

Abstract
Narrow‐leafed lupin (Lupinus angustifolius L.) cultivation was transformed by 2 dominant vernalization‐insensitive, early flowering time loci known as Ku and Julius (Jul), which allowed expansion into shorter season environments. However, reliance on these loci has limited genetic and phenotypic diversity for environmental adaptation in cultivated lupin. We recently predicted that a 1,423‐bp deletion in the cis‐regulatory region of LanFTc1, a FLOWERING LOCUS T (FT) homologue, derepressed expression of LanFTc1 and was the underlying cause of the Ku phenotype. Here, we surveyed diverse germplasm for LanFTc1 cis‐regulatory variation and identified 2 further deletions of 1,208 and 5,162 bp in the 5' regulatory region, which overlap the 1,423‐bp deletion. Additionally, we confirmed that no other polymorphisms were perfectly associated with vernalization responsiveness. Phenotyping and gene expression analyses revealed that Jul accessions possessed the 5,162‐bp deletion and that the Jul and Ku deletions were equally capable of removing vernalization requirement and up‐regulating gene expression. The 1,208‐bp deletion was associated with intermediate phenology, vernalization responsiveness, and gene expression and therefore may be useful for expanding agronomic adaptation of lupin. This insertion/deletion series may also help resolve how the vernalization response is mediated at the molecular level in legumes.
Keywords: cis‐regulation, insertion/deletion (INDEL), variant series, FLOWERING LOCUS T (FT)
Short abstract
“Greater genetic and phenotypic diversity for flowering time would enhance the adaptation of narrow‐leafed lupin to Australian and northern European agricultural environments. We studied the major floral integrator gene in narrow‐leafed lupin, LanFTc1, and found a series of three overlapping independent deletions within the promoter region that reduce vernalization responsiveness, shorten flowering time, and heighten expression of LanFTc1. The smallest deletion results in a unique intermediate phenotype, which may be valuable for breeding purposes and expanding the agronomic adaptation of lupin to new and existing production regions. Additionally, the insertion/deletion series may also help to increase knowledge of how signalling within the vernalization pathway is mediated at a molecular level in legumes.”
1. INTRODUCTION
Narrow‐leafed lupin (Lupinus angustifolius L.) is one of three fully domesticated Old World Lupinus species originating from the Mediterranean and northern Africa (Gladstones, 1974). It is predominantly grown in Australia and several northern European countries, including Poland, Russia, Germany, Ukraine, and Belarus, as a winter and summer annual pulse crop, respectively (FAO, 2014; Gladstones, 1970). The grain is most commonly marketed as a high‐protein livestock and aquaculture feed. However, in light of the nutritional and metabolomics properties of narrow‐leafed lupin seed (Lima‐Cabello et al., 2017) and the associated benefits to human health and disease prevention (Foyer et al., 2016; Kouris‐Blazos & Belski, 2016), it is also being promoted in the human food market. In addition to high‐protein grain production, narrow‐leafed lupin has great agricultural value as a superior break crop. Lupin crops mobilize soil‐bound phosphorus through carboxylate exudation (Lambers, Clements, & Nelson, 2013) and improve soil nitrogen through symbiosis, which, together with disease and weed control, is beneficial to the performance of subsequent crop rotations (Seymour, Kirkegaard, Peoples, White, & French, 2012).
Following domestication, one of the most important achievements in narrow‐leafed lupin breeding has been the manipulation of phenology. In most wild populations, a prolonged period of exposure to cold winter temperatures, known as vernalization, is required to promote the transition from vegetative to reproductive growth (Rahman & Gladstones, 1972). However, this trait caused great difficulty in the global transitioning of the species from a green manure and fodder crop into a broad acre grain crop. Within key production zones of Australia, winter temperatures are often mild and incapable of reliably saturating the vernalization requirement from year to year, leading to delayed phenology, susceptibility to terminal drought stress and reduced final yields (Berger, Buirchell, Luckett, Palta, et al., 2012; Gladstones, 1977). Similarly, strong vernalization requirement was also a problem for summer cropping of lupin in northern Europe, where cool and wet autumn conditions hindered grain maturation, particularly if sowing was delayed by the late arrival of spring or growing regions receive high amounts of summer rainfall (Kubok, 1988; Mikołajczyk, 1966). Two naturally occurring dominant mutations were discovered independently during the 1960s in Australia and northern Europe and effectively removed the requirement for vernalization, thereby improving the adaptation and yield stability of narrow‐leafed lupin in short season environments. The most widely adopted of these is Ku, which arose as a spontaneous mutant in the cultivar “Borre” and is capable of advancing flowering time by up to 5 weeks in Australia (Gladstones & Hill, 1969). It has been selected in almost all elite Australian cultivars released since the 1970s (Cowling, 1999; Stefanova & Buirchell, 2010) and has also been commonly used in European breeding programs (Boersma, Buirchell, Sivasithamparam, & Yang, 2007). The second mutation, Julius (Jul), was first discovered in the Russian bred cultivar, Krasnolistny, and became an important source of early phenology in Polish breeding during the 1980s (Kubok, 1988; Mikołajczyk, 1966). Due to striking similarity in photoperiodic as well as vernalization responsiveness, Ku and Jul were thought to be controlled by the same gene (Rahman & Gladstones, 1972).
Global production of narrow‐leafed lupin grain has stagnated in recent years with a leading cause being the limited genetic and adaptive diversity available within domesticated breeding pools of narrow‐leafed lupin (Berger, Buirchell, Luckett, & Nelson, 2012). This lack of diversity largely stems from the species' recent domestication, which was based on a small number of founding individuals, and subsequent strong and persistent selection for key traits, such as phenology, all of which have resulted in severe genetic bottlenecks (Berger, Buirchell, Luckett, & Nelson, 2012; Cowling, 1999; Stefanova & Buirchell, 2010). Among the consequences for these bottlenecks is a reduction in genetic variation for flowering time within a domestic background. Furthermore, any variation that is present is hidden by the dominant overriding effect of the Ku and Jul loci.
A lack of phenological diversity is problematic for two main reasons. First, despite the profound influence Ku and Jul have had in adapting narrow‐leafed lupin to short‐season production environments, many regions still lack options for cultivars adequately matched to their respective climates (Berger, Buirchell, et al., 2008; Berger, Buirchell, Luckett, Palta, et al., 2012). The southwest of Western Australia and the eastern states of Australia are two such regions, where longer growing seasons coupled with reduced evapotranspiration could support later flowering cultivars and achieve higher yields (C. Chen, Fletcher, Lawes, Berger, & Roberston, 2017). Second, the reliance on Ku and lack of genetic diversity due to domestication bottlenecks will reduce the ability of breeders to select stress tolerant cultivars adapted to climate change, which threatens future yield potential of lupins and crop harvestability (Nelson, Berger, & Erskine, 2010). New genetic variation for phenology is required to increase the adoption of narrow‐leafed lupin in longer season environments and to maintain or extend feasible production zones in the face of climate change.
Recently, the genetic identify of Ku was revealed as a FLOWERING LOCUS T (FT) homologue, LanFTc1 (Nelson et al., 2017). In Arabidopsis, FT has a well‐defined role as a floral integrator gene, coordinating signals from the vernalization, photoperiod, and circadian clock pathways to promote flowering at an opportune time (Turck, Fornara, & Coupland, 2008). A 1,423‐bp deletion in the promoter region of LanFTc1 was implicated as the casual mutation for the Ku allele, as its presence was perfectly predictive of vernalization responsiveness in 216 wild and domesticated accessions and was associated with derepressed expression of LanFTc1 in the absence of vernalization (Nelson et al., 2017). Given the demonstrated capacity for mutations in the noncoding sequence of this gene to modify its expression and plant phenology, we endeavoured to find other polymorphisms that may affect cis‐regulation of LanFTc1 and provide alternative sources of flowering time variation. Here, we report the discovery of a series of insertion/deletions (INDELs) in the 5' regulatory region of this gene, which are associated with altered vernalization responsiveness, flowering time, and LanFTc1 gene expression in the absence of vernalization.
2. MATERIALS AND METHODS
2.1. Screening for polymorphisms within the genomic region of LanFTc1 in diverse germplasm
Polymorphisms in the LanFTc1 genomic region were explored in a panel of 48 narrow‐leafed lupin accessions (Table S1) comprising (a) the species reference genome cultivar, Tanijl (Hane et al., 2017); (b) 43 accessions, including 30 genetically diverse wild accessions representing the natural geographic range of the species throughout the Mediterranean Basin (Mousavi‐Derazmahalleh et al., 2017) and 13 fully domesticated or semidomesticated accessions from Australia and Europe, all for which short‐read sequencing data were generated to assemble the LanFTc1 region; (c) Krasnolistny, the first cultivar described as carrying the Jul early flowering time locus (Mikołajczyk, 1966); and (d) three Polish cultivars (Kazan, Mirela, and Sur) with Krasnolistny in their pedigree (Kubok, 1988).
The genomic sequence encompassing roughly 7‐Kb upstream and 2‐Kb downstream of the LanFTc1 coding region was extracted from the Tanjil narrow‐leafed lupin reference genome (Hane et al., 2017) plus the 43 accessions with short‐read sequencing data by aligning Illumina Paired End reads from each accession to the Tanjil reference using Bowtie2 v2.2.9 (‐‐sensitive; Langmead & Salzberg, 2012). Variants were called using samtools and bcftools (Li, 2011; Li et al., 2009), which were then filtered to remove artefactual sequence variant calls arising from misalignments close to large (>1,000 bp) INDELs and polymorphisms that were physically disrupted by others.
Both Tanjil and the P27255 wild‐type LanFTc1 sequence (GenBank ID KT862491) served as references to genotype the 1,423‐bp INDEL polymorphism previously identified by Nelson et al. (2017) in the 5' regulatory region and to survey for other alternative INDEL variations in this same region. After discovering additional INDELs in the 7‐Kb sequence upstream of LanFTc1, PCR primers were designed in the immediately adjacent sequences to screen for presence/absence of these INDELs for those accessions (Krasnolistny, Kazan, Mirela, and Sur) for which no re‐sequencing data were obtained. A summary of the INDELs, the different PCR primers assay, and the conditions for PCR amplification are provided in Table S2. To confirm the size and boundary of the newly identified INDELs, as determined from alignment of the short‐read sequences to the P27255 and Tanjil references, the nucleotide sequence of PCR products was determined by Sanger sequencing.
Additional INDELs and single nucleotide polymorphisms (SNPs) were also assessed in the coding and remaining noncoding sequences within the extracted LanFTc1 genomic region in the 44 wild and domestic accessions using Tanjil as the reference genome. The P27255 wild‐type LanFTc1 sequence (GenBank ID KT862491), encompassing approximately 5‐Kb upstream and 800‐bp downstream of the LanFTc1 coding region, was used as the reference to call variants within the 1,423‐bp sequence in the 5' regulatory region that is deleted in the Tanjil reference genome.
