813/lj (ex MPURLS) (Fig. 3, Supplementary Video S1) stored at MUST.
Figure 3.

Crocodylus checchiai14 from As Sahabi. Specimen sn813/lj in dorsal (a1,a2), ventral (b1,b2), right lateral (c1,c2) and left lateral (d1,d2) views. Anatomical abbreviations: bo, basioccipital; bs, basisphenoid; ect, ectopterygoid; en, external naris; f, frontal; if, incisive foramen; itf, infratemporal fenestra; j, jugal; l, lacrimal; m, maxilla; n, nasal; o, orbit; pa, parietal; pal, palatine; pf, prefrontal; pm, premaxilla; po, postorbital; pt, pterygoid; q, quadrate; qj, quadratojugal; soc, supraoccipital; sof, suborbital fenestra; sq, squamosal; stf, supratemporal fenestra; t, tooth. Scale bar: 10 cm.
Occurrence
Albeit the age of the As Sahabi vertebrate fauna has long been debated, we concur with most recent studies20 in which the geological context suggests a Messinian age, around 7 Ma, for As Sahabi vertebrate bearing levels. According to Bernor and Rook21 and Bernor et al.22 in respect to faunal biogeography and biochronology, As Sahabi is best correlated with the Upper Nawata of Lothagam (Kenya) and MN13 Mammal Unit in the European Neogene Mammal biochronological time scale23.
Emended diagnosis
Crocodylus checchiai differs from all the other extant or extinct Crocodylus species because of the combination of the following characters: presence of medial rostral boss (the only African species so far described with a medial rostral boss); broad contact of lacrimal with nasal without a posterior process of the maxilla; flattened (or nearly so) posterolateral margin of the squamosal; quadratojugal extension to superior angle of infratemporal fenestra. Noteworthy is that C. checchiai is characterized by 5 premaxillary and 13 maxillary alveoli. According to Maccagno14 this species has 12 dentary alveoli.
Description
Preservation and general features
The specimen sn813/lj (Fig. 3, Supplementary Video S1) is a relatively well-preserved, minimally distorted mesorostral skull, whose dorsal length from the preserved anterior tip of the premaxillae to posterior tip of the supraoccipital corresponds to 47.0 cm. Pterygoids and basisphenoid are incomplete. The anterior tip of the nasals is missing. A small lateral area on the left maxilla, at the level of the posterior most alveoli, is damaged. The dorsal surface of the skull hosts a complex pattern of pits and ridges that build up a dense and evident ornamentation. Most of the surface is however covered by a thin layer of arenaceous crusts that hinders some of the sutures and fine morphological details. The rostrum shows a marked festooning in both dorsal and lateral views. The lateral surface of the snout is distinctly constricted in dorsal view at the level of the premaxillary-maxillary suture. The slight asymmetry visible in dorsal view is even more evident on the ventral surface, where, although there is a distinct lateral notch on both sides, the (anatomically) right one is better defined. On the right side there are two low longitudinal ridges delimiting, dorsally and ventrally, an elongated, lateral shallow concavity. A distinct convexity develops on the dorsal surface of both maxillae in correspondence of the root of the fifth maxillary tooth. The most striking character of the rostrum is the presence of a mid-rostral boss that involves anteriorly the posterior area of the nasals, but also the prefrontals, lacrimals (where a sort of accessory bump is present) and the anterior tip of the frontal process posteriorly. Canthi rostralii are absent, as well as marked preorbital ridges and antorbital fenestrae. The trapezoidal skull table is relatively small and flat (slightly concave in posterior view if the small bump of the right squamosal is considered), but a breakage, followed by partial displacement, lowered the area anterior to the right supratemporal fenestra.
Fenestrae and major openings
The external naris opens flush with dorsal surface of premaxillae and is slightly anterposteriorly elongated, but not prominently teardrop-shaped (therefore character 83 was scored as 0). Its posterior rim is broken off for about 1 cm exactly in the area in which presumably the nasals reached the naris. This is supported by the fact that the remaining portion of the nasals still protrude into the naris below the missing dorsal surface. The orbits are larger than the infratemporal fenestrae; their rim is slightly upturned and their ventral edge is nearly circular. The infratemporal fenestrae are posteriorly incomplete due to the breakage of the thin quadratojugal lamina; the apparent quadratojugal spine visible on the right fenestra is interpreted here as an artifact of preservation. The supratemporal fenestrae are almond-shaped and have a smooth anteromedial corner (the fossa is not shallow in that area). The bones delimiting the fenestrae do not overhang the fossa. The medial parietal wall of the supratemporal fenestra is imperforate. The posterior margin of otic aperture is probably bowed, but the presence of matrix in the area and a possible breakage renders this statement as tentative. The foramen magnum is as broad as the occipital condyle. The incisive foramen is relatively small and abuts the premaxillary tooth row. The suborbital fenestrae extend anteriorly to the ninth alveolus. The internal choana is entirely surrounded by pterygoids and projects postero-ventrally. Its rim and internal structure is significantly altered and therefore it is not possible to evaluate the presence or absence of a septum, but the posterior area is clearly flush with the pterygoid surface and there is no evidence for a notch. Concerning the smaller foramina, the foramen of the XII cranial nerve is very small, the foramen vagi is hosted in a very wide depression, and the posterior carotid foramen is located in the depression of the foramen vagi, but is much smaller and placed below it.
