

Abstract
Background:
Diagnosis of bacterial meningitis (BM) is challenging in newborn infants. Presently, biomarkers of BM have limited diagnostic accuracy. Analysis of cerebrospinal fluid (CSF) metabolites may be a useful diagnostic tool in BM.
Methods:
In a nested case-control study, we examined over 400 metabolites in CSF of uninfected infants and infants with culture-confirmed BM using gas- and liquid-chromatography mass spectrometry. Preterm and full-term infants in a Level III or IV Neonatal Intensive Care Unit were prospectively enrolled when evaluated for serious bacterial infection.
Results:
Over 200 CSF metabolites significantly differed in uninfected infants and infants with BM. Using machine-learning, we found as few as 6 metabolites distinguished infants with BM from uninfected infants in this pilot cohort. Further analysis demonstrated 3 metabolites associated with Group B Streptococcal meningitis.
Conclusions:
We report the first comprehensive metabolic analysis of CSF in infants with BM. In our pilot cohort, we derived a metabolic signature that predicted the presence or absence of BM, irrespective of gestational age, postnatal age, sex, race and ethnicity, presence of neurosurgical hardware, white blood cell count in CSF, and red blood cell contamination in CSF. Metabolic analysis may aid diagnosis of BM and facilitate clinical decision-making in infants.
INTRODUCTION
Bacterial meningitis (BM) is a devastating illness affecting 0.1-0.4 neonates per 1000 live births, with a higher incidence in preterm and chronically hospitalized infants1,2. Of previously healthy infants presenting to care with fever in the first and second month of life, 1.2% and 0.4%, respectively, will be diagnosed with BM3. Up to 10-20% of infants die, and 20-50% of survivors develop seizures, cognitive deficiencies, motor abnormalities, and hearing and visual impairments4. Despite declines in mortality, morbidity has not improved in decades5. The impact of neonatal BM is even greater in resource-limited settings worldwide, where mortality estimates are in excess of 50-60%, with a third of survivors exhibiting neurodevelopmental impairment6.
Prompt identification of BM is crucial to institute appropriate antimicrobial therapy and optimize outcomes7. The gold standard for diagnosis of BM is culture of cerebrospinal fluid (CSF). However, lumbar puncture (LP) is often deferred until after the administration of antibiotics in critically ill infants making culture results unreliable. In these cases, diagnosis of BM hinges on interpretation of CSF white blood cell counts (WBC) and differential, glucose, and protein. LP in infants is technically challenging, and 30% of CSF obtained is contaminated with enough blood to make interpretation of WBC counts difficult or impossible8. Further, neonatal BM may occur in the presence of normal CSF parameter9. Multiple alternative diagnostic biomarkers have been proposed, including acute phase reactants and cytokines, but none has yet achieved sufficient diagnostic accuracy to be employed in clinical practice10,11. As a result, in the absence of interpretable laboratory studies and culture of CSF, infants are often treated conservatively with long courses of intravenous antibiotics. There is a critical need for novel, accurate methods to minimize injury to the infant not only from BM but from associated interventions, including exposure to unnecessary broad-spectrum antimicrobial agents and prolonged need for central venous catheters.
Mechanisms underlying brain injury in infants with BM are poorly understood, limiting our ability to predict clinical outcomes. Furthermore, no therapies are currently available to protect the developing brain in the setting of BM. Comprehensive metabolic analysis, or metabolomics, has recently emerged as a powerful tool for the identification of novel biomarkers for diagnosis and prediction of disease severity and outcome. Recent investigations have demonstrated the ability of plasma metabolites to predict outcomes in adults with bacterial sepsis12,13. Additionally, limited metabolic analysis of CSF was shown to have some success segregating adults with BM from those with viral meningitis or uninfected subjects14. However, no investigations of the CSF metabolome in infants with meningitis have been performed to date15.
We hypothesized that measurement of CSF metabolites would aid in diagnosis of BM in infants. In this pilot study, we aimed to identify individual metabolites, as well as metabolic pathways, significantly altered in infants with culture-proven BM compared to uninfected infants. A deeper understanding of the metabolic milieu present in infant CSF has the potential to bolster our diagnostic approach to meningitis and also suggest novel therapies to protect the developing brain from lasting damage due to infection, thus promoting better outcomes.
METHODS
Study population and setting
Subjects were recruited from the Neonatal Intensive Care Units (NICUs) at Children’s Hospital of Philadelphia, Hospital of the University of Pennsylvania, and Pennsylvania Hospital. Recruited subjects included infants 0-12 months old who underwent LP or withdrawal of CSF from indwelling neurosurgical hardware (ventriculo-peritoneal [VP] shunt or Ommaya reservoir) for evaluation of possible meningitis. The Institutional Review Boards at the Children’s Hospital of Philadelphia and the University of Pennsylvania approved this study. Informed consent was obtained from families prior to analysis of CSF samples and data acquisition. From this larger, prospective cohort, we designed a nested case-control study to include infants with BM and uninfected infants matched by gestational age, postnatal age, sex and race.
