

Abstract
目的
探讨血清淀粉样蛋白A1(SAA1)基因rs4638289、rs7131332位点多态性与川崎病(KD)患儿发病及其并发冠状动脉病变(CAL)的相关性。
方法
随机选取2013~2017年间住院治疗的105例汉族KD患儿为KD组,另选取同期行健康体检的100例汉族儿童为对照组。KD组根据是否合并CAL,分为CAL组(n=23)和无冠状动脉病变(NCAL)组(n=82)。运用聚合酶链反应-限制性片段长度多态性分析法分析各组SAA1基因rs4638289、rs7131332位点多态性。
结果
KD组SAA1基因rs4638289位点AA、AT、TT基因型分布和A、T等位基因分布与对照组比较差异均无统计学意义(P > 0.05)。CAL组rs4638289位点AA、AT、TT基因型分布与NCAL组比较差异有统计学意义(P=0.016),A、T等位基因分布比较差异无统计学意义(P > 0.05);携带AT基因型是KD并发CAL的保护因素(OR=0.276,95% CI:0.099~0.772,P=0.011)。KD组SAA1基因rs7131332位点AA、AG、GG基因型分布和A、G等位基因分布与对照组比较差异无统计学意义(P > 0.05)。CAL组rs7131332位点AA、AG、GG基因型分布和A、G等位基因分布与NCAL组比较差异无统计学意义(P > 0.05)。
结论
SAA1基因rs4638289、rs7131332位点多态性与KD发病无关,但rs4638289位点多态性与KD并发CAL有关,携带AT基因型可能使KD并发CAL风险降低。
Keywords: 川崎病, 血清淀粉样蛋白A1, 基因多态性, 冠状动脉病变, 儿童
Abstract
Objective
To study the association of the polymorphisms of the serum amyloid A1 (SAA1) gene at rs4638289 and rs7131332 loci with Kawasaki disease (KD) and its complication coronary artery lesion (CAL) in children.
Methods
A total of 105 Han children with KD who were hospitalized and treated from 2013 to 2017 were enrolled as the KD group. A total of 100 Han children who underwent physical examination were enrolled as the control group. According to the presence or absence of CAL, the KD group was further divided into a CAL group with 23 children and a non-CAL (NCAL) group with 82 children. Polymerase chain reaction-restriction fragment length polymorphism was used to investigate the polymorphisms of the SAA1 gene at rs4638289 and rs7131332 loci.
Results
For the locus rs4638289 of the SAA1 gene, there were no significant differences between the KD and control groups in the genotype frequencies of AA, AT, and TT and the allele frequencies of A and T (P > 0.05). But there were significant differences between the CAL and NCAL groups in the genotype frequencies of AA, AT, and TT (P=0.016), while there were no significant differences in the allele frequencies of A and T (P > 0.05). AT genotype was a protective factor against CAL (OR=0.276, 95%CI:0.099-0.772, P=0.011). For the locus rs7131332 of the SAA1 gene, there were no significant differences between the KD and control groups in the genotype frequencies of AA, AG, and GG and the allele frequencies of A and G (P > 0.05). There were also no significant differences between the CAL and NCAL groups in the genotype frequencies of AA, AG, and GG and the allele frequencies of A and G (P > 0.05).
Conclusions
Polymorphisms of the SAA1 gene at loci rs4638289 and rs7131332 are not associated with the onset of KD, while the polymorphism at the locus rs4638289 is associated with CAL in KD patients. KD patients with genotype AT may have a reduced risk of CAL.