Skeletal elements
The dorsal surface of each premaxilla, close to the anterior edge, is pierced by the hole produced by the occlusion of the first dentary tooth. The surface lateral to naris is smooth. The posterior projections of the premaxillae are short: they do not reach the level of the third alveolus on the dorsal surface and just the first alveolus on the ventral surface. Each premaxilla has five alveoli. The largest alveolus is the fourth. There are 13 alveoli on the right maxilla and probably the same number on the left side. All the alveoli are circular in cross-section (slightly compressed the last ones). The maxillary tooth row is rather linear. The largest alveolus is the fifth and the penultimate alveolus has a diameter less than twice the diameter of the last one. The maxilla terminates in palatal view anterior to the lower temporal bar and its medial margin is linear and adjacent to the suborbital fenestra. The nasals are relatively broad and dorsally curved in their posterior sector (in the area hosting the rostral boss). The posterior rim of the external naris is incomplete and therefore it is not possible to directly assess the relationships between the nasals and the naris that however seem to contact each other (see the section “Fenestrae and major openings”).
The lacrimal is much longer than the prefrontal, and it makes a broad contact with nasal without any insertion of the posterior processes of maxilla. The jugal forms the posterior angle of infratemporal fenestra. The medial jugal foramen is covered by concretion, but thanks to the CT scan it is possible to state that it is small. The prefrontal dorsal surface is rather flat, without any discrete knob-like processes. Prefrontals are separated from each other by frontals and nasals. Both the prefrontal pillars are preserved, but the left one is still embedded in the matrix and does not offer any detailed morphological information. Conversely, the right pillar is clearly anteroposteriorly expanded dorsally and bears a medial process anteroposteriorly expanded and constricted at the base. The prefrontal pillar is solid without pneumatic recess. The frontal is smooth between orbits (it does not host a midsagittal crest). The frontoparietal suture is wavy and asymmetrical and fully lies on the skull table. The frontal has an acute and sharp anterior termination. The postorbital bar is slender and has a short and not prominent process. The ventral margin of postorbital bar is inset to the lateral jugal surface. The dorsal margin is not continuous with anterolateral edge of skull table, but clearly inset. The parietal is deeply constricted by the supratemporal fenestrae. The supraoccipital is rather small and it is only minimally exposed on the skull table at the back of the parietal.
The dorsal surface of the squamosals is rather flat and does not develop a marked convexity at the posterolateral edge. The squamosal does not extend ventrolaterally to lateral extent of the paroccipital process. The dorsal and ventral rims of the squamosal groove for the ear valve musculature are parallel to each other. The squamosal-quadrate suture extends dorsally along the posterior margin of the external auditory meatus. The quadrate ramus has no crest on the dorsal surface. The foramen aëreum is small, and located on the mediodorsal angle of the quadrate. The quadrate ramus has, on its ventral surface, the attachment scar for the posterior mandibular adductor muscle. The medial hemicondyle is expanded. The presence and development of the quadratojugal spine was not evaluated for preservational reasons. The putative spine visible in the right infratemporal fenestra is very likely the result of the breakage of the medial rim of the fenestra. The quadratojugal does not show an anterior process along the lower temporal bar, but extends to the superior angle of the infratemporal fenestra. The palatine process extends significantly beyond the anterior end of the suborbital fenestra and terminates by curving gently. The left palatine-pterygoid suture clearly does not reach the posterior corner of the suborbital fenestra, but the morphology is less clear on the left side. The lateral edges of palatines are parallel posteriorly. The ectopterygoid abuts maxillary tooth row. The ectopterygoid maxillary ramus forms less than two thirds of the lateral margin length of suborbital fenestra. Its anterior tip terminates at the level of the third last alveolus and is clearly forked on both sides. The left ectopterygoid extends along the medial face of the postorbital bar, and the right one extends slightly beyond. The pterygoid ramus of the ectopterygoid is bowed and the posterolateral margin of suborbital fenestra is concave. Since the pterygoid flanges are broken and missing, it is not possible to assess the ectopterygoid extension along the pterygoid. The pterygoid surface lateral and anterior to the choana is rather flush with the choanal margin. The basisphenoid is clearly thin and does not show any sulcus on the rostrum. The laterosphenoid process is oriented anteroposteriorly.
Dentition and occlusal pattern
There are 5 and 13 alveoli for each premaxilla and maxilla (Fig. 3). Teeth are preserved in the following positions: 3, 4, 5 and 3, 4 respectively on the right and left premaxilla; 1, 2, 4, 5, 9, 10, 12 and 3, 4, 5, 9, 10, 11 respectively on the right and left maxilla. The shape of the crown varies according to its position in the tooth row. All premaxillary teeth are slender, whereas those of the dentary show a significant variation, being the posterior teeth much lower and stouter that the anterior teeth. The crowns are variably preserved, but no one is complete. However, it is clear that they are not distinctly labiolingually compressed and characterized by a mesiodistal carina, devoid of serration.
