

Abstract
Dengue is a major public health problem worldwide with distinct clinical manifestations: an acute presentation (dengue fever, DF) similar to other febrile illnesses (OFI) and a more severe, life-threatening form (severe dengue, SD). Due to nonspecific clinical presentation during the early phase of dengue infection, differentiating DF from OFI has remained a challenge, and current methods to determine severity of dengue remain poor early predictors. We present a prospective clinical cohort study conducted in Caracas, Venezuela from 2001–2005, designed to determine whether clinical and hematological parameters could distinguish DF from OFI, and identify early prognostic biomarkers of SD. From 204 enrolled suspected dengue patients, there were 111 confirmed dengue cases. Piecewise mixed effects regression and nonparametric statistics were used to analyze longitudinal records. Decreased serum albumin and fibrinogen along with increased D-dimer, thrombin-antithrombin complex, activated partial thromboplastin time and thrombin time were prognostic of SD on the day of defervescence. In the febrile phase, the day-to-day rates of change in serum albumin and fibrinogen concentration, along with platelet counts, were significantly decreased in dengue patients compared to OFI, while the day-to-day rates of change of lymphocytes (%) and thrombin time were increased. In dengue patients, the absolute lymphocytes to neutrophils ratio showed specific temporal increase, enabling classification of dengue patients entering the critical phase with an area under the ROC curve of 0.79. Secondary dengue patients had elongation of Thrombin time compared to primary cases while the D-dimer formation (fibrinolysis marker) remained always lower for secondary compared to primary cases. Based on partial analysis of 31 viral complete genomes, a high frequency of C-to-T transitions located at the third codon position was observed, suggesting deamination events with five major hot spots of amino acid polymorphic sites outside in non-structural proteins. No association of severe outcome was statistically significant for any of the five major polymorphic sites found. This study offers an improved understanding of dengue hemostasis and a novel way of approaching dengue diagnosis and disease prognosis using piecewise mixed effect regression modeling. It also suggests that a better discrimination of the day of disease can improve the diagnostic and prognostic classification power of clinical variables using ROC curve analysis. The piecewise mixed effect regression model corroborated key early clinical determinants of disease, and offers a time-series approach for future vaccine and pathogenesis clinical studies.
Author summary
Dengue fever results in a self-limiting, non-specific febrile illness. In approximately 10% of cases, the disease progresses to a severe, life-threatening syndrome. While hematological derangement is a key indicator of dengue, the mechanisms by which pathophysiological changes occur over the course of infection remain unclear. Additionally, there are limited clinical algorithms to facilitate rapid prognosis of dengue. We conducted a prospective clinical cohort study in Caracas, Venezuela to determine whether clinical and hematological parameters could distinguish dengue fever from other febrile illnesses, and identify early prognostic biomarkers of severe disease. Piecewise linear mixed effects regression models demonstrate that rates of change of albumin, fibrinogen, lymphocytes, platelets and thrombin time were significantly different between dengue and other febrile illnesses, and that the absolute value of albumin, fibrinogen, thrombin-antithrombin complex, thrombin time and partial thromboplastin time were prognostic of severe dengue on the day of defervescence. Our study offers extended insights into dengue pathogenesis and provides new approaches to dengue diagnosis and severity prognosis.
Introduction
Dengue virus (DENV) is a mosquito-transmitted flavivirus that endemically circulates as four antigenically distinct serotypes (DENV1-4). Incidence has grown dramatically worldwide in recent decades, with an estimated 390 million annual infections, of which 96 million manifest clinically [1–3]. The most common clinical presentation is a self-limiting illness characterized by fever, headache, myalgia and arthralgia (dengue fever, DF) [4]. In approximately 10% of cases, the disease progresses to severe dengue (SD), characterized by increased vascular permeability resulting in capillary leakage, and can lead to lethal dengue shock syndrome (DSS) [4].
In 2000, DENV-3 re-emerged in Venezuela causing a prolonged, major outbreak with more than 83,000 reported cases [5]. We conducted a prospective clinical cohort study in Caracas between 2001–2005 on patients with suspected dengue to better understand the pathogenesis and progression of the disease with respect to immune cell activation and hemostasis. We aimed to correlate changes in selected blood biomarkers with clinical severity to identify diagnostic and prognostic biomarkers of DF and SD.
Dengue disease severity is affected by both viral and human immune factors [6, 7]. Primary exposure to any one of four DENV serotypes induces lasting immunity against reinfection by the same serotype [8–10]. However, epidemiological studies have shown an increased risk of developing SD after secondary exposure to a different serotype [11, 12]. An antibody dependent enhancement hypothesis has been proposed wherein preexisting antibodies bind but do not neutralize virions of the subsequent infecting serotype [13]. Instead, these virus-immune complexes are recognized by Fcγ receptor-bearing cells that facilitate virus entry and replication, resulting in increased disease severity [14, 15]. Host protein cross-reactivity with DENV proteins has also been suggested to contribute to the pathogenesis of dengue [16–18].
Cytokine secretion and complement activation impair the endothelial barrier resulting in the leakage and loss of plasma components into the perivascular space, causing an increase in hematocrit and triggering the coagulation system. Studies on dengue patients indicate endothelial injury and/or activation by increased concentrations of von Willebrand factor (vWF) [19–22]. In addition, various processes contribute to thrombocytopenia including depression in bone marrow function, increase in megakaryocytes in the bone marrow, shortened platelet survival, and increased platelet consumption [23–27]. Neutropenia, lymphocytosis, as well as other elevated mediators including C3a, C5a, interleukins, tumor necrosis factor α, interferon γ, TRAIL, monocyte chemotactic protein 1, soluble suppression of tumorigenicity 2 (sST2, a member of the interleukin-1 receptor family), CXCL10 and histamine have also been associated with dengue pathobiology [28–40].
In SD, defects in coagulation such as prolonged activated partial thromboplastin time (aPTT), prothrombin time (PT), or thrombin time (TT) have been reported [41–43]. In addition, hyperfibrinolysis may occur in SD patients indicated by decreased plasma concentration of fibrinogen and increased concentration of its degradation products such as D-dimer (DD) [44–46]. A panel of peripheral white blood cell (WBC) and platelet counts, aPTT and PT, and blood chemistry including concentration of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) was recently found to be predictive of SD diagnosis [47–49].
Current guidelines separate the course of the disease into three phases: (i) the febrile phase, from onset of symptoms to the day preceding defervescence, (ii) the critical phase, comprised of the day of defervescence and the two following days and (iii) the subsequent recovery phase [4, 50]. Whereas most studies in the field consider the day of onset of symptoms as day 1, we and others defined the day of defervescence as day 0 (D0) [51, 52]. This standardization of disease progression around a pivotal event allows better comparison of the physiological processes around the clinically important critical phase and permits a more robust statistical analysis as the variance due to different febrile phase length is eliminated. However, the practical clinical use of any predictors of DF or SD in this analytical context would require the means to estimate the day of disease at admission.