2.2. Measuring degree‐days to flowering and vernalization responsiveness in diverse germplasm
The panel of 48 diverse accessions was phenotyped for time to flowering in two partially replicated trials (n = 1–3, as outlined in Table S1) for preliminary assessment of LanFTc1 polymorphism genotype effect on vernalization responsiveness and time to flowering. The first trial included 40 accessions representing the 0‐, 1,208‐, and 1,423‐bp INDEL variants. Data were gathered for all accessions except for P22603, which failed to germinate. In the second trial, six accessions carrying the 5,162‐bp INDEL were compared with 10 representatives of the three other INDEL variants.
All seeds were germinated in Jiffy‐7® peat pellets within a controlled environment room (CER) at The University of Western Australia (Perth, Australia), which was maintained at 20 °C constant temperature and with a 14‐hr photoperiod. Two vernalization treatments were provided: (a) a full vernalization treatment in which 2‐day old seedlings were transferred to a 4 °C room (14‐hr photoperiod) for 32 days before transferring to the CER for a further 140 days and (b) a mild, partial vernalization treatment, whereby 7‐day old seedlings were transferred to the 4 °C room for a total of 8 days before transferring to the CER for a further 140 days. A mild vernalization treatment was preferred to a fully non‐vernalizing treatment where accessions with a strong vernalization requirement would not flower at all. Thus, the mild vernalization treatment was designed to allow the degree of vernalization responsiveness to be measureable in the most strongly vernalization responsive accessions. All plants were transferred to the CER on the same day with approximately the same accumulated degree‐days (approximately 190 degree‐days, with 0 °C as the baseline temperature) and were placed in a randomized block design.
Flowering was scored immediately after anthesis, which was indicated by an erect standard petal (i.e., open flower) or the changing colour of petals. To accommodate the few accessions that did not flower within the allocated time of the experiment, even with mild vernalization treatment, flowering time was transformed to rate of flowering by taking the reciprocal of the degree‐days to flowering. Three‐way analysis of variance (ANOVA) was performed on the rate to flowering data using Genstat V.18, with vernalization treatment and deletion category as main effects, and accessions nested within deletion category to subdivide variance among and within categories. Category effects were compared using orthogonal contrasts by least significant difference. ANOVA was performed separately for each phenotyping trial, and residual plots used to identify outliers and check that errors were randomly and independently distributed.
2.3. Assessing the relationship of INDEL variation in the 5' regulatory region and LanFTc1 gene expression
Following the preliminary analysis of LanFTc1 polymorphism effect on phenotype, a subset of accessions representing four variants of prominent INDELs in the 5' regulatory region of LanFTc1 was selected from the original panel of 48 accessions to phenotype more precisely flowering time and vernalization responsiveness and to correlate these traits with LanFTc1 gene expression. The representative subset included the following narrow‐leafed lupins: (a) P27255, a wild Moroccan accession that is highly responsive to vernalization; (b) 83A:476, a vernalization‐insensitive Australian breeding line; (c) P22660, a wild Israeli accession with mild sensitivity to vernalization; (d) P29039, a vernalization‐insensitive Belarussian breeding line; and (e) Russian cultivar, Krasnolistny, also insensitive to vernalization. P27255 and 83A:476 are the parents for a wild × domestic F8 recombinant inbred line mapping population (Boersma et al., 2005; Kroc, Koczyk, Święcicki, Kilian, & Nelson, 2014; Nelson et al., 2006; Nelson, Moolhuijzen, et al., 2010).
Seeds were scarified and imbibed in Milli‐Q water for 6 hr before being immediately sown (non‐vernalized treatment) or incubated in a darkened room at 4 °C for 21 days in petri dishes (vernalized treatment). On the day of sowing the vernalized seeds, both treatments had accumulated approximately equal degree‐days (calculated using baseline temperature of 0 °C). All plants were grown in a phytotron located at The University of Western Australia (Perth, Australia) with a diurnal temperature range of 18 ± 0.5 °C (day) to 14 ± 0.5 °C (night) and exposed to natural photoperiod (10‐ to 12‐hr daylight during May to October 2017). Flowering time and degree‐days to flowering were scored immediately after anthesis and the data analysed as outlined above.
The four uppermost fully emerged leaves were harvested for gene expression analyses from three biological replicates per treatment per accession at five growth stages: 4‐leaf, 8‐leaf, 12‐leaf, 16‐leaf, and flowering. Samples were harvested between 12:00 and 14:00 hr and immediately snap‐frozen in liquid nitrogen. RNA isolation, cDNA synthesis, and quantitative reverse transcription PCR were conducted according to the methods of Taylor, Jost, Erskine, and Nelson (2016) and Nelson et al. (2017). Briefly, the relative expression of LanFTc1 was calculated as the average cycle threshold (CT) for two primer pairs, which had previously been designed by Nelson et al. (2017) using transcript sequences from the draft Tanjil reference genome assembly (Kamphuis et al., 2015) to be specific to LanFTc1 and to target unique portions of the coding sequence for gene‐wide transcription assessment. The average LanFTc1 CT value was then normalized against Ubiquitin C, which had been validated as a robust reference gene under the same experimental conditions (Taylor et al., 2016), and relative expression was finally expressed as 40‐ΔCT. This method reports relative transcript abundance on a Log2 scale, where a value of 40 represents the mean level of expression of Ubiquitin C and the fold difference between treatments is calculated as 2ΔΔCT (where ΔΔCT is equal to difference in average ΔCT between vernalized and non‐vernalized treatments) when primer efficiency is approximately 2.0 (Bari, Pant, Stint, & Scheible, 2006).
Nested ANOVA and polynomial linear regression of relative gene expression over degree‐days to flowering were performed in Genstat V.18 as described above. Nested polynomial linear regression demonstrated nonsignificant differences between accessions within INDEL size categories, and therefore, only the INDEL size category results are presented (Figure 3a,b). The regressions were compared using orthogonal contrasts. The regression equations generated by Genstat were then used to plot smooth fitted curves, and we included the biological replicate data points for context (Figure 3a,b).
2.4. Characterizing the LanFTc1 promoter region
Relative to P27255, representing the wild‐type LanFTc1 sequence, three large, distinct INDELs were identified in the 5' regulatory region. Two of the INDELs, which were associated with modified gene expression and phenology, were assessed relative to the wild‐type for the presence/absence of transcription factor binding site motifs previously identified by Nelson et al. (2017). Putative binding site motifs were identified in that study using two open‐access web‐interface platforms, including JASPAR 2014, which contains CORE Plantae matrix models (Mathelier et al., 2014), and PLACE, which contains cis‐acting regulatory DNA elements in plants (Higo, Ugawa, Iwamoto, & Korenaga, 1999).
2.5. Assessment of linkage disequilibrium in the LanFTc1 genomic region
To determine the likelihood of other polymorphisms within the LanFTc1 genomic region being involved in modifying the response to vernalization, we measured the association of each polymorphism with vernalization responsiveness among the 44 accessions previously genotyped for polymorphisms in the LanFTc1 genomic region. This was done by measuring pairwise linkage disequilibrium (r 2) of SNP and INDEL variants identified relative to the P27255 wild‐type LanFTc1 reference sequence (see above) with the vernalization responsiveness phenotype, which was scored as a multi‐allelic genotype (unresponsive, mildly responsive, and responsive). An r 2 value was also calculated for the four INDEL variants as a single multi‐allelic polymorphism (0, 1,208, 1,423, and 5,162 bp). All pairwise r 2 values (‐‐ld‐window‐r2 0) were calculated using PLINK v1.9 (Purcell et al., 2007). Default filtering settings in PLINK were used to remove markers with low quality or that were almost monomorphic from analysis. A linear adjusted association analysis was also conducted in PLINK v1.9 to determine the significance and strength of the association between the sequence variants and vernalization responsiveness phenotype.
3. RESULTS
3.1. Screening for polymorphisms within the genomic region of LanFTc1 in diverse germplasm
A 1,423‐bp deletion between 4,248‐ and 2,826‐bp upstream of the ATG start codon of LanFTc1 was previously hypothesized as the causal sequence variant modifying vernalization responsiveness in the breeding line, 83A:476 (Ku), compared with the wild‐type LanFTc1 sequence represented by P27255, and no polymorphisms were observed between accessions in the coding sequence (Nelson et al., 2017). To dissect this further, we surveyed the LanFTc1 genomic region, from approximately 7‐Kb upstream to 2‐Kb downstream of the coding region in the Tanjil reference genome (Hane et al., 2017), for polymorphisms in 44 accessions of narrow‐leafed lupin. This analysis revealed a total of 260 SNPs and 56 INDELs relative to Tanjil, excluding the 1,423‐bp INDEL (Tables 1 and S3a). Importantly, no polymorphisms were found in the coding sequence. The genomic region features containing the most SNPs and INDELs (all less than 30 bp in length) were the large third intron and the 5' regulatory region, which contained approximately 50% and 32% of all polymorphisms. Variant calling using the P27255 wild‐type LanFTc1 sequence revealed a further 17 SNPs and eight small INDELs (less than 20 bp in length) within the 1,423‐bp sequence in the 5' regulatory region, which is deleted in the Tanjil reference genome (Table S3b).
Table 1.
A summary of SNP and INDEL polymorphisms (excluding the large INDEL within the 5' regulatory region) observed in the genomic region from approximately 7‐Kb upstream to 2‐Kb downstream of LanFTc1 in 44 accessions of narrow‐leafed lupin relative to the Tanjil narrow‐leafed lupin reference genome
| LanFTc1 genomic region feature | Coordinates on pseudochromosome NLL‐10 | Coordinates on scaffold_276_44 | Number of SNP polymorphisms | Number of INDEL polymorphisms |
|---|---|---|---|---|
| 5' regulatory region | 8,016,843–8,023,566 | 3,823–10,546 | 91 | 11 |
| 5' UTR | 8,023,567–8,023,842 | 10,547–10,822 | 1 | 1 |
| Exon 1 (CDS) | 8,023,843–8,024,040 | 10,823–11,020 | 0 | 0 |
| Intron 1 | 8,024,041–8,024,162 | 11,021–11,142 | 1 | 0 |
| Exon 2 (CDS) | 8,024,163–8,024,225 | 11,143–11,205 | 0 | 0 |
| Intron 2 | 8,024,226–8,024,381 | 11,206–11,361 | 2 | 3 |
| Exon 3 (CDS) | 8,024,382–8,024,420 | 11,362–11,400 | 0 | 0 |
| Intron 3 | 8,024,421–8,030,939 | 11,401–17,919 | 127 | 29 |
| Exon 4 (CDS) | 8,030,940–8,031,155 | 17,920–18,135 | 0 | 0 |
| 3' UTR | 8,031,156–8,031,424 | 18,136–18,404 | 3 | 2 |
| 3' regulatory region | 8,031,425–8,033,155 | 18,405–20,135 | 35 | 10 |
| Total | 260 | 56 |
Note. Genomic region features include regulatory regions adjacent to the coding sequence (CDS), and the untranslated regions (UTRs), exons (coding sequences), and introns (noncoding intragenic sequences) of LanFTc1. SNP = single nucleotide polymorphism; INDEL = insertion/deletion.