For preservational reasons (mostly the presence of the arenaceous crust in the interalveolar spaces), the only evident occlusal pit is located in the sixth interalveolar space of the right maxilla. The absence of pits medial to the tooth row in the posterior sector of the maxillae contributes in indicating that the general occlusal pattern was characterized by the dentary teeth occluding in line with the maxillary tooth row.
Results
In the first analysis (see results in Fig. 4) we obtained 6 equally parsimonious trees 311 steps long and with adjusted homoplasy = 29.5, Consistency Index = 0.47 and Retention index = 0.71. Strict consensus tree is 327 steps long with adjusted homoplasy = 31.25, Consistency Index = 0.45 and Retention index = 0.69. Crocodylus checchiai is placed in a clade comprehending all South American crocodiles; it is arranged in polytomy with C. intermedius and basal to a polytomic clade including C. moreletii, C. rhombifer and C. acutus.
Figure 4.
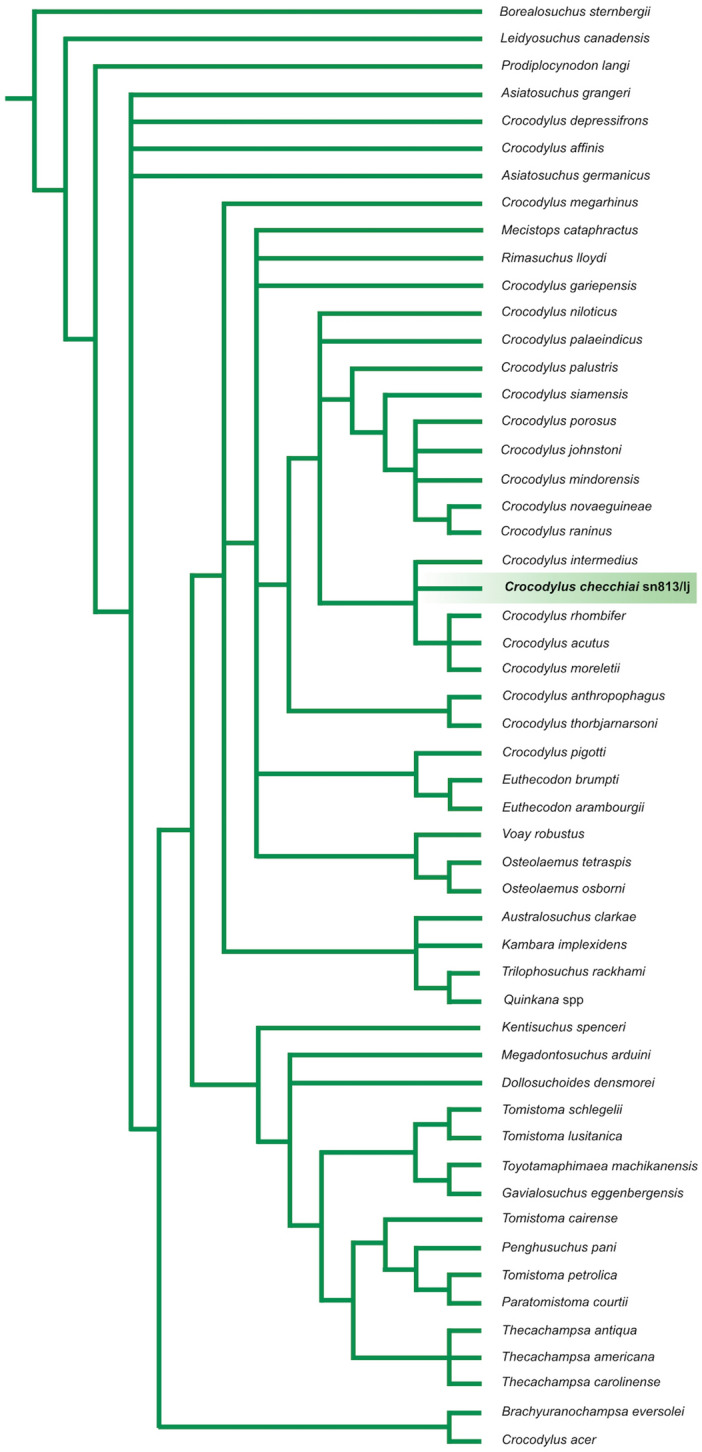
Implied weighting tree of a matrix comprising 53 taxa and 189 characters based on Brochu and Storrs9. Crocodylus checchiai sn813/lj is retrieved in a polytomy along with all the four extant American Crocodylus.
In the second analysis (see results in Fig. 5) we obtained 1 tree 64 steps long and with adjusted homoplasy = 5.99, Consistency Index = 0.55 and Retention index = 0.70. Crocodylus niloticus is the outgroup of the clade including, as successive ingroups, C. checchiai, C. intermedius, C. falconensis and all the other extant Crocodylus species from America.