Study definitions
Subjects for whom CSF culture results were positive for a bacterial pathogen and whom clinicians treated with a prolonged course of antibiotics were defined as having BM. Subjects whose CSF samples demonstrated no laboratory evidence of meningitis and whose CSF culture results were negative were defined as uninfected. To ensure CSF culture results of uninfected infants were truly negative, we selected from a large pool of culture-negative samples only those not receiving antibiotics prior to LP. Due to the critical and unstable nature of many of the infants in our cohort, 12/19 of the infants with BM received antibiotics prior to LP. Six of 12 had CSF samples obtained within 24 hours of receiving antibiotics. The remainder of the BM samples were acquired from 2-5 days into antibiotic therapy due to clinical situation or limited sample availability.
Study procedures and sample collection
We obtained an additional aliquot of CSF at the time of LP or shunt/reservoir tap performed due to suspicion for serious bacterial infection. Alternatively, frozen (−20°C), unused CSF samples were retrieved from our clinical laboratories following parental consent. CSF samples were coded and stored at −80°C. Given the exploratory nature of this study, sample size was driven by availability of positive samples for this relatively infrequent condition.
Sample Analysis
Specimens deidentified relative to patient information and disease status were shipped on dry ice to Metabolon, Inc. where metabolites were extracted and characterized by semi-quantitative liquid-and gas-chromatography mass spectrometry (LC/GC-MS)16,17. Volumes of specimens required to perform analysis were 50-100μL. Compounds were identified using Metabolon’s proprietary visualization and interpretation software, in which MS peaks representing individual compounds in CSF were cross-referenced against a library of over 4,000 known compounds16. A vast array of breakdown products of sugars, proteins, fats, cholesterols, and nucleic acids are detected using this technique. After MS peaks were identified, relative quantification of MS peaks was performed using area under the curve. Subsequent QC and data curation processes were performed to ensure accurate, consistent identification, as well as to minimize system artifacts, mis-assignments, and background noise. Library matches for each compound were verified for each sample.
Data collection
We abstracted demographic, clinical and laboratory data including risk factors for infection, culture results, and treatment details from the medical records of enrolled patients. Data were maintained in a coded electronic database.
Statistical Analysis
We computed summary statistics for demographic variables, risk factors, and laboratory parameters. Continuous variables were presented as medians and interquartile ranges, while categorical variables were presented as proportions and/or percentages. P values were calculated with Graphpad Prism 7 using the Mann-Whitney U test, Chi-square, or Fisher’s exact test, as applicable.
To compare metabolomics data between desired groups, raw compound data, expressed as relative intensities, were log2-transformed. We then used a linear model with the “limma” package in R18 to determine compounds whose relative intensities significantly differed between groups. Given the risk of false positives with a large data set undergoing multiple comparisons to find differentially abundant metabolites, p values were adjusted for the false discovery rate (FDR)19. Only those compounds whose adjusted p values were less than 0.05 were considered to significantly differ between study groups.
The machine learning classification algorithm random forests excels at creating decision trees in the setting of many input variables20. In our case, the decision tree of interest was whether a sample should be classified as BM or uninfected. Given the size of our cohort, random forests have the additional advantage of being less prone to overfitting than other classification algorithms. In order to generate a decision tree, random forests repeatedly “train” on a proportion of individuals in a cohort (20% in our case) then use the remaining proportion of the cohort to “test” the decision tree. While not equivalent to use of an independent cohort to test the validity of a signature of BM, this represents a form of internal validation. It is important to note that a signature of BM was not derived and tested on exactly the same subjects. We took a conservative and unbiased approach to derive a signature of BM with 3 times repeated 10-fold cross-validation and the random forests algorithm using the R package Caret21. Metabolites were then gradually eliminated from the prediction model with recursive feature elimination to test the performance of progressively more focused metabolic signatures of BM. The R package “random Forest”22 was used to construct a visual model of the prediction algorithm.
RESULTS
Patient characteristics
Subjects included 19 infants with culture-confirmed BM and 19 uninfected infants with no significant differences in gestational age, birthweight, postnatal age, sex, and race and ethnicity (Table 1). Overall, 58% of the cohort was male, and 68% was preterm. Seven infants with BM and 2 uninfected infants had bloodstream infections at the time of LP. Nine infants with BM had a VP shunt or Ommaya reservoir in place, compared to only 1 uninfected infant with indwelling neurosurgical hardware. CSF white blood cell count (WBC) and protein were significantly elevated, while CSF glucose was significantly decreased, in infants with BM versus uninfected infants. CSF red blood cell count (RBC) was significantly higher in subjects with BM. However, the proportion of subjects in each group with substantial red blood cell contamination of CSF, defined as ≥500 RBC/mm3 8, was not significantly different. Most bacterial isolates from CSF were gram positive (68%) (Supplemental Table S1). The most common pathogens identified in CSF were Streptococcus agalactiae (n=7) and Staphylococcus aureus (n=5). Of note, one infant with Staphylococcus epidermidis isolated from CSF was deemed infected by the medical team and received a full course of treatment.