We then used P27255 and Tanjil as references to genotype the 1,423‐bp INDEL and determine if there were any other major INDEL variations in the 5' regulatory region in 42 additional wild and domestic accessions. The wild‐type LanFTc1 sequence was present in a total of 29 wild accessions from the Mediterranean and two vernalization responsive Australian cultivars (all known to carry the ku allele), whereas the 1,423‐bp deletion was present in 10 Australian and Polish Ku cultivars. Importantly, two new large INDEL variants were identified. The first of these was a 1,208‐bp deletion observed between 3,970‐ and 2,763‐bp upstream of the ATG start codon in the Israeli accession, P22660, relative to the P27255 wild‐type LanFTc1 sequence. The majority of this smaller deletion overlapped with that of the 1,423‐bp variant. However, as determined from Sanger sequencing (GenBank ID MH166758), the first 277 bp at the 5' end of the 1,423‐bp deletion was retained in P22660, whereas a further 62 bp immediately downstream of the 1,423‐bp variant had been deleted (Figure 1). The second INDEL variant was a prominent deletion of 5,162 bp in P29039 (a Belarussian breeding line) and Emir (a Polish cultivar) positioned between 6,209‐ and 1,048‐bp upstream of the ATG start codon relative to the P27255 reference. Sanger sequencing supported a shared origin of the 5,162‐bp deletion as both P29039 and Emir had identical deletion breakpoints (GenBank ID MH166759). This large deletion completely spanned the 1,423‐ and 1,208‐bp deletions (Figure 1).
Figure 1.
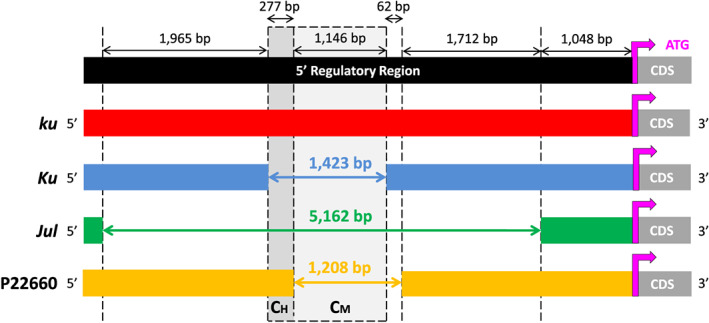
A schematic illustrating the positions of insertion/deletion genotypes in the 5' regulatory region of LanFTc1 relative to the start codon (ATG) of the coding sequence (CDS). The wild‐type LanFTc1 sequence (ku) was obtained from P27255, a wild Moroccan accession, and the 1,423‐bp deletion (Ku) from the Tanjil reference genome. A 5,162‐bp deletion (Julius or Jul) was found in several European breeding lines and cultivars, including Krasnolistny (Russian cultivar), P29039 (Belarussian breeding line), and Emir (Polish cultivar). A 1,208‐bp deletion was identified in P22660, a wild accession from Israel. Critical regions of the regulatory region, if deleted, enable high (CH, shaded dark grey) and moderately high (CM, shaded light grey) levels of LanFTc1 expression, respectively, relative to the wild‐type sequence
To determine if the narrow‐leafed lupin cultivars carrying Jul contain the wild‐type sequence or one of the three deletion genotypes, a PCR‐marker (Table S2) approach was used. This confirmed that Krasnolistny, the original Jul cultivar, and three Polish cultivars known to descend from it (Kazan, Mirela, and Sur) contain the 5,162‐bp deletion as detected in P29039 and Emir (Figure S1).
3.2. Measuring degree‐days to flowering and vernalization responsiveness in diverse germplasm
To explore whether the four prominent INDEL variants in the 5' regulatory region of LanFTc1 may affect vernalization responsiveness and phenology in narrow‐leafed lupin, we phenotyped rate to flowering (reciprocal of degree‐days to flowering) in the full panel of 48 accessions under both mildly and fully vernalizing conditions across two trials, with 39 accessions in the first trial (Figure 2a) and 17 accessions in the second trial (Figure 2b; Table S1). In both trials, there were strong INDEL variant by vernalization treatment interactions (p < .001), in which vernalization response was consistently proportional to flowering time (Figure 2a,b). Thus, the strongest vernalization response was observed in the late flowering wild‐type accessions (0‐bp INDEL), followed by the intermediate flowering 1,208‐bp INDEL accession and finally, the early flowering 1,423‐ and 5,162‐bp accessions, both of which had a small response to vernalization (Figure 2b). Accordingly, the large differences in rate to flowering observed between the 0‐, 1,208‐, 1,423‐ and 5,162‐bp variants under mild vernalization were greatly reduced under full vernalization. Thus, under fully vernalizing conditions, the strongly vernalization responsive wild‐types (0‐bp deletions) flowered at the same rate as the intermediate 1,208‐bp INDEL variant and only marginally slower than the weakly vernalization responsive accessions with the 1,423‐ and 5,162‐bp deletions.
Figure 2.
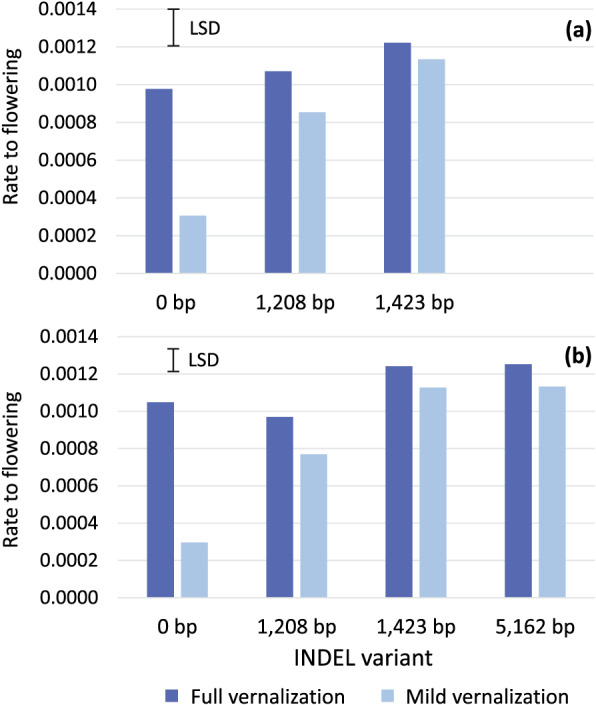
Average rate to flowering (reciprocal of degree‐days to flowering) in mildly and fully vernalizing conditions for narrow‐leafed lupins possessing various insertion/deletions (INDELs) in the 5' regulatory region of LanFTc1 in two phenotyping trials. (a) Trial 1 included narrow‐leafed lupins carrying the 0‐bp deletion (n = 29), 1,208‐bp deletion (n = 1), and 1,423‐bp deletion (n = 9). (b) Trial 2 included narrow‐leafed lupins with the 0‐bp deletion (n = 2), 1,208‐bp deletion (n = 1), 1,423‐bp deletion (n = 8), and 5,162‐bp deletion (n = 6). The least significant difference (LSD) value is provided to compare responses within and between vernalization treatments in each phenotyping trial ([a] LSD = .00020; [b] LSD = .00011)
3.3. Assessing the relationship of INDEL variation in the 5' regulatory region upon LanFTc1 gene expression
On the basis of the association between phenology and vernalization responsiveness with INDEL variation in the 5' regulatory region of LanFTc1, we measured LanFTc1 gene expression in five representative accessions, grown with and without vernalization treatment. Four of the accessions included P27255, P22660, 83A:476, and P29039, representing the 0‐bp (ku), 1,208‐bp, 1,423‐bp (Ku), and 5,162‐bp deletions, respectively. Krasnolistny was also included to gain further evidence implicating the 5,162‐bp deletion in the 5' regulatory region of LanFTc1 as the causal mutation for the Jul locus.
3.3.1. Flowering time is earlier, and vernalization responsiveness is reduced or effectively lost in accessions with a deletion genotype
Orthogonal contrasts revealed that the flowering behaviour of the representative subset was consistent with the larger association study (Figure 2). INDEL genotype by vernalization treatment interactions was highly significant (p < .001) and ranked in the same order as previously. Thus, the wild‐type 0‐bp deletion was much more vernalization responsive than the 1,208‐bp deletion (p diff < .001), which in turn was more responsive than the 1,423‐ and 5,162‐bp INDELs (p diff < .001), which had a similar low response to vernalization (p diff = .883; Table 2). The two accessions with the 5,162‐bp deletion genotype also had a similar low response to vernalization (p diff = .450), although P29039 was always slightly later flowering than Krasnolistny (p diff < .001; Table 2).
Table 2.
Average days and degree‐days to flowering in vernalized and non‐vernalized treatments for narrow‐leafed lupins representing the 5' regulatory region wild‐type sequence (ku) for LanFTc1 and three major deletion variants of 1,208 bp, 1,423 bp (Ku), and 5,162 bp (Jul)
| Days to flowering | Degree‐days to flowering | ||||
|---|---|---|---|---|---|
| Accession | Deletion genotype | Vernalized | Non‐vernalized | Vernalized | Non‐vernalized |
| P27255 | 0 bp (ku) | 53.7 | 134.3 | 1,023.2 | 2,434.8 |
| P22660 | 1,208 | 49.0 | 67.0 | 941.5 | 1,256.5 |
| 83A:476 | 1,423 bp (Ku) | 51.0 | 49.0 | 976.5 | 941.5 |
| P29039 | 5,162 (Jul) | 58.3 | 55.7 | 1,104.8 | 1,058.2 |
| Krasnolistny | 5,162 (Jul) | 52.3 | 50.7 | 999.8 | 970.7 |
Note. Differences between accession means within and across treatments greater than 1.9 days and 33.64 degree‐days are significant (least significant difference p < .05).
3.3.2. LanFTc1 is expressed to varying degrees in accessions with a deletion genotype independently of vernalization treatment
All accessions had high gene expression under vernalizing conditions during vegetative growth, starting from a common high basal level at the 4‐leaf stage (approximately 277 degree‐days, Figure 3a). The deletion size categories (1,208, 1,423, and 5,162 bp) had similar curvilinear increases in relative transcript abundance in the late vegetative stage (approximately 750 degree‐days), whereas the wild‐type (0‐bp deletion) showed a flat slope, with no change in transcript abundance over time (p = .851).
Figure 3.