Due to nonspecific clinical manifestations during the early phase of dengue infection, differentiating DF from OFI has remained a challenge. Current laboratory-based methods to diagnose dengue include serological tests and virus antigen and nucleic acid detection, but their use is limited, especially in regions without access to sophisticated equipment and well-trained personnel. As there is currently no specific treatment for DF and SD, timely and proper hydration via intravenous fluid replacement remains the most effective way to reduce mortality, but requires early detection of patients at risk. However, the criteria commonly used to differentiate DF and SD, such as hemoconcentration and platelets count, remain poor predictors of SD [53]. In 2009, the World Health Organization [4] suggested new guidelines for the classification of dengue fever based on warning signs and evidence of plasma leakage [4]. Although the sensitivity of this new classification to detect cases of SD is high, its specificity remains low. Dengvaxia is the first FDA-approved vaccine for dengue; however, vaccination is limited to people with a prior history of dengue, as it can exacerbate disease [13, 54, 55]. A better understanding of the disease pathogenesis and the identification of early prognostic biomarkers are therefore essential for the development of future diagnostic tests and supportive measures, and to define end-points for vaccination validation studies.
Materials and methods
Study population
Between 2001–2005, 204 patients with suspected dengue were enrolled from outpatient clinics within and around Caracas and referred to the Universidad Central de Venezuela, Centro Nacional de Hemofilia, Banco Municipal de Sangre. Inclusion criteria for the study were two or more days of febrile illness with one or more of the following symptoms: fever ≥38.5°C, headache, myalgia, or a maculopapular rash, and no sign of other obvious infection (Fig 1B). Exclusion criteria were recumbent systolic blood pressure <85 mmHg, significant bleeding prior to study enrollment, and patients unlikely to attend or who did not attend follow up visits.
Fig 1.

A. Diagram of the time-line for data collection. B. Flow diagram representation of study inclusion and exclusion criteria. Exclusion criteria included recumbent systolic blood pressure <85 mmHg, significant bleeding prior to study enrollment, and patients unlikely to attend or that did not attend follow-up visits. 149 patients completed the protocol. 1 Headache, myalgia or maculopapular rash; 2 Routine laboratory testing: complete blood counts, PT, PTT, TT, albumin, AST and ALT. Specialized coagulation studies: TAT, F1+2, fibrinogen, D-D and vWF; 3 Signs of hypovolemia were defined using the following criteria: (heart rate / systolic blood pressure) ≥ 1 or (systolic–diastolic blood pressure) ≤ 20 mmHg; HI: hemagglutination inhibition; RT-PCR: reverse transcription polymerase chain reaction.
Ethics statement
All adult patients provided written informed consent, and a parent or guardian of any child participant provided written informed consent on the child’s behalf. The study was approved by the ethics review board of the Universidad Central de Venezuela, Centro Nacional de Hemofilia, Banco Municipal de Sangre (IRB number H-3693).
Data collection
Schematics of data collection are shown in Fig 1A. Briefly, after the initial visit and enrollment to the study, each patient attended the outpatient clinic daily. On every visit, medical history and physical findings, including hemorrhagic signs, were recorded on standardized case report forms and entered into a secured, centralized database. The patients also provided a venous blood specimen at each of these visits, prior to receiving IV saline solution. The blood samples were used to determine: the complete blood cell counts, the concentrations of albumin and liver enzymes and the activity parameters of coagulation and fibrinolysis. Patients were instructed to keep a record of oral temperatures three times a day and independently recorded these values at home. The patients were asked to return to the study clinic for a convalescent visit 2 weeks after enrollment for endpoint measurements as proxies for baseline status. Serological analyses for dengue immunoglobulin G and M (IgG/IgM) were performed on the day of study enrollment per protocol of the Instituto Nacional de Salud, 2 days after defervescence and on the convalescent visit.
sST2 concentrations were also measured in a randomly selected sample of 31 patients. One serum sample for each enrolled patient was processed for RT-PCR and a subset for exon sequencing. The archived data was utilized to generate multiple comparisons using MAFFT, with default settings. The consensus sequence was calculated using the default options of the EMBOSS Cons program (https://www.ebi.ac.uk/Tools/msa/emboss_cons/). The sequences analyzed are deposited at NCIB gene bank: FJ373304.1, FJ373303.1, FJ182015.1, EU854292.1, EU854291.1, EU660420.1, EU569691.1, EU569690.1, EU569689.1, EU569688.1, EU529691.1, EU529690.1, EU529689.1, EU529688.1, EU529687.1, EU529686.1, EU529685.1, EU529684.1, EU482614.1, EU482613.1, EU482612.1, KF955486.1, KF955487.1, KF955447.1, KF955330.1, KF955449.1, KF955450.1, KF955331.1, KF955452.1, KF955451.1, KF955453.1, KF955454.1. We confirmed that KF955487 and FJ182015 are two identical genome sequences from a duplicate of the same patient sample. The list contains a total of 31 unique DENV-3 genomes, out of 43 samples that were used in the analysis.
Laboratory tests
Blood samples were collected in Vacutainer tubes (2% EDTA, 3.2% sodium citrate, and tubes without anticoagulant) and immediately used for analyses. Blood was tested for WBC and platelet counts, hemoglobin and hematocrit concentration (Beckman-Coulter). Reticulocytes were counted with a supravital stain. Plasma was used to perform coagulation tests including aPTT, PT, and TT, and concentration of fibrinogen was measured (Diagnostica Stago). Plasma concentrations of vWF (Diagostica Stago) prothrombin fragment 1+2 (F1+2, Dade-Behring), thrombin-antithrombin complex (TAT, Dade-Behring), DD (Diagnostica Stago), and sST2 were measured (MBL ELISA) according to the manufacturer’s instructions. Concentrations of AST, ALT and albumin were measured in an automated analyzer (Wiener Lab, Argentina). Dengue IgM was measured by MAC ELISA and IgG titer with a hemagglutination inhibition assay (HI) from the Instituto Nacional de Salud in Venezuela [56]. Additionally, IgG ELISAs were utilized from commercially available sources (PanBio).
Dengue virus genomic RNA was isolated from febrile serum samples using the QIAmp Viral RNA kit (QIAGEN) and DENV serotype-specific reverse transcription and polymerase chain reaction (RT-PCR, QIAGEN) was performed as previously described [57]. The 31 sequenced viral genomes were uploaded to Genbank (S1 Table).
Classification criteria
Patients were classified as dengue patients or as OFI based on the detection of genomic dengue RNA using RT-PCR, presence of IgM antibodies or seroconversion (four-fold increase in HI levels in the second sample compared to the first). The HI levels were used to further classify dengue patients as primary infection (HI titer ≤ 1:1280) or secondary infection (HI titer > 1:1280 in the first sample) [38].
The criteria used to classify a patient as having SD were genomically or serologically confirmed dengue infection plus at least one of the following: presence of any type of bleeding, presence of effusion or edema, signs of hypovolemia defined as pulse pressure (systolic blood pressure–diastolic blood pressure) ≤ 20 mmHg and/or heart rate ≥ systolic blood pressure. This corresponds to a risk-averse interpretation of the 2009 WHO criteria for SD, placing an emphasis on endothelial dysregulation.
Statistical analysis
Statistical analyses were performed using R (v.3.5.0) and data completion using SAS 9.4. Two-sided Mann-Whitney U tests were performed due to the non-normal distributions of many variables, without correction for multiple comparisons. Statistics were only performed when at least five observations were present in each group for a given day and variable.
To examine longitudinal trends of outcome variables adjusted for patient-related covariates, we used piecewise linear mixed effects regression models as previously described [58–61]. The time period under observation was divided into two main stages: pre- (D-3 to D-1, ttime.before) and post- (D0 to D+3, ttime.after) defervescence. The equation for the piecewise regression model is as follows:
where Yit indicates longitudinal measurements for a variable of interest for i-patient at t-time; ttime.before indicates the pre-defervescence day; ttime.after indicates the post-defervescence day; and β0 is the intercept of the model and a population estimate on the day of defervescence (D0). The regression parameters indicate the slopes, or rate of change per day for pre- (βb) and post-defervescence (βa) time period, utilizing D0 as the baseline. In this context, a statistically significant difference in slopes between two groups indicates a different day-to-day rate of change at patient level for a given biomarker. A statistically significant difference in the intercept indicates a difference in absolute value at D0 and at population level for a given biomarker.