Table 1. Demographic data for enrolled infants with culture-proven bacterial meningitis and uninfected infants without meningitis.
P values were calculated with Graphpad Prism 7 using the Mann-Whitney U test, Chi-square, or Fisher’s exact test, as applicable. BM: bacterial meningitis, IQR: interquartile range, VP: ventriculo-peritoneal, WBC: white blood cell count, RBC: red blood cell count.
| Variables | BM (N=19) | Uninfected (N=19) | p |
|---|---|---|---|
| Gestational age, weeks+days, median (IQR) | 35w6d (27w0d-38w0d) | 35w0d (26w5d-38w0d) | >0.99 |
| Postnatal age, days, median (IQR) | 40 (21-57) | 44 (30-65) | 0.86 |
| Birthweight, grams, median (IQR) | 2211 (905-3160) | 2210 (640-3090) | 0.73 |
| Preterm (<37 weeks gestation), n (%) | 13 (68%) | 13 (68%) | >0.99 |
| Male sex, n (%) | 11 (58%) | 13 (68%) | 0.74 |
| Race/Ethnicity, n (%) | 0.78 | ||
| Black, non-Hispanic | 8 (42%) | 6 (32%) | |
| White, non-Hispanic | 9 (47%) | 11 (58%) | |
| Other (Hispanic, Asian, or unknown) | 2 (11%) | 2 (10%) | |
| VP shunt or Ommaya reservoir, n (%) | 9 (47%) | 1 (5%) | 0.0078 |
| Culture-proven bacteremia, n (%) | 7 (37%) | 2 (10%) | 0.12 |
| CSF WBC (cells/mm3), median (IQR) | 215 (57-923) | 2 (0-4) | <0.0001 |
| CSF Glucose (g/dL), median (IQR) | 31 (20-46) | 52 (42-64) | <0.0001 |
| CSF Protein (g/dL), median (IQR) | 317 (171-709) | 76 (43-122) | <0.0001 |
| CSF RBC (cells/mm3), median (IQR) | 124 (29-2870) | 15 (3-623) | 0.042 |
| Greater than 500 cells/mm3 RBC, n (%) | 9 (47%) | 5 (26%) | 0.19 |
Derivation of a metabolic signature of BM
We first broadly indentified and quantified biochemicals in CSF of infants with BM and uninfected infants. Mass-spectrometry revealed that 210 of 422 individual metabolites detected in CSF significantly differed (adjusted p value <0.05) between infants with and without BM (Fig. 1a and Supplemental Table S2). Of the 210 metabolites, 196 were enriched in BM, while 14 were reduced.
Figure 1. Many individual metabolites in CSF differ between infants with bacterial meningitis and uninfected infants.
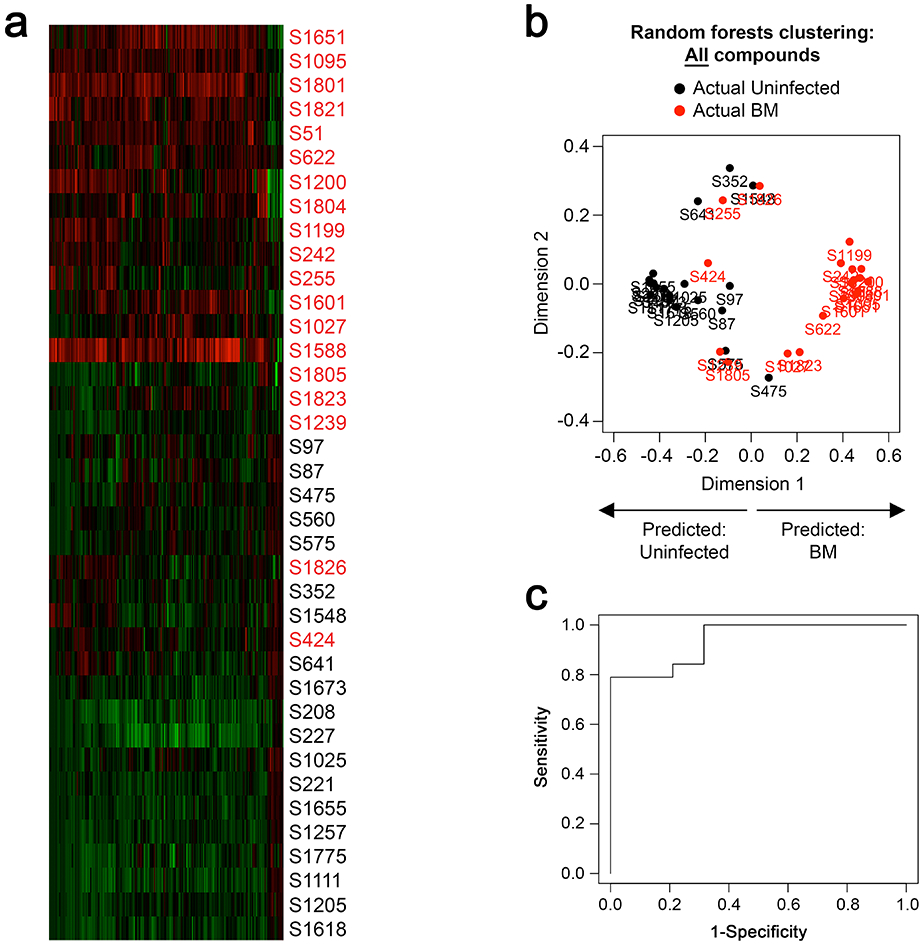
(a) Heat map in which columns represent individual compounds that significantly differ (adjusted p<0.05) between infants with BM and uninfected infants. Columns highlighted in red represent compounds significantly increased. Columns highlighted in green represent compounds significantly decreased. Rows represent individual subjects with BM (red text) and uninfected infants (black text). Samples are organized by Euclidian clustering based on levels of all significantly different compounds. (b) Multi-dimensional scaling (MDS) plot of uninfected infants (black dots) and infants with BM (red dots), organized by random forests based on levels of all 422 detected metabolites. Points closer together indicate similarity in levels of metabolites, while points farther apart indicate dissimilarity. Points in the left hemisphere of the plot are predicted to be uninfected infants. Points in the right hemisphere of the plot are predicted to have BM. (c) Receiver operating characteristic (ROC) curve for random forests modeling of disease status and 10-fold cross-validation with 3 repeats. Area under the curve (AUC) = 0.94.
We next sought to understand whether detectable metabolites, in aggregate, predicted presence or absence of bacterial meningitis. We took an unbiased approach using 3 times repeated 10-fold cross-validation and random forests to generate a prediction algorithm using combinations of metabolites. Based on levels of all 422 detected metabolites, random forests segregated infants with BM from uninfected infants with an AUC of 0.94 (Figs. 1b and 1c).