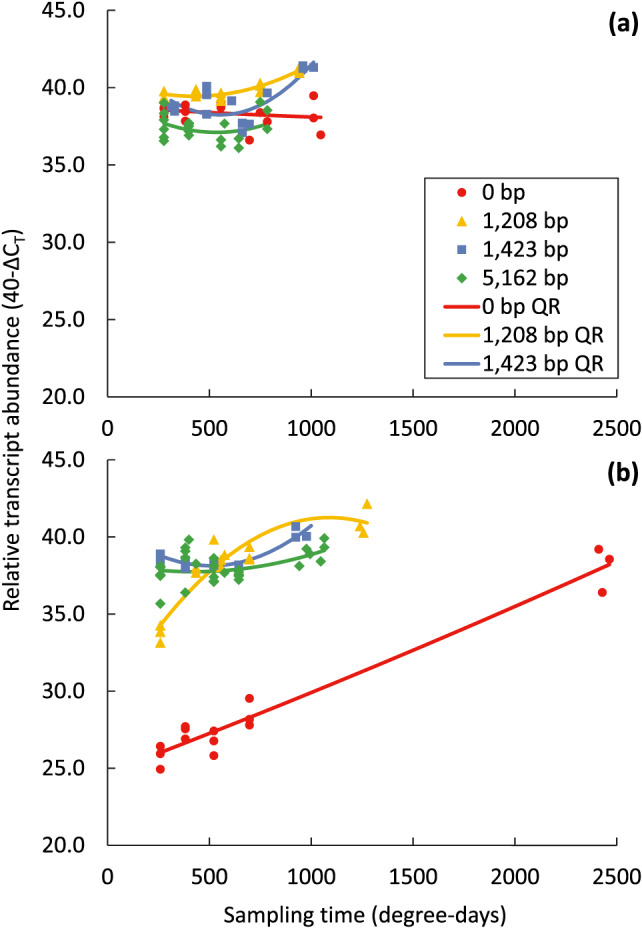
Relative expression of LanFTc1 at various degree‐days from 4‐leaf stage (approximately 277 degree‐days) to first flowering in deletion categories of narrow‐leafed lupin (red circle, 0‐bp deletion wild‐type; yellow triangle, 1,208‐bp deletion; blue square, 1,423‐bp deletion; and green diamond, 5,162‐bp deletion) with (a) and without (b) vernalization. The quadratic regression (QR) model captured 94.7% of variance and indicated significant intercept, linear, and quadratic slope differences (p < .001) between category/vernalization treatment combinations. The final sampling time is at first flower and therefore varies widely between treatment combinations
Relative LanFTc1 expression varied greatly among the accessions in the absence of vernalization (Figure 3b). Those with the 1,423‐ and 5,162‐bp deletions behaved similarly to their respective vernalized treatments, with similar curvilinear increases in transcript abundance in the late vegetative stage. By contrast, relative expression levels in the accession (P22660) with the smaller 1,208‐bp deletion rose rapidly from an intermediate basal level and reached similar levels as the larger deletion categories by the mid‐vegetative stage (Figure 3b). Finally, the wild‐type (0‐bp deletion) had a slow linear increase in relative gene expression throughout the vegetative phase, starting from the lowest level of expression at the 4‐leaf stage, and reached similar levels to the three deletion categories by the onset of flowering (Figure 3b).
3.4. Characterizing the LanFTc1 promoter region
The gene expression profiles of the three deletion variants indicate that sections of the 5' regulatory region are critical for regulating flowering time via the vernalization pathway and LanFTc1. It appears that the 1,423‐ and 5,162‐bp deletions are functionally equivalent, as they both result in insensitivity to vernalization and similar LanFTc1 expression profiles (Figure 3a,b). Therefore, crucial regulatory elements responsible for full gene suppression in the absence of vernalizing conditions should reside within the 1,423‐bp INDEL sequence (Figure 1). The functional activity of the 1,208‐bp INDEL further refines this critical region. As the first 277 bp at the 5' end of the 1,423‐bp INDEL is not also deleted within the 1,208‐bp INDEL, it suggests that this 277‐bp region is critical for establishing complete derepression of LanFTc1 (CH; critical region for high early expression and vernalization insensitivity). Additionally, as the 62‐bp sequence at the 3' end of the 1,208‐bp deletion is conserved in the wild‐type, this indicates that the 1,146‐bp sequence common to the 1,208‐, 1,423‐, and 5,162‐bp INDELs is responsible for establishing a moderate level of early gene activity without vernalization (CM; critical region for moderate early expression and moderate vernalization responsiveness).
We next explored variation in the CH and CM critical regions that may explain differences in vernalization responsiveness and LanFTc1 expression. Both critical regions were screened for candidate transcription factor binding motifs that may have roles in the repression of LanFTc1 within the P27255 representative wild‐type sequence from the comprehensive list compiled by Nelson et al. (2017). A total of 31 individual motifs were found within the CH region, including six in which the motif overhangs the CH region and the adjacent 5' wild‐type sequence and/or contains an alternative SNP allele in one or more vernalization responsive accession(s) (Tables S4 and S5). Meanwhile, as many as 168 individual motifs were identified within the CM region, including seven for which one or more vernalization responsive accession(s) have SNPs (Tables S4 and S5). Among all of the motifs identified upstream of the coding sequence within the 5' regulatory region and 5' untranslated region (UTR), the binding sites for three and five types of transcription factors were unique to the CH and CM regions, respectively, including several reported to have roles in determining flowering time (Table 3). Only a single type of transcription factor, named BRI1‐EMS‐SUPPRESSOR 1 (BES1), was common to both critical regions yet absent in the remainder of the adjacent 5' UTR and 5' regulatory region.
Table 3.
A list of candidate transcription factor (TF) binding site motifs unique to sequences within the 5' regulatory regions critical for establishing moderate (CM) or high (CH) levels of derepressed LanFTc1 expression during early vegetative growth
| Motifs present in the wild‐type sequence | Motifs present in the CH sequence | Motifs present in the CM sequence | Role of TF in flowering within other angiosperms | References |
|---|---|---|---|---|
| AGL9 | 1a | 0 | AGAMOUS‐LIKE 9 (AGL9), also known as SEPALLATA3 (SEP3), is a MADS‐box TF that is involved in establishing identity of petals, stamens, and carpels in Arabidopsis. In rice (Oryza sativa), knock out of two AGL9/SEP3 homologues, OsMADS7 and OsMADS8, also results in delayed flowering. |
Mandel and Yanofsky (1998) Pelaz, Ditta, Baumann, Wisman, and Yanofsky (2000) Cui et al. (2009) |
| ATHB5 | 1 | 0 | ARABIDOPSIS THALIANA HOMEOBOX PROTEIN 5 (ATHB5) is a homeodomain leucine zipper TF that forms a heterodimer with its family member, ARABIDOPSIS THALIANA HOMEOBOX PROTEIN 16, which regulates photoperiodic responsiveness in Arabidopsis. |
Johannesson, Wang, and Engström (2001) Y. Wang et al. (2003) De Smet et al. (2013) |
| PI | 1 | 0 | PISTILLATA (PI) is a MADS‐box TF that is involved in establishing identity of petals and stamens in Arabidopsis. |
Hill and Lord (1989) Bowman, Smyth, and Meyerowitz (1989) |
| AT3G20750 | 0 | 1 | AT3G20750 (also known as GATA29) is a member of the GATA protein family and contains a HAN domain, which has roles in regulating cell differentiation and speciation, including for floral organs, in Arabidopsis. The rice (Oryza sativa) AT3G20750 homologue, NECK LEAF 1 (NL1), has a similar role in floral organ identity, and its overexpression is thought to affect regulation of Hd3a, a rice FT homologue, and delay flowering. |
Reyes, Muro‐Pastor, and Florencio (2004) Zhao et al. (2004) Behringer and Schwechheimer (2015) L. Wang et al. (2009x) Tamaki, Matsuo, Wong, Yokoi, and Shimamoto (2007) |
| C1 | 0 | 1 | C1 is a MYB TF involved in anthocyanin biosynthesis, thus flower colouration, in maize (Zea mays). |
Paz‐Ares, Ghosal, Wienand, Peterson, and Saedler (1987) Sainz, Grotewold, and Chandler (1997) Mola, Grotewold, and Koesa (1998) |
| MADSA | 0 | 1 | MADSA, also known as AGAMOUS‐LIKE 20 and SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1) in Arabidopsis, is a MADS‐box TF activated in the shoot apical meristems during the transition from vegetative to floral development, and which integrates signals from the giberellin pathway, and also the photoperiod pathway through CONSTANS via FT. |
Borner et al. (2000) Yoo et al. (2005) |
| NAC6 | 0 | 1 | NAC6 (also known as NAC2 or AtNAC2) is a member of the NAC TF family, which has roles in regulating morphogenesis and stress responses, and is itself involved in the regulation of stamen development in Arabidopsis. |
Ooka et al. (2003) Mandaokar et al. (2006) |
| WRKY | 0 | 1 | WRKY20 is a member of the WRKY TF family that has roles in plant stress responses and development. Wild soybean (Glycine soja) homologue WRKY20 is abundantly expressed in flowers and floral meristems and is thought to be involved in positive regulation of the autonomous pathway. Its overexpression in Arabidopsis results in early flowering and is associated with up‐regulation of floral integrator genes, including FT and SOC1. | Luo et al. (2013) |
| BES1 | 1 | 2 | BES1 is a member of the BES1/BZR1 TF family and interacts with EARLY FLOWERING 6 and RELATIVE OF EARLY FLOWERING 6 in Arabidopsis to repress the photoperiodic pathway and FLOWERING LOCUS C, a repressor of FT. |
Noh et al. (2004) Yu et al. (2008) |
Note that six of seven nucleotides forming this motif are located within the CH region and that one or more vernalization responsive accessions contain a single nucleotide polymorphism within the motif.
3.5. Assessment of linkage disequilibrium in the LanFTc1 genomic region
To rule out the involvement of other polymorphisms in the LanFTc1 gene region being involved in modifying vernalization responsiveness, we measured the association of each polymorphism with the vernalization responsiveness phenotype. A total of 48 INDELs and 206 SNPs were used for pairwise calculation of linkage disequilibrium (r 2) with the vernalization responsiveness genotype. Low r 2 values of .35 or less were observed for the vast majority of polymorphisms (Figure 4). One INDEL point at 752 bp in the P27255 wild‐type reference sequence, which represents the 1,423‐bp deletion in the 5' regulatory region, was the only variant with an r 2 value of 1.0 and perfect linkage with the vernalization responsiveness phenotype (Figure 4). An r 2 value of 1.0 was also achieved when grouping the 1,423‐, 5,162‐, and 1,208‐bp deletions together as a single multi‐allelic INDEL. The major association between the 1,423‐bp INDEL and vernalization responsiveness phenotype was highly significant (PLINK linear regression, coefficient t statistic = 32.58, Bonferroni adjusted p = 6.60e−27). There was an additional small association with vernalization responsiveness for a 1‐bp INDEL at position 3,215 bp relative to the P27255 wild‐type sequence (PLINK linear regression, Bonferroni adjusted p = .04; Figure 4).
Figure 4.