We performed receiver operator characteristic (ROC) curves to determine whether knowledge about the day of illness could improve the DF/SD discrimination power of a particular variable. Optimal cutoffs were identified for each day between D-2 and D+1, for the indiscriminate first day of consultation and for the median value of each patient, using the Youden index (J = sensitivity + specificity -1). The areas under the curve (AUC) were calculated.
To perform initial alignment of DENV genomes, we used Prank (v.150803). The alignment was then submitted to codeml program in the PAML (v.4.8) suite of tools [62, 63]. From the codons output by codeml, separate alignments of first, second, and third codon positions were derived. For each column in each alignment, the changes from the ancestral sequence state to each other state were counted separately and divided by the number of isolates represented by nucleotides in the column to get a proportion for each mutation. Gap characters due to incomplete sequences were not counted. The graph was drawn using Plotnine, an implementation of ggplot in Python.
Results
Of 204 patients enrolled, 149 completed the protocol. The median age was 20 years (range 5 to 74) and 61% were female. We confirmed dengue infection in 111 cases with 52 patients (47%) classified as DF and 59 (53%) as SD. All received supportive therapy and survived. 90 (81%) patients experienced primary and 19 (17%) secondary dengue, while 2 (2%) could not be classified (Table 1). The patients were observed between 0 and 5 days (median = 1) before and between 1 and 7 days (median = 3) after defervescence (including defervescence day, D0). Overall, 70% of all daily designated clinical assessments and laboratory tests were completed, ranging from 3% to 97% completion. We determined that 65 out of 78 RT-PCR positive cases (83%) were infected with dengue serotype three.
Table 1. Description of the study population.
| Caracas Cohort | |||||
|---|---|---|---|---|---|
| Severe Dengue | Dengue Fever | Other Febrile Illnesses | Excluded | Total | |
| Total Group | 59 | 52 | 38 | 55 | 204 |
| Age Median (Q25-Q75) | 18 (12–33.5) | 23.5 (14–37) | 23.5 (12.3–39.5) | 20 (14–35) | 20 (13–35) |
| Age, <7 / ≥7 | 2/57 | 1/51 | 1/37 | 4/51 | 8/196 |
| Sex, F / M | 30/29 | 19/33 | 17/21 | 25/30 | 91/113 |
| Serology, 1° / 2° dengue/Unknown | 46/11/2 | 44/8/0 | NA | NA | NA |
Non-parametric statistics
We performed comparative statistics on hematological test results obtained from patients with dengue (DF and SD) versus OFI. In the febrile phase, the dengue group had significantly decreased platelets and WBC median counts while having increased AST, ALT, DD and TAT complex median plasma concentrations. On D0, median plasma concentrations of fibrinogen and albumin as well as WBC absolute and neutrophils relative counts were decreased, whereas concentration of TAT, DD, AST, ALT, vWF and sST2 were elevated in the dengue group before defervescence compared to the OFI group. aPTT and TT were also prolonged in dengue cases compared to the OFI group. In the post-defervescence phase, dengue patients had decreased albumin and fibrinogen median plasma concentration, a decreased PT, an increased TT as well as elevated AST, ALT, vWF plasma concentration and relative lymphocytes count. (Figs 2A and 3A and S1 Fig).
Fig 2. Overview of the blood biomarkers changes between the different groups.

A. Significant changes in blood markers for each day using Mann-Whitney U test. D0 represent the day of defervescence. In the comparison dengue VS other febrile illnesses, dengue comprises both dengue fever and severe dengue cases. B. Significant changes in blood biomarkers kinetic and intercept using the piecewise linear mixed effect regression model. Before and after represent two slopes while intercept is the biomarker’s value on D0. Blue, significant decrease. Red, significant increase.
Fig 3. Evolution of selected blood biomarkers for selected groups comparison over the course of disease.

A. Evolution of the medians (and interquartile ranges) with comparison between dengue and other febrile illness patients. B. Evolution of the biomarkers at patient level using piecewise linear mixed effects models with comparison between dengue and other febrile illness patients. C. Evolution of the medians (and interquartile ranges) with comparison between dengue fever and severe dengue patients. In A and C, the linking lines are displayed for a better visualization but do not represent the evolution of the biomarkers at patient level. D0 is the day of defervescence.
When comparing patients with primary dengue to patients with secondary dengue, the albumin and hemoglobin concentration as well as the relative neutrophils and monocytes counts were significantly decreased on the day of defervescence. The WBC count and the relative lymphocytes count, representing lymphoproliferations, as well as the TT were elevated in secondary cases compared to primary cases (Fig 2A and S2 Fig).
On the day of defervescence (D0), SD patients had significantly decreased median plasma concentration of serum albumin and fibrinogen compared to DF, while DD plasma concentration was increased and the TT was elongated. The median plasma concentration of sST2 were elevated in the SD group between D-1 and D+1 (190.4 vs 152.6 at D-1, 256.4 vs 109.4 at D0 and 98.1 vs 64.5 at D+1), but the results were not statistically significant. (Figs 2A and 3C and S3 Fig). In the post-defervescence phase, the SD group had significantly decreased albumin and increased DD median plasma concentration compared to the DF group. SD patients had significantly higher heart rates and lower systolic and diastolic blood pressure around D0 compared to DF cases.
The full comparative statistics, including medians, quartiles and p-values for each variable, at each day and in each group can be found in S2 Table (DF vs OFI), S3 Table (primary vs secondary dengue) and S4 Table (DF vs SD).
Piecewise linear mixed effects regression
We next performed a piecewise linear mixed effects model analysis to investigate the evolution of each variable over time (fixed effect) and its expected value at D0 (intercept). Compared to OFI, dengue cases had a sharper increase in relative lymphocyte count and TT and a sharper decrease in platelet count during the pre-defervescence phase. Plasma concentrations of albumin and fibrinogen decreased in dengue patients while increased in OFI. On D0, the dengue group had lower albumin and fibrinogen plasma concentrations, as well as lower WBC, neutrophil and platelets counts. Plasma concentrations of AST, ALT and vWF as well as relative lymphocytes count, aPTT and TT were higher in the dengue group. During the post-defervescence phase, WBC counts increased in dengue patients while decreased in OFI cases (Figs 2B and 3B and S4 Fig).
The linear mixed effects models demonstrated that, compared to DF cases, SD cases had a steeper increase (slope is statistically different) in plasma concentration of AST during the febrile phase. On D0, SD patients had lower plasma concentrations of albumin and fibrinogen, serum concentrations of AST and TAT were higher, and prolonged aPTT and TT were observed (Fig 2B and S5 Fig). No statistically significant differences were observed during the post-defervescence phase. The model could not be applied to some clinical variables like DD due to a relative lack of data points compared to the total number of random effects. The full model statistics can be found in S5 Table (dengue vs OFI) and S6 Table (DF vs SD).