To find the minimum number of compounds whose levels best distinguished infants with BM from uninfected infants, we performed recursive feature elimination with random forests to identify the compounds most important to prediction of BM status or uninfected status. A “signature”, consisting of 9 of the most important metabolites, best segregated infants with BM and uninfected infants with an AUC of 0.97 (Fig. 2a). These 9 metabolites included: proline, N6-acetyllysine, taurine, cytidine, 2-hydroxyglutarate, ornithine, thymine, glutamate, α-ketoglutarate (Fig. 2b). Of note, we found that a signature of BM consisting only of the top 6 metabolites was similarly effective in classifying infants with BM and uninfected infants (AUC=0.97, Fig. 2a).
Figure 2. A focused metabolic signature associated with BM in our cohort of infants.

(a) Each point represents an individual AUC value (y-axis) of a predictive model using a progressively larger number of metabolites (x-axis). The highest AUC of 0.97 is seen using either the top 6 or top 9 metabolites to distinguish infants with BM from uninfected infants. (b) Standard box and whisker plots showing relative levels of the 9 compounds composing the optimal signature to distinguish uninfected infants from those with BM. Individual p values are: proline (p=0.000003); N6-AL, N6-acetyllysine (p=0.000008); taurine (p=0.00001); cytidine (p=0.000001); 2-HG, 2-hydroxygluatarate (p=0.0002); ornithine (p=0.0002); thymine (p=0.000007); glutamate (p=0.000003); and αKG, α-ketoglutarate (p=0.000007).
Metabolic signature of BM is independent of GA, PMA, CSF WBC, CSF RBC, and presence of neurosurgical hardware
We next assessed for metabolites that distinguished preterm and term infants with BM, but we found no metabolites that significantly differed between these two groups (Supplemental Table S3). We also tested whether metabolites in BM differed on the basis of postnatal age (PNA) and found that no metabolites significantly differed between infants older or younger than 30 days PNA with BM (Supplemental Table S4). Further, no compounds significantly differed between infants with BM and CSF WBC less than or equal to 100 versus those with CSF WBC greater than 100 (Supplemental Table S5). Likewise, no compounds significantly differed between infants with BM and CSF RBC less than 500 versus those with CSF RBC greater than 500 (Supplemental Table S6). Finally, no compounds significantly differed between infants with BM who did or did not have indwelling neurosurgical hardware (Supplemental Table S7, Supplemental Figure S1).
A secondary signature associated with GBS meningitis
As Streptococcus agalactiae (Group B Streptococcus, GBS) was the most common pathogen in our cohort, we sought to determine whether CSF metabolites in GBS BM differed from CSF metabolites in BM caused by other organisms. Three compounds were significantly altered in GBS BM compared to all other BM: α-hydroxyisocaproic acid (HICA), 2-hydroxy-3-methylvaleric acid (HMVA), and sucrose (Fig. 3). Shown both with boxplots (Fig. 3a) and graphically (Fig. 3b), GBS BM was associated with relatively higher levels of HICA and HMVA and relatively lower levels of sucrose. Non-GBS BM, on the other hand, was characterized by relatively lower levels of HICA and HMVA and relatively higher levels of sucrose. As this represented only a subset of our cohort of 19 infants, we do not depict the diagnostic value of these data with ROC curves. Of note, no metabolites significantly differed between gram-positive BM versus gram-negative BM (Supplemental Table S8).