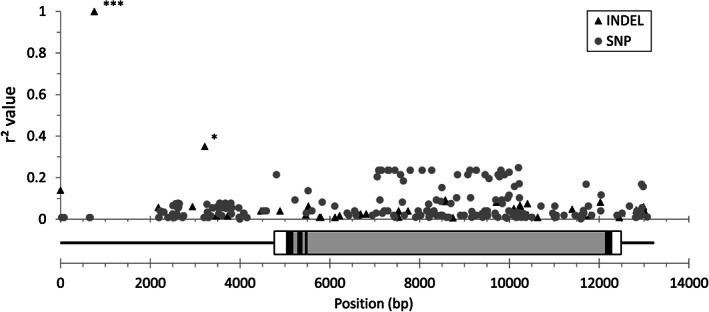
Linkage (represented as r 2) of insertion/deletions (INDELs; black triangles) and single nucleotide polymorphisms (SNPs; grey circles) identified in the narrow‐leafed lupin LanFTc1 wild‐type genomic sequence, represented by Moroccan accession P27255, with vernalization responsiveness. The positions of polymorphisms are indicated relative to the base pair (bp) position along the wild‐type LanFTc1 genomic sequence (GenBank ID KT862491) and a schematic of the LanFTc1 genomic features, including regulatory regions (solid black line), untranslated regions (solid white bar with black border), exons (solid black bar with black border), and introns (solid grey bar with black border). An r 2 value of 1.0 represents perfect linkage with vernalization response phenotype. Asterisks denote significant associations between polymorphisms and the vernalization responsiveness phenotype (Bonferroni adjusted p values: *.01 < p < .05; **.001 < p < .01; ***p < .001)
4. DISCUSSION
4.1. A series of cis‐regulatory variants in a legume FT homologue
A 1,423‐bp deletion in the 5' regulatory region of LanFTc1, an FT homologue, was recently hypothesized as the causal mutation behind the Ku locus that has been significant in establishing narrow‐leafed lupin as a viable pulse crop in Australia and northern Europe (Nelson et al., 2017). Here, we have shown that another two independently occurring mutations, namely, 1,208‐ and 5,162‐bp deletions, overlap the same region of the promoter, creating a series of cis‐regulatory variants that derepress LanFTc1 expression to varying extents. Additionally, we have shown that the 1,423‐bp deletion, plus all three deletions when scored as a singular multi‐allelic variant at the same position, are the only sequence variants in the LanFTc1 wild‐type sequence that are perfectly associated with vernalization response and, as a consequence, early flowering time under non‐vernalizing conditions. The remaining 206 SNPs and 48 small INDELs within the 13‐Kb wild‐type genomic region are not associated with vernalization responsiveness (r 2 ≤ .35; Figure 4), with the exception of one small INDEL (1 bp), which is weakly linked (r 2 = .35; Figure 4). This finding compliments those of Nelson et al. (2017), where presence/absence of the 1,423‐bp deletion was perfectly predictive of vernalization responsiveness in 216 accessions and provides compelling evidence that the INDEL series are the likely causal mutations modifying vernalization responsiveness and, thus, phenology. However, we have not yet ruled out the possibility that other variants outside of this 13‐Kb genomic region are also strongly or perfectly associated with vernalization responsiveness. Lastly, we found strong evidence that the 5,162‐bp deletion corresponds to Jul, supporting previous speculation that these two dominant early flowering time genes discovered independently in Australian and European breeding programs (Rahman & Gladstones, 1972) are in fact different alleles of the same gene, LanFTc1. We are currently developing biparental genetic populations to confirm this conclusion.
To the best of our knowledge, this is the first report of a naturally occurring series of mutations in the noncoding region of a floral integrator gene in any legume species. However, it adds to a growing list of literature similarly reporting series of cis‐regulatory variants of vernalization and photoperiodic pathway genes in Arabidopsis and cereal crops. The largest and most widely published allelic series identified to date involves VRN‐1, a MADS‐box transcription factor that is orthologous to APETALA1 in Arabidopsis (Yan et al., 2003) and that is involved in maintaining down‐regulation of floral repressors following vernalization within members of the Poaceae family (A. Chen & Dubcovsky, 2012). Within the promoter and intronic regions, a staggering number of INDELs ranging in size from 20 to 6,850 bp, in addition to a single SNP, have been identified in the A, B, D, and G genomes of various diploid, tetraploid, and hexaploid wild and domestic wheats and their progenitors (Fu et al., 2005; Golovnina, Kondratenko, Blinov, & Goncharov, 2010; Konopatskaia, Vavilova, Kondratenko, Blinov, & Goncharov, 2016; Milec, Tomková, Sumíková, & Pánková, 2012; Muterko, Balashova, Cockram, Kalendar, & Sivolap, 2015; Santra, Santra, Allan, Campbell, & Kidwell, 2009; Shcherban, Efremova, & Salina, 2012; Takumi, Koyamam, Fujiwara, & Kobayashi, 2011; Yan et al., 2004; Zhang, Gao, Wang, Chen, & Cui, 2015), plus the H genome of barley (Hordeum vulgare L.; Fu et al., 2005). Our study also adds to others in Arabidopsis (Liu et al., 2014; Schwartz et al., 2009), perennial ryegrass (Lolium perenne; Skøt et al., 2011), and wheats and barley (F. Chen et al., 2013; Yan et al., 2006) showing that FT orthologues have similarly been a common target for the evolution of natural flowering time variation in a range of plant families. Lastly, a series of 7‐bp tandem repeat INDELs has also been identified in Arabidopsis to modify the cis‐regulation of CONSTANS (CO), a gene which encodes a zinc‐finger transcription factor responsive to the photoperiodic and circadian clock flowering pathways (Rosas et al., 2014).
4.2. Implications for breeding and expansion of the adaptive range of narrow‐leafed lupin
Discovery of an INDEL series in the promoter region of LanFTc1 has major practical implications in light of its demonstrated capacity to modify vernalization responsiveness and flowering time in narrow‐leafed lupin. The Ku (1,423‐bp deletion) and Jul (5,162‐bp deletion) alleles have already been widely incorporated into domestic breeding programs in Australia and Europe. However, the 1,208‐bp deletion present in the Israeli wild accession, P22660, represents a new form of valuable variation that has the potential to delay flowering time by approximately 2.5 weeks in the absence of vernalization. Such variation would be extremely beneficial in expanding the production range of narrow‐leafed lupin, plus increasing crop adaptation and yield potential in current environments with longer seasons, such as the southern Western Australian and eastern Australian growing regions. The predominance of the Ku and Jul alleles in breeding programs means that, without prior knowledge of the 1,208‐bp INDEL variant, it would not be easily identified in the early stages of segregation from hybrids with breeding lines containing the dominant early alleles, Ku or Jul. The PCR marker designed by Nelson et al. (2017) will serve as a useful resource to screen for the 1,208‐bp INDEL in future breeding (Table S2a).
Similar to the 1,208‐bp deletion identified in wild germplasm from Israel, it is interesting to note that Jul is thought to have originated from the same region of the Middle East (Mikołajczyk, 1966). Evaluation studies of previous germplasm collection trips (Clements & Cowling, 1994; Gladstones & Crosbie, 1979) and a recent genetic and adaptive diversity analysis (Mousavi‐Derazmahalleh et al., 2017) have identified the Eastern Mediterranean and Northern Africa (including but not limited to parts of Morocco, the Middle East, and Aegean islands) as key geographic regions associated with early phenology in the natural habitat of L. angustifolius. The lower elevation and latitude of these regions, in combination with reduced, variable rainfall and increased seasonal temperatures, results in shorter growing seasons with heightened abiotic stresses that drive phenological evolution (Berger, Ludwig, & Buirchell, 2008; Berger, Shrestha, & Ludwig, 2017). Therefore, it is possible that valuable cis‐regulatory variations of LanFTc1 or other genes regulating time to flowering exist in wild populations of narrow‐leafed lupin from these origins. We are currently exploring this possibility in our research activities.
4.3. Understanding the regulation of FT homologues and the mediation of vernalization responsiveness in the legume family
In addition to the potential benefits to narrow‐leafed lupin breeding, the discovery of two new INDEL variants within the 5' regulatory region has also enabled us to explore which part of the promoter is critical for retaining normal repression of LanFTc1 in the absence of vernalization and, which if manipulated, is capable of modifying phenology. This critical region has been further divided into two zones, one of which is critical for establishing a medium level of derepressed gene expression (CM), whereas the second enables high and completely derepressed transcriptional state (CH). However, at this stage, it remains unclear as to why these regions are critical and what role(s) the deleted sequences play in the wild‐type ku allele.
As evidenced from variant series in other species and flowering time genes, cis‐regulatory changes are mediated by polymorphisms in a number of different ways. The classical FT promoter in Arabidopsis contains four major blocks (Adrian et al., 2010; Liu et al., 2014), comprising first, the A block, positioned roughly 400‐bp upstream of the ATG start codon and which contains a number of transcriptional elements, such as those bound by CO (Tiwari et al., 2010); second, the B block, located approximately 1.8‐Kb upstream of the coding sequence and containing two conserved binding sequences for basic helix‐loop‐helix proteins (Adrian et al., 2010); the distal C block, located roughly 5.2‐Kb upstream of the start codon and which contains binding elements for proteins involved in delivering CO to motifs within Block A (Cao et al., 2014); and lastly, an intermittent sequence roughly 3.7‐Kb upstream of the ATG transcription start site that includes a block known as ID, which is involved in establishing physical proximity of the A and C blocks for photoperiod responsiveness. It is the latter in which two INDELs influencing promoter efficiency have evolved in Arabidopsis (Liu et al., 2014). The first of these includes an approximately 1.1‐Kb insertion near Block ID that causes an excessive physical distance between Blocks A and C, preventing their normal interaction. Meanwhile, a smaller deletion of approximately 250 bp contrastingly provides sufficient proximity of Blocks A and C, such that the ID block is redundant. Previous research from Książkiewicz et al. (2016) has indicated a lack of sequence conservation between the 1,423‐bp deletion (Ku) and ID block, therefore suggesting that the INDEL series in LanFTc1 is unlikely to operate in a similar manner to that of the FT series in Arabidopsis. The discovery of the 5,162‐bp deletion in Jul accessions in this study also suggests that this is not the case, as a significantly large proportion of the promoter that may correspond to other blocks has been deleted. However, further research to characterize the sequences either side of the 5,162‐bp INDEL may be required to firmly eliminate the improvement of promoter efficiency by modification to regulatory element proximity as one possible consequence of the INDEL series in LanFTc1.
Alternative ways in which the INDEL series may instead modify cis‐regulation of LanFTc1 is through changing the profile of transcription factor binding sites within the promoter region or their capacity to be bound. A straightforward explanation is the complete removal of transcription factor binding site motifs from within the three deletions. We refined a list of candidate transcription factor motifs from Nelson et al. (2017), revealing a total of 168 and 31 individual motifs present in the wild‐type promoter sequence yet which are absent in the CM and CH regions, respectively (Tables S4 and S5). However, this is still an extremely large number of candidate transcription factor motifs, and it will be very difficult to further resolve which may or may not have functional roles in LanFTc1 regulation, especially if no further variants are found overlapping this region.
Copy number of transcription factor binding sites also represents another possibility of cis‐regulatory modification. As has been demonstrated in Arabidopsis, increasing from three to four tandem repeats of a 7‐bp motif for CYCLING DOF FACTOR 1 in the promoter of CO increases the day‐time repression of this gene and significantly delays flowering time (Rosas et al., 2014). We identified motifs for the binding site of a single transcription factor, named BES1, present once within the CH and twice within the CM critical regions (Tables 2, S4, and S5), yet nowhere else in the 5' UTR and 5' regulatory region of LanFTc1. In Arabidopsis, BES1 interacts with EARLY FLOWERING 6 and RELATIVE OF EARLY FLOWERING 6 proteins to respectively repress the photoperiodic pathway, through unknown means, and FLOWERING LOCUS C, a repressor of FT that is itself repressed by vernalization (Noh et al., 2004; Yu et al., 2008). Therefore, although both FLOWERING LOCUS C and RELATIVE OF EARLY FLOWERING 6 are apparently absent from the narrow‐leafed lupin genome (Hane et al., 2017), there is precedence for BES1 involvement in the regulation of flowering time, and it is conceivable that it could be involved in the direct regulation of LanFTc1 through partnership with other flowering time genes. In such a scenario, deletion of two copies of the BES1 binding site motif via the 1,208‐bp INDEL genotype would be sufficient to elevate LanFTc1 expression to an intermediate level, and deletion of all three motifs via the 1,423‐ or 5,162‐bp INDELs would fully derepress expression. With genome editing tools, such as the CRISPR/Cas‐9 system (Bortesi & Fischer, 2015), and more efficient transformation protocols in narrow‐leafed lupin (Barker et al., 2016), it may be feasible to modify BES1 binding site motifs in the wild‐type sequence to test the validity of this hypothesis in the future. If BES1 has a role in regulating LanFTc1, BES1's known involvement within the photoperiodic pathway could explain why cultivars with the 1,423‐bp deletion are also less responsive to inductive long days than wild‐types without the large deletion in the promoter region of LanFTc1 (J. D. Berger, unpublished data).