ROC statistics
We further conducted ROC analysis using the longitudinal clinical and hematological data to evaluate diagnostic and prognostic test value for candidate biomarkers. Counts of WBC (AUC = 0.89 at D-1), reticulocytes (AUC = 0.77 at D-1), and platelets (AUC = 0.88 at D+1), TT (AUC = 0.85 at D0), and plasma concentrations of AST (AUC = 0.82 at D0), ALT (AUC = 0.86 at D-2), vWF (AUC = 0.77 at D-1), fibrinogen (AUC = 0.75 at D0), TAT (AUC = 0.75 at D-1) and D-dimer (AUC = 0.80 at D-2) appeared as the best candidate biomarkers to differentiate between dengue and OFI. Plasma concentrations of albumin (AUC = 0.71 at D0), fibrinogen (AUC = 0.69 at D0) and D-dimer (AUC = 0.70 at D0) could be useful in distinguishing between DF and SD. The full tables with AUC, best cutoffs, sensitivity, specificity and confidence interval are accessible in S7 Table (DF vs OFI) and S8 Table (DF vs SD). We noticed large variation and difference of up to 0.21 in AUC when comparing the ROC of a biomarker at a defined day of illness to the ROC at the indiscriminate day of first contact with healthcare (mostly between D-3 and D0 (Table 2). Nevertheless, none of these differences were statistically significant.
Table 2. Evolution of the receiver operating characteristics statistics for selected biomarkers over the course of disease.
Classification between the dengue and OFI groups. First contact: day of first contact with healthcare (independently of the day of fever). Median days -2 to 1: a posteriori calculation of the biomarker’s median value between D-2 and D+1. AUC: area under the curve; CI: confidence interval.
| Day | |||||||
|---|---|---|---|---|---|---|---|
| Variables | First Contact | -2 | -1 | 0 | 1 | Median days -2 to 1 | |
| White blood cells, *1000/μl | Best cutoff, Youden index | 5.25 | 5.55 | 3.95 | 3.35 | 4.25 | 4.08 |
| AUC | 0.76 | 0.82 | 0.89 | 0.79 | 0.67 | 0.8 | |
| 95% CI AUC | 0.66–0.86 | 0.70–0.94 | 0.82–0.96 | 0.7–0.87 | 0.56–0.77 | 0.72–0.89 | |
| Neutrophils, % | Best cutoff, Youden index | 84.5 | 57.5 | 72 | 51.5 | 47.5 | 51.75 |
| AUC | 0.5 | 0.6 | 0.58 | 0.63 | 0.7 | 0.66 | |
| 95% CI AUC | 0.38–0.61 | 0.37–0.83 | 0.44–0.72 | 0.52–0.74 | 0.6–0.8 | 0.56–0.76 | |
| Lymphocytes, % | Best cutoff, Youden index | 11.5 | 36.5 | 25.5 | 33.5 | 51.5 | 38.25 |
| AUC | 0.51 | 0.61 | 0.57 | 0.65 | 0.72 | 0.67 | |
| 95% CI AUC | 0.40–0.63 | 0.39–0.82 | 0.43–0.71 | 0.54–0.76 | 0.62–0.82 | 0.57–0.77 | |
| Platelets, *1000/μl | Best cutoff, Youden index | 194.5 | 199.5 | 170 | 157.5 | 166.5 | 160 |
| AUC | 0.74 | 0.76 | 0.81 | 0.85 | 0.88 | 0.84 | |
| 95% CI AUC | 0.64–0.83 | 0.61–0.90 | 0.7–0.92 | 0.78–0.92 | 0.81–0.95 | 0.76–0.92 | |
| Thrombin Time, difference in sec | Best cutoff, Youden index | 3.3 | 0.1 | 2.45 | 2.8 | 1.95 | 1.75 |
| AUC | 0.7 | 0.69 | 0.73 | 0.85 | 0.71 | 0.8 | |
| 95% CI AUC | 0.59–0.80 | 0.49–0.88 | 0.62–0.85 | 0.77–0.92 | 0.6–0.82 | 0.72–0.88 | |
The above results suggest that better knowledge of the day of illness could improve the classification power of certain tests and provide clinicians with valuable insights about the onset of the critical phase. We therefore assessed the ability of biomarkers to discern the day of illness and identified the absolute lymphocytes/neutrophils ratio as a potential time predictor in dengue patients. In patients with confirmed dengue infection, there is a quasi-linear correlation between days (D-2 to D+1) and the proposed ratio (Fig 4). ROC statistics and optimal cutoffs were calculated to differentiate the critical day (D0) from previous days of illness in febrile dengue patients. The AUC was 0.79 and the best cutoff using the Youden index was 0.537 with 65% specificity and 80% sensitivity. Using a cutoff of 0.978, patients entering the critical phase could be identified with 90% specificity and 47% sensitivity.
Fig 4. Evolution of neutrophils and lymphocytes absolute counts and their ratio.
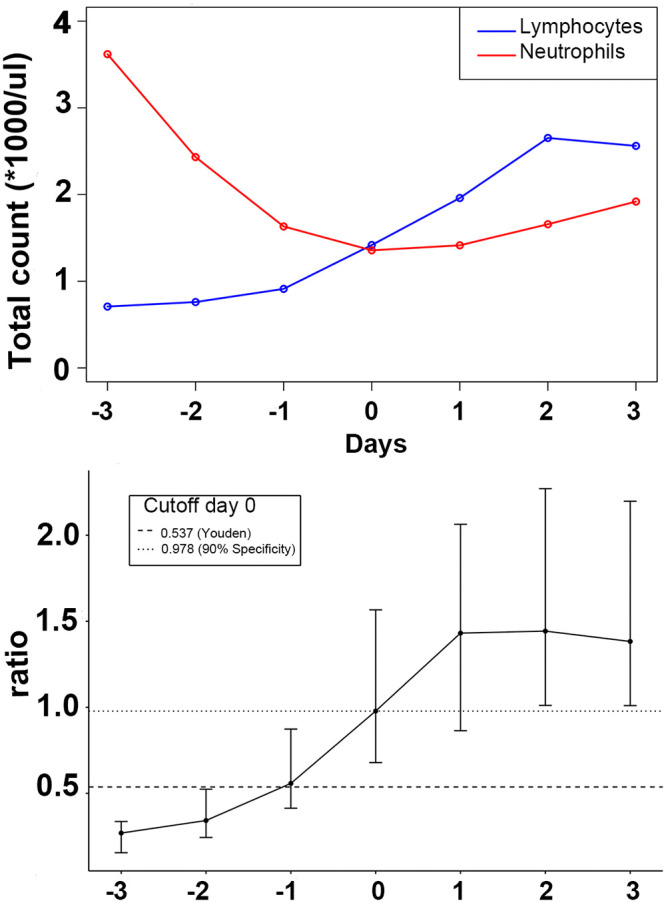
A. Evolution of the neutrophils and lymphocytes absolute counts’ medians for dengue patients. B. Evolution of the neutrophils over lymphocytes ratio and selected cutoffs (method used) to differentiate between D0 and any previous day. The intervals represent the range between the first and the third quartiles. (- - -) sensitivity = 0.80, specificity = 0.65; (·····) sensitivity = 0.47, specificity = 0.90.
Genome analysis
In the 31 sequenced DENV3 genomes out of 78 RT-PCR positives (40%), we observed 49 amino acid mutations well distributed across the genome. We observed higher frequency of T-to-C/C-to-T substitutions than other mutations, and these transitions were 7.6-fold higher at the third codon position (S6A Fig). The limited number of collected specimens with completed sequence data precluded formal assessment of the association between mutations and disease severity. The four serotypes of dengue co-circulated during the dengue outbreak according to RT-PCR data, where dengue 3 represented the majority (83%). All the sequenced DENV-3 genomes were classified as Genotype V (S6B Fig).