Figure 3. Secondary metabolites associated with Group B Streptococcal meningitis.

(a) Standard box and whisker plots showing relative levels of the 3 individual compounds whose levels significantly differ between infants with GBS BM and infants with non-GBS BM. Individual adjusted p values are: α-hydroxyisocaproic acid (HICA, p=0.02), 2-hydroxy-3-methylvaleric acid (HMVA, p=0.02), and sucrose p=0.03. (b) 3-dimensional plot shows clustering of infants with GBS BM and non-GBS BM based on levels of HICA, HMVA, and sucrose. Blue dots represent individuals with GBS BM, and red dots represent individuals with non-GBS BM.
Evidence for dysregulation of key metabolic pathways in BM
We next investigated whether metabolites distinguishing infants with BM from uninfected infants belonged to common metabolic pathways. Using all 210 metabolites that significantly differed between infants with BM and uninfected infants, we performed pathway analysis with MetaboAnalyst23. Several metabolic pathways were significantly over-represented in BM (Fig. 4). Among the significantly over-represented pathways, alanine, aspartate, and glutamate metabolism demonstrated the highest impact score, signifying that key members of this pathway were those dyregulated in BM.
Figure 4. Families of metabolites distinguish subjects with BM from uninfected infants.

All compounds significantly altered (increased or decreased) between subjects with BM and uninfected infants were subjected to pathway analysis using MetaboAnalyst. All pathways found to be significantly over-represented among metabolites enriched in BM are shown. For each pathway, corresponding p values adjusted for the false discovery rate (FDR) are shown on the right-hand axis. The number of metabolites matched to each pathway are shown in white, embedded within bars depicting the impact score of each pathway. A higher impact score signifies that perturbations in matched compounds are likely to have a greater impact on the performance of the overall pathway.
DISCUSSION
We combined GC/LC-MS and machine learning to comprehensively characterize metabolites in CSF of infants undergoing evaluation for BM. We derived an optimal predictive model, composed of as few as 6 metabolites, that distinguished infants with BM from uninfected infants in our pilot cohort. Such a focused signature may be useful in future targeted clinical investigations, especially to help guide management in infants with sterile cultures but on antibiotics prior to LP. In secondary analysis of our cohort, 3 CSF metabolites were associated with BM caused by Group B Streptococcus, a highly prevalent organism among infected neonates and infants in the first months of life4.
Surprisingly, the derived metabolic signature predicted BM equally well in preterm and term infants of varying postnatal ages. We found no metabolites that significantly differed between preterm and term subjects with BM, despite known differences in development of the brain and immune system between preterm and term infants24,25. Also surprising was our finding that no metabolites significantly differed between infants with shunt- or reservoir-associated BM and infants with BM in the absence of indwelling neurosurgical hardware. Altogether, these data support a common pathway for the host response to BM in infants and speak to the potential clinical utility of using metabolites to diagnose BM in a wide range of infants.
Metabolites in CSF have aided diagnosis of BM for decades, but no individual metabolite or group of metabolites is perfectly sensitive or specific. Lactate and glucose were the first metabolites to be examined in CSF of patients with meningitis26. A meta-analysis supports that elevated CSF lactate, in particular, is associated with BM in adults and children27. Underscoring the validity of our approach, we also found significantly elevated lactate and decreased glucose in CSF of infants with BM relative to uninfected infants in our cohort. However, neither lactate nor glucose were among the most important metabolites found to distinguish subjects with BM from uninfected subjects. These data highlight the urgent unmet need for novel tools to diagnose BM in infants, especially in light of the well-known imperfect sensitivity and specificity of traditional laboratory values used to diagnose BM in infants, including CSF glucose9,28.
The compounds that compose the optimal predictive model of BM in our cohort are: proline, N6-acetyllysine, taurine, cytidine, 2-hydroxyglutarate, ornithine, thymine, glutamate, and α-ketoglutarate. Consistent with our data, others recently found that proline was among the top five amino acids that best segregated children with confirmed tuberculous meningitis from uninfected infants29. Our findings are also in agreement with prior investigations showing that glutamate and GABA were increased in CSF of children and adults with bacterial meningitis30-32. These data suggest that our metabolic signature may have broader applicability to other patient populations, but this remains to be tested. The data also highlight the power of broad metabolomic analysis, as proline, glutamate, and GABA, either alone or in combination, were not sufficient to predict disease state30-33. Only with simultaneous detection of several metabolites not reported in prior investigations could we distinguish subjects with BM from uninfected infants in our cohort.