Figure 5.
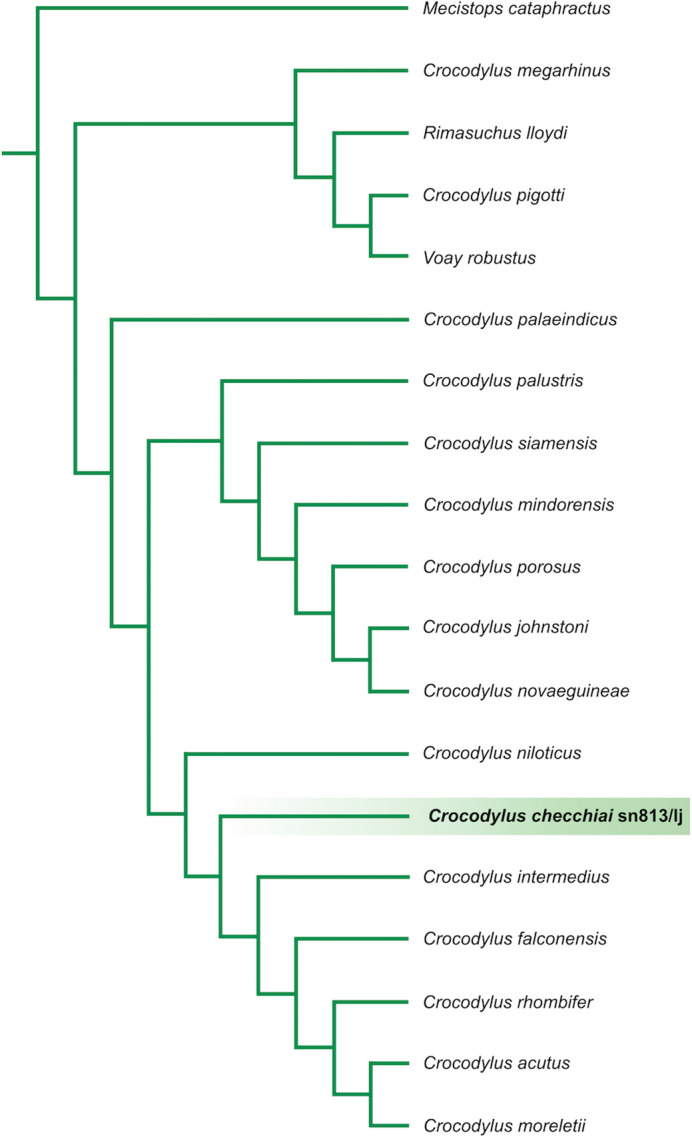
The addition of the coding of Crocodylus checchiai based on the late Miocene Libyan specimen sn813/lj to an updated version of the matrix by Scheyer et al.19 provides (“implicit enumeration search” with implied weighting) a topology with C. checchiai in intermediate position between the African Crocodylus niloticus and American crocodiles, with the early Pliocene Crocodylus falconensis nested within the American clade and not at its base as in Scheyer et al.19.
Discussion
The phylogenetic relationships of Crocodylus checchiai
Despite the four American Crocodylus species share with the African extinct Crocodylus checchiai an evident, unique morphological character, the mid rostral boss, a close phylogenetic relationship among them was not retrieved in earlier studies.
Maccagno14 was aware that the median boss is present in at least some of the American crocodile species (she listed “Cr. americanus” and “Cr. rombifer” [sic]) but since she discussed this and other characters in a taxonomic context only, she simply discarded the conspecificity of the Libyan taxon without facing its phylogenetic relationships. On the contrary, on the basis of a mixture of characters of little or no phylogenetic value, and not taking into consideration intra and interspecific variability and the effects of postmortem deformation, Maccagno15 identified in the extinct Crocodylus palaeindicus and the Asian taxa the closest relationships of the Libyan crocodylians. As already remarked by Brochu6, Tchernov24 considered C. checchiai as the sister taxon of C. niloticus, whereas according to Leakey et al.25 these two taxa were even synonyms. Hect13 correctly underlined a link with the American taxa by stating that “the similarity of the dorsal skull morphology of Crocodylus checchiai to the two forms of American freshwater crocodilians, Crocodylus moreletti [sic] and C. rhombifera [sic], indicated that these two forms may be the relics of a more widespread species distribution”. He concluded that even if it could be a homoplastic character, the promontorium should be considered a synapomorphy of the group on the basis of the knowledge of that time.
The first, and up to now only, phylogenetic analysis that evaluated the relationships of C. checchiai9,26 confirmed the inclusion of this species in the genus Crocodylus, but, having retrieved a broad politomy, lacks of enough resolution to provide precise indications.