Discussion
Our findings demonstrate detailed differences in laboratory features between OFI, DF, and SD during a dengue epidemic in Caracas, Venezuela. The results of the mixed effects models were consistent with the results obtained using non-parametric statistics. At D0, 12 different laboratory parameters had classifier effects for dengue vs OFI and 6 when comparing DF with SD. The results of the piecewise mixed effects model indicate statistically significant differences in the rates of change of 5 biomarkers before, and one after, defervescence in dengue vs OFI and of one single biomarker in the pre-defervescence phase for DF vs SD.
At D0, coagulation biomarkers were important differentiators between DF and OFI, including increased TT, increased concentrations of vWF and TAT complex, and decreased concentrations of fibrinogen and DD. Additional diagnostic biomarkers were decreased concentration of albumin, increased concentration of sST2, decreased WBC and platelets counts, increased percentage of lymphocytes, and decreased percentage of neutrophils. Most of these biomarkers were also diagnostic during the febrile phase.
The results highlighted seven hematological biomarkers (albumin, AST, fibrinogen, DD and TAT-complex concentrations as well as aPTT and TT coagulation tests) as prognostic of SD at D0. These findings suggest an exacerbation of the dengue-related coagulation disorders in patients experiencing SD. The majority of our findings are consistent with the existing literature indicating coagulation factors can be used to define the severity of the disease. One noticeable exception is the hematocrit, which was found to be lower in SD patients compared to DF, where it would be expected to be higher due to hemoconcentration. This could be explained by the hydration protocol in place during this hospital study. Another novel finding is that fibrinolysis is not as prominent in secondary dengue cases compared to primary cases as the concentration if DD was consistently lower compared to primary infections. This could be due to the more prominent extravasation of fibrinogen and lower serum levels, as well as a possible decrease in the hepatic synthesis of fibrinogen during secondary dengue infections. Notably, the differences in DD between primary and secondary were not statistically significant; rather, the results observed are general trends.
Some of the observations derived from our time-series analysis provide a better understanding of dengue pathogenesis. The elongation in TT and aPTT, coupled with decreased levels of fibrinogen and increased DD suggest a procoagulatory state with an associated activation of fibrinolysis as a reaction mechanism. This hypothesis is further supported by elevated TAT complex and F1+2 concentrations, suggesting an enhanced conversion rate from prothrombin to thrombin. Endothelial damage, as demonstrated by the elevated levels of vWF, is likely to be a trigger for the procoagulatory state alongside elevated tissue factor (TF) whose plasma levels have been shown to increase during febrile phases [22, 64]. The decreased levels of fibrinogen could be explained by an increased consumption due to the procoagulatory state but also by the formation of immune complexes with antibodies against viral proteins (e.g. anti-NS1). We therefore support the use of blood coagulation biomarkers (such as coagulation times, Fibrinogen, TAT complex, vWF, and DD), often not considered, to define severity end points in future dengue studies.
We observed that the timing of neutropenia was associated with platelet decrease. A possible explanation for the loss of neutrophils from the circulation during the febrile phase is neutrophil attachment, rolling, and extravasation, potentially mediated by increased expression of P-selectin in the vascular endothelium. This hypothesis is supported by an increase of P-selection mRNA in endothelial cells infected with dengue virus in vitro [65]. Clinical data in humans reporting endothelium activation and neutrophil extravasation during neutropenia further support this hypothesis [66]. To test this hypothesis, it would be useful to quantitatively evaluate the loss of neutrophils due to extravasation versus attrition or suppressed production as previously described in animal models [67].
To date, hematological studies have been primarily focused on dengue patients from Asia. Our study is the first to provide detailed information about the timing of hemostatic biomarker changes at each time point in a large dengue cohort in Latin America. In addition, we examined longitudinal trends of outcome variables in patients by uniquely utilizing piecewise linear mixed effects regression models. The rates of change of albumin, fibrinogen, lymphocytes, platelets and TT were shown to be significantly different between dengue and OFI patients in the febrile phase. From a clinical perspective, these findings suggest that at least two consecutive days of consultation and laboratory tests could increase the diagnostic power of several biomarkers.
Given the variation in accuracy of many biomarkers over the course of disease and the importance of close monitoring during the critical phase, we hypothesized that additional information about the day of illness relative to D0 would benefit clinicians and patients. This could improve the diagnostic and prognostic accuracy of specific biomarkers while simultaneously improving risk and patient flow management. Given a day of first contact with the healthcare system approximately evenly distributed between D-3 and D0 and a febrile phase lasting between 2 and 7 days, any prediction about the onset of the critical phase based only on the day of first symptoms or of first consultation remains unreliable. In our cohort, the ratio of decreasing neutrophils over increasing lymphocytes (absolute counts) was progressively more pronounced over the course of the disease for patients infected with dengue virus. We propose this ratio as a surrogate to identify the day of illness relative to D0. A ratio over 1.0 classifies febrile patients entering the critical phase with a specificity of 90%. In resource limited settings, a better anticipation of the onset of the critical phase could help manage follow-up consultation and inpatient admissions. Moreover, the costs for such additional inference is minimal as the needed biomarker data are already routinely collected as part of the care process. The clinical use of this ratio would need confirmation in additional, larger cohorts.
The higher frequency of C-to-T/T-to-C substitutions at the third codon position observed in the genome analysis is consistent with our previously published studies on Zika and Ebola viruses [68, 69]. These data suggest that host RNA editing enzymes (e.g. ADARs) and chemical deamination may contribute to viral RNA diversity; however, further investigation is required to determine the exact mechanisms of the observed deaminations. Our phylogenetic analysis suggested that the epidemic DENV-3 strain originated in the Caribbean in 1998 (S6B Fig).
Our study has some limitations. Most of the patients in our cohort were infected with DENV-3 and generalization of our results would need confirmation in populations infected with other DENV serotypes. Due to the outpatient care setting of the study, patients presenting with more than 48 hours of fever, significant bleeding (based on clinical history and physician judgment) or signs of shock during the first visit, were excluded. This might have influenced the recruitment of SD patients. Moreover, reliable information about underlying comorbidities, which could explain abnormal biomarker levels, were available as per clinical history but not confirmed with other tests outside the protocol established in the study. Due to the requirement of a long 4 kb open reading frame for RT-PCR procedures (Schmidt et al, 2011), of the 43 samples submitted for sequencing at genomic facility, 25 resulted in complete and 6 in incomplete genomes, for a total of 31 unique genome sequences for dengue serotype 3. This represented 72% of the total number of samples submitted but only 40% of the 78 PCR positive patient samples acquired. The techniques for sequencing RNA viral genomes at that time resulted in a loss of some full genomic sequences and thereby precluded inference about the association between point mutations and disease severity. We provide the successfully sequenced genomes and their corresponding disease outcome in S1 Table to support findings of genome variability among patients’ samples from the same outbreak.
The combination of classic and novel time-series statistics confirms and expands existing knowledge, offers extended insights into dengue pathogenesis, and provides possible new approaches to dengue diagnosis and severity prognosis that should have applicability in future studies, including those for vaccine trials.
Supporting information
DF: dengue fever; SD: severe dengue.
(XLSX)
(XLSX)
(XLSX)
(XLSX)
β0: intercept and value at D0; βb: slope time before [D-3 to D0); βa: slope time after [D0 to D+3); SD: standard deviation; 1s-t-test: 1 sample t-test; 2s-t-test: 2 samples t-test.