Lastly, the location of INDELs relative to transcription factor binding site motifs can influence the affinity for transcription factor binding, as demonstrated in the case of the FT homologue allelic series in perennial ryegrass (Skøt et al., 2011). Relative to the wild‐type sequence designated as the C haplotype, a deletion of five nucleotides positioned 7‐ to 11‐bp downstream of a conserved motif (the A haplotype) resulted in a 2‐day delay in flowering time, whereas a six nucleotide deletion positioned 1 to 6 bp directly 3' of the conserved motif (the B haplotype) resulted in a 7‐day flowering time delay. Here, we have identified three classes of transcription factor binding site motifs, which are disrupted by the 5' end of the CH region and which are completely absent in the CM region. However, motifs for these same transcription factors are also found on several occasions elsewhere within the LanFTc1 genomic region, and small INDEL polymorphisms present in vernalization responsive accessions without the 1,208‐, 1,423‐, or 5,162‐bp INDELs can also be found disrupting some of these motifs. Therefore, it seems unlikely that any of these motifs are functional or critical to LanFTc1; however, further research will be required to better characterize other motifs adjacent to the large deletions.
Despite our lack of knowledge as to how they impact gene expression, the discovery of the INDEL variant series in the 5' regulatory region of LanFTc1 has provided us with a rare opportunity to better explore possible ways in which FT homologues are regulated, and vernalization responsiveness is mediated, at the molecular level outside of the Brassicaceae and Poaceae. At present, our greatest understanding concerning vernalization response within the legume family comes from Medicago truncatula (Weller & Ortega, 2015). In this model species, an FTa1 homologue is up‐regulated following the return of warm conditions post‐vernalization, and loss‐of‐function mutations within the coding sequence render plants insensitive to vernalization (Laurie et al., 2011). Three induced mutant lines with dominant early, vernalization‐insensitive flowering have been shown to contain transposon insertions in the large third intron or the 3' regulatory region that result in up‐regulated expression of FTa1, suggesting that these genomic regions are important sites for conferring transcriptional repression in the wild‐type (Jaudal et al., 2013). However, similar to the present story in narrow‐leafed lupin, it is unknown what elements within these regions are important for the vernalization pathway and FTa1 transcription. Thus far, it appears that methylation in the FTa1 genomic region is unlikely to play a role, with no differences observed between the mutants with induced transposon insertions and wild‐type plants (Jaudal et al., 2013). The discovery of further INDEL variants in LanFTc1 could provide further clues of how signalling mediated through the vernalization pathway is centred on FT homologues at the molecular level in the Fabaceae.
Supporting information
Figure S1. PCR markers to assay four major INDEL variants (0 bp, 1,423 bp, 1,208 bp and 5,162 bp) in the promoter region of LanFTc1, a FLOWERING LOCUS T homologue of narrow‐leafed lupin.
Table S1. Narrow‐leafed lupin (Lupinus angustifolius L.) germplasm genotyped for polymorphisms in the genomic region (approximately 7 Kb upstream and 2 Kb downstream of the coding sequence) of LanFTc1, a vernalisation responsive FT homologue.
Table S2. a) Primers, b) PCR reaction, and c) cycling conditions used for PCR marker genotyping of the INDEL in the 5' regulatory region of LanFTc1, an FT homologue in narrow‐leafed lupin (Lupinus angustifolius L.).
Table S3a. A list of SNP (single nucleotide polymorphism) and INDEL (insertion/deletion) polymorphisms in 43 accessions of narrow‐leafed lupin (Lupinus angustifolius L.) in the LanFTc1 genomic region (encompassing 7 kb upstream and 2 kb downsteam of the gene) relative to the Tanjil reference genome (Hane et al. 2017).
Table S3b. A list of polymorphisms in 42 accessions of narrow‐leafed lupin (Lupinus angustifolius L.) within the 1,423 bp sequence of the 5' regulatory region of LanFTc1, a FLOWERING LOCUS T homologue, which is deleted in the Tanjil reference genome (Hane et al. 2017).
Table S5. Transcription factor motifs identified in the wild‐type sequence (ku; represented by accession P27255) of LanFTc1, the vernalisation responsive FT homologue in narrow‐leafed lupin (Lupinus angustifolius L.).
Table S6. A list of candidate transcription factor binding site motifs for sequences within the 5’ regulatory region and 5' UTR of LanFTc1 that are critical for establishing moderate (CM) or high (CH) levels of de‐repressed LanFTc1 expression during early vegetative growth.
ACKNOWLEDGMENTS
This research was generously funded by Grants DAW00238 and UWA00147 from the Grains Research and Development Corporation (GRDC), Australia and funding from The University of Western Australia. Our thanks go to Michał Książkiewicz (Polish Academy of Sciences) for the kind provision of seed for a number of Russian and Polish cultivars used in this study.
Taylor CM, Kamphuis LG, Zhang W, et al. INDEL variation in the regulatory region of the major flowering time gene LanFTc1 is associated with vernalization response and flowering time in narrow‐leafed lupin (Lupinus angustifolius L.). Plant Cell Environ. 2019;42:174–187. 10.1111/pce.13320
REFERENCES
- Adrian, J. , Farrona, S. , Reimer, J. , Albani, M. , Coupland, G. , & Turck, F. (2010). cis‐regualtory elements and chromatin state coordinately control temporal and spatial expression of FLOWERING LOCUS T in Arabidopsis . The Plant Cell, 22, 1425–1440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bari, R. , Pant, B. D. , Stint, M. , & Scheible, W.‐R. (2006). PHO2, MicroRNA399, and PHR1 define a phoshate‐signaling pathway in plants. Plant Physiology, 141, 988–999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barker, S. J. , Si, P. , Hodgson, L. , Ferguson‐Hunt, M. , Khentry, Y. , Krishnamurthy, P. , … Erskine, W. (2016). Regeneration selection improves transformation efficiency in narrow‐leaf lupin. Plant Cell Tissue and Organ Culture, 126, 219–228. [Google Scholar]
- Behringer, C. , & Schwechheimer, C. (2015). B‐GATA transcription factors—Insights into their structure, regulation, and role in plant development. Frontiers in Plant Science, 6, 90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berger, J. , Buirchell, B. , Palta, J. , Luckett, D. , Ludwig, C. , & Shrestha, D. (2008). G x E analysis of narrow‐leafed lupin historical trials indicates little specific adaptation among Australian cultivars In Palta J. A., & Berger J. D. (Eds.), Lupins for Health and Wealth (pp. 317–320. Proceedings of the 12th International Lupin Conference, 14‐18 September 2008, Fremantle, Western Australia). Canterbury, New Zealand: International Lupin Association. [Google Scholar]
- Berger, J. , Ludwig, C. , & Buirchell, B. (2008). Ecogeography of the Old World lupins: Characterising the habitat range In Palta J. A., & Berger J. D. (Eds.), Lupins for Health and Wealth (pp. 355–361. Proceedings of the 12th International Lupin Conference, 14‐18 September 2008, Fremantle, Western Australia). Canterbury, New Zealand: International Lupin Association. [Google Scholar]
- Berger, J. , Shrestha, D. , & Ludwig, C. (2017). Reproductive strategies in Mediterranean legumes: Trade‐offs between phenology, seed size and vigor within and between wild and domesticated Lupinus species collected along aridity gradients. Frontiers in Plant Science, 8, 548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berger, J. D. , Buirchell, B. J. , Luckett, D. J. , & Nelson, M. N. (2012). Domestication bottlenecks limit genetic diversity and constrain adaptation in narrow‐leafed lupin (Lupinus angustifolius L.). Theoretical and Applied Genetics, 124, 637–652. [DOI] [PubMed] [Google Scholar]
- Berger, J. D. , Buirchell, B. J. , Luckett, D. J. , Palta, J. A. , Ludwig, C. , & Liu, D. L. (2012). How has narrow‐leafed lupin changed in its 1st 40 years as an industrial, broad‐acre crop? A G×E‐based characterization of yield‐related traits in Australian cultivars. Field Crops Research, 126, 152–164. [Google Scholar]
- Boersma, J. G. , Buirchell, B. J. , Sivasithamparam, K. , & Yang, H. (2007). Development of a sequence‐specific PCR marker linked to the Ku gene which removes the vernalization requirement in narrow‐leafed lupin. Plant Breeding, 126, 306–309. [Google Scholar]
- Boersma, J. G. , Pallotta, M. , Li, C. , Buirchell, B. J. , Sivasithamparam, K. , & Yang, H. (2005). Construction of a genetic linkage map using MFLP and identification of molecular markers linked to domestication genes in narrow‐leafed lupin (Lupinus angustifolius L.). Cellular and Molecular Biology Letters, 10, 331–344. [PubMed] [Google Scholar]
- Borner, R. , Kampmann, G. , Chandler, J. , Gleißner, R. , Wisman, E. , Apel, K. , & Melzer, S. (2000). A MADS domain gene involved in the transition to flowering in Arabidopsis . The Plant Journal, 24, 591–599. [DOI] [PubMed] [Google Scholar]
- Bortesi, L. , & Fischer, R. (2015). The CRISPR/Cas9 system for plant genome editing and beyond. Biotechnology Advances, 33, 41–52. [DOI] [PubMed] [Google Scholar]
- Bowman, J. L. , Smyth, D. R. , & Meyerowitz, E. M. (1989). Genes directing flower development in Arabidopsis . The Plant Cell, 1, 37–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao, S. , Kumimoto, R. , Gnesutta, N. , Calogero, A. , Mantovani, R. , & Holt, B. III (2014). A distal CCAAT/NUCLEAR FACTOR Y complex promotes chromatin looping at the FLOWERING LOCUS T promoter and regulates the timing of flowering in Arabidopsis . The Plant Cell, 26, 1009–1017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, A. , & Dubcovsky, J. (2012). Wheat TILLING mutants show that the vernalization gene VRN1 down‐regulates the flowering repressor VRN2 in leaves but is not essential for flowering. PLoS Genetics, 8, e1003134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, C. , Fletcher, A. , Lawes, R. , Berger, J. , & Roberston, M. (2017). Modelling phenological and agronomic adaptation options for narrow‐leafed lupins in the southern grainbelt of Western Australia. European Journal of Agronomy, 89, 140–147. [Google Scholar]
- Chen, F. , Gao, M. , Zhang, J. , Zuo, A. , Shang, X. , & Cui, D. (2013). Molecular characterization of vernalization and response genes in bread wheat from the Yellow and Huai Valley of China. BMC Plant Biology, 13, 199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clements, J. , & Cowling, W. (1994). Patterns of morphological diversity in relation to geographical origins of wild Lupinus angustifolius from the Aegean region. Genetic Resources and Crop Evolution, 41, 109–122. [Google Scholar]
- Cowling, W. A. (1999). Pedigrees and characteristics of narrow‐leafed lupin cultivars released in Australia from 1967 to 1998. Bulletin 4365. Agriculture Western Australia, Perth, Australia.