Many of the most important compounds distinguishing subjects with BM from uninfected infants are intermediates of either glutamate, aspartate, and alanine metabolism or arginine and proline metabolism. These two pathways are intimately linked, both contributing to synthesis of glutamate, the canonical excitatory neurotransmitter in the central nervous system. Several compounds in our signature are intermediates of glutamate metabolism, including glutamate itself. Both ornithine and proline can be metabolized to glutamate through the intermediate pyrroline-5-carboxylate34. 2-hydroxyglutarate and its precursor, α-ketoglutarate, are also precursors of glutamate. Alpha-ketoglutarate is consumed, and glutamate is produced, by branched-chain amino acid (BCAA) metabolism, in which leucine is converted to HICA and isoleucine to HMVA. Both HICA and HMVA composed the “secondary” metabolic signature of GBS BM. Altogether, our data support the notion that glutamate dysregulation may play a key role in diagnosis and pathophysiology of BM in infants.
Glutamate acts at many postsynaptic receptors, including the AMPA and NMDA receptors. Perturbations in glutamate signaling can be caused by bacterial-derived products and have been shown in animal models of bacterial meningitis to cause profound neurotoxicity, which may be reversed by NMDA receptor antagonism35. Further, in a neonatal rat model of BM, nonspecific inhibition of excitatory amino acid receptors, including glutamate receptors, conferred protection against cortical and hippocampal injury36. GABA and 2-HG have also been shown to signal through the NMDA receptor and may contribute to neonatal neuronal excitotoxicity37,38. These data support the notion that direct or indirect blockade of potentially unbridled glutamate receptor activation in the setting of BM may limit brain injury in the setting of BM in infants. Basic investigations into the metabolic changes we uncovered in the setting of BM may lead to adjunctive therapies to antibiotics to minimize lifelong brain injury to infants with BM. Although many of the infants in our cohort study had multiple comorbidities related to prematurity or congenital malformations, future studies will also be directed at understanding a role for metabolites in predicting outcomes related to BM.
It is noteworthy that brains of infants with hypoxic-ischemic encephalopathy (HIE) also exhibit well-characterized elevations in glutamate levels, and many lines of evidence support a neurotoxic effect of this excess glutamate in the pathogenesis of brain injury in HIE39-41. Interestingly, glutamate was found to be reduced by magnetic resonance spectroscopy in brains of infants with HIE during the period of therapeutic hypothermia, relative to the period after rewarming42. In the same study, levels of taurine were elevated during cooling. Taurine, another one of the top nine compounds we found that composed the optimal predictive model of BM, has been implicated as a counter-regulatory compound to glutamate and may serve as a neuroprotectant in the setting of brain injury43. These data support that different insults to the neonatal brain may ultimately lead to similar mechanisms of injury and raise the question of whether therapeutic hypothermia also may be of benefit in BM, as it is in HIE.
This study has several strengths. Using a pilot cohort of infants, we describe a novel metabolic signature associated with BM in infants and a secondary signature associated with GBS BM. In this multi-center study, the population of infants was heterogeneous, and the organisms isolated reflect the prevalent bacterial pathogens in this age group. However, this study also has several limintations. As BM is a rare condition in infants, we recognize that our study is limited by small sample size and need for prospective validation of metabolites enriched in BM in an independent larger cohort of infants. We also have yet to test the performance of the derived signature of BM in subj ects other than infants, in a setting other than the NICU, in a cohort with more gram-negative organisms represented, and in a body fluid other than CSF. We also acknowledge that the preterm infants in our cohort were of advanced postnatal age, and thus the metabolic signature derived may more closely reflect that of more mature infants. Finally, as 12/19 of the infants in the BM group received antibiotics prior to LP, it will be important in future studies to validate our findings in a cohort of infants undergoing LP at initial presentation of illness, prior to initiation of antibiotic therapy.
In addition, a large proportion of infants with BM in our study had VP shunts or Ommaya reservoirs, perhaps reflecting the population of infants admitted to our main study site, a Level IV NICU. A local, rather than likely hematogenous, spread of infection from neurosurgical hardware may explain why only a third of the infants with BM in our cohort had concurrent sepsis, as opposed to higher rates of concurrent sepsis previously reported in the neonatal literature44,45. Nevertheless, we found the metabolic signature detected to be independent of the presence of neurosurgical hardware. The metabolic profile of such brains may well differ at baseline from those not requiring surgery, but this remains to be addressed in future studies.
CONCLUSIONS
We developed a metabolic signature in CSF that predicted BM in a heterogeneous pilot cohort of infants. Our findings have the potential to support the future development of a validated clinical tool to rapidly assay for BM with the potential to expedite diagnosis and therapy. Further analysis of metabolites in additional subjects may also provide insights into pathophysiologic mechanisms of brain injury in BM and suggest therapeutic targets to minimize morbidity and mortality in infants.
Supplementary Material
Bullet points:
Key message of the article: In a pilot cohort, metabolites in cerebrospinal fluid distinguished infants with bacterial meningitis from uninfected infants.
Adds to the existing literature: We report the first comprehensive metabolic analysis of cerebrospinal fluid in infants with bacterial meningitis.