(XLSX)
β0: intercept and value at D0; βb: slope time before [D-3 to D0); βa: slope time after [D0 to D+3); SD: standard deviation; 1s-t-test: 1 sample t-test; 2s-t-test: 2 samples t-test.
(XLSX)
(XLSX)
(XLSX)
The linking lines are displayed for a better visualization but do not represent the evolution of the biomarkers at patient level. *p-value ≤ 0.05; **p-value ≤ 0.01; ***p-value ≤ 0.001.
(TIF)
The lower and upper error bars represent the first and third quartiles respectively. The linking lines are displayed for a better visualization but do not represent the evolution of the biomarkers at patient level. *p-value ≤ 0.05; **p-value ≤ 0.01; ***p-value ≤ 0.001.
(TIF)
The lower and upper error bars represent the first and third quartiles respectively. The linking lines are displayed for a better visualization but do not represent the evolution of the biomarkers at patient level. *p-value ≤ 0.05; **p-value ≤ 0.01; ***p-value ≤ 0.001.
(TIF)
No results displayed for D-dimer, monocytes, reticulocytes and sST2 as the models could partially not be fitted. *p-value ≤ 0.05; **p-value ≤ 0.01; ***p-value ≤ 0.001.
(TIF)
No results displayed for D-dimer as the models could partially not be fitted. *p-value ≤ 0.05; **p-value ≤ 0.01; ***p-value ≤ 0.001.
(TIF)
A. Distribution of amino acid mutations across the DENV-3 genome, separated by codon positions. B. Phylogenetic tree showing Caracas samples collected in 2001 in red, other Caracas samples in blue, and isolates from nearby locations in grey. Tree was found using RAxML rapid bootstrapping with 100 bootstrap replicates.
(TIFF)
Acknowledgments
BV thanks AHS and IB for their supervision and mentoring. The authors are grateful to Lee Gehrke at the Massachusetts Institute of Technology for support of the work of BV and IB. To Alan Rothman and Francis Ennis for their help in protocol design and support to IB and NB. The authors thank Ana Aleman, Mariella Lilue, Carmen Carpio and Jose Angel Ilarraza for patient enrollment and data collection; the personnel of Instituto Nacional de Higiene Rafael Rangel for support on serologic testing; Isabel Gonzalez- Bocco for manuscript editorial support. Finally, we thank the medical and scientific staff of the Banco Municipal de Sangre and all of the patients and families who participated in the study.
Data Availability
All relevant data are within the manuscript and its supporting information files.
Funding Statement
This work was supported by grant U01 AI45440 from the National Institutes of Health and funding from the Ministerio de Salud y Desarollo Social, Venezuela to IB and NB. BV was supported by a Mercator Fellowship and is currently a Berrow Foundation Lord Florey scholar at Lincoln College, Oxford. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Knipe DM, Howley PM, Griffin DE. Fundamental virology. 4th ed Philadelphia: Lippincott Williams & Wilkins; 2001. xi, 1395 p. p. [Google Scholar]
- 2.Guzman MG, Harris E. Dengue. Lancet. 2015;385(9966):453–65. 10.1016/S0140-6736(14)60572-9 [DOI] [PubMed] [Google Scholar]
- 3.Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, et al. The global distribution and burden of dengue. Nature. 2013;496(7446):504–7. 10.1038/nature12060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.WHO. Dengue: guidelines for diagnosis, treatment, prevention and control: World Health Organization; 2009. [PubMed]
- 5.Brathwaite Dick O, San Martin JL, Montoya RH, del Diego J, Zambrano B, Dayan GH. The history of dengue outbreaks in the Americas. Am J Trop Med Hyg. 2012;87(4):584–93. 10.4269/ajtmh.2012.11-0770 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rothman AL. Immunity to dengue virus: a tale of original antigenic sin and tropical cytokine storms. Nat Rev Immunol. 2011;11(8):532–43. 10.1038/nri3014 [DOI] [PubMed] [Google Scholar]
- 7.Screaton G, Mongkolsapaya J, Yacoub S, Roberts C. New insights into the immunopathology and control of dengue virus infection. Nat Rev Immunol. 2015;15(12):745–59. 10.1038/nri3916 [DOI] [PubMed] [Google Scholar]
- 8.Thomas SJ, Nisalak A, Anderson KB, Libraty DH, Kalayanarooj S, Vaughn DW, et al. Dengue plaque reduction neutralization test (PRNT) in primary and secondary dengue virus infections: How alterations in assay conditions impact performance. Am J Trop Med Hyg. 2009;81(5):825–33. 10.4269/ajtmh.2009.08-0625 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Halstead SB, Rojanasuphot S, Sangkawibha N. Original antigenic sin in dengue. Am J Trop Med Hyg. 1983;32(1):154–6. 10.4269/ajtmh.1983.32.154 [DOI] [PubMed] [Google Scholar]
- 10.van Panhuis WG, Gibbons RV, Endy TP, Rothman AL, Srikiatkhachorn A, Nisalak A, et al. Inferring the serotype associated with dengue virus infections on the basis of pre- and postinfection neutralizing antibody titers. J Infect Dis. 2010;202(7):1002–10. 10.1086/656141 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Libraty DH, Acosta LP, Tallo V, Segubre-Mercado E, Bautista A, Potts JA, et al. A prospective nested case-control study of Dengue in infants: rethinking and refining the antibody-dependent enhancement dengue hemorrhagic fever model. PLoS Med. 2009;6(10):e1000171 10.1371/journal.pmed.1000171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kliks SC, Nimmanitya S, Nisalak A, Burke DS. Evidence that maternal dengue antibodies are important in the development of dengue hemorrhagic fever in infants. Am J Trop Med Hyg. 1988;38(2):411–9. 10.4269/ajtmh.1988.38.411 [DOI] [PubMed] [Google Scholar]
- 13.Halstead SB, O’Rourke EJ. Antibody-enhanced dengue virus infection in primate leukocytes. Nature. 1977;265(5596):739–41. 10.1038/265739a0 [DOI] [PubMed] [Google Scholar]
- 14.Halstead SB, Udomsakdi S, Simasthien P, Singharaj P, Sukhavachana P, Nisalak A. Observations related to pathogenesis of dengue hemorrhagic fever. I. Experience with classification of dengue viruses. Yale J Biol Med. 1970;42(5):261–75. [PMC free article] [PubMed] [Google Scholar]
- 15.Guzman MG, Alvarez M, Halstead SB. Secondary infection as a risk factor for dengue hemorrhagic fever/dengue shock syndrome: an historical perspective and role of antibody-dependent enhancement of infection. Arch Virol. 2013;158(7):1445–59. 10.1007/s00705-013-1645-3 [DOI] [PubMed] [Google Scholar]
- 16.Cheng HJ, Lei HY, Lin CF, Luo YH, Wan SW, Liu HS, et al. Anti-dengue virus nonstructural protein 1 antibodies recognize protein disulfide isomerase on platelets and inhibit platelet aggregation. Mol Immunol. 2009;47(2–3):398–406. 10.1016/j.molimm.2009.08.033 [DOI] [PubMed] [Google Scholar]