- Cui, R. , Han, J. , Zhao, S. , Su, K. , Wu, F. , Du, X. , … Meng, Z. (2009). Functional conservation and diversification of class E floral homeotic genes in rice (Oryza sativa). The Plant Journal, 61, 767–781. [DOI] [PubMed] [Google Scholar]
- De Smet, I. , Lau, S. , Erismann, J. S. , Axiotis, I. , Kolb, M. , Kientz, M. , … Jürgens, G. (2013). Transcriptional repression of BODENLOS by HD‐ZIP transcription factor HB5 in Arabidopsis thaliana . Journal of Experimental Botany, 64, 3009–3019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FAO (2014) FAOSTAT. Food and Agriculture Organization of the United Nations, Rome, Italy.
- Foyer, C. H. , Lam, H.‐M. , Nguyen, H. T. , Siddique, K. H. M. , Varshney, R. K. , Colmer, T. D. , … Considine, M. J. (2016). Neglecting legumes has compromised human health and sustainable food production. Nature Plants, 2, 16112. [DOI] [PubMed] [Google Scholar]
- Fu, D. , Szűcs, P. , Yan, L. , Helguera, M. , Skinner, J. S. , von Zitzewitz, J. , … Dubcovsky, J. (2005). Large deletions within the first intron in VRN‐1 are associated with spring growth habit in barley and wheat. Molecular Genetics and Genomics, 273, 54–65. [DOI] [PubMed] [Google Scholar]
- Gladstones, J. S. (1970). Lupins as crop plants. Field Crop Abstracts, 23, 123–148. [Google Scholar]
- Gladstones, J. S. (1974). Lupins of the Mediterranean region and Africa, Western Australian Department of Agriculture, Technical Bulletin No. 26.
- Gladstones, J. S. (1977). The narrow‐leafed lupin in Western Australia. Western Australia Department of Agriculture, Bulletin 3990.
- Gladstones, J. S. , & Crosbie, G. B. (1979). Lupin wild types introduced into Western Australia to 1973. Department of Agriculture, Western Australia, Technical Bulletin No. 43.
- Gladstones, J. S. , & Hill, G. D. (1969). Selection for economic characters in Lupinus angustifolius and L. digitatus. 2. Time of flowering. Australian Journal of Experimental Agriculture and Animal Husbandry, 9, 213–220. [Google Scholar]
- Golovnina, K. , Kondratenko, E. , Blinov, A. , & Goncharov, N. (2010). Molecular characterization of vernalization loci VRN1 in wild and cultivated wheats. BMC Plant Biology, 10, 168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hane, J. K. , Ming, Y. , Kamphuis, L. G. , Nelson, M. N. , Garg, G. , Atkins, C. A. , … Singh, K. B. (2017). A comprehensive draft genome sequence for lupin (Lupinus angustifolius), an emerging health food: Insights into plant‐microbe interactions and legume evolution. Plant Biotechnology Journal, 15, 318–330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Higo, K. , Ugawa, Y. , Iwamoto, M. , & Korenaga, T. (1999). Plant cis‐acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Research, 27, 297–300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill, J. P. , & Lord, E. M. (1989). Floral development in Arabidopsis thaliana: A comparison of the wild type and the homeotic pistillata mutant. Canadian Journal of Botany, 67, 2922–2936. [Google Scholar]
- Jaudal, M. , Yeoh, C. C. , Zhang, L. , Stockum, C. , Mysore, K. S. , Ratet, P. , & Putterill, J. (2013). Retroelement insertions at the Medicago FTa1 locus in spring mutants eliminate vernalisation but not long‐day requirements for early flowering. The Plant Journal, 76, 580–591. [DOI] [PubMed] [Google Scholar]
- Johannesson, H. , Wang, Y. , & Engström, P. (2001). DNA‐binding and dimerization preferences of Arabidopsis homeodomain‐leucine zipper transcription factors in vitro . Plant Molecular Biology, 45, 63–73. [DOI] [PubMed] [Google Scholar]
- Kamphuis, L. G. , Hane, J. K. , Nelson, M. N. , Gao, L. , Atkins, C. A. , & Singh, K. B. (2015). Transcriptome sequencing of different narrow‐leafed lupin tissue types provides a comprehensive uni‐gene assembly and extensive gene‐based molecular markers. Plant Biotechnology Journal, 13, 14–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Konopatskaia, I. , Vavilova, V. , Kondratenko, E. , Blinov, A. , & Goncharov, N. (2016). VRN1 genes variability in tetraploid wheat species with a spring growth habit. BMC Plant Biology, 16, 244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kouris‐Blazos, A. , & Belski, R. (2016). Health benefits of legumes and pulses with a focus on Australian sweet lupins. Asia Pacific Journal of Clinical Nutrition, 25, 1–17. [DOI] [PubMed] [Google Scholar]
- Kroc, M. , Koczyk, G. , Święcicki, W. , Kilian, A. , & Nelson, M. N. (2014). New evidence of ancestral polyploidy in the Genistoid legume Lupinus angustifolius L. (narrow‐leafed lupin). Theoretical and Applied Genetics, 127, 1237–1249. [DOI] [PubMed] [Google Scholar]
- Książkiewicz, M. , Rychel, S. , Nelson, M. , Wyrwa, K. , Naganowska, B. , & Wolko, B. (2016). Expansion of the phosphatidylethanolamine binding protein family in legumes: A case study of Lupinus angustifolius L. FLOWERING LOCUS T homologs, LanFTc1 and LanFTc2 . BMC Genomics, 17, 820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubok, I. (1988). The history of lupine breeding in Poland. Plant Breeding and Acclimatization Institute, Radzików, Poland.
- Lambers, H. , Clements, J. C. , & Nelson, M. N. (2013). How a phosphorus‐acquisition strategy based on carboxylate exudation powers the success and agronomic potential of lupines (Lupinus, Fabaceae). American Journal of Botany, 100, 263–288. [DOI] [PubMed] [Google Scholar]
- Langmead, B. , & Salzberg, S. L. (2012). Fast gapped‐read alignment with Bowtie 2. Nature Methods, 9, 357–359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laurie, R. E. , Diwadkar, P. , Jaudal, M. , Zhang, L. , Hecht, V. , Wen, J. , … Macknight, R. C. (2011). The Medicago FLOWERING LOCUS T homolog, MtFTa1, is a key regulator of flowering time. Plant Physiology, 156, 2207–2224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, H. (2011). A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics, 27, 2987–2993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, H. , Handsaker, B. , Wysoker, A. , Fennell, T. , Ruan, J. , Homer, N. , … 1000 Genome Project Data Processing Subgroup . (2009). The sequence alignment/map format and SAMtools. Bioinformatics, 25, 2078–2079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lima‐Cabello, E. , Alche, V. , Foley, R. , Andrikopoulos, S. , Morahan, G. , Singh, K. , … Jimenez‐Lopez, J. (2017). Narrow‐leafed lupin (Lupinus angustifolius L.) β‐conglutin proteins modulate the insulin signaling pathway as potential type 2 diabetes treatment and inflammatory‐related disease amelioration. Molecular Nutrition & Food Research, 61, 1600819. [DOI] [PubMed] [Google Scholar]
- Liu, L. , Adrian, J. , Pankin, A. , Hu, J. , Dong, X. , von Korff, M. , & Turck, F. (2014). Induced and natural variation of promoter length modulates the photoperiodic response of FLOWERING LOCUS T . Nature Communications, 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo, X. , Sun, X. , Liu, B. , Zhu, D. , Bai, X. , Cai, H. , … Zhu, Y. (2013). Ectopic expression of a WRKY homolog from Glycine soja alters flowering time in Arabidopsis . PLoS ONE, 8, e73295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mandaokar, A. , Thines, B. , Shin, B. , Lange, B. M. , Choi, G. , Koo, Y. J. , … Browse, J. (2006). Transcriptional regulators of stamen development in Arabidopsis identified by transcriptional profiling. The Plant Journal, 46, 984–1008. [DOI] [PubMed] [Google Scholar]
- Mandel, M. A. , & Yanofsky, M. F. (1998). The Arabidopsis AGL9 MADS box gene is expressed in young flower primordia. Sexual Plant Reproduction, 11, 22–28. [Google Scholar]
- Mathelier, A. , Zhao, X. , Zhang, A. W. , Parcy, F. , Worsley‐Hunt, R. , Arenillas, D. J. , … Wasserman, W. W. (2014). JASPAR 2014: An extensively expanded and update open‐access database of transcription factor binding profiles. Nucleic Acids Research, 42, D1423–D1147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mikołajczyk, J. (1966). Genetic studies in Lupinus angustifolius. Part. III. Inheritance of the alkaloid content, seed hardness and length of the growing season in blue lupin. Genetica Polonica, 7, 181–196. [Google Scholar]
- Milec, Z. , Tomková, L. , Sumíková, T. , & Pánková, K. (2012). A new multiplex PCR test for the determination of Vrn‐B1 alleles in bread wheat (Triticum aestivum L.). Molecular Breeding, 30, 317–323. [Google Scholar]
- Mola, J. , Grotewold, E. , & Koesa, R. (1998). How genes paint flowers and seeds. Trends in Plant Science, 3, 212–217. [Google Scholar]
- Mousavi‐Derazmahalleh, M. , Bayer, P. , Nevado, B. , Hurgobin, B. , Filatov, D. , Kilian, A. , … Nelson, M. (2017). Exploring the genetic and adaptive diversity of a pan‐Mediterranean crop wild relative: Narrow‐leafed lupin. Theoretical and Applied Genetics, 10.1007/s00122-017-3045-7 In press, 131, 887–901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muterko, A. , Balashova, I. , Cockram, J. , Kalendar, R. , & Sivolap, Y. (2015). The new wheat vernalization repsonse allele Vrn‐D1s is caused by DNA transposon insertion in the first intron. Plant Molecular Biology Reporter, 33, 294–303. [Google Scholar]
- Nelson, M. N. , Berger, J. D. , & Erskine, W. (2010). Flowering time control in annual legumes: Prospects in a changing global climate. CAB Reviews: Perspectives in Agriculture, Veterinary Science, Nutrition and Natural Resources, 5, 017. [Google Scholar]
- Nelson, M. N. , Książkiewicz, M. , Rychel, S. , Besharat, N. , Taylor, C. M. , Wyrwa, K. , … Wolko, B. (2017). The loss of vernalization requirement essential to domestication in narrow‐leafed lupin is associated with a deletion in the promoter and de‐repressed expression of an FT homologue. New Phytologist, 213, 220–232. [DOI] [PubMed] [Google Scholar]