The scoring of C. checchiai by Brochu and Storrs9 was based on the two skulls KNM-LT 23108 and KNM-LT 26618 as well as postcranial material from the late Miocene-early Pliocene of Tanzania and differs from the scoring we used (based on sn813/lj) for a few characters (for further discussion see Supplementary Note 1). In particular, two characters were scored differently in the two matrixes (and 29 characters were scored in Brochu and Storrs9, but not by us; ten characters were scored by us for the first time). The anterior process of the ectoperygoids is cleary forked in sn813/lj but was scored as pointed in Brochu and Storrs9. Remarkable is the relationship between the nasals and the external naris because on one side Maccagno clearly stated that in both the now lost holotype of C. checchiai and sn813/lj (as we confirmed; see Fig. 6) the nasals participated in the posterior rim of the external naris, as described and shown by Delfino18 for specimen 30P24A likewise coming from As Sahabi (further inspection of the latter confirmed that the nasals separate the premaxillae behind the naris for about 10 mm). On the other side, Hect13 described and figured the opposite condition in 3P84A from the same area. The fact that the material from Tanzania shows nasals not reaching (at least externally) the naris should not by itself grant specific separation, but requires further evaluation of the possible polymorphy of the character. The CT scans did not allow us to confirm the presence of the linear array of pits in the caviconchal recesses in sn813/lj, whose presence was considered as unknown in C. checchiai9 even if character 101 was scored as 1 (presence of a linear array of pits) in their matrix.
Figure 6.

CT investigation of the relationship between the nasals and the premaxillae in sn813/lj. Top left: anterior tip of the snout in dorsal view showing the planes of the sections in (A), (B) and (C). (A) anterior transversal section do not involve the nasals; (B): posterior transversal section cutting the nasals and showing that the premaxillae are entirely separated by the nasals; (C): sagittal section showing that the nasals are slightly lowered in the area of the posterior process of the premaxillae. Note in (A) and (B) how the nasals are raised to form the medial boss of the snout. Scale bar: 3 cm.
Towards the origin of the Neotropical crocodiles
According to the results obtained by Oaks2, there are two possible scenarios for the dispersal of Crocodylus to the Neotropics: from Australasia to the Neotropics and then to Africa or, alternatively, from Australasia to Africa and then to the Neotropics.
Thanks to our results, the late Miocene Crocodylus checchiai could offer a geographic and phylogenetic link between the African Crocodylus niloticus and the American clade whose oldest member is C. falconensis from the early Pliocene, providing direct evidence for the dispersal from Africa to America and suggesting to discard the other option. As shown in Fig. 4, resulting from the analysis based on the data matrix by Brochu and Storrs9, but with the scoring of the African C. checchiai sn813/lj from Libya, the latter belongs to the American clade, even if the resolution of the retrieved trees does allow us to place it, as theoretically expected because of its age, at the base of the extant Neotropical taxa.
Conversely, the application of the implied weighting method to an updated version of the smaller matrix (only 32 characters and 18 taxa) published by Scheyer et al.19, with the addition of our scoring of C. checchiai, provides a well-resolved topology (Fig. 5) that places C. niloticus at the base of a branch including as successive ingroups C. checchiai, Crocodylus intermedius, Crocodylus falconensis, and then the other extant American species (for further discussion see Supplementary Note 2). The extinct C. falconensis was recently described from the early Pliocene of Venezuela19 and, representing the geologically oldest palaeontological evidence of the presence of Crocodylus in America, provides a term ante quem for the dispersal of Crocodylus. As for the fossil record of Crocodylus in America, to the remains mentioned by Brochu6 we should add the recently described fragmentary remains from the late Pliocene Ware Formation (Colombia) referred to Crocodylus sp. by Moreno-Bernal et al.27. None of them is older than the early Pliocene and, in fact, Nicolai and Mazke3 considered as Pliocene the dispersal of Crocodylus from Africa to America.
The late Miocene age of the African Crocodylus checchiai slightly predates previous estimates and is however fully congruent with a westward dispersal towards America and the appearance of Crocodylus in America in the early Pliocene fossil record. It seems likely that at the same time Crocodylus dispersed northward from Africa to Europe across the Tethys (or alternatively from Asia to Europe3) as testified by several remains coming from different late Miocene southern European localities. At least the remains from the fissures fillings of Gargano promontory28 can be confidently referred to Crocodylus sp., but also the remains from Montebamboli29 and Scontrone30 have been tentatively referred to the same taxon. Interestingly enough, the presence of C. checchiai has been cited for the Iberian late Miocene site of Venta del Moro31, but a recent revision of this fragmentary material did not confirm the specific identification and suggested that the material could belong to an undetermined crocodile (Delfino, personal observations).
Remarkably, thanks to the evolution of range-expansion-promoting traits (salt tolerance32, but not only, see Nicolai and Mazke3) Crocodylus started to disperse across the globe very quickly around the end of the Miocene and successfully replaced the local taxa that went extinct because of climatic changes and related environmental remodeling6,19,27,30,33,34,35.