- 17.Cheng HJ, Luo YH, Wan SW, Lin CF, Wang ST, Hung NT, et al. Correlation between serum levels of anti-endothelial cell autoantigen and anti-dengue virus nonstructural protein 1 antibodies in dengue patients. Am J Trop Med Hyg. 2015;92(5):989–95. 10.4269/ajtmh.14-0162 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sun DS, Chang YC, Lien TS, King CC, Shih YL, Huang HS, et al. Endothelial Cell Sensitization by Death Receptor Fractions of an Anti-Dengue Nonstructural Protein 1 Antibody Induced Plasma Leakage, Coagulopathy, and Mortality in Mice. J Immunol. 2015;195(6):2743–53. 10.4049/jimmunol.1500136 [DOI] [PubMed] [Google Scholar]
- 19.Borchiellini A, Fijnvandraat K, ten Cate JW, Pajkrt D, van Deventer SJ, Pasterkamp G, et al. Quantitative analysis of von Willebrand factor propeptide release in vivo: effect of experimental endotoxemia and administration of 1-deamino-8-D-arginine vasopressin in humans. Blood. 1996;88(8):2951–8. [PubMed] [Google Scholar]
- 20.Basuki PS. A glance at the von Willebrand factor in dengue virus infection. Southeast Asian J Trop Med Public Health. 2003;34(3):559–63. [PubMed] [Google Scholar]
- 21.Chuansumrit A, Puripokai C, Butthep P, Wongtiraporn W, Sasanakul W, Tangnararatchakit K, et al. Laboratory predictors of dengue shock syndrome during the febrile stage. Southeast Asian J Trop Med Public Health. 2010;41(2):326–32. [PubMed] [Google Scholar]
- 22.Sosothikul D, Seksarn P, Pongsewalak S, Thisyakorn U, Lusher J. Activation of endothelial cells, coagulation and fibrinolysis in children with Dengue virus infection. Thromb Haemost. 2007;97(4):627–34. [PubMed] [Google Scholar]
- 23.Srichaikul T, Nimmannitya S. Haematology in dengue and dengue haemorrhagic fever. Baillieres Best Pract Res Clin Haematol. 2000;13(2):261–76. 10.1053/beha.2000.0073 [DOI] [PubMed] [Google Scholar]
- 24.Kho LK, Wulur H, Himawan T. Blood and bone marrow changes in dengue haemorrhagic fever. Paediatr Indones. 1972;12(1):31–9. [PubMed] [Google Scholar]
- 25.Putintseva E, Vega G, Fernandez L. Alterations in thrombopoiesis in patients with thrombocytopenia produced by dengue hemorrhagic fever. Nouv Rev Fr Hematol. 1986;28(5):269–73. [PubMed] [Google Scholar]
- 26.Krishnamurti C, Peat RA, Cutting MA, Rothwell SW. Platelet adhesion to dengue-2 virus-infected endothelial cells. Am J Trop Med Hyg. 2002;66(4):435–41. 10.4269/ajtmh.2002.66.435 [DOI] [PubMed] [Google Scholar]
- 27.Srichaikul T, Nimmannitya S, Sripaisarn T, Kamolsilpa M, Pulgate C. Platelet function during the acute phase of dengue hemorrhagic fever. Southeast Asian J Trop Med Public Health. 1989;20(1):19–25. [PubMed] [Google Scholar]
- 28.Malasit P. Complement and dengue haemorrhagic fever/shock syndrome. Southeast Asian J Trop Med Public Health. 1987;18(3):316–20. [PubMed] [Google Scholar]
- 29.Suvatte V. Immunological aspects of dengue haemorrhagic fever: studies in Thailand. Southeast Asian J Trop Med Public Health. 1987;18(3):312–5. [PubMed] [Google Scholar]
- 30.Ruangjirachuporn W, Boonpucknavig S, Nimmanitya S. Circulating immune complexes in serum from patients with dengue haemorrhagic fever. Clin Exp Immunol. 1979;36(1):46–53. [PMC free article] [PubMed] [Google Scholar]
- 31.Lin CF, Chiu SC, Hsiao YL, Wan SW, Lei HY, Shiau AL, et al. Expression of cytokine, chemokine, and adhesion molecules during endothelial cell activation induced by antibodies against dengue virus nonstructural protein 1. J Immunol. 2005;174(1):395–403. 10.4049/jimmunol.174.1.395 [DOI] [PubMed] [Google Scholar]
- 32.Diamond MS, Roberts TG, Edgil D, Lu B, Ernst J, Harris E. Modulation of Dengue virus infection in human cells by alpha, beta, and gamma interferons. J Virol. 2000;74(11):4957–66. 10.1128/jvi.74.11.4957-4966.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Avirutnan P, Malasit P, Seliger B, Bhakdi S, Husmann M. Dengue virus infection of human endothelial cells leads to chemokine production, complement activation, and apoptosis. J Immunol. 1998;161(11):6338–46. [PubMed] [Google Scholar]
- 34.Chen YC, Wang SY. Activation of terminally differentiated human monocytes/macrophages by dengue virus: productive infection, hierarchical production of innate cytokines and chemokines, and the synergistic effect of lipopolysaccharide. J Virol. 2002;76(19):9877–87. 10.1128/jvi.76.19.9877-9887.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Bokisch VA, Top FH Jr., Russell PK, Dixon FJ, Muller-Eberhard HJ. The potential pathogenic role of complement in dengue hemorrhagic shock syndrome. N Engl J Med. 1973;289(19):996–1000. 10.1056/NEJM197311082891902 [DOI] [PubMed] [Google Scholar]
- 36.Butthep P, Chunhakan S, Yoksan S, Tangnararatchakit K, Chuansumrit A. Alteration of cytokines and chemokines during febrile episodes associated with endothelial cell damage and plasma leakage in dengue hemorrhagic fever. Pediatr Infect Dis J. 2012;31(12):e232–8. 10.1097/INF.0b013e31826fd456 [DOI] [PubMed] [Google Scholar]
- 37.Warke RV, Martin KJ, Giaya K, Shaw SK, Rothman AL, Bosch I. TRAIL is a novel antiviral protein against dengue virus. J Virol. 2008;82(1):555–64. 10.1128/JVI.01694-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Becerra A, Warke RV, de Bosch N, Rothman AL, Bosch I. Elevated levels of soluble ST2 protein in dengue virus infected patients. Cytokine. 2008;41(2):114–20. 10.1016/j.cyto.2007.11.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Houghton-Trivino N, Salgado DM, Rodriguez JA, Bosch I, Castellanos JE. Levels of soluble ST2 in serum associated with severity of dengue due to tumour necrosis factor alpha stimulation. J Gen Virol. 2010;91(Pt 3):697–706. 10.1099/vir.0.012971-0 [DOI] [PubMed] [Google Scholar]
- 40.Guerrero CD, Arrieta AF, Ramirez ND, Rodriguez LS, Vega R, Bosch I, et al. High plasma levels of soluble ST2 but not its ligand IL-33 is associated with severe forms of pediatric dengue. Cytokine. 2013;61(3):766–71. 10.1016/j.cyto.2012.12.024 [DOI] [PubMed] [Google Scholar]
- 41.Hsieh CC, Cia CT, Lee JC, Sung JM, Lee NY, Chen PL, et al. A Cohort Study of Adult Patients with Severe Dengue in Taiwanese Intensive Care Units: The Elderly and APTT Prolongation Matter for Prognosis. PLoS Negl Trop Dis. 2017;11(1):e0005270 10.1371/journal.pntd.0005270 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Laoprasopwattana K, Binsaai J, Pruekprasert P, Geater A. Prothrombin Time Prolongation was the Most Important Indicator of Severe Bleeding in Children with Severe Dengue Viral Infection. J Trop Pediatr. 2017;63(4):314–20. 10.1093/tropej/fmw097 [DOI] [PubMed] [Google Scholar]