Keywords: Kawasaki disease, Serum amyloid A1, Polymorphism, Coronary artery lesion, Child
川崎病(Kawasaki disease, KD)是一种急性血管炎性疾病,主要侵犯中、小动脉,以冠状动脉病变(coronary artery lesion, CAL)为主。KD多发于婴幼儿和学龄前儿童,男女之比约(1.5~1.8): 1[1]。未经治疗的KD患儿的心脏损害高达25%,约3%~5%接受治疗的患儿仍会发展为CAL[2]。KD在全世界各地区发病率呈逐年上升趋势[3],在发达国家和一些发展中国家已经取代风湿性心脏病,成为儿童获得性心脏病的最常见原因[4]。KD发病机制至今尚不明确,目前认为与感染、免疫功能异常和遗传易感性因素等相关[5]。血清淀粉样蛋白A(serum amyloid A, SAA)是由肝脏产生的炎症性载脂蛋白家族,可引起全身系统性炎症。Alwaili等[6]发现SAA基因多态性与冠状动脉性疾病的发生密切相关,可更有效地预测冠状动脉扩张的发生。目前国内外尚无SAA基因多态性与KD相关性的研究,SAA1基因rs4638289、rs7131332位点多态性在国内外研究较多。Zhang等[7]发现与非肥胖儿童相比,SAA基因rs4638289位点多态性频率在肥胖儿童中显著增加。Xie等[8]发现SAA基因rs4638289位点与非糖尿病患者血浆葡萄糖水平显著相关。Zhang等[9]发现SAA基因rs7131332位点多态性与脑梗死显著相关,rs7131332位点AA基因型可能是脑梗死患者的高危因素。本研究对SAA基因rs4638289、rs7131332位点多态性进行分析,旨在探讨该基因位点多态性与KD发病及其并发CAL之间是否存在关联,进而从分子遗传学角度为探索KD的遗传易感性提供参考。
1. 资料与方法
1.1. 研究对象
随机选取2013~2017年间在中南大学湘雅医学院附属株洲医院治疗的105例汉族KD患儿为KD组,其中男66例,女39例,年龄7个月至6岁,平均年龄30±17个月。所有患儿均符合KD诊断标准[10],入院前均未经丙种球蛋白和阿司匹林治疗。KD组再根据是否合并CAL[10],分为CAL组和无冠状动脉损伤(non-CAL, NCAL)组。CAL组男13例,女10例,平均年龄29±16个月;NCAL组男53例,女29例,平均年龄31±17个月。选取同期在该院体检的汉族儿童100例为对照组,其中男60例,女40例,年龄7个月至6岁,平均年龄33±17个月。对照组儿童均无感染性疾病、风湿性疾病、心血管疾病及既往KD病史。KD组与对照组之间的性别构成(χ2=0.177,P=0.674)及年龄(t=-1.250,P=0.213)比较差异均无统计学意义,CAL组和NCAL组之间的性别构成(χ2=0.506,P=0.626)及年龄(t=0.407,P=0.685)比较差异均无统计学意义。各组SAA1基因rs4638289、rs7131332位点基因型分布符合Hardy-Weinberg遗传平衡定律(P > 0.05),表明选取对象具有群体代表性,样本量符合统计学计算最低要求。本研究已获得该院医学伦理委员会批准,伦理审批号:(2018)伦审(K)第(0324)号。全部入选对象均提前征得患者儿监护人知情同意,并签署知情同意书。
1.2. 标本采集
患儿于入院第1天在其家长或监护人知情同意下,使用一次性采血管(含抗凝剂5%的EDTA-K2)抽取2 mL静脉血,立即提取DNA或置于-80℃冰箱保存,2个月内提取DNA。
1.3. 基因组DNA的提取
应用Promega DNA提取试剂盒,按照说明书要求提取血液基因组DNA,并进行浓度和纯度的测定,置于-80℃冰箱保存。
1.4. 基因多态性的检测
应用聚合酶链-限制性片段长度多态性技术(polymerase chain reaction-restriction fragment length polymorphism, PCR-RFLP)鉴定KD组和对照组SAA1基因rs4638289、rs7131332位点基因型。SAA1基因rs4638289位点上游引物:5'-GGTTGTTGAAGTCTATGTTATGCC-3';下游引物:5'-CTCCTCGTTCATTCTCTTCTCAGT-3',片段长度为399 bp。SAA1基因rs7131332位点上游引物:5'-GTAGGGACTTTGTTGAGAAGCC-3';下游引物:5'-GGCTTGGGCTGTACTCTGAA-3',片段长度为385 bp。引物由上海生工生物工程技术服务有限公司合成。PCR反应体系25 μL:T3 Super PCR Mix 22 μL,上下游引物各1 μL,DNA 1 μL。PCR反应条件均为:98℃预变性2 min;98℃变性10 s,59℃退火10 s,72℃延伸5 s,35个循环;72℃终末延伸2 min,4℃保存。反应在PCR扩增仪(美国Applied Biosystems公司)上进行。扩增产物进行琼脂糖凝胶电泳并于紫外灯下照相,初步鉴定扩增是否成功。扩增产物送测序公司(上海铂尚生物技术有限公司)进行基因测序。测序结果在Chromas软件中进行阅读分析。
1.5. 统计学分析
采用SPSS 22.0软件对数据进行统计分析,符合正态分布计量资料采用均数±标准差(x±s)表示,基因型和等位基因频率采用百分率(%)表示,两组间比较采用χ2检验。使用优势比(OR)评估相对危险度并计算95%置信区间(95%CI)。P < 0.05为差异有统计学意义。
2. 结果
2.1. 临床资料
KD组临床表现有:发热105例(100%),皮疹73例(69.5%),非化脓性眼结合膜充血82例(78.1%),口腔黏膜充血93例(88.6%),非化脓性淋巴结肿大67例(63.8%),手足硬肿64例(61.0%),指趾端脱皮65例(61.9%),肛周脱皮42例(40.0%)。在KD组中,有冠状动脉扩张者23例(21.9%)。
2.2. SAA1基因PCR扩增
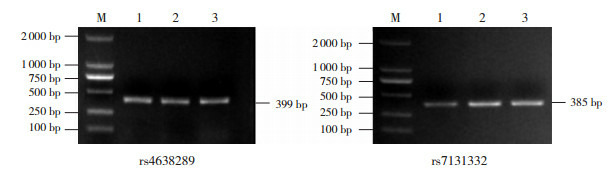
结果按PCR反应体系及条件扩增,经1.5%琼脂糖凝胶电泳,以Marker为核酸分子质量参照标准,PCR产物条带位于300~400 bp之间,扩增产物为目标片段,且附近没有其他条带,符合实验要求,见图 1。
1.
SAA1基因PCR电泳图
M:Marker;1~3泳道:SAA1基因片段的PCR产物。
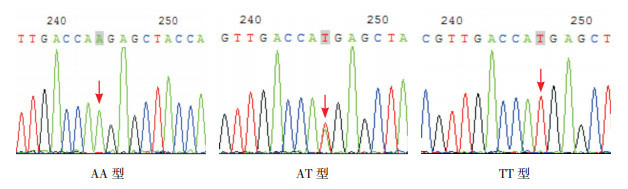
2.3. SAA1基因测序结果
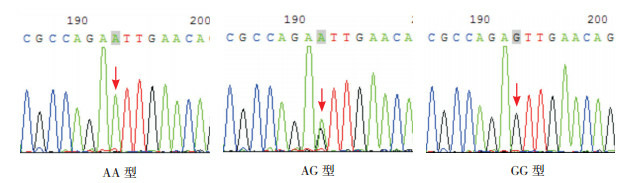
SAA1基因rs4638289位点多态性表现为A突变为T,基因型为AA、AT、TT。rs7131332位点多态性表现为A突变为G,基因型为AA、AG、GG。见图 2~3。
2.
SAA1基因rs4638289位点测序图
箭头所指为多态位点,单峰为纯合子基因型,套峰为杂合子基因型。
3.
SAA1基因rs7131332位点测序图
箭头所指为多态位点,单峰为纯合子基因型,套峰为杂合子基因型。
2.4. SAA1基因rs4638289位点基因型及等位基因分布
KD组rs4638289位点AA、AT、TT基因型分布和A、T等位基因分布与对照组比较差异无统计学意义(P > 0.05,表 1)。CAL组rs4638289位点AA、AT、TT基因型分布与NCAL组比较差异有统计学意义(P=0.016);A、T等位基因分布差异无统计学意义(P > 0.05,表 2)。携带AT基因型是KD并发CAL的保护因素(OR=0.276,95%CI:0.099~0.772,P=0.011),见表 3。
1.
KD组及对照组rs4638289位点基因型及等位基因分布比较 [例(%)]
| 组别 | 例数 | 基因型 | 等位基因 | ||||
| AA | AT | TT | A | T | |||
| 对照组 | 100 | 5(5.0) | 43(43.0) | 52(52.0) | 53(26.5) | 147(73.5) | |
| KD组 | 105 | 8(7.6) | 52(49.5) | 45(42.9) | 68(32.4) | 142(67.6) | |
| χ2值 | 1.929 | 1.703 | |||||
| P值 | 0.394 | 0.196 | |||||
2.
CAL组及NCAL组rs4638289位点基因型及等位基因分布比较 [例(%)]
| 组别 | 例数 | 基因型 | 等位基因 | ||||
| AA | AT | TT | A | T | |||
| NCAL组 | 82 | 4(4.9) | 46(56.1) | 32(39.0) | 54(32.9) | 110(67.1) | |
| CAL组 | 23 | 4(12.5) | 6(26.1) | 13(56.5) | 14(30.4) | 32(69.6) | |
| χ2值 | 8.241 | 0.102 | |||||
| P值 | 0.016 | 0.750 | |||||
3.
CAL组及NCAL组rs4638289位点基因型及等位基因分布风险分析 [例(%)]
| 组别 | 例数 | 基因型 | 等位基因 | ||||
| AA | AT | TT | A | T | |||
| NCAL组 | 82 | 4(4.9) | 46(56.1) | 32(39.0) | 54(32.9) | 110(67.1) | |
| CAL组 | 23 | 4(12.5) | 6(26.1) | 13(56.5) | 14(30.4) | 32(69.6) | |
| χ2值 | 3.348 | 6.471 | 2.245 | 0.102 | |||
| P值 | 0.067 | 0.011 | 0.134 | 0.750 | |||
| OR值 | 4.105 | 0.276 | 2.031 | 0.891 | |||
| 95%CI | 0.940~17.923 | 0.099~0.772 | 0.796~5.180 | 0.439~1.808 | |||
2.5. SAA1基因rs7131332位点基因型及等位基因分布
KD组rs7131332位点AA、AG、GG基因型分布和A、G等位基因分布与对照组比较差异均无统计学意义(P > 0.05,表 4)。CAL组rs7131332位点AA、AG、GG基因型分布和A、G等位基因分布与NCAL组比较差异均无统计学意义(P > 0.05,表 5)。
4.
KD组及对照组rs7131332位点基因型及等位基因分布比较 [例(%)]
| 组别 | 例数 | 基因型 | 等位基因 | ||||
| AA | AG | GG | A | G | |||
| 对照组 | 100 | 31(31.0) | 49(49.0) | 20(20.0) | 111(55.5) | 89(44.5) | |
| KD组 | 105 | 40(38.1) | 49(46.7) | 16(15.2) | 129(61.4) | 81(38.6) | |
| χ2值 | 1.464 | 1.483 | |||||
| P值 | 0.481 | 0.223 | |||||
5.
CAL组及NCAL组rs7131332位点基因型及等位基因分布比较 [例(%)]
| 组别 | 例数 | 基因型 | 等位基因 | ||||
| AA | AG | A | G | ||||
| NCAL组 | 82 | 30(36.6) | 39(47.6) | 13(15.9) | 99(60.4) | 65(39.6) | |
| CAL组 | 23 | 10(43.5) | 10(43.5) | 3(1.3) | 30(65.2) | 16(34.8) | |
| χ2值 | 0.388 | 0.357 | |||||
| P值 | 0.811 | 0.550 | |||||
3. 讨论
KD可在全球所有种族中发生,但在亚洲种族中发病率最高[11]。在美国,亚洲婴儿的患病率比黑人高3倍,比白人高6倍[12],显然遗传因素是影响KD发病的重要因素[13]。免疫系统的异常激活被认为是KD的主要特征,细胞因子和炎性介质相互影响,以增强免疫效果;基因多态性影响KD中细胞因子的表达,从而进一步影响KD的发病。目前已经发现多个细胞因子基因多态性与KD易感性有关,如肿瘤坏死因子基因、白细胞介素相关基因、血小板内皮细胞黏附分子-1等一些调节免疫、炎症的单核苷酸多态性位点[14]。多个多态性等位基因会影响KD的易感性,不同种族的人群具有不同的等位基因表达模式。SAA是一个包括SAA1~4同源体的炎症性载脂蛋白,其在炎症的急性反应期中显著上调(> 1 000倍),已被用作各种炎症和自身免疫性疾病的生物标志物[15]。SAA基因簇位于11号染色体的短臂上,位于p15.1处,只有SAA1和SAA2基因分别编码产生的SAA1和SAA2蛋白起主要作用。SAA1基因的多态性可赋予免疫应答灵活性,其基因的变化可影响SAA的表达从而影响相应细胞因子的功能[16]。
脂质代谢紊乱是KD急性期的表现,并最终导致血清总胆固醇降低和三酰甘油增加[2],这种脂质状态改变可能在KD诊断后3年内持续存在。超声显示患有KD病史的年轻成年人的血管弹性降低和颈动脉内膜中层厚度异常[17],并且尸检研究表明KD患者可能患有更严重的动脉粥样硬化病变[18]。Xu等[19]发现SAA1基因与中国人群的血脂水平有关。Carty等[20]发现SAA基因多态性与心血管疾病和高密度脂蛋白(HDL)相关。Xie等[21-22]发现SAA基因多态性与颈动脉内膜中层厚度和冠状动脉疾病有关。Whitin等[23]发现KD患儿急性期的SAA水平明显高于其他疾病,SAA可能在KD病理生理学中起重要作用。
本研究对KD患儿及健康儿童进行病例对照研究,比较SAA1基因型分布情况。临床资料显示,KD患儿中CAL 23例,发生率为21.9%。KD组与对照组比较,SAA1基因rs4638289、rs7131332位点基因型和等位基因分布比较差异均无统计学意义。CAL组与NCAL组比较,rs4638289位点AA、AT、TT基因型分布差异有统计学意义,SAA1基因rs4638289位点多态性可能与KD并发CAL有关,携带AT基因型可能使KD并发CAL风险降低。SAA在脂质代谢中起重要作用,并参与细菌清除、炎症调节和肿瘤发病机制。一方面,SAA作为一种载脂蛋白,与脂质水平相关,例如80%的SAA可与HDL结合,富含SAA的HDL抗氧化作用减弱,可导致动脉粥样硬化的形成[24]。SAA的亲脂性有助于将其与脂质运输和代谢及CAL联系起来。另一方面,SAA作为细胞因子样蛋白的功能已在细胞间通讯及炎症、免疫、肿瘤和保护途径的反馈中得到公认[25]。Sun等[26]发现SAA通过依赖MYD88的机制改变巨噬细胞表型并调节巨噬细胞功能,从而影响炎症反应的程度和持续时间。SAA促进了中性粒细胞的成熟和动员[27],同时上调内皮及单核-巨噬细胞系中IL-1、IL-6、IL-23、TNF-α等众多炎症因子的表达[28],加重机体的炎症水平,同时诱导抗原细胞和单核细胞组织因子释放,启动凝血系统,促使其在动脉血管壁内的沉积增加,影响冠状动脉及血管疾病的发生发展。
综上所述,本研究发现SAA1基因rs4638289位点与KD并发CAL有关,携带AT基因型可能使KD并发CAL风险降低。能否阻断KD患儿体内SAA的表达,从而减轻或抑制KD的发病,为临床诊疗提供了一个新思路。但SAA1基因多态性与血脂水平之间的关系尚不清楚,仍未阐明SAA如何驱动CAL的确切机制。本研究结果尚受到研究对象来源的地区、种族及样本量较小的影响,KD及其CAL与SAA1基因多态性的相关性,还有待进一步深入研究。
Biography
陈颖, 女, 硕士研究生, 住院医师
Funding Statement
湖南省自然科学基金(S2019JJSSLH0064)
References
- 1.Wu MH, Lin MT, Chen HC, et al. Postnatal risk of acquiring Kawasaki disease:a nationwide birth cohort database study. J Pediatr. 2017;180:80–86. doi: 10.1016/j.jpeds.2016.09.052. [DOI] [PubMed] [Google Scholar]
- 2.Marchesi A, Tarissi de Jacobis I, Rigante D, et al. Kawasaki disease:guidelines of the Italian Society of Pediatrics, part I-definition, epidemiology, etiopathogenesis, clinical expression and management of the acute phase. Ital J Pediatr. 2018;44(1):102. doi: 10.1186/s13052-018-0536-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Singh S, Vignesh P, Burgner D. The epidemiology of Kawasaki disease:a global update. Arch Dis Child. 2015;100(11):1084–1088. doi: 10.1136/archdischild-2014-307536. [DOI] [PubMed] [Google Scholar]
- 4.Martin M, Wrotniak BH, Hicar M. Suppressed plasmablast responses in febrile infants, including children with Kawasaki disease. PLoS One. 2018;13(3):e0193539. doi: 10.1371/journal.pone.0193539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Del Principe D, Pietraforte D, Gambardella L, et al. Pathogenetic determinants in Kawasaki disease:the haematological point of view. J Cell Mol Med. 2017;21(4):632–639. doi: 10.1111/jcmm.12992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Alwaili K, Bailey D, Awan Z, et al. The HDL proteome in acute coronary syndromes shifts to an inflammatory profile. Biochim Biophys Acta. 2012;1821(3):405–415. doi: 10.1016/j.bbalip.2011.07.013. [DOI] [PubMed] [Google Scholar]
- 7.Zhang X, Tang QZ, Wan AY, et al. SAA1 gene variants and childhood obesity in China. Lipids Health Dis. 2013;12:161. doi: 10.1186/1476-511X-12-161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Xie X, Ma YT, Yang YN, et al. SAA1 genetic polymorphisms are associated with plasma glucose concentration in non-diabetic subjects. http://cn.bing.com/academic/profile?id=feedb6c640098da84167508d54757523&encoded=0&v=paper_preview&mkt=zh-cn. Clin Chem Lab Med. 2013;51(12):2331–2334. doi: 10.1515/cclm-2013-0097. [DOI] [PubMed] [Google Scholar]
- 9.Zhang LJ, Yuan B, Li HH, et al. Associations of genetic polymorphisms of SAA1 with cerebral infarction. Lipids Health Dis. 2013;12:130. doi: 10.1186/1476-511X-12-130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.江 载芳, 申 昆玲, 沈 颖. 诸福棠实用儿科学. 北京: 人民卫生出版社; 2015. pp. 778–789. [Google Scholar]
- 11.Okubo Y, Nochioka K, Sakakibara H, et al. National survey of pediatric hospitalizations due to Kawasaki disease and coronary artery aneurysms in the USA. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=ae24bd8845476cf175a31ff479cd40ad. Clin Rheumatol. 2017;36(2):413–419. doi: 10.1007/s10067-016-3512-6. [DOI] [PubMed] [Google Scholar]
- 12.Rowley AH. Kawasaki disease:novel insights into etiology and genetic susceptibility. Annu Rev Med. 2011;62:69–77. doi: 10.1146/annurev-med-042409-151944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kuo HC, Hsu YW, Wu MS, et al. Intravenous immunoglobulin, pharmacogenomics, and Kawasaki disease. J Microbiol Immunol Infect. 2016;49(1):1–7. doi: 10.1016/j.jmii.2014.11.001. [DOI] [PubMed] [Google Scholar]
- 14.Xie X, Shi X, Liu M. The roles of genetic factors in Kawasaki disease:a systematic review and Meta-analysis of genetic association studies. http://cn.bing.com/academic/profile?id=348ba2cc7dee2e730b074c0ea5485fc5&encoded=0&v=paper_preview&mkt=zh-cn. Pediatr Cardiol. 2018;39(2):207–225. doi: 10.1007/s00246-017-1760-0. [DOI] [PubMed] [Google Scholar]
- 15.Burgess EJ, Hoyt LR, Randall MJ, et al. Bacterial lipoproteins constitute the TLR2-stimulating activity of serum amyloid A. J Immunol. 2018;201(8):2377–2384. doi: 10.4049/jimmunol.1800503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kapelski P, Skibinska M, Maciukiewicz M, et al. Association study of functional polymorphisms in interleukins and interleukin receptors genes:IL1A, IL1B, IL1RN, IL6, IL6R, IL10, IL10RA and TGFB1 in schizophrenia in Polish population. Schizophr Res. 2015;169(1-3):1–9. doi: 10.1016/j.schres.2015.10.008. [DOI] [PubMed] [Google Scholar]
- 17.McCrindle BW, McIntyre S, Kim C, et al. Are patients after Kawasaki disease at increased risk for accelerated atherosclerosis? J Pediatr. 2007;151(3):244–248. doi: 10.1016/j.jpeds.2007.03.056. [DOI] [PubMed] [Google Scholar]
- 18.Marchesi A, Tarissi de Jacobis I, Rigante D, et al. Kawasaki disease:guidelines of Italian Society of Pediatrics, part II-treatment of resistant forms and cardiovascular complications, follow-up, lifestyle and prevention of cardiovascular risks. Ital J Pediatr. 2018;44(1):103. doi: 10.1186/s13052-018-0529-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Xu XL, Sun XT, Pang L, et al. Rs12218 in SAA1 gene was associated with serum lipid levels. Lipids Health Dis. 2013;12:116. doi: 10.1186/1476-511X-12-116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Carty CL, Heagerty P, Heckbert SR, et al. Association of genetic variation in serum amyloid-A with cardiovascular disease and interactions with IL6, IL1RN, IL1beta and TNF gene in the cardiovascular health study. J Atheroscler Thromb. 2009;16(4):419–430. doi: 10.5551/jat.No968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Xie X, Ma YT, Yang YN, et al. Polymorphisms in the SAA1/2 gene are associated with carotid intima media thickness in healthy Han Chinese subjects:the cardiovascular risk survey. PLoS One. 2010;5(11):e13997. doi: 10.1371/journal.pone.0013997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Xie X, Ma YT, Yang YN, et al. Genetic polymorphisms of serum amyloid A1 and coronary artery disease risk. Tissue Antigens. 2015;85(3):168–176. doi: 10.1111/tan.12516. [DOI] [PubMed] [Google Scholar]
- 23.Whitin JC, Yu TS, Ling XB, et al. A novel truncated form of serum amyloid A in Kawasaki disease. PLoS One. 2016;11(6):e0157024. doi: 10.1371/journal.pone.0157024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Xie X, Ma YT, Yang YN, et al. Serum uric acid levels are associated with polymorphism in the SAA1 gene in Chinese subjects. PLoS One. 2012;7(6):e40263. doi: 10.1371/journal.pone.0040263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sack GH Jr. Serum amyloid A-a review. http://cn.bing.com/academic/profile?id=cfaf155c7db4d9118e489236310b01ff&encoded=0&v=paper_preview&mkt=zh-cn. Mol Med. 2018;24(1):46. doi: 10.1186/s10020-018-0047-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sun L, Zhou H, Zhu Z, et al. Ex vivo and in vitro effect of serum amyloid A in the induction of macrophage M2 markers and efferocytosis of apoptotic neutrophils. J Immunol. 2015;194(10):4891–4900. doi: 10.4049/jimmunol.1402164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Murdoch CC, Espenschied ST, Matty MA, et al. Intestinal serum amyloid A suppresses systemic neutrophil activation and bactericidal activity in response to microbiota colonization. PLoS Pathog. 2019;15(3):e1007381. doi: 10.1371/journal.ppat.1007381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lucherini OM, Lopalco G, Cantarini L, et al. Critical regulation of Th17 cell differentiation by serum amyloid-A signalling in Behcet's disease. Immunol Lett. 2018;201:38–44. doi: 10.1016/j.imlet.2018.10.013. [DOI] [PubMed] [Google Scholar]