- Nelson, M. N. , Moolhuijzen, P. M. , Boersma, J. G. , Chudy, M. , Lesniewska, K. , Bellgard, M. , … Ellwood, S. R. (2010). Aligning a new reference genetic map of Lupinus angustifolius with the genome sequence of the model legume, Lotus japonicus . DNA Research, 17, 73–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson, M. N. , Phan, H. T. T. , Ellwood, S. R. , Moolhuijzen, P. M. , Hane, J. , Williams, A. , … Cowling, W. A. (2006). The first gene‐based map of Lupinus angustifolius L.‐location of domestication genes and conserved synteny with Medicago truncatula . Theoretical and Applied Genetics, 113, 225–238. [DOI] [PubMed] [Google Scholar]
- Noh, B. , Lee, S.‐H. , Kim, H.‐J. , Yi, G. , Shin, E.‐A. , Lee, M. , … Noh, Y.‐S. (2004). Divergent roles of a pair of homologous jumonji/zinc‐finger‐class transcription factor proteins in the regulation of Arabidopsis flowering time. The Plant Cell, 16, 2601–2613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ooka, H. , Satoh, K. , Doi, K. , Nagata, T. , Otomo, Y. , Murakami, K. , … Kikuchi, S. (2003). Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana . DNA Research, 10, 239–247. [DOI] [PubMed] [Google Scholar]
- Paz‐Ares, J. , Ghosal, D. , Wienand, U. , Peterson, P. A. , & Saedler, H. (1987). The regulatory c1 locus of Zea mays encodes a protein with homology to myb proto‐oncogene products and with structural similarities to transcriptional activators. The EMBO Journal, 6, 3553–3558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelaz, S. , Ditta, G. S. , Baumann, E. , Wisman, E. , & Yanofsky, M. F. (2000). B and C floral organ identity functions require SEPALLATA MADS‐box genes. Nature, 405, 200–203. [DOI] [PubMed] [Google Scholar]
- Purcell, S. , Neale, B. , Todd‐Brown, K. , Thomas, L. , Ferreira, M. A. R. , Bender, D. , … Sham, P. C. (2007). PLINK: A tool set for whole‐genome association and population‐based linkage analyses. The American Journal of Human Genetics, 81, 559–575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rahman, M. S. , & Gladstones, J. S. (1972). Control of lupin initiation by vernalization, photoperiod and temperature under controlled envrionment. Australian Journal of Experimental Agriculture and Animal Husbandry, 12, 638–645. [Google Scholar]
- Reyes, J. C. , Muro‐Pastor, M. I. , & Florencio, F. J. (2004). The GATA family of transcription factors in Arabdiopsis and rice. Plant Physiology, 134, 1718–1732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosas, U. , Mei, Y. , Xie, Q. , Banta, J. , Zhou, R. , Seufferheld, G. , … Purugganan, M. (2014). Variation in Arabidopsis flowering time associated with cis‐regulatory variation in CONSTANS . Nature Communications, 5, 3651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sainz, M. B. , Grotewold, E. , & Chandler, V. L. (1997). Evidence for direct activation of an anthocyanin promoter by the maize C1 protein and comparison of DNA binding by related Myb domain proteins. The Plant Cell, 9, 611–625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santra, D. , Santra, M. , Allan, R. , Campbell, K. , & Kidwell, K. (2009). Genetic and molecular characterization of vernalization genes Vrn‐A1, Vrn‐B1, and Vrn‐D1 in spring wheat germplasm from the Pacific Northwest region of the U.S.A. Plant Breeding, 28, 576–584. [Google Scholar]
- Schwartz, C. , Balasubramanian, S. , Warthmann, N. , Michael, T. , Lempe, J. , Sureshkumar, S. , … Weigel, D. (2009). Cis‐regulatory changes at FLOWERING LOCUS T mediate natural variation in flowering responses in Arabidopsis thaliana . Genetics, 183, 723–732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seymour, M. , Kirkegaard, J. , Peoples, M. , White, P. , & French, R. (2012). Break‐crop benefits to wheat in Western Australia—Insights from over three decades of research. Crop & Pasture Science, 63(1), 16. [Google Scholar]
- Shcherban, A. , Efremova, T. , & Salina, E. (2012). Identification of a new Vrn‐B1 allele using two near‐isogenic wheat lines with difference in heading time. Molecular Breeding, 29, 675–685. [Google Scholar]
- Skøt, L. , Sanderson, R. , Thomas, A. , Skøt, K. , Thorogood, D. , Latypova, G. , … Armstead, I. (2011). Allelic variation in the perennial ryegrass FLOWERING LOCuS T gene is associated with changes in flowering time across a range of populations. Plant Physiology, 155, 1013–1022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stefanova, K. T. , & Buirchell, B. (2010). Multiplicative mixed models for genetic gain assessment in lupin breeding. Crop Science, 50, 880–891. [Google Scholar]
- Takumi, S. , Koyamam, K. , Fujiwara, K. , & Kobayashi, F. (2011). Identification of a large deletion in the first intron of the VRN‐D1 locus, associated with loss of vernalization requirement in wild wheat progenitor Aegilops tauschii Coss. Genes & Genetic Systems, 86, 183–195. [DOI] [PubMed] [Google Scholar]
- Tamaki, S. , Matsuo, S. , Wong, H. L. , Yokoi, S. , & Shimamoto, K. (2007). Hd3a protein is a mobile flowering signal in rice. Science, 316, 1033–1036. [DOI] [PubMed] [Google Scholar]
- Taylor, C. , Jost, R. , Erskine, W. , & Nelson, M. (2016). Identifying stable reference genes for qRT‐PCR normalisation in gene expression studies of narrow‐leafed lupin (Lupinus angustifolius L.). PLoS ONE, 11, e0148300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tiwari, S. , Shen, Y. , Chang, H.‐C. , Hou, Y. , Harris, A. , Fong Ma, S. , … Ratcliffe, O. (2010). The flowering time regulator CONSTANS is recruited to the FLOWERING LOCUS T promoter via a unique cis‐element. New Phytologist, 187, 57–66. [DOI] [PubMed] [Google Scholar]
- Turck, F. , Fornara, F. , & Coupland, G. (2008). Regulation and identity of florigen: FLOWERING LOCUS T moves center stage. Annual Review of Plant Biology, 59, 573–594. [DOI] [PubMed] [Google Scholar]
- Wang, L. , Yin, H. , Qian, Q. , Yang, J. , Huang, C. , Hu, X. , & Luo, D. (2009). NECK LEAF 1, a GATA type transcription factor, modulates organogenesis by regulating the expression of multiple regulatory genes during reproductive development in rice. Cell Research, 19, 598–611. [DOI] [PubMed] [Google Scholar]
- Wang, Y. , Henriksson, E. , Söderman, E. , Nordin Henriksson, K. , Sundberg, E. , & Engström, P. (2003). The Arabidopsis homeobox gene, ATHB16, regulates leaf development and the sensitivity to photoperiod in Arabdiopsis. Developmental Biology, 264, 228–239. [DOI] [PubMed] [Google Scholar]
- Weller, J. L. , & Ortega, R. (2015). Genetic control of flowering time in legumes. Frontiers in Plant Science, 6, 207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan, L. , Fu, D. , Li, C. , Blechl, A. , Tranquilli, G. , Bonafede, M. , … Dubcovsky, J. (2006). The wheat and barley vernalization gene VRN3 is an orthologue of FT . Proceedings of the National Academy of Sciences, USA, 103, 19581–19586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan, L. , Helguera, M. , Kato, K. , Fukuyama, S. , Sherman, J. , & Dubcovsky, J. (2004). Allelic variation at the VRN‐1 promoter region in polyploid what. Theoretical and Applied Genetics, 109, 1677–1686. [DOI] [PubMed] [Google Scholar]
- Yan, L. , Loukoianov, A. , Tranquilli, G. , Helguera, M. , Fahima, T. , & Dubcovsky, J. (2003). Positional cloning of the wehat vernalization gene VRN1 . Proceedings of the National Academy of Sciences, USA, 100, 6263–6268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoo, S. K. , Chung, K. S. , Kim, J. , Lee, J. H. , Hong, S. M. , Yoo, S. J. , … Ahn, J. H. (2005). CONSTANS activates SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 through FLOWERING LOCUS T to promote flowering in Arabidopsis. Plant Physiology, 139, 770–778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu, X. , Li, L. , Li, L. , Guo, M. , Chory, J. , & Yin, Y. (2008). Modulation of brassinosteroid‐regulated gene expression by jumonji domain‐containing proteins ELF6 and REF6 in Arabidopsis . Proceedings of the National Academy of Sciences, USA, 105, 7618–7623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, Z. , Gao, M. , Wang, S. , Chen, F. , & Cui, D. (2015). Allelic variation at the vernalization and photoperiod sensitivity loci in Chinese winter wheat cultivars (Triticum aestivum L.). Frontiers in Plant Science, 6, 470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao, Y. , Medrano, L. , Ohashi, K. , Fletcher, J. C. , Yu, H. , Sakai, H. , & Meyerowitz, E. M. (2004). HANABA TARANU is a GATA transcription factor that regulates shoot apical meristem and flower development in Arabdiopsis. The Plant Cell, 16, 2586–2600. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. PCR markers to assay four major INDEL variants (0 bp, 1,423 bp, 1,208 bp and 5,162 bp) in the promoter region of LanFTc1, a FLOWERING LOCUS T homologue of narrow‐leafed lupin.
Table S1. Narrow‐leafed lupin (Lupinus angustifolius L.) germplasm genotyped for polymorphisms in the genomic region (approximately 7 Kb upstream and 2 Kb downstream of the coding sequence) of LanFTc1, a vernalisation responsive FT homologue.
Table S2. a) Primers, b) PCR reaction, and c) cycling conditions used for PCR marker genotyping of the INDEL in the 5' regulatory region of LanFTc1, an FT homologue in narrow‐leafed lupin (Lupinus angustifolius L.).
Table S3a. A list of SNP (single nucleotide polymorphism) and INDEL (insertion/deletion) polymorphisms in 43 accessions of narrow‐leafed lupin (Lupinus angustifolius L.) in the LanFTc1 genomic region (encompassing 7 kb upstream and 2 kb downsteam of the gene) relative to the Tanjil reference genome (Hane et al. 2017).
Table S3b. A list of polymorphisms in 42 accessions of narrow‐leafed lupin (Lupinus angustifolius L.) within the 1,423 bp sequence of the 5' regulatory region of LanFTc1, a FLOWERING LOCUS T homologue, which is deleted in the Tanjil reference genome (Hane et al. 2017).
Table S5. Transcription factor motifs identified in the wild‐type sequence (ku; represented by accession P27255) of LanFTc1, the vernalisation responsive FT homologue in narrow‐leafed lupin (Lupinus angustifolius L.).
Table S6. A list of candidate transcription factor binding site motifs for sequences within the 5’ regulatory region and 5' UTR of LanFTc1 that are critical for establishing moderate (CM) or high (CH) levels of de‐repressed LanFTc1 expression during early vegetative growth.