Methods
Tomographic analysis
Given the rarity of complete and well preserved skulls of C. checchiai, the non-destructive method of CT scanning36–39 was chosen to observe and analyse the internal anatomical features of the specimen sn813/lj. The fossil was scanned in its entirety in the coronal slice plane from front to back using a Philips Brilliance CT 64-channel scanner at M.G. Vannini Hospital (Rome). The scanning resulted in 670 slices with dimensions of 531 × 531 pixels. The slice thickness is 0.8 mm with an interslice space of 0.4 mm. The CT data were processed using OsiriX 5.5.2 and Materialise Mimics 20.0. The final 3D model was rendered with ZBrush 4R6.
Phylogenetic analysis
We performed two different phylogenetic analyses in TNT software40 (Supplementary Data Files S1-S2). The first analysis was based on Brochu and Storrs9: we replaced their coding of Crocodylus checchiai based on Tanzanian material with our coding of the same taxon based on the Libyan specimen sn813/lj (Supplementary Data Files S1 and Supplementary Note 3; for an alternative analysis with both Libyan and Tanzanian codings see Supplementary Note 4; Supplementary Fig. S1 and Supplementary Data File S3), thus building a matrix comprising 53 taxa and 189 characters. Borealosuchus sternbergii was set as outgroup. We performed a “new technology search”41. Characters were weighted equally, and multistate characters were left unordered.
In the second analysis, we added our coding of Crocodylus checchiai (Supplementary Note 5) to an updated version of the matrix published in Scheyer et al.19 that includes also C. falconensis, i.e. an extinct species that represent the oldest Crocodylus from South America. The matrix comprised therefore 19 taxa and 32 characters (Supplementary Data File S2). In this case we performed an “implied weighting” with ùk parameter = 341. Multistate characters were left unordered.
Supplementary information
Acknowledgements
We gratefully thank M.R. Palombo, M. Lustrino and L. Riti for access to the material. Sincere appreciation is expressed to M. Danti which gave us access, assistance and facilities for the making of the CT scanning. We thank R.L.Bernor and S. Scardigli for discussion and for sharing information about the Sahabi site, and Carlo Petrocchi historical data. L.C.M. Wencker kindly provided the phylogenetic analysis included in the Supplementary Information. This research was supported by: Fondi di Ateneo (Sapienza Università di Roma) 2015 – Finisterrae (C26A153RNH) and Grants Sapienza PaleoFactory Lab (A.C. 07.04.010) to RS; Progetti di Ricerca finanziati dall’Università degli Studi di Torino (ex 60% – 2018–2019), Generalitat de Catalunya (CERCA Program) and Spanish Agencia Estatal de Investigación (CGL2016-76431-P, AEI/FEDER, EU) to MD; Fondi di Ateneo (Università degli Studi di Firenze) and NGS (8788–10) to LR. LR coordinates the Italian participation within the frame of the ELNRP (East Libya Neogene Research Project, directed by N.T. Boaz). We hope that the social and political troubles affecting the Libyan people will end as soon as possible allowing, among others, a proper safeguard of the exceptional cultural heritage of their country.
Author contributions
M.D., D.A.I. and B.M. designed the research. M.D. wrote the manuscript with input from the other authors. D.A.I. processed and analysed the CT images. B.M. and P.P. performed the phylogenetic analyses. L.R. and R.S. give critical ideas and revision comments on data analysis and manuscript writing. All authors contributed equally to the interpretation of results and discussed the manuscript.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Massimo Delfino, Email: massimo.delfino@unito.it.
Dawid A. Iurino, Email: dawid.iurino@uniroma1.it
Supplementary information
is available for this paper at 10.1038/s41598-020-68482-5.
References
- 1.Uetz, P. & Hošek, J. The Reptile Database. https://www.reptile-database.org (2019). Accessed 5 Nov 2019.