- 43.Chua MN, Molanida R, de Guzman M, Laberiza F. Prothrombin time and partial thromboplastin time as a predictor of bleeding in patients with dengue hemorrhagic fever. Southeast Asian J Trop Med Public Health. 1993;24 Suppl 1:141–3. [PubMed] [Google Scholar]
- 44.Huang YH, Liu CC, Wang ST, Lei HY, Liu HL, Lin YS, et al. Activation of coagulation and fibrinolysis during dengue virus infection. J Med Virol. 2001;63(3):247–51. 10.1002/1096-9071(200103)63:3<247::aid-jmv1008>3.0.co;2-f [DOI] [PubMed] [Google Scholar]
- 45.Marchi R, Nagaswami C, Weisel JW. Fibrin formation and lysis studies in dengue virus infection. Blood Coagul Fibrinolysis. 2009;20(7):575–82. 10.1097/MBC.0b013e32832fb1cf [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Chuang YC, Lei HY, Lin YS, Liu HS, Wu HL, Yeh TM. Dengue virus-induced autoantibodies bind to plasminogen and enhance its activation. J Immunol. 2011;187(12):6483–90. 10.4049/jimmunol.1102218 [DOI] [PubMed] [Google Scholar]
- 47.Fernando S, Wijewickrama A, Gomes L, Punchihewa CT, Madusanka SD, Dissanayake H, et al. Patterns and causes of liver involvement in acute dengue infection. BMC Infect Dis. 2016;16:319 10.1186/s12879-016-1656-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nguyen TL, Nguyen TH, Tieu NT. The impact of dengue haemorrhagic fever on liver function. Res Virol. 1997;148(4):273–7. 10.1016/s0923-2516(97)88364-1 [DOI] [PubMed] [Google Scholar]
- 49.Laoprasopwattana K, Jundee P, Pruekprasert P, Geater A. Outcome of Severe Dengue Viral Infection-caused Acute Liver Failure in Thai Children. J Trop Pediatr. 2016;62(3):200–5. 10.1093/tropej/fmv099 [DOI] [PubMed] [Google Scholar]
- 50.Dutta AK, Biswas A, Baruah K, Dhariwal AC. National guidelines for diagnosis and management of dengue fever/dengue haemorrhagic fever and dengue shock syndrome. J Indian Med Assoc. 2011;109(1):30–5. [PubMed] [Google Scholar]
- 51.Srikiatkhachorn A, Ajariyakhajorn C, Endy TP, Kalayanarooj S, Libraty DH, Green S, et al. Virus-induced decline in soluble vascular endothelial growth receptor 2 is associated with plasma leakage in dengue hemorrhagic Fever. J Virol. 2007;81(4):1592–600. 10.1128/JVI.01642-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Libraty DH, Endy TP, Houng HS, Green S, Kalayanarooj S, Suntayakorn S, et al. Differing influences of virus burden and immune activation on disease severity in secondary dengue-3 virus infections. J Infect Dis. 2002;185(9):1213–21. 10.1086/340365 [DOI] [PubMed] [Google Scholar]
- 53.Potts JA, Rothman AL. Clinical and laboratory features that distinguish dengue from other febrile illnesses in endemic populations. Trop Med Int Health. 2008;13(11):1328–40. 10.1111/j.1365-3156.2008.02151.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Sridhar S, Luedtke A, Langevin E, Zhu M, Bonaparte M, Machabert T, et al. Effect of Dengue Serostatus on Dengue Vaccine Safety and Efficacy. N Engl J Med. 2018;379(4):327–40. 10.1056/NEJMoa1800820 [DOI] [PubMed] [Google Scholar]
- 55.Aguiar M, Halstead SB, Stollenwerk N. Consider stopping dengvaxia administration without immunological screening. Expert Rev Vaccines. 2017;16(4):301–2. 10.1080/14760584.2017.1276831 [DOI] [PubMed] [Google Scholar]
- 56.Velasco-Salas ZI, Sierra GM, Guzman DM, Zambrano J, Vivas D, Comach G, et al. Dengue seroprevalence and risk factors for past and recent viral transmission in Venezuela: a comprehensive community-based study. Am J Trop Med Hyg. 2014;91(5):1039–48. 10.4269/ajtmh.14-0127 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lanciotti RS, Calisher CH, Gubler DJ, Chang GJ, Vorndam AV. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J Clin Microbiol. 1992;30(3):545–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Bruno VD, Benedetto U. Linear mixed-effect models in longitudinal data analysis: Shaken not stirred. J Thorac Cardiovasc Surg. 2018. [DOI] [PubMed] [Google Scholar]
- 59.Ross RA, Lee ML, Delaney ML, Onderdonk AB. Mixed-effect models for predicting microbial interactions in the vaginal ecosystem. J Clin Microbiol. 1994;32(4):871–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Naumova EN, Must A, Laird NM. Tutorial in Biostatistics: Evaluating the impact of ‘critical periods’ in longitudinal studies of growth using piecewise mixed effects models. Int J Epidemiol. 2001;30(6):1332–41. 10.1093/ije/30.6.1332 [DOI] [PubMed] [Google Scholar]
- 61.Bates D, Martin M, Bolker B, Walker S. Fitting Linear Mixed-Effects Models Using lme4. Journal of Statistical Software. 2015;67(1):57446. [Google Scholar]
- 62.Yang Z. PAML 4: phylogenetic analysis by maximum likelihood. Mol Biol Evol. 2007;24(8):1586–91. 10.1093/molbev/msm088 [DOI] [PubMed] [Google Scholar]
- 63.Loytynoja A. Phylogeny-aware alignment with PRANK. Methods Mol Biol. 2014;1079:155–70. 10.1007/978-1-62703-646-7_10 [DOI] [PubMed] [Google Scholar]
- 64.Schmidt DJ, Pickett BE, Camacho D, Comach G, Xhaja K, Lennon NJ, et al. A phylogenetic analysis using full-length viral genomes of South American dengue serotype 3 in consecutive Venezuelan outbreaks reveals a novel NS5 mutation. Infect Genet Evol. 2011;11(8):2011–9. 10.1016/j.meegid.2011.09.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Warke RV, Xhaja K, Martin KJ, Fournier MF, Shaw SK, Brizuela N, et al. Dengue virus induces novel changes in gene expression of human umbilical vein endothelial cells. J Virol. 2003;77(21):11822–32. 10.1128/jvi.77.21.11822-11832.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Basu A, Chaturvedi UC. Vascular endothelium: the battlefield of dengue viruses. FEMS Immunol Med Microbiol. 2008;53(3):287–99. 10.1111/j.1574-695X.2008.00420.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Palabrica T, Lobb R, Furie BC, Aronovitz M, Benjamin C, Hsu YM, et al. Leukocyte accumulation promoting fibrin deposition is mediated in vivo by P-selectin on adherent platelets. Nature. 1992;359(6398):848–51. 10.1038/359848a0 [DOI] [PubMed] [Google Scholar]
- 68.Dudas G, Carvalho LM, Bedford T, Tatem AJ, Baele G, Faria NR, et al. Virus genomes reveal factors that spread and sustained the Ebola epidemic. Nature. 2017;544(7650):309–15. 10.1038/nature22040 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Metsky HC, Matranga CB, Wohl S, Schaffner SF, Freije CA, Winnicki SM, et al. Zika virus evolution and spread in the Americas. Nature. 2017;546(7658):411–5. 10.1038/nature22402 [DOI] [PMC free article] [PubMed] [Google Scholar]