Impact: Our findings may be used to improve diagnosis of bacterial meningitis and to offer mechanistic insights into the pathophysiology of bacterial meningitis in infants.
Acknowledgments
Statement of financial support: Dr. Gordon received support for this work from the American Academy of Pediatrics, Children’s Hospital of Philadelphia, and NIH grant T32 AI 118684. Drs. Srinivasan and Harris received support for this work from The Foerderer Awards at the Children’s Hospital of Philadelphia.
Footnotes
Patient consent statement: Informed consent was obtained from families prior to analysis of infant CSF samples and data acquisition.
Disclosure statement: The authors declare no conflicts of interest.
Category of study: Basic science
REFERENCES
- 1.Okike IO et al. Incidence, etiology, and outcome of bacterial meningitis in infants aged <90 days in the United kingdom and Republic of Ireland: prospective, enhanced, national population-based surveillance. Clin Infect Dis. 59, el 50–7 (2014). [DOI] [PubMed] [Google Scholar]
- 2.Lawn JE, Cousens S, Zupan J Lancet Neonatal Survival Steering Team. 4 million neonatal deaths: when? Where? Why? Lancet 365, 891–900 (2005). [DOI] [PubMed] [Google Scholar]
- 3.Biondi EA et al. Prevalence of Bacteremia and Bacterial Meningitis in Febrile Neonates and Infants in the Second Month of Life. JAMA Netw Open 2, el90874–12 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mann K & Jackson MA Meningitis. Pediatr Rev 29, 417–29– quiz 430 (2008). [DOI] [PubMed] [Google Scholar]
- 5.Libster R et al. Long-term Outcomes of Group B Streptococcal Meningitis. Pediatrics 130, e8–el5 (2012). [DOI] [PubMed] [Google Scholar]
- 6.Seale AC et al. Neonatal severe bacterial infection impairment estimates in South Asia, sub-Saharan Africa, and Latin America for 2010. Pediatr Res 74, 73–85 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Heath PT & Okike IO Neonatal bacterial meningitis: an update. Paediatrics and Child Health 20, 526–530 (2010). [Google Scholar]
- 8.Thomson J et al. Cerebrospinal Fluid Reference Values for Young Infants Undergoing Lumbar Puncture. Pediatrics 141, e20173405 (2018). [DOI] [PubMed] [Google Scholar]
- 9.Garges HP et al. Neonatal meningitis: what is the correlation among cerebrospinal fluid cultures, blood cultures, and cerebrospinal fluid parameters? Pediatrics 117, 1094–1100 (2006). [DOI] [PubMed] [Google Scholar]
- 10.Srinivasan L, Kilpatrick L, Shah SS, Abbasi S & Harris MC Cerebrospinal fluid cytokines in the diagnosis of bacterial meningitis in infants. Pediatr Res 80, 566–572 (2016). [DOI] [PubMed] [Google Scholar]
- 11.Wei T-T et al. Diagnostic Accuracy of Procalcitonin in Bacterial Meningitis Versus Nonbacterial Meningitis: A Systematic Review and Meta-Analysis. Medicine 95, e3079 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Langley RJ et al. An integrated clinico-metabolomic model improves prediction of death in sepsis. Sci Transl Med 5, 195ra95–195ra95 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Rogers AJ et al. Metabolomic Derangements Are Associated with Mortality in Critically 111 Adult Patients. PLoS ONE 9, e87538–7 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Coen M, O'Sullivan M, Bubb WA, Kuchel PW & Sorrell T Proton nuclear magnetic resonance-based metabonomics for rapid diagnosis of meningitis and ventriculitis. Clin Infect Dis. 41, 1582–1590 (2005). [DOI] [PubMed] [Google Scholar]
- 15.Gordon SM, Srinivasan L & Harris MC Neonatal Meningitis: Overcoming Challenges in Diagnosis, Prognosis, and Treatment with Omics. Front. Pediatr 5, 139 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Evans AM, DeHaven CD, Barrett T, Mitchell M & Milgram E Integrated, Nontargeted Ultrahigh Performance Liquid Chromatography/Electrospray Ionization Tandem Mass Spectrometry Platform for the Identification and Relative Quantification of the Small-Molecule Complement of Biological Systems. Anal. Chem 81, 6656–6667 (2009). [DOI] [PubMed] [Google Scholar]
- 17.Langley RJ et al. Integrative ‘Omic’ Analysis of Experimental Bacteremia Identifies a Metabolic Signature That Distinguishes Human Sepsis from Systemic Inflammatory Response Syndromes. Am J Respir Crit Care Med 190, 445–455 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ritchie ME et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 43, e47 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Benjamini Y & Hochberg Y Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. Journal of the Royal Statistical Society: Series B (Methodological) 57, 289–300 (1995). [Google Scholar]
- 20.Breiman L Random Forests. Machine Learning 45, 5–32 (2001). [Google Scholar]
- 21.Kuhn M Building predictive models in R using the caret package. Journal of Statistical Software 28, (2008). [Google Scholar]