- 2.Oaks JR. A time-calibrated species tree of Crocodylia reveals a recent radiation of the true crocodiles. Evolution. 2011;65(11):3285–3297. doi: 10.1111/j.1558-5646.2011.01373.x. [DOI] [PubMed] [Google Scholar]
- 3.Nicolaï MPJ, Matzke NJ. Trait-based range expansion aided in the global radiation of Crocodylidae. Glob. Ecol. Biogeogr. 2019;1:2. doi: 10.1111/geb.12929. [DOI] [Google Scholar]
- 4.Hekkala E, et al. An ancient icon reveals new mysteries: mummy DNA resurrects a cryptic species within the Nile crocodile. Mol. Ecol. 2011;20:4199–4215. doi: 10.1111/j.1365-294X.2011.05245.x. [DOI] [PubMed] [Google Scholar]
- 5.Meredith RW, Hekkala ER, Amato G, Gatesy J. A phylogenetic hypothesis for Crocodylus (Crocodylia) based on mitochondrial DNA: evidence for a trans-Atlantic voyage from Africa to the New World. Mol. Phylogenetics Evol. 2011;60:183–191. doi: 10.1016/j.ympev.2011.03.026. [DOI] [PubMed] [Google Scholar]
- 6.Brochu CA. Phylogenetic relationships and divergence timing of Crocodylus based on morphology and the fossil record. Copeia. 2000;2000(3):657–673. doi: 10.1643/0045-8511(2000)000[0657:PRADTO]2.0.CO;2. [DOI] [Google Scholar]
- 7.Brochu CA. Congruence between physiology, phylogenetics and the fossil record on crocodylian historical biogeography. In: Grigg GC, Seebacher F, Franklin CE, editors. Crocodilian Biology and Evolution. New South Wales: Surrey Betty & Sons; 2001. pp. 9–28. [Google Scholar]
- 8.Brochu CA. Phylogenetic approaches toward crocodylian history. Annu. Rev. Earth Planet. Sci. 2003;31:357–397. doi: 10.1146/annurev.earth.31.100901.141308. [DOI] [Google Scholar]
- 9.Brochu CA, Storrs GW. A giant crocodile from the Plio-Pleistocene of Kenya, the phylogenetic relationships of Neogene African crocodylines, and the antiquity of Crocodylus in Africa. J. Vertebr. Paleontol. 2012;32(3):587–602. doi: 10.1080/02724634.2012.652324. [DOI] [Google Scholar]
- 10.Fermino BR, et al. Shared species of crocodilian trypanosomes carried by tabanid flies in Africa and South America, including the description of a new species from caimans Trypanosoma kaiowa n. sp. Parasites Vectors. 2019;12(1):225. doi: 10.1186/s13071-019-3463-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hekkala ER, Amato G, DeSalle R, Blum MJ. Molecular assessment of population differentiation and individual assignment potential of Nile crocodile (Crocodylus niloticus) populations. Conserv. Genet. 2010;11:1435–1443. doi: 10.1007/s10592-009-9970-5. [DOI] [Google Scholar]
- 12.Schmitz A, et al. Molecular evidence for species level divergence in African Nile Crocodiles Crocodylus niloticus (Laurenti, 1786) C. R. Palevol. 2003;2(8):703–712. doi: 10.1016/j.crpv.2003.07.002. [DOI] [Google Scholar]
- 13.Hecht MK. Fossil snakes and crocodilians from the Sahabi Formation of Libya. In: Boaz NT, El-Arnauti A, Wahid Gaziry A, De Heinzelin J, Boaz DD, editors. Neogene Palaeontology and Geology of Sahabi. New York: Alan R. Liss. Inc.; 1987. pp. 101–106. [Google Scholar]
- 14.Maccagno AM. Descrizione di una nuova specie di “Crocodilus” del Giacimento di Sahabi (Sirtica) Atti Accad. Naz. Lincei Mem. Classe Sci. Fis. Mat. Nat. Sez. II Fis. Chim. Geol. Paleontol. Mineral. Ser. VIII. 1947;1(2):61–96. [Google Scholar]
- 15.Maccagno AM. I coccodrilli di Sahabi. Rend. Accad. Naz. XL. 1952;4(3):73–117. [Google Scholar]
- 16.Rook L. The discovery of Sahabi site: Ardito Desio or Carlo Petrocchi? In Boaz, N.T. El-Arnauti, A., Pavlakis, P. & Salem, M. (eds) Circum-mediterranean neogene geology and biotic evolution: the perspective from Libya. Garyounis Sci. Bull. 2008;5:11–18. [Google Scholar]
- 17.Pandolfi L, Rook L. The latest Miocene Rhinocerotidae from Sahabi (Lybia) C. R. Palevol. 2019;18:442–448. doi: 10.1016/j.crpv.2019.03.002. [DOI] [Google Scholar]
- 18.Delfino M. New remains of Crocodylus checchiai Maccagno, 1947 (Crocodylia, Crocodylidae) from the Late Miocene of As Sahabi, Libya. In Boaz N. T., El-Arnauti A., Pavlakis P. & Salem M. (eds). Circum-Mediterranean geology and biotic evolution during the Neogene Period: the perspective from Libya. Garyounis Sci. Bull. 2008;5:111–118. [Google Scholar]
- 19.Scheyer TM, et al. Crocodylian diversity peak and extinction in the late Cenozoic of the northern Neotropics. Nat. Commun. 2013;4:1907. doi: 10.1038/ncomms2940. [DOI] [PubMed] [Google Scholar]
- 20.El-Shawaihdi MH. Stratigraphy of the Neogene Sahabi units in the Sirt Basin, northeast Libya. J. Afr. Earth Sci. 2016;118:87–106. doi: 10.1016/j.jafrearsci.2016.02.015. [DOI] [Google Scholar]
- 21.Bernor RL, Rook L. A current view of Sahabi large mammal biogeographic relationships. In Boaz, N. T., El-Arnauti, A., Pavlakis, P. & Salem, M. (eds) Circum-mediterranean neogene geology and biotic evolution: the perspective from Libya. Garyounis Sci. Bull. 2008;5:285–292. [Google Scholar]
- 22.Bernor RL, Boaz NT, Rook L. Eurygnathohippus feibeli (Perissodacyla: Mammalia) from the Late Miocene of As Sahabi (Libya) and its evolutionary and biogeographic significance. Boll. Soc. Paleont. Ital. 2012;51:39–48. [Google Scholar]
- 23.Gradstein F, Ogg J, Smith A. A Geologic Time Scale 2012. Cambridge: Cambridge University Press; 2012. [Google Scholar]
- 24.Tchernov E. Evolution of the Crocodiles in East and North Africa. Paris: CNRS; 1986. [Google Scholar]
- 25.Leakey MG, et al. Lothagam: a record of faunal change in the Late Miocene of East Africa. J. Vertebr. Paleontol. 1996;16(3):556–570. doi: 10.1080/02724634.1996.10011339. [DOI] [Google Scholar]
- 26.Brochu CA. Pliocene crocodiles from Kanapoi, Turkana Basin Kenya. J. Hum. Evol. 2020;140:102410. doi: 10.1016/j.jhevol.2017.10.003. [DOI] [PubMed] [Google Scholar]
- 27.Moreno-Bernal JW, Head J, Jaramillo CA. Fossil crocodilians from the High Guajira Peninsula of Colombia: Neogene faunal change in northernmost South America. J. Vertebr. Paleontol. 2016;36(3):e1110586. doi: 10.1080/02724634.2016.1110586. [DOI] [Google Scholar]
- 28.Delfino M, Böhme M, Rook L. First European evidence for transcontinental dispersal of Crocodylus (late Neogene of southern Italy) Zool. J. Linn. Soc. 2007;149:293–307. doi: 10.1111/j.1096-3642.2007.00248.x. [DOI] [Google Scholar]
- 29.Delfino M, Rook L. African crocodylians in the Late Neogene of Europe. A revision of Crocodylus bambolii Ristori, 1890. J. Paleontol. 2008;82(2):336–343. doi: 10.1666/06-079.1. [DOI] [Google Scholar]
- 30.Delfino M, Rossi MA. Fossil crocodylid remains from Scontrone (Tortonian, Southern Italy) and the Late Neogene Mediterranean biogeography of crocodylians. Geobios. 2013;46:25–31. doi: 10.1016/j.geobios.2012.10.006. [DOI] [Google Scholar]
- 31.Montoya P, et al. Las nuevas excavaciones (1995–2006) en el yacimiento del Mioceno final de Venta del Moro Valencia. Estud. Geol. 2006;62:313–326. doi: 10.3989/egeol.0662128. [DOI] [Google Scholar]
- 32.Taplin LE, Grigg GC. Historical zoogeography of the eusuchian crocodilians: a physiological perspective. Am. Zool. 1989;29:885–901. doi: 10.1093/icb/29.3.885. [DOI] [Google Scholar]
- 33.Brochu CA, Nieves-Rivera ÁM, Vélez-Juarbe J, Daza-Vaca JD, Santos H. Tertiary crocodylians from Puerto Rico: evidence for Late Tertiary endemic crocodylians in the West Indies? Geobios. 2007;40:51–59. doi: 10.1016/j.geobios.2005.10.008. [DOI] [Google Scholar]
- 34.Mannion PD, et al. Climate constrains the evolutionary history and biodiversity of crocodylians. Nat. Commun. 2015;6:8438. doi: 10.1038/ncomms9438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Díaz Aráez JL, et al. New remains of Diplocynodon (Crocodylia: Diplocynodontidae) from the Early Miocene of the Iberian Peninsula. C. R. Palevol. 2017;16:12–26. doi: 10.1016/j.crpv.2015.11.003. [DOI] [Google Scholar]
- 36.Iurino DA, Danti M, Della Sala SW, Sardella R. Modern techniques for ancient bones: vertebrate palaeontology and medical CT analysis. Boll. Soc. Paleontol. Ital. 2013;52:145–155. [Google Scholar]
- 37.Iurino DA, Sardella R. CT scanning analysis of Megantereon whitei (Carnivora, Machairodontinae) from Monte Argentario (Early Pleistocene, central Italy): evidence of atavistic teeth. Naturwissenschaften. 2014;101(12):1099–1106. doi: 10.1007/s00114-014-1249-0. [DOI] [PubMed] [Google Scholar]
- 38.Lautenschlager S, Butler RJ. Neural and endocranial anatomy of Triassic phytosaurian reptiles and convergence with fossil and modern crocodylians. PeerJ. 2016;4:e2251. doi: 10.7717/peerj.2251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Cherin M, et al. Synchrotron radiation reveals the identity of the large felid from Monte Argentario (Early Pleistocene, Italy) Sci. Rep. 2018;8(1):8338. doi: 10.1038/s41598-018-26698-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Goloboff PA, Farris JS, Nixon KC. TNT, a free program for phylogenetic analysis. Cladistics. 2008;24(5):774–786. doi: 10.1111/j.1096-0031.2008.00217.x. [DOI] [Google Scholar]
- 41.Goloboff PA. Parsimony and weighting: a reply to Turner and Zandee. Cladistics. 1995;11(1):91–104. doi: 10.1111/j.1096-0031.1995.tb00006.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.