- 22.Liaw A & Wiener M Classification and regression by randomForest. R News 2, 18–22 (2002). [Google Scholar]
- 23.Chong J & Xia J MetaboAnalystR: an R package for flexible and reproducible analysis of metabolomics data. Bioinformatics 34, 4313–4314 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Melville JM & Moss TJM The immune consequences of preterm birth. Front Neurosci 7, 79 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Volpe JJ, Kinney HC, Jensen FE & Rosenberg PA The developing oligodendrocyte: key cellular target in brain injury in the premature infant. Int. J. Dev. Neurosci 29, 423–440 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.De Sanctis AG, Killian JA & Garcia T Lactic acid of spinal fluid in meningitis: Practical diagnostic and prognostic value. Am J Dis Child 46, 239–249 (1933). [Google Scholar]
- 27.Huy NT et al. Cerebrospinal fluid lactate concentration to distinguish bacterial from aseptic meningitis: a systemic review and meta-analysis. Critical Care 14, R240 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Srinivasan L, Harris MC & Shah SS Lumbar Puncture in the Neonate: Challenges in Decision Making and Interpretation. Semin. Perinatol 36, 445–453 (2012). [DOI] [PubMed] [Google Scholar]
- 29.Mason S Cerebrospinal Fluid Amino Acid Profiling of Pediatric Cases with Tuberculous Meningitis. 1–8 (2017). doi: 10.3389/fnins.2017.00534 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Buryakova AV & Sytinsky IA Amino acid composition of cerebrospinal fluid in acute neuroinfections in children. Arch. Neurol 32, 28–31 (1975). [DOI] [PubMed] [Google Scholar]
- 31.Kuroda H et al. Cerebrospinal fluid GABA levels in various neurological and psychiatric diseases. J. Neurol. Neurosurg. Psychiatry 45, 257–260 (1982). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Spranger M et al. Excess glutamate in the cerebrospinal fluid in bacterial meningitis. J. Neurol. Sci 143, 126–131 (1996). [DOI] [PubMed] [Google Scholar]
- 33.Mason S, Reinecke CJ & Solomons R Cerebrospinal Fluid Amino Acid Profiling of Pediatric Cases with Tuberculous Meningitis. Front Neurosci 11, 248–8 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kanehisa M & Goto S KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 28, 27–30 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wippel C et al. Bacterial cytolysin during meningitis disrupts the regulation of glutamate in the brain, leading to synaptic damage. PLoS Pathog 9, e1003380 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Leib SL, Kim YS, Ferriero DM & Täuber MG Neuroprotective effect of excitatory amino acid antagonist kynurenic acid in experimental bacterial meningitis. J Infect Dis 173, 166–171 (1996). [DOI] [PubMed] [Google Scholar]
- 37.Ben-Ari Y, Khazipov R, Leinekugel X, Caillard O & Gaiarsa JL GABAA, NMDA and AMPA receptors: a developmentally regulated 'ménage à trois'. Trends Neurosci. 20, 523–529 (1997). [DOI] [PubMed] [Google Scholar]
- 38.Kölker S et al. NMDA receptor activation and respiratory chain complex V inhibition contribute to neurodegeneration in d-2-hydroxyglutaric aciduria. European Journal of Neuroscience 16, 21–28 (2002). [DOI] [PubMed] [Google Scholar]
- 39.Choi DW & Rothman SM The role of glutamate neurotoxicity in hypoxic-ischemic neuronal death. Annu. Rev. Neurosci 13, 171–182 (1990). [DOI] [PubMed] [Google Scholar]
- 40.Barks JD & Silverstein FS Excitatory amino acids contribute to the pathogenesis of perinatal hypoxic-ischemic brain injury. Brain Pathol. 2, 235–243 (1992). [DOI] [PubMed] [Google Scholar]
- 41.Pu Y et al. Increased detectability of alpha brain glutamate/glutamine in neonatal hypoxic-ischemic encephalopathy. AJNR Am J Neuroradiol 21, 203–212 (2000). [PMC free article] [PubMed] [Google Scholar]
- 42.Wisnowski JL et al. The effects of therapeutic hypothermia on cerebral metabolism in neonates with hypoxic-ischemic encephalopathy: An in vivo 1H-MR spectroscopy study. J Cereb Blood Flow Metab 36, 1075–1086 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Idrissi El, A. & Trenkner E. Growth factors and taurine protect against excitotoxicity by stabilizing calcium homeostasis and energy metabolism. J. Neurosci 19, 9459–9468 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wiswell TE, Baumgart S, Gannon CM & Spitzer AR No lumbar puncture in the evaluation for early neonatal sepsis: will meningitis be missed? Pediatrics 95, 803–806 (1995). [PubMed] [Google Scholar]
- 45.Stoll BJ et al. To Tap or Not to Tap: High Likelihood of Meningitis Without Sepsis Among Very Low Birth Weight Infants. Pediatrics 113, 1181–1186 (2